overexpression of polygalacturonase in transgenic apple trees leads to a range of novel
TRANSCRIPT

Overexpression of Polygalacturonase in Transgenic AppleTrees Leads to a Range of Novel Phenotypes InvolvingChanges in Cell Adhesion1
Ross G. Atkinson, Roswitha Schroder*, Ian C. Hallett, Daniel Cohen, and Elspeth A. MacRae
The Horticulture and Food Research Institute of New Zealand, Mount Albert Research Centre, Private Bag92 169, Auckland, New Zealand
Polygalacturonases (PGs) cleave runs of unesterified GalUA that form homogalacturonan regions along the backbone ofpectin. Homogalacturonan-rich pectin is commonly found in the middle lamella region of the wall where two adjacent cellsabut and its integrity is important for cell adhesion. Transgenic apple (Malus domestica Borkh. cv Royal Gala) trees wereproduced that contained additional copies of a fruit-specific apple PG gene under a constitutive promoter. In contrast toprevious studies in transgenic tobacco (Nicotiana tabacum) where PG overexpression had no effect on the plant (K.W.Osteryoung, K. Toenjes, B. Hall, V. Winkler, A.B. Bennett [1990] Plant Cell 2: 1239–1248), PG overexpression in transgenicapple led to a range of novel phenotypes. These phenotypes included silvery colored leaves and premature leaf sheddingdue to reduced cell adhesion in leaf abscission zones. Mature leaves had malformed and malfunctioning stomata thatperturbed water relations and contributed to a brittle leaf phenotype. Chemical and ultrastructural analyses were used torelate the phenotypic changes to pectin changes in the leaf cell walls. The modification of apple trees by a single PG genehas offered a new and unexpected perspective on the role of pectin and cell wall adhesion in leaf morphology and stomataldevelopment.
Polygalacturonases (PGs) are expressed in a widerange of tissues and developmental stages in plantsand are encoded by relatively large gene families(e.g. approximately 52 genes in Arabidopsis; TheArabidopsis Genome Initiative, 2000). PGs are asso-ciated with fruit ripening, cell separation processessuch as leaf and flower abscission, pod and antherdehiscence, pollen grain maturation, pathogen de-fense, plant-host interactions, and processes of cellexpansion, growth, and xylogenesis (for review, seeHadfield and Bennett, 1998; Bergey et al., 1999; Sitritet al., 1999; Torki et al., 1999; Wang et al., 2000).Endo-PGs cleave runs of unesterified GalUA thatform homogalacturonan regions along the backboneof pectin. Homogalacturonan-rich pectin is com-monly found in the middle lamella region of the wallwhere two adjacent cells abut (for example, see Knoxet al., 1990; Steele et al., 1997). The best characterized(fungal) endo-PG enzyme requires four to five con-secutive runs of unesterified GalUA residues forcleavage; however, little is known about the enzymeactivities and specificities of most cloned PGs fromplants.
Transgenic plants have been used to study the roleof endo-PGs in vivo. In tomato (Lycopersicon esculen-tum), down-regulation of the fruit-specific PG genepTOM6 under the control of the constitutive cauli-flower mosaic virus 35S promoter showed reduceddepolymerization of pectin polymers in fruit (Smithet al., 1990). Overexpression of PG in the ripening-inhibited mutant rin background restored PG activityand pectin degradation in fruit (Giovannoni et al.,1989). In both cases, only the fruit was affected by thetransgene expression; therefore, the gene product iso-lated from tomato fruit appeared to have fruit-specific PG activity. Further experiments where thepTOM6 gene was overexpressed in tobacco (Nicotianatabacum; Osteryoung et al., 1990) showed that thetomato protein was properly processed and localizedin the cell walls of leaves in tobacco. The enzymeshowed activity when extracted from transgenic to-bacco leaves and tested against tobacco cell wallextracts in vitro. However, no changes in leaf pheno-type were observed, nor were there any alterations tothe pectins in the tobacco cell walls in vivo.
Apple (Malus domestica Borkh. cv Royal Gala) rip-ens very differently than tomato and many otherfruits in not undergoing cell wall swelling as part ofthe cell wall modifications during ripening (Redg-well et al., 1997). There is minimal change in viscosityof cell walls, and minimal pectin solubilization ordegradation during fruit ripening. This implies thatany endo-PG isolated from ripening fruit of applemay have different characteristics to that isolatedfrom ripening tomato fruit. Therefore, results relat-
1 This work was supported by the Foundation for Research,Science, and Technology of New Zealand (grant nos. C06817 andC06X001).
* Corresponding author; e-mail [email protected];fax 0064 –9 – 815– 4201.
Article, publication date, and citation information can be foundat www.plantphysiol.org/cgi/doi/10.1104/pp.010986.
122 Plant Physiology, May 2002, Vol. 129, pp. 122–133, www.plantphysiol.org © 2002 American Society of Plant Biologists
Dow
nloaded from https://academ
ic.oup.com/plphys/article/129/1/122/6110110 by guest on 17 February 2022

ing to overexpression of tomato fruit endo-PG maynot be the same as overexpression of apple fruitendo-PG.
PG enzyme in ripe apple fruit has been isolatedand biochemically characterized as an endo-PG (Wuet al., 1993). The corresponding cDNA (MdPG1, for-merly GDPG1; Atkinson, 1994) was isolated fromapple cv Golden Delicious and shown to hybridize toan mRNA present in ripe fruit but not in developingfruit or flowers (Atkinson et al., 1998). MdPG1 en-coded a protein with 52% amino acid identity to thetomato fruit-specific clone pTOM6. Analysis of thepromoter of MdPG1 showed that 532- and 1,460-bpfragments conferred �-glucuronidase expression inripe tomato fruit, but not in flowers, leaves, or de-veloping fruit (Atkinson et al., 1998).
The MdPG1 cDNA was overexpressed in applewith the expectation that disruption of cell wall me-tabolism would occur in ripening fruit. However, wereport in this paper a range of novel phenotypes inother plant tissues associated with endo-PG overex-pression, providing new information on the involve-ment of pectin in cell-cell adhesion and stomatalfunction.
RESULTS
Apple Plants Overexpressing Fruit-Specific PG HaveIncreased PG mRNA, Protein, and Activity inMature Leaves
Three independent PG transformants (MdPGS-2,-3, and -4) were identified that exhibited a novelphenotype in tissue culture, characterized by palegreen leaves showing necrosis around the edges. TheMdPGS transformants were micropropagated toform transgenic lines. DNA gel-blot analysis revealedthat MdPGS-2 and -4 each contained a single inte-grated copy of the PG transgene, whereas MdPGS-3contained two copies (Fig. 1a).
RNA gel-blot analysis showed that the MdPGStransformants produced easily detectable amounts ofthe fruit-specific PG mRNA (Fig. 1b) in rapidly ex-panding leaves. Wild-type apple leaves of equivalentage did not express a homologous message. Antibod-ies raised against endo-PG from ripening tomatofruit reacted with a polypeptide at 46 kD in MdPGSleaves (Fig. 1c), indicating a correctly translated andprocessed protein from the PG mRNA. Very lowlevels of a cross-reacting polypeptide of the same sizewere observed in control leaves. The increased PGexpression and protein in the transformants was re-flected in the measured enzyme activity (Table I). Inmature leaves of MdPGS-2 and -4, PG activity wasalmost doubled, and in MdPGS-3, activity increasedby approximately 35% above wild-type levels. Inwild-type leaves, low enzyme activity was detected(Table I), probably reflecting the very faint immuno-reactive band (Fig. 1c).
PG Overexpression Leads to Changes in Leaf Color,Water Relations, and Leaf Abscission
Leaves of PG-overexpressing plants showed a dis-tinct change in leaf color phenotype during matura-tion compared with wild-type controls. MdPGS-2and -4 had mature leaves in which the silver colorextended throughout the leaf blade (Fig. 2a), whereasin MdPGS-3, the silver color was centered on the leafveins but did not extend fully into the leaf blade. The
Figure 1. Analysis of MdPGS transformant and wild-type (WT) appleplants by DNA gel blot, RNA gel blot, and western blot. a, DNAgel-blot analysis was performed using genomic DNA (10 �g) digestedwith BamHI and size fractionated on 0.8% (w/v) agarose gels. Mem-branes were probed at the left border with an NptII-specific probe(a1) and at the right border with a cauliflower mosaic virus 35S-specific probe (a2). Membranes were washed at a stringency allow-ing 15% mismatch (0.5� SSC and 1% [w/v] SDS, 65°C). Size markersare in kb. b, RNA gel-blot analysis was performed using RNA (5 �g)probed with a GDPG1-specific probe (GDPG1 nucleotides 1–603).Membranes were washed at a final stringency of 0.1� SSC � 0.1%(w/v) SDS and exposed for 16 h. Signals were analyzed using aphospho-imaging system and ImageQuant software analysis and cor-rected for any small differences in RNA loading by comparison withribosomal RNA hybridization. c, For western-blot analysis, PG en-zyme was extracted as described in “Materials and Methods” andseparated on denaturing 10% (w/v) polyacrylamide gels (loadingapproximately 11 �g protein per lane). Blots were incubated withantibodies raised against endo-PG from ripening tomato fruit todetect cross-reacting proteins. Endo-PG purified from ripening to-mato fruit (DellaPenna et al., 1986) was used as a positive control.Arrows indicate position of Mr markers.
Overexpression of Polygalacturonase in Apple
Plant Physiol. Vol. 129, 2002 123
Dow
nloaded from https://academ
ic.oup.com/plphys/article/129/1/122/6110110 by guest on 17 February 2022

difference in leaf color could be quantified using theL value on a colorimeter (Table II). In all MdPGStransformants, the L value was higher than for wild-type controls. Young leaves did not show a quantifi-able change in phenotype compared with controls(data not shown). Microscopic examination of leavesfrom the MdPGS transformants indicated that thesilvery areas of leaves had more airspaces betweenthe epidermis and the palisade cells of the mesophyll,indicating poorer adhesion of cells (Fig. 2c). Stainingwith ruthenium red showed less pectin on the epi-dermal cell walls that face palisade cells in all trans-formants (Fig. 2c). Young transgenic leaves did notshow this phenotype.
Because leaves of MdPGS plants appeared morebrittle and wilted and abscised more easily, photo-synthetic rate and leaf diffusive conductance werecompared in silvery leaves of MdPGS-3 and leaves ofa control plant. Although photosynthetic rates werecomparable (13.5 � 1.4 versus 13.2 � 0.8 �mol m�2
s�1 for MdPGS-3 and control, respectively), leaf-diffusive conductance was approximately double inthe silvery leaves (0.17 � 0.01 versus 0.08 � 0.01 molm�2 s�1 for MdPGS-3 and control, respectively). Inaddition, pulse-amplitude-modulated fluorescencemeasurements showed that calculated maximumelectron transport rate (ETR) and quantum yieldwere similar in silvery and normal leaves (data notshown).
The force required for removal of leaves fromMdPGS-2 and -4 plants was significantly less (P �0.05) than that of wild type (Table II), whereas that ofMdPGS-3 was intermediate. The measurement wasconsistent for mature leaves throughout the growingseason, with a tendency for the phenotype to increaseas the plants reached senescence (data not shown).Because PG only acts on nonesterified homogalactu-ronan regions of pectin, putative abscission zones ofwild-type and transformant petioles were labeledwith antibodies JIM5 (low methyl-esterified pectin,degree of esterification [DE] � 35%) and JIM7 (highmethyl-esterified pectin; Knox et al., 1990). There wasmore low-esterified pectin in abscission zones oftransformants than in wild type, and it was visiblethroughout most of the cell walls in this region (Fig.2b). In wild type, only cell junctions and middlelamellae were labeled. Although mature enough,none of the MdPGS transformants developed flowersand, as a consequence, did not set any fruit.
PG Overexpression Alters Stomatal Functioning andCreates Lesions and Disordered Internal Structure inMature Apple Leaves
When wild-type and transformant leaves werefreshly harvested and placed in the dark for 30 min,only the stomata on mature wild-type leaves closed(Fig. 3a). A similar result was found with abscisicacid (ABA) treatment (data not shown). Closer exam-ination of the stomata revealed a disruption in cellmorphology with holes at one or both ends of thestomatal aperture in transformants (Fig. 3, b and c).The holes varied in size but invariably penetratedthrough the epidermis, and guard cells and neigh-boring epidermal cells were often separated (Fig. 2, dand e). In young transformant leaves, only a fewstomata had holes (Fig. 3d). The pattern of stomataldevelopment looked similar to that in wild-typeleaves, although in some cases surface irregularitiescould be seen adjacent to guard cells.
The lower epidermis of MdPGS leaves was fragileand collapsed frequently before, or during, fixation.When stained with ruthenium red to visualize pectin,staining was reduced compared with wild type (Fig.2d) and labeling with JIM5 antibodies indicated areduction in low-esterified pectin (Fig. 2f). Spongymesophyll cells and cells around the vasculature inMdPGS leaves were more enlarged and unevenlyshaped than those in wild type (data not shown).Mature MdPGS leaves labeled with JIM7 antibodiesin a similar manner to wild type (data not shown).There was little difference in labeling of palisade cellsbetween wild type and MdPGS leaves (data notshown).
Guard cells of both wild-type and transformantstomata showed labeling of highly esterified pectin(JIM7) across the cell wall in a similar manner (Fig.2h), although stomata were frequently malformed inthe transformants. Low-esterified pectin (JIM5) inwild type was also distributed across the whole wall,with areas of increased intensity at the pore edge andnear the adjacent epidermal cell (Fig. 2g). In thetransformants, JIM5 labeling was strong close to theplasma membrane edge of the guard cell wall andweak to nonexistent near the outer edge.
Overexpression of a Fruit-Specific PG Alters Cell WallChemistry in Mature Leaves
Overexpression of fruit-specific PG led to signifi-cant changes in the chemistry of leaf cell walls of theMdPGS transformants. In most cell wall chemicalanalyses, results for MdPGS-3 tended to be the sameor similar to wild-type values, whereas MdPGS-2 and-4 were significantly different than wild type. This isconsistent with the relative endo-PG enzyme activi-ties and mRNA expression levels in thesetransformants.
On a fresh weight basis, MdPGS leaves had 15% to22% less cell wall material (CWM) than wild-type
Table I. PG activity in mature leaves of wild-type apple andMdPGS transformants
GenotypePG Activity
Extraction I Extraction II Average
mmol reducing groups � 100 mg�1 CWM
Wild type 13.9 15.4 14.7MdPGS-2 24.9 27.9 26.4MdPGS-3 20.8 18.7 19.8MdPGS-4 23.1 26.9 25.0
Atkinson et al.
124 Plant Physiol. Vol. 129, 2002
Dow
nloaded from https://academ
ic.oup.com/plphys/article/129/1/122/6110110 by guest on 17 February 2022

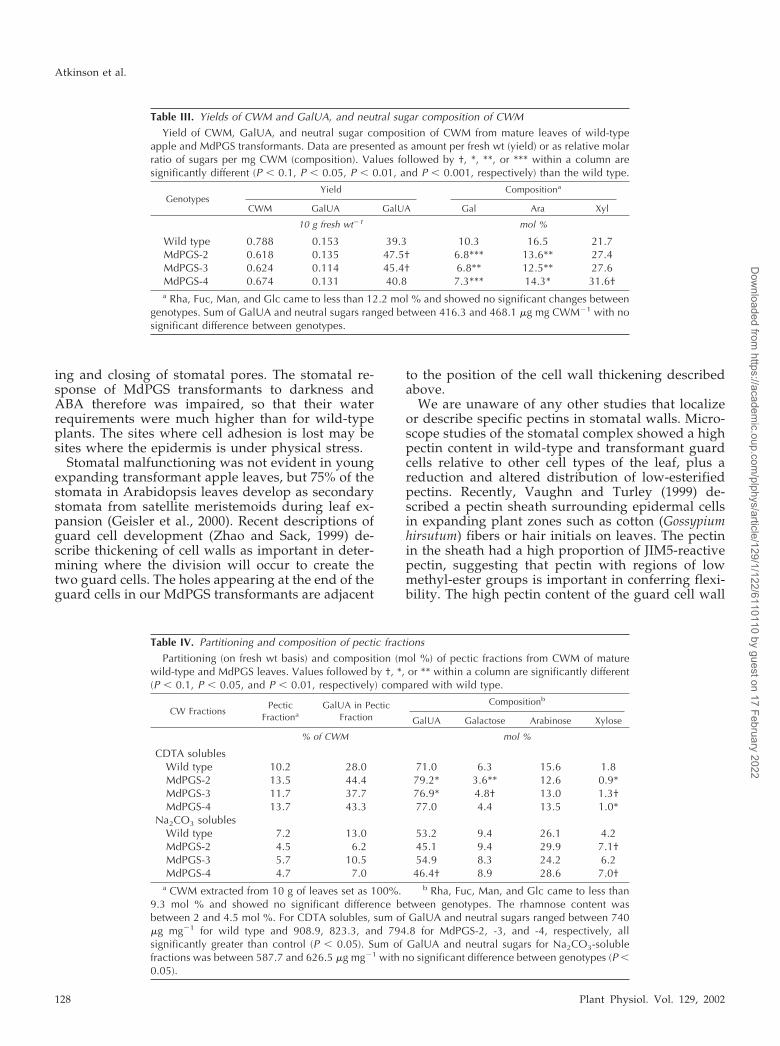
leaves (Table III), and GalUA, the major componentof pectin, was reduced by 12% to 26%. However, themolar composition showed an increased proportionof GalUA compared with wild type. Gal and Aracontents of the CWM were markedly reduced in thetransformants, but the Xyl content was higher than inwild-type leaves.
To determine which type of pectin had been influ-enced by overexpression of PG, CWM was extractedsequentially with trans-1,2-diaminocyclohexane-N,N,N�,N�-tetra-acetic acid (CDTA) and Na2CO3.CDTA is a chelator that extracts pectins held in thewall by ionic bonds, whereas the alkaline reagentNa2CO3 extracts covalently linked pectin. There wasa difference in partitioning of polymers and in theircomposition between the subfractions of the CWM oftransformants compared with wild type (Table IV).Transformants had a higher yield of CDTA solublesthat contained more GalUA on a fresh weight basisthan wild type. In comparison, there was a reduction
in yield of the Na2CO3 solubles in MdPGS leaves,which also contained less GalUA. Compositionalanalyses of the two pectic fractions showed a veryhigh abundance of GalUA and the neutral monosac-charides Gal, Ara, and Xyl, which are commonlyfound as side chains of apple-derived pectins (Scholset al., 1995). This and the very low presence of Glc,Man, and Fuc, sugars that are commonly found innon-pectic polysaccharides, indicate that the CDTAand Na2CO3 fractions consist mostly of pectins.CDTA-soluble pectins of transformants had a highermolar ratio of GalUA than the wild type. For Na2CO3solubles, the molar ratio was lower for MdPGS-2 and4, but not for MdPGS-3. The Gal, Ara, and Xyl con-tent of the CDTA-soluble pectins was reduced intransformants, whereas for Na2CO3-soluble pectins,Ara and Gal content was unchanged. Xyl was ele-vated, particularly in MdPGS-2 and -4. These resultsshow that the elevated GalUA levels and reducedlevels of Gal and Ara seen in CWM of transformants
Figure 2. Phenotype of leaves from MDPGStransformants compared with wild type (WT). a,Silvery leaf phenotype. Scale bar � 10 mm. b,Putative petiole abscission zone showing distri-bution of low-esterified pectin by labeling withJIM5 antibodies. c, Cross section of upper epi-dermis and palisade cells of leaves demonstrat-ing reduced cell adhesion in the silvery leafphenotype (stained with ruthenium red for pec-tin). d, Cross section of entire leaves, demon-strating the reduced ruthenium red staining ofthe lower epidermis and holes next to guardcells or stomata. e, Cross section of the lowerleaf epidermis showing the hole next to theguard cell in the transformant (stained with to-luidine blue). f, Cross section of leaves labeledwith JIM5 antibodies, showing reduced labelingof lower epidermal cells in the transformant. g,Cross section of the lower epidermis of leaveslabeled with JIM5 antibodies showing altereddistribution of label in guard cells of transfor-mant stomata. h, Cross section of the lowerepidermis of leaves labeled with JIM7 antibodiesshowing labeling in the guard cells and neigh-boring cells. Scale bars in b through h � 10 �m.
Overexpression of Polygalacturonase in Apple
Plant Physiol. Vol. 129, 2002 125
Dow
nloaded from https://academ
ic.oup.com/plphys/article/129/1/122/6110110 by guest on 17 February 2022

could be attributed to ionically bound pectins,whereas elevated Xyl levels in CWM of transfor-mants were found to be primarily in the cell wallresidue after extraction (data not shown).
Strength of cell-to-cell adhesion is to a large partdetermined by length and DE of pectin molecules ofthe middle lamella (Jarvis, 1984). Because endo-PGshydrolyze pectins to smaller chains, the size ofCDTA-soluble pectins of all genotypes was com-pared (Fig. 4). In all MdPGS transformants, these hada lower average molecular mass than the correspond-ing wild-type pectins, with the elution peak of wild-type CDTA pectin at approximately 350 kD, com-pared with 200 to 250 kD for MdPGS plants. The sizedifference was more pronounced in MdPGS-2 and -4than in MdPGS-3. Because labeling with JIM5 anti-bodies differed markedly between wild-type leavesand the transformants, we compared the DE ofCDTA-soluble pectins from each plant. MdPGS-2pectin was significantly (P � 0.05) less esterified(16%) than the wild-type (33.5%), whereas MdPGS-3and -4 were intermediate (23.4% and 25.3%). There-fore, CDTA-soluble pectins of all genotypes would berecognized by JIM 5. The DE of other cell wall frac-tions cannot be analyzed because they are de-esterified during alkaline extraction.
In summary, overexpression of fruit-specific PGhas caused a change in partitioning of pectic poly-mers and in their structure, including composition,length, and DE. Cell wall extractions of young ex-panding leaves showed the same tendencies as for
mature leaves but differences were less marked (datanot shown).
DISCUSSION
Constitutive overexpression of a fruit-specific PGin apple plants has led to a range of novel pheno-types including disrupted leaf organization, per-turbed water relations, malformed and malfunction-ing stomata, silvery leaves, and changes in leafabscission. The modification of apple trees by thissingle fruit gene, therefore, has offered a new andunexpected perspective on a number of physiologicaland developmental processes.
The Role of Pectin in Development and Cell Adhesion
Overexpression of PG in apple led to abnormal cellseparation within the leaf. Loss of cell adhesion wascommon to a number of phenotypes. It occurredbetween upper epidermis and palisade parenchyma,resulting in the silvery color of leaves, between cellsof the lower epidermis resulting in stomatal dysfunc-tion, and in the abscission zone of the petioles, re-sulting in more easily detachable leaves. These phe-notypes are consistent with the predicted action ofPG. Plant cells are held together in the middle la-mella region by pectin forming a gel via Ca2� cross-links (Jarvis, 1984; Knox et al., 1990; Knox, 1992). Thestrength of adhesion of this gel is largely determinedby the length of pectin molecules and their DE. InMdPGS transformants, the CDTA-soluble pectinsheld in the wall by Ca2� cross-links had shorterchains compared with wild type. This results in lessefficient binding between polymers, a weaker gel,and thus poorer cell adhesion, even though the lowerDE of the CDTA fraction favors Ca2� cross-linking.The altered monosaccharide content of the sidechains of the CDTA pectins may also have an impacton cell adhesion.
Analyses of mutants in several plant species haveshown alterations to pectin positioning and structurethat affect cell-to-cell adhesion and normal tissuedevelopment. In the Arabidopsis EMB30 mutant,pectins (especially low-esterified ones) appear to bemisdirected so they accumulate in intercellularspaces. Cells of leaves and callus adhere poorly, andare irregular and larger than wild type (Shevell et al.,2000). Unlike the MdPGS transformants, however,the EMB30 mutant has unaltered neutral sugar com-position in the cell wall.
In the Cnr tomato mutant, pectin structure and cell-to-cell adhesion in fruit is altered through a disruptionin the deposition of (135)-�-l-arabinan that forms thebranched sidechains of rhamnogalacturonan-I (Orfilaet al., 2001). As a consequence, ripe-stage Cnr fruitshave stronger cell walls throughout the pericarp thatdo not swell, and there is extensive intercellular spacein the inner pericarp in comparison with wild type.Orfila et al. (2001) showed that maturation processes
Table II. Characterization of silver leaf phenotype and abscissionforce
The color of mature apple leaves from wild-type and MdPGStransformants was measured using a chromameter. Data (given as Lvalues, where higher values indicate lighter colors) are means de-rived from 15 leaves per plant. The force (N) required to remove amature apple leaf from its stem in wild-type and MdPGS transfor-mants was measured for six to seven leaves per plant using a Mate-rials Testing Machine. Values followed by one or two asterisks aresignificantly different (P � 0.05 and P � 0.01, respectively) than wildtype.
Genotype Plant L Value Force (N)
Wild type 1 37.35 2.762 38.45 2.293 38.81 1.67
Mean 38.13 2.24MdPGS-2 1 41.47 0.74
2 43.15 0.803 42.41 0.554 43.30 0.56
Mean 42.52* 0.66**MdPGS-3 1 41.34 2.20
2 41.03 2.023 41.49 1.67
Mean 41.24 1.96MdPGS-4 1 43.82 0.67
2 43.98 0.93Mean 43.87* 0.82**
Atkinson et al.
126 Plant Physiol. Vol. 129, 2002
Dow
nloaded from https://academ
ic.oup.com/plphys/article/129/1/122/6110110 by guest on 17 February 2022

involving middle lamella pectins are altered in Cnrfruit, resulting in the absence or a low level of pectin-/calcium-based cell adhesion.
The Role of PG and Pectin in Stomatal Function
Our study of the stomata in transgenic MdPGSapple plants has confirmed the important role of the
cell wall in stomatal functioning (Ziegenspeck, 1938).As a consequence of PG overexpression, guard cellsand neighboring epidermal cells are often separatedfrom the lower epidermis of mature transformantleaves. Moreover, holes appeared at each end of theguard cells during leaf growth. These changes re-duced the ability of the cells of the stomatal complexto act against each other, which is necessary for open-
Figure 3. Stomata from MdPGS transformant leaves compared with wild type (WT). a, Light microscope view of stomata 30min after placement in the dark. b and c, Holes associated with stomata in mature transgenic leaves compared withwild-type. d, Stomata in young, developing leaves of transgenic and wild-type plants. Scale bars in a, b, and d � 100 �m;scale bar in c � 10 �m.
Overexpression of Polygalacturonase in Apple
Plant Physiol. Vol. 129, 2002 127
Dow
nloaded from https://academ
ic.oup.com/plphys/article/129/1/122/6110110 by guest on 17 February 2022
ing and closing of stomatal pores. The stomatal re-sponse of MdPGS transformants to darkness andABA therefore was impaired, so that their waterrequirements were much higher than for wild-typeplants. The sites where cell adhesion is lost may besites where the epidermis is under physical stress.
Stomatal malfunctioning was not evident in youngexpanding transformant apple leaves, but 75% of thestomata in Arabidopsis leaves develop as secondarystomata from satellite meristemoids during leaf ex-pansion (Geisler et al., 2000). Recent descriptions ofguard cell development (Zhao and Sack, 1999) de-scribe thickening of cell walls as important in deter-mining where the division will occur to create thetwo guard cells. The holes appearing at the end of theguard cells in our MdPGS transformants are adjacent
to the position of the cell wall thickening describedabove.
We are unaware of any other studies that localizeor describe specific pectins in stomatal walls. Micro-scope studies of the stomatal complex showed a highpectin content in wild-type and transformant guardcells relative to other cell types of the leaf, plus areduction and altered distribution of low-esterifiedpectins. Recently, Vaughn and Turley (1999) de-scribed a pectin sheath surrounding epidermal cellsin expanding plant zones such as cotton (Gossypiumhirsutum) fibers or hair initials on leaves. The pectinin the sheath had a high proportion of JIM5-reactivepectin, suggesting that pectin with regions of lowmethyl-ester groups is important in conferring flexi-bility. The high pectin content of the guard cell wall
Table IV. Partitioning and composition of pectic fractions
Partitioning (on fresh wt basis) and composition (mol %) of pectic fractions from CWM of maturewild-type and MdPGS leaves. Values followed by †, *, or ** within a column are significantly different(P � 0.1, P � 0.05, and P � 0.01, respectively) compared with wild type.
CW FractionsPectic
FractionaGalUA in Pectic
Fraction
Compositionb
GalUA Galactose Arabinose Xylose
% of CWM mol %
CDTA solublesWild type 10.2 28.0 71.0 6.3 15.6 1.8MdPGS-2 13.5 44.4 79.2* 3.6** 12.6 0.9*MdPGS-3 11.7 37.7 76.9* 4.8† 13.0 1.3†MdPGS-4 13.7 43.3 77.0 4.4 13.5 1.0*
Na2CO3 solublesWild type 7.2 13.0 53.2 9.4 26.1 4.2MdPGS-2 4.5 6.2 45.1 9.4 29.9 7.1†MdPGS-3 5.7 10.5 54.9 8.3 24.2 6.2MdPGS-4 4.7 7.0 46.4† 8.9 28.6 7.0†
a CWM extracted from 10 g of leaves set as 100%. b Rha, Fuc, Man, and Glc came to less than9.3 mol % and showed no significant difference between genotypes. The rhamnose content wasbetween 2 and 4.5 mol %. For CDTA solubles, sum of GalUA and neutral sugars ranged between 740�g mg�1 for wild type and 908.9, 823.3, and 794.8 for MdPGS-2, -3, and -4, respectively, allsignificantly greater than control (P � 0.05). Sum of GalUA and neutral sugars for Na2CO3-solublefractions was between 587.7 and 626.5 �g mg�1 with no significant difference between genotypes (P �0.05).
Table III. Yields of CWM and GalUA, and neutral sugar composition of CWM
Yield of CWM, GalUA, and neutral sugar composition of CWM from mature leaves of wild-typeapple and MdPGS transformants. Data are presented as amount per fresh wt (yield) or as relative molarratio of sugars per mg CWM (composition). Values followed by †, *, **, or *** within a column aresignificantly different (P � 0.1, P � 0.05, P � 0.01, and P � 0.001, respectively) than the wild type.
GenotypesYield Compositiona
CWM GalUA GalUA Gal Ara Xyl
10 g fresh wt�1 mol %
Wild type 0.788 0.153 39.3 10.3 16.5 21.7MdPGS-2 0.618 0.135 47.5† 6.8*** 13.6** 27.4MdPGS-3 0.624 0.114 45.4† 6.8** 12.5** 27.6MdPGS-4 0.674 0.131 40.8 7.3*** 14.3* 31.6†a Rha, Fuc, Man, and Glc came to less than 12.2 mol % and showed no significant changes between
genotypes. Sum of GalUA and neutral sugars ranged between 416.3 and 468.1 �g mg CWM�1 with nosignificant difference between genotypes.
Atkinson et al.
128 Plant Physiol. Vol. 129, 2002
Dow
nloaded from https://academ
ic.oup.com/plphys/article/129/1/122/6110110 by guest on 17 February 2022

has also been suggested to act as a temporary storageof K� ions during stomatal movements (Bowling andSmith, 1990).
The leaves of MdPGS transformants have a brittlephenotype, indicative of dehydration. Malfunction-ing stomata would accentuate this phenotype. Mur1Arabidopsis mutants lacking Fuc in the cell wallshow increased brittleness in elongating inflores-cence stems (Reiter et al., 1993) and potato (Solanumtuberosum) plants with reduced Man in cell wall poly-mers have a phenotype of leaf dehydration followingthe development of black spots on the surface (Kelleret al., 1999). The PG transformants, however, did notshow any differences in these sugars compared withwild type.
PG Overexpression and the Silvery Leaf Phenotype
The silvery leaf phenotype of MdPGS transformantleaves resembles the “silverleaf” phenotype that oc-curs in apple in response to infection by the systemicfungus Chondrostereum purpureum. The silverleafsymptoms develop in the leaves as they mature andare believed to be due to airspaces caused by partialseparation of the epidermal cells from the palisadecells (Tetley, 1932; Spiers et al., 1987) as was seen inthe MdPGS transformants. Endo-PG produced by thefungus is responsible for the development of thepathogenic symptoms (Miyairi et al., 1977). A com-parison of infected apple leaves with healthy leavesshowed altered pectin partitioning, a high content ofmethyl-esterified pectin, and some low-Mr pectin oc-curring in infected leaves (Miyairi et al., 1997). In thesilvery leaves of MdPGS transformants, we alsofound altered pectin structure and partitioning.
In MdPGS transformants, young leaves were notsilvery, did not detach easily, and stomata were stillfunctioning. Changes in cell wall chemistry were al-ready present but not as marked as in mature leaves.These results suggest that endo-PG can interrupt cellwall metabolism only at a particular stage in leafdevelopment. It is possible that PG has no substrateavailable in developing leaves. Pectin is synthesizedand incorporated into the cell wall in a highly me-thoxylated form (Zhang and Staehelin, 1992), andonly during development are the methyl-groups re-moved by pectin-methyl-esterase, thereby creating asubstrate for PG (Kim and Carpita, 1992). PG is alsoimmobilized in specific sections in the cell wall(Steele et al., 1997). If the cell wall structure loosensduring maturation, PG may then be able to come incontact with potential substrates. Pectins have beenreported as mediators of wall porosity in soybeans(Glycine max), and a mild treatment with PG appar-ently enlarged the channels (Baron-Epel et al., 1988).Another possibility is that a continuous modificationof pectin structure in the transformant leaves (be-cause of the use of a constitutive promoter) results inaltered cell-cell association that is “revealed” only ascell expansion continues. Interestingly, fungalendo-PG applied to mature apple leaves has no effect(silvering does not occur and the polysaccharidesremain large), whereas if young expanding leaves aretreated with the fungal enzyme, symptoms developand the polysaccharides are degraded (Miyairi et al.,1997).
The Role of PG in Leaf Abscission
Our results also provide the first direct evidencethat expression of a single nonabscission-related PGgene can have a significant role in cell-to-cell adher-ence in the abscission region, leading to a loss ofstructural strength.
Endo-PGs have previously been identified andcharacterized in the abscission zone of several plantspecies including tomato (Taylor et al., 1990;Kalaitzis et al., 1997; Hong et al., 2000) and Brassicanapus (Petersen et al., 1996). Taylor et al. (1993) cor-related increases in PG activity only with the onset ofabscission. Experiments with transgenics contributedto the theory that PG action was not one of the primedeterminants of abscission in leaves. Antisensedown-regulation of the tomato fruit-specific PG genepTOM6 in tomato did not appear to affect leaf ab-scission (Taylor et al., 1990). Constitutive overexpres-sion of the same gene in tobacco did not result in anincrease in leaf abscission although PG activity waselevated (Osteryoung et al., 1990), suggesting thatsubstrate-enzyme interactions were not appropriatefor fruit-specific PGs to act in the leaf abscissionzone.
In wild-type apple, in regions where the abscissionzone develops, low-esterified pectin was only
Figure 4. Molecular distribution of CDTA-soluble pectin. Mr distri-bution was analyzed by gel-permeation chromatography on a Sepha-rose CL-2B column (65 � 2 cm; eluent 0.05 M Na-acetate, pH 6.0;0.125 M NaCl; 0.05% [w/v] chlorobutanol; fraction size 20 min; flowrate 8.4 mL h�1). The column was calibrated using dextran molecularstandards T2000, T500, and T10 (Pharmacia Biotech, Piscataway,NJ), and the elution profile monitored using the GalUA assay (Ahmedand Labavitch, 1977). Na2CO3-soluble pectins of all genotypes werenot soluble after freeze drying and therefore could not be subjectedto gel permeation chromatography.
Overexpression of Polygalacturonase in Apple
Plant Physiol. Vol. 129, 2002 129
Dow
nloaded from https://academ
ic.oup.com/plphys/article/129/1/122/6110110 by guest on 17 February 2022

present in cell junctions and middle lamellae. In theMdPGS transformants, levels of low-esterified pec-tins were increased and distributed almost across thewhole cell wall. This should improve cell adhesion(Jarvis, 1984). However, if modifications to the pec-tins of these cell walls occurred in a similar manner tothose in the leaves, then the reduction in cell adher-ence during detachment could be due to the decreasein size of the CDTA-soluble pectins, which are moreabundant in the transformants. With a DE � 35% inall genotypes, they would still be recognized by JIM5.Earlier examinations of the abscission zone in variousorgans have shown that during the separation phase,pectins are solubilized and the middle lamella swells(Sexton and Roberts, 1982). This fits well with ourmicroscopic studies with localization of the low-esterified pectin in the abscission zone of the petiole.
CONCLUSIONS
Constitutive overexpression of a fruit-specific PGin apple plants led to profound effects on leaf mor-phology, plant water relations, stomatal structureand function, and leaf attachment. This has not beenthe case in other transgenic plant studies. In tobacco,constitutive expression of a tomato fruit PG had littleor no effect on tobacco plant morphology or devel-opment, although the enzyme was appropriatelyprocessed and showed activity in vitro (Osteryounget al., 1990). Is the difference between tomato fruit PGand apple fruit PG one of mobility in the cell wall, ofsubstrate availability, or of substrate preference? Werecently have transformed Arabidopsis with thesame apple gene and find a similar set of phenotypicchanges in Arabidopsis leaves as observed in trans-genic apple trees. Hence, it may be that apple fruitPG is less specific in the choice of its substrates thantomato fruit PG. Our results in apple suggest that itwould be useful to carry out a fuller examination ofthe action of specific PGs on pectins, and the changesin pectins during leaf development.
MATERIALS AND METHODS
Plant Transformation and Growth
The apple (Malus domestica Borkh. cv Royal Gala) PGcDNA clone MdPG1, formerly GDPG1 (Atkinson, 1994),was used as the starting point for vector construction.Initially, the NotI site present in pSportI was removed byblunting, then the PG cDNA insert was excised with BamHIand SmaI, blunted, and cloned into the SmaI-site of pART7(Gleave, 1992). This construct was digested with NotI andcloned into the binary pART27. The binary was electropo-rated into Agrobacterium tumefaciens strain LBA4404.
Transgenic apple shoots were produced using themethod of Yao et al. (1995). Independent transgenic shootswere micropropagated from axillary buds to form trans-genic lines. Transgenic lines were maintained in a contain-ment greenhouse under identical conditions (ambient light
and temperature) to wild-type plants, except MdPGS trans-formants were given supplementary watering. Plants were3 years old at the time of experimentation.
DNA and RNA Gel-Blot Analyses
Because previous transformations of apple cv “RoyalGala” with the empty binary vector pART27 (Yao et al.,1995) showed that all transformants were similar to wildtype, we used wild-type plants as controls for these and allfurther analyses.
Apple genomic DNA was extracted from young, ex-panding leaves as described in Dellaporta et al. (1983).DNA gel-blot analysis, apple leaf RNA extractions, andRNA gel-blot analysis were performed as described bySchroder et al. (1998).
PG Extraction, Activity Assays, and Western Blotting
PG enzyme was extracted using the method of Bergey etal. (1999). Aliquots of enzyme (600 �L) were incubatedwith 1% (w/v) poly-GalUA, pH 4.5 (100 �L) and 0.3 mNa-acetate, pH 4.5 (150 �L). Duplicate samples boiled inwater for 5 min before addition of the substrate served asblanks (assay modified after Pressey, 1986). The mixtureswere incubated overnight at 30°C, and PG activity ana-lyzed by measuring reducing end groups using4-hydroxybenzoic acid hydrazide with GalUA as standard(Lever, 1972). After subtraction of the corresponding blank,PG activity was expressed in mmol reducing end groupsgenerated per 100 mg CWM. Protein concentration wasestimated using the protein assay (Bio-Rad Laboratories,Hercules, CA).
For immunoblotting, proteins were separated on 1.5 mm10% (w/v) SDS-Tris-Tricine gels (Schagger and van Jagow,1987) using a Mini-Protean II electrophoresis system (Bio-Rad). Proteins were electroblotted onto Immobilon-P trans-fer membranes (Millipore, Bedford, MA) using a Trans-BlotSD Semi-Dry transfer cell (Bio-Rad) following the manu-facturer’s instructions (Millipore). Blots were blocked over-night at 4°C, with TBS-T20 (10 mm Tris base, 150 mm NaCl,and 0.1% [v/v] Tween 20) containing 5% (w/v) nonfatdried milk powder and incubated with tomato (Lycopersi-con esculentum) endo-PG antibody (1:1,000 [w/v] dilution)in blocking solution for 1 h at ambient temperature. Afterwashing in TBS-T20, the blots were incubated with alkalinephosphatase-conjugated goat anti-rabbit IgE (1:1,000[w/v], Sigma, St. Louis) in blocking solution. After washingin TBS-T20 (3 � 15 min), the bound antibodies were visual-ized using 1-Step nitroblue tetrazolium/5-bromo-4-chloro-3-indolyl phosphate (Pierce Chemical, Rockford, IL).
Measurement of Force for Leaf Detachment
Petioles of mature, fully expanded leaves were wrappedthree times with string using slipknots. The string wasfirmly attached with glue to the petiole, leaving at least 1cm to the leaf abscission zone. The other end of the stringwas attached to a hook attachment on a Materials Testing
Atkinson et al.
130 Plant Physiol. Vol. 129, 2002
Dow
nloaded from https://academ
ic.oup.com/plphys/article/129/1/122/6110110 by guest on 17 February 2022

Machine (model MN 44, Instron, Canton, MA) with a 100 nload cell. The petiole was positioned at right angles to thedirection in which the breakforce was applied. Tensileforce was applied at a constant 10 mm min�1. The maxi-mum force required for leaf removal was recorded for atleast five leaves from three plants of each transgenic lineand the wild-type control. The experiment was carried outin spring, summer, and early autumn.
Color Readings and Photosynthesis, Leaf-DiffusiveConductance, ETR, and Photon Efficiency
The silvery leaf phenotype was measured on matureleaves on the upper surface using a Chromameter (Minolta,Osaka) and the CIELAB model for hue (h) and lightness (L)(ASTM, 1966) and the standard illuminant D65 (emits theequivalent of average daylight including UV region withcorrelated color temperature of 6504K). C (chromaticity), L,and h values were measured on seven leaves of each tree ofeach transgenic line.
All leaf gas-exchange measurements were made with aclimate-controlled minicuvette system (CMS 400, Walz, Ef-feltrich, Germany) as described by Buwalda et al. (1991).The dewpoint temperature of air entering the cuvette wasset at 5.2°C and the air temperature in the cuvette was setat 24°C. The reported photosynthetic rate and leaf diffusiveconductance were the values recorded at a photosyntheti-cally active radiation of 900 �mol m�2 s�1. Maximum ETRand photon efficiency of electron transport were measuredusing a MINIPAM (Walz, Effeltrich, Germany) accordingto the method of Laing et al. (2000).
Microscopy
Light microscopy observations were carried out on aVanox AHT 3 microscope (Olympus, Tokyo) fitted with a35-mm film camera and a CoolSnap digital camera (RoperScientific, Tucson, AZ). Lamina tissue from young andmature leaves and petiole tissue from mature leaves wereexcised and fixed in either 2% (v/v) formaldehyde with0.1% (v/v) glutaraldehyde in 0.1 m phosphate buffer, or 2%(v/v) formaldehyde with 2.5% (v/v) glutaraldehyde in 0.1m phosphate buffer. Tissue was subjected to light vacuumto infiltrate airspaces, washed in buffer, dehydrated withan alcohol series, and embedded in LR White resin (Hallettet al., 1992). The embedded material was sectioned at athickness of 0.5 or 1 �m and stained with 0.05% (w/v)toluidine blue in borate buffer (pH 4.4) for gross morpho-logical examination and 0.02% (w/v) ruthenium red forgeneral localization of pectic material. Antibodies JIM5 andJIM7 were used to localize low-esterified pectin and high-esterified pectin, respectively (Sutherland et al., 1999). Di-rect observation of intact unfixed leaves of wild-type andtransformed leaves was carried out after exposure to light,up to 45 min in the dark and exposure for 30 min to 3 �mABA brushed over the epidermis.
For low-temperature scanning electron microscopy, leaftissue material was mounted on copper specimen stubsand rapidly frozen in liquid nitrogen and either processed
and observed immediately or stored under liquid nitrogen.Material was processed and transferred for observation ona cryostage in a Philips PSEM 505 scanning electron micro-scope (Philips, Eindhoven, The Netherlands) using anEMScope SP2000 sputter cryo-system (Falloon et al., 1989).
Cell Wall Analyses
Tissue from mature leaves (10 g, midribs excised) wasground in liquid N2 and homogenized by polytroning in 40mL of methanol:chloroform:water:formic acid (16:5:1:1[v/v]). Homogenates were left at �20°C overnight andcentrifuged (6,000g, 10 min). Pellets were resuspended bypolytroning using 50 mL of methanol:chloroform:water(16:5:1 [v/v]) and centrifuged. Extraction of the twice morewashed pellet with dimethyl sulfoxide to give CWM, andsequential extraction of CWM with CDTA and Na2CO3 togive CDTA solubles, Na2CO3 solubles, and residue wascarried out after Redgwell et al. (1988). Fractions weredialyzed extensively against water for 7 d using dialysismembranes with a molecular mass cutoff of 3.5 kD (Spec-tra/Por, Spectrum) to minimize loss of pectic fragmentspossibly created by the action of PG, and then freeze dried.The composition of neutral monosaccharides was analyzedby gas liquid chromatography according to Albersheim etal. (1967). GalUA content was quantified (Blumenkrantzand Asboe-Hansen, 1973; Ahmed and Labavitch, 1977) us-ing d-GalUA as a standard. The DE was measured afterWood and Siddiqui (1971).
ACKNOWLEDGMENTS
We thank Jialong Yao for carrying out the apple trans-formation and observing some of the unusual phenotypes.We would also like to thank a number of our colleagues(Roger Harker, Teresa Wegrzyn, Jun Hong Ma, Ping HuaHe, Roy van der Brink, Julie Nicholls, Bret Morris, BiljanaPetrovich-Stojanovska, John Meekings, Tony Greaves, Wil-liam Laing, and Adrian Spiers) for helpful discussions ortechnical support. The JIM antibodies were a gift from Dr.Paul Knox, and the tomato PG antibody from the late Dr.Colin Brady.
Received October 30, 2001; returned for revision December19, 2001; accepted February 8, 2002.
LITERATURE CITED
Ahmed AER, Labavitch JM (1977) A simplified method foraccurate determination of cell wall uronide content. JFood Biochem 1: 361–365
Albersheim P, Nevins DJ, English PD, Karr A (1967) Amethod for the analysis of sugars in plant cell-wall poly-saccharides by gas-liquid chromatography. CarbohydrRes 5: 340–345
ASTM (1996) ASTM Standards on Color and AppearanceMeasurement, Ed 5. ASTM, West Conshohocken, PA
Atkinson RG (1994) A cDNA clone for endopolygalactu-ronase from apple. Plant Physiol 105: 1437–1438
Overexpression of Polygalacturonase in Apple
Plant Physiol. Vol. 129, 2002 131
Dow
nloaded from https://academ
ic.oup.com/plphys/article/129/1/122/6110110 by guest on 17 February 2022

Atkinson RG, Bolitho KM, Wright MA, Iturriagagoitia-Bueno T, Reid SJ, Ross GS (1998) Apple ACC-oxidaseand polygalacturonase: ripening-specific gene expres-sion and promoter analysis in transgenic tomato. PlantMol Biol 38: 449–460
Baron-Epel O, Gharyal PK, Schindler M (1988) Pectins asmediators of wall porosity in soybean cells. Planta 175:389–395
Bergey DR, Orozco-Cardenas M, de Moura DS, Ryan CA(1999) A wound- and systemin-inducible polygalacturo-nase in tomato leaves. Proc Natl Acad Sci USA 96:1756–1760
Blumenkrantz N, Asboe-Hansen G (1973) New methodfor quantitative determination of uronic acids. Anal Bio-chem 24: 484–489
Bowling DJF, Smith GN (1990) Apoplastic transport in theleaf epidermis in relation to stomatal activity. BiochemPhysiol Pflanz 186: 309–316
Buwalda JG, Meekings JS, Smith GS (1991) Seasonalchanges in photosynthetic capacity of leaves of kiwifruit(Actinidia deliciosa) vines. Physiol Plant 83: 93–98
DellaPenna D, Alexander DC, Bennett AB (1986) Molec-ular cloning of tomato fruit polygalacturonase: analysisof polygalacturonase mRNA levels during ripening. ProcNatl Acad Sci USA 83: 6420–6424
Dellaporta SL, Wood J, Hicks JB (1983) A plant DNAminipreparation: version II. Plant Mol Biol Rep 1: 19–21
Falloon RE, Sutherland PW, Hallett IC (1989) Morphologyof Erysiphe pisi on leaves of Pisum sativum. Can J Bot 67:3410–3416
Geisler M, Nadeau J, Sack FD (2000) Oriented asymmetricdivisions that generate the stomatal spacing pattern inArabidopsis are disrupted by the too many mouths muta-tion. Plant Cell 12: 2075–2086
Giovannoni JJ, DellaPenna D, Bennett AB, Fischer RL(1989) Expression of a chimeric polygalacturonase genein transgenic rin (ripening inhibitor) tomato fruit resultsin polyuronide degradation but not fruit softening. PlantCell 1: 53–63
Gleave AP (1992) A versatile binary vector system with aT-DNA organisational structure conducive to efficientintegration of cloned DNA into the plant genome. PlantMol Biol 20: 1203–1207
Hadfield KA, Bennett AB (1998) Polygalacturonases:many genes in search of a function. Plant Physiol 117:337–343
Hallett IC, MacRae EA, Wegzryn TF (1992) Changes inkiwifruit cell wall ultrastructure and cell packing duringpostharvest ripening. Int J Plant Sci 153: 49–60
Hong S-B, Sexton R, Tucker ML (2000) Analysis of genepromoters for two tomato polygalacturonases expressedin abscission zones and the stigma. Plant Physiol 123:869–882
Jarvis MC (1984) Structure and properties of pectin gels inplant cell walls. Plant Cell Environ 7: 153–164
Kalaitzis P, Solomos T, Tucker ML (1997) Three differentpolygalacturonases are expressed in tomato leaf andflower abscission, each with a different temporal expres-sion pattern. Plant Physiol 113: 1303–1308
Keller R, Springer F, Renz A, Kossman J (1999) Antisenseinhibition of the GDP-mannose pyrophosphorylase re-duced the ascorbate content in transgenic plants leadingto developmental changes during senescence. Plant J 19:131–141
Kim J-B, Carpita NC (1992) Changes in esterification of theuronic acid groups of cell wall polysaccharides duringelongation of maize coleoptiles. Plant Physiol 98: 646–653
Knox JP (1992) Cell adhesion, cell separation and plantmorphogenesis. Plant J 2: 137–141
Knox JP, Linstead PJ, King J, Cooper C, Roberts K (1990)Pectin esterification is spatially regulated both withincell walls and between developing tissues of root apices.Planta 4: 512–521
Laing W, Greer D, Sun O, Beets P, Lowe A, Payn T (2000)Physiological impacts of Mg deficiency in Pinus radiata:growth and photosynthesis. New Phytol 146: 47–57
Lever M (1972) A new reaction for colorimetric determina-tion of carbohydrates. Anal Biochem 47: 273–279
Miyairi K, Fujita K, Okuno T, Sawai K (1977) A toxicprotein causative of silver-leaf disease symptom on ap-ple trees. Agric Biol Chem 41: 1897–1902
Miyairi K, Ito K, Okuno T (1997) Preparation and charac-terization of pectic polysaccharides from silvered leavesof apple trees infected with Stereum purpureum. Bull FacAgric Hirosaki Univ 61: 1–14
Orfila C, Seymour GB, Willats WGT, Huxham IM, JarvisMC, Dover CJ, Thompson AJ, Knox JP (2001) Alteredmiddle lamella homogalacturonan and disrupted depo-sition of (135)-�-l-arabinan in the pericarp of Cnr, aripening mutant of tomato. Plant Physiol 126: 210–221
Osteryoung KW, Toenjes K, Hall B, Winkler V, BennettAB (1990) Analysis of tomato polygalacturonase expres-sion in transgenic tobacco. Plant Cell 2: 1239–1248
Petersen M, Sander L, Child R, van Onckelen H, UlvskovP, Borkhardt B (1996) Isolation and characterization of apod dehiscence zone-specific polygalacturonase fromBrassica napus. Plant Mol Biol 31: 517–527
Pressey R (1986) Extraction and assay of tomato polyga-lacturonases. HortScience 21: 490–492
Redgwell RJ, MacRae EA, Hallett I, Fischer M, Perry J,Harker R (1997) In vivo and in vitro swelling of cell wallsduring fruit ripening. Planta 203: 162–173
Redgwell RJ, Melton LD, Brasch DJ (1988) Cell-wall poly-saccharides of kiwifruit (Actinidia deliciosa): chemical fea-tures in different tissue zones of the fruit at harvest.Carbohydr Res 182: 241–258
Reiter W-D, Chapple CS, Somerville CR (1993) Alteredgrowth and cell walls in a fucose-deficient mutant ofArabidopsis. Science 261: 1032–1035
Schagger H, van Jagow G (1987) Tricine-sodium dodecylsulfate-polyacrylamide gel electrophoresis for the sepa-ration of proteins in the range from 1 to 100 kDa. AnalBiochem 166: 368–379
Schols HA, Vierhuis E, Bakx EJ, Voragen AGJ (1995)Different populations of pectic hairy regions occur inapple cell walls. Carbohydr Res 275: 343–360
Schroder R, Atkinson RG, Langenkamper G, Redgwell RJ(1998) Biochemical and molecular characterization of xy-
Atkinson et al.
132 Plant Physiol. Vol. 129, 2002
Dow
nloaded from https://academ
ic.oup.com/plphys/article/129/1/122/6110110 by guest on 17 February 2022

loglucan endotransglycosylase from ripe kiwifruit.Planta 204: 242–251
Sexton R, Roberts JA (1982) Cell biology of abscission. AnnRev Plant Physiol 33: 133–162
Shevell DE, Kunkel T, Chua N-H (2000) Cell wall alter-ations in the Arabidopsis emb30 mutant. Plant Cell 12:2047–2059
Sitrit Y, Hadfield KA, Bennett AB, Bradford KJ, DownieAB (1999) Expression of a polygalacturonase associatedwith tomato seed germination. Plant Physiol 121:419–428
Smith CJS, Watson CF, Bird CR, Ray J, Schuch W, Grier-son D (1990) Expression of a truncated tomato polyga-lacturonase gene inhibits expression of the endogenousgene in transgenic plants. Mol Gen Genet 224: 477–481
Spiers AG, Edwards WRN, Hopcroft DH (1987) Effects ofsilverleaf infection on ultrastructure of foliage of Prunus,Rosa and Populus. N Z J Bot 25: 411–423
Steele NM, McCann MC, Roberts K (1997) Pectin modifi-cation in cell walls of ripening tomatoes occurs in distinctdomains. Plant Physiol 114: 373–381
Sutherland P, Hallett I, Redgwell R, Benhamou N, Mac-Rae E (1999) Localization of cell wall polysaccharidesduring kiwifruit (Actinidia deliciosa) ripening. Int J PlantSci 160: 1099–1109
Taylor JE, Tucker GA, Lasslett Y, Smith CJS, Arnold CM,Watson CF, Schuch W, Grierson D, Roberts JA (1990)Polygalacturonase expression during leaf abscission ofnormal and transgenic plants. Planta 183: 133–138
Taylor JE, Webb STJ, Coupe SA, Tucker GA, Roberts JA(1993) Changes in polygalacturonase activity and solu-bility of polyuronides during ethylene-stimulated leafabscission in Sambucus nigra. J Exp Bot 44: 93–98
The Arabidopsis Genome Initiative (2000) Analysis of thegenome sequence of the flowering plant Arabidopsis thali-ana. Nature 408: 796–815
Tetley U (1932) The development and cytology of theleaves of healthy and “silvered” Victoria plum-trees.Ann Bot 46: 633–652
Torki M, Mandaron P, Thomas F, Quigley F, Mache R,Falconet D (1999) Differential expression of a polygalac-turonase gene family in Arabidopsis thaliana. Mol GenGenet 261: 948–952
Vaughn KC, Turley RB (1999) The primary walls of cottonfibers contain an ensheathing pectin layer. Protoplasma209: 226–237
Wang Z-Y, MacRae EA, Wright MA, Bolitho KM, RossGS, Atkinson RG (2000) Polygalacturonase gene expres-sion in kiwifruit: relationship to fruit softening and eth-ylene production. Plant Mol Biol 42: 317–328
Wood PJ, Siddiqui IR (1971) Determination of methanoland its application to measurement of pectin methylester content and pectin methylesterase activity. AnalBiochem 39: 418–428
Wu Q, Szakacs-Dobozi M, Hemmat M, Hrazdina G (1993)Endopolygalacturonase in apples (Malus domestica) andits expression during fruit ripening. Plant Physiol 102:219–225
Yao J-L, Cohen D, Atkinson R, Richardson K, Morris B(1995) Regeneration of transgenic plants from the com-mercial apple cultivar Royal Gala. Plant Cell Rep 14:407–412
Zhang GF, Staehelin LA (1992) Functional compartmenta-tion of the Golgi apparatus of plant cells. Plant Physiol99: 1070–1083
Zhao L, Sack FD (1999) Ultrastructure of stomatal devel-opment in Arabidopsis (Brassicaceae) leaves. Am J Bot 86:929–939
Ziegenspeck H (1938) Die Micellierung der turgeszenMechanismen. Teil 1. Die Spaltoffnungen (mit phyloge-netischen Ausblicken). Bot Arch 39: 268–309, 332–372
Overexpression of Polygalacturonase in Apple
Plant Physiol. Vol. 129, 2002 133
Dow
nloaded from https://academ
ic.oup.com/plphys/article/129/1/122/6110110 by guest on 17 February 2022