organization of afferent and efferent projections of the ... · projections of th e nucleus b...
TRANSCRIPT

THE JOURNAL OF COMPARATIVE NEUROLOGY 365:306-328 (1996)
Organization of Afferent and Efferent Projections of the Nucleus Basalis
Prosencephali in a Passerine, Taeniopygia guttata
J. MARTIN WILD AND SUSAN M. FAKABAUGH Department of Anatomy, School of Medicine, University of Auckland, Auckland, New Zealand
ABSTRACT The connections of nucleus basalis (NB) of the rostral forebrain of the zebra finch were
investigated electrophysiologically and with anterograde and retrograde tracing methods to determine their functional organization, the sources of their pontine afferents, and the targets of their telencephalic efferents. The nucleus was found to be partitioned into three major components, a rostral lingual part that received a hypoglossal projection via a lateral subnucleus of the principal sensory trigeminal nucleus (PrV), a middle beak part that received a trigeminal projection via a medial subnucleus of PrV, and a caudal auditory part that received a short latency auditory projection via the intermediate nucleus of the lateral lemniscus. Beak NB also received a projection from a paralateral lemniscal nucleus, and the dorsocaudal part of auditory NB and the medially adjacent neostriatum also received a projection from a lateral subnucleus of the superior vestibular nucleus (VS). The efferent projections of each of the three major parts of NB were mainly to the adjacent neostriatum frontale (NF), which then provided projections to the lobus parolfactorius (exclusive of area X), the lateral archistriatum intermedium (Ail), and the lateral neostriatum caudale (NC1). Ail received a projection from NC1 and provided terminal fields to the contralateral NCl and the NF. The major projections of Ail, however, descended bilaterally through the brainstem via the occipitomesencephalic tracts, with dense terminations in the medial spiriform nucleus and with extensive bilateral terminations throughout the lateral reticular formation of the pons and medulla. For the most part, jaw, tongue, and tracheosyringeal motor nuclei did not receive terminations.
The results suggest that NB in zebra finch, like NB in pigeon and duck, is likely to be a major component of trigeminal sensorimotor circuitry involved in feeding and in other oral- manipulative behaviors. Results also show that the auditory component of NB is not directly linked to the vocal control system at telencephalic levels, but the possibility remains that the lingual, beak, and auditory parts of NB play a role in vocalization by multisynaptic influences on cranial nerve motor nuclei innervating various parts of the vocal tract. 0 1996 ~i ’ i l ey -~ i s s , Inc.
Indexing terms: zebra finch, trigeminal, auditory, song system, vocal control
In birds, nucleus basalis (NB) is a rostral forebrain structure that has long been known to be concerned functionally with sensory input from the beak and the oral cavity (Wallenberg, 1903; Witkovsky et al., 1973; Berk- houdt et al., 1981; Dubbeldam et al., 1981; Wild et al., 1985; Felix and Roesch, 1986). This input is largely derived from the principal sensory nucleus of the trigeminal complex (PrV), which projects directly to NB without an intervening thalamic relay. In some species with specialized tongues and feeding habits, this input includes a lingual component, which, in ducks, is mediated by glossopharyngeal and minor facial projections to a specific dorsomedial portion of PrV (Dubbeldam, 1980; Dubbeldam et al., 1979, 1981; Dubbel-
dam and Den Boer-Visser, 1994) and which, in other species (e.g., finches and parrots), possibly is mediated by hypoglos- sal projections to a circumscribed lateral portion of PrV (Wild, 1981, 1990; Cam and Wild, 1984a). There is thought to be no trigeminal innervation of the tongue in birds (Cords, 19041, although this may not be true for all species (Carr and Wild, 1984b). NB in pigeons, ducks, and geese projects to adjacent regions of the frontal “neostriatum” (Wild et al., 1985; Veenman and Gottschaldt, 1986; Dubbel-
Accepted August 3,1995 Address reprint requests to Dr. J.M. Wild, Department of Anatomy, School
of Medicine, university of Auekland, Auckland, New Zealand.
D 1996 WILEY-LISS, INC.

NUCLEUS BASALIS IN ZEBRA FINCH 307
dam and Visser, 1987), which, in turn, projects to the caudolateral neostriatum and to a circumscribed “somatic sensorimotor” portion of the archistriatum intermedium (Ai; Zeier and Karten, 1971). In ducks and geese, NB has other projections as well. In pigeons and ducks, Ai is the source of long descending projections to regions of the rhombencephalic lateral reticular formation that contain premotor neurons for the trigeminal and facial motor nuclei, which innervate the jaw muscles (Wild and Zeigler, 1980; Arends and Dubbeldam, 1982; Berkhoudt et al., 1982). These connections complete a trigeminal sensorimo- tor circuit that mediates control of the jaw during feeding and other activities involving the beak, the bird’s multipur- pose prehensile organ (Zeigler, 1989; Dubbeldam and Den Boer-Visser, 1994; Zeigler et al., 1994).
In addition to inputs from PrV, NB in birds has been reported to receive a variety of other inputs. The best known of these is a short-latency auditory projection (Maekawa, 1981, 1987; Schall and Delius, 1986; Schall, 1987) that originates in the intermediate nucleus of the lateral lemniscus and terminates in caudal regions of NB (Kirsch, 1983; Arends and Zeigler, 1986; Schall et al., 1986; Hall et al., 1993; Striedter, 1994). This represents a second pathway for auditory information to reach the telencepha- lon in birds, the first or classical pathway relaying through nucleus ovodalis (Ov) of the thalamus to field L, which is the primary telencephalic auditory field (Karten, 1968; Wild et al., 1993). The functional significance of the audi-
13 Ail Am An BC c0 DMN X DT E FAT FLM HA Hb HD H v HVC
ICo I 0 IX-x
La LH LLI LLIC LLIr lMAN LMD LoC LPO MC Mv N NB NC NC1 NF NIII N N N v
tory pathway to NU has been controversial, but, in the budgerigar, it has been suggested to be the primary source of auditory input for the neural control of vocal learning and vocal production. This is because, in this species, NB has some indirect connections with the vocal control nuclei (Striedter, 19941, whereas connections of field L with these nuclei have not been found (Hall et al., 1993). Furthermore, lesions of NB, but not of field L, impair the production of learned vocalizations in budgerigars (Hall et al., 1994). In contrast, the vocal learning and vocal control system of oscine songbirds receives auditory input from field L (For- tune and Margoliash, 1992; Margoliash et d., 1994; Wild, unpublished observations); however, whether NB in song- birds also contributes inputs to the vocal control system is not known, because information on the connections of NB in passerines is minimal (Kirsch, 1983). Based on these differences in auditory-vocal connections and on differences in histochemical properties of the vocal control nuclei of songbirds and parrots (Ball, 1990; Ball and Castro, 1991; Gahr et al., 1993), Striedter (1994) contends that the control systems for learned vocalizations have arisen inde- pendently in these two orders of birds. Additional informa- tion relevant to the assessment of this important compara- tive hypothesis would be provided by an investigation like the present one, which was designed specifically to deter- mine whether there are any direct or indirect links of the auditory component of NB with the vocal control system in a passerine.
Abbreviations
lamina 13 of the tectum achistriatum intermedium, pars lateralis nucleus ambiguus nucleus angularis brachium conjunctiwm chiasma opticum nucleus motorius dorsalis nervi vagi dorsal thalamus ectostriatum tractus frontoarchistriatalis fasiculus longitudinalis medialis hyperstriatum accessoriu m nucleus habenularis hyperstriatum dorsale hyperstriatum ventrale high vocal center (formerly hyperstriatum ventrale, pars
nucleus intercollicularis nucleus isthmoopticus nucleus nervi glossopharyngei et nucleus motorius dorsalis
nucleus laminaris lamina hyperstriatica nucleus lemnisci lateralis intermedius nucleus lemnisci lateralis intermedius, pars caudalis nucleus lcmnisci lateralis intermedius, pars rostralis lateral magnocellular nucleus of the anterior neostriatum lamina medullaris dorsalis locus coeruleus lobus parolractorius nucleus magnocellularis nucleus motorius nervi trigemini neostriatum nucleus basalis neostriatum caudale neostriaturn caudale, pars lateralis neostriatum frontale n e m s oculomotorii nervus trochlearis nervus trigeminus
caudale)
nervi vagi
NVI NVII 0 1 OM 0s o v PA PL PLL PM P P PrV PT R RA RAm RF RPgc Ru S SpL SpM SSP TPc TS TSM TTD Uva VeD VeL VeM VI VIId VIIi VIIV vs X XI1 XI1 1 XIIts
n e m s abducens nervus facialis nucleus olivaris inferior tractus occipitomcsencephalicus nucleus olivaris superior nucleus ovoidalis paleostriatum augmentatum nucleus pontis lateralis nucleus paralemnisci lateralis nucleus pontis medialis paleostriatum primitivum nucleus sensorius principalis nervi trigemini nucleus pretectalis nucleus raphes nucleus robustus archistriatalis nucleus retroambigualis reticular formation nucleus reticularis pontis, pars gigantocellularis nucleus ruber nucleus solitarius nucleus spiriform lateralis nucleus spiriform medialis nucleus supraspinalis nucleus tcgmenti pedunculopontinus, pars compacta tractus solitarius tractus septomesencephalicus nucleus et tractus descendens nervi trigemini nucleus uvaeformis nucleus vestibularis decendens nucleus vestibularis lateralis nucleus vestibularis medialis nucleus abducens nucleus motorius nervi facialis, pars dorsalis nucleus motorius nervi facialis, pars intermedialis nucleus motorius ncrvi facialis, pars ventralis nucleus vestibularis superior area X of the parolfactory lobe nucleus nervi hypoglossi nucleus nervi hypoglossi, pars Iingualis nucleus nervi hypoglossi, pars tracheosyringealis

308 J.M. WILD AND S.M. FARABAUGH
A second motive for our study was to provide an anatomi- cal basis for future neurobehavioral studies that would assess the role NB might play in the coordination of beak and tongue movements during vocalization. Although it has generally been thought that vocal tract resonances play no part in bird vocalizations, recent studies show that the songbird’s vocal tract acts as an acoustic filter that is actively coordinated with the output of the syrinx on a note-by-note basis (Nowicki and Marler, 1988; Hausberger et al., 1991; Westneat et al., 1993). A bird might actively alter the resonant frequency of its vocal tract by lengthen- ing the trachea, by flaring the beak and glottis, or possibly by altering the position of the tongue. Video analysis of singing swamp sparrows (Melospizu melodza) shows that birds open their beaks wider for notes of’ higher frequency (Nowicki and Marler, 1988; Westneat et al., 1993); thus, by possessing a sensory representation of the beak and tongue as well as an auditory input, NB may well play a role in the sensory control or coordination of orolingual and syringeal movements during vocalization. In the present study, the intra- and extratelencephalic connections of NB were exam- ined in the zebra finch, a readily available oscine passerine for which we possess the most detailed knowledge of the vocal control system in aves (reviewed in Konishi, 1989) and that has a well-developed feeding apparatus, including a large seed-cracking beak and a highly specialized tongue (Wild, 1990). Zebra finches also have a learned, stereotyped, repetitive song that is composed of high- and low-frequency notes. Preliminary data generated by the use of an on-line beak-gape monitoring system (Deich et al., 1985) during vocalization in the zebra finch indicate that beak move- ments are also stereotyped and repetitive (Suthers, Ber- mejo, Goller, Wild, and Zeigler, unpublished observations), rendering the mechanisms by which zebra finches coordi- nate beak movements and vocalizations more amenable to future anatomical and functional analyses.
MATERIALS AND METHODS Subjects were 27 adult male zebra finches (Taenzopygiu
guttutu). Each subject was anesthetized with an intramus- cular injection of ketamine (50 mg/kg) and xylazine (6 mgikg) and was placed in a Kopf-type stereotaxic apparatus equipped with custom-made hollow ear bars and beak bar. An unpublished stereotaxic atlas of the zebra finch brain (generously donated by M. Konishi) was used to aid in the location of structures for electrophysiological recording and injections of tracers. In the coordinate system of this atlas, the beak is placed at 45 degrees below the horizontal plane, and the confluence of the superior sagittal and cerebellar sinuses (the “Y’ sinus) is placed 0.3 mm caudal to intraau- ral zero. Electrophysiological recordings were made system- atically from NB by using tungsten microelectrodes (Freder- ick Haer; 3-5 M n ) . An indifferent electrode was either inserted into a neck muscle or attached to the head skin. Tactile stimuli in the form oftouch, pressure, stroking, etc., were applied to the beak, oral cavity, and tongue by using a fine brush, a pointed probe, or tweezers. Auditory stimuli were presented in the form of 10 msec clicks delivered at 1 Hz via a 1.3 m plastic tube attached to the hollow ear bar. Other auditory stimuli in the form of human whistles, voice, claps, key jingles, etc., were delivered in the “free field,” but there was no attempt to present these in any particular order or to quantify responses evoked by them. The principal objective of these procedures was simply to
define and distinguish somatosensory and auditory regions of NB and, if possible, to distinguish oral (trigeminall from lingual (hypoglossal) components of the somatosensory region. Evoked responses were monitored on an oscillo- scope and over a loud speaker in conventional fashion, and auditory evoked potentials were recorded and averaged by using a MacLab14 data acquisition system, a Macintosh I1 computer, and Scope software (Analog Digital Ltd.). Follow- ing recordings, the tungsten electrode was removed, a glass micropipette (outer diameter, 20 Fm) attached to an elec- trode carrier was lowered to the same coordinate, and injections of tracer (n = 11) were made at the recording site. These were either air-pressure injections (picospritzer) or iontophoretic injections (positive current; 7 seconds on/7 seconds off for 10-20 minutes) of 10% biotinylated dextran amine (BDA; Molecular Probes, Inc.) in phosphate-buffered saline (PBS; pH 7.4; Veenman et al., 1992) or picospritzer injections of 1.5% cholera toxin B-chain (CTB; List Labora- tories, Inc.) in PBS. To confirm the boundaries of NB by anterograde transport, unilateral injections of BDA or CTB were made into or near the PrV or into the intermediate nucleus of the lateral lemniscus (LLI) in 12 birds following electrophysiological recordings of multiunit activity evoked either by tactile stimulation of the beaks (for PrV) or by auditory stimuli (clicks for LLI). To validate the efferent projections of NB by retrograde labeling and to define subsequent intra- and extratelencephalic projections of the “basalis-neostriatal complex” (Dubbeldam and Visser, 1987), other injections of BDA or CTB were made into the frontal neostriatum (NF) surrounding NB (n = 6 ) , into the hyperstriatum ventrale (HV; n = 31, into the caudolateral part of the neostriatum (NCI; n = 81, or into the lateral archistriatum (n = 2). The variability in numbers of injections per region was a function of the accuracy with which terminal fields or nuclei could be targeted. In 13 of the birds, injections of BDA and CTB were made into the same or opposite sides of the brain in order to maximize results and to minimize the total number of birds used. The survival time was 4 days, after which the birds were deeply anesthetized and were perfused through the heart with saline followed by 4% paraformaldehyde in PBS, pH 7.4. The brains were postfixed for 2-5 hours, blocked in the same transverse plane as that used for recording and injection, and then equilibrated in 25% sucrose buffer overnight. Serial transverse sections were cut frozen at 40 km, collected in PBS in three series, washed again in PBS, and then treated with 50% methanol and 0.1%) H202 for 30 minutes to aid subsequent penetration of the antibody and to quench endogenous peroxidase activity. BDA was visual- ized by incubating sections in ExtrAvidin peroxidase conju- gate (Sigma Chemical Company; 1:850 in 0.3% Triton PBS) for 1 hour at room temperature followed by 3-3‘-diamino- benzidine (DAB; 0.025%) and HzOz. CTB was visualized using sequential incubations in a goat anti-CTB antibody (List Laboratories; 1:30,000; 15 hours at PC), rabbit anti-goat IgG (Sigma; 1:200), and ExtrAvidin peroxidase conjugate (1:850). In cases receiving both BDA and CTB injections, BDA was visualized first, and the reaction product was colored black by the addition of 0.002% CoClz to the DAB mixture. CTB was visualized second, and the reaction product was colored brown by DAB without CoCI,. At least one series of sections in each case was subsequently counterstained with Giemsa on the slide for the identifica- tion of nuclear groups.

NUCLEUS BASALIS IN ZEBRA FINCH 309
RESULTS Normal anatomy of NB
In Nissl-stained normal material, NB in the zebra finch appears as a compact nucleus of small, densely packed cells situated on the lateral aspect of the paleostriatal complex and is clearly distinguishable from the larger, less densely packed cells of the surrounding NF (Fig. 1). At rostral and middle levels of NB, these latter cells are arranged around NB in two layers, with the layer nearest to NB in the form of radially aligned columns. The very caudal pole of NB is situated under the horizontal or concave ventral border of the rostral ectostriatum, and the overlap of these two structures lies in the same rostrocaudal plane as the rostral extent of the optic chiasm. NB then extends rostrally for just over 2 mm though the level of the rostral pole of area X, the large comma-shaped nucleus within the lobus parolfac- torius (LPO) that is clearly visible in well-counterstained sections of the male brain. In the transverse plane, NB is crescent shaped throughout much of its rostrocaudal ex- tent, measuring approximately 0.5 mm in width at its widest point. The concavity of the crescent is applied to the lateral border of the paleostriatum augmentatum (PA) at more caudal levels and of LPO more rostrally, whereas its convexity bulges into the laterally adjacent neostriatum. At the most rostral levels, however, the crescent of NB be- comes a narrow wedge that protrudes laterally from the border of LPO for approximately 1 mm; thus, it takes the form of a “tongue” of cells that, in fact, does contain the representation of the tongue (see below).
Functional subdivisions of NB Electrophysiologically, NB was found to be divided into
three distinct parts, with auditory responses confined to caudal regions, somatosensory responses to tactile stimula- tion of the beaks confined to middle regions, and responses to tactile stimulation of the tongue confined to rostral regions. The most caudal point of NB from which auditory responses could be evoked was found immediately ventral to the most rostral level of the ectostriatum, from which visual responses could be elicited simply by interrupting the light beam of the microscope that illuminated the prepara- tion. Robust click-evoked responses of very short latency ( < 3 msec) were recorded from this caudal pole of NB and from passes made successively rostrally at 1-200 p,m intervals through the next 0.75 mm of the nucleus (Fig. 2). Typical stereotaxic coordinates corresponding to the extent of what is henceforth called auditory NB were A2.1-2.8, L4.0-3.7, and 2.7-3.5 below the dura. Although the use of different auditory stimuli did not change our overall assess- ment of the physical extent of auditory NB, not all stimuli were equally effective in evoking responses. Clicks evoked responses most reliably; sometimes, they were the only stimulus capable of evoking a response, possibly because of their broad bandwidth. In other cases, however, human voice was the most effective stimulus, and this produced changes in the level of activity of multiunit responsiveness that seemed to “follow” faithfully the natural frequency (rather than the amplitude) modulations of the talking voice. Qualitative and preliminary though these observa- tions are, they suggest that auditory NB units may be tuned to rather specific frequency ranges. Clearly, further detailed investigations of single units in auditory NB under con- trolled acoustic conditions would prove rewarding.
Rostra1 to auditory NB for approximately 0.9 mm through middle levels of the nucleus, only responses to tactile stimulation of the beak and oral cavity, exclusive of the tongue, could be elicited. The most effective stimulus was one that moved relative to the beak (e.g., a stroke) rather than a touch at a single point. Receptive fields were frequently small and were confined to particular areas of either the upper or the lower beak. In general, ipsilaterally applied stimuli evoked stronger responses than contralater- ally applied stimuli. Again, however, a detailed somatotopic mapping of the nucleus was beyond the scope of the study; rather, our intention was to gain an estimate of the extent of the nuclear region from which responses to the applica- tion of any somatosensory stimulus to the beaks could be elicited. This region is henceforth called beak NB and has the following typical stereotaxic coordinates: A2.f?-3.7, L3.5-3.0, and 3.3-4.0 below the dura.
Extending rostral to beak NB for another 0.5 mm was a dorsoventrally narrow region from which only responses to tactile stimulation of the tongue could be elicited. Again, small strokes or scrapes applied to the tongue surface were the most effective stimuli, but ipsicontralaterality could not be assessed because of restricted access to the inside of the mouth. Responses from what is henceforth called lingual NB were confined to a region corresponding to the following stereotaxic coordinates: A3.7-4.2, L3.2-2.4, and 3.5-3.8 below the dura.
Topography of afferent projections to NB Injections of BDA that were largely confined to NB
produced retrograde labeling of cell bodies in the pons and in the H V . In the pons, the specific location of labeled cells depended on which part of NB was injected, and, in each case, labeled cells were bilaterally distributed and were located in the same nucleus or part of a nucleus on each side, with a slight preponderance of ipsilaterally labeled cells. When injections were made into lingual NB (Fig. 4A), labeled cells were found predominantly in the lateral part of PrV, and a few were found along the dorsal border of the medial part (Fig. 3A,a). The lateral part of PrV forms a large, distinct, oval subnucleus separated from the medial part of PrV by a continuous border of cells. This lateral part of PrV and the dorsal border of the medial part receive a major primary afferent projection from the tongue via the hypoglossal nerve (Wild, 1990). When injections were largely confined to beak NB (Fig. 4B), labeled cells were found predominantly in the medial part of PrV, excluding its dorsal border (Fig. 3B,b). When injections were made into auditory NU (Fig. 4C), labeled cells were confined to the intermediate nucleus of the lateral lemniscus (LLI) (Fig. 3C,c). The bulk of this nucleus lies rostrally at isthmic levels and is known as LLIr (Arends and Zeigler, 1986). At these levels, it is round in cross section, but it has a tapering tail that extends caudally as far as the ventral border of PrV, where it is known as LLIc. Injections into beak NB or into both tongue and beak NB also retrogradely labeled a previously unidentified nucleus situated dorsomedial to LLIr and dorsal to the rostral pole of the trigeminal motor nucleus, where it lies in the path of the axons of LLIr neurons (Fig. 3D,d). Because it appears to form a dorsome- dial extension of LLIr, it will be called here thepuruluterul lemniscal nucleus (PLL), although it can clearly be distin- guished from LLIr in Nissl-stained material(Fig. 3c) and on the basis of retrograde labeling from different parts of NB. For example, in a case in which a BDA injection was made

Figure 1

NUCLEUS BASALIS IN ZEBRA FINCH 311
80
40 n > rl
Q)
(d
v
0 0 c. - >o
-40
-80
0 5 10 15 20
Time (ms)
Fig. 2. Averaged evoked potential recorded from the locus shown in the caudal part of NB (at right) to 50 presentations of a 10 msec click presented at 1 Hz to the contralateral ear. The click was presented at
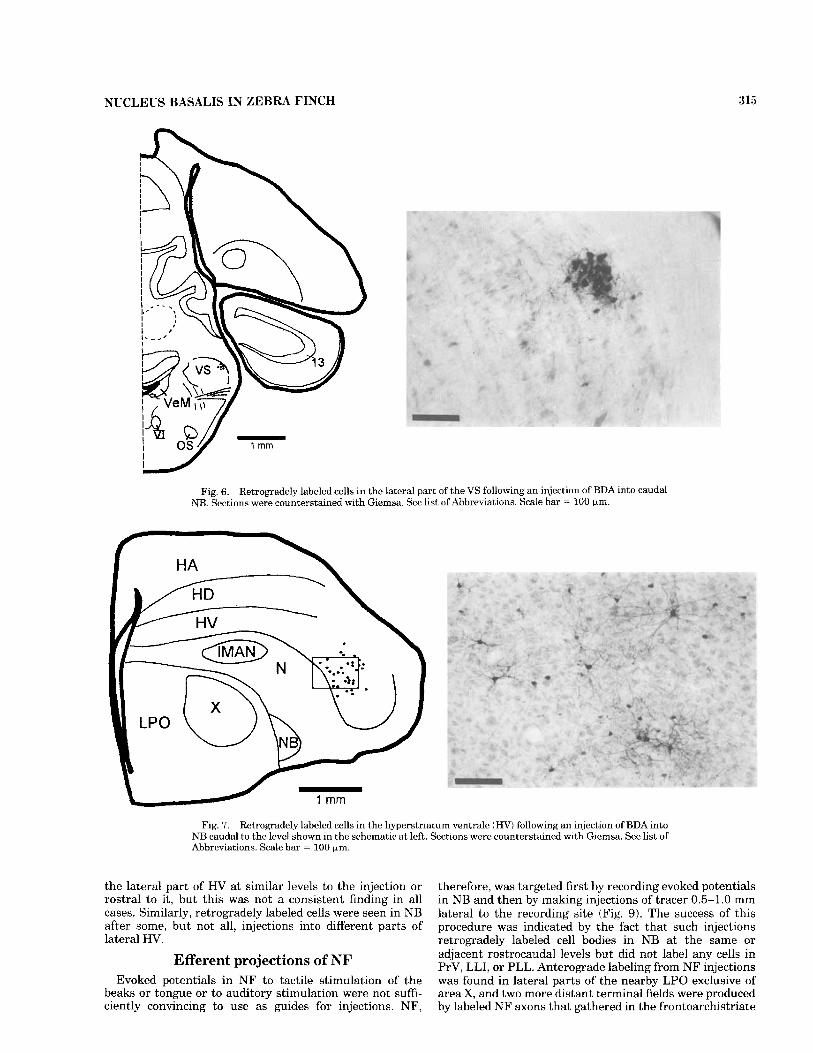
into auditory NB, and an injection of CTB was made into the lingual and beak parts of NB, there were no double- labeled cells in either PrV or LLI. The black cells (see Materials and Methods) labeled from the injection of BDA into auditory NB were confined to LLI, whereas brown cells labeled from the CTB injection into the lingual and beak parts of NB were located in both lateral and medial parts of PrV and in PLL (Fig. 3D,d). In addition to labeled cells in PrV, LLI, and PLL, injections into caudal, but not into rostral, NB retrogradely labeled a small cluster of cells bilaterally in the lateral part of the superior vestibular nucleus (VS) (Fig. 6).
Following injections into NB, labeled cells were also found in the lateral part of H V , in each case slightly rostral to the injection site in each of the three functional parts of NB (Fig. 7). The projection of HV to NB was confirmed by injections into HV that produced terminal labeling in NB, but these injections also produced denser terminal labeling in NF immediately adjacent to NB.
Topography of efferent projections to NB Definition of the extent and borders of NB and of its
functional subdivisions was also provided by injections of tracer into either NB or PrV. The clearest and most comprehensive topographical results were produced by unilateral injections into electrophysiologically defined re- gions of NB, and some of these were the same injections
Fig. 1. Transverse sections through the nucleus basalis (NB) of the zebra finch at rostral (A,a), middle (B,b), and caudal (C,c) levels. The regions of the photomicrographs shown in a-c are boxed in the drawings of the hemisections shown in A-C; respectively. Sections were counterstained with Giemsa. See list of Abbreviations. Scale bars = 200 pm in a x .
25
time 0, and the latency to the first deflection of the potential included 3.3 msec of travel time from the click generator to the external auditory meatus. See list of Abbreviations.
that also produced the retrograde labeling described above (see ‘I’opography of afferent projections to NB). Each injec- tion produced a terminal field in the contralateral NB, the position, shape and extent of which reflected the position and extent of the injection (Fig. 4); that is, the terminal field produced by each injection that was largely confined to a specific functional subdivision of NB was also largely confined to the same subdivision on the opposite side of the brain. These contralateral terminal fields were not pro- duced by commissural, reciprocal projections of NB but were the result of somatopetal (retrograde) transport to the parent cell bodies in the pons and subsequent somatofugal (anterograde) transport to the contralateral NB via branched axons (see Discussion).
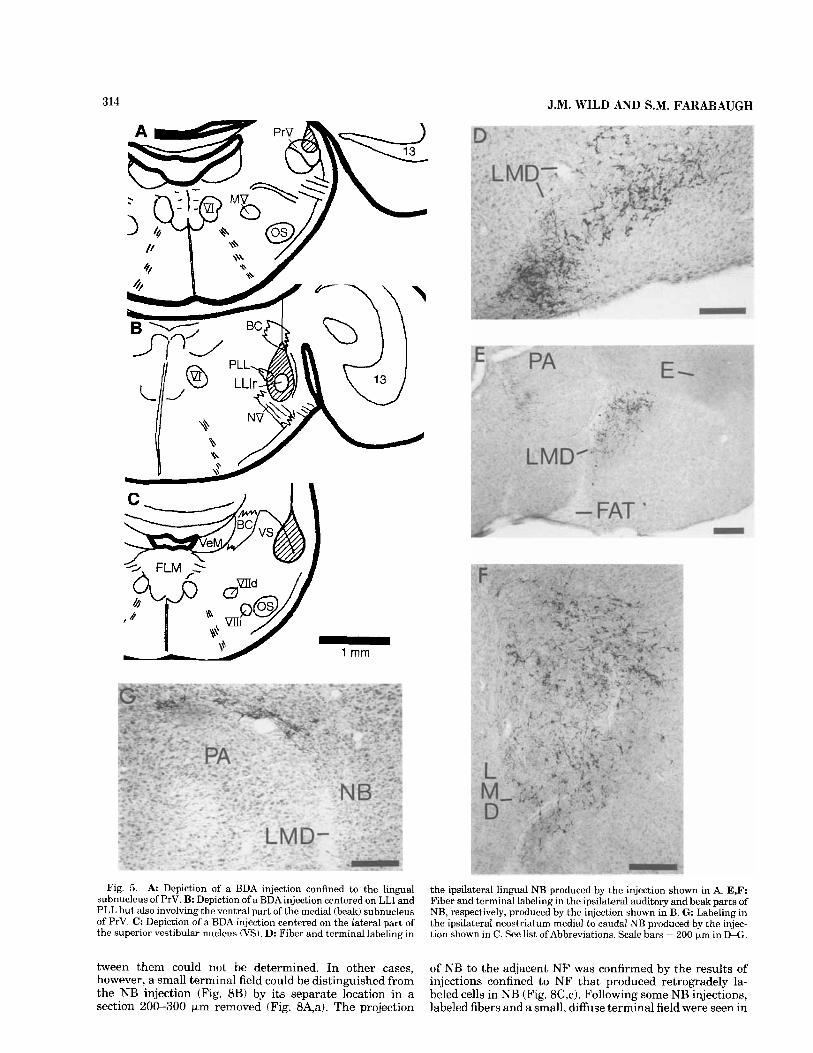
Because of the conformation of the brain at isthmic levels, it proved difficult to confine injections to every subnuclear component of the sources of afferents to NB; although, in general, the results were complementary to, and thus confirmed, the results of the retrograde tracing studies described above. Injections into PrV showed that the terminations in NB were bilateral and were limited to the cytoarchitecturally defined nucleus as described above. Judged by eye, ipsilateral terminations were only slightly heavier than contralateral terminations. Specifically, an electrophysiologically controlled injection confined to the lingual part of PrV produced terminal labeling in NB that was confined to its lingual portion (Fig. 5A,D). In contrast, an injection that was centered on LLI but that also included PLL and ventral portions of the medial (beak) part of PrV (Fig. 5B) produced terminal labeling throughout auditory NB and parts of beak NB but did not label any part of lingual NB (Fig. 5E,F). In three cases with injections aimed at PrV, the center of the injection was located caudal to the intended level and covered the lateral aspect of VS (Fig. 5C). In these cases, the projections to the telencephalon were specifically limited to the most dorsocaudal border of NB
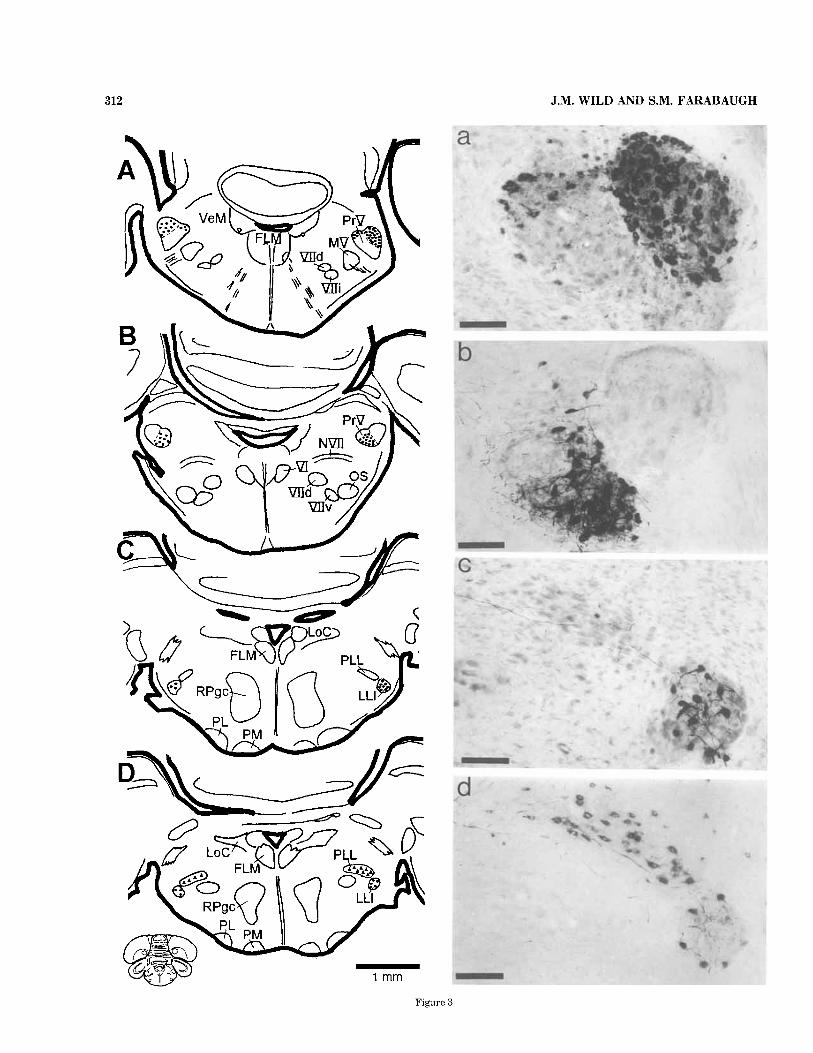
312 J.M. WILD AND S.M. FARABAUGH
Figure 3

NUCLEUS BASALIS IN ZEBRA FINCH 313
Fig. 4. A-C: Depictions of injections into the right lingual. beak, and auditory NB, respectively. The terminal labeling in the contralat- eral NB produced by these injections via somatopetal and somatofugal transport (see text) is depicted on the left in A-C. Note that the terminal labeling shown in C is at the very caudal pole of auditoiy NB (compare to Fig. 5E). These injections also produced the pattern of
retrograde labeling shown in Figure 3. D: Photomicrograph of terminal labeling in the contralateral lingual NB produced by the BDA injection shown in A. E: Photomicrograph of terminal labeling in the largest part of the contralateral auditory NB produced by a large CTB injection centered at about the same location as the small BDA injection shown in C. See list ofAbbreviations. Scale bars = 200 pm in D,E.
Fig. 3. Retrograde labeling in PrV, LLI, and PPL following injec- tions of BDA or CTB into NB depicted in Figure 4A-C. A,a: Cells in the lateral nucleus of PrV and in the dorsal part of the medial nucleus following an injection of CTB into rostral NB (see Fig. 4A). B,b: Cells in the medial nucleus of PrV following an injection of BDA into the middle part of NB (see Fig. 4B). C,c: Cells in LLI following an injection of BDA into caudal NB (see Fig. 4C). D,d: Cells in LLI (indicated by solid triangles in schematic and colored black in the original; see Materials and Methods) following an injection of BDA into caudal PiB (see Fig. 4C) and into PLL (indicated by solid circles in schematic and colored brown in the original) following an injection of CTB into the middle part of NB (not shown in Fig. 4). There was no admixture of brown and black cells in either LLI or PLL. Note that, in all cases, the labeling is bilateral, but it is predominantly ipsilateral. Note also that, in c and d, the axons of LLI cells pass through PLL as they leave the nucleus to head medially and rostrally. Giemsa counterstain was used in a x , and d was not counterstained. See list of Abbreviations. Scale bars = 100 pm in a-d.
and the medially adjacent neostriatum bordering the lamina medullaris dorsalis (Fig. 5G).
Efferent projections of NB Apart from the terminal field in the opposite NB, which
was not produced by NB efferents but by somatopetal and somatofugal transport to and from pontine cell bodies (see above), the only other suggestion of terminal fields follow- ing injections into NB were located in N F laterally or rostrolaterally adjacent to the injection site. In some cases, these apparent terminal fields consisted of labeled fibers and varicosities that extended laterally from the injec- tion site as far as the lamina hyperstriatica (LH), which separates the neostriatum from the H V . But, because there was no unlabeled region between the injection into NB and the putative terminal field more laterally, the border be-
314 J.M. WILD AND S.M. FARABAUGH
Fig. 5. A: Depiction of a BDA injection confined to the lingual subnucleus of PrV. B: Depiction of a BDA injection centered on LLI and PLL but also involving the ventral part of the medial (beak) subnucleus of PrV. C: Depiction of a BDA injection centered on the lateral part of the superior vestibular nucleus (VS). D: Fiber and tcrminal labeling in
the ipsilateral lingual NB produced by the injection shown in A. E,F: Fiber and terminal labeling in the ipsilateral auditory and beak parts of NB, respectively, produced by the injection shown in B. G Labeling in the ipsilateral neostriatum medial to caudal NB produced by the injec- tion shown in C. See list of Abbreviations. Scale bars = 200 p,m in D-G.
tween them could not be determined. In other cases, however, a small terminal field could be distinguished from the NB injection (Fig. 8B) by its separate location in a section 200-300 pm removed (Fig. 8A,a). The projection
of NB to the adjacent N F was confirmed by the results of injections confined to NF that produced retrogradely la- beled cells in NB (Fig. 8C,c). Following some NB injections, labeled fibers and a small, diffuse terminal field were seen in
NUCLEUS BASALIS IN ZEBRA FINCH 315
Fig. 6. Retrogradely labeled cells in the lateral part of the VS following an injection of BDA into caudal NB. Sections were counterstained with Giemsa. See list of Abbreviations. Scale har = 100 pm.
Fig. 7. Retrogradely labeled cells in the hyperstriatum ventrale ( H V ) following an injection of BDA into NB caudal to the level shown in the schematic at left. Sections were counterstained with Giemsa. See list of Abbreviations. Scale bar = 100 wm.
the lateral part of H V at similar levels to the injection or rostra1 to it, but this was not a consistent finding in all cases. Similarly, retrogradely labeled cells were seen in NB after some, but not all, injections into different parts of lateral H V .
Efferent projections of NF Evoked potentials in NF to tactile stimulation of the
beaks or tongue or to auditory stimulation were not suffi- ciently convincing to use as guides for injections. NF,
therefore, was targeted first by recording evoked potentials in NB and then by making injections of tracer 0.5-1.0 mm lateral to the recording site (Fig. 9). The success of this procedure was indicated by the fact that such injections retrogradely labeled cell bodies in NB at the same or adjacent rostrocaudal levels but did not label any cells in PrV, LLI, or PLL. Anterograde labeling from NF injections was found in lateral parts of the nearby LPO exclusive of area X, and two more distant terminal fields were produced hy labeled NF axons that gathered in the frontoarchistriate

316 J.M. WILD AND S.M. FARABAUGH
, . ... . ..~.. . ,
NB
1 mm
Fig. 8. a: Darkfield photomicrograph showing terminal labeling in the neostriatum frontale (NF) at the location boxed in A following the injection of BDA into NB at a more caudal level \also shown in B). c:
Retrograde labeling in NR at the location hnxed in C following an injection of BDA into the laterally adjacent NF (also shown in C ) . See list ofhbbreviations. Scale bars = 200 krn in a, 100 brn in c.

NUCLEUS BASALIS IN ZEBRA FINCH
tract (FAT) and passed caudally. A smaller field was located in the lateral archistriatum at the level of nucleus robustus, and a much larger and more diffiise one was located in the lateral part of NCl, slightly more caudally (Fig. 9). The projections of different parts of NF corresponding to each of the three parts of NB (auditory, beak, and lingual, i.e., caudal, middle, and rostral parts of NF, respectively) had slightly different but partly overlapping territories of termi- nation within both the lateral archistriatum intermedium (Ail) and the NC1. The projections of the caudal NF terminated most ventrally in Ail and most caudolaterally in NC1 (Figs. 9C, lOB), the projections of middle regions of NF terminated slightly more dorsally in Ail and more dorsally in NC1 (Fig. 9B), and the projections of rostral NF termi- nated slightly more laterally in Ail and medial to the projections of mid-NF in NCl (Figs. 9A, 10A).
Retrograde confirmation of the NF projections was pro- vided by injections of either BDA or CTB into the lateral archistriatum (Fig. 11) or into NCl (Fig. 12). Injections into either site produced a similar pattern of retrograde labeling within the lateral parts of NF that surround NB, but, for the most part, this labeling did not extend into the circum- ferential strip of radially aligned columns of NF cells immediately adjacent to NB and did not label any cells in NB itself. Both Ail and NC1 injections also labeled numer- ous neurons in other parts of the ipsilateral frontal pallium. In the case of Ail injections, retrogradely labeled cells in the lateral magnocellular nucleus of the anterior neostriatum (1MAN) were likely caused by interruption of MAN axons that pass through the dorsolateral part of the caudal neostriatum en route to the robust nucleus of the archistria- tum (RA, Bottjer et al., 1989). In the case of NCl injections, it is unclear whether the retrogradely labeled cells outside NF resulted from the injections in the region of the NF terminal fields or from spread of the injection to regions of the caudolateral neostriatum outside those fields. NCl injections also produced retrogradely labeled cells in the contralateral, but not the ipsilateral, Ail (not shown), a putative Ad projection that was confirmed in anterograde tracing experiments (see below and Fig. 14). Injections into Ail also produced retrograde, but not anterograde, labeling in the contralateral Ail (Fig. 13C), but it is believed that these cells were labeled by the interruption of Ail s o n s passing though the site of injection en route to NC1 (note preceding sentence and Fig. 14).
317
Efferent projections of NCl and lateral archistriatum
The injections of CTB into NC1 produced terminal fields in the ipsilateral lateral archistriatum that overlapped the terminal fields of the NF projection (Fig. 12D). These projections were confirmed by massive retrograde labeling in NCl following injections of BDA into the ipsilateral lateral archistriatum (Fig. 11D). Both of the two injections into the lateral archistriatum produced anterograde, but not retrograde, labeling in the contralateral NC1 (Figs. 13C, 14) and in the contralateral NF (Figs. 11, 14). The Ail projection to the contralateral NCl was confirmed by the presence of retrogradely labeled cell bodies in Ail following injections into the contralateral NCl (see above). However, for reasons that are not entirely clear, injections of BDA into NF failed to produce retrograde labeling of cells in the contralateral Ail. The route of the putative Ail projection to the contralateral NF was via the anterior commissure and then through the contralateral paleostriatum, the LPO,
and the FAT. In both Ail cases, the distribution of the anterograde labeling in contralateral NF resembled the distribution of retrogradely labeled cell bodies on the ipsilateral side (Fig. 11E,F). The Ail injections also pro- duced bilateral terminal labeling within lateral regions of LPO itself, but this labeling completely avoided area X (Fig. 11B,C). The bulk of the lateral archistriatal efferents, however, was bilaterally directed into the brainstem via the occiptomesencephalic tracts (OM), although the labeling was heavier on the ipsilateral side. The projections in the two cases were very similar in terms of their trajectories and patterns of termination, although the larger pico- spritzer injection (shown in Fig. 13) produced denser and more extensive labeling than the smaller iontophoretic injection. Labeled fibers left the hemisphere in the lateral aspect of the OM (Fig. 13A,B). In the posterior thalamus, there was a dense terminal field confined to the ventral or medial part of nucleus spiriform medialis (SpM; Fig. 13B,b), and there was more diffuse labeling more dorsally. At pontine levels (Fig. 13C), labeled fibers left OM to surround PrV and encroach on its medial border, and the fifth and seventh motor nuclei, which innervate the jaw muscles (Wild and Zeigler, 19801, were surrounded particularly dorsally by labeled fibers passing caudally and ventrally, although there were a few terminations within the body of the nuclei themselves. More caudally, the major terminal field of the lateral archistriatal efferents was extensively distributed throughout the parvocellular reticular forma- tion of the caudal pons (Fig. 13D), and, at these levels, fibers crossed through the middle of the tegmentum to terminate relatively densely on the contralateral side. At medullary levels (Fig. 13E-G), there was a further crossing over of labeled fibers, particularly in the commissura infima dorsal to the dorsal motor nucleus of the vagus (Fig. 13F), and, at these levels, there were conspicuous terminal fields ventro- lateral to the hypoglossal nucleus and sparser fields more ventrally throughout the medullary reticular formation. Fibers continued caudally in OM to reach as far as the upper cervical spinal cord, where they terminated on the lateral aspect of the intermediate gray bilaterally (Fig. 13H).
DISCUSSION Figure 14 summarizes the pathways that were delineated
in the present study, and these are discussed below. From the technical point of view, it can be noted that, although the ascendingprojections to NB were defined on the basis of both anterograde and retrograde tracing methods, the origins of these projections and the topographic organiza- tion of their terminations (in the contralateral NB) were defined primarily on the basis of somatopetal and somatofu- gal transport to and from retrogradely labeled neurons. Somatopetal-somatofugal transport to and from cell bodies with axon collaterals is a well-known phenomenon in the nervous system generally (Mesulam, 1982), but it has not been described frequently for ascending sensory relay neu- rons with specific bilateral projections. Like the results in the present study, such bidirectional transport was noted following injections into NB of the budgerigar (Hall et al., 1993) and following injections into the thalamic auditory nucleus of the pigeon (Wild, 1987). Potentially, this consti- tutes a useful “technique” for the identification of terminal fields of specific identified populations of neurons projecting bilaterally via branched axons. In fact, we would contend

318 J.M. WILD AND S.M. FARABAUGH
- 1 mm
Fig. 9. Schematic representation (left) of terminal fields in the lateral archistriatum intermedium (Ail) and lateral neostriatum cau- dale (NCI) following injections of BDA into rostral (A), middle (B), and
caudal (C) NF and NB (right). Note the terminal labeling in the lobus parolfactorius (LPO) produced by the injections. See list of Abbrevia- tions.

J.M. WILD AND S.M. FARABAUGH 319
in NCl (B) following the-injection shown in Figure‘9C Fig. 10. Photomicroaauhs ofterminal labelingin Ail (A) following the injection shown in Firmre 9A and
, , ,. I Scale bars = 150 km in A, 200 pm in B.
that, in a case such as the present one, where terminal fields (e.g., in NB) can be identified physiologically and can be targeted with greater accuracy than the subnuclei of origin, the exploitation of a neuron’s capacity to transport tracers somatopetally and somatofugally is not only fully justified but actually constitutes the method of choice for determin- ing the specific anatomical locations and extents of those terminal fields. However, whether the contralateral termi- nal field will be labeled completely will partly depend, of course, on the proportion of neurons in the nucleus of origin that project solely to the ipsilateral side. In the present study, there were always a few more PrV or LLI cells retrogradely labeled on the ipsilateral than on the contralateral side, suggesting that, in fact, there was a small proportion of cells that did not project contralaterally.
Ascending projections to NB The results show that NB receives ascending bilateral
inputs from PrV, LLI, and PLL. In addition, NB may also receive a small projection from VS. For the most part, these results generally confirm results in other species (Wallen- berg, 1903; Dubbeldam et al., 1981; Wild et al., 1985; Arends and Zeigler, 1986; Schall et al., 1986; Hall et al., 1993; Striedter, 19941, although there are interesting differ- ences, which are discussed below.
The PrV of finches is divided primarily into two major divisions, lateral and medial. In both finches and parrots, the lateral portion of PrV receives a major primary afferent projection from the tongue via the hypoglossal nerve (Wild, 1981, 1990; Bottjer and Arnold, 1982). The cell bodies of these fierents are located in the jugular ganglion together with those of the glossopharyngeal and vagus nerves, and their peripheral processes innervate the orderly arrays of tactile mechanoreceptor complexes in the tip and dorsum of the tongue. These complexes are involved principally in the species-specific feeding habits of the birds, which require a highly sensitive receptor surface for seed selection, husk- ing, and intraoral manipulation (Krulis, 1978). The tracheo- syringeal branch of the hypoglossal nerve, which contains sensory fibers innervating the syrinx (Bottjer and Arnold, 19821, does not appear to contribute to the projections to PrV (Wild, unpublished observations). Whether PrV in finches also receives a glossopharyngeal projection from the
tongue, as it does in ducks and parrots (Dubbeldam, 1980; Wild, 1981), is not known.
The present study has shown that the lateral “hypoglos- sal” portion of PrV in the zebra finch projects to the rostral part of NB, and this would seem to be the first report of a sensory hypoglossal representation in the telencephalon of a vertebrate. Interestingly, the rostral part of NB in the zebra finch is shaped like the finch’s tongue (i.e., wedge shaped; see Wild, 19901, with its base medially adjacent to the edge of the paleostriatum and with its apex protruding laterally into the neostriatum. It would be interesting to determine whether the proximodistal rows of mechanorecep- tor complexes in the tongue are mapped to lingual NB in a medial-to-lateral fashion, thereby representing their respec- tive arrangement along the length of the tongue.
The bulk of the medial part of PrV in the zebra finch is never labeled by injections of tracer into the tongue (Wild, 1990) and presumably receives its predominant input from the three branches of the trigeminal nerve that innervate the beak and buccal cavity, exclusive of the tongue, similar to all or most of PrV in pigeon and duck (Dubbeldam and Karten, 1978; Dubbeldam, 1980; Wild and Zeigler, 1995). This medial part of PrV then projects upon middle regions of NB, but it has not yet been determined whether there are specific territories of representation of each of the three trigeminal branches in NB, as there are in the mallard (Berkhoudt et al., 1981).
The projections from the distinct medial and lateral parts of PrV in the zebra finch appear to be specific to those parts, judging from the pattern of retrograde labeling, which suggests that the input from the beak and tongue is separately maintained in NB. However, it is not known whether the lateral part of PrV that receives the hypoglos- sal projection from the tongue also receives a trigeminal projection from the beak. The electrophysiological record- ings that were made from different parts of NB in the present study, however, would suggest that this is not the case, because responses to stimulation of the beak or tongue were exclusive to middle and rostral regions of NB, respec- tively. In the mallard, too, the region of PrV to which the lingual (glossopharyngeal) afferents project is not en- croached upon by trigeminal afferents (Dubbeldam, 19801, and the region of NB to which the lingual component of PrV projects is located at the rostral end of the nucleus, similar
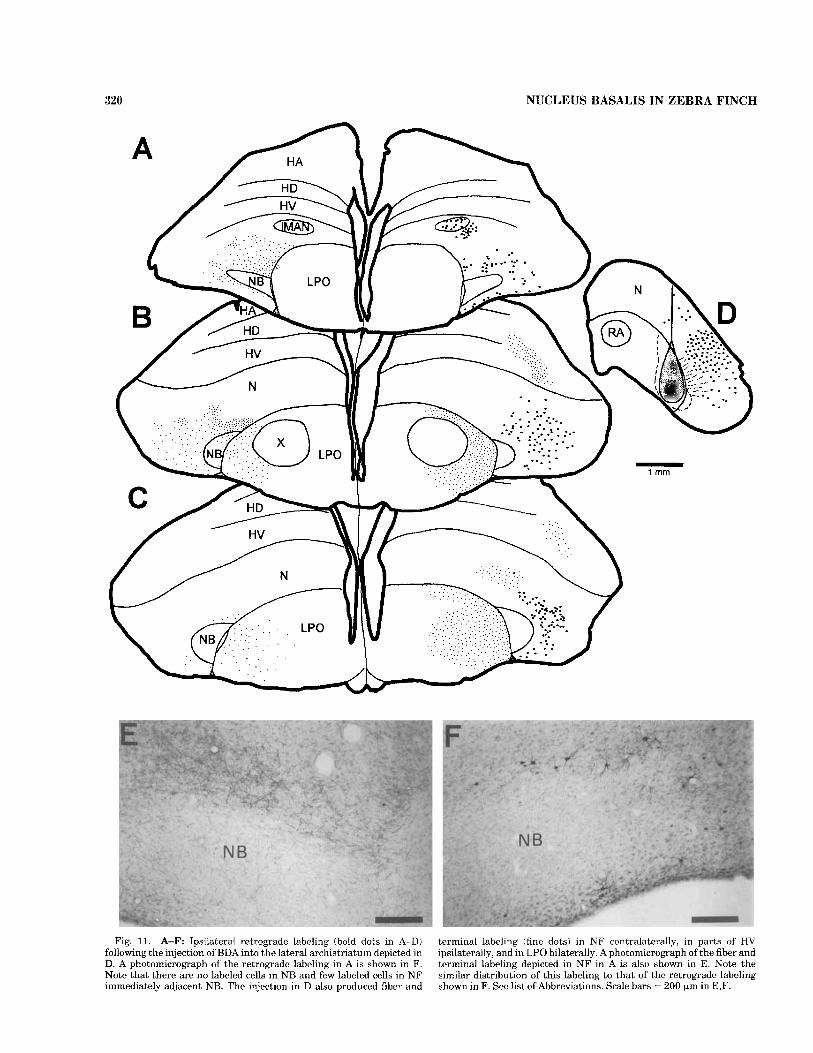
320 NUCLEUS BASALIS IN ZEBRA FINCH
A
Fig. 11. A-F: Ipsilateral retrograde labeling (bold dots in A-DI following the injection of BDA into the lateral archistriatum depicted in D. A photomicrograph of the retrograde labeling in A is shown in F. Note that there are no labeled cells in NB and few labeled cells in NF immediately adjacent NB. The injection in D also produced fiber arid
terminal labeling (fine dots) in N F contralaterally, in parts of' HV ipsilaterally, and in LPO bilaterally. Aphotomicrograph of the fiber and terminal labeling depicted in NF in A is also shown in E. Note the similar distribution of this labeling to that of thc rctrogradc labeling shown in F. See list of Abbreviations. Scale bars = 200 pm in E.F.

J.M. WILD AND S.M. FAKABAUGH 321
Fig. 12. A-E: Retrograde labeling (bold dots in A-C) in the lateral NF and HV following the injection of CTB into the NCI depicted in D. A photomicrograph of the labeling depicted in C is also shown in E. Note that there are no labeled cells in NR and few labeled cells in NF
immediately adjacent to NB. Note also that the injection into NCI produces a terminal field (fine dots) in the Ail (D) and the LPO (A-C). See list ofAbbreviations. Scale bar = 200 Km in E.

322 NUCLEUS BASALIS IN ZEBRA FINCH
to the zebra finch, and is responsive to stimulation of the tongue but not of the beak (Berkhoudt et al., 1981). As these authors point out, this organization of the representa- tions of the different oral nerves suggests that the informa- tion derived from functionally cooperative regions of the beak and tongue is not integrated at the level of NB.
Another nucleus that was retrogradely labeled from injections into beak NB was PLL. Although nothing posi- tive is known of the afferents to this nucleus, its location close to LLIr might suggest that it receives an auditory input like the lateral lemniscal nuclei in other species (Leibler, 1975; Conlee and Parks, 1986; Takahashi and Konishi, 1988); but, if this is the case, then this input was not readily detectable in evoked potentials or in multiunit recordings from the beak part of NB in the present study. Furthermore, no retrogradely labeled cells were ever found in PLL following injections into auditory NB. It is known that PLL in the zebra finch does not receive a primary afferent input from the tongue (Wild, 1990), and this finding is consistent with retrograde labeling in PLL from the beak rather than from the lingual part of NB. The evidence suggests, therefore, that PLL is related to the trigeminal system, but further work is required to confirm this.
Unlike PLL, LLI (both rostra1 and caudal parts) was retrogradely labeled following injections into auditory NB, and these results confirm those found in starling (Kirsch, 1983), pigeon (Arends and Zeigler, 1986; Schall et al., 1986; Maekawa, 1987), and budgerigar (Hall et al., 1993; Striedter, 1994). Because LLI in the zebra finch receives a projection from the cochlear nucleus angularis and possibly from nucleus laminaris as well (Wild, unpublished observations), it appears to be the source of the short latency auditory projection to caudal NB, as was suggested on electrophysi- ological grounds in pigeon and in other species before the anatomy was known (see Delius et al., 1979).
A final source of possible brainstem projections to the most dorsocaudal region of NB and medially adjacent neostriatum is a specific, small, lateral part of VS. Follow- ing injections of tracer into NB, retrogradely labeled VS neurons were first reported in pigeon by Schall et al. (1986), and, although their exact distribution is difficult to deter- mine from the figures, they were depicted as lying in a plane 3.00 mm lateral to the midline. According to the atlas of Karten and Hodos (19671, this locates them on the lateral margin of VS, where the labeled VS cells were also located in the zebra finch. The input to the lateral part of VS in the zebra finch is unknown, but, in pigeon, a specific projection from the lagenar nerve terminates on the quadrangular division of VS, a division that is located on the lateral aspect the nucleus (Boord and Karten, 1974). Thus, the VS cells projecting to NB in pigeon and in zebra finch could conceiv- ably mediate lagenar input, but the specific nature of this input has not been determined. In pigeon, evoked potentials in NB have been recorded following both electrical stimula- tion of the sagittal semicircular canal (Schall and Delius, 1986) and rotation of the head about the transverse axis, or pitch (Schall, 1987). This not only confirms the presence of a vestibular projection to NB; it also suggests that, at least in pigeon, there are other VS regions in addition to the quadrangular division that are involved in relaying vestibu- lar information to NB, because these other regions receive inputs from other parts of the vestibular apparatus, such as the maculae of the semicircular ampullae (Wilson and Felpel, 1972; Wold, 1975).
Telencephalic projections to NB From within the telencephalon, NB receives a projection
from the ipsilateral HV, with which NB appears to be reciprocally connected. Such an arrangement is also pre- sent in the mallard (Dubbeldam and Visser, 1987) and the goose (Veenman and Gottschaldt, 19861, but it was not reported in the pigeon (Wild et al., 1985). The NB-HV connection is difficult to interpret meaningfully, partly because it is not clear what telencephalic structure or cell group in mammals is homologous with HV in birds. It can be noted, however, that HV is one of the few telencephalic sources of input to primary sensory areas of the telencepha- lon in birds. Because these primary sensory areas are considered to be homologous with specific populations of neocortical lamina IV neurons in mammals (Karten, 1969; Karten and Shimizu, 19891, it may be that specific popula- tions of HV neurons are comparable to one or more supragranular neocortical lamina that project upon lamina IV. This assumes, of course, that NB itself is a telencephalic nucleus, an assumption that has been challenged in the past (Cohen and Karten, 1974). Aresolution of the question of whether NB should be considered a diencephalic or a telencephalic structure will probably have to wait for a clearer understanding of the developmental origins of specific nuclear groups in the avian forebrain.
Efferents of NB Like the results in a similar study in pigeon (Wild et al.,
1985), terminal fields in NF resulting from injections into NB were often difficult to identify or to interpret in the zebra finch because of the narrow width of NB in the transverse plane and due to the fact that it lies adjacent to NF. Thus, the injection and the terminal field were not well separated. In the larger brained mallard and goose, NB was shown to project upon the adjacent ventral layer of the neostriatum frontale (NF,v; Veenman and Gottschaldt, 1986; Dubbeldam and Visser, 19871, a region that probably corresponds with the layer of radially aligned NF neurons surrounding NB in the zebra finch. Injections placed in NF lateral to NB in the present study consistently resulted in retrogradely labeled cells in NB, thus confirming the NB-NF projection. However, the zebra finch data suggest that NB projects not only to the inner NF layer of radially aligned neurons but also to the outer layer of NF as well, although this conclusion must remain tentative in view of the technical problems noted at the beginning of this section. In the mallard, NB appears to project only to NF,v, with NF,v then providing the projection to the overlying dorsal layer of NF (NF,d; Dubbeldam and Visser, 1987).
These short axonal projections from NB to the surround- ing NF are presumably those that mediate the short latency auditory projection to the NF region that have been re- ported for several species of birds (Harman and Philips, 1967; reviewed in Delius et al., 1979). However, the audi- tory region that was found in NF of starling by Kirsch et al. (1980) had a latency that was too long (13-20 msec) to be considered part of this system and deserves further atten- tion.
NF efferents Three terminal fields were observed following injections
into NF, a relatively sparse and diffuse one in the ventrolat- era1 LPO, a small and nuclear specific one in Ail, and a large, diffuse, and dense one in NC1. The first terminal field

NUCLEUS BASALIS IN ZEBRA FINCH
avoided area X completely, suggesting that this projection is not concerned with direct access to the “recursive loop” of song-learning nuclei (Bottjer et al., 1989; Williams, 1989). However, the lobus parolfactorius is a significant source of input to the midbrain tegmentum via descending basal ganglia pathways, and there are also some projections upon the ventral area of Tsai (Kitt and Brauth, 1981). Because the ventral area of Tsai projects upon area X in the zebra finch (Lewis et al., 19811, there could be some modulation of ascending inputs to the song system via descending projec- tions originating in NF. The second and third terminal fields are the end stations of caudally directed components of the FAT. The specific terminal field in Ail is similar to that described in the pigeon (Wild et al., 1985) and mallard (Dubbeldam and Visser, 1987); but, in the zebra finch, the use of small BDA injections into different rostrocaudal parts of N F has allowed us to delineate a topography in the projections of NF to the archistriatum that was not ob- served in either pigeon, goose, or mallard (Wild et al., 1985; Veenman and Gottschaldt, 1986; Dubbeldam and Visser, 1987). In the present study, we have shown that, in addition to a trigeminal contribution to Ail from the beak part of NB, there are contributions both from a more rostra1 tongue or hypoglossal part of NB and from a more caudal auditory part of NB, these parts relaying in their adjacent NF regions. These same NF regions also project to NCI, where the pattern of termination in Ail is duplicated; i.e., here are distinct but partly overlapping terminal fields. Again, although N F projections to the NCI have been reported in pigeon and in mallard, their topographic organi- zation has not been observed previously. We do not have clear evidence, however, that shows whether the different NC1 regions project topographically back to Ail, although Ail is certainly a target of these NCI regons, as it is in pigeon (Wild et al., 1985).
A novel finding in the present study was the presence of a terminal field in the contralateral NF following injections of BDA in Ail. The location of this field generally resembled that of the origins of the ipsilateral N F projection to Ail. Although the putative NF-Ail projection was not confirmed by injections of BDA into NF, we feel that it is likely to be a real projection because of the absence of any evidence of projections to NF from any region adjacent or near to Ail, including NC1. Possibly, the absence of retrograde confirma- tion could be explained technically by the fact that rela- tively small iontophoretic injections of RDA were made into the region of a diffuse terminal field. Ail was also found to project to the contralateral NC1, and this projection was confirmed by the presence of retrogradely labeled cells in Ail following injections of either CTB or BDA into NCI. Cells in a similar location in the Ai of pigeon were retro- gradely labeled (bilaterally) by injections of horseradish peroxidase in the Wulst (Bagnoli and Burkhalter, 19831, suggesting that ventrolateral regions of the Ai of aves are a major source of the widespread projections to the contralat- era1 telencephalic hemisphere originally described as a component of the anterior commissure by Zeier and Karten (1971, 1973). The specific function of these projections is unknown, but their organization naturally would suggest a general role in bilateral integration by feedback modulation of sensory activity. In this respect, it is interesting to note that transection of the anterior cornmissure has been reported to reduce the amplitude of auditory evoked poten- tials recorded near NB in the frontal forebrain by 90% (see Delius et al., 1979).
323
The Ail in the zebra finch is the source of a major bilaterally descending pathway with extensive terminations in the posterior thalamus and brainstem as far as the upper cervical spinal cord. The extent to which the origins of this pathway precisely coincide with the region of the lateral archistriatum that receives projections from the ipsilateral NF and NC1 (viz. Ail) and that projects contralaterally upon them is not yet known. It may well be that the cells of origin of the descending pathway described in the present report include more than Ail. This pathway is a component of the occipitomesencephalic tract (OM), and it crosses not only via the anterior commissure before descending, but it continues to cross throughout its extent at several levels of the brainstem, particularly through the commissura infima of the closed medulla. Its most conspicuous and dense terminal field at thalamic levels is in the ventral or medial portion of the SpM, a nucleus that, in catfish and in pigeon, projects upon the cerebellum and, in pigeon, projects upon the red nucleus, but it has no obvious counterpart in mammals (Karten and Finger, 1976; Wild, 1992). At pon- tine levels, the tract appears for the most part not to terminate on the trigeminal or facial motor nucleus; thus, its influence on jaw motoneurons is presumably largely via interneurons, which was thought to be the case in pigeon and in duck (Arends and Dubbeldam, 1982; Berkoudt et al., 1982). However, this conclusion must remain somewhat tentative in view of the facts that a few terminations were observed in these nuclei and that another archistriatal nucleus (viz. nucleus robustus) projects directly upon an- other cranial nerve motor nucleus (viz. the tracheosyrin- geal motor nucleus of NXII) in songbirds, including the zebra finch (Wild, 1993b). At medullary levels, the Ail projections are preferentially directed into the lateral parts of the reticular formation and do not terminate upon either the lingual or the tracheosyringeal portions of the hypoglos- sal nucleus. Any influence the projections may have on vocal tract structures, therefore, must also be mediated via interneurons. In its course and terminations, the tract labeled by injections into Ail resembles that described by Zeier and Karten (1971) in pigeon as being similar to the bundle of Bagley of ungulates (Haartsen and Verhaart, 1967). It is distinctly different from the occipitomesence- phalic tract. pars hypothalami (HOM), which arises from the caudal and the most medial parts of the archistriatum, has major projections upon the medial hypothalamus, and lacks a more caudal trajectory (Zeier and Karten, 1971). But it is also distinctly different from tracts that arise from other parts of A, such as that arising from its ventromedial part in pigeon (Aivm; Wild et al., 1993) and that arising from the medially situated RA in songbirds, including the zebra finch (Wild, 1993b). This latter tract makes up a medial component of OM, has a distinct terminal field in the dorsomedial nucleus of the intercollicular region, and has major projections upon glossopharyngeal (laryngeal) and hypoglossal (tracheosyringeal) cranial nerve motor nuclei and premotor neurons involved in the control of vocalization and respiration (Nottebohm et al., 1976; Vi- cario, 1993; Wild, 1993a,b). Clearly, the archistriatum of birds gives rise to several distinct descending pathways that target largely nonoverlapping sets of nuclei involved in a variety of different behaviors.
Hodology of NB and vocalization A primary motivation for the present study was to
confirm the presence and location in a songbird of an

324 J.M. WILD AND S.M. FARARAUGH
Figure 13

NUCLEUS BASALIS IN ZEBRA FINCH 325
C
Fig. 13. A-H: Charts of the anterograde projections (fine dots) and retrogradely labeled cells (bold dots) resulting from thc injcction of BDA into Ail depicted in C (the same injection is shown in Fig. 1 In). The very dense and specific projection to the ventral or medial part of
the ipsilateral mcdial spiriform nucleus (SpM) is shown in photomicro- graph b, which corresponds to the level of B. See list of Abbreviations. Scale bar = 200 pm in b.

326 J.M. WILD AND S.M. FARABAUGH
Fig. 14. Schematic diagram summarizing the connections delin- eated in the present study. See list of &4bbreviations.
auditory component of NB (Kirsch, 1983) and to determine whether the intra- or extratelencephalic connections of NR had any obvious links with the vocal system. It was found that, in the zebra finch, NB definitely has an auditory component located in the caudal part of the nucleus, but obvious links of the output of any part of NB with any part of the vocal control system (as it is usually conceived) were not identified. That is, there were no direct or indirect connections with other auditory regions of the telencepha- lon, such as field L, with the high vocal center (previously known as the hyperstriatum ventrale, pars caudalis or HVc) or the RA in the telencephalon, and there were no projec- tions upon respiratory-vocal nuclei of the brainstem (Wild, 1994). However, in view of recent evidence implicating nonsyringeal components of the vocal tract (namely, the beak and possibly the tongue) in vocal production both in songbirds and in nonsongbirds (Hausberger et al., 1991; Westneat et al., 19931, our current concept of “the vocal system” is likely to need considerable expansion. This is because, in both songbirds and nonsongbirds, there is a marked correlation between the size of the gape and sound frequency; therefore, it is clear that, in many birds, vocaliza- tion involves a control of the jaw and possibly the tongue musculature. (In fact, the involvement of the beak in singing and a correlation of gape size with frequency are obvious upon careful inspection of a number of songbirds.) Neural control of the avian jaw has been the subject of intensive investigation over many years, but this investiga- tion has been concerned largely with the trigeminal sensori- motor control of feeding in pigeon and in duck (Zeigler, 1989; Dubbeldam and Den Boer-Visser, 1994; Zeigler et al.,
1994). Lesions of NB in pigeons disrupt both the responsive- ness to food and the organization of the ingestive response sequence (involving grasp, stationing, and intraord trans- port components; Zeigler and Karten, 19731, and these effects are mediated via intra- and extratelencephalic tri- geminal sensorimotor circuitry (Berkhoudt et al,, 1982; Wild et al., 1985; reviews in Zeigler, 1989; Zeigler et al., 1994). Similar pathways delineated in the present study that originate in NB and that eventually terminate in the rhombencephalic lateral reticular formation are likely to be involved in the control of feeding in the zebra finch, but, in songbirds, they may also be implicated in the sensorimotor control of vocalization by their possible influence on jaw and tongue motoneurons. Furthermore, it does not seem unreasonable to suggest that an auditory input to this circuitry could play a significant part in this control, although other interpretations of the functional impor- tance of the auditory input to NB are possible (Schall and Delius, 1986). Thus, whereas the neural control of feeding may involve primarily trigeminal sensorimotor circuitry (and other sensory inputs from the tongue that are routed through PrV in different species), singing in songbirds may involve a dual sensory control of the jaw and tongue by trigeminal (and other orosensory, e.g., lingual, inputs routed through PrV) and auditory inputs. Evidence for this sugges- tion is provided by a report that lesions of NB disrupt vocalization in the budgerigar (Hall et al., 1993). However, these authors interpreted their lesion-induced deficits solely in terms of the effects on the auditory component of NB. The possibility of a contribution to these effects from lesions of the lingual or trigeminal components either was not considered or was discounted, respectively, in view of the authors’ assumption that song production is generally thought to be a sole function of syringeal and respiratory activities. In fact, the study of Hall et al. (1993) did not distinguish between the various components of NB; thus, further studies are required to assess the possible different roles of these components in the vocalizations of both the budgerigar and songbirds. The present study has provided the necessary anatomical foundation for such a neurobehav- ioral study in the zebra finch.
Finally, with reference to Striedter’s (19941 suggestion that songbirds and psittacines have evolved their vocal control systems independently, the present results permit a more detailed comparison of the intratelencephalic connec- tions of NB in zebra finch and budgerigar than has been possible hitherto. One difference that was not appreciated previously is that the terminations in the archistriatum that are such a consistent feature of the projections of NF in zebra finch (and in pigeon and duck) have not been found in budgerigar. However, NF in budgerigar does project upon a region of the lateral hemisphere called by Striedter the supracentral nucleus of the lateral neostriatum (NLs; Striedter, 1994), which, in turn, projects upon the archistria- tum, as NF does in zebra finch, via NCl. Admittedly, the regions of archistriatum that receive the projections from NLs or from NC1 in the two species are very different from each other topographically, which is indicated by the fact that, in budgerigar, Striedter (1994) called the archistriatal nucleus in question the medial nucZeus of the anterior nrchistriatum (AAm). However, the conformation of the archistriaturn and its relations to the surrounding neostria- tum in psittacines are so different from those in passerines that comparisons of nuclei based on relative positions are rendered highly questionable. Clearly, a more meaningful

NUCLEUS BASALIS IN ZEBRA FINCH 327
comparison of AAm and Ail can only be made when the efferent projections of AAm are known. If they turn out to be similar, then the superficial similarity of NLs of budgeri- gar to NC1 of zebra finch would take on added significance, especially when it is realized that the projections of NLs to AAm of the archistriatum in budgerigar (as depicted by Striedter, 1994) are heavier than those to the central nucleus of the lateral neostriatum (NLc), which provides the input to the vocal system. In fact, the weakest link in the chain of projections joining auditory NB with the vocal system in budgerigar (as depicted by Striedter, 1994) is the link between NLs and NLc, because the dense terminal field that would be expected in NLc following injections of tracer into NLs was conspicuous by its absence, as were clusters of retrogradely labeled cells in NLs following injections of tracer into NLc. Therefore, we believe that the hypothesis that the auditory input to the vocal system in budgerigars is provided primarily, if not exclusively, via projections of NB and NF (Striedter, 1994) should remain a hypothesis pending the provision of further evidence of a substantial projection from NLs to NLc. These reservations notwith- standing, there are so many other differences between the vocal control systems of budgerigar and songbirds (as cogently documented by Striedter, 1994) that Striedter’s more general hypothesis of the independent evolution of those systems seems justified.
ACKNOWLEDGMENTS This work was supported by a grant to J.M.W. from
Whitehall Foundation, Inc. The authors thank Han-Xu Liu for histological and technical assistance.
LITERATURE CITED Arends, J.J.A., and J.L. Dubbledam (1982) Exteroceptive and proprioceptive
afferents of the trigeminal and facial motor nuclei in the mallard (Anus plnfyrhynchos LJ. J. Comp. Neurol. 209:313-329.
Arends, J.J.A., and H.P. Zeigler (1986) Anatomical identification of an auditory pathway from a nucleus of the lateral lemniscal system to the frontal telencephalon (nucleus basalis). Brain Res. 398r375-381.
Bagnoli, P., and A. Burkhalter (1983) Organization of the afferent projec- tions to the wulst in the pigeon. J. Comp. Neurol. 214r103-113.
Ball, G.F. (1990) Chemical neuroanatomical studies of the steroid-sensitive songbird vocal control system: A comparative approach. In J. Balthazart (ed.!: Hormones, Brain and Behavior in Vertebrates. 1. Sexual Differen- tiation, Neuroanatomical Aspects, Neurotransmitters and Neuropep- tides (Comp. Physiol., Vol. 81. Basel: Karger, pp. 148-167.
Ball, G.F., and J.M. Castro (1991) Autoradiographic localization of NMDA receptors in the avian song control system using [’HI MK-801. Soc. Neurosci. Abstr. 17~1053.
Berkhoudt, H., J.L. Dubbeldam, and S. Zcilstra (19813 Studies on the somatotopy of the trigeminal systcm in the mallard, Anasplatyrhynchos I,. IV. Tactile representation in the nucleus basalis. J. Comp. Neurol. 196:407420.
Berkhoudt, H., B.G. Klein, and I1.P. Zeigler (1982) Afferents to the trigeminal and facial motor nuclei in the pigeon iCulumba l i v i ~ L.): Central connections ofjaw motoneurons. J. Comp. Neurol. 209r301-312.
Boord, R.L., and H.J. Karten (1974) The distribution of primary lagenar fibers within the vestibular nuclear complex of the pigeon. Brain Behav. Evol. 10~228-235.
Bottjer, S.W., and A.P. Arnold (1982) Afferent neurons in the hypoglossal nerve of the zebra finch (Poephcla guttutu): Localization with horserad- ish peroxidase. J. Comp. Neurol. 21Or190-198.
Bottjer, S.W., S.A. Halscma, S.A. Brown, and E.A. Miesner (1989) Axonal connections of a forebrain nucleus involved with vocal learning in zebra finches. J. Comp. Neurol. 279~312326.
Carr. R.A., and J.M. Wild (1984a) Cranial nerve afferents to the principal sensory trigeminal nucleus in the galah (Cacatua roseicupilla). Neurosci. Lett. (Suppl.) 15r824.
Carr, R.A., and J.M. Wild (1984b) Sensory trigeminal innervation of the tongue of the galah ( C Q C Q ~ ~ U roseicapilla). Soc. Neurosci. Abstr. 10:lll.
Cohen, D.H., and H.J. Karten (1974) The structural organization of the avian brain: An overview. In I.J. Goodman and M.W. Schein (eds.): Birds, Brain and Behavior. New York Academic Press, pp. 29-73.
Conlee, J.W., and T.N. Parks (1986) Origin of ascending auditory projections to the nucleus mesencephalicus lateralis, pars dorsalis in the chicken. Brain Res. 367:96-113.
Cords, E. (1904) Beitrage zur Lehre vom Kopfnervensystem der Vogel. Z. Anat. Rntwick. 26:49-100.
Deich, J.D., D. Houben, R.W. Allan, and H.P. Zeigler (19851 “On-line” monitoringofjaw movements in the pigeon. Physiol. Behav. 3a5:307-311.
Delius, J.D., T.E. Runge, and H. Oeckinghaus (1979) Short-latency auditory projection to the frontal telencephalon of the pigeon. Exp. Neurol. 63~594-609.
Dubbeldam, J.L. (1980) Studies on the somatotopy of the trigerninal system in the mallard, Anus placyrhynchos L. 11. Morphology of the principal sensory nucleus. J. Comp. Neurol. 291.357-571.
Dubbeldam, J.L., and A.M. Den Boer-Visser (1994) Organization of “feeding circuits” in birds: Pathways for the control ofbcak and head movements. Eur. J. Morphol. 32r127-133.
Dubbeldam, J.L., and H.J. Karten (1978) The trigeminal system in the pigeon (Columba lzuza). I. Projections of the gasserian ganglion. J. Comp. Neurol. 180:661-678.
Dubbeldam, J.L., and A.M. Visser (1987) The organization of the nucleus hasalis-neostriatum complex in the mallard (Anasplatyrhynchos L.) and its connections with the archistriatum and paleostriatum complex. Neuroscience 21:487-517.
Dubbeldam, J.L., E.R. Brus, S.B.J. Menken, and S. Zeilstra (1979) The central projections of the glossopharyngeal and vagus ganglia in the mallard, Anasplatyrhynchos L. J. Comp. Neurol. 183:14%168.
Dubbeldam, J.L., C.S.M. Brauch, and A. Don (1981) Studies on the somatotopy of the trigeminal system in the mallard, Anas platyrhynchos L. 111. Afferents and organization of the nucleus basalis. J. Comp. Neurol. 196r391-405.
Felix, B., and ’r. Roesch (1986) Telencephalic bill projections in the Landes goose. Somatosens. Res. 4:141-152.
Fortune, E.S., and D. Margoliash (19Y2j Multiple auditory pathways into HVc. Soc. Neurosci. Abstr. 18~1193.
Gahr, M., H.-R. Guttinger, and D.E. Kroodsma (1993) Estrogen receptors in the avain brain: Survey reveals general distribution and forebrain areas unique to songbirds. J. Comp. Neurol. 327:112-122.
Haartsen, A.B., and W.J.C. Verhaart (1967) Cortical projections to the brainstem and spinal cord in the goat by way of the pyramidal tract and bundle of Bagley. J. Comp. Neurol. 129r189-202.
Hall, W.S., P.L. Cohen, and S.E. Brauth (1993) Auditory projections to the anterior telencephalun in the budgerigar (Meiopszttacus undulatus). Brain Behav. Evol. 41:97-116.
Hall, W.S., S.E. Brauth, and J.T Heaton (1994) Comparison of the effects of lesions in nucleus basalis and field “L” on vocal learning and perfor- mance in the budgerigar (Melopsittams undulatusj. Brain Behav. Evol. 44r133-148.
Harman, A.L., and R.E. Philips (1967) Responses in the avian midbrain, thalamus and forebrain evoked by click stimuli. Exp. Neurol. 18:276-286.
Hausberger, M., J.M. Black, and J. Richard (1991) Bill opening and sound spectrum in barnacle goose loud calls: Individuals with “wide mouths” have higher pitched voxes. Anim. Behav. 42:319-~322.
Karten, H.J. (1968) The ascendingauditorypathway in the pigeon (Columba liuia) 11. Telencephalic projections of the nucleus ovidalis thalami. Brain Kes. 11~134-153.
Karten, H.J. (19691 The organization of the avian telencephalon and some speculations on the phylogeny of the amniote telencephalon. Ann. N.Y. Acad. Sci. 167:164-179.
Karten, H.J., and T.E. Finger (1976) A direct thalamo-cerebellar pathway in pigeon and catfish. Brain Res. 10Zr335-338.
Karten, H.J.. and W. Hodos (1967) A Stereotaxic Atlas of the Brain of the Pigeon (Columba hocaJ. Baltimore: Johns Hopkins Press.
Karten, H.J., and T. Shimizu (1989) The origins of the neocortex: Connec- tions and lamination as distinct events in e~olution. J. Cogn. Neurosci. 1.291-301.
Kirsch, M. I 19831 Antworteigenschaften und Verbindungen des akustischen Zentrums mit kurzer Latenz im frontalen Vorderhirn des Staren. Verh. Deutsch. 2001. Gcs. 296396.

328 J.M. WILD AND S.M. FARABAUGH
Kirsch, M., R.B. Coles, and H.-J. Leppelsack (19801 Unit recordings from a new auditory area in the frontal neostriatum o f the awake starling (Sturnus uuigaris:. Exp. Brain Res. 38:375-380.
Kitt, C.A., and S.E. Brauth (1981) Projections ofthe paleostriatum upon the midbrain tegmentum in the pigeon. Neuroscience 6:1551-1556.
Konishi, M. (1989) Birdsong for neurobiologists. Neuron 3:541-549. Krulis, V. (1978) Struktur und Verteilung von Tastrezeptoren im Schnabel-
Zungenbereich vim Singvogeln, im besondern der Fringillidae. Rev. Suisse Zool.85:385-447.
Leibler, L.M. (1975) Monaural and Binaural Pathways in the Ascending Auditory System of the Pigeon. Ph.D. Dissertation, Massachusetts Institute nf Technology.
Lewis, J.W., S.M. Ryan, A.P. Arnnld, and L.L. Butcher (1981) Evidence for a catecholaminergic projection to area X in the zebra finch. J. Comp. Neurol. 196:347-354.
Maekawa, M (1981) The early auditory response in the frontal forebrain of the pigeon. J. Physiol. SOC. Jpn. 43358.
Maekawa, M. (1987) Auditory responses in the nucleus basalis of the pigeon. Hearing Res. 27:231-237.
Margoliash, D., E.S. Fortune, M.L. Sutter, A.C. Yu, B.D. Wrenhardin, and A. Dave (1994) Distributed representation in the song system of oscines: Evolutionary implications and functional consequences. Brain Behav. Evol. 4-5:247-264.
Mesulam, M.-M. (1982) Tracing Neural Connections with Horseradish Peroxidase. Chichester: John Wiley and Sons.
Nottebohm, F., T.M. Stokes, and C.M. Leonard (1976) Central control of song in the canary, Serinus canaria. J. Comp. Neurol. 207:344-357.
Nowicki, S., and P. Marler (1988) How do birds sing? Music Percept. 5:391-426.
Shal l , U. (1987) Vestibular, olfactory, and vibratory responses of nucleus basalis prosencephali neurons in pigeons. Neurosci. Res. 4:376-384.
Schall, U., and J.D. Delius (1986) Sensory inputs to the nucleus basalis prosencephali, a feeding-pecking ceni.re in the pigeon. J. Comp. Physiol. 159:3341.
Schall, U., 0. Gunturkun, and J.D. Delius (1986) Sensory projections to the nucleus basalis prosencephali of the pigeon. Cell Tissue Res. 245:539- 546.
Striedter, G.F. (1994) The vocal control pathways in budgerigars differ from those in songbirds. J. Comp. Neurol. 343:35-56.
Takahashi, T.T., and M. Konishi (1988) Projections of nucleus angularis and nucleus laminaris to the lateral lemniscal nuclear complex of the harn owl. J. Comp. Neurol. 274:212-238.
Veenman, C.L., and K.-M. Gottschaldt (1986) The nucleus basalis- neostriatum complex in the goose (Anser anser L.). Adv. Anat. Embryol. Cell Bid. 96:l-85.
Veenman, C.L., A. Ileiner, and M.G. IIonig (1992) Biotinylated dextran amine as an anterograde tracer for single- and double-labeling studies. J. Neurosci. Methods 41:239-254.
Vicario, D.S. (1993) A new brainstem pathway for vocal control in the zebra finch song system. Neuroreport. 4:983-986.
Wallenberg, A. (19033 Der Ursprung des tractus isthmo-striatus (oder bulbo-striatus) der Taube. Neurol. Zcntralb. 22:9%101.
Westneat, M.W., J.H. Long Jr., W. Hoese, and S. Nowicki (1993) Kinematics of birdsong: Functional correlation of cranial movements and acoustic features in sparrows. J. Exp. Bid. I82:147-171.
Wild, J.M. (1981) Identification and localization of the motor nuclei and sensory projections of the glossopharyngeal. vagus, and hypoglossal nerves of the cockatoo (Cacatua ruseicapilla), Cacatuidae. J. Comp. Neurol. 20.?:351-377.
Wild, J.M. (1987) Nuclei of the lateral lemniscus project directly to the thalamic auditory nuclei in the pigeon. Brain Res. 408:303-307.
Wild, J.M. (1990) Peripheral and central terminations of hypoglossal afferents innervating lingual tactile mechanoreceptor complexes in FrzngiZZidae. J. Comp. Neurol. 298:157-171.
Wild, J.M. (7992) Direct and indirect “cortico”-rubral and rubro-cellular cortical projections in the pigeon. J. Comp. Neurnl. 326S23-636.
Wild, J.M. (1993a) The avian nucleus retroambigualis: A nucleus for breathing. singing and calling. Brain Res. 606:319-324.
Wild, J.M. i1993b) Descending projections of the songbird nucleus robustus archistriatalis. J. Comp. Neurnl. d38:225-241.
Wild, J.M. (1994) The auditory-vocal-respiratory axis in birds. Brain Behav. Evol. 44:192-209.
Wild, J.M., and H.P. Zeigler (1980) Central representation and somatotopic organization of the jaw muscles within the facial and trigeminal nuclei of the pigeon (Colurnba lzuia). J. Comp. Neurol. 192175-201.
Wild, J.M., and H.P. Zeigler (1996) Central projections and somatotopic organization of trigeminal primary afferents in pigeon (Cnlumba liuia). J. Comp. Neurol. (in press).
Wild, J.M., J.J.A. Arends, and H.P. Zeigler (1985) Telencephalic connections of the trigeminal system in the pigeon (Colunba liiiia): A trigeminal sensorimotor circuit. J. Comp. Neurol. 234:441-464.
Wild, J.M., H.d. Karten, and B.J. Frost (1993) Connections of the auditory forebrain in thr pigeon (CnZurnha liuia). J. Comp. Neurol. 337~32-62.
Williams, H. (1989) Multiple representations and auditory-motor interac- tions in the avian song system. Ann. N.Y. Acad. Sci. 56‘3:14%164.
Wilson, V.J., and L.P. Felpel (1972) Specificity of semicircular input to neurons in the pigeon’s vestibular nuclei. J. Neurophysiol. 35253-264.
Witkovsky, P., R. Silver, and H.P. Zeigler 1,1973) The nucleus basalis of the pigeon: A singlc unit analysis. J. Comp. Neurol. 147:119-128.
Wold, J.E. (19753 The vestibular nuclei in the domestic hen (GaZZns dom~stieus). 11. Primary afferents. Brain Kes. 95:531-543.
Zeier, H.. and H.J. Karten (1971) The archistriatum of the pigeon: Organiza- tion of afferent and efferent connections. Brain Res. 31:313-326.
Zeier, H., and H.J. Karten (1973) Connections of the anterior commissure in the pigeon (Colurnba liuia). J. Comp. Neurol. 150:201-216.
Zeigler, H.P. (1989, Neural control of the jaw and ingestive behavior: Anatomical and behavioral studies of a trigeminal sensorimotor circuit. Ann. N.Y. Acad. Sci. 563:69-86.
Zeigler, H.P., and H.J. Karten (1973) Brain mechanisms and feeding behavior in the pigeon (Colzrmba liuia). Quintn-frontal structures. J. Comp. Neurol. I,52:59-81.
Zeigler, H.P., R. Bermejo, and R. Bout (1994) Ingestive behavior and the sensorimotor control of the jaw. In M.N.O. Davis and P.R. Green (eds.): Perception and Motor Control in Birds. Berlin: Springer-Verlag, pp. 182-200.