oncogenic bystander radiation effects in ...sirr2.it › uploads › agosto2008.pdfsiblings showed...
TRANSCRIPT

Volume XI n. 2
Periodico della Società Italiana per le Ricerche sulle Radiazioni
Agosto 2008
ONCOGENIC BYSTANDER RADIATIONEFFECTS IN PATCHED HETEROZYGOUSMOUSE CEREBELLUM
MODELLIZZAZIONE DELL’EFFETTO BYSTANDER:COME LA RADIAZIONE MODULA LA COMUNICAZIONE CELLULARE
MOVIMENTI INTRA-FRAZIONE IN TRATTAMENTI DI PROTONTERAPIA CON“SPOT-SCANNING”: EFFETTI DOSIMETRICI E METODI DI COMPENSAZIONE
CITOTOSSICITÀ DI UNA VARIANTE ISOFORME DI MANGANESE SUPEROSSIDO DISMUTASI E SUA INTERAZIONE CON LARADIOSENSIBILITÀ IN CELLULE UMANE
Pos
teIta
liane
S.p
.A.-
Spe
d.in
Abb
.Pos
tale
-D.L
.353
/200
3(C
onv.
inL.
27/0
2/20
04n.
46)a
rt.1
com
.2-D
CB
-Rom
a

2
RADIAZIONI Ricerca e Applicazioni Anno XI n. 2/2008
SOMMARIORadiazioni Ricerca e ApplicazioniPeriodico della Società Italianaper le Ricerche sulle RadiazioniPubblicazione Periodica QuadrimestraleAgosto 2008 - Vol. XI n. 2
Direttore ResponsabileFrancesca BallariniDipartimento di Fisica Nucleare e TeoricaUniversità di Paviae-mail: [email protected]
Direttore EditorialeRaffaele De VitaSezione Tossicologia e Scienze BiomedicheENEA Centro Ricerche Casaccia, Romae-mail: [email protected]
Capo RedattoreLorenzo MantiDipartimento di FisicaUniversità Federico II, Napolie-mail: [email protected]
Comitato di RedazioneMaurizio AmichettiAgenzia Provinciale per la ProtonTerapiaTrentoe-mail: [email protected]
Francesca AntonelliDipartimento Tecnologia e SaluteIstituto Superiore di Sanità, Romae-mail:[email protected]
Mauro BonardiUniversità degli Studi di Milanoe-mail: [email protected]
Martino GrandolfoDipartimento Tecnologie e SaluteIstituto Superiore di Sanità, Romae-mail: [email protected]
Per Informazioni e CorrispondenzaFrancesca BallariniTel. 0382 987949Tel. 02 50317399 Fax 02 50317630e-mail: [email protected]
Editrice: Società Italiana per le Ricerche sulle Radiazioni
Registrazione del Tribunale di Roma n. 406 del 6 Agosto 1998
Grafica: Renato Cafieri
Stampa:
Pubblicità:
ONCOGENIC BYSTANDER RADIATION EFFECTSIN PATCHED HETEROZYGOUS MOUSE CEREBELLUM 3Mariateresa Mancuso and Anna Saran
MODELLIZZAZIONE DELL’EFFETTO BYSTANDER: COME LA RADIAZIONE MODULALA COMUNICAZIONE CELLULARE 6Luca Mariotti
MOVIMENTI INTRA-FRAZIONE IN TRATTAMENTI DI PROTONTERAPIA CON “SPOT-SCANNING”: EFFETTI DOSIMETRICI EMETODI DI COMPENSAZIONE 9Silvia Tomaselli, Marco Schwarz
CITOTOSSICITÀ DI UNA VARIANTE ISOFORMEDI MANGANESE SUPEROSSIDO DISMUTASI ESUA INTERAZIONE CON LA RADIOSENSIBILITÀIN CELLULE UMANE 13Maria Rosaria Rusciano
Redazione EditorialeSocietà Italiana per le Ricerche sulle Radiazioni Sezione Tossicologia e Scienze BiomedicheENEA Centro Ricerche Casaccia, s.p. 016Via Anguillarese, 301 - 00123 ROMA% 06/30484671 Fax 06/30484891e-mail: [email protected]://www.sirr.unina.it
In copertina: Radiation damage by expected scatter dose in exposed vs bystan-der EGL. Immagine fornita da Mariateresa Mancuso and Anna Saran, Biotech-nology Unit, ENEA C.R.-Casaccia, Rome.
Segreteria S.I.R.R.Sezione Tossicologia e Scienze BiomedicheENEA Centro Ricerche Casaccia, s.p. 016Via Anguillarese, 301 - 00123 Roma% 06/30484442 Fax 06/30486559e-mail: [email protected]

3
RADIAZIONI Ricerca e ApplicazioniAnno XI n. 2/2008
ONCOGENIC BYSTANDER RADIATIONEFFECTS IN PATCHED HETEROZYGOUS
MOUSE CEREBELLUMMariateresa Mancuso and Anna SaranBiotechnology Unit, ENEA C.R.-Casaccia, Rome
e-mail: [email protected]@enea.it
Ionizing radiation is a well-known genotoxic agent andhuman carcinogen that causes different short- and long-term effects [1,2]. A longstanding paradigm for biologi-cal radiation effects has been that radiation traversalthrough the cell nucleus is required for genetic damageand biological responses. However, in the last twodecades this view has been challenged by observationsthat genetic/epigenetic changes occur in unexposed“bystander cells” neighboring directly-hit cells, due tocell-to-cell communication or soluble factors releasedby irradiated cells. To date, the vast majority of theseeffects are described in cell-culture systems, whilst in-vivo validation and assessment of biological conse-quences within an organism remain uncertain [3-6].Hemizygous Ptch1 mice have many features of Gorlinsyndrome, including radiosentivity to X-ray exposure.Neonatal irradiation greatly accelerates medulloblasto-ma, and promotes skin basal cell carcinoma (BCC)precursor lesions to progress to infiltrative BCC [7,8].To prevent early medulloblastoma mortality andimprove characterization of the skin phenotype in irra-diated Ptch1+/- mice, we irradiated progenies ofPtch1+/- and wild-type (wt) mice at postna-tal day 2 (P2) using expressly-designedcylindrical lead shields providing head pro-tection (SH groups; Fig. 1A) while expo-sing the remainder of the body to 3 Gy of Xrays. Shielding was checked by dosimetryand Monte Carlo simulation. The dose dueto primary photons beneath the shields wasfound to be < 0.26%. Shielded tissues,however, receive scattered radiation due toX-ray deflection through irradiated tissues.A conservative value of the dose (attenuatedand scattered) of 1.2% of the total dose toshielded tissues was estimated. Thus,another cohort of mice was whole-body (WB) exposedto 0.036 Gy as internal control. Three additional
cohorts were WB exposed to 3 Gy of X rays, sham irra-diated (SI), or left untreated (CN).Ptch1+/- mice were placed on a lifetime study andmonitored for tumor development (Fig. 1B). A highpercentage of WB-irradiated mice (62%) died ofaggressive disease by 23 weeks, with median survivalof 14 weeks (P < 0.0001 vs CN, log rank test). Signifi-cantly, we also observed a remarkably increasedmedulloblastoma rate (39%) in SH-irradiated Ptch1+/-
mice. Although this tumor response was lower than inWB-exposed animals (P = 0.0011), SH mice develo-ped significantly more tumors with highly reducedlatency relative to CN mice (P = 0.0003), showing thatpartial-body irradiation promotes Ptch1-driven tumori-genesis in shielded cerebellum. Mice WB exposed tothe estimated scatter dose (0.036 Gy) have not shownsigns of disease above background level. Biallelic Ptch1 loss (LOH) represents the majorpathway to medulloblastoma development in Ptch1+/-
mice. We analyzed tumors from irradiated and shieldedmice for Ptch1 allelic imbalance by exploiting a T/Cpolymorphism at position 4016 of Ptch1, that discrimi-
Fig. 1 (A) Irradiation set up for shielded irradiation. (B)Kaplan-Meier kinetic analysis of medulloblastoma. (C)Molecular analyses.

4
RADIAZIONI Ricerca e Applicazioni Anno XI n. 2/2008
nates the 129Sv-derived mutant from the CD1-derivedwt allele. Similar to WB-irradiated Ptch1+/- mice,medulloblastomas from SH–exposed mice showed lossof the wt-CD1 allele (Fig. 1C). To assess Ptch1 loss ofheterozigosity (LOH) mechanisms, panels of medullo-blastomas from SH or WB mice were analyzed using aminimum of 22 informative microsatellite markersspanning mouse chr-13. All medulloblastomas had achr-13 interstitial region in which several contiguousloci showed LOH (Fig. 1C). Overall, the pattern lackedhighly stringent breakpoint clustering, supporting agenetic mechanism based on loss of a critical genewithin the chr-13 minimal deleted region where welocalized Ptch1. Hence, our results suggest that bothdirect and bystander radiation damage promote tumo-rigenesis by facilitating LOH at the Ptch1 locus,implying that genetic damage is an important compo-nent of in-vivo oncogenic bystander responses.In the developing brain, radiosensitivity is highlydependent on developmental stage, being higher inneural granule cell progenitors (GCPs) compared withpostmitotic neurons [9]. In proliferating GCPs of P2cerebellum we have analyzed induction of γ-H2AXfoci, an endpoint that strongly correlates with radia-tion-induced DSBs. Intense γ-H2AX staining, invol-ving over 85% of GCPs, was present by immunohisto-chemistry in the EGL at 0.5 h post-3Gy WB irradiation(Fig. 2A). In contrast, no γ-H2AX staining was detec-ted in cerebellum of lead-shielded mice (Fig. 2B). γ-H2AX staining declined in irradiated EGL at latertimes (3, 6, and 18 h: 5.2, 2.9, and 2.5%), whilst remai-ning very low or undetectable in shielded EGL (0.05,0.15, and 0%). The finding of very low levels ofγ-H2AX induction by 3 Gy in shielded cerebel-lum, in contrast with highly increased tumor response,could be explained by a low mean number of DSBs percell. Slower kinetics of disappearance of DSBs in neu-ral progenitors relative to neurons, in addition, mightcontribute to a persistent low level of unrepairedDSBs. As evidence of persisting damage, a steepincrease in apoptosis was detected at 6 h in the EGL ofshielded mice relative to internal controls (WB-0.036Gy) or sham-irradiated mice (Fig. 2C-H).To confirm that the genetic damage to cerebellum suf-fered by shielded mice did not result from insufficientshielding or dose scatter and to maximize downstreambystander responses, we compared the magnitude andkinetics of DNA DSBs and of apoptosis in mice thathad been either SH-irradiated with 8.3 Gy (SH-8.3Gy),or WB exposed to the estimated 0.1 Gy scatter dose tocerebellum (WB-0.1Gy). Lacking additional indirecteffects, shielded EGL receiving 0.1 Gy through dosescatter should theoretically suffer biological damage
similar to the EGL of mice receiving 0.1 Gy to the totalbody. γ-H2AX focus formation was very low or unde-tectable in cerebellum directly exposed to 0.1 Gy (Fig.3B,C). Instead, highly significant induction of γ-H2AXfoci occurred in SH cerebellum at 4.5 (P = 0.0015) and6 h (P = 0.0139) after exposure to 8.3 Gy of theremainder of the body (Fig. 3A,C), with decrease tocontrol level at 18 h. A statistically significant apopto-tic response in shielded relative to 0.1-Gy exposedEGL was already evident at 4.5 h (P = 0.0001), with a3.2-fold bystander-related enhancement over the scat-ter-dose group (Fig. 3E-G), increasing to 7-fold at 6 h(P = 0.0001). Thus, significant bystander genotoxicresponses occur in shielded cerebellum in-vivo subse-quent to X-ray exposure of the remainder of the body.Significantly, short-term cellular responses were notspecific of radiosensitive Ptch1+/- mice, as wt CD1siblings showed identical bystander phenomena inneural precursors of P2 cerebellum (not shown).One of the major challenges in the field is to under-stand the mechanisms of non-targeted effects. In-vitrostudies have shown that, in confluent cultures, physicalcontacts through gap-junctional intercellular commu-nication (GJIC) between irradiated and non-irradiatedcells are essential for the process [10,11]. On the otherhand, in low-density cultures bystander effects mayrely on soluble factors released in the culture mediumby irradiated cells [12]. Here, we provide direct evi-dence that a Cx43-mediated gap-junction transfer ofthe bystander signal in vivo mediates short-term cellu-lar responses that may subsequently trigger longer-term carcinogenic effects in mouse CNS. In fact, TPAtreatment abrogated DNA-DSB responses in the EGLpost-8.3Gy SH irradiation (Fig. 3D,H). Moreover,apoptotic damage was reduced by 3.3-fold (Fig. 3H).In contrast, suppression of COX-2 activity in bystandercerebellum by the chemical inhibitor nimesulide didnot influence bystander damage, suggesting thatinflammation/oxidative stress is not central in theeffects observed here (Fig. 3D,H). In conclusion, these results describe the neonatalmouse cerebellum as an accurate in-vivo model todetect, quantify and mechanistically dissect radiation-bystander responses. DNA double-strand breaks andapoptotic cell death were induced in bystander cere-bellum in-vivo. Accompanying these genetic events,we report bystander-related tumor induction in cere-bellum of radiosensitive Ptch1 heterozygous mice sub-sequent to X-ray exposure of the remainder of thebody. We further show that genetic damage is animportant component of in-vivo oncogenic bystanderresponses, and provide direct evidence for the role ofGJICs in transmission of bystander signals. If ongoing

5
RADIAZIONI Ricerca e ApplicazioniAnno XI n. 2/2008
and future studies confirm the tumorigenic role ofbystander effects, this will have significant impact onfuture assessment of cancer risks associated with occu-pational, environmental and medical exposures.
References1. J.B. Little (2000) Radiation carcinogenesis. Carcinogene-
sis 21, 397-404. 2. M. Sowa, B.J. Arthurs, B.J.Estes and W.F. Morgan (2006)
Effects of ionizing radiation on cellular structures, indu-ced instability and carcinogenesis. EXS 96, 293-301.
3. H. Nagasawa and J.B. Little (1992) Induction of sisterchromatid exchanges by extremely low doses of alpha-particles. Cancer Res 52, 6394-6396.
4. W.F. Morgan, A. Hartmann, C.L. Limoli, S. Nagar and B.Ponnaiya (2002) Bystander effects in radiation-inducedgenomic instability. Mutat Res 504, 91-100.
5. O.V. Belyakov et al. (2005) Biological effects in unirra-diated human tissue induced by radiation damage up to 1mm away. Proc Natl Acad Sci USA 102, 14203-14208.
6. I. Koturbash et al. (2008) In vivo bystander effect: cranialX-irradiation leads to elevated DNA damage, altered cel-lular proliferation and apoptosis, and increased p53 levelsin shielded spleen. Int J Radiat Oncol Biol Phys 70: 554-562.
7. S. Pazzaglia et al. (2202) High incidence of medullobla-stoma following X-ray-irradiation of newborn Ptc1 hete-rozygous mice. Oncogene 21: 7580-7584.
8. M. Mancuso et al. (2004) Basal cell carcinoma and itsdevelopment: insights from radiation-induced tumors inPtch1-deficient mice. Cancer Res 64: 934-941.
9. E. Nowak et al. (2206) Radiation-induced H2AX pho-sphorylation and neural precursor apoptosis in the deve-loping brain of mice. Radiat Res 165: 155-164.
10. E.I. Azzam, S.M. de Toledo and J.B. (2001) Little Directevidence for the participation of gap junction-mediated
Fig. 2 Induction of DSBs and apoptosis in irradiated andbystander cerebellum.
Fig. 3 (A-F) Radiation damage by expected scatter dose in exposed vs bystander EGL. (G,H) Mechanism of non-targeted effects.
intercellular communication in the transmission of dama-ge signals from alpha -particle irradiated to nonirradiatedcells. Proc Natl Acad Sci USA 98: 473-478.
11. B. Hu et al. (2006). The time and spatial effects of bystan-der response in mammalian cells induced by low doseradiation. Carcinogenesis 27: 245-251.
12. Z. Goldberg and B.E. Lehnert (2002) Radiation-inducedeffects in unirradiated cells: a review and implications incancer. Int J Oncol 21: 337-349.

6
RADIAZIONI Ricerca e Applicazioni Anno XI n. 2/2008
1) l’irraggiamento di una piccola frazione di cellulegrazie all’utilizzo di una bassissima fluenza di par-ticelle alfa [2];
2) l’utilizzo di microbeam (dimensioni lineari del fa-scio dell’ordine di alcuni micron) in grado di irrag-giare selettivamente alcune cellule o, all’interno diuna singola cellula, di irraggiare selettivamente ilcitoplasma [5];
3) condivisione del terreno di coltura di cellule irraggia-te (e.g. attraverso utilizzo di inserti “Transwell”) [6].
Dalle prime evidenze ad oggi sono stati realizzatinumerosi studi fenomenologici che hanno permesso distudiare quali endpoints biologici sono presenti nellecellule bystander (e.g morte clonogenica, apoptosi,micronuclei, foci γ-H2AX, mutazioni) e quali possibi-li molecole segnale possano essere coinvolte nei mec-canismi bystander (e.g. ROS, NO e citochine).Sono stati inoltre realizzati una serie di esperimentti adhoc al fine di ottenere alcune delle caratteristiche cru-ciali necessarie per identificare i possibili segnali, tracui: il timing nella produzione di danni in cellulebystander [6-7], il timing nella produzione di segnaliextracellulari [8] e il timing di “scomparsa” del segna-le dal mezzo; è stato inoltre studiato il ruolo del dannoal DNA nelle cellule irraggiate, ritenuto necessariocome “trigger” per l’emissione di segnali o richiestonella risposta biologica [9].
Possibili meccanismi dell’effetto bystanderTra le specie chimiche che sono state proposte comemediatrici dell’effetto bystander ci sono le ROS (spe-cie reattive dell’ossigeno), l’NO (ossido nitrico) e alcu-ni segnali proteici, tra cui interleuchine e fattori di cre-scita (TGF-β e TNF-α) [10-12].In particolare sono stati realizzati alcuni esperimenti incui gli effetti bystander erano notevolmente attenuatigrazie all’utilizzo di opportuni enzimi “scavengers” in
IntroduzioneSecondo il paradigma classico della radiobiologia, glieffetti indotti dalle radiazioni ionizzanti sono il risul-tato di deposizioni di energia nel DNA presente nelnucleo della cellula. Tuttavia con il passare degli anni(fine anni ’80 - inizio anni ‘90) si è cominciata ariscontrare una serie di evidenze sperimentali di dannida radiazioni, in particolare alle basse dosi, per celluleche non erano state attraversate direttamente. La sco-perta di questi effetti, chiamati non-targeted effects inquanto non dovuti all’irraggiamento diretto del nucleodella cellula, ha portato a un cambiamento radicale delparadigma classico e all’abbandono della concezioneDNA-centrica del danno biologico da radiazioni.In realtà già nel 1954 si ebbe evidenza di effetti non-targeted [1], ma è solo agli inizi degli anni ’90 con gliesperimenti di Nagasawa e Little [2] e di Mothersill eSeymour [3] che, usando linee cellulari completamen-te differenti e differenti endpoints, vennero riscontratidei danni biologici (scambio di cromatidi fratelli emorte clonogenica) in cellule non irraggiate (effettobystander).
Effetto bystanderAd oggi si ritiene che l’effetto bystander indotto dallaradiazione si verifichi quando una cellula irraggiatacomunica con cellule non irraggiate attraverso la secre-zione di determinati fattori (nell’ambiente extracellula-re e/o attraverso gap junctions), inducendo in questeultime una risposta biologica [4].Tali effetti, riscontrati utilizzando sia diversi tipi diradiazioni ionizzanti (e.g. raggi X e raggi γ, particellealfa e altri ioni) sia diversi tipi cellulari, potrebberoavere un impatto significativoo nella valutazione delrischio da bassi dosi.Sono principalmente tre i tipi di esperimenti realizzatiper studiare gli effetti bystander:
MODELLIZZAZIONE DELL’EFFETTO BYSTANDER: COME LA RADIAZIONE
MODULA LA COMUNICAZIONE CELLULARE
Luca MariottiUniversità degli Studi di Pavia, Dipartimento di Fisica Nucleare e Teorica, Pavia
e-mail: [email protected]

7
RADIAZIONI Ricerca e ApplicazioniAnno XI n. 2/2008
grado di rimuovere selettivamente solo alcune speciechimiche (DMSO per le specie ROS ed il c-PTIO perl’NO) [13-14]. Diversi meccanismi sono stati propostiper spiegare l’effetto bystander, e in alcuni di questi lecitochine sembrano avere un ruolo chiave [15]. In particolare Lehnert e collaboratori [10] hanno pro-posto un possibile meccanismo di induzione del dannoin cellule non irraggiate così strutturato: in seguitoall’irraggiamento diretto della cellula si ha la produ-zione di ROS intracellulare; la presenza di ROS intra-cellulare può indurre la trascrizione del fattore nuclea-re Nf-kB, che a sua volta può indurre la sintesi disegnali proteici (e.g. IL-8); tali citochine sono in gradodi diffondere dalla cellula irraggiata nell’ambienteextra-cellulare e quindi di raggiungere cellule nellevicinanze. Una volta internalizzata nella cellulabystander mediante un opportuno recettore selettivo,tale citochina può attivare la proteina di membranaNADH+, che è in grado di rilasciare H2O2 nell’am-biente intracellulare. Si è giunti in questo modo allapresenza di radicali liberi (indotti dalla comunicazionecellulare via citochine) in grado di danneggiare ilmateriale genetico in cellule non irraggiate.
ModelliL’attività di modellizzazione dell’effetto bystander sipropone di portare ad una quantificazione dei parame-tri cruciali e dei meccanismi coinvolti nell’effettobystander (prima in vitro, poi in vivo) sia per sistemi2D sia per sistemi 3D [16-19]. In questo contesto, pres-so l’Università di Pavia sono stati sviluppati un model-lo meccanicistico e un codice Monte Carlo in grado disimulare il rilascio, la diffusione e l’interazione di cito-chine (e.g IL-6) in colture di fibroblasti AG1522 semi-nati ad un’opportuna densità.I risultati delle simulazioni sono stati confrontati conesperimenti ad hoc costruiti per indagare sul rilascio el’“assorbimento” di IL-6 da parte di una coltura difibroblasti dopo il cambio del terreno di coltura, inseguito a irraggiamenti da 0.25 Gy e 1 Gy di radiazio-ne γ. Gli irraggiamenti sono stati effettuati presso illaboratorio di radioterapia del Policlinico S.Matteo diPavia mediante una sorgente 60Co.La modellizzazione del rilascio, diffusione e interazio-ne delle citochine è stata realizzata mediante la costru-zione di una geometria che ricalcasse, in scala, l’insie-me dei fibroblasti presenti sulla base della flasca uti-lizzata negli esperimenti. Si sono quindi implementatii seguenti tre processi a cui possono essere sottopostele citochine:(a) rilascio di citochine con un rate costante nel tempo;(b) loro trasporto mediante un modello di diffusione
libera (moto Browniano);
(c) “eliminazione” dal terreno di coltura, medianteinternalizzazione da parte delle cellule raggiuntedalle molecole segnale (depletion).
È stato in questo modo possibile effettuare una valida-zione dei tre processi ipotizzati (rilascio, diffusione edepletion) che stanno alla base dell’andamento tempo-rale della concentrazione di citochine in una coltura difibroblasti, ed una quantificazione del rate di rilasciosia in assenza (controllo) sia in presenza di radiazione.I risultati della simulazione mostrati in figura sono statiottenuti con tre diversi rate di rilascio nel caso speri-mentale di 10,000 cellule/cm2 seminate ed irraggiate
Fig. 1. Andamento temporale della concentrazione di Inter-leuchina 6 nel caso di 250,000 fibroblasti “sham-irradiated”ed in seguito ad un irragiamentoγ da 0.25 Gy e 1 Gy. (I puntisono dati sperimentali, le linee sono simulazioni MonteCarlo).
con tre differenti dosi di radiazione γ. Si è ottenuto unrilascio da parte di singola cellula di 0.013 citochine/snel caso senza radiazione, di 0.016 citochine/s nel casodi 0.25 Gray e di 0.012 citochine/s nel caso di 1 Gray.Si può quindi concludere che le cellule rispondono inmaniera diversa ad una differente dose di radiazione γ,con un rilascio maggiore di citochine in seguito all’ir-raggiamento a basse dosi (0.25 Gy).
ConclusioniL’approccio Monte Carlo, raffrontato con i risultatisperimentali, ha suggerito che i meccanismi coinvoltinella comunicazione cellulare (mediata dalle citochi-ne) possano essere quelli di rilascio e di internalizza-zione continua da parte dei fibroblasti: in particolarenel caso di un irraggiamento a bassa dose (0.25 Gy) siè confermato un incremento nel rilascio delle citochinepari circa al 20% rispetto al caso ”sham-irradiated”.
Referenze1. Pearson J.(1954) Changes in sternal bone marrow fol-
lowing roentgen-ray therapy to the spleen in chronic gra-

8
RADIAZIONI Ricerca e Applicazioni Anno XI n. 2/2008
nulocytic leukemia. Cancer. 4: 179-1892. Nagasawa H, Little JB.(1992) Induction of sister chroma-
tid exchanges by extremely low doses of α-particles.Cancer Res. 52: 6394–96.
3. Mothersill C, Seymour C.(1997) Medium from irradiatedhuman epithelial cells but not human fibroblasts reducesthe clonogenic survival of irradiated cells. Int J RadiatBiol; 71: 421–27.
4. Morgan WF (2003) Is there a common mechanismunderlying genomic instability, bystander effects andother non targeted effects of exposure of ionizing radia-tion? Oncogene; 22:7094-7099
5. Shao C, Folklard M, Barry M, Prise M. (2004) Targetedcytoplasmatic irradiation induces bystander response.PNAS; 101:37
6. H Yang, V Anzenberg and K Held (2007) The time depen-dence of bystander responses induced by iron-ion radia-tion in normal human skin fibroblasts. Radiat Res; 168:292-298
7. B Hu, L Wu, W Han, L L Zhang, S P Chen, A Xu, T K.Heiand Z Yu.(2006) The time and spatial effects of bystanderresponse in mammalian cells induced by low dose radia-tion. Carcinogenesis; 27: 245–251
8. Facoetti A et al (2008) Hotspots for experimentalists andmodellers to study cytokine release for radiation inducedbystander effects. Submitted Int J Radiat Biol
9. Prise K M. et al.(2007) New insight on radiation inducedbystander signaling and its relationship to DNA repair.International Congress Series; 1299:121-127
10. Lehnert BE, Goodwin EH.(1997) Extracellular factor(s)following exposure to alpha-particles can cause sisterchromatid exchanges in normal human cells. Cancer Res;
QUOTA ASSOCIATIVA S.I.R.R. 2008...E QUELLE ARRETRATE!
Carissimo Socio,come sai, la quota sociale, oltre ad essere la principale fonte di finanziamento per il funziona-mento della nostra Società, è anche un segno annuale di adesione e partecipazione.La quota sociale, attualmente ad un livello minimo, è un dovere che ogni Socio deve assolve-re entro il 31 marzo di ogni anno, onde evitare che la gestione delle quote con relativi solle-citi e verifiche abbia un costo superiore alla stessa quota. La quota per il 2008 è di € 30,00 e potrà essere versata tramite assegno circolare o bancario,non trasferibile, intestato a S.I.R.R. oppure tramite versamento in contanti alla Segreteria oppu-re mediante bonifico bancario: c/c n. 14688 c/o Banca Nazionale del Lavoro 6385 RomaCasaccia Via Anguillarese 301 - 00123 Roma. Coordinate bancarie IBAN: IT 19T0100503385000000014688Con l'intento di favorire i cosidetti "non strutturati" (studenti, borsisti, etc.) la quota sociale èridotta a € 15,00, chi si trova in questa condizione dovrà esplicitamente dichiararlo medianteautocertificazione contestualmente all'invio della quota annuale.Fiduciosi della tua collaborazione e partecipazione, cogliamo l'occasione per inviarti i nostripiù cari saluti.
LA SEGRETERIA
57: 2164–71.11. Iyer R, Lehnert BE.(2000) Factors underlying the cell
growth-related bystander responses to alpha particles.Cancer Res; 60: 1290–98.
12. Portess D I, Bauer G, Hill M A, O’Neill P.(2007) Low-Dose Irradiation of Nontransformed Cells Stimulates theSelective Removal of Precancerous Cells via IntercellularInduction of Apoptosis. Cancer Res; 63: 1246-1253
13. Shao C, Stewart V, Folkard M, et al.(2004) Nitric oxide-mediated signaling in the bystander response of indivi-dually targeted glioma cells. Cancer Res; 63: 8437–42.
14. W Han, L Wu et al (2007) Constitutive nitric oxide actingas a possible intercellular signaling molecule in the inita-tion of radiation induced DNA double strand breaks innon-irradiated bystander cells. Oncogene; 26: 2330-2339
15. Bentzen SM. (2006). Preventing or reducing late sideeffects of radiation therapy: radiobiology meets molecularpathology. Nat Rev Cancer; 6:702-13.
16. M P Little et al (2005) A model for radiation-inducedbystander effects, with allowance for spatial position andthe effects of cell turnover. J Theoretical Biology; 232:329-338
17. H Schoellnberger (2007) Detrimental and protectivebystander effects: A model approach. Radiat Res;168:614-626
18. F Ballarini et al (2006) Modelling radiation-inducedbystander effect and cellular communication. Radiat ProtDosim; 122: 244-251
19. I Shuryak, R Sachs, D Brenner (2007) Biophysicalmodels of radiation induced bystander effects:1) Spatialeffects in three dimensional tissue. Radiat Red; 168: 741-749

9
RADIAZIONI Ricerca e ApplicazioniAnno XI n. 2/2008
MOVIMENTI INTRA-FRAZIONE IN TRATTAMENTI DI PROTONTERAPIA CON
‘SPOT-SCANNING’: EFFETTI DOSIMETRICIE METODI DI COMPENSAZIONE
Silvia Tomaselli, Marco SchwarzAgenzia Provinciale per la Protonterapia (ATreP) - Trento
e-mail: [email protected]@atrep.it
IntroduzioneI protoni hanno proprietà fisiche che permettono diottenere una migliore localizzazione della dose nel tar-get rispetto ai fotoni, risparmiando i tessuti sani adia-centi.Tra le tecniche di erogazione di fasci di protoni, attual-mente riscuote un particolare interesse la scansioneattiva di fasci sottili di protoni (‘beam scanning’); conquesta tecnica, infatti, è possibile risolvere alcuni pro-blemi delle tecniche convenzionali di beam scattering,creando distribuzioni in cui la dose depositata da unfascio scansionato ad ogni punto nel tessuto è in buonamisura indipendente dalla dose in qualsiasi altro punto.Un aspetto critico delle tecniche di scanning è la sensi-bilità della distribuzione di dose rispetto al movimentod’organo intra-frazione, che causa divergenze tra ladistribuzione di dose pianificata e quella effettivamen-te depositata. La questione è di particolare interesse peri trattamenti di protonterapia con ‘spot-scanning’ in sititumorali soggetti al movimento intra-frazione, comeper esempio i polmoni. Ci sono diverse possibilità per compensare le distor-sione nella distribuzione di dose causate del movimen-to intra-frazione, che si producono nell’irradiazionecon scansione attiva di fasci sottili di protoni [1,2]. Unadi queste è il ‘repainting’, che consiste nello scansio-nare più volte il volume bersaglio depositando una fra-zione della dose totale ad ogni scansione. Se gli effettidi interplay sono casuali, ovvero se il movimentorespiratorio e il sistema di irradiazione non sono corre-lati, ci si aspetta che tali effetti siano mediati su uncerto numero di scansioni.Un obiettivo di questo lavoro è stato quello di studiaregli effetti di interplay tra movimento d’organo intra-frazione e irradiazione con sistema di scansione attiva.Abbiamo poi analizzato l’efficacia del repainting nel
compensare le disomogeneità della distribuzione didose, rispetto alla singola scansione.Infine, abbiamo esaminato quale schema di repaintingpermette di ottimizzare il tempo di trattamento, cherimane un parametro critico in radioterapia.
Tecniche di irradiazioneNelle simulazioni abbiamo implementato il metodo discansione applicato al Paul Scherrer Institute (PSI),vicino a Zurigo [3]. Abbiamo usato due diversi schemidi scansione, che qui chiameremo ‘gantry1’ e ‘gan-try2’. Nel primo, utilizzato nelle pratica clinica al PSI, ladose è depositata nel volume bersaglio in piani paral-leli all’asse del fascio (Fig.1). Il fascio è scansionatoprima lungo una linea alla stessa profondità utilizzan-do l’azione di un magnete. Poi si diminuisce la profon-dità degli spot variando l’energia del fascio con unsistema di piatti inseriti individualmente nel percorsodel fascio. Il terzo asse di scansione, nella secondadirezione laterale, è controllato dal movimento deltavolo del paziente. Durante gli spostamenti il fascioviene disattivato mediante un altro magnete.Nel metodo di scansione progettato per il nuovo gantry(‘gantry2’) il fascio sarà scansionato in piani perpendi-colari all’asse del fascio, perchè ci sarà un secondomagnete per la scansione veloce che sostituirà il movi-mento lento del tavolo (Fig.1). I tempi morti per gli spostamenti del fascio da unaposizione alla successiva con il magnete ‘sweeper’, ilsistema di ‘range shifter’ e il movimento del tavolosono rispettivamente 4 ms, 40 ms e 1 s. Il tempo tota-le per trattare un volume di 1 litro è di circa 4 minuti,e questo implica che i problemi causati dal movimentointra-frazione dovrebbero essere considerati.Considerando il metodo di scansione del gantry1 si

10
RADIAZIONI Ricerca e Applicazioni Anno XI n. 2/2008
Calcolo della dose 4DLa distribuzione di dose in assenza di movimentointra-frazione (caso statico) è calcolata con un’ espres-sione che può essere riassunta come segue:
possono implementare due principali schemi di repain-ting, che indicheremo con ‘repainting 3D’ e ‘2D’. Nelprimo caso, per ogni scansione viene irradiato l’interovolume prima di passare alla scansione successiva.Questo significa che per ogni scansione viene effettua-to il massimo spostamento lungo il tavolo. Con ilrepainting 2D invece lo spostamento del tavolo delpaziente viene minimizzato, poiché per ogni piano per-pendicolare allo spostamento del tavolo si eseguonotutte le scansioni previste, prima di passare al pianosuccessivo.
Impostazione delle simulazioniAbbiamo pianificato un unico campo con angolo delgantry di -90° e angolo del tavolo di 0°, i.e. il primomagnete sweeper agisce nella direzione Cranio-Cauda-le (CC), il range shifter in direzione Latero-Laterale(LL), mentre il tavolo del paziente (per gantry1) o ilsecondo magnete sweeper (per gantry2) controllano ilmovimento in direzione Anteriore-Posteriore (AP).Abbiamo simulato un trattamento in seduta unica,assumendo l’assenza di errori di set-up e delineando untarget sferico (diametro 2 cm), in un fantoccio d’acquacubico, su cui abbiamo pianificato una distribuzione didose omogenea. Le informazioni in uscita del Sistemadi Pianificazione del Trattamento (TPS) relative all’ir-radiazione (posizione e numero degli spot) sono statemanipolate per diverse combinazioni di numero direpainting, percorso di scansione e frazione del nume-ro totale di spot sottoposto a repainting. Per ogni sche-ma di repainting è stata creata una sequenza di irradia-zione che permettesse di associare un ‘timestamp’ adogni spot. Parallelamente, sono stati calcolati i vettoridi movimento in base all’anatomia del paziente simu-lata. Il movimento intra-frazione è stato simulato indue modi: attraverso una funzione sinusoidale, cos4,modificata in modo che la posizione mediata sul tempofosse uguale a zero [5,6], o utilizzando dati di movi-mento generati da respiro reale ottenuti da misure diMRI 4D [7].
dove Dx è il valore di dose sull’asse centrale, allaprofondità x, e i due termini esponenziali rappresenta-no il contributo di dose nelle due direzioni ortogonali ze y. Per tener conto dell’effetto del movimento intra-frazio-ne abbiamo modificato questo algoritmo, sviluppandoun modello che calcola la dose non solo in funzionedelle coordinate spaziali ma anche del tempo. L’equa-zione per calcolare la cosidetta ‘dose 4D’ è:
dove dz(t) e dy(t) sono rappresentazioni vettoriali dellavariazione, in funzione del tempo t, della posizionelungo z e lungo y di ogni punto della griglia per il cal-colo della dose. Nelle simulazioni abbiamo implemen-tato movimenti nelle direzioni laterali z e y.Il TPS produce gli Istogrammi Dose-Volume (DVH)del caso statico e in presenza di movimento.Per l’analisi dei DVH abbiamo calcolato la dosemedia, la dose ricevuta da almeno il 99% del volume ela dose ricevuta da almeno l’1% del volume, che indi-cheremo rispettivamente con <D>, D99 e D1. Inoltreper quantificare il livello di uniformità della distribu-zione di dose abbiamo scelto l’omogeneità di dose,definita come segue:
Fig. 1
dove Di è la dose del singolo voxel e N è il numero divoxel nel volume bersaglio. Infine, per valutare la risposta del tumore alle diversedistribuzioni di dose abbiamo usato il TCP (TumorControl Probability) secondo la formula di Webb-Nahum[8].
RisultatiL’unico parametro statistico che ha mostrato variazio-ni relativamente ridotte in presenza di movimentod’organo intra-frazione in trattamenti in singola scan-sione è <D> (±2.5%). Tutti gli altri variano significa-tivamente (tra 30% e 40%), portando a distribuzioni didose inaccettabili nella pratica clinica. Per esempio, un movimento intra-frazione di 10 mm(ampiezza picco-picco) riduce l’omogeneità di doseanche del 20% rispetto al caso statico e il TCP passa
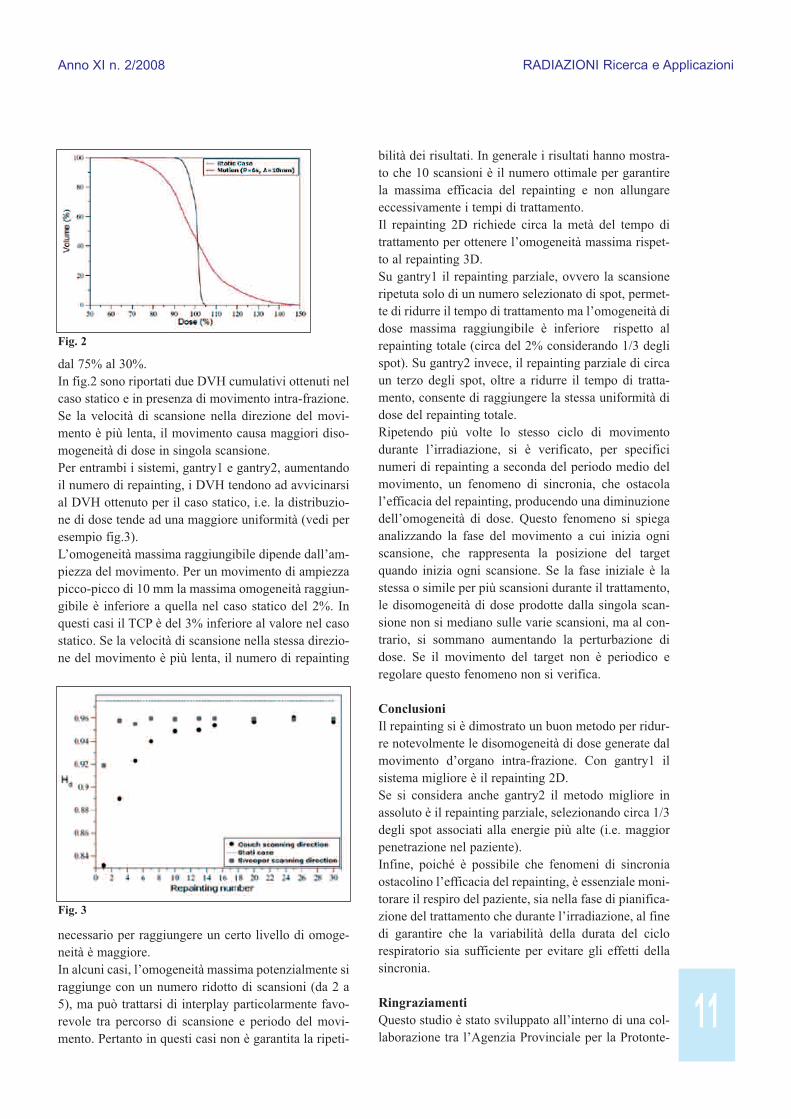
11
RADIAZIONI Ricerca e ApplicazioniAnno XI n. 2/2008
bilità dei risultati. In generale i risultati hanno mostra-to che 10 scansioni è il numero ottimale per garantirela massima efficacia del repainting e non allungareeccessivamente i tempi di trattamento.Il repainting 2D richiede circa la metà del tempo ditrattamento per ottenere l’omogeneità massima rispet-to al repainting 3D.Su gantry1 il repainting parziale, ovvero la scansioneripetuta solo di un numero selezionato di spot, permet-te di ridurre il tempo di trattamento ma l’omogeneità didose massima raggiungibile è inferiore rispetto alrepainting totale (circa del 2% considerando 1/3 deglispot). Su gantry2 invece, il repainting parziale di circaun terzo degli spot, oltre a ridurre il tempo di tratta-mento, consente di raggiungere la stessa uniformità didose del repainting totale.Ripetendo più volte lo stesso ciclo di movimentodurante l’irradiazione, si è verificato, per specificinumeri di repainting a seconda del periodo medio delmovimento, un fenomeno di sincronia, che ostacolal’efficacia del repainting, producendo una diminuzionedell’omogeneità di dose. Questo fenomeno si spiegaanalizzando la fase del movimento a cui inizia ogniscansione, che rappresenta la posizione del targetquando inizia ogni scansione. Se la fase iniziale è lastessa o simile per più scansioni durante il trattamento,le disomogeneità di dose prodotte dalla singola scan-sione non si mediano sulle varie scansioni, ma al con-trario, si sommano aumentando la perturbazione didose. Se il movimento del target non è periodico eregolare questo fenomeno non si verifica.
ConclusioniIl repainting si è dimostrato un buon metodo per ridur-re notevolmente le disomogeneità di dose generate dalmovimento d’organo intra-frazione. Con gantry1 ilsistema migliore è il repainting 2D.Se si considera anche gantry2 il metodo migliore inassoluto è il repainting parziale, selezionando circa 1/3degli spot associati alla energie più alte (i.e. maggiorpenetrazione nel paziente).Infine, poiché è possibile che fenomeni di sincroniaostacolino l’efficacia del repainting, è essenziale moni-torare il respiro del paziente, sia nella fase di pianifica-zione del trattamento che durante l’irradiazione, al finedi garantire che la variabilità della durata del ciclorespiratorio sia sufficiente per evitare gli effetti dellasincronia.
RingraziamentiQuesto studio è stato sviluppato all’interno di una col-laborazione tra l’Agenzia Provinciale per la Protonte-
dal 75% al 30%. In fig.2 sono riportati due DVH cumulativi ottenuti nelcaso statico e in presenza di movimento intra-frazione.Se la velocità di scansione nella direzione del movi-mento è più lenta, il movimento causa maggiori diso-mogeneità di dose in singola scansione.Per entrambi i sistemi, gantry1 e gantry2, aumentandoil numero di repainting, i DVH tendono ad avvicinarsial DVH ottenuto per il caso statico, i.e. la distribuzio-ne di dose tende ad una maggiore uniformità (vedi peresempio fig.3).L’omogeneità massima raggiungibile dipende dall’am-piezza del movimento. Per un movimento di ampiezzapicco-picco di 10 mm la massima omogeneità raggiun-gibile è inferiore a quella nel caso statico del 2%. Inquesti casi il TCP è del 3% inferiore al valore nel casostatico. Se la velocità di scansione nella stessa direzio-ne del movimento è più lenta, il numero di repainting
necessario per raggiungere un certo livello di omoge-neità è maggiore.In alcuni casi, l’omogeneità massima potenzialmente siraggiunge con un numero ridotto di scansioni (da 2 a5), ma può trattarsi di interplay particolarmente favo-revole tra percorso di scansione e periodo del movi-mento. Pertanto in questi casi non è garantita la ripeti-
Fig. 2
Fig. 3

12
RADIAZIONI Ricerca e Applicazioni Anno XI n. 2/2008
rapia (ATreP) di Trento ed il Paul Scherrer Institute(PSI) di Villigen.
Referenze1. Keall P J, Mageras G S, Balter J M, Emery R S, Forster K
M, Jiang S B, Kapatoes J M, Low D A, Murphy M J, Mur-ray B R and others (2006) The management of respiratorymotion in radiation oncology. Report of AAPM TaskGroup 76 Med. Phys. 33 3874-900
2. Webb S (2006) Motion effects in (intensity modulated)radiation therapy: a review. Phys. Med. Biol. 51 R403-25
3. Pedroni E, Scheib S, Boehringer T, Coray A, GrossmannM, Lin S and Lomax A (2005) Experimental characteri-zation and physical modelling of the dose distribution ofscanned proton pencil beams. Phys. Med. Biol. 50(3)541-61
4. Lambert J, Suchowerska N, McKenzie D R and Jackson M(2005) Intrafractional motion during proton beam scan-ning. Phys. Med. Biol. 50(20) 4853-62
5. Lujan A E, Larsen W E, Balter J M, Ten Haken R K (1999)A method for incorporating organ motion due to breatinginto 3D dose calculations. Med. Phys. 26(5) 715-720
6. Seppenwoolde Y, Shirato H, Kitamura K, Shimizu S, vanHerk M, Lebesque J, Miyasaka K (2002) Precise andreal-time measurement of 3D tumor motion in lung due tobreathing and heartbeat, measured during radiotherapy.Int. J. Radiat. Oncol. Biol. Phys. 53(4) 822-834
7. von Siebenthal M, Székely G, Gamper U, Boesiger P,Lomax A, Cattin P (2007) 4D MR imaging of
respiratory organ motion and its variability. Phys. Med.Biol. 52(6) 1547-64
8. Webb S and Nahum A (1993) A model for calculatingtumour control probability in radiotherapy including theeffects on inhomogeneous distributions of dose and clo-nogenic cell density. Phys. Med. Biol 38(6) 653-666
X INTERNATIONAL CONFERENCE ON ENVIRONMENTAL MUTAGENS
The Renaissance of Environmental Mutagenesis
Palazzo dei Congressi Firenze, 20-25 Agosto 2009
Per informazioniconsultare il sito: www.icem2009.org
Segreteria Congressuale: OIC srl, e-mail:[email protected]

13
RADIAZIONI Ricerca e ApplicazioniAnno XI n. 2/2008
IntroduzioneLa radiazione ionizzante causa diversi danni ai tessutiviventi attraverso una serie di meccanismi molecolariche dipendono dall’energia della radiazione. Dalmomento che i tessuti umani contengono acqua perl’80%, il danno principale della radiazione è dovuto airadicali liberi dell’acqua, prodotti dall’interazione dellaradiazione con le molecole di acqua. Questi reagisconocon diversi bersagli nel sistema biologico e causanodisfunzioni e mortalità cellulari. Il danno cellulareradioindotto è potenziato o mitigato per l’azione dialcuni fattori, come la presenza dell’ossigeno, cherende le cellule più sensibili alle radiazioni, di compo-sti sulfidrilici, che hanno una funzione protettiva neiconfronti dell’esposizione di cellule alle radiazioni, e dialtre molecole presenti nel mezzo cellulare. Il dannoossidativo al materiale genetico cellulare, come ilDNA, gioca un ruolo fondamentale nei processi dimutagenesi e carcinogenesi. I radicali dell’ossigeno altamente reattivi prodotti dallaradiazione ionizzante causano lesioni al DNA, che pos-sono portare alla morte cellulare e a mutazioni. Enzimicome la superossido dismutasi, la glutatione perossida-si e le catalasi proteggono le cellule di mammifero daldanno ossidativo della radiazione. Tali enzimi agisco-no trasformando l’anione superossido in perossido diidrogeno, che è conseguentemente detossificato e tra-sformato in acqua e ossigeno molecolare. Le celluletumorali crescono rapidamente e invadono i tessuti cheli circondano formando delle regioni necrotiche e ipos-siche che rendono la radiazione poco efficace in questearee. Per poter quindi controllare l’invasività tumoralee evitare la formazione di metastasi, bisogna sommini-strare alte dosi di radiazioni. Questo non è possibileclinicamente. Infatti dal momento che i tessuti norma-
li che circondano le cellule tumorali sono ben irrorati eossigenati, sono molto più sensibili alle radiazioniionizzanti, e quindi molto più predisposti al danno daradiazione. Di qui la necessità di utilizzare sostanzeche proteggano i tessuti normali dai danni radioindottie aumentino invece la sensibilità alle radiazioni delbersaglio critico. Oggetto di questo lavoro è stato lostudio di una proteina, isolata dal dott. A. Mancini del-l’Istituto Nazionale dei Tumori Fondazione G. Pascaledi Napoli. Tale proteina è una variante della MnSODisolata da cellule di un liposarcoma (LSA) pleiomorfi-co umano e pertanto definita LSA-type-MnSOD. Essaè stata prodotta in modo ricombinante usando lisati direticolociti di coniglio o virus di vaccini ricombinantiin cellule di fibroblasti A9 di topo, ed è stata pertantodefinita L-rSOD. Essendo una superossido dismutasi,questa proteina ha la stessa funzione radioprotettivadelle SOD espresse normalmente in tutte le celluledegli organismi aerobi. Il suo meccanismo d’azionequindi consiste nella conversione dei radicali liberidell’ossigeno, prodotti sia dal metabolismo cellularesia dalla radiazione ionizzante, in perossido di idroge-no. Il perossido di idrogeno è poi trasformato in acquae ossigeno molecolare per l’azione degli enzimi catala-si o alternativamente dagli enzimi glutatione perossi-dasi. La L-SOD tuttavia differisce dalle SOD conven-zionali perché non è localizzata come queste ultimenella matrice mitocondriale ma è secreta dalle celluleche la producono ed è pertanto veicolabile; infatti essapuò essere facilmente somministrata in vitro o in vivoper via sistemica. Per meccanismi molecolari nonancora totalmente noti essa ha la capacità di penetraresolo nelle cellule tumorali, dove esercita la sua funzio-ne enzimatica; ciò la rende particolarmente versatileper il trattamento terapeutico delle neoplasie. La pro-
CITOTOSSICITÀ DI UNA VARIANTEISOFORME DI MANGANESE
SUPEROSSIDO DISMUTASI E SUAINTERAZIONE CON LA RADIOSENSIBILITÀ
IN CELLULE UMANEMaria Rosaria Rusciano
Dipartimento di Biologia e Patologia Cellulare e Molecolare L. CalifanoUniversità degli Studi di Napoli “Federico II”
e-mail: [email protected]

14
RADIAZIONI Ricerca e Applicazioni Anno XI n. 2/2008
teina presenta, infatti, una attività citotossica specificae selettiva nei riguardi delle sole cellule tumorali.Dopo essere entrata nelle cellule tumorali essa trasfor-ma tutti i radicali disponibili in perossido di idrogeno.La concentrazione di questo perossido è così elevatache la scarsa quantità di Catalasi presente in una cellu-la tumorale non riesce a neutralizzare la quantità diperossido creata dalla L-rSOD, provocando la morteper necrosi delle cellule tumorali.
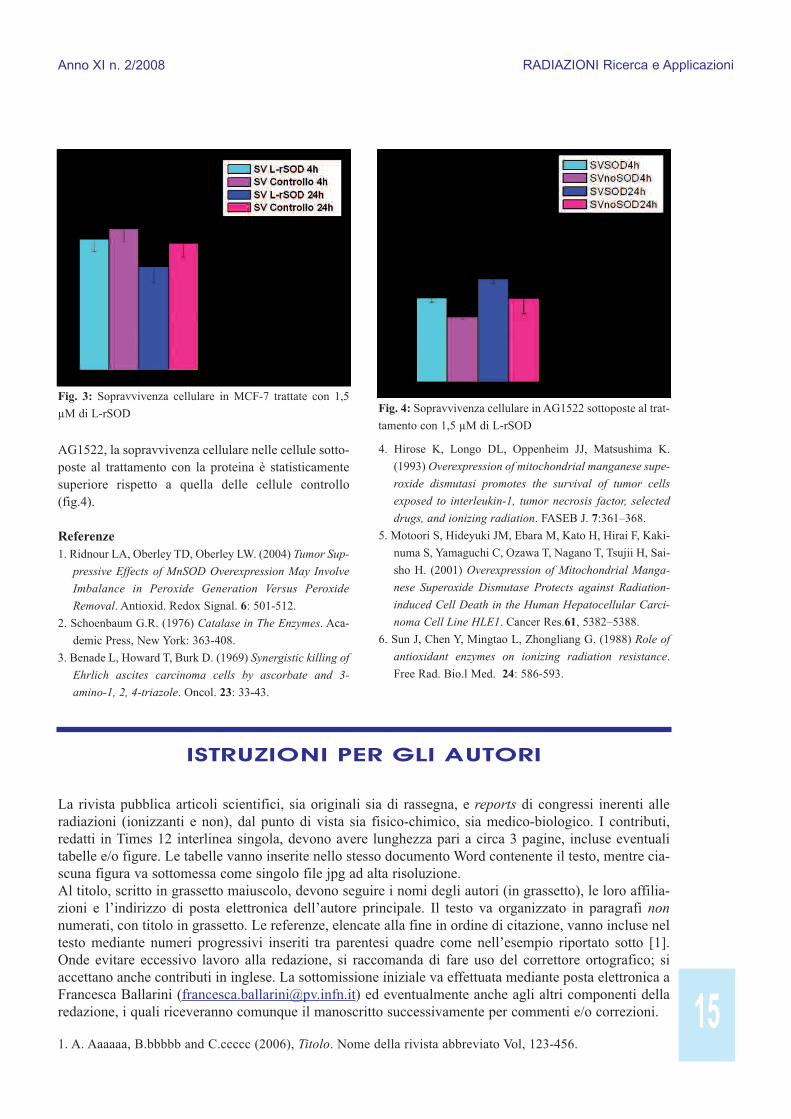
RisultatiAl fine di valutare gli effetti della L-rSOD su celluleumane è stato effettuato un test clonogenico su duediverse linee cellulari in presenza della proteina ricom-binante. Le linee cellulari scelte per tale esperimentosono state MCF-7 e AG1522. Il test clonogenico è statoeffettuato utilizzando la proteina ricombinante a duediverse concentrazioni: 0,4 µM e 1,5 µM. Gli effettidel trattamento con la proteina sono stati osservati 4 e24 ore dopo la somministrazione. I risultati ottenutihanno dimostrato che alla concentrazione più bassa laL-rSOD non aveva alcun effetto sulle capacità di cre-scita cellulare sia nelle cellule tumorali MCF-7 sia inquelle normali AG1522. Risultati molto interessantisono stati ottenuti utilizzando una concentrazione pro-teica di 1,5 µM. A tale concentrazione la proteina haindotto morte clonogenica nelle cellule tumorali, infat-ti la misura dell’efficienza di clonaggio diminuiscenelle cellule trattate con L-rSOD rispetto al controllo(fig.1).Tale effetto, invece, non è stato osservato nelle cellulenormali; infatti la misura dell’efficienza di clonaggiodelle cellule trattate con L-rSOD non differisce daquella delle cellule a cui non è stata somministrata laproteina (fig.2). La L-rSOD mostra un’azione specifi-ca e selettiva su cellule di tumori estrogeno-dipenden-ti. Tale azione si esplica in una induzione della necrosidella massa tumorale. Ciò è dovuto ad un meccanismoabbastanza chiaro secondo cui la proteina ricombinan-te, nei mitocondri, converte i radicali liberi dell’ossi-geno in acqua ossigenata. L’acqua ossigenata, che nelle cellule normali è trasfor-mata in acqua e ossigeno molecolare grazie all’azionedegli enzimi catalasi, nelle cellule tumorali si accumu-la per una totale mancanza o diminuzione delle catala-si stesse [1-3].Dal momento che l’acqua ossigenata è citotossica, ilrisultato del trattamento con la proteina ricombinantenelle cellule tumorali, quali MCF-7, è la morte dellacellula. Le cellule AG1522 non subiscono invece l’a-zione necrotica della proteina ricombinante perché,essendo una linea cellulare normale, esprimono glienzimi catalasi che trasformano il perossido di idroge-
no in acqua e ossigeno molecolare. Infine è stato valu-tato l’effetto radioprotettivo della L-rSOD. È statoinfatti osservato, in diversi studi [4-6], che l’over-espressione del gene della MnSOD protegge le celluledalla morte indotta dalla radiazione ionizzante. Il mec-canismo radioprotettivo attraverso cui agisce laMnSOD-2 si attua nella conversione del radicale supe-rossido in perossido di idrogeno, che è successivamen-te trasformato in acqua e ossigeno molecolare perintervento delle catalasi. L’effetto della proteina ricom-binante, utilizzata ad una concentrazione di 1,5 µM, inrisposta alla citotossicità radioindotta è stato valutatosulle linee cellulari AG1522 e MCF-7. In entrambe lelinee cellulari la proteina è stata aggiunta 10 minutiprima dall’irraggiamento, che è stato poi effettuatoesponendo le cellule a raggi X di 250 kVp ad una dosedi 3 Gy. L’effetto delle proteina è stato valutato a 4 e24 ore dall’irraggiamento. I risultati ottenuti per lalinea cellulare MCF-7 hanno mostrato che l’uso com-binato di radiazione ionizzante e proteina ricombinan-te non ha un effetto sinergico nel ridurre la sopravvi-venza cellulare (fig.3). Diversamente nelle cellule
Fig. 1: Efficienza di clonaggio in cellule MCF-7 sotto-poste a trattamento con [1,5 µM] di L-rSOD
Fig. 2: Efficienza di clonaggio in cellule AG1522 sottopostea trattamento con [1,5 µM] di L-rSOD
15
RADIAZIONI Ricerca e ApplicazioniAnno XI n. 2/2008
AG1522, la sopravvivenza cellulare nelle cellule sotto-poste al trattamento con la proteina è statisticamentesuperiore rispetto a quella delle cellule controllo(fig.4).
Referenze1. Ridnour LA, Oberley TD, Oberley LW. (2004) Tumor Sup-
pressive Effects of MnSOD Overexpression May InvolveImbalance in Peroxide Generation Versus PeroxideRemoval. Antioxid. Redox Signal. 6: 501-512.
2. Schoenbaum G.R. (1976) Catalase in The Enzymes. Aca-demic Press, New York: 363-408.
3. Benade L, Howard T, Burk D. (1969) Synergistic killing ofEhrlich ascites carcinoma cells by ascorbate and 3-amino-1, 2, 4-triazole. Oncol. 23: 33-43.
Fig. 3: Sopravvivenza cellulare in MCF-7 trattate con 1,5µM di L-rSOD Fig. 4: Sopravvivenza cellulare in AG1522 sottoposte al trat-
tamento con 1,5 µM di L-rSOD
ISTRUZIONI PER GLI AUTORI
La rivista pubblica articoli scientifici, sia originali sia di rassegna, e reports di congressi inerenti alleradiazioni (ionizzanti e non), dal punto di vista sia fisico-chimico, sia medico-biologico. I contributi,redatti in Times 12 interlinea singola, devono avere lunghezza pari a circa 3 pagine, incluse eventualitabelle e/o figure. Le tabelle vanno inserite nello stesso documento Word contenente il testo, mentre cia-scuna figura va sottomessa come singolo file jpg ad alta risoluzione. Al titolo, scritto in grassetto maiuscolo, devono seguire i nomi degli autori (in grassetto), le loro affilia-zioni e l’indirizzo di posta elettronica dell’autore principale. Il testo va organizzato in paragrafi nonnumerati, con titolo in grassetto. Le referenze, elencate alla fine in ordine di citazione, vanno incluse neltesto mediante numeri progressivi inseriti tra parentesi quadre come nell’esempio riportato sotto [1].Onde evitare eccessivo lavoro alla redazione, si raccomanda di fare uso del correttore ortografico; siaccettano anche contributi in inglese. La sottomissione iniziale va effettuata mediante posta elettronica aFrancesca Ballarini ([email protected]) ed eventualmente anche agli altri componenti dellaredazione, i quali riceveranno comunque il manoscritto successivamente per commenti e/o correzioni.
1. A. Aaaaaa, B.bbbbb and C.ccccc (2006), Titolo. Nome della rivista abbreviato Vol, 123-456.
4. Hirose K, Longo DL, Oppenheim JJ, Matsushima K.(1993) Overexpression of mitochondrial manganese supe-roxide dismutasi promotes the survival of tumor cellsexposed to interleukin-1, tumor necrosis factor, selecteddrugs, and ionizing radiation. FASEB J. 7:361–368.
5. Motoori S, Hideyuki JM, Ebara M, Kato H, Hirai F, Kaki-numa S, Yamaguchi C, Ozawa T, Nagano T, Tsujii H, Sai-sho H. (2001) Overexpression of Mitochondrial Manga-nese Superoxide Dismutase Protects against Radiation-induced Cell Death in the Human Hepatocellular Carci-noma Cell Line HLE1. Cancer Res.61, 5382–5388.
6. Sun J, Chen Y, Mingtao L, Zhongliang G. (1988) Role ofantioxidant enzymes on ionizing radiation resistance.Free Rad. Bio.l Med. 24: 586-593.