notes on the unusual megalopae of the ghost crab ocypode quadrata and related species (decapoda:...
TRANSCRIPT

Eagle Hill Institute
Notes on the Unusual Megalopae of the Ghost Crab Ocypode quadrata and Related Species(Decapoda: Brachyura: Ocypodidae)Author(s): John J. McDermottSource: Northeastern Naturalist, Vol. 16, No. 4 (2009), pp. 637-646Published by: Eagle Hill InstituteStable URL: http://www.jstor.org/stable/27744600 .
Accessed: 18/06/2014 07:29
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Eagle Hill Institute is collaborating with JSTOR to digitize, preserve and extend access to NortheasternNaturalist.
http://www.jstor.org
This content downloaded from 185.2.32.60 on Wed, 18 Jun 2014 07:29:44 AMAll use subject to JSTOR Terms and Conditions

2009 NORTHEASTERN NATURALIST 16(4):637-646
Notes on the Unusual Megalopae of the Ghost Crab
Ocypode quadrata and Related Species (Decapoda: Brachyura: Ocypodidae)
John J. McDermott*
Abstract - The postlarval stage, or megalopa, of Ocypode quadrata (Atlantic Ghost
Crab) has distinctive morphological characteristics, with large size being its most
recognizable character. Nevertheless, there is little biological or ecological informa
tion on this critical stage in the crab's life cycle. Over a 20-year period (1980-2000) of field work, only five specimens have been collected incidentally along the sandy beaches of southern New Jersey, near the northern limit of its geographic range.
Megalopae were obtained via seine, hand net, and sediment core samples from Sep tember 6 to October 6, even though some of the locations were sampled throughout the year. Mean carapace width (CW) and length (CL) for the five megalopae were
4.85 ? 0.59 mm by 6.14 ? 0.68 mm, respectively; CW ranged from 4.01-5.43 mm
and CL from 5.18-7.01; mean CW/CL ratio was 0.790 ? 0.023. Carapace dimensions
of New Jersey specimens were compared with megalopae recorded in the literature
from the shores of Long Island, NY and North Carolina, as well as data from two
eastern Pacific species, Ocypode gaudichaudii and O. occidentalis. Morphology of
the megalopa appears to be adaptive for survival through metamorphosis into the first
crab stage. Recruitment of O. quadrata to sandy beaches in the northern part of its
range is probably in the fall.
Introduction
"The large size and peculiar structure of this megalops render it one of the
most interesting forms of the group of larvae to which it belongs. "
S.I. Smith
(1873a) reference to the megalopa o? Ocypode quadrata.
Ghost crabs, Ocypode spp., are well-known, semi-terrestrial, generally warm-water inhabitants of worldwide marine sandy beaches, where older animals live in burrows primarily well above mean high-tide level, and ju veniles occur closer to the water line. Ocypode quadrata (Fabricius) appears to be the only species of this genus found on beaches of the western Atlantic
Ocean, ranging from Long Island Sound, NY to Brazil (Cowles 1908; Haley 1969, 1972; Milne and Milne 1946; Williams 1984). Other species of Ocy
pode are found on the eastern coast of Africa (Monod 1956) and the eastern Pacific and Indo-West Pacific coasts (Crane 1940, Poore 2004).
Laboratory-raised zoeal stages (five instars) and the megalopa of O. quadrata from North Carolina were described by Diaz and Cost
low (1972). They distinguished these larvae from those of four other
*Department of Biology, Franklin and Marshall College, Lancaster, PA 17604-3003;
This content downloaded from 185.2.32.60 on Wed, 18 Jun 2014 07:29:44 AMAll use subject to JSTOR Terms and Conditions

638 Northeastern Naturalist Vol. 16, No. 4
species, O. cordimanus Desmarest, O. gaudichaudii (Milne Edwards and
Lucas), O. occidentalis Stimpson, and O. platytarsis (Milne Edwards).
Although complete zoeal series were not known for the latter four species, the gross morphological characteristics of their distinctive, large megalopae
were very similar to those of O. quadrata.
Little biological information is available for the Ocypode megalopa, con
sidered by Hines (1986) to be among the largest of all known brachyurans
(Fig. 1). However, the megalopae of the vent crab Bythograea thermydron Williams are larger (Williams 1980). Few megalopae of O. quadrata have
been reported from the planktonic zone or at the surf-beach interface where
metamorphosis into the first crab stage takes place (Say 1817; Smith 1873a,
b; but see Smith 1880). They are easily recognized in plankton samples, sieved sediment, or in seine samples because of their extraordinary size,
globose body, large blue eyes and often white or mottled coloration. Thomas
Say (1817), confused as to the nature of preserved museum specimens of
the O. quadrata megalopae, described them (without illustrations) as adult
crabs new to science. The megalopa was named Monolepis inermis (genus: one scale, which refers to caudal lamellae on each side of the telson; spe cies: unarmed or defenseless). Smith (1873a, b) recognized the megalopae he collected in 1870 on Fire Island beach (south shore of Long Island) as
those of O. quadrata, the largest of which was 5.6 mm by 6.4 mm in CW
Figure 1. Photograph of a living megalopa of O. quadrata, antero-dorsal view, col
lected 2007 from a sandy beach in Fort Macon State Park, NC, courtesy of Park
Ranger Randy Newman. Specimen not measured, but was approximately 5 mm wide
(R. Newman, pers. comm.).
This content downloaded from 185.2.32.60 on Wed, 18 Jun 2014 07:29:44 AMAll use subject to JSTOR Terms and Conditions

2009 J.J. McDermott 639
and CL, respectively. He did not give the dimensions of any other specimens. Megalopae of O. quadrata were apparently much less numerous than the just metamorphosed first-crab stages in the same location. The latter were 6.1 to 6.5 mm CW and 5.6 to 6.0 mm CL (width now greater than length). Smith
(1873a, b) described the basic morphological characters of the megalopa (without illustrations) as follows: large eyes; carapace convex above, deep body, posterior broader, no dorsal spine; front sharply deflexed, with long me
dial tooth; sides of body impressed to receive the first three pairs of walking legs; small last pair of legs folded to fit into grooves on the latero-posterior sides of the carapace. He further described the first crab, also without illustra
tions, as follows: carapace broader than long and convex; front broader than in the megalopa; margin of the orbit inclined obliquely backward; posterior pair of walking legs proportionately smaller than in the adult.
Megalopae of O. quadrata were found initially on the shore of Long Island by Smith (1873a, b) in August, and by the middle of September, they
were particularly common. Later Smith (1880) reported that "... a great number of the megalops have been taken in Vineyard Sound by V.N. Ed
wards, and, early in September, 1875,1 took some specimens swimming at the surface in the same region," but no size data were provided. A specimen kept alive for "some days", burrowed in sand and produced a partial molt.
Presently there is not much published information on the role of the mega lopae in the natural history and recruitment of the ghost crab to its sandy beach habitat.
Crane (1940) described and illustrated the megalopae of O. quadrata and those of the eastern Pacific species O. gaudichaudii and O. occidentalis. Her
drawings of O. quadrata correspond in basic morphology to illustrations of Diaz and Costlow (1972) for laboratory-raised specimens. Diaz and Costlow
(1972) showed that megalopae of the eastern Pacific species and those of two Indo-West Pacific species (Raja Bai Naidu 1951, 1954) were very similar, concluding that the Ocypode megalopa was a distinctly unique brachyuran larval stage.
The purpose of this paper is to provide information on the occurrence and carapace dimensions of Ocypode quadrata megalopae collected over a 20-year period (1980-2000) from sandy beaches on the southern coast of New Jersey. These data will be compared with those from the literature on the same and other species within the same subfamily (Ocypodinae), and the
adaptive morphological and behavioral features of these brachyuran mega lopae will be discussed.
Methods
Data on ghost crab megalopae were obtained during organized year long ecological studies of the sandy beach surf zone, primarily in early fall, on the Cape May peninsula (McDermott 1983, 2001). Using a beach seine
This content downloaded from 185.2.32.60 on Wed, 18 Jun 2014 07:29:44 AMAll use subject to JSTOR Terms and Conditions

640 Northeastern Naturalist Vol. 16, No. 4
(15 x 1.5 m; 6.5-mm mesh), a sediment corer, and a hand net, specimens were collected from Stone Harbor and Hereford Inlet on the east shore of
the peninsula, and Delaware Bay on the west shore. Mean surface water
temperature was 21.9 ? 1.3 ?C, similar to fall temperatures measured over
many years. All specimens, except one, were preserved in 10% seawater
formalin and stored in 70% ethanol. All were measured (C W x CL) by ocular
micrometer to 0.01 mm, and CW/CL ratios were determined.
Results
Only five megalopae of Ocypode quadrata were collected during a 20
year span (Table 1). All were found in the months of September and October.
Two were collected in a seine on an exposed sandy beach at Stone Harbor, another in a sediment core sample from the same beach, one with a hand net
in sand inside of Hereford Inlet, and one from a sandy beach in Delaware
Bay. One early crab stage was collected in a seine haul on the Stone Harbor
beach during the same time period. Mean carapace measurements and the mean CW/CL ratio for the five
megalopae are given in Table 1. Ranges in CW and CL were 4.01-5.43 mm
and 5.18-7.01 mm, respectively. All larvae had the morphological char
acteristics described by Crane (1940) and Diaz and Costlow (1972) for
O. quadrata. A megalopa collected 7 September 1999 at Stone Harbor, maintained in the laboratory with periodic feeding oiArtemia nauplii, died
after 34 days without molting. An early crab stage (possibly a first crab) collected 9 September 1994 was 7.52 mm CW and 6.68 mm CL with a 1.13
CW/CL ratio.
Discussion
Morphometric comparisons A comparison of carapace width and length measurements of Ocypode
quadrata with data from the literature (Table 2) shows that the CW/CL
ratio for the one cultured specimen taken from the drawing of Diaz and
Table 1. Carapace dimensions and mean width/length ratios for megalopae of Ocypode quadra ta (Atlantic Ghost Crab) collected on sandy beaches in New Jersey.
Megalopa Date of Collection _Carapace_ number* collection site** Width (mm) Length (mm) W/L ratio
1 6 Sep 1980 DB 530 6AS (X82 2 19 Sep 1981 HI 5.01 6.18 0.81
3 13 Sep 1985 SH 4.01 5.18 0.77
4 7 Sep 1999 SH 5.43 7.01 0.78
5 6Oct2000 SH 4.51 5.85 0.77
MeaniSD 4.85 ? 0.59 6.14 ?0.69 0.790 ? 0.023
* Listed chronologically. ** DB = Delaware Bay, HI = Hereford Inlet, SH - Stone Harbor.
This content downloaded from 185.2.32.60 on Wed, 18 Jun 2014 07:29:44 AMAll use subject to JSTOR Terms and Conditions

2009 J.J. McDermott 641
Costlow (1972) corresponds to the mean value for the five megalopae from New Jersey, but not those of Smith (1873a, b) and Crane (1940).
Carapace dimensions for the two eastern Pacific species of Ocypode (Ta ble 2) show that CL is also greater than CW, but the ratios appear slightly
higher than for those of O. quadrata. Smith (1873a) reported that the
largest specimen of O. quadrata from Fire Island beach, NY was 5.6 mm
CW by 9.4 mm CL for a ratio of 0.596. Smith (1873b) recorded the larg est megalopa from Fire Island as 5.6 by 6.4 mm. As his two papers were
nearly identical except for the 9.4 mm value, most likely a measurement
or typographical error was involved.
Carapace measurements of the small juvenile Ocypode quadrata from
New Jersey implied that at metamorphosis (or shortly thereafter) the cara
pace width became the greater dimension as is also the case in the adult
(Williams 1984). However, it is possible that this crab was not the first
crab instar. On Fire Island beach, Smith (1973a, b) measured the carapace of the smallest crabs, which ranged from 6.1 to 6.5 mm CW by 5.6 to 6.0 mm CL (number of crabs measured not given). Thus, my specimen (7.52 x 6.68 mm) was most likely a second instar. In contrast, Haley (1972) measured what he considered to be a first crab instar from a summer
brood of O. quadrata specimens along the middle Texas coast at a CW of 7 mm in October. However, Haley measured only one of these first crabs, and unless megalopae of O. quadrata are larger in this southern part of their range, the 7-mm CW individual may also represent a second instar.
Alternatively, 7 mm CW could have been the upper CW limit of first
stage crabs at his field site. Hines (1986) sought correlations between larval patterns and life-history
characteristics among seven families of brachyurans, which included the
Ocypodidae. In his analysis, megalopa size (CW) was positively correlated
Table 2. Carapace dimensions and mean width/length ratios, determined by other authors, for the
megalopae o? Ocypode quadrata and two species within the same genus from the eastern Pacific.
Carapace
Species
No. of
specimens
Width (mm)
Length
(mm) W/L ratio References
O. quadrata* 1 5.6 6.4 0.88 Smith (1873a, b) 3** 4.9 5.9 0.83 Crane (1940)
1 5.9 7.4 0.79 Diaz and Costlow (1972)*** O. gaudichaudii If 4.7 5.6 0.85 Crane (1940)
5**n 4.6 5.3 0.87 Crane (1940) O. occidentalis lw 4.3 5.0 0.86 Crane (1940)
*all specimens from northwestern Atlantic Ocean, United States. **size ranges given from which "mean values" were determined.
**Measurements from their line drawing, Figure 9.
Specimen from Costa Rica,
specimens from Colombia.
mspecimen from Mexico.
This content downloaded from 185.2.32.60 on Wed, 18 Jun 2014 07:29:44 AMAll use subject to JSTOR Terms and Conditions

642 Northeastern Naturalist Vol. 16, No. 4
with adult crab size only if data for the unusually large megalopae of three
species o? Ocypode species were eliminated from the other crab species, i.e., Uca spp., within the Ocypodidae.
Natural history of the megalopae Say (1817) collected "several specimens" of the megalopae o? Ocypode
quadrata on the eastern sandy shore of Maryland. Deposited on the beach as the tide receded, they burrowed into the sand, protecting them from sun
and minimizing water loss, presumably prior to metamorphosis. Smith
(1873a, b) collected megalopae in late August at Fire Island beach. He observed "many more young crab instars" that had metamorphosed from
megalopae. He conjectured that molting as soon as possible would tend to minimize mortality in a terrestrial environment. Smith further noted that the megalopae of late August were only half the size of those collected in the middle of September, suggesting that the disparity in size might be due to multiple broods during the reproductive season. The present data from
New Jersey beaches and those of Smith for the beaches of Long Island sug gest that early fall may be the usual period for recruitment O. quadrata in this section of the Atlantic coast.
Crane ( 1940) studied the morphology of megalopal stages of three spe cies of Ocypode. She concluded that the obese megalopae of Ocypode had a thicker exoskeleton than megalopae of other brachyuran species. This fea ture and the curious behavior of folding their five pairs of legs close to the
c?phalothorax (Fig. 2) where grooves accommodate these folded legs (Crane 1940, Diaz and Costlow 1972), led Crane to see these as adaptive character istics to coming ashore in the turbulent surf zone of sandy beaches. With legs folded, the animal becomes semispherical and compact (Fig. 2b), preventing legs from injury as it is rolled by the surf onto the beach. The thick exoskel eton would give added protection to the delicate internal organs, and the
capacity of the megalopa to burrow at least superficially into the sand would serve to minimize water loss. Once on shore, it would seem imperative that
megalopae transform as soon as possible into the semi-terrestrially adapted first-crab stage with its functional walking legs and feeding appendages.
Are the megalopae of ghost crabs that are deposited on the shore com
petent to metamorphose, i.e., have they reached the developmental stage, as megalopae, necessary for a successful molt? Diaz and Costlow (1972) showed that the time for development of the megalopa in the laboratory was a minimum of 34 days following hatching as first zoeae. Their megalopae, however, failed to metamorphose into first crabs. Possible chemical or physi cal cues to induce metamorphosis were not considered by Diaz and Costlow.
Therefore, the length of time required for the megalopa to reach competency is not known.
Presumably, megalopae are tossed onto less favorable shore locations than the preferred sandy beaches. Can competent megalopae of Ocypode
This content downloaded from 185.2.32.60 on Wed, 18 Jun 2014 07:29:44 AMAll use subject to JSTOR Terms and Conditions
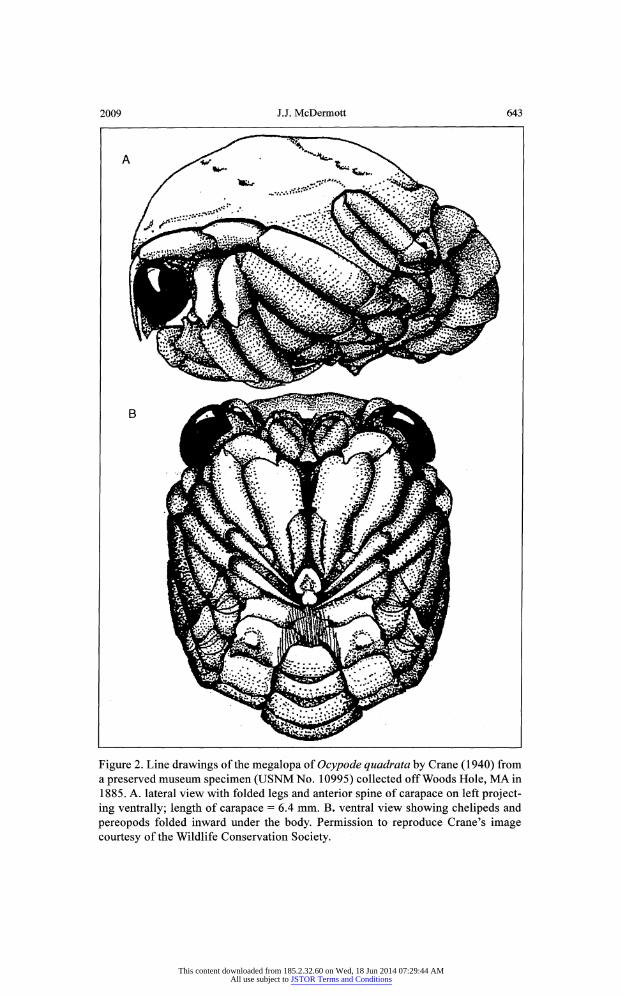
2009 J.J. McDermott 643
Figure 2. Line drawings of the megalopa of Ocypode quadrata by Crane (1940) from a preserved museum specimen (USNM No. 10995) collected off Woods Hole, MA in 1885. A. lateral view with folded legs and anterior spine of carapace on left project
ing ventrally; length of carapace = 6.4 mm. B. ventral view showing chelipeds and
pereopods folded inward under the body. Permission to reproduce Crane's image
courtesy of the Wildlife Conservation Society.
This content downloaded from 185.2.32.60 on Wed, 18 Jun 2014 07:29:44 AMAll use subject to JSTOR Terms and Conditions

644 Northeastern Naturalist Vol. 16, No. 4
postpone metamorphosis after landing in such unfavorable locations? Will
they reenter the water column to be carried elsewhere to metamorphose? I
have not been able to find any information on delayed metamorphosis in
Ocypode. The fact that one of the megalopae collected in New Jersey was
able to postpone metamorphosis for 34 days indicates that such a process is possible and may be adaptive. Certainly, the adaptive nature of delayed
metamorphosis has been demonstrated for many crustaceans and other major invertebrate groups (Bishop et al. 2006, Butman 1987, Pechenik 1990).
Do megalopae have some mechanism for remaining planktonic in order to
minimize being swept ashore before competency? Chemical or physical en
vironmental cues on sandy beaches that might control metamorphosis should
be considered (Steinberg et al. 2007, 2008). There is much to learn about the enigmatic, bug-eyed megalopa of Ocy
pode and its critical role in the recruitment of new generations to sandy beaches. Metamorphic data are needed on the size range of this larval stage in relation to its metamorphosis into the first crab. How do megalopae man
age to return from a planktonic existence to populate sandy beaches? Are the
unusually large eyes of megalopae of particular importance in this naviga tion? How do they stay out of the surf zone until they are competent to molt
into first crabs? More intense field studies and laboratory observation of
cultured larvae are needed to answer these and other questions dealing with
the natural history of ghost crabs.
Acknowledgments
I thank the following for checking their crustacean collections for megalopae of
Ocypode quadrata: J. Dineen, Smithsonian Marine Station, Fort Pierce, FL; A.H.
Hines, Smithsonian Environmental Research Center, Edgewater, MD; and D.M.
Knott, Southeast Regional Taxonomic Center Marine Resources Research Insti
tute, Charleston, SC. D.M. Knott also provided me with a photo of an O. quadrata
megalopa, which was taken by R. Newman, a park ranger at Macon State Park, NC.
Critical reviews of an early draft of this paper by J. Dineen and D.M. Knott were most
helpful, as were the comments on the manuscript by two anonymous reviewers. I ap
preciate discussions with W.S. Johnson, Goucher College, Towsen, MD, concerning his Zooplankton studies along the mid-Atlantic coast. J. Norenburg and C. Walter,
Smithsonian Institution, Washington, DC, were helpful in retrieving information
from files of the US National Museum, and J.T. Thompson, Franklin and Marshall
College, provided technical assistance. Franklin and Marshall College continues to
aid and encourage my research.
Literature Cited
Bishop, CD., M.J. Huggett, A. Heyland, J.Hodin, and B.P. Brandhorst. 2006. In
terspecific variation in metamorphic competence in marine invertebrates: The
significance for comparative investigations into the timing of metamorphosis.
Integrative and Comparative Biology 46:662-682.
This content downloaded from 185.2.32.60 on Wed, 18 Jun 2014 07:29:44 AMAll use subject to JSTOR Terms and Conditions

2009 J.J. McDermott 645
Butman, CA. 1987. Larval metamorphosis of soft sediment invertebrates: The
spatial scales of pattern explained by active habitat selection and the emerging role of hydrodynamical processes. Oceanography and Marine Biology Annual
Review 25:113-165.
Cowles, R.P. 1908. I. Habits, reactions, and associations in Ocypoda arenaria. Pa
pers from the Tortugas Laboratory of the Carnegie Institution of Washington No.
3,2:1-41.
Crane, J. 1940. Eastern Pacific Expedition of the New York Zoological Society. XVIII. On the post-embryonic development of brachyuran crabs of the genus
Ocypode. Zool?gica 25:65-82.
Diaz, H., and J.D. Costlow. 1972. Larval development o? Ocypode quadrata (Brachyu ra: Crustacea) under laboratory conditions. Marine Biology 15:120-131.
Haley, S.R. 1969. Relative growth and sexual maturity of the Texas Ghost Crab, Ocy
pode quadrata (Fabr.) (Brachyura, Ocypodidae). Crustaceana 17:285-297.
Haley, S.R. 1972. Reproductive cycling in the ghost crab, Ocypode quadrata (Fabr.)
(Brachyura, Ocypodidae). Crustaceana 23:1-11.
Hi?es, A.H. 1986. Larval patterns in the life histories of brachyuran crabs (Crusta
cea, Decapoda, Brachyura). Bulletin of Marine Science 39:444-466.
McDermott, J.J. 1983. Food web in the surf zone of an exposed sandy beach along the mid-Atlantic coast of the United States. Pp. 529-538, In A. McLachlan and
T. Erasmus (Eds.). Sandy Beaches as Ecosystems. Dr. W Junk, The Hague, The
Netherlands. 757 pp.
McDermott, J.J. 2001. Biology oiChiridotea caeca (Say, 1818) (Isopoda: Idoteidae) in the surf zone of exposed sandy beaches along the coast of southern NewJersey,
USA. Ophelia 55:123-135. Milne, L.J., and M.J. Milne. 1946. Notes on the behavior of the ghost crab. American
Naturalist 80:362-380.
Monod, T. 1956. Hippidea et Brachyura ouest-africains. M?moires de L'Institut
Fran?ais D'Afrique Noire No. 45:1-674.
Pechenik, J.A. 1990. Delayed metamorphosis by larvae of benthic marine inverte
brates: does it occur? Is there a price to pay? Ophelia 32:63-95.
Poore, G.C.B. 2004. Marine Decapod Crustacea of Southern Australia: A Guide
to Identification. CSIRO Publishing, Collingwood, Victoria, Australia. 574
pp., 32 pis.
Raja Bai Naidu, K.G. 1951. Some stages in the development and bionomics of Ocy
pode platy tar sis. Proceedings of the Indian Journal of Sciences 33:32-40, 3 pis.
Raja Bai Naidu, K.G. 1954. The post-larval development of the shore crab Ocypode
platytarsis (M. Edwards) and Ocypode cardimana (Demarest). Proceedings of
the Indian Journal of Sciences (Section B) 40:89-101.
Say, T. 1817. An account of the Crustacea of the United States. Journal of the Acad
emy of Natural Sciences of Philadelphia 1:155-160 Smith, S.I. 1873a. III. Zoology and botany. 1. The megalops stage of Ocypoda.
American Journal of Science 3:67-68.
Smith, S.I. 1873b. The metamorphoses of the lobster, and other Crustacea. Pp.
522-537, In A.E. Verrill (Ed.). Report Upon the Invertebrate Animals of Vine
yard Sound and the Adjacent Waters, with an Account of the Physical Characters
of the Region. In S.F. Baird (Ed.). Report on the Conditions of the Sea Fisheries
of the South Coast of New England in 1871 and 1872. United States Commission of Fish and Fisheries, Washington, DC.
This content downloaded from 185.2.32.60 on Wed, 18 Jun 2014 07:29:44 AMAll use subject to JSTOR Terms and Conditions

646 Northeastern Naturalist Vol. 16, No. 4
Smith, S.I. 1880. Occasional occurrence of tropical and subtropical species of de
capod Crustacea on the coast of New England. Transactions of the Connecticut
Academy of Arts and Sciences 4:254-267.
Steinberg, M.K., CE. Epifanio, and A.U. Andon. 2007. A highly specific chemical
cue for metamorphosis of the Asian Shore Crab, Hemigrapsus sanguineus. Jour
nal of Experimental Marine Biology and Ecology 347:1-7.
Steinberg, M.K., L.S. Krimsky, and CE. Epifanio. 2008. Induction of metamorpho sis in the Asian Shore Crab Hemigrapsus sanguineus: Effects of biofilms and
substratum texture. Estuaries and Coasts 31:738-744.
Williams, A.B. 1980. A new crab family from the vicinity of submarine thermal vents
on the Galapagos Rift (Crustacea: Decapoda: Brachyura). Proceedings of the
Biological Society of Washington 93:443-472. Williams, A.B. 1984. Shrimps, Lobsters, and Crabs of the Atlantic Coast of the East
ern United States, Maine to Florida. Smithsonian Institution Press, Washington, DC. 550 pp.
This content downloaded from 185.2.32.60 on Wed, 18 Jun 2014 07:29:44 AMAll use subject to JSTOR Terms and Conditions