nmr spektroskopie biologicky aktivních molekul - vscht.cz · strategie pro určování struktur...
TRANSCRIPT

NMR spektroskopie biologicky aktivních molekul

Jak vidí současné a budoucí uplatnění NMR spektroskopie profesor Richard Ernst.
J.W. Emsley:“NMR started as the plaything of the physicists, it became the favourite toy of the chemists and finally went on to seduce biochemists.”
MedicineBiochemistry
Chemistry
Physics
Nobel prize in chemistry 1991

Kurt WüthrichNobel Price Winner in Chemistry 2002
G. Wagner, K. Wüthrich. 1982. Sequential resonance assignments in protein 1Hnuclear magnetic resonance spectra. Basic pancreatic trypsin inhibitor.J. Mol. Biol. 155, 347-366.

1. Jaké typy biologický aktivních molekul ?
peptidy a proteiny
nukleové kyseliny
oligosacharidy
2. Jaký typ informace může být pomocí NMR získán?
identifikace substrátu
prostorová struktura molekuly
studium dynamického chování systému
prostorová struktura komplexu
zkoumání vazby ligandu a substrátu
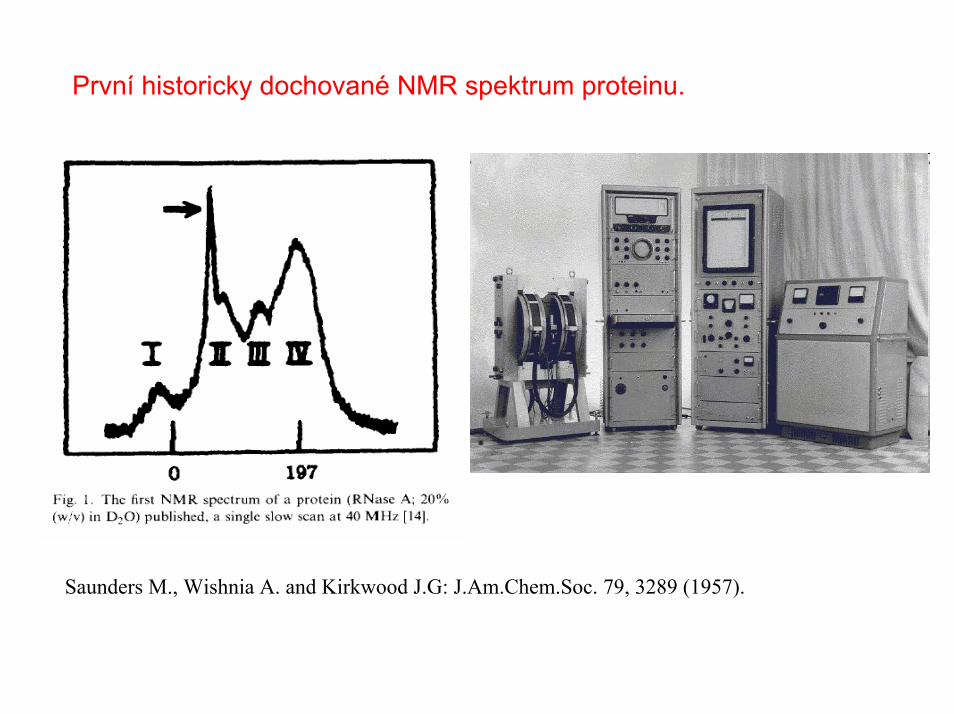
První historicky dochované NMR spektrum proteinu.
Saunders M., Wishnia A. and Kirkwood J.G: J.Am.Chem.Soc. 79, 3289 (1957).

900 MHz 1H NMR spektrumlysozymu
900 MHz magnet firmy Bruker
900 MHz magnet firmy Varian

Strategie pro určování struktur biomolekul
Obecné informace o molekule (primární struktura, kovalentnívazby…)
NMR vzorek
NMR experimenty
NMR spektra Přiřazení signálů
Přiřazení experimentálních NMR parametrů (NOE…)
Odhad přibližné struktury
Oprava přiřazení NMR parametrů, signálů
Zhodnocení kvality struktur
Výpočet souboru struktur
Výpočet statistických údajůpro soubor konečných struktur
Výpočet NOESY spekterPorovnání s databázemi(Procheck, Whatif….)

Příprava vzorku proteinu pro NMR měření
1. Získání DNA proteinu
2. Příprava plasmidové DNA
3. Exprese rekombinantního proteinu v E.Coli
4. Izolace a čištění
5. Zakoncentrování vzorku
6. Testování vzorku na dlouhodobou stabilitu
7. Zopakování procesu s médiem obohaceným o izotopy 13C, 15N,případně i 2H

Vzorek pro NMR experimenty
Úspěšné řešení bezpodmínečněvyžaduje kvalitní spolupráci mezi NMR spektroskopiky a biochemiky !
rozpouštědlo H2O, resp. 90-95% H2O a 5-10% D2O
Vzorek musí zůstat aktivní a nedenaturovaný během NMR experimentů!
pH kompromis mezi minimalizací chemické výměny mezi signály labilních protonů a signálem vody a optimem pro studovaný protein (4.0 - 7.0)
pufr fosfátový pufr neobsahuje žádné protonyacetátový pufr (nutno připravit deuterovaný)
teplota podle požadavků studovaného materiálu (15 – 40°C)
aditiva nutná aditiva je možné zaměnit za deuterovaná analoga
koncentrace pro NMR experimenty musí být v rozsahu alespoň 0.5-2.0 mM, vzoreknesmí podléhat agregaci, koagulaci, sebezničení v tomto konc. rozmezí
stabilita nutná dlouhodobá stabilita v rozsahu minimálně několika týdnů
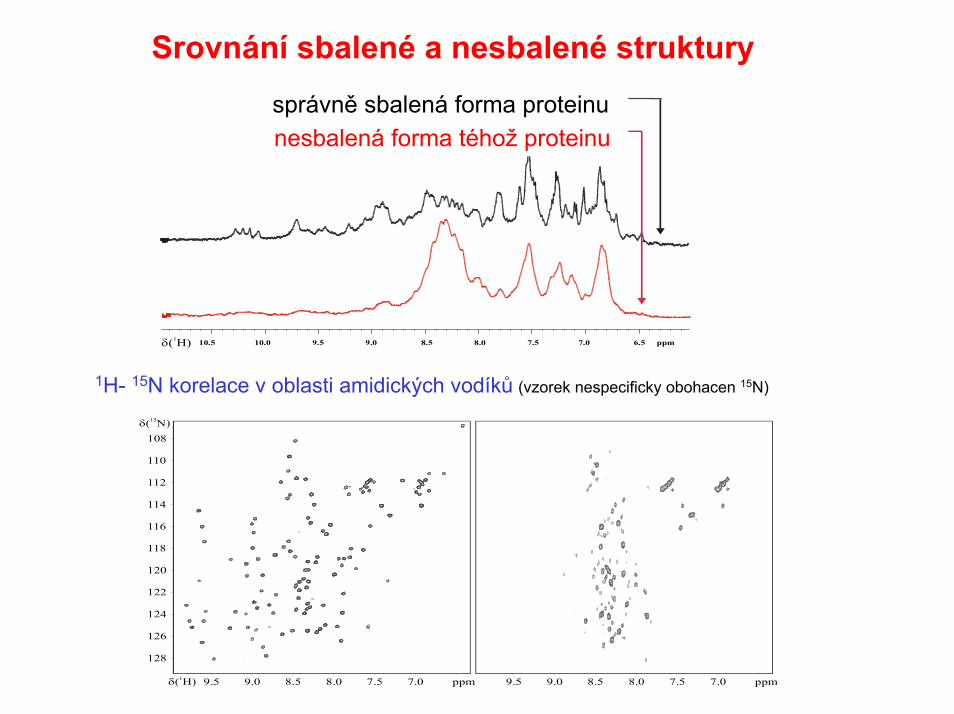
Srovnání sbalené a nesbalené struktury
10.5 10.0 9.5 9.0 8.5 8.0 7.5 7.0 6.5 ppm
WVQPI 107 AA (12 kDa) IMMCS
WVQPI 107 AA (12 kDa) IMMCS
83 AA (9 kDa)
δ(1H)
správně sbalená forma proteinu nesbalená forma téhož proteinu
1H- 15N korelace v oblasti amidických vodíků (vzorek nespecificky obohacen 15N)
7.07.58.08.59.09.5 ppm
108
110
112
114
116
118
120
122
124
126
128
7.07.58.08.59.09.5 ppmδ(1H)
δ(15N)

Strategie pro určování struktur biomolekul
Obecné informace o molekule (primární struktura, kovalentnívazby…)
NMR vzorek
NMR experimenty
NMR spektra Přiřazení signálů
Přiřazení experimentálních NMR parametrů (NOE…)
Odhad přibližné struktury
Oprava přiřazení NMR parametrů, signálů
Zhodnocení kvality struktur
Výpočet souboru struktur
Výpočet statistických údajůpro soubor konečných struktur
Výpočet NOESY spekterPorovnání s databázemi(Procheck, Whatif….)

Biomolekulární NMR spektroskopie: měřená jádra
vysoké přirozené zastoupení (99.98%)vysoká citlivost (1.00)malá disperze chemických posunů NMR signálů (~15.0 ppm)
velká disperze chemických posunů NMR signálů (~200.0 ppm)nízké přirozené zastoupení (1.108%), možné uměle navýšit až na 100%nízká citlivost (1.76x10-4), po 100%ním izotopovém obohacení(1.59x10-2)
menší počet atomů než 13Cstřední disperze chemických posunů NMR signálů (~30.0 ppm) (oproti 13C nezávislost na typu aminokyseliny)nízké přirozené zastoupení (0.37%), možné uměle navýšit až na 100%velmi nízká citlivost (3.85x10-6), po 100%ním izotopovém obohacení (1.04x10-3)
používá se pro speciální účely
1H
13C
15N
2H

Potlačení signálu vody
Signál H2O je 104-105 násobně intenzivnější než odezva měřené molekuly.
Metoda presaurace
Během relaxační doby ozařujeme signál vody slabým RF polem.
Proč H2O?1. Voda je fyziologické prostředí2. Nelze použít D2O z důvodů chemické výměny s amidickými protony.
CW -ozařování
90 deg

1H spektrum proteinu po presaturaci H2O
zbytkový signál H2O
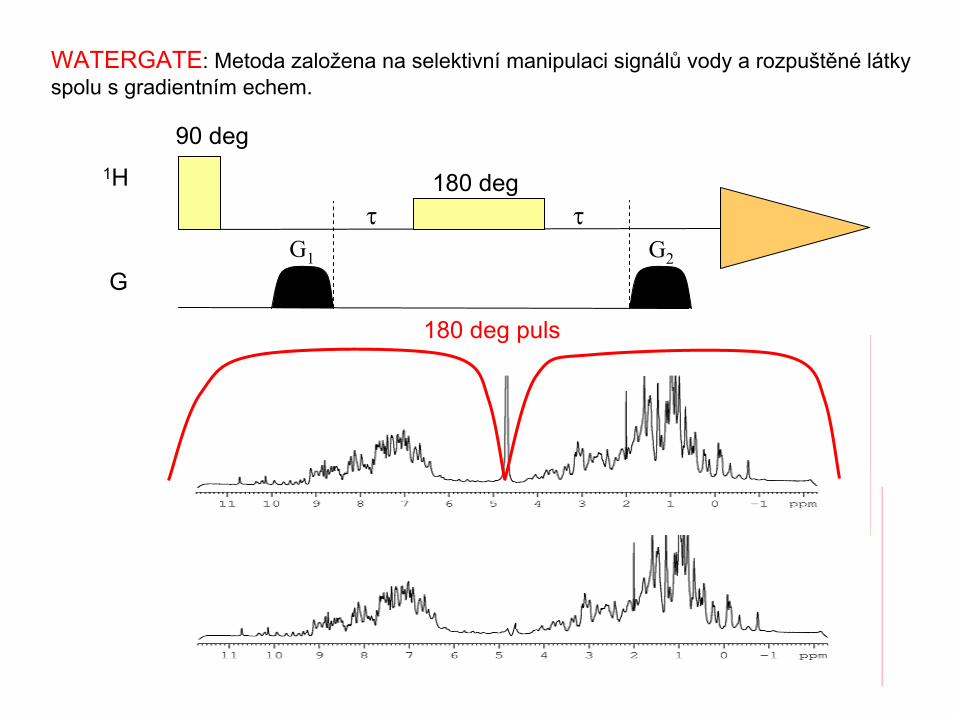
WATERGATE: Metoda založena na selektivní manipulaci signálů vody a rozpuštěné látky spolu s gradientním echem.
90 deg
180 deg puls
τ τG1 G2
180 deg1H
G

Potlačení signálu vody: metoda WATERGATE
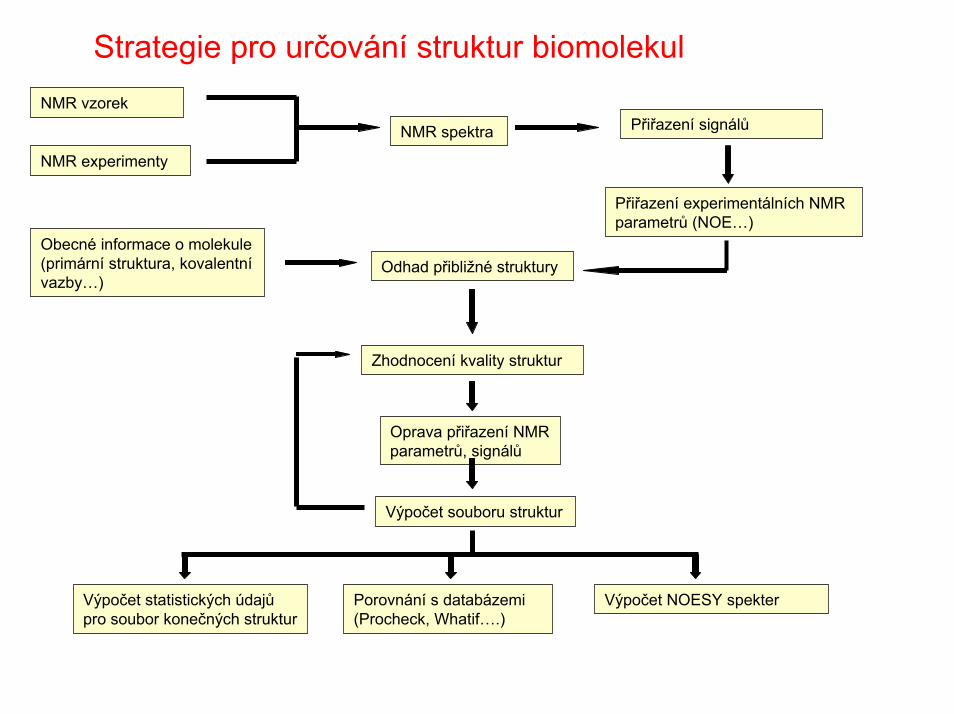
Strategie pro určování struktur biomolekul
Obecné informace o molekule (primární struktura, kovalentnívazby…)
NMR vzorek
NMR experimenty
NMR spektra Přiřazení signálů
Přiřazení experimentálních NMR parametrů (NOE…)
Odhad přibližné struktury
Oprava přiřazení NMR parametrů, signálů
Zhodnocení kvality struktur
Výpočet souboru struktur
Výpočet statistických údajůpro soubor konečných struktur
Výpočet NOESY spekterPorovnání s databázemi(Procheck, Whatif….)
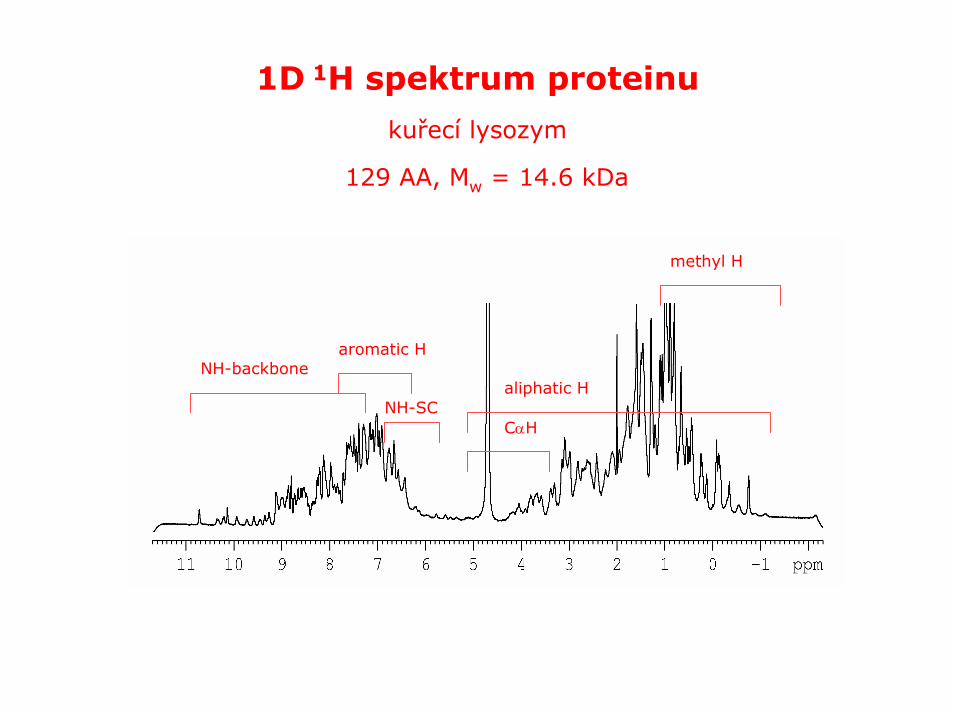
1D 1H spektrum proteinu
kuřecí lysozym
129 AA, Mw = 14.6 kDa
aromatic HNH-backbone
NH-SCCαH
aliphatic H
methyl H

Multidimensionální NMR spektroskopie jakonástroj pro zjednodušení spekter
3D
F1(1H)
1D
F2(X)2D
F2(1H)F1(1H/X) F1(1H/X)
F3(1H)
Lepší rozlišení je ve vícedimenzionálních spektrech zajištěno využitím izotopového obohacení 15N a 13C.
F3(X)4D
F2(X)
F1(1H)F4(1H)

Přiřazování rezonancí• NMR experimenty pro přiřazení signálů pracují se dvěma nebotřemi různými jádry najednou (experimenty s trojnásobnou rezonancí),tato jádra jsou navzájem zkorelována.
• Názvy takovýchto experimentů se tvoří podle typu jader, která korelují:
HNCA koreluje amidický vodík s příslušným dusíkem a uhlíkem vpozici α.
HN(CO)CA koreluje stejné typy atomů (jader) jako HNCA, ale přesCO. To naznačuje směr korelace, tj. H a N i-té aminokyseliny a Cα aminokyseliny v pozici i-1.
• Směr přenosu magnetizace je v případě těchto experimentůH N Cα a zpět. Experimenty se nazývají „out and back“
• Naproti tomu přenos magnetizace u experimentů např. CBCA(CO)NHzačíná na atomu CΒ (i-1) aminokyseliny a končí na amidickém Haminokyseliny následující, tj. experimenty „out and stay“.

Přiřazování rezonancí
13Cβ13Cβ
13Cγ
13C’ 15N
Hβ Hβ
Hα HαHN
13C’13Cα13Cα
55Hz 15Hz
90Hz
35Hz
7Hz<1Hz
130Hz
140Hz
35Hz
35Hz
11Hz 55Hz
HNCA experiment
aminokyselinový zbytek Iaminokyselinový zbytek I-1

HNCA experiment
Korelace ve spektru: HN
i - Ni - Cαi
HNi - Ni – Cα
i-1-1
x x φ2 x x x x x x
t2/2
1H
φ1 xφ4
xx
τ τ τ τδδt1/2 t1/2
x
t2/2
x
x x
acq t3
dec
φ4
15N
φ3
13Cα
13CO
vývojová periodapřenos magnetizace

Strategie pro určování struktur biomolekul
Obecné informace o molekule (primární struktura, kovalentnívazby…)
NMR vzorek
NMR experimenty
NMR spektra Přiřazení signálů
Přiřazení experimentálních NMR parametrů (NOE…)
Odhad přibližné struktury
Oprava přiřazení NMR parametrů, signálů
Zhodnocení kvality struktur
Výpočet souboru struktur
Výpočet statistických údajůpro soubor konečných struktur
Výpočet NOESY spekterPorovnání s databázemi(Procheck, Whatif….)

Konstrukce multidimensionálních NMR spekter3D HNCA
F2(15N )
I
I-1
F1(13Cα)
F2(15N )
F1(13Cα)
F3(1HN)
F3(1HN)
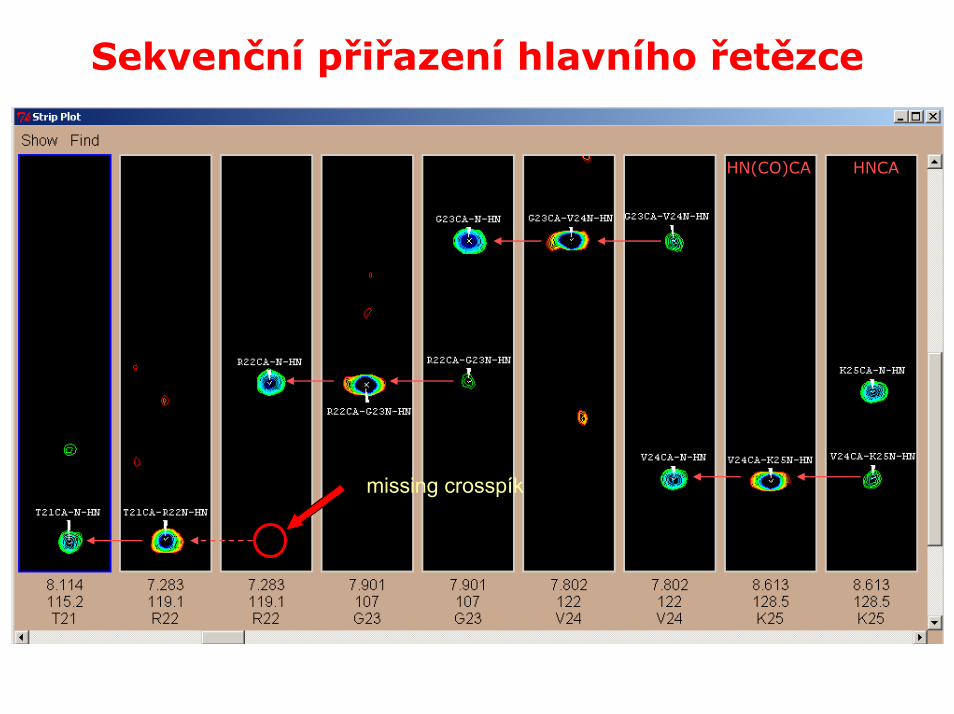
Sekvenční přiřazení hlavního řetězce
HNCAHN(CO)CA
missing crosspík

Přiřazování rezonancí
13Cβ13Cβ
13Cγ
13C’ 15N
Hβ Hβ
Hα HαHN
13C’13Cα13Cα
55Hz 15Hz
90Hz
35Hz
7Hz<1Hz
130Hz
140Hz
35Hz
35Hz
11Hz 55Hz
HN(CO)CA experiment

Konstrukce multidimensionálních NMR spekter3D HNCA/HN(CO)CA
F3(1HN)
F2(15N )
F1(13C)
F2(15N )
I-1
F2(15N )
I
I-1
F1(13C)
F3(1HN)

Sekvenční přiřazení hlavního řetězce
HNCAHN(CO)CA
missing crosspík

Přiřazování rezonancí postranních řetězců
Cβ
C’ N
Hβ Hβ
HαHN
C’Cα
Hα
Cα
CγHγ Hγ
CβHβ Hβ
CγHγ Hγ
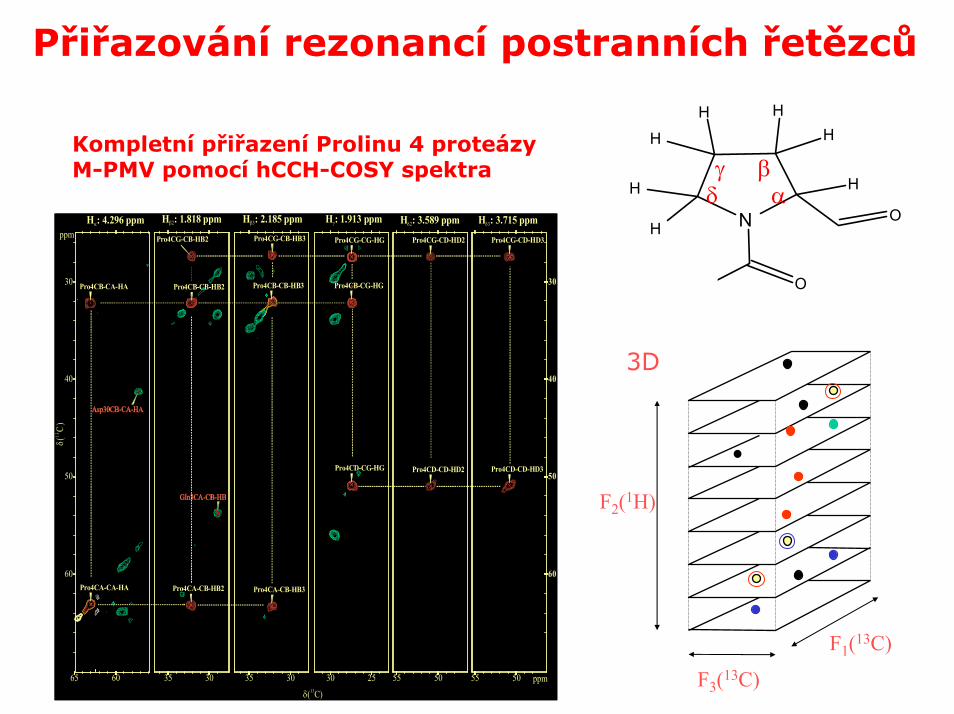
Přiřazování rezonancí postranních řetězců
ON
H
HH
H
H
H
H
OCH 3
αβγ
δ
Kompletní přiřazení Prolinu 4 proteázyM-PMV pomocí hCCH-COSY spektra
55 50 55 50
60
50
40
30
35 30 35 30 30 2565 60
60
50
40
30
ppm
ppm
Pro4CG-CD-HD2
Pro4CD-CD-HD2
Pro4CG-CD-HD3
Pro4CD-CD-HD3
Pro4CA-CB-HB2
Pro4CG-CB-HB2
Pro4CB-CB-HB2
Pro4CA-CB-HB3
Pro4CB-CB-HB3
Pro4CG-CB-HB3
Pro4CB-CG-HG
Pro4CD-CG-HG
Pro4CG-CG-HG
Pro4CB-CA-HA
Pro4CA-CA-HA
H : 4.296 ppmα H : 1.818 ppmβ2 H : 2.185 ppmβ3 H : 1.913 ppmγ H : 3.589 ppmδ2 H : 3.715 ppmδ3
δ(13
C)
δ(13C)
3D
F2(1H)
F1(13C)
F3(13C)

Práce s extra velkými molekulami Mw> 25 kDa
Práce s velkými molekulami způsobuje dvojí komplikaci
• velmi komplikovaná spektra
• rychlá spin-spinová relaxace
)](....'[8 6
22)(
cfsJrCH
CDH τγγ h=2R
γH/ γD ~ 6.6
Řešení: výměna atomů vodíku za deuterium
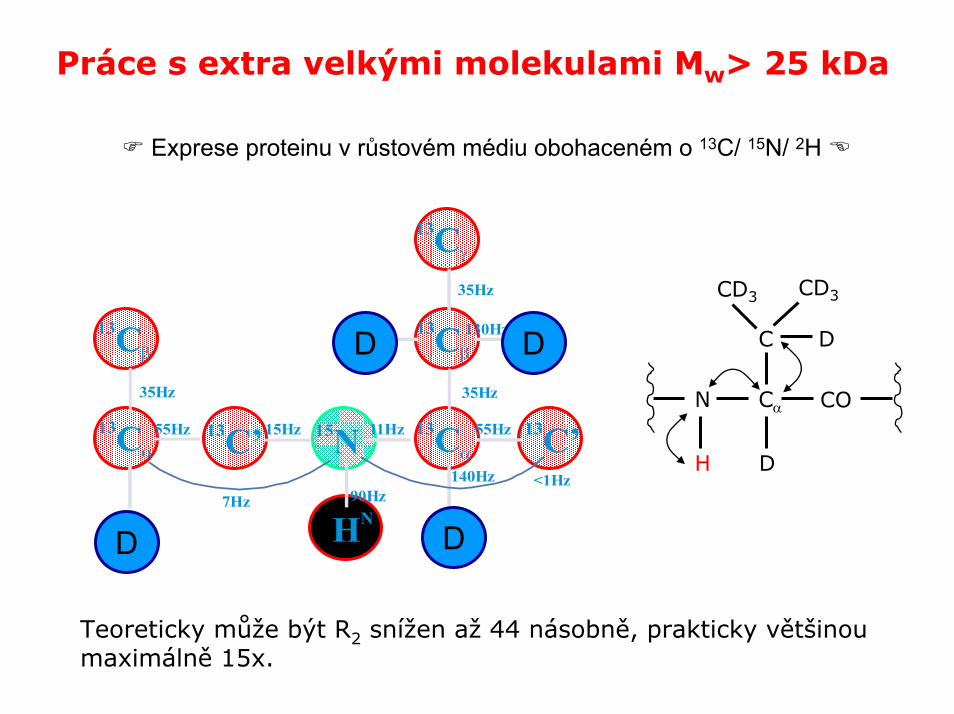
Práce s extra velkými molekulami Mw> 25 kDa
Exprese proteinu v růstovém médiu obohaceném o 13C/ 15N/ 2H
13Cβ13Cβ
13Cγ
13C’ 15N
Hβ Hβ
Hα HαHN
13C’13Cα13Cα
55Hz 15Hz
90Hz
35Hz
7Hz<1Hz
130Hz
140Hz
35Hz
35Hz
11Hz 55Hz
D D
DD
CD3 CD3
C D
Cα
D
CON
H
Teoreticky může být R2 snížen až 44 násobně, prakticky většinou maximálně 15x.

Fully protonated versus perdeuterated EIN protein

Fully protonated versus perdeuterated EIN protein
Missing crosspeaks are marked

Strategie pro určování struktur biomolekul
Obecné informace o molekule (primární struktura, kovalentnívazby…)
NMR vzorek
NMR experimenty
NMR spektra Přiřazení signálů
Přiřazení experimentálních NMR parametrů (NOE…)
Odhad přibližné struktury
Oprava přiřazení NMR parametrů, signálů
Zhodnocení kvality struktur
Výpočet souboru struktur
Výpočet statistických údajůpro soubor konečných struktur
Výpočet NOESY spekterPorovnání s databázemi(Procheck, Whatif….)

Nukleární Overhauserův efekt
σIS - rychlost křížové relaxace, nárůstu NOEτc - korelační časrIS - meziatomová vzdálenostω - pracovní frekvence NMR spektrometru
HH
rIS < 5Ǻ
dipól - dipólováinterakce mezi atomy
62
422
416
104−
2 ⎥⎦
⎤⎢⎣
⎡−
+⎟⎠⎞
⎜⎝⎛= ISc
c
cIS rh τ
τωτγ
πµσ ο
6−
⎟⎟⎠
⎞⎜⎜⎝
⎛=
cal
IS
cal
IS
rr
σσ { } 6−
⎟⎟⎠
⎞⎜⎜⎝
⎛=
cal
IS
cal
I
rr
fSf
Poměr intenzit NOE efektů fI{S}/fcal je úměrný poměru vzdáleností příslušných atomů vodíku pouze pro velmi krátké časy!!!

Převod intenzity NOE krospíků na vzdálenost mezi atomy.
Dolní mez :1.8 ǺJedná se o součet vzdáleností van der Waalsovskýchpoloměrů dvou interagujících atomů vodíku
Horní mez :Nastavuje se podle intenzity příslušného krospíku. Pro větší molekuly se používá max. vzdálenost až 6 Å.
1.8 Ǻ ≤ r ≤ 2.5 Ǻ
1.8 Ǻ ≤ r ≤ 3.5 Ǻ
1.8 Ǻ ≤ r ≤ 5.0 Ǻ

Editovaná NOESY spektra4D 13C/ 15N-editované NOESY
1H1H
13C
15N
NOE1H 1H
15N 13CJHCJHN
15N= 106.4 ppm 15N= 106.4 ppm13C= 45.8 ppm
15N= 106.4 ppm13C= 56.1 ppm
G78HN-G78Hα G78HN-S77Hα
3D 15N-editované NOESY 4D 13C/15N-editované NOESY

Nepřímá spin-spinová interakční konstantaExperimentální omezení dihedrálních úhlů
Karplusova rovnice3J = A cos2Θ + Β cosΘ + C
Vztah mezi interakční konstantou a dihedrálními úhly peptidu
3J
H
O
O
N
Cα
CβC
C
ψ
φχ1
χ2
Cγ
H
H
H
ω
[Hz]-120 600 120
Θ deg
H-NCα-H
CO-NCα-H
H-NCα-CβH-NCα-CO
10
8
6
4
2
0

Typické hodnoty interakčních konstant 3JHH pro dihedrální úhel φ
α-helix φ ∼ − 60 deg
3 ≤ J ≤ 6 Hz
typické nastavení pro úhel φ:
−110 ≤ φ ≤ −10 deg
β-struktura skládanéholistu φ ∼ −120ο
6 ≤ J ≤ 9 Hz
typické nastavení pro úhel φ:
−170 ≤ φ ≤ −70 deg

Stereospecifické přiřazení diastereotopních atomů v CβH2 skupinách
- Jαβ coupling- HN-Hβ NOE
N
COHα
R
Hβ3
Hβ2
N
COHα
R
Hβ2 Hβ3
N
COHα
RHβ3
Hβ2
tg (g-) gg (g+)gt (t)
Jαβ = 9.5 cos2θ −1.6 cos θ +1.8
0 5 10 15
Jαβ2
0
180 (gt)
60 (gg) -60 (tg)
0
15
10
Jαβ
3 5

Vodíkové vazby
OC HN
Měření: - výměnné experimenty s D2O- teplotní závislost výměnitelných
protonů (NH, OH…)
NMR experimenty:- malé molekuly -COSY- velké molekuly - 1H-15N HSQC
Z NMR experimentů je možné získat pouze informaci o donoru!! Akceptory jsou většinou určeny až z molekulárního modelování a výpočtů!!

Vodíkové vazby v pravidelných strukturáchα-helix
β-sheet
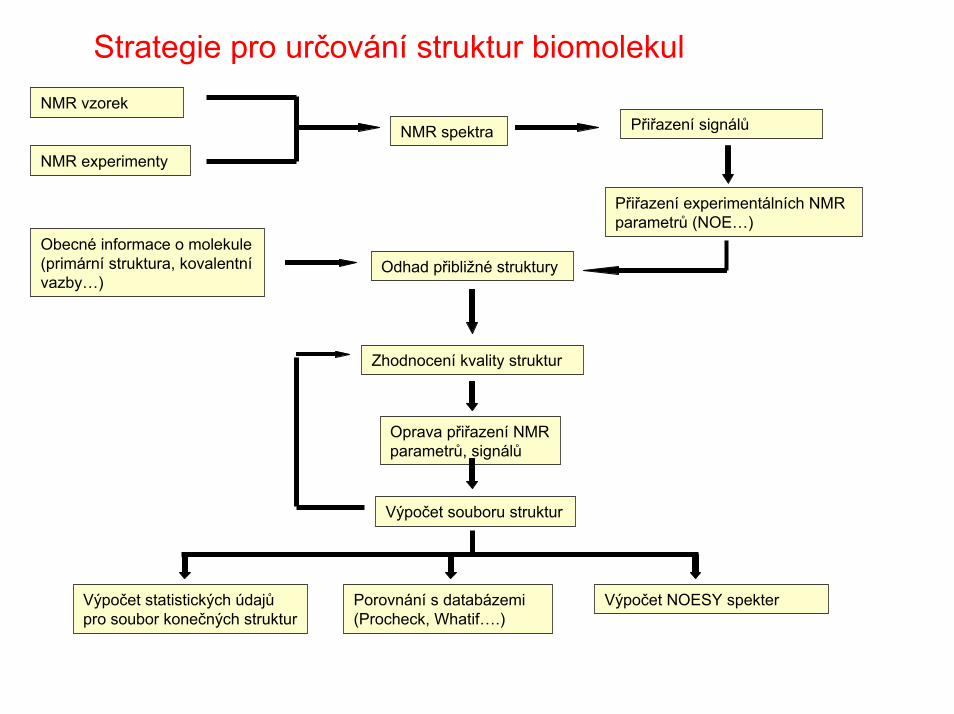
Strategie pro určování struktur biomolekul
Obecné informace o molekule (primární struktura, kovalentnívazby…)
NMR vzorek
NMR experimenty
NMR spektra Přiřazení signálů
Přiřazení experimentálních NMR parametrů (NOE…)
Odhad přibližné struktury
Oprava přiřazení NMR parametrů, signálů
Zhodnocení kvality struktur
Výpočet souboru struktur
Výpočet statistických údajůpro soubor konečných struktur
Výpočet NOESY spekterPorovnání s databázemi(Procheck, Whatif….)

Jak vše poskládat dohromady ????
Cray T3E
Info o kovalentnístruktuře
Omezení vzdáleností(NOEs)
Omezení dihedrálních úhlů(interakční konst.)
potkintot EEE +=
Výpočetní algoritmus: Molekulární mechanika
simulované žíhání s experimentálními omezeními (vzdálenosti,
dihedrální úhly…)
- molekula se ohřeje na vysokou teplotu (2000 – 50 000 K)
- pomalu se ochladí na teplotu blízkou nule
simulované žíhání v Kartézském prostoru (Newtonovy pohybové
rovnice)
simulované žíhání v prostoru torsních úhlů (Lagrangeovy rovnice)

Růstový modulátor Granulin 1eCyprinus carpio
Růstový modulátor Granulin 1eCyprinus carpio

Studium dynamických jevů proteinů pomocí NMR. Proč?
Molekuly nejsou statické, vykonávají pohyby v různých časových škálách.
Vypočtená statická struktura je často průměrem skutečných stavůmolekuly.
Funkce mnoha biologicky aktivních molekul závisína jejich flexibilitě.
V roztoku (fyziologické prostředí) podléhají biologicky aktivní molekuly přirozeným pohybům, které nejsou v krystalové mřížce patrné.
Výhoda NMR spektroskopie nad rentgenovou krystalografií.


Studium dynamických jevů proteinů pomocí NMRVztah relaxační rychlostí k molekulárním pohybům v různé časové škále:NMR parametr časová škála
podélná relaxace R1 1012 – 108 s-1
podélná relaxace běhemspin-locku R1ρ 106 – 103 s -1
příčná relaxace R2 103 – 10-3 s -1
Měřená jádra:
1H – téměř se neměří (obtížně definovatelné)
15N – dynamika páteře proteinu (dobře měřitelné, dobře definovatelné)
13C – dynamika postranních řetězců i páteře (obtížněji měřitelné, dobře definovatelné)
2H - měří se ve speciálních případech (CH3, obtížněji připravitelný vzorek, není jednoduché měřit, dobře definovatelné

Lipari-Szabóův přístupZpracování výsledků
Pro analýzu je nutný model pohybu molekuly jako celku a jejích částí.
Jeden z nejúspěšnějších je Lipari-Szabóův „bezmodelový“ přístup (“model-free“ approach)
Předpoklady Lipari-Szabóova modelu:
• relaxace je modulována dvěma pohyby: globálním a lokálním
• oba pohyby jsou statisticky nezávislé
• globální reorientace je izotropní
• molekulární pohyb je charakterizován parametry:
tM korelační čas globálního pohybu
S2 parametr uspořádanosti (hodnota 0 - 1)
te korelační čas lokálního pohybu
Rex rychlost chemické (konformační) výměny
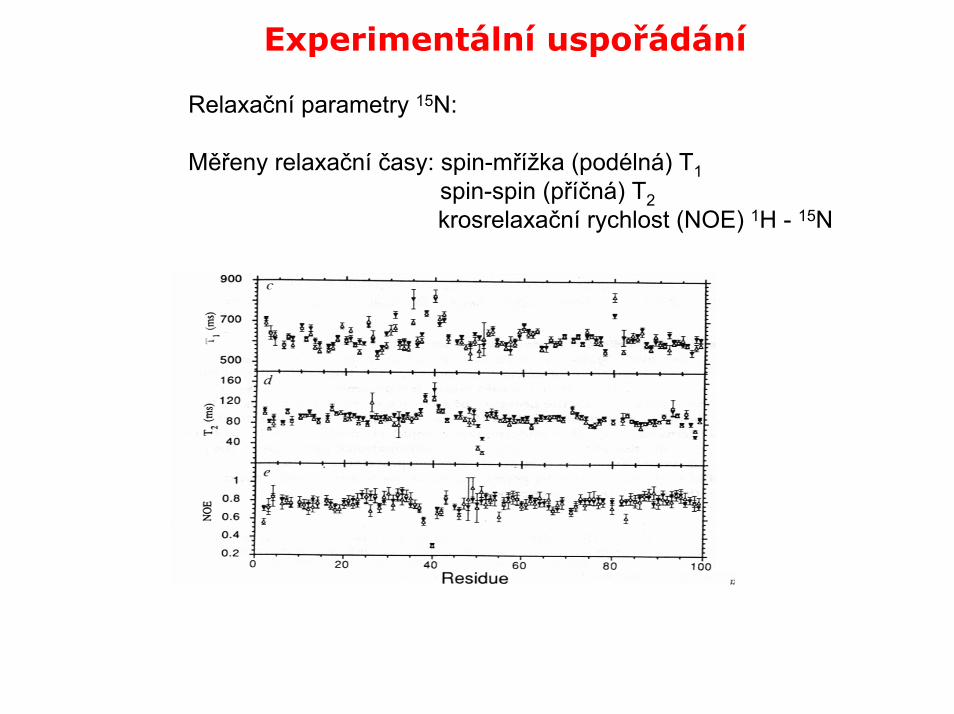
Experimentální uspořádání
Relaxační parametry 15N:
Měřeny relaxační časy: spin-mřížka (podélná) T1spin-spin (příčná) T2krosrelaxační rychlost (NOE) 1H - 15N
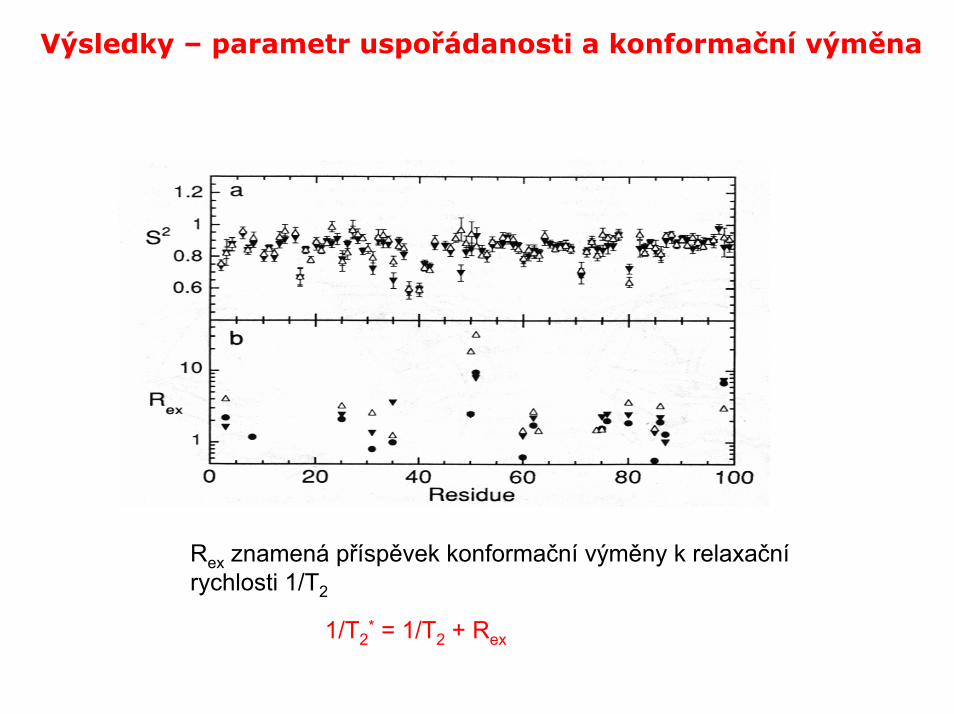
Výsledky – parametr uspořádanosti a konformační výměna
Rex znamená příspěvek konformační výměny k relaxačnírychlosti 1/T2
1/T2* = 1/T2 + Rex

Interpretace výsledků měření dynamiky páteře HIV-1 PR
Aminokyselinové zbytky podléhající rychlým pohybům v pikosekundové časovéškále (1-100 ps) o velké amplitudě
Aminokyselinové zbytky podléhající pomalým pohybům (konformační výměny v mikro- až milisekundové časové škále.

HIV-1 proteáza M-PMV proteáza (12 kDa)
Problém: Vyskytuje se M-PMV PR též jako homodimer nebojen v monomerní formě?
Metoda řešení pomocí NMR: Studium dynamiky proteinu.

Srovnání relaxačních vlastností 15N proteáz HIV-1 a M-PMV
M-PMV protease (C7/A, D26/N, C106/A) HIV-1 protease
NOE
-1.5
-1
-0.5
0
0.5
1
0 10 20 30 40 50 60 70 80 90 100 110
T2
00.050.1
0.150.2
0.250.3
0 10 20 30 40 50 60 70 80 90 100 110
T1
00.20.40.60.8
11.2
0 10 20 30 40 50 60 70 80 90 100 110
Závěr: U proteázy viru M-PMV chybí čtyřvláknový ß-sheet („ground floor“),který představuje hlavní stabilizační faktor homodimeru.M-PMV PR (12 kDa) se vyskytuje jako monomer.

HIV-1 protease with indicated “ground floor”
NN’C C’