nitric oxide promotes the wound-healing response of potato leaflets
TRANSCRIPT

Plant Physiol. Biochem. 45 (2007) 80e86www.elsevier.com/locate/plaphy
Research article
Nitric oxide promotes the wound-healingresponse of potato leaflets
Ramiro Parıs a, Lorenzo Lamattina b, Claudia A. Casalongue a,b,*
a Instituto de Investigaciones Biologicas, Universidad Nacional de Mar del Plata, Funes 3250, 7600 Mar del Plata, Argentinab Departamento de Biologıa, Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Mar del Plata,
Funes 3250, 7600 Mar del Plata, Argentina
Received 11 May 2006; accepted 5 December 2006
Available online 28 December 2006
Abstract
Nitric oxide (NO) is an essential regulatory molecule in several developmental and (patho) physiological processes. In this work, it isdemonstrated that NO participates in the wound-healing response of potato leaves. The experimental approaches showed that the depositionof the cell-wall glucan callose was induced by the application of the NO donor sodium nitroprusside (SNP), and such induction was additiveto the wound-induced callose production. Additionally, the expression of wound-related genes as phenylalanine ammonia-lyase (PAL) andextensin showed an accumulation of their transcript levels by SNP treatment. Moreover, the SNP-mediated increase of the PAL transcript levelwas additive to the induction mediated by wounding. These results indicate that increased levels of NO might potentiate the healing responses inplants leading to a rapid restoration of the damaged tissue.� 2006 Elsevier Masson SAS. All rights reserved.
Keywords: Callose; Healing; Nitric oxide; PAL; Solanum tuberosum; Wounding
1. Introduction
Wounding is a common damage that occurs to plants as aresult of biotic and abiotic stress factors. Generally, this dam-age not only physically destroys plant tissues, but also pro-vides a pathway for pathogen invasion. When plant tissue isdamaged, physiological, biochemical and anatomical changesthat take place are associated with defense and wound-healingmechanisms. Therefore, the wound-healing response is aimedat restoring the physiological status of the damaged tissue and
Abbreviations: cPTIO, Carboxy-2-phenyl-4,4,5,5-tetramethylimidazoli-
none-3-oxide-1-oxyl; DAF-FM DA, 4,5-diaminofluorescein-FM diacetate;
GSNO, nitrosoglutathione; JA, jasmonic acid; NO, nitric oxide; PAL, phenyl-
alanine ammonia-lyase; SA, salicylic acid; SNP, sodium nitroprusside.
* Corresponding author. Instituto de Investigaciones Biologicas, Universidad
Nacional de Mar del Plata, Funes 3250, 7600 Mar del Plata, Argentina. Tel.:
þ54 223 475 30 30; fax: þ54 223 475 31 50.
E-mail address: [email protected] (C.A. Casalongue).
0981-9428/$ - see front matter � 2006 Elsevier Masson SAS. All rights reserve
doi:10.1016/j.plaphy.2006.12.001
it is critical to prevent further lesions. Most of the inducedresponses, that include the induction of wound-healing relateddefense genes, appear between minutes to hours after wound-ing [25]. In addition, downstream events occurring in a timewindow of days after wounding are characterized by the depo-sition of different substances as suberin, tannins, and phenols.These compounds could act as precursors of lignin-like mate-rial deposited at the wounded site making tissues relativelyimpermeable and preventing the loss of water and microbialinvasion [3,27]. Consequently, some of the events underlyingthe plant wound-healing response are as follows: (i) repairof the damaged plant tissue, (ii) production of substancesthat could inhibit the action of possible predators and patho-gens, (iii) activation of wound-healing defense signaling path-ways; (iv) regulation of plant metabolism to the imposednutritional demands. However, these responses differ betweenspecies and organs [25].
Several studies have shown that different plant hormonesand signaling molecules are involved in wounding responses
d.

81R. Parıs et al. / Plant Physiol. Biochem. 45 (2007) 80e86
and its interacting networks [6]. Among them, jasmonic acid(JA) [26], salicylic acid (SA) [22], abscisic acid [5], andethylene [30] are important for the action of the woundingresponse [6]. In both, animal and plant systems, nitric oxide(NO) is an essential regulatory molecule involved in severaldevelopmental processes and stress responses [24]. In plants,NO action is mediated by signaling cascades that share com-mon elements with plant hormones, cell response mediatorsand second messengers, denoting the versatility an ubiquityof this molecule in diverse (patho)physiological processes[29]. Recent findings indicate that wounding causes a localand fast accumulation of NO in Cucumis sativus hypocotyls[33], and leaves [14] and epidermal cell layers [17] of Arabi-dopsis plants. Therefore, the possible role of NO as a modula-tor of plant wounding responses has been addressed throughits cross-talk with JA and SA signaling pathways [10,17,21].
Based on those evidences, it was hypothesized that NOprobably participates in the wound-healing response of leaftissues. In order to get a clue on this point, the effect ofSNP treatments on some parameters related to the wound-healing response was tested. Thereafter, results are discussedin terms of understanding the participation of NO in thewound-healing responses of leaf tissues.
2. Results and discussion
The experimental approach consisted in a mechanical stressapplied on potato leaflets at several sites (Fig. 1, lower panel).NO production was visualized in real-time at tissue level usingthe NO-selective fluorescent probe 4,5-diaminofluorescein-FM
diacetate (DAF-FM DA) (Fig. 1, upper panel). An initial fluo-rescence microscopy examination showed that autofluores-cence was undetectable in wounded leaflets without theapplication of the probe (Fig. 1A). Induction of NO (greenfluorescence) was visualized and restricted at wounding siteswhen potato leaflets were loaded with DAF-FM DA(Fig. 1B). Notice that non-wounded areas do not show in-crease on the fluorescence, meaning that NO productionoccurs specifically in response to wounding. Wounding plus100 mM SNP, used as NO donor, resulted in a significant in-crease of NO-detected fluorescence (Fig. 1C). Carboxy-2-phe-nyl-4,4,5,5-tetramethylimidazolinone-3-oxide-1-oxyl (cPTIO)is highly specific membrane-permeable NO scavenger. Itdoes not react with ROS and has been used to scavenge NOproduction as well as blocker of NO-dependent events[10,9]. Fig. 1D shows that cPTIO almost completely sup-pressed the elicited burst of fluorescence after wounding.Since cPTIO scavenges NO in an equimolar ratio [1], it isspeculated that under the experimental conditions used inthis assay and due to the strength of the NO burst in responseto wounding, the NO concentration could overstep the scav-enging capacity of cPTIO. On a previous report, Orozco-Car-denas and Ryan [31] showed that wounding did not result inenhanced NO production, probably due to the very differentdetection limit of the applied NO assay. In this work, NO pro-duction was visualized as the total NO accumulated during90 min after wounding and it was consistent with previous re-ports which indicate that the NO production was registered asearly as 10 min up to 3 h after wounding in epidermal cells[17] and entire leaves [14] of Arabidopsis plants.
Fig. 1. Nitric oxide (NO) detection in wounded potato leaflets. Potato leaflets were wounded with a tip and incubated with the specific NO fluorescent probe, DAF-
FM DA (DAF) (5 mM). (AeD) Fluorescence microscopy images were taken at 90 min after wounding. (A) Autofluorescence of wounded leaflets without DAF. (B)
Fluorescence indicative of NO production is localized as bright green spots at the surrounding of wounding sites. (C) Wounded leaflets and then treated with
100 mM SNP, a strong green fluorescence which spreads over the wounding sites is visualized. (D) In the presence of the NO scavenger, 1 mM cPTIO, reduced
fluorescence is observed. (EeH) Bright-field images corresponding to panels (A)e(D). Bar¼ 0.2 mm.

82 R. Parıs et al. / Plant Physiol. Biochem. 45 (2007) 80e86
In order to evaluate the action of NO in the wound-healingresponse, potato leaflets were pretreated with two different NOdonors, SNP and GSNO, and wounded after 24 h. In this case,wounding treatments were performed with a forceps to in-crease the damaged area. Fig. 2 shows better tissue integrity(Fig. 2A) and sustained chlorophyll levels (Fig. 2B) at 4days after wounding on GSNO-pretreated leaflets. Similar re-sults were obtained using SNP as NO donor. Furthermore, thespectrophotometric quantification of methanol extracts fromwounded leaflets indicated higher chlorophyll levels in SNP-pretreated leaflets compared with water-pretreated ones,22.7 mg ml�1� 3.05 and 13.5 mg ml�1� 3.60, respectively.
On the other hand, callose is a b-1,3 glucan deposited be-tween the plasma membrane and the cell wall after exposureof plants to a range of abiotic and biotic stresses. It is believedthat callose could partially obliterate the vascular system toavoid loss of turgor and disappear if not lasting damage oc-curs. Fig. 3A shows that wounding induces the accumulationof callose in potato leaflets, as it was previously reported forother plant species [19]. Fig. 3A also shows that, the SNP-pre-treatment was sufficient to increase approximately 50% thecallose content compared with water-pretreated leaflets. How-ever, higher amounts of callose were measured upon woundingin SNP-pretreated leaves, denoting a potentiated wound-heal-ing response when exogenous SNP was applied. Duringnormal plant growth and development, callose is found asa transitory component of the cell plate in dividing cells[34]. Fabbri et al. [11] described that in potato tuber tissuewounding enhances mitotic activity in specific meristematiccell populations. Consequently, augmented callose content inpotato leaflets tissue stimulated by SNP-pretreatment could
be due to an NO-stimulation mediated stimulation of eitherthe callose synthesis at the preexisting cell wall matrix and/or a stimulation of the cell division rate with the concomitantincrease on the number of cell plates where callose is also de-posited [8,32]. Hence, in order to discern between these twohypotheses, the specific cell-localization of callose depositionafter SNP and wounding treatments should be investigatedusing callose-specific dyes.
Previous studies have also provided a strong support in theknowledge of lignifying cells as part of the plant defense re-sponse. Lignins are complex cell wall phenolic heteropoly-mers, which result from the oxidative polymerization of theseveral monolignols. Gabaldon et al. [13] described that theeffect of NO on cell wall lignification is rather complex. Inour system, lignin was measured and neither quantitative norqualitative analyses by derivatization with thioglycolic acidor bright-field microscopy using phloroglucinol method, re-spectively, showed reproducible differences after woundingand/or SNP treatments (not shown). However, the depositionof a brown material on the SNP-pretreated leaflets at thewound boundary layer in comparison to water-pretreated leaf-lets was evident (Fig. 3B). Moreover, the measurement of totalsoluble phenols by Folin-Ciocalteu’s method indicated nodifference or a slight decay of total phenols after woundingand SNP treatment, compared with control (not shown).Thereafter, it is speculated that a proportion of the solublephenols turns oxidized into quinones and then, it becomes neg-ligible to the soluble-phenols detection method after wounding[12]. The polyphenol oxidase enzyme could catalyze thatoxidation and the determination of its activity could reinforcethis hypothesis [35].
Fig. 2. Effect of wounding and NO in the wound-healing response. Potato leaflets were pretreated for 24 h with 50 mM GSNO (GSNO) or 50 mM GSH (control)
before wounding. (A) Images were taken 4 days post-wounding before (upper panel) or upon chlorophyll removal by ethanol (lower panel). Bar¼ 1 mm. (B)
Chlorophyll was extracted with methanol and quantified from potato leaflets 4 days after wounding. Values are expressed as a percentage of the water-pretreated
sample.

83R. Parıs et al. / Plant Physiol. Biochem. 45 (2007) 80e86
In addition to polysaccharides, plant cell walls containmany proteins, as extensins, a family of hydroxyproline-richcell wall glycoproteins. These proteins become insolubilizedat the cell wall, probably by the formation of intermolecularcross-linkages. The deposition and cross-linking of these pro-teins have been proposed to increase the mechanical strengthof the cell wall [20]. In addition to its physiological action,extensin proteins and transcripts accumulate in response towounding [36]. In this work, potato leaflets were pretreatedwith 100 mM SNP or water for 24 h and then submitted towounding. To perform Northern blot assays, tissue from thearea surrounding the wounded sites was collected at 6 and24 h post-wounding (hpw). Fig. 4 shows that the extensin tran-script level increased 1.5-fold at 6 h post-wounding. This in-crease reached almost 2-fold after 24 h post-wounding, as itwas previously reported for potato tubers [4] and Phaseolusvulgaris [36]. Furthermore, SNP-pretreatment increased theextensin transcript level 3-fold. In SNP-pretreated and thenwounded leaflets, the extensin transcript level reached anincrease of 2-fold as it was shown for non-pretreated leafletsat 24 h post-wounding.
Other accepted marker of wounding is the expression ofphenylalanine ammonia-lyase (PAL) gene. In higher plants,PAL activity varies greatly with the stage of development,with cell and tissue differentiation and upon the exposure todifferent stresses. Their products operate in the very earlysteps in the phenylpropanoid pathway and lead to a varietyof defense and healing compounds such as SA, phytoalexinsand lignin-like polymers as the polyaromatic domains of su-berin, among others [15]. Different reports have demonstratedthat NO modulates the expression of PAL in Arabidopsisplants [16], soybean cell cultures [9], and tobacco leaves[10]. Consequently, the PAL transcript level was monitored
Fig. 3. Influence of NO and wounding treatments on callose contents. (A)
Callose was extracted and quantified from potato leaflets. Callose content is
expressed as the percentage of the total content of non-wounded and water-
treated leaflets (control 100%). The mean average value of three independent
experiments is indicated with standard deviation. (B) Potato leaflets were pre-
treated for 24 h with water or 100 mM SNP (SNP) before wounding. Images
were taken 4 days post-wounding upon chlorophyll removal by ethanol.
Bar¼ 1 mm.
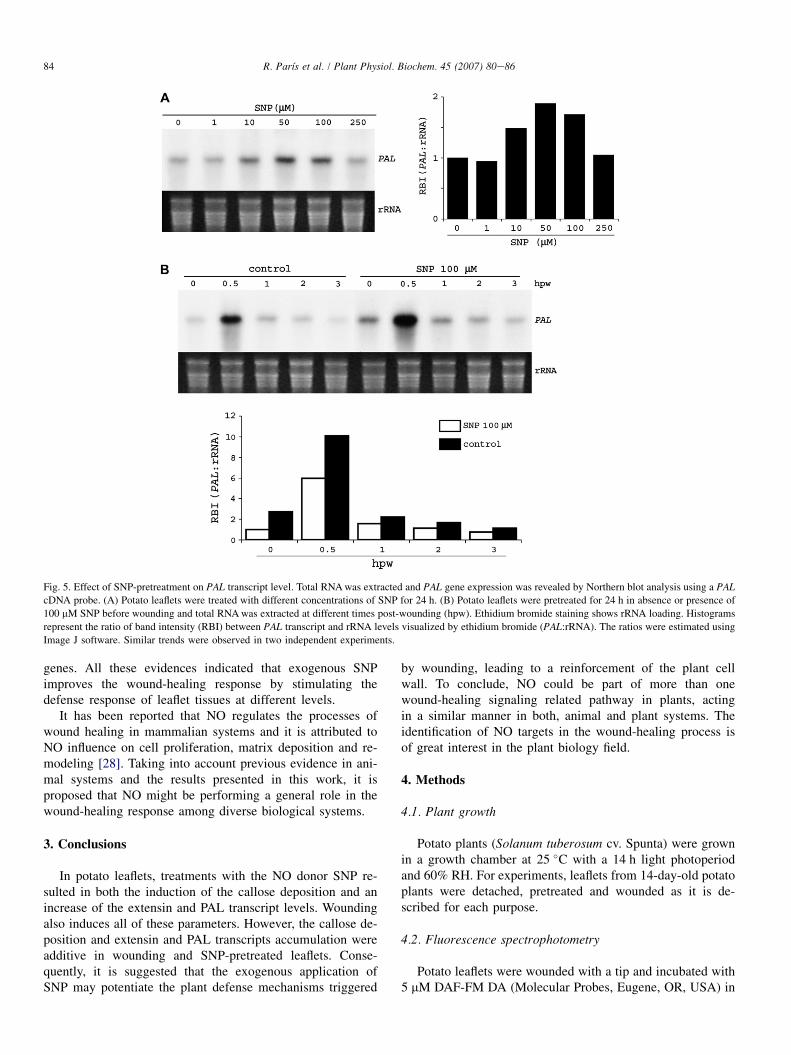
in wounded potato leaflets. First, a dose response was esti-mated using different SNP concentrations and the maximumlevel of PAL transcript was detected at 50 and 100 mM SNPafter 24 h of treatment (Fig. 5A). Fig. 5B shows the typicalPAL response to wounding, with a sharp and strong transcriptaccumulation at 0.5 h, returning to the basal level 1 h later(Fig. 5B), as was previously described by Joos and Hahlbrock[18]. However, the PAL transcript level was significantlyhigher in 24 h SNP-pretreated leaflets than in water-pretreatedones at 0.5 h post-wounding. These results suggest an additiveeffect on the increase of PAL transcript levels by both stimuliSNP and wounding.
Ours results suggested a positive correlation betweenwound-healing response and NO signaling. Others studies[16,17] demonstrated a fast accumulation of NO after wound-ing and the involvement of NO in the defense responses. In-terestingly, a previous work [31] showed that NO acts asa negative regulator of some others wound-inducible defensegene expression as the proteinase inhibitor I. However, in thiswork NO does not down regulate the early wound-inducible
Fig. 4. Effect of SNP-pretreatment on extensin transcript level. Potato leaflets
were pretreated for 24 h in absence (control) or presence of 100 mM SNP
before wounding. Total RNA was extracted at different times post-wounding
(hpw). (A) Northern blot assay was performed using a potato cDNA probe
encoding extensin. Ethidium bromide staining shows rRNA loading. (B) The
histogram represents the ratio between the extensin transcript level and
rRNA visualized by ethidium bromide (EXT:rRNA). The ratios were estimated
using Image J software. Similar trends were observed in two independent
experiments.
84 R. Parıs et al. / Plant Physiol. Biochem. 45 (2007) 80e86
Fig. 5. Effect of SNP-pretreatment on PAL transcript level. Total RNA was extracted and PAL gene expression was revealed by Northern blot analysis using a PALcDNA probe. (A) Potato leaflets were treated with different concentrations of SNP for 24 h. (B) Potato leaflets were pretreated for 24 h in absence or presence of
100 mM SNP before wounding and total RNA was extracted at different times post-wounding (hpw). Ethidium bromide staining shows rRNA loading. Histograms
represent the ratio of band intensity (RBI) between PAL transcript and rRNA levels visualized by ethidium bromide (PAL:rRNA). The ratios were estimated using
Image J software. Similar trends were observed in two independent experiments.
genes. All these evidences indicated that exogenous SNPimproves the wound-healing response by stimulating thedefense response of leaflet tissues at different levels.
It has been reported that NO regulates the processes ofwound healing in mammalian systems and it is attributed toNO influence on cell proliferation, matrix deposition and re-modeling [28]. Taking into account previous evidence in ani-mal systems and the results presented in this work, it isproposed that NO might be performing a general role in thewound-healing response among diverse biological systems.
3. Conclusions
In potato leaflets, treatments with the NO donor SNP re-sulted in both the induction of the callose deposition and anincrease of the extensin and PAL transcript levels. Woundingalso induces all of these parameters. However, the callose de-position and extensin and PAL transcripts accumulation wereadditive in wounding and SNP-pretreated leaflets. Conse-quently, it is suggested that the exogenous application ofSNP may potentiate the plant defense mechanisms triggered
by wounding, leading to a reinforcement of the plant cellwall. To conclude, NO could be part of more than onewound-healing signaling related pathway in plants, actingin a similar manner in both, animal and plant systems. Theidentification of NO targets in the wound-healing process isof great interest in the plant biology field.
4. Methods
4.1. Plant growth
Potato plants (Solanum tuberosum cv. Spunta) were grownin a growth chamber at 25 �C with a 14 h light photoperiodand 60% RH. For experiments, leaflets from 14-day-old potatoplants were detached, pretreated and wounded as it is de-scribed for each purpose.
4.2. Fluorescence spectrophotometry
Potato leaflets were wounded with a tip and incubated with5 mM DAF-FM DA (Molecular Probes, Eugene, OR, USA) in

85R. Parıs et al. / Plant Physiol. Biochem. 45 (2007) 80e86
300 ml buffer 20 mM HEPES pH 7.7 for 90 min at roomtemperature. Wounded leaflets were also pre-incubated with1 mM cPTIO (Molecular Probes, Eugene, OR, USA) or100 mM SNP (Merck & Co., Whitehouse Station, NJ, USA) dur-ing 30 min before DAF-FM DA incubation. Preparations wereobserved with an epi-fluorescence microscopy Nikon ECLIPSEE200 (excitation at 450e190 nm; emission at 535e540 nm) andphotographs were taken using a Nikon Coolpix 990 digitalcamera.
4.3. Extraction and quantitative analysisof chlorophyll and callose
To measure the callose content, potato leaflets were pre-treated with water or 100 mM SNP for 24 h, wounded witha forceps and floated in sterile petri dishes containing the cor-responding pretreatment solutions for 4 days.
The chlorophyll content was measured in leaflets pretreatedwith 50 mM GSNO (GSNO) or 50 mM GSH (control) for 24 hbefore wounding. Extraction and measurements of chlorophylland callose were carried out as described by Beligni andLamattina [2] and Kohler et al. [23], respectively.
4.4. RNA gel blot analysis
Potato leaflets were pretreated as described above andwounded with a scissor blade. Total RNA from the adjacentarea to the wounding site (2 mm) was isolated using Trizol Re-agent according to the manufacturer instructions (Invitrogen,Carlsbad, CA, USA). Total RNA (15 mg) was denaturizedwith glyoxal and subsequently separated on 1.5% agarosegel electrophoresis. Then, the gel was stained with ethidiumbromide and transferred onto Hybond-XL (Amersham Biosci-ences, Piscataway, NJ, USA) membranes. Membranes wereprehybridized and hybridized in Church’s solution [7] (0.5 MNaPOH4 buffer pH 7.2, 7% SDS and 10 mM EDTA) contain-ing 32P-labelled PAL [18], or extensin [4] nucleotide sequencesat 65 �C overnight. These probes were generated using Mega-prime DNA Labeling System as described by the manufacturer(Amersham Biosciences, Piscataway, NJ, USA) using [a-32P]dCTP. Autoradiograms were exposed at�80 �C (Agfa, Mortsel,Belgium) and then subjected to scanning and densitometricanalysis.
Acknowledgements
We thank Dr Hahlbrock and Dr Bown for providing PALand extensin cDNA clones, respectively, and Lic. Marıa Ceci-lia Terrile for her excellent technical assistance. This work wassupported by the Agencia Nacional de Promocion CientıficaTecnologica (ANPCyT), Consejo Nacional de InvestigacionesCientıficas y Tecnicas (CONICET) and UNMdP, Argentina.RP was recipient of a fellowship from the CONICET.CC and LL are researchers from CONICET. LL is a fellowfrom J.S. Guggenheim Foundation.
References
[1] T. Akaike, M. Yoshida, Y. Miyamoto, K. Sato, M. Kohno, K. Sasamoto,
K. Miyazaki, S. Ueda, H. Maeda, Antagonistic action of imidazolineoxyl
N-oxides against endothelium-derived relaxing factor/�NO through a rad-
ical reaction, Biochemistry 32 (1993) 827e832.
[2] M.V. Beligni, L. Lamattina, Nitric oxide protects against cellular damage
produced by methylviologen herbicides in potato plants, Nitric Oxide 3
(1999) 199e208.
[3] A.M. Boudet, D.P. Goffner, J. Grima-Pettenati, Lignins and lignification:
recent biochemical and biotechnological developments, Plant Biol.
Pathol. C.R. Acad. Sci. Paris (1996) 317e331.
[4] D.P. Bown, G.P. Bolwell, J.A. Gatehouse, Characterisation of potato
(Solanum tuberosum L.) extensins: a novel extensin-like cDNA from
dormant tubers, Gene 134 (1993) 229e233.
[5] E. Carrera, S. Prat, Expression of the Arabidopsis abi1-1 mutant allele
inhibits proteinase inhibitor wound-induction in tomato, Plant J. 15
(1998) 765e771.
[6] Y.H. Cheong, H.S. Chang, R. Gupta, X. Wang, T. Zhu, S. Luan,
Transcriptional profiling reveals novel interactions between wounding,
pathogen, abiotic stress, and hormonal responses in Arabidopsis, Plant
Physiol. 129 (2002) 661e677.
[7] G.M. Church, W. Gilbert, Genomic sequencing, Proc. Natl. Acad. Sci.
U.S.A. 81 (1984) 1991e1995.
[8] N. Correa-Aragunde, M. Graziano, C. Chevalier, L. Lamattina, Nitric
oxide modulates the expression of cell cycle regulatory genes during
lateral root formation in tomato, J. Exp. Bot. 57 (2006) 581e588.
[9] M. Delledonne, Y. Xia, R.A. Dixon, C. Lamb, Nitric oxide functions as
a signal in plant disease resistance, Nature 394 (1998) 585e588.
[10] J. Durner, D. Wendehenne, D.F. Klessig, Defense gene induction in
tobacco by nitric oxide, cyclic GMP, and cyclic ADP-ribose, Proc.
Natl. Acad. Sci. U.S.A. 95 (1998) 10328e10333.
[11] A.A. Fabbri, C. Fanelli, M. Reverberi, A. Ricelli, E. Camera,
S. Urbanelli, A. Rossini, M. Picardo, M.M. Altamura, Early physiologi-
cal and cytological events induced by wounding in potato tuber, J. Exp.
Bot. 51 (2000) 1267e1275.
[12] M. Friedman, Chemistry, biochemistry and dietary role of potato poly-
phenols, J. Agric. Food Chem. 45 (1997) 1523e1540.
[13] C. Gabaldon, L.V. Gomez Ros, M.A. Pedreno, A. Ros Barcelo, Nitric
oxide production by the differentiating xylem of Zinnia elegans, New
Phytol. 165 (2005) 5e7.
[14] H. Garces, D. Durzan, M.C. Pedroso, Mechanical stress elicits nitric
oxide formation and DNA fragmentation in Arabidopsis thaliana, Ann.
Bot. 87 (2001) 553e707.
[15] K. Hahlbrock, D. Scheel, Physiology and molecular biology of phenyl-
propanoid metabolism, Annu. Rev. Plant Physiol. Plant Mol. Biol. 40
(1989) 347e369.
[16] X. Huang, E. Kiefer, U. von Rad, D. Ernst, I. Foissner, J. Durner, Nitric
oxide burst and nitric oxide-dependent gene induction in plants, Plant
Physiol. Biochem. 40 (2002) 625e631.
[17] X. Huang, K. Stettmaier, C. Michel, P. Hutzler, M.J. Mueller, J. Durner,
Nitric oxide is induced by wounding and influences jasmonic acid
signaling in Arabidopsis thaliana, Planta 218 (2004) 938e946.
[18] H.J. Joos, K. Hahlbrock, Phenylalanine ammonia-lyase in potato (Sola-
num tuberosum L.). Genomic complexity, structural comparison of two
selected genes and modes of expression, Eur. J. Biochem. 204 (1992)
621e629.
[19] H. Kauss, Callose and callose synthase, in: D.J. Bowles, S. Gurr,
M. McPherson (Eds.), Practical Approaches to Molecular Plant Pathol-
ogy, Oxford University Press, Oxford, 1992, pp. 1e8.
[20] M.J. Kieliszewski, D.T. Lamport, Extensin: repetitive motifs, functional
sites, post-translational codes, and phylogeny, Plant J. 5 (1994)
157e172.
[21] D.F. Klessig, J. Durner, R. Noad, D.A. Navarre, D. Wendehenne, D. Kumar,
J.M. Zhou, J. Shah, S. Zhang, P. Kachroo, Y. Trifa, D. Pontier, E. Lam,
H. Silva, Nitric oxide and salicylic acid signaling in plant defense, Proc.
Natl. Acad. Sci. U.S.A. 97 (2000) 8849e8855.

86 R. Parıs et al. / Plant Physiol. Biochem. 45 (2007) 80e86
[22] D.F. Klessig, J. Malamy, The salicylic acid signal in plants, Plant Mol.
Biol. 26 (1994) 1439e1458.
[23] A. Kohler, S. Schwindling, U. Conrath, Extraction and quantitative deter-
mination of callose from Arabidopsis leaves, Biotechniques 28 (2000)
1084e1086.
[24] L. Lamattina, C. Garcia-Mata, M. Graziano, G.C. Pagnussat, Nitric
oxide: the versatility of an extensive signal molecule, Annu. Rev. Plant
Biol. 54 (2003) 109e136.
[25] J. Leon, E. Rojo, J.J. Sanchez-Serrano, Wound signaling in plants, J. Exp.
Bot. 52 (2001) 1e9.
[26] J. Leon, E. Rojo, E. Titarenko, J.J. Sanchez-Serrano, Jasmonic acid-
dependent and -independent wound signal transduction pathways are
differentially regulated by Ca2þ/calmodulin in Arabidopsis thaliana,
Mol. Gen. Genet. 258 (1998) 412e419.
[27] E. Lulai, D. Corsini, Differential deposition of suberin phenolic and
aliphatic domains and their roles in resistance to infection during potato
tuber (Solanum tuberosum L.) wound-healing, Physiol. Mol. Plant
Pathol. 53 (1998) 209e222.
[28] J.D. Luo, A.F. Chen, Nitric oxide: a newly discovered function on wound
healing, Acta Pharmacol. Sin. 26 (2005) 259e264.
[29] S. Neill, R. Desikan, J. Hancock, Nitric oxide signals in plants, New
Phytol. 159 (2003) 11e35.
[30] P.J. O’Donnell, C. Calvert, R. Atzorn, C. Wasternack, H.M.O. Leyser,
D.J. Bowles, Ethylene as a signal mediating the wound response of
tomato plants, Science 274 (1996) 1914e1917.
[31] M.L. Orozco-Cardenas, C.A. Ryan, Nitric oxide negatively modulates
wound signaling in tomato plants, Plant Physiol. 130 (2002) 487e493.
[32] K. Otvos, T.P. Pasternak, P. Miskolczi, M. Domoki, D. Dorjgotov,
A. Szucs, S. Bottka, D. Dudits, A. Feher, Nitric oxide is required for,
and promotes auxin-mediated activation of, cell division and embryo-
genic cell formation but does not influence cell cycle progression in
alfalfa cell cultures, Plant J. 43 (2005) 849e860.
[33] G.C. Pagnussat, M. Simontacchi, S. Puntarulo, L. Lamattina, Nitric oxide
is required for root organogenesis, Plant Physiol. 129 (2002) 954e956.
[34] A.L. Samuels, T.H. Giddings Jr., L.A. Staehelin, Cytokinesis in tobacco
BY-2 and root tip cells: a new model of cell plate formation in higher
plants, J. Cell Biol. 130 (1995) 1345e1357.
[35] P. Thipyapong, M.D. Hunt, J.C. Steffens, Antisense downregulation of
polyphenol oxidase results in enhanced disease susceptibility, Planta
220 (2004) 105e117.
[36] K.L. Wycoff, P.A. Powell, R.A. Gonzales, D.R. Corbin, C. Lamb,
R.A. Dixon, Stress activation of a bean hydroxyproline-rich glycoprotein
promoter is superimposed on a pattern of tissue-specific developmental
expression, Plant Physiol. 109 (1995) 41e52.