nghiÊn cỨu Ảnh hƯỞng cỦa chẤt kÍch khÁng lÊn sỰ...
TRANSCRIPT

ĐẠI HỌC HUẾ
TRƯỜNG ĐẠI HỌC KHOA HỌC
TRƯƠNG THỊ PHƯƠNG LAN
NGHIÊN CỨU ẢNH HƯỞNG CỦA CHẤT KÍCH KHÁNG LÊN SỰ BIỂU HIỆN CỦA MỘT SỐ GEN
THAM GIA QUÁ TRÌNH SINH TỔNG HỢP CURCUMINOID
Ở TẾ BÀO NGHỆ ĐEN (Curcuma zedoaria Roscoe)
LUẬN ÁN TIẾN SĨ SINH HỌC
Chuyên nganh: SINH LÝ HỌC THỰC VẬT
Ma sô: 9420112
Người hướng dẫn khoa học:
GS.TS. NGUYỄN HOÀNG LỘC

HUẾ - NĂM 2019
LỜI CÁM ƠN
Hoàn thành luận án này, trước hết chúng tôi xin bày tỏ lòng biết ơn sâu
sắc đến GS.TS Nguyễn Hoàng Lộc đã quan tâm giúp đỡ và hướng dẫn tận tình.
Xin được bày tỏ lòng biết ơn tới cán bộ giảng viên của Bộ môn Sinh học
Ứng dụng, Bộ môn Công nghệ Sinh học, Khoa Sinh học, Trường Đại học Khoa
học, Đại học Huế; Phòng thí nghiệm Các hợp chất thứ cấp, Viện tài nguyên,
Môi trường và Công nghệ sinh học, Đại học Huế (giai đoạn 2013-2014); Viện
Công nghệ Sinh học, Đại học Huế đã giúp đỡ chúng tôi trong suốt thời gian
thực hiện đề tài.
Xin cám ơn Ban Giám đốc, Ban đào tạo Sau đại học của Đại học Huế; Ban
Giám hiệu, Phòng Nghiên cứu Khoa học và hợp tác Quốc tế; Phòng Đào tạo
Sau đại học -Trường đại học Khoa học; Ban chủ nhiệm Khoa sinh học-Trường
Đại học Khoa học- Đại học Huế ; Ban Giám hiệu, Khoa Dược, Khoa cơ bản,
Bộ môn Sinh học-Trường Đại học Y Dược Huế đã có nhiều giúp đỡ quý báu,
tạo mọi điều kiện tốt nhất để chúng tôi hoàn thành luận án.
Xin cám ơn các đồng nghiệp, bạn bè đã nhiệt tình động viên, hỗ trợ chúng
tôi hoàn thành luận án.
Cuối cùng, xin được bày tỏ lòng biết ơn đến những người thân trong gia
đình đã luôn giúp đỡ, động viên và khích lệ cả vật chất lẫn tinh thần.
Xin trân trọng cảm ơn!
Huế, ngày tháng năm 2019
Tác giả
Trương Thị Phương Lan

i
LỜI CAM ĐOAN
Tôi xin cam đoan đây là công trình nghiên cứu của riêng tôi. Các số liệu,
kết quả nêu trong luận án là trung thực, khách quan, nghiêm túc và chưa từng
được ai công bố trong bất kỳ công trình nào khác. Nếu có gì sai sót tôi xin chịu
hoàn toàn trách nhiệm.
Tác giả luận án
Trương Thị Phương Lan

ii
BẢNG CHÚ THÍCH CHỮ VIẾT TẮT
BAP 6-benzylaminopurine
CoA coezyme A
cs cộng sự
CTAB hexadecyltrimethylammonium bromide
CzDCS Curcuma zedoaria DCS
CzCURS1 Curcuma zedoaria CURS1
CzCURS2 Curcuma zedoaria CURS2
CzCURS3 Curcuma zedoaria CURS3
ĐC đối chứng
DPPH 1-diphenyl-2-picrylhydrazyl
dw dry weight (khối lượng khô)
FRAP ferric reducing antioxidant power
fw fresh weight (khối lượng tươi)
HPLC high-performance liquid chromatography
IBA 3-indolebutyric acid
KIN kinetin
MAPK mitogen-activated protein kinase
MeJA methyl jasmonate
MS Murashige and Skoog (1962)
NAA naphthaleneacetic acid
NO nitric oxide
PAA phenyl acetic acid
ROS reactive oxygen species
SA salicylic acid
SNP sodium nitroprusside
YE yeast extract (dịch chiết nấm men)

iii
MỤC LỤC
LỜI CAM ĐOAN .............................................................................................. i
BẢNG CHÚ THÍCH CHỮ VIẾT TẮT ............................................................ ii
MỤC LỤC ........................................................................................................ iii
DANH MỤC CÁC BẢNG.............................................................................. vii
DANH MỤC CÁC HÌNH ẢNH .................................................................... viii
MỞ ĐẦU ........................................................................................................... 1
1. TÍNH CẤP THIẾT CỦA ĐỀ TÀI ................................................................ 1
2. MỤC TIÊU NGHIÊN CỨU.......................................................................... 2
3. NỘI DUNG VÀ PHẠM VI NGHIÊN CỨU ................................................ 3
Chương 1. TỔNG QUAN TÀI LIỆU ............................................................... 5
1.1. CÂY NGHỆ ĐEN ...................................................................................... 5
1.1.1. Đặc điểm thực vật học......................................................................... 5
1.1.2. Phân bố ................................................................................................ 5
1.1.3. Thành phần hóa học chính của củ nghệ đen ....................................... 6
1.1.3.1. Tinh dầu ........................................................................................... 6
1.1.3.2. Curcuminoid ..................................................................................... 7
1.1.4. Công dụng của nghệ đen ..................................................................... 8
1.1.4.1. Hoạt tính giảm đau ........................................................................... 8
1.1.4.2. Hoạt tính kháng ung thư ................................................................... 9
1.1.4.3. Hoạt tính bảo vệ gan ........................................................................ 9
1.1.4.4. Hoạt tính kháng viêm và chống loét ................................................ 9
1.1.4.5. Hoạt tính chống oxy hóa ................................................................ 10
1.1.4.6. Hoạt tính kháng khuẩn và kháng nấm............................................ 10
1.1.4.7. Các hoạt tính khác .......................................................................... 11
1.3. ELICITOR VÀ CÁC ỨNG DỤNG ......................................................... 15

iv
1.3.1. Elicitor ............................................................................................... 15
1.3.1.1. Khái niệm ....................................................................................... 15
1.3.1.2. Phân loại ......................................................................................... 15
1.3.1.3. Cơ chế kích kháng .......................................................................... 16
1.3.1.4. Các yếu tố ảnh hương đến sự kích kháng ...................................... 18
1.3.2. Ứng dụng của elicitor ........................................................................ 20
1.3.2.1. Elicitor sinh học ............................................................................. 20
1.3.2.2. Elicitor phi sinh học ....................................................................... 26
1.4. CÁC GEN THAM GIA TỔNG HỢP CURCUMINOID ........................ 27
1.4.1. Các con đường sinh tổng hợp curcuminoid ...................................... 27
1.4.2. Vai trò của các gen tham gia chu trình tổng hợp curcuminoid ......... 31
1.4.2.1. Gen mã hóa enzyme diketide-CoA synthase (DCS) ...................... 31
1.4.2.2. Gen mã hóa enzyme curcumin synthase (CURS) .......................... 31
1.4.2.3. Gen mã hóa enzyme Curcuminoid synthase .................................. 32
1.4.2.4. Gen mã hóa enzyme Chalcone synthase ........................................ 33
1.4.2.5. Các gen khác .................................................................................. 34
1.4.3. Tổng hợp curcuminoid theo phương thức tái tổ hợp ........................ 35
1.4.4. Cải thiện sự biểu hiện gen bằng xử lý elicitor .................................. 37
Chương 2. ĐỐI TƯỢNG VÀ PHƯƠNG PHÁP NGHIÊN CỨU .................. 39
2.1. ĐỐI TƯỢNG NGHIÊN CỨU ................................................................. 39
2.2. PHƯƠNG PHÁP NGHIÊN CỨU ............................................................ 39
2.2.1. Nuôi cấy in vitro cây nghệ đen ......................................................... 40
2.2.1.1. Khử trùng mẫu vật ......................................................................... 40
2.2.1.2. Tái sinh chồi và tạo rễ in vitro ....................................................... 41
2.2.1.3. Nuôi cấy callus ............................................................................... 41
2.2.1.4. Nuôi cấy tế bào .............................................................................. 41
2.2.2. Phân lập các gen tổng hợp curcuminoid ........................................... 42
2.2.2.1. Tách chiết DNA tổng số................................................................. 42

v
2.2.2.2. Khuếch đại PCR ............................................................................. 43
2.2.2.3. Tạo dòng và chú giải gen ............................................................... 43
2.2.2.4. Xây dựng cây phả hệ ...................................................................... 45
2.2.3. Xác định sự biểu hiện của các gen tổng hợp curcuminoid ............... 46
2.2.3.1. Xử lý elicitor .................................................................................. 46
2.2.3.2. Phân tích RT-PCR .......................................................................... 46
2.2.3.3. Phân tích HPLC ............................................................................. 48
2.2.4. Xử lý thống kê ................................................................................... 49
Chương 3. KẾT QUẢ NGHIÊN CỨU ........................................................... 50
3.1. THIẾT LẬP NUÔI CẤY TẾ BÀO .......................................................... 50
3.1.1. Nhân giống cây nghệ đen in vitro ..................................................... 50
3.1.1.1. Tái sinh chồi ................................................................................... 50
3.1.1.2. Tạo rễ in vitro ................................................................................. 52
3.1.2. Nuôi cấy callus .................................................................................. 54
3.1.2.1. Ảnh hương của 2,4-D và KIN ........................................................ 54
3.1.2.2. Ảnh hương của 2,4-D và NAA ...................................................... 55
3.1.2.3. Ảnh hương của 2,4-D và AgNO3 ................................................... 55
3.1.3. Nuôi cấy tế bào nghệ đen .................................................................. 57
3.2. NHẬN DẠNG CÁC GEN TỔNG HỢP CURCUMINOID .................... 59
3.2.1. Phân lập gen ...................................................................................... 59
3.2.2. Phân tích biểu hiện của các gen tổng hợp curcuminoid.................... 68
3.3. ẢNH HƯỞNG CỦA ELICITOR LÊN QUÁ TRÌNH SINH TỔNG HỢP
CURCUMINOID ............................................................................................ 72
3.3.1. Thăm dò ảnh hương của các elicitor lên biểu hiện của các gen tổng
hợp curcuminoid ......................................................................................... 72
3.3.2. Biểu hiện của gen tổng hợp curcuminoid ......................................... 74
3.3.3. Tích lũy curcumin ............................................................................. 76
Chương 4. BÀN LUẬN .................................................................................. 79

vi
4.1. THIẾT LẬP NUÔI CẤY TẾ BÀO .......................................................... 79
4.1.1. Nhân giống cây nghệ đen in vitro ..................................................... 79
4.1.2. Nuôi cấy callus và tế bào cây nghệ đen ............................................ 81
4.2. PHÂN LẬP CÁC GEN TỔNG HỢP CURCUMINOID ......................... 83
4.2.1. Phân lập gen tổng hợp curcuminoid ................................................. 83
4.2.2. Sự biểu hiện của các gen tổng hợp curcuminoid .............................. 85
4.3. NGHIÊN CỨU ẢNH HƯỞNG CỦA ELICITOR LÊN SỰ BIỂU HIỆN
CỦA CÁC GEN TỔNG HỢP CURCUMINOID ........................................... 85
4.3.1. Đặc điểm của các elicitor sử dụng trong nghiên cứu ........................ 85
4.3.1.1. Dịch chiết nấm men ....................................................................... 85
4.3.1.2. Salicylic acid .................................................................................. 87
4.3.2. Ảnh hương của elicitor lên sự sinh trương của tế bào ...................... 89
4.3.3. Ảnh hương của elicitor lên mức độ biểu hiện gen ............................ 90
4.3.4. Ảnh hương của các elicitor lên khả năng sản xuất curcumin ........... 92
KẾT LUẬN VÀ KIẾN NGHỊ......................................................................... 95
KẾT LUẬN ..................................................................................................... 95
KIẾN NGHỊ .................................................................................................... 95
DANH MỤC CÁC CÔNG BỐ LIÊN QUAN ĐẾN LUẬN ÁN .................... 96
TÀI LIỆU THAM KHẢO ............................................................................... 97

vii
DANH MỤC CÁC BẢNG
Bảng 1.1. Phân loại các elicitor khác nhau ..................................................... 17
Bảng 1.2. Một số ứng dụng elicitor để cải thiện khả năng tích lũy các hợp chất
thứ cấp trong nuôi cấy in vitro. ....................................................................... 21
Bảng 2.1. Trình tự các primer được dùng để khuếch đại vùng CDS của các gen
tổng hợp curcuminoid ơ nghệ đen ................................................................... 45
Bảng 2.2. Trình tự các primer được dùng để khuếch đại vùng chỉ thị của các
gen tổng hợp curcuminoid ơ nghệ đen ............................................................ 47
Bảng 3.1. Ảnh hương của AgNO3 lên khả năng tái sinh chồi mới của chồi in
vitro nguyên vẹn. ............................................................................................. 51
Bảng 3.2. Ảnh hương của AgNO3 lên khả năng tái sinh chồi mới của chồi in
vitro đã được chẻ đôi ....................................................................................... 52
Bảng 3.3. Ảnh hương của AgNO3 lên khả năng tạo rễ của chồi in vitro ........ 53
Bảng 3.4. Ảnh hương của KIN từ 0,5-2 mg/L và 2,4-D 1 mg/L lên sinh trương
của callus nghệ đen ......................................................................................... 55
Bảng 3.5 Ảnh hương của NAA từ 0,5-2 mg/L và 2,4-D 1 mg/L lên sinh trương
của callus nghệ đen ......................................................................................... 56
Bảng 3.6. Ảnh hương của 2,4-D 1 mg/L và AgNO3 từ 0,5-2 mg/L lên sinh
trương của callus nghệ đen. ............................................................................. 56
Bảng 3.7. Mật độ băng DNA từ phân tích RT-PCR vùng đặc hiệu của các gen
CzDCS, CzCURS1, CzCURS2 và CzCURS3 trong 2 loại mô khác nhau của nghệ
đen ................................................................................................................... 70
Bảng 3.8. Mật độ các băng DNA của vùng đặc hiệu ơ các gen tổng hợp
curcuminoid ..................................................................................................... 76
Bảng 3.9. Ảnh hương của các elicitor lên sinh trương và tích lũy curcumin trong
tế bào nghệ đen ................................................................................................ 77

viii
DANH MỤC CÁC HÌNH ẢNH
Hình 1.1. Cấu trúc hóa học của các curcumin .................................................. 8
Hình 1.2. Con đường sinh tổng hợp các curcuminoid trong cây nghệ vàng .. 30
Hình 1.3. Vai trò của các gen DCS và CURS trong tổng hợp curcuminoid ơ
nghệ vàng. ....................................................................................................... 32
Hình 2.1. Cây nghệ đen. .................................................................................. 39
Hình 2.2. Sơ đồ thí nghiệm. ............................................................................ 40
Hình 2.3. Vector pGEM®-T Easy (Promega, Mỹ). ......................................... 44
Hình 2.4. Vị trí của primer xuôi và ngược trên gen CzCURS1 ...................... 48
Hình 3.1. Cây nghệ đen in vitro 2 tháng tuổi. ................................................. 53
Hình 3.2. Callus nghệ đen 2 tuần tuổi sinh trương trên môi trường có 2,4-D 1
mg/L kết hợp với KIN 1,5 mg/L (A) và đối chứng sinh trương trên môi trường
có 2,4-D 3 mg/L kết hợp với BAP 3 mg/L (B). .............................................. 57
Hình 3.3. Đường cong sinh trương của tế bào nghệ đen. ............................... 58
Hình 3.4. Tế bào nghệ đen nuôi trong bình tam giác chứa môi trường MS bổ
sung 2,4-D 3 mg/L và BAP 3 mg/L ................................................................ 59
Hình 3.5. Sinh khối tươi (A) và khô (B) của tế bào nghệ đen. ....................... 59
Hình 3.6. Sản phẩm PCR của các gen tổng hợp curcuminoid khuếch đại từ DNA
tổng số của nghệ đen ....................................................................................... 61
Hình 3.7. Sơ đồ sắp xếp của các intron/exon trên 4 gen sinh tổng hợp
curcuminoid ơ nghệ đen. ................................................................................. 61
Hình 3.8. So sánh trình tự nucleotide (nu) vùng CDS của gen CzDCS
(MF663785) ơ nghệ đen và DCS ơ nghệ vàng (AB495006.1). ...................... 62
Hình 3.9. So sánh trình tự nucleotide (nu) vùng CDS của gen CzCURS1
(MF402846) ơ nghệ đen và CURS1 ơ nghệ vàng (AB495007.1). ................. 63
Hình 3.10. So sánh trình tự nucleotide (nu) vùng CDS của gen CzCURS2
(MF402846) ơ nghệ đen và CURS2 ơ nghệ vàng (AB506762.1). .................. 64

ix
Hình 3.11. So sánh trình tự nucleotide (nu) vùng CDS của gen CzCURS3
(NCBI: MF987835) ơ nghệ đen và CURS3 ơ nghệ vàng (NCBI: AB506763.1).
......................................................................................................................... 65
Hình 3.12. So sánh trình tự amino acid suy diễn của 2 gen CzDCS và DCS
(C0SVZ5.1). .................................................................................................... 66
Hình 3.13. So sánh trình tự amino acid suy diễn của 2 gen CzCURS1 và CURS1
(AJF45913.1)................................................................................................... 66
Hình 3.14. So sánh trình tự amino acid suy diễn của 2 gen CzCURS2 và CURS2
(BAW81545.1). ............................................................................................... 67
Hình 3.15. So sánh trình tự amino acid suy diễn của 2 gen CzCURS3 và CURS3
(AJF45914.1)................................................................................................... 67
Hình 3.16. Cây phả hệ của các gen sinh tổng hợp curcuminoid. .................... 68
Hình 3.17. Phân tích RT-PCR vùng đặc hiệu của các gen CzDCS, CzCURS1,
CzCURS2 và CzCURS3 trong 2 loại mô khác nhau của nghệ đen. ................ 70
Hình 3.18. Phổ HPLC của curcumin chuẩn. ................................................... 71
Hình 3.19. Phổ HPLC của dịch chiết củ nghệ đen. ......................................... 71
Hình 3.20. Phổ HPLC của dịch chiết callus nghệ đen. ................................... 72
Hình 3.21. RNA tổng số của các mẫu tế bào sau khi được xử lý elicitor ....... 73
Hình 3.22. Phân tích RT-PCR vùng đặc hiệu của các gen CzCURS1 (A) và
CzCURS3 (B) sau khi được xử lý elicitor. ...................................................... 74
Hình 3.23. Sản phẩm RT-PCR vùng đặc hiệu của các gen sinh tổng hợp
curcuminoid ơ nghệ đen. ................................................................................. 75
Hình 3.24. Phổ HPLC của curcumin chuẩn. ................................................... 78
Hình 3.25. Phổ HPLC của dịch chiết tế bào được xử lý với YE 1 g/L sau 5 ngày
nuôi cấy. .......................................................................................................... 78
Hình 3.26. Phổ HPLC của dịch chiết tế bào được xử lý với SA 100 µM sau 5
ngày nuôi cấy. ................................................................................................. 79

1
MỞ ĐẦU
1. TÍNH CẤP THIẾT CỦA ĐỀ TÀI
Cây nghệ đen (Curcuma zedoaria Roscoe) thuộc họ Gừng (Zingiberaceae)
còn được gọi là nga truật, tam nại hay ngải tím là loài thảo dược bản địa ơ Ấn
Độ và Indonesia, nhưng cũng được trồng nhiều ơ Trung Quốc, Nhật Bản,
Brazil, Nepal, Thái Lan [78] và Việt Nam [7]. Nghệ đen từ lâu đã được sử dụng
trong y học cổ truyền của nhiều nước để điều trị các chứng viêm, đau nhức, các
bệnh về da như các vết thương và các vết lơ loét, cũng như sự bất thường của
chu kỳ kinh nguyệt [157].
Nhóm chất màu curcuminoid bao gồm curcumin và các dẫn xuất của nó
như demethoxycurcumin và bisdemethoxycurcumin là nhóm hợp chất chính
tạo nên các hoạt tính sinh học quan trọng của củ nghệ [76]. Các nghiên cứu gần
đây cho thấy curcuminoid, đặc biệt là curcumin, có phạm vi tác dụng dược lý
rộng như kháng viêm, kháng khuẩn, kháng virus, chống oxy hóa mạnh, chống
tia tử ngoại, ức chế phát triển khối u và bảo vệ thần kinh (kháng β-amyloid)
[14], [58], [116]. Curcuminoid được chiết xuất từ củ (thân rễ) của các loài nghệ
khác nhau, chẳng hạn C. caesia [24], C. amada [50], C. longa [67], C.
aromatica [74] và C. zedoaria [78]. Curcuminoid hiện đang được sử dụng như
dược chất trong nghiên cứu lâm sàng cho các bệnh nhân ung thư phổi, ung thư
trực tràng, viêm khớp dạng thấp, bệnh Alzheimer, bệnh vảy nến… [35]. Nhiều
nghiên cứu cũng đã cho thấy curcuminoid có độ an toàn cao, dung nạp tốt với
cơ thể, không độc đến liều 8 g/kg thể trọng [46].
Hiện nay, các gen tham gia trong quá trình tổng hợp curcuminoid ơ nghệ
vàng (C. longa) đã được xác định và phân tích mức độ biểu hiện, bao gồm hai
nhóm gen mã hóa enzyme type III polyketide synthase là diketide-CoA
synthase (DCS) và các curcumin synthase (CURS1, CURS2 và CURS3) [66],
[67]. Trước đó, Brand và cs. (2006) cũng đã mô tả một gen mã hóa enzyme

2
type III polyketide synthase khác là chalcone synthase (CHS) có ơ cây
Wachendorfia thyrsiflora, và gen này cũng tham gia vào quá trình tổng hợp
curcuminoid [28]. Behar và cs. (2016) khi phân tích biểu hiện của các gen tham
gia tổng hợp curcuminoid ơ C. caesia bao gồm DCS, CURS, CURS2, CURS3
và CHS1 đã nhận thấy mức độ biểu hiện của chúng trong củ cao hơn ơ lá [24].
Tuy nhiên, các gen tham gia tổng hợp curcuminoid ơ loài nghệ đen đến nay vẫn
chưa được công bố.
Elicitor là những chất hóa học được dùng để tác động vào con đường
chuyển hóa thứ cấp nhằm tăng cường sinh tổng hợp các chất có giá trị dược
phẩm trong nuôi cấy tế bào thực vật [62], [111]. Theo Abraham và cs (2011),
dịch chiết nấm men (YE) đã được ứng dụng trong nuôi cấy in vitro thực vật do
khả năng kích thích cơ chế bảo vệ, tăng sản sinh các chất chuyển hóa thứ cấp
có hoạt tính sinh học [9]. Salicilic acid (SA) được xem là một trong những tín
hiệu quan trọng trong phản ứng tự vệ của cây và cũng được sử dụng rộng rãi
trong sản xuất các chất chuyển hóa từ nuôi cấy tế bào thực vật [22]. Methyl
jasmonate (MeJA) cũng đã được chứng minh là một chất đóng vai trò quan
trọng trong việc truyền tín hiệu điều chỉnh khả năng phòng vệ của thực vật và
có thể kích thích sự sản sinh các chất chuyển hóa thứ cấp trong nuôi cấy tế bào
[158], [169].
Từ những lý do trên, chúng tôi thực hiện đề tài “Nghiên cứu ảnh hương
của chất kích kháng lên sự biểu hiện của một số gen tham gia vào quá trình
tổng hợp curcuminoid ơ tế bào nghệ đen (Curcuma zedoaria Roscoe)” nhằm
xác định các loại elicitor và nồng độ thích hợp của chúng để điều hòa tăng biểu
hiện của các gen mã hóa enzyme type III polyketide synthase trong con đường
phenylpropanoid ơ tế bào nghệ đen. Kết quả nghiên cứu của chúng tôi sẽ cung
cấp bằng chứng đầu tiên về vai trò của SA, YE và MeJA như là những chất
điều hòa dương tính của biểu hiện gen ơ loài dược liệu có giá trị này.
2. MỤC TIÊU NGHIÊN CỨU

3
Mục tiêu lý thuyết
Cải thiện mức độ biểu hiện của các gen CzDCS, CzCURS1, CzCURS2 và
CzCURS3 tham gia trong con đường chuyển hóa phenylpropanoid sinh tổng
hợp curcuminoid ơ tế bào nghệ đen nuôi cấy in vitro bằng một số elicitor.
Mục tiêu thực nghiệm
Tăng hiệu suất sinh tổng hợp curcumin, một thành phần chính của nhóm
chất curcuminoid được ứng dụng nhiều trong dược phẩm, ơ tế bào nghệ đen
nuôi cấy in vitro.
3. NỘI DUNG VÀ PHẠM VI NGHIÊN CỨU
Nội dung nghiên cứu
- Nuôi cấy sinh khối tế bào nghệ đen in vitro hiệu suất cao bằng cách bổ
sung AgNO3 vào môi trường nuôi cấy ơ quy mô phòng thí nghiệm.
- Phân lập các gen CzDCS, CzCURS1, CzCURS2 và CzCURS tham gia
trong quá trình tổng hợp curcuminoid (các gen curcuminoid) ơ nghệ đen.
- Nghiên cứu ảnh hương của một số elicitor như YE, SA và MeJA lên mức
độ biểu hiện của các gen tổng hợp curcuminoid và khả năng tích lũy curcumin
trong tế bào nghệ đen nuôi cấy in vitro.
4. ĐÓNG GÓP MỚI CỦA LUẬN ÁN
Luận án có các đóng góp mới như sau:
- Các nghiên cứu nuôi cấy mô và tế bào cây nghệ đen trước đây đều chưa
sử dụng AgNO3 để tăng hiệu quả nuôi cấy, trong nghiên cứu này, bổ sung
AgNO3 1,5 mg/L vào môi trường nuôi cấy (bao gồm tạo cây in vitro, nuôi cấy
callus) đều làm tăng hiệu quả của quá trình nuôi cấy so với các nghiên cứu
trước đây.
- Đã phân lập thành công 4 gen tham gia vào quá trình tổng hợp
curcuminoid, các gen này tương đồng 99% so với các gen tương ứng ơ cây
nghệ vàng và đã được đăng ký trên ngân hàng gen với các mã số lần lượt là

4
MF663785, MF402846, MF402847 và MF987835. Các gen này đều có sự biểu
hiện ơ trong củ và callus của cây nghệ đen.
- Gen DCS có vai trò lớn nhất trong các gen tham gia vào quá trình sinh
tổng hợp curcumin ơ cây nghệ đen, mức độ biểu hiện của gen này quyết định
trực tiếp đến hàm lượng curcumin thu được.
- Đã nghiên cứu được ảnh hương của một số loại elicitor (dịch chiết nấm
men và salicilic acid) lên khả năng tích lũy curcumin và mức độ biểu hiện của
các gen liên quan. Giá trị tốt nhất thu được khi xử lý dịch chiết nấm men (1
g/L) sau 5 ngày nuôi cấy, mức độ biểu hiện cao hơn 2,78 lần so với đối chứng.

5
Chương 1
TỔNG QUAN TÀI LIỆU
1.1. CÂY NGHỆ ĐEN
1.1.1. Đặc điểm thực vật học
Nghệ đen là cây thân thảo sống nhiều năm hoặc hàng năm, cao khoảng 1-
1,5m; có củ hình trứng, có khía chạy dọc, củ tỏa theo hình chân vịt; cây mẫm
và chắc. Cây có nhiều củ, ngoài củ chính ra còn có những củ phụ, vỏ củ có màu
xám, bên trong củ có màu vàng lưu huỳnh nhạt hoặc vàng sáng, khi già có nhiều
vòng màu xanh tím. Củ nghệ khô có mùi thơm camphor nhẹ và có vị đắng hơi
cay. Chồi lá nghệ đen có thể cao tới 1 m với 5 lá. Lá có bẹ ôm vào thân cây
phía dưới, dài 30-60 cm, rộng 7-8 cm, dọc theo gân chính giữa có những đốm
màu đỏ, cuống lá ngắn hay hầu như không có. Cụm hoa mọc ngang, dài 15-20
cm. Lá bắc phía dưới hình trứng hay hình mác tù, màu xanh lục nhạt, mép đỏ;
lá bắc phía trên màu vàng nhạt, đầu lá màu đỏ, không mang hoa. Hoa màu vàng,
đài hoa có thùy hình mác tù dài 15 mm, thùy giữa nhọn, cánh môi hẹp ơ phía
dưới nhưng hơi mơ rộng phía trên [59], [105].
Trong hai tháng 3 và 4, các cụm hoa từ chồi nách của thân và chồi đỉnh
của củ bắt đầu vươn lên mặt đất. Trên các điểm gần cụm hoa thường phát triển
các chồi và từ đó bắt đầu hình thành các nhánh mới. Vào mùa thu, lá trên mặt
đất bắt đầu lụi. Từ tháng 11 đến tháng 12, các chất chuyển hóa thứ cấp ơ củ bắt
đầu được tích lũy [7].
1.1.2. Phân bô
Cây nghệ đen được xem như là cây bản địa của vùng Đông Bắc Ấn Độ
nhưng hiện đang được trồng khắp nơi ơ Ấn Độ, Malaysia, Nhật Bản, Trung
Quốc, Thái Lan và Việt Nam. Ở Việt Nam, nghệ đen phân bố ơ các tỉnh miền
núi và trung du phía Bắc như Hà Giang, Lào Cai, Yên Bái… và một số tỉnh ơ

6
miền Trung. Hiện nay, cây nghệ đen cũng đã được trồng tại một số địa phương
khác của miền Bắc và Tây Nguyên [6].
1.1.3. Thanh phần hóa học chính của củ nghệ đen
Nghệ đen là loài thảo dược có củ chứa các nhóm chất chủ yếu như tinh
dầu (sesquiterpene và monosesquiterpene) và curcuminoid (curcumin,
demethoxycurcumin và bisdemethoxycurcumin). Ngoài ra, cây nghệ còn chứa
một số hợp chất khác như tinh bột, chất dẻo và các chất có vị đắng như tannin,
flavonoiod [76].
1.1.3.1. Tinh dầu
Từ năm 1928, Rao và cs đã khảo sát sơ bộ thành phần hóa học của tinh
dầu củ nghệ đen và tìm thấy các hợp chất như α-pinen, borneol, camphen,
camphor và cineol bên cạnh các sesquiterpene, nhưng không phân lập và xác
định được loại sesquiterpene nào. Xingyi (1999) khi nghiên cứu tinh dầu nghệ
đen ơ Trung Quốc nhận thấy chúng chứa 37 thành phần khác nhau, trong đó
chủ yếu là curzerenone (45,02%), curcumenol (8,31%), β-elemene (5,79%) và
isocurcumenol (4,05%) [164]. Trong tinh dầu nghệ đen sinh trương ơ vùng
Đông Bắc Ấn Độ, Tohda và cs (2006) đã thu được 37 hợp chất, chiếm 87,7%
lượng tinh dầu tổng số, chủ yếu là curzerenone (22,3%), tiếp đến là 1,8-cineole
(15,9%) và germacrone (9%), β-tumerone (19,88%) và zingiberene (7,84%)
[155]. Duke và cs (2003) đã tìm thấy trong củ nghệ đen một số sesquiterpenoid
như ar-turmerone zederone, β-turmerone, curcumadiol, curcumenol, curcumol,
curcolone, curdione, curzerene, curzerenone, dehydrocurdione,
epicurzerenone, furanodiene, isocurcumenol, procurcumenol và zingiberene
[41].
Ở Việt Nam, khảo sát thành phần hóa học của nghệ đen cũng đã và đang
được quan tâm nghiên cứu. Thành phần chính trong tinh dầu củ nghệ thu được
tại Sóc Sơn (Hà Nội) là zurumbon (chiếm 79,08%) [3], trong khi tinh dầu nghệ

7
đen ơ Đô Lương (Nghệ An) và Hương Sơn (Hà Tĩnh) đều giàu epicurzerenon
và germacrone. Các hợp chất khác trong tinh dầu có hàm lượng thấp hơn đó là
α-cadinol, β-elemen, β-pinen, δ-cadinen, 1,8-cineol, 2,4-diisopropenyl-1-vinyl-
cyclohexan, benzofuran-6-ethyxyl-4,5,6,7-tetrahydro-3,6-dimethyl-5
isopropyl, camphor, germacrene, isoborneol, T-muurolol và zingiberen [1].
Thành phần hóa học của tinh dầu củ nghệ đen trồng ơ Đà Lạt có chứa các hợp
chất như γ-elemen (14,18-18,79%), curzeren (14,28-16,67%), và germacrone
(22,53-24,28%) [4].
1.1.3.2. Curcuminoid
Bên cạnh các nghiên cứu về thành phần hóa học của tinh dầu, các nghiên
cứu về thành phần hợp chất màu vàng có trong nghệ đen cũng được quan tâm.
Syu và cs (1998) nhận thấy dịch chiết ethanol của củ nghệ đen có chứa các hợp
chất nhóm curcuminoid là curcumin, demethoxycurcumin và
bisdemethoxycurcumin (Hình 1.1) [149].
Curcumin (C21H20O6) là một hợp chất dạng tinh thể, màu vàng cam, có
trong các loài thực vật thuộc chi Curcuma, tan tốt trong các dung môi hữu cơ
như acetone, methanol, ethanol và isopropanol nhưng không tan trong nước.
Các dung môi hòa tan thích hợp để tách chiết curcumin là acetone,
dichloromethan, ethanol ethyl acetate, methanol, n-butanol và hexane.
Curcumin có thể được thu hồi bằng cách kết tinh từ dịch chiết [2], [60].

8
Hình 1.1. Cấu trúc hóa học của các curcumin.
1.1.4. Công dụng của nghệ đen
Cây nghệ đen được trồng phổ biến dùng làm rau hoặc đồ gia vị ơ các nước
vùng Đông và Nam Á bao gồm Ấn Độ, Nhật Bản, Trung Quốc, Malaysia, Thái
Lan và Việt Nam [59], [135]. Từ lâu, nghệ đen đã được sử dụng như một vị
thuốc trong bài thuốc Đông y cổ truyền để chữa bệnh. Nghệ đen cũng được kê
trong các đơn thuốc dùng để chữa bệnh dạ dày, điều trị hội chứng “Oketsu” gây
ra do tắc nghẽn mạch máu, điều kinh và cải thiện kinh nguyệt với nhiều dạng
pha chế khác nhau [96]. Ở Thái Lan, nghệ đen được dùng để làm dịu cơn đau
dạ dày, chống tiêu chảy, chống nôn mửa và sốt hoặc làm se các vết thương ơ
ngoài da [135]. Người Ấn Độ đã dùng củ nghệ đen để trị chứng viêm da, bong
gân, ung nhọt và vết thương [41]. Ngoài ra, củ nghệ đen còn được dùng để chữa
giun sán ơ trẻ em; bột nghệ dùng để chống dị ứng; lá nghệ có tác dụng chữa
bệnh phù hay phong hủi [64], [109]. Ngày nay, nhiều hoạt chất sinh học của
nghệ đen đã được chứng minh có tác dụng giảm đau, bảo vệ gan, kháng ung
thư, kháng viêm, kháng khuẩn, chống oxy hóa, chống đột biến…
1.1.4.1. Hoạt tính giảm đau
Curcumin
H3CO
HO
OCH3
OH
O O
HO
OCH3
OH
O O
Demethoxycurcumin
HO OH
O O
Bisdemethoxycurcumin

9
Shin và cs (1994) khảo sát hoạt tính dược lý của hai loại sesquiterpene là
cuzerenone (I) và curcumenol (II) từ củ nghệ đen trên thỏ và chuột thực nghiệm.
Kết quả cho thấy, cả hai chất này đều thể hiện hoạt tính giảm đau tương đối
[144]. De Navarro và cs (2002) khi nghiên cứu hoạt tính giảm đau của nghệ
đen ơ Brazil đã nhận thấy hợp chất curcumenol có tác dụng giảm đau cao hơn
nhiều lần so với các loại thuốc giảm đau thông thường như aspirin và dipyrone
[38].
1.1.4.2. Hoạt tính kháng ung thư
Nghiên cứu của Hong và cs (2002) cho thấy, dịch chiết nghệ đen có hoạt
tính chống ung thư và kháng viêm. Khả năng ức chế sinh tổng hợp các chất
prostalandin và NO của dịch chiết nghệ được cho là có tiềm năng trong việc
kháng viêm và trong hóa trị liệu chống ung thư [59]. Seo và cs (2005) nhận
thấy dịch chiết nước của củ nghệ có hoạt tính chống di căn phổi của các tế bào
khối u ác tính B16 [140]. Syu và cs (1998) đã nghiên cứu hoạt tính kháng ung
thư của các curcuminoid thu được từ dịch chiết ethanol của củ nghệ. Kết quả
cho thấy, chúng có hoạt tính gây độc đối với các tế bào ung thư buồng trứng
OVCAR-3 ơ người [149]. Các nghiên cứu của Jiang và cs (1996), Hanif và cs
(1997) cũng cho thấy curcuminoid phân lập từ nghệ đen có khả năng ức chế
sinh trương khối u và gây độc cho các dòng tế bào ung thư ruột kết và ung thư
biểu mô gan ơ người [55], [63].
1.1.4.3. Hoạt tính bảo vệ gan
Matsuda và cs (1998) nhận thấy dịch chiết acetone-nước của củ nghệ đen
có hoạt tính bảo vệ gan, sesquiterpene và curcumin chống lại sự hình thành D-
galactosamine/lipopolysaccharide làm giảm tổn thương gan ơ chuột [95]. Kết
quả nghiên cứu của Kim và cs (2005) cho thấy, nghệ đen có thể được sử dụng
như một loại thuốc tiềm năng trong điều trị chứng xơ gan mãn tính [70].
1.1.4.4. Hoạt tính kháng viêm và chống loét

10
Nghệ đen còn được sử dụng như là phương thuốc chủ yếu để điều trị các
vị trí bị loét trong hệ tiêu hóa. Nghiên cứu của Raghuveer và cs (2003) cho thấy
dịch chiết nghệ đen có khả năng chống lại tình trạng tiết nhiều acid và viêm
loét dạ dày [122]. Watanabe và cs (1986) đã nghiên cứu hoạt tính kháng loét
của 8 loại dịch chiết từ nghệ đen trồng ơ Yakushima (Nhật Bản) trên chuột gây
viêm loét dạ dày cấp tính. Các hợp chất furanogermenone và (4S, 5S)-(+)
germacrone 4,5-epoxide phân lập từ tinh dầu nghệ đen có hoạt tính ức chế sự
hình thành vết loét trên chuột thực nghiệm [161]. Jang và cs (2001) nhận thấy
ba hợp chất epiprocurcumenol, procurcumenol và 7-bis (4-hydroxyphenyl)-
1,4,6-heptatrien-3-one từ dịch chiết củ nghệ đen có hoạt tính kháng viêm [61].
1.1.4.5. Hoạt tính chống oxy hóa
Theo Matsuda và cs (1993), các hợp chất curcumin, demethoxycurcumin
và bisdemethoxycurcumin phân lập từ củ nghệ đen có hoạt tính chống oxy hóa
và kháng viêm tương đương với các chất này thu được từ củ nghệ vàng [94].
Dịch chiết ethanol của củ nghệ đen ơ Thái Lan có hoạt tính bắt gốc tự do DPPH
với nồng độ ức chế 50% (EC50) từ 18,29-40,33 μg/mL (trung bình 25,71
μg/mL). Khả năng bắt gốc tự do của các curcuminoid phân tách từ dịch chiết
ethanol của nghệ đen lần lượt là: curcumin (EC50 = 7,89 μg/mL),
demethoxycurcumin (EC50 = 9,52 μg/mL) và bisdemethoxycurcumin (EC50 =
149,09 μg/mL) [114]. Trần Thị Việt Hoa và cs (2007) nhận thấy tinh dầu nghệ
đen trồng ơ Đà Lạt ơ nồng độ 20 mg/mL có khả năng chống oxy hóa tương đối
cao, từ 74,8-77,8% [4]. Nghiên cứu của Loc và cs (2008) cho thấy hoạt tính của
các enzyme chống oxy hóa như peroxidase, superoxide dismutase và catalase
trong tế bào nghệ đen đạt giá trị cao nhất sau 14 ngày nuôi cấy, lần lượt là
0,63 U/mg, 16,60 U/mg và 19,59 U/mg protein [81].
1.1.4.6. Hoạt tính kháng khuẩn và kháng nấm
Hoạt tính kháng các loại vi khuẩn và nấm gây bệnh ơ người và thực vật
của nghệ đen cũng được quan tâm nghiên cứu. Theo Wilson (2005), dịch chiết
từ củ nghệ đen có khả năng kháng năm chủng vi khuẩn khác nhau là Bacillus

11
subtilis NCIM 2603, Escherichia coli NCIM 2574, Klebsiella pneumoniae
NCIM 2957, Micrococcus luteus NCIM 2103 và Proteus mirabilis NCIM
2300, và hai chủng nấm là Aspergillus niger NCIM 596 và Candida albicans
NCIM 3102 [162]. Dịch chiết ethanol của nghệ đen trồng ơ Ấn Độ có khả năng
kháng B. cereus ơ nồng độ 1.000 μg/mL và có hoạt tính ức chế tương đối với
C. albicans và K. pneumoniae [145]. Nghiên cứu của Giang và cs (2000) cho
thấy, dịch chiết củ nghệ đen trồng ơ Việt Nam có phổ kháng khuẩn rộng. Các
chất như a-humulene, humulene-8-hydroperoxide, zerumbone và zerumbone-
2,3-epoxide có hoạt tính kháng lại các loài vi khuẩn Gram (-) và Gram (+) được
thử nghiệm [49].
1.1.4.7. Các hoạt tính khác
Ansari và Ahmad (1991) nhận thấy dịch chiết alcohol từ củ nghệ đen nồng
độ 1-10 mg/mL có khả năng ức chế sự phát triển của amoeba (Entamoeba
histolytica) [16]. Champakaew và cs (2007) khi phân tích thành phần hóa học
và thử hoạt tính kháng muỗi Aedes aegypti mang virus gây bệnh sốt Dengue
của tinh dầu nghệ đen đã nhận thấy chúng thật sự có hiệu quả trong việc giết
chết các ấu trùng muỗi, đây được xem là nguồn tinh dầu thay thế triển vọng để
phát triển các loại thuốc diệt ấu trùng muỗi trong tương lai [30]. Daduang và cs
(2005) nghiên cứu khả năng kháng nọc độc rắn của dịch chiết nghệ đen cho
thấy nó có tác dụng ngăn cản nọc độc di chuyển trước khi bổ sung kháng thể
kháng độc [37].
1.2. NUÔI CẤY MÔ TẾ BÀO THỰC VẬT TRONG SẢN XUẤT HỢP
CHẤT THỨ CẤP
1.2.1. Sự tích lũy các hợp chất thứ cấp trong nuôi cấy tế bao thực vật
Kỹ thuật nuôi cấy tế bào thực vật có nhiều triển vọng và ứng dụng lâu dài
trong việc sản xuất các hợp chất tự nhiên, đặc biệt là các chất dùng trong y học.
Hướng nghiên cứu này sẽ dẫn đến sự ổn định về mặt chất lượng và số lượng
sản phẩm, ít phụ thuộc vào tự nhiên. Đồng thời, là nguồn nguyên liệu cho những

12
thí nghiệm sinh lý, hóa sinh và ứng dụng để tách chiết các hợp chất thứ cấp
khác nhau [5].
Các nghiên cứu cho thấy rằng nuôi cấy tế bào thực vật có khả năng sản
xuất các sản phẩm thứ cấp với hàm lượng lớn hơn so với các chất đó được chiết
từ cây ngoài tự nhiên. Ưu điểm của chúng là có thể cung cấp sản phẩm một
cách liên tục và đáng tin cậy dựa trên cơ sơ:
- Các tế bào thực vật có thể được nuôi cấy trong các điều kiện nhân tạo
nên không phụ thuộc vào thời tiết và địa lý, không cần phải vận chuyển và bảo
quản một số lượng lớn các nguyên liệu thô.
- Có thể kiểm soát chất lượng và hiệu suất của sản phẩm bằng cách loại
bỏ các trơ ngại trong quá trình sản xuất thực vật.
- Phủ định ảnh hương sinh học đến các sản phẩm là hợp chất thứ cấp trong
tự nhiên (vi sinh vật và côn trùng).
- Một số sản phẩm trao đổi chất có thể được sản xuất từ nuôi cấy dịch
huyền phù có chất lượng cao hơn trong cây hoàn chỉnh [5].
- Chọn lọc các giống cây trồng cho nhiều loại hợp chất thứ cấp khác nhau.
- Với việc tự động hóa điều khiển sự sinh trương của tế bào và điều hòa
quá trình chuyển hóa, chi phí có thể giảm và lượng sản phẩm sẽ tăng lên [108].
Thực vật có khả năng sản xuất số lượng lớn các hợp chất thứ cấp và sự
phân bố các hợp chất thứ cấp ơ mức độ tế bào phụ thuộc vào con đường sinh
tổng hợp và đặc điểm cấu trúc của chúng [112]. Trong đó, các không bào dự
trữ thường chiếm 40-90% thể tích tế bào thực vật, chúng đóng vai trò then chốt
trong sự tích lũy các hợp chất thứ cấp ơ thực vật. Sự tích lũy các hợp chất thứ
cấp trong không bào có ít nhất hai vai trò được xác định đó là lưu trữ tạm thời
các hoạt chất sinh học nội sinh trong tế bào và bảo vệ chúng khỏi quá trình dị
hóa [52]. Các nghiên cứu cũng cho thấy có hai cơ chế vận chuyển các hợp chất
thứ cấp chủ yếu trong không bào đó là vận chuyển theo gradient H+ qua kênh

13
vận chuyển ion H+ và vận chuyển sơ cấp cần năng lượng trực tiếp bơi các chất
mang dạng hình hộp liên kết với phân tử ATP [93]. Một số nghiên cứu khác
cũng nhận thấy, gen không những liên quan đến sinh tổng hợp các hợp chất thứ
cấp mà còn liên quan đến các yếu tố vận chuyển chúng. Kết quả nghiên cứu
này sẽ hữu ích trong công nghệ trao đổi chất nhằm tăng khả năng sản xuất các
hợp chất thứ cấp có giá trị ơ thực vật [165].
1.2.2. Các phương pháp nuôi cấy sử dụng trong sản xuất hợp chất thứ
cấp từ thực vật
1.2.2.1. Nuôi cấy callus
Nuôi cấy callus là quá trình nuôi cấy các tế bào thực vật không biệt hóa
được hình thành bằng cách nuôi cấy các mô trên môi trường chứa nồng độ
auxin cao hoặc kết hợp auxin và cytokinin trong điều kiện in vitro. Nuôi cấy
callus đã được ứng dụng trong sản xuất các hợp chất thứ cấp thực vật đặc biệt
là các hợp chất flavonoid. Quy trình nuôi cấy callus ổn định và tối ưu là bước
quan trọng trong giai đoạn đầu của quá trình nuôi cấy huyền phù để sản xuất
hợp chất thứ cấp ơ quy mô lớn hơn [153].
1.2.2.2. Nuôi cấy huyền phù tế bào
Nuôi cấy huyền phù tế bào là quá trình nuôi cấy các tế bào hay một khối
tế bào nhỏ không biệt hóa của thực vật trong môi trường lỏng và được duy trì
trong điều kiện sục khí, kích thích, ánh sáng, nhiệt độ và các thông số vật lý
thích hợp. Các tế bào nuôi cấy không chỉ có thể tạo ra các hợp chất hóa sinh
tiêu chuẩn xác định với khối lượng lớn mà còn loại bỏ sự hiện diện của nhiều
hợp chất không mong muốn khác với trong cây tự nhiên. Nuôi cấy huyền phù
tế bào được ứng dụng phổ biến nhất để sản xuất thứ cấp ơ quy mô lớn. Một số
loại bioreactor khác nhau đã được sử dụng cho quá trình nuôi ấy. Ứng dụng
thương mại đầu tiên về nuôi cấy tế bào quy mô lớn được thực hiện trong các

14
bioreactor bể khuấy có công suất 200 lít và 750 lít để sản xuất shikonin từ
Lithospermum erythrorhizon. Tế bào của Catharanthus roseus, Dioscorea
deltoidea, Digitalis lanata, Panax notoginseng, Taxus wallichiana và
Podophyllum hexandrum đã được nuôi cấy trong các bioreactor khác nhau để
sản xuất các sản phẩm thực vật thứ cấp. Những tiến bộ trong lĩnh vực nuôi cấy
tế bào để sản xuất các hợp chất dược phẩm đã tạo ra nhiều loại dược phẩm như
alkaloids, terpenoids, steroid, saponin, phenolics, flavanoid. Bằng cách cải biến
quy trình nuôi cấy, sản lượng hợp chất thứ cấp thu được có thể tăng cao (dẫn
theo Plunkett và cs. 2004) [119].
1.2.2.3. Nuôi cấy rễ tơ
Hệ thống nuôi cấy rễ tơ (hairy root) đã trơ nên phổ biến trong hai thập
kỷ qua như một phương pháp để sản xuất các chất chuyển hóa thứ cấp được
tổng hợp trong rễ. Rễ tơ là các rễ đột biến trong quá trình nuôi cấy biệt hóa
được tạo ra bơi sự lây nhiễm của Agrobacterium rhizogenes vào các loài thực
vật bậc cao bị tổn thương. Tác nhân gây bệnh này gây ra sự nhiễm bệnh dẫn
đến sự phát triển các khối u của rễ được đặc trưng bơi tốc độ tăng trương cao
trong môi trường không chứa hormone và sự ổn định di truyền khi nuôi cấy
trong một thời gian dài. Rễ tơ có con đường tổng hợp các hợp chất cũng giống
với ơ các cơ quan hoang dại nhưng lại cho năng suất cao hơn nhiều lần. Chính
đặc tính ổn định và năng suất cao cho phép khai thác rễ tơ là công cụ công nghệ
sinh học có giá trị để sản xuất các chất chuyển hóa thứ cấp của thực vật [113],
[119].
1.2.3. Vai trò của AgNO3 trong nuôi cấy mô tế bao thực vật
Nano bạc bao gồm các hạt bạc có kích thược nano, khoảng từ 1-100 nm,
thông thường kích thước đo được khoảng 25 nm. Theo Kumar và cs (2009) và

15
Sandra & Maira (2013), AgNO3 khi được bổ sung vào môi trường nuôi cấy mô
thực vật có tác dụng ức chế hoạt động của ethylene nội sinh, vì thế quá trình
sinh trương của tế bào sẽ được thuận lợi hơn [73], [134].
Đến nay, AgNO3 đã được sử dụng rộng rãi trong nuôi cấy mô thực vật để
tái sinh chồi, tăng hệ số nhân giống, phát sinh cơ quan, cải thiện các thông số
hóa sinh và thậm chí chuyển gen thông qua Agrobacterium… (Kumar và cs.
2009) [73], Sandra và Maira. 2013 [134], Sgamma và cs. 2015 [141],
Sarropoulou và cs. 2016 [137], Mohiuddin và cs. 1997 [104], Tamini và cs.
2015 [151], Harathi và cs. 2016 [56], Da Silva và cs. 2013 [152]). Park và cs
(2016) cho rằng YE và AgNO3 có thể cảm ứng làm tăng mức độ biểu hiện của
gen sinh tổng hợp phenylpropanoid và tăng cường tích lũy rosmarinic acid
trong tế bào cây hoắc hương núi (Agastache rugosa) [115].
1.3. ELICITOR VÀ CÁC ỨNG DỤNG
1.3.1. Elicitor
1.3.1.1. Khái niệm
Elicitor (hay chất kích kháng) được định nghĩa như là một chất cơ bản mà
khi đưa một lượng nhỏ vào hệ thống tế bào sống thì có thể khơi động hoặc cải
thiện sự sinh tổng hợp các hợp chất thứ cấp của tế bào [111]. Sự kích kháng
thực vật (elicitation) là quá trình cảm ứng tăng cường sinh tổng hợp các chất
chuyển hóa thứ cấp nhờ tác động của các elicitor, giúp cho cây chống lại các
yếu tố bất lợi của ngoại cảnh như tác nhân gây bệnh hoặc điều kiện sinh thái
khắc nghiệt [143].
1.3.1.2. Phân loại
Elicitor có thể được phân loại dựa trên bản chất tự nhiên của chúng là
elicitor phi sinh học (abiotic elicitor) và elicitor sinh học (biotic elicitor), hoặc
dựa vào nguồn gốc của chúng là elicitor ngoại sinh (exogenous elicitor) và
elicitor nội sinh (endogenous elicitor). Elicitor phi sinh học là các chất có nguồn

16
gốc không thuộc sinh vật, chủ yếu là các muối vô cơ và các tác nhân vật lý, ví
dụ: các ion Cu2+, Cd2+, Ca2+, độ pH cao... Elicitor sinh học là các chất có nguồn
gốc từ sinh vật, bao gồm các polysacharide của thành tế bào thực vật (cellulose
hoặc pectin) và vi sinh vật (chitin hoặc glucan), hoặc các glycoprotein, G-
protein hay các protein nội bào có chức năng gắn các receptor và tác động bằng
cách hoạt hóa hay bất hoạt một số các enzyme hoặc các kênh ion. Elicitor ngoại
sinh là các chất có nguồn gốc bên ngoài tế bào như các acid béo, các
polysaccharide và polyamine. Ngược lại, elicitor nội sinh là các chất có nguồn
gốc bên trong tế bào như là galacturonide, hepta-β-glucoside... (Bảng 1.1)
[111].
1.3.1.3. Cơ chế kích kháng
Elicitor là các chất có nguồn gốc khác nhau, có khả năng gây nên các đáp
ứng về mặt hình thái, sinh lý và tích lũy phytoalexin (chất được sinh ra khi thực
vật chịu tác động của các tác nhân gây bệnh). Chúng có thể là các elicitor phi
sinh học như ion kim loại, hợp chất vô cơ hoặc elicitor sinh học có nguồn gốc
từ nấm, vi khuẩn hoặc động vật ăn cỏ, mảnh vỡ của thành tế bào thực vật cũng
như các chất được giải phóng ra tại vị trí tổn thương của thực vật do mầm bệnh
hoặc động vật tấn công.
Thực vật được xử lý elicitor hoặc bị mầm bệnh tấn công gây ra một loạt
các phản ứng phòng vệ, bao gồm sự tích lũy các hợp chất thứ cấp bảo vệ ơ cả
trong cây tự nhiên cũng như trong nuôi cấy in vitro. Mặc dù đã có nhiều nghiên
cứu về cơ chế ảnh hương của elicitor lên quá trình sinh tổng hợp các chất thứ
cấp ơ thực vật nhưng cơ chế của sự kích kháng vẫn chưa được hiểu đầy đủ. Có
nhiều giả thuyết đã được đưa ra như cơ chế truyền tin bơi Ca2+, các yếu tố ảnh
hương đến sự nguyên vẹn của màng tế bào, các con đường ức chế/hoạt hóa nội
bào hay sự thay đổi áp suất thẩm thấu [111].
Bảng 1.1. Phân loại các elicitor khác nhau [111]
A. Theo bản chất

17
Elicitor sinh học Elicitor phi sinh học
- Được giải phóng trực tiếp từ vi sinh
vật và được nhận diện bơi tế bào thực
vật (các enzyme, các mảnh vỡ của
thành tế bào).
- Được tạo thành bơi hoạt động của vi
sinh vật trên thành tế bào thực vật
(pectin).
- Được tạo ra từ hoạt động của
enzyme thực vật trên thành tế bào vi
khuẩn (chitosan, glucan).
- Các hợp chất nội sinh được hình
thành hoặc tiết ra bơi tế bào thực vật
khi đáp ứng các kích thích khác nhau.
- Tác dụng của các tác nhân vật lý
hoặc hóa học tự nhiên theo đường nội
sinh tạo thành các elicitor sinh học.
- Tia UV.
- Protein biến tính (RNase).
- Đông và rã đông.
- Các thành phần không thiết yếu của
môi trường (agarose).
- Kim loại nặng.
- Các loại hóa chất có ái lực cao với
DNA, có hoạt tính phá vỡ màng tế
bào, thuốc diệt nấm (benomyl,
butylamin, maneb), và thuốc diệt cỏ
(acifluorofen).
B. Theo nguồn gôc
Elicitor ngoại sinh Elicitor nội sinh
- Hình thành từ bên ngoài tế bào, bao
gồm các phản ứng trực tiếp hoặc qua
các chất nội sinh trung gian.
- Các polysaccharide: chitosan,
glucan, và glucomanose.
- Peptide và chuỗi các ion dương:
glycoprotein, monilicolin, polyamine,
và poly-L-lysine.
- Các enzyme: polygalacturonase,
cellulase, và endo-polygalacturonase
acid lyase.
- Các acid béo: acid arachidonic và
acid eicosapentanoic.
- Hình thành qua các phản ứng thứ
cấp, cảm ứng bằng một tín hiệu sinh
học hoặc phi sinh học trong tế bào.
- Alginate oligomer.
- Dodeca-β-1,4-D-galacturonide.
- Hepta-β-glucoside.
Một số nghiên cứu đã giả thiết có sự liên kết của elicitor vào các thụ thể

18
trên màng sinh chất để kích hoạt quá trình chống chịu [32], [54]. Gelli và cs
(1997) cho rằng khi có kích kháng, Ca2+ di chuyển vào tế bào chất từ bên ngoài
tế bào [48]. Một số tác giả nhấn mạnh đến sự thay đổi nhanh chóng của quá
trình phosphoryl hóa protein và kích hoạt kinase chính là cơ chế của quá trình
kích kháng [47], [129]. Trong khi đó, nhiều tác giả khác nhận thấy có sự tích
lũy mitogen-activated protein kinase (MAPK) và kích hoạt G-protein trong quá
trình kích kháng [10], [40]. Armero và Tena (2001) cho rằng, có sự acid hóa
màng nguyên sinh chất gây ra bơi sự bất hoạt H+-ATPase, trong khi giảm sự
phân cực của màng, tăng pH bên ngoài màng được quan sát thấy khi xử lý
elicitor [19]. Apostol và cs (1989), Bolwell và cs (1995) và Pugin và cs (1997)
giải thích rằng, việc sản xuất các ROS như anion superoxide và H2O2 tạo ra
hiệu ứng kháng khuẩn trực tiếp cũng như góp phần tạo ra các dẫn xuất của acid
béo có hoạt tính sinh học [17]. Tương tự, ROS tham gia vào quá trình liên kết
với protein giàu proline gắn trên thành tế bào sau đó hoạt động như là tín hiệu
thứ cấp và kích hoạt dịch mã các gen bảo vệ [89].
Theo một giả thuyết khác, sự tích lũy của các protein liên quan đến việc
bảo vệ thực vật khỏi các tác nhân gây bệnh như chitinase, glucanase và
endopolygalacturonase góp phần giải phóng các pectic oligomer tín hiệu
(elicitor nội sinh), glycoprotein giàu hydroxyproline và chất ức chế protease
[25]. Ngoài ra, sự kích hoạt phiên mã của các gen bảo vệ trong quá trình kích
kháng cũng đã được công bố [99], [139].
1.3.1.4. Các yếu tố ảnh hương đến sự kích kháng
Tăng cường sản xuất hợp chất thứ cấp có hoạt tính sinh học từ nuôi cấy tế
bào thực vật bằng cách xử lý chúng với các elicitor thích hợp đã mơ ra một
hướng nghiên cứu mới trong công nghiệp dược phẩm. Các thông số như thành
phần và nồng độ elicitor, thời gian xử lý, tuổi của tế bào, dòng tế bào, chất điều
hòa sinh trương, môi trường dinh dưỡng và đặc điểm thành tế bào phù hợp có

19
thể tăng khả năng tích lũy các sản phẩm thứ cấp [111].
Nồng độ elicitor: Nồng độ elicitor đóng vai trò rất quan trọng trong quá
trình kích kháng. Namdeo và cs (2002, 2007) nhận thấy sự tích lũy ajmalicine
cao trong tế bào cây dừa cạn (Catharanthus roseus) được xử lý dịch chiết nấm
Trichoderma viride, Aspergillus niger và Fusarium moniliforme ơ các nồng độ
khác nhau. Lượng ajmalicine trong tế bào được xử lý elicitor nồng độ cao (5%)
lớn hơn so với nồng độ thấp (0,5%). Tuy nhiên, nồng độ elicitor >10% đã ảnh
hương bất lợi lên sự tích lũy ajmalicine do chúng gây ra hiện tượng đáp ứng
quá ngưỡng, dẫn đến chết tế bào [111], [110].
Thời gian xử lý elicitor: Nghiên cứu của Namdeo và cs (2002, 2007) cho
thấy lượng ajmalicine tăng khoảng 3 lần ơ tế bào dừa cạn được xử lý dịch chiết
của T. viride trong 48 giờ, trong khi đó chất này chỉ tăng 2 lần ơ tế bào được
xử lý dịch chiết của A. niger và F. moniliforme trong cùng thời gian. Khi thời
gian xử lý lớn hơn 96 giờ khả năng tích lũy ajmalicine của tế bào đã giảm rõ
rệt [111], [110].
Tuổi của tế bao: Thời điểm cấy chuyển là một yếu tố quan trọng trong
sản xuất các chất chuyển hóa sinh học từ các tế bào được xử lý elicitor. Namdeo
và cs (2002, 2007) nhận thấy lượng ajmalicine đạt cao nhất (166 µg/g khối
lượng khô (dw)) ơ tế bào 20 ngày tuổi được xử lý dịch chiết của T. viride [111],
[110].
Môi trường dinh dưỡng: Thành phần môi trường cũng đóng một vai trò
quan trọng trong quá trình kích kháng. Theo Namdeo và cs (2002, 2007),
ajmalicine được tích lũy ơ tế bào sinh trương trong môi trường Zenk nhiều hơn
so với môi trường MS [111], [110].
Ngoài ra, sự kích kháng cũng phụ thuộc vào đặc tính của các elicitor, dòng
tế bào hoặc chủng vi sinh vật được sử dụng làm elicitor, sự có mặt của các chất
điều hòa sinh trương cũng như các điều kiện nuôi cấy khác [111].

20
1.3.2. Ứng dụng của elicitor
Đến nay, đã có nhiều công trình ứng dụng elicitor để kích thích sản xuất
các hợp chất thứ cấp từ nuôi cấy in vitro thực vật. Các nghiên cứu đều cho thấy
sự tích lũy của những chất này đã tăng lên đáng kể so với đối chứng sau khi
được xử lý với loại elicitor và nồng độ thích hợp của chúng (Bảng 1.2).
1.3.2.1. Elicitor sinh học
Thông thường, các elicitor có nguồn gốc phi sinh học hoặc kết hợp elicitor
sinh học và phi sinh học được sử dụng phổ biến hơn. Tuy nhiên, cũng có tác giả
chỉ sử dụng elicitor sinh học trong các nghiên cứu của mình. Chẳng hạn:
Cousins và cs (2010) đã nuôi cấy cây nghệ vàng (C. longa) trong hệ lên
men thể tích 2,5 L có bổ sung nhiều loại elicitor khác nhau để cải thiện khả
năng sinh tổng hợp các chất chuyển hóa thứ cấp của chúng. Chẳng hạn: 1)
phenylalanine được bổ sung vào môi trường nuôi cấy từ tuần thứ 12-17; và 2)
proline, dịch chiết cá giàu proline hoặc chitosan được bổ sung vào tuần thứ 20.
Ở trường hợp 1, sinh khối của cây và sự tích lũy các chất chống oxy hóa của
chúng đã giảm sau 5 tuần xử lý. Ở trường hợp 2, hàm lượng các hợp chất phenol
tăng lên 4,7% dw (đối chứng 4,1% dw) khi xử lý nitrogen ơ nồng độ thấp sau
1,5 tuần [36].
Khi nuôi cấy in vitro cây nghệ C. mangga, Abraham và cs (2011) nhận
thấy xử lý YE không ảnh hương lên sinh khối và hình thành chồi ơ cây con.
Ngoài ra, cây con có hình thái bất thường khi được xử lý YE ơ nồng độ cao
(>3,5 mg/L). Tuy nhiên, ơ các môi trường có bổ sung YE, cây con tích lũy
nhiều chất có hoạt tính bắt gốc tự do. Cây con được nuôi trên môi trường có bổ
sung chitosan 150 mg/L và YE 3,5 mg/L có hàm lượng chất bắt gốc tự do nhiều
hơn các môi trường khác [9].

21
Bảng 1.2. Một sô ứng dụng elicitor để cải thiện khả năng tích lũy các hợp chất thứ cấp trong nuôi cấy in vitro [111]
Loai thực vật Mẫu vật
nuôi cấy
Elicitor Sản phẩm thứ cấp Ham lượng
Không xử lý Xử lý
Capsicum annum Tế bào T. viride (thô) Capdisol 0 1 mg/bình
Carthamus tinctorius Rễ tơ Vi khuẩn lam (thô) Kinobeon A 0,6 mg/L 5,78 mg/L
C. roseus Rễ tơ Penicillium sp. (thô) Alkaloids (indole) 3 mg/g dw 9 mg/g dw
C. roseus Tế bào Alteromonas macleodii, alginate Phosphodiesterase 0,022 U/mL <0,235 U/mL
C. roseus Tế bào Pythium sp. (thô) Ajmalicine 0 400 µg/L
C. roseus Tế bào T. viride Ajmalicine 79 µg/g dw 166 µg/g dw
C. roseus Tế bào Pythium aphanidermatum (thô) Alkaloids (indole) 50 µmol/L 75 µmol/L
Cupressus usitanica Tế bào Nấm β-thujaplicin 0 187 µg/g dw
Datura stramonium Tế bào Phytopthora megasperma (thô) Alkaloids (tropane) 0,85 mg/g dw 4,27 mg/g dw
D. stramonium Rễ tơ CuSO4 Rishitin 0 Vết
D. stramonium Rễ tơ CdCl2 Sesquiterpenes 0 140 nM/g
Dioscorea deltoidea Tế bào Rhizopus arrhizus (thô) Diosgenin 134 mg/L 230 mg/L
Eschscholtzia californica Tế bào Dịch chiết nấm men (thô) Sanguinarine 20 mg/L 60 mg/L

22
Hyoscyamus muticus Rễ tơ Rhizoctonia solani Sesquiterpenes 0 1 mg/10 g fw
Lithospermum
erythrorhizon
Tế bào Các tác nhân nội sinh (thô) Shikonin 0 28 µg/10mL
Lotus corniculatus Rễ tơ Glutathione Isoflavonoids 0 160 µg/g
Morinda citrifolia Tế bào Chitin 50 (tinh khiết) Anthraquinones 3 µg/g dw 7 µg/g dw
Nicotiana tabacum Rễ tơ Dịch chiết nấm men Sesquiterpenes 1 µg/g dw 87 µg/g dw
N. tabacum Tế bào P. cryptogea (thô) Capsidiol 0 25 µg/mL
Panax ginseng Rễ tơ Selenium Ginseng saponin Đối chứng Tăng 1,33 lần
Papaver bracteatum Tế bào Dendryphion (thô) Sanguinarine 50 µg/g dw 450 µg/g fw
Sanguinaria canadensis Tế bào Verticillium dahliae (tế bào) Dopamine 3 mg/g fw 15 mg/g fw
S. canadensis Tế bào V. dahliae (thô) Sanguinarine 3 µg/g fw 12 µg/g fw
Tagetes patula Rễ tơ F. conglutanis (thô) Thiophene 0,2 g/100g dw 0,55 g/100 g
dw
T. patula Rễ tơ A. niger (thô) Thiophene 1,5 µM/g dw 3,5 µM/g dw
Taxus chinensis Tế bào Trifluoroethyl salicylate Taxuyunnanine C 14 mg/g dw 21,9 mg/g dw
Thalictrum rugosum Tế bào S. cerevisiae (thô) Berberine 0,5% dw 2% dw

23
Ảnh hương của các oligosaccharide DP4, DP7 và DP10 tách chiết từ
chủng nấm F. oxysporum Dzf17 lên khả năng tích lũy diosgenin ơ D.
zingiberensis đã được Li và cs (2011) nghiên cứu. Tế bào được xử lý với hỗn
hợp oligosaccharide nói trên ơ nồng độ 20 mg/L sau 26 ngày nuôi và thu sinh
khối vào ngày thứ 32. Hàm lượng diosgenin đạt được cao nhất là 2,187 mg/L,
gấp 5,65 lần đối chứng. Xử lý từng oligosaccharide riêng lẻ từ 2-10 mg/L cho
thấy DP7 ơ 6 mg/L cho hiệu quả cao nhất, lượng diosgenin đạt 3,202 mg/L,
gấp 8,27 lần đối chứng. Khi xử lý DP7 hai lần vào ngày 24 và 26 và thu vào
ngày 30, lượng diosgenin còn tăng lên cao hơn nữa đạt 4,843 mg/L, gấp 12,38
lần đối chứng [77].
Bota và cs (2012) dùng dịch chiết của hai chủng nấm Botrytis và
Sclerotinia để tăng hàm lượng flavonoid ơ Digitalis lanata. Bổ sung 1-2 mL
dịch chiết nấm vào 100 mL dịch tế bào của dòng số 11 và 13 đã cải thiện sự
tích lũy flavonoid theo thời gian và nồng độ xử lý. Hàm lượng flavonoid thu
được sau 96 giờ nuôi dòng số 11 đạt 1.000 mg% (dịch chiết Botrytis) và 999,81
mg% (dịch chiết Sclerotinia), trong khi dòng số 13 đạt 1051,65 mg% (Botrytis)
và 1025,43 mg% (Sclerotinia) [27].
Khi bổ sung dịch chiết của A. niger, F. oxysporum và YE vào môi trường
nuôi cấy Hypericum triquetrifolium ơ nồng độ từ 0,1-0,75 mg/L, Azeez và cs
(2013) nhận thấy các elicitor này có vai trò khác nhau trong việc sản xuất các
hợp chất thứ cấp. Dùng YE 0,5 mg/L đã tăng hàm lượng p-OH-benzoic acid và
chlorgenic acid lên đáng kể so với đối chứng. Sinh tổng hợp caffeic acid và
tannic acid giảm ơ tất cả công thức xử lý nhưng catechin lại tăng khi bổ sung
dịch chiết A. niger. Hypersoid, quercitin và rutin cũng tăng lên nhiều trong các
công thức thử nghiệm [20].
Nuôi cấy rễ tơ cây rau sam (Portulaca oleracea) cũng đã được Pirian và
cs (2013) thực hiện. YE ơ các nồng độ 125, 250, 500 và 1.000 mg/L được sử
dụng để cải thiện khả năng tích lũy noradrenaline với thời gian xử lý 2 ngày.

24
Các tác giả nhận thấy YE ơ nồng độ 250 và 500 mg/L là thích hợp nhất, hàm
lượng noradrenaline đã tăng 3-4 lần so với đối chứng [117].
Admeh và cs (2014) đã dùng dịch chiết của A. niger, P. notatum, YE và
chitosan để kích thích sản xuất psoralen trong nuôi cấy tế bào cây Psoralea
corylifolia. Bổ sung dịch chiết A. niger đã tăng lượng psoralen lên 9 lần so với
đối chứng. Trong khi dùng P. notatum, YE và chitosan lượng psoralen thu được
ít hơn 4-7 lần. Nói chung, dịch chiết A. niger 1% cho hiệu quả cao nhất, hàm
lượng psoralen đạt 9.850 μg/g sinh khối khô [12].
Nghiên cứu của Hasanloo (2014) cho thấy, chitosan có khả năng kích thích
tổng hợp silymarin trong nuôi cấy rễ tơ của loài Silybum marianum. Rễ tơ sau
30 ngày nuôi cấy được xử lý với chitosan có khối lượng phân tử trung bình.
Hàm lượng silymarin tổng số tăng lên 5,26 lần và khối lượng khô của rễ tơ đạt
cao nhất là 0,535 g sau 96 giờ xử lý với 30 mg/50 mL chitosan [57].
Hiệu quả của các elicitor sinh học cũng được Ebrahimi (2015) thử nghiệm
trong nuôi cấy tế bào Peganum harmala để tăng sinh tổng hợp hai loại alkaloid
β-carboline là harmaline và harmine. Nuôi cấy đã được xử lý với dịch chiết của
các loại nấm (Alternaria alternate, A. flavus, Coriolus versicolor, F.
oxysporum, Mucor sp., P. notatum và Rhizopus stonifer), dịch thủy phân casein
và YE ơ các nồng độ khác nhau. Lượng harmine thu được cao nhất khi bổ sung
YE 1.000 mg/L là 91,2 μg/g dw, gấp 1,68 lần đối chứng. Khi bổ sung dịch thủy
phân casein từ 75-100 mg/L vào nuôi cấy, sinh khối tế bào đã tăng mạnh, lượng
harmaline và harmine cũng tăng lần lượt gấp 1,61 và 1,46 lần đối chứng [43].
Silene vulgaris là một loài thực vật được phân bố rộng rãi ơ Bắc Mỹ chứa
các saponin loại oleanane có hoạt tính sinh học. Khả năng sản xuất saponin
bằng nuôi cấy rễ tơ của loài này đã được Kim và cs (2015) nghiên cứu thành
công. Rễ tơ của S. vulgaris được hình thành bằng cách lây nhiễm các mẫu lá
với năm chủng Agrobacterium rhizogenes khác nhau (LBA9402, R1000, A4,
13333 và 15834). Khi bổ sung MeJA vào môi trường nuôi cấy, khả năng tích

25
lũy saponin triterpenoid trong rễ tơ có sự thay đổi đáng kể. Lượng segetalic
acid và gypsogenic acid cao gấp 5 và 2 lần tương ứng so với đối chứng [71].
Arican (2016) nghiên cứu ảnh hương của các loại elicitor sinh học đến khả
năng tổng hợp triterpene trong nuôi cấy tế bào Alstonia scholaris. Nuôi cấy
được xử lý dịch chiết các loại nấm Candida albicans, F. oxysporum, P.
avelanium và YE. Các elicitor gây ra sự kích thích nhanh chóng quá trình trao
đổi chất thứ cấp của các tế bào A. scholaris dẫn đến tăng tổng hợp triterpenoid.
Khi tiếp xúc trong thời gian dài, một số dòng tế bào có khả năng tăng gấp đôi
sản lượng của triterpene. YE là elicitor tốt nhất cho tất cả các dòng tế bào được
khảo sát với lượng ursolic acid và oleanolic acid tích được tương ứng là 5 và 7
mg/g dw [18].
Hyoscyamine và scopolamine là hai loại alkaloid tropane rất có giá trị,
thường được sử dụng để làm chất chống đông máu, chống co thắt, chống dị
ứng, giảm đau và làm thuốc an thần. Các chất này được phát hiện có nhiều
trong loài Hyoscyamus reticulatus. Để tìm phương pháp sản xuất hyoscyamine
và scopolamine tốt nhất, Moharrami (2017) đã thử nghiệm nuôi cấy rễ tơ của
cây nảy mầm từ hạt được biến nạp chủng A. rhizogenes A7 có xử lý elicitor
sinh học ơ các nồng độ và thời điểm khác nhau. Kết quả thu được cho thấy, YE
ơ 500 và 250 mg/L sau 48 giờ xử lý là tốt nhất, hàm lượng hyoscyamine và
scopolamine đã tăng gấp 2 và 2,5 lần tương ứng so với đối chứng [103].
Năm 2018, Lee và cs đã nghiên cứu phương thức tăng khả năng tổng hợp
saponin trong nuôi cấy tế bào Kalopanax septemlobus bằng cách sử dụng
elicitor. Khi không bổ sung elicitor, hàm lượng saponin tổng số thu được là
1,56 mg/60 mL sau 15 ngày nuôi cấy. Khi bổ sung coronatine 1 μM, lượng
saponin được tích lũy tăng lên 160%. Ngoài ra, các tác giả nhận thấy coronatine
còn làm tăng biểu hiện sinh tổng hợp β-amyrin từ đó dẫn đến tích lũy oleanolic
acid (tiền thân của oleanan-một loại saponin triterpene) [75].

26
1.3.2.2. Elicitor phi sinh học
Các elicitor phi sinh học thường được dùng phổ biến hơn elicitor sinh học,
nhiều công trình nghiên cứu đã được tiến hành theo hướng này. Rezaei và cs
(2011) đã nghiên cứu sự tích lũy taxol trong nuôi cấy tế bào của cây thủy tùng
(Taxus baccata) bằng cách bổ sung SA từ 25-50 mg/L kết hợp với sóng siêu
âm có tần số 40 kHz trong 2 phút. Hàm lượng taxol thu được ơ môi trường có
SA 50 mg/L cao gấp 8 lần đối chứng [126].
MeJA đã được Wang và cs (2015) sử dụng để tăng sản xuất flavonoid
trong nuôi cấy tế bào Hypericum perforatum. Khi bổ sung MeJA 100 μmol/L
vào môi trường trong 15 ngày đã thu được lượng flavonoid cao nhất là 280
mg/L, gấp 2,7 lần đối chứng. Nguyên nhân có thể do MeJA ức chế hoạt tính
catalase nhưng lại tăng hoạt tính phenylalanine ammonia lyase (PAL), những
enzyme liên quan đến quá trình sinh tổng hợp flavonoid, từ đó tăng tổng hợp
chất này trong quá trình nuôi cấy [159].
Năm 2016, Khojasteh và cs đã sản xuất thành công rosmarinic acid bằng
nuôi cấy tế bào Satureja khuzistanica trong nồi phản ứng sinh học (bioreactor).
Hai elicitor là MeJA (100 μM) và cyclodextrin (40 mM) đã được thử nghiệm
riêng rẻ hoặc kết hợp. Kết quả cho thấy chỉ có MeJA khi dùng riêng rẻ đã tăng
tích lũy rosmarinic acid gấp 3 lần đối chứng, đạt 3,9 g/L. Khi chuyển sang nuôi
cấy trong các bioreactor, lượng rosmarinic acid tích lũy tối đa đạt 3,1 g/L [69].
Manivannan và cs (2016) nghiên cứu ảnh hương của MeJA, SA và SNP
lên khả năng sinh tổng hợp các chất chống oxy hóa và các chất chuyển hóa thứ
cấp trong nuôi cấy tế bào Scrophularia kakudensis. Tất cả các elicitor đều có
ảnh hương tích cực đến khả năng tích lũy các chất này trong tế bào. Trong đó,
MeJA từ 150-200 μM cho hiệu quả tổng hợp các chất nhóm phenol và flavonoid
cao nhất [91].
Saeed (2017) khi nuôi cấy tế bào của rễ cây Ajuga bracteosa đã bổ sung
thêm MeJA và PAA. Kết quả cho thấy sinh khối tế bào đã tăng đáng kể, đạt tối

27
đa 8,88 g/L dw ơ ngày thứ 32 khi bổ sung MeJA 0,6 mg/L, và 8,24 g/L dw ơ
ngày thứ 40 khi bổ sung PAA 1,2 mg/L. Ngoài ra, khả năng tích lũy các hợp
chất phenol, flavonoid và hoạt tính chống oxy hóa cũng tăng lên nhiều lần khi
sử dụng hai loại elicitor nói trên [131].
Estrada-Soto và cs (2018) đã nghiên cứu ảnh hương của MeJA và SA
trong nuôi cấy callus lá của Leptochinia caulescens để sản xuất ursolic và
oleanolic acid. Nhìn chung, cả hai loại elicitor nói trên đều có ảnh hương tích
cực lên quá trình sản xuất ursolic và oleanolic acid. Bổ sung MeJA trong 8 giờ
đã cho kết quả tốt nhất, hàm lượng các triterpen tăng gấp 5 lần đối chứng [45].
Bên cạnh việc sử dụng riêng lẻ các loại elicitor sinh học và phi sinh học,
rất nhiều công trình nghiên cứu đã sử dụng kết hợp các elicitor để mang lại hiệu
quả cao hơn. Artemisinin là chất thường được sử dụng để sản xuất thuốc chống
sốt rét ơ các nước châu Á và châu Phi. Ahlawat và cs (2014) đã thiết lập nuôi
cấy rễ tơ cây Artemisia annua để sản xuất bằng cách bổ sung MeJA, dịch chiết
nấm (A. alternate, Curvularia limata, F. solani và Piriformospora indica),
farnesyl pyrophosphate và miconazole vào môi trường nuôi cấy. Kết quả xử lý
riêng rẽ các elicitor cho thấy hiệu quả cao nhất thu được khi sử dụng dịch chiết
P. indica, hàm lượng artemisinin cao gấp 1,97 lần đối chứng. Tuy nhiên, khi
kết hợp xử lý MeJA và P. indica, hàm lượng artemisinin đã tăng lên cao hơn,
đạt 2,44 lần [11].
1.4. CÁC GEN THAM GIA TỔNG HỢP CURCUMINOID
1.4.1. Các con đường sinh tổng hợp curcuminoid
Năm 1973, Roughly và Whiting đưa ra hai con đường sinh tổng hợp
curcumin ơ thực vật dựa trên các dữ liệu phân tích 14C. Ở con đường thứ nhất,
cinnamic acid kết hợp với malonyl-CoA (có nguồn gốc từ malonic acid) trong
một chuỗi phản ứng mà cuối cùng được aryl hóa thành một curcuminoid. Trong
con đường thứ hai, hai đơn vị muối cinnamate được kết hợp với nhau bằng

28
malonyl-CoA. Cả hai cơ chế cùng sử dụng cinnamic acid có nguồn gốc từ
phenylalanine như điểm khơi đầu của chúng [72], [130].
Bằng cách sử dụng những chất đánh dấu đồng vị phóng xạ, một số nghiên
cứu cho thấy các hợp chất nhóm curcuminoid có nguồn gốc từ các chất trung
gian trong con đường phenylpropanoid kết hợp với các phân tử khác từ con
đường tổng hợp acetate hay chuỗi ngắn và vừa của acid béo. Dựa trên kết quả
này, Schröder (1997) cho rằng các enzyme tương tự như polyketide synthase
đóng vai trò quan trọng trong việc tổng hợp các cấu trúc khung của curcuminoid
và sử dụng các CoA có nguồn gốc từ các hợp chất trung gian. Từ đó, hai con
đường sinh tổng hợp curcuminoid đã được đưa ra. Curcuminoid có thể được
hình thành từ sự kết hợp của hai phân tử p-coumaroyl-CoA với một phân tử
malonyl-CoA nhờ hoạt động của polyketide synthase (hoặc tương tự), có thể
do bổ sung diketide trung gian (Berbd và Schneider 2006). Kết quả,
bisdemethoxycurcumin được chuyển thành demethoxycurcumin, rồi thành
curcumin theo hai vòng liên tục của sự hydro hóa và methyl hóa. Mặt khác, có
khả năng enzyme tổng hợp curcuminoid sử dụng các dạng ester CoA của 2
phân tử p-coumaric acid và ferulic acid làm cơ chất. Trong tường hợp này, sự
hydro hóa và methyl hóa dẫn tới sự hình thành các nhóm chức methoxyl trong
phân tử curcumin tương tự như các phản ứng phát hiện trong con đường
phenylpropanoid (Hình 1.2) [124].
Để tìm hiểu quá trình sinh tổng hợp curcuminoid ơ cây nghệ vàng (C.
longa), Kita và cs (2008) đã thiết lập hệ thống nuôi cấy in vitro cho loài dược
liệu này với các tiền chất bổ sung vào môi trường chứa 13C. Kết quả phân tích
hàm lượng demethoxycurcumin và các dạng curcuminoid khác bằng phân tích
cộng hương từ hạt nhân 13C cho thấy một phân tử acetic acid hoặc malonic acid
và hai phân tử phenylalanine hoặc phenylpropanoid (không phải tyrosine) kết
hợp tạo thành demethoxycurcumin. Sự kết hợp với các cơ chất giống nhau để
tạo thành demethoxycurcumin hay curcumin là tương tự nhau, theo thứ tự

29
malonic acid > acetic acid, và cinnamic acid > p-coumaric acid > ferulic acid.
Kết quả này cho thấy có thể hai phân tử cinnamoyl-CoA và một phân tử
malonyl-CoA đã tạo nên curcuminoid, các gốc hydroxy và methoxy trên vòng
thơm được gắn vào sau khi hình thành cấu trúc khung của curcuminoid [72].
Xie và cs (2009) đã nghiên cứu các cơ chế điều hòa biểu hiện gen tổng
hợp các hợp chất thứ cấp ơ cây nghệ vàng và nhận thấy có một cơ chế điều hòa
sinh tổng hợp ba dạng curcuminoid. Sự tồn tại của cơ chế điều hòa này đã chứng
minh cho giả thuyết rằng nhóm 3-methoxyl trên vòng thơm của phân tử
curcuminoid được hình thành trước sự hình thành của khung heptanoid trong
quá trình sinh tổng hợp curcumin cũng như sự tồn tại của nhiều gen mã hóa
enzyme thuộc nhóm polyketide synthases với nhiều cơ chất khác nhau, hình
thành nhiều diarylheptanoid trong cây nghệ vàng [163].

30
Hình 1.2. Con đường sinh tổng hợp các curcuminoid trong cây nghệ vàng. PAL: phenylalanine ammonia lyase. C4H: cinnamate 4-
hydroxylase. 4CL: 4-coumarate-CoA ligase. CST: p-coumaroyl shikimate transferase. CS3’H: p-coumaroyl 5-O-shikimate 3’-
hydroxylase. CCOMT: caffeoyl-CoA O-methyltransferase [124].

31
1.4.2. Vai trò của các gen tham gia chu trình tổng hợp curcuminoid
Tham gia vào chu trình sinh tổng hợp curcuminoid ơ nghệ vàng có rất
nhiều enzyme khác nhau xúc tác cho nhiều phản ứng trung gian khác nhau,
phần lớn các gen/enzyme này đều đã được nghiên cứu cả về đặc điểm, chức
năng cũng như biểu hiện tái tổ hợp. Trong số đó, các enzyme thuộc nhóm
polyketide synthase type III đóng vai trò hết sức quan trọng. Đây là họ enzyme
thiết yếu trong quá trình sinh tổng hợp flavonoid và các hợp chất polyphenol ơ
thực vật bằng cách gắn các nhóm acetyl từ malonyl-CoA lên vị trí liên kết
thioester trên trung tâm hoạt động của polyketide synthase (Hình 1.3) [125].
1.4.2.1. Gen mã hóa enzyme diketide-CoA synthase (DCS)
Gen DCS mã hóa diketide-CoA synthase là một enzyme thuộc nhóm
polyketide synthase type III xúc tác tạo thành feruloyldiketide-CoA từ feruloyl-
CoA và malonyl-CoA (Hình 1.3) [66], [67].
1.4.2.2. Gen mã hóa enzyme curcumin synthase (CURS)
Gen CURS mã hóa curcumin synthase là enzyme xúc tác tạo thành
curcuminoid từ cinnamoyldiketide-N-acetylcysteamine và feruloyl-CoA. Hoạt
động đồng thời của DCS và CURS khi có sự hiện diện của feruloyl-CoA và
malonyl-CoA sẽ tạo ra nhiều curcumin, trong khi một mình CURS cho hiệu quả
tổng hợp curcumin thấp cùng với sự hiện diện của feruloyl-CoA và malonyl-
CoA. Như vậy, DCS tổng hợp feruloyldiketide-CoA trong khi CURS xúc tác
chuyển hóa diketide-CoA thành curcuminoid [66].
Hiện nay, các nhà khoa học đã xác định được có ba gen CURS mã hóa ba
enzyme CURS với các vai trò khác nhau. CURS1 xúc tác hình thành curcumin
từ feruloyl-CoA và feruloyldiketide-CoA (được tạo ra bơi hoạt động của gen
DCS). Hai enzyme DCS và CURS1 cùng tham gia xúc tác tạo thành curcumin,
cả hai gen đều sử dụng p-coumaroyl-CoA nhưng hiệu quả thấp, chúng cũng có

32
khả năng sinh tổng hợp demethoxycurcumin từ feruloyl-CoA và malonyl-CoA
theo con đường tạo thành p-coumaroyldiketide-CoA. CURS2 tương tự CURS1
về tính đặc hiệu cơ chất với feruloyl-CoA là cơ chất khơi đầu. CURS3 cũng có
tính đặc hiệu cơ chất như CURS1 và CURS2, sử dụng cả p-coumaroyl-CoA và
feruloyl-CoA làm cơ chất khơi đầu. Giống như CURS1, CURS2 chủ yếu xúc
tác quá trình tạo thành curcumin và demethoxycurcumin bằng cách gắn
feruloyl-CoA với p-coumaroyldiketide-CoA hoặc feruloyldiketide-CoA. Trong
khi đó, CURS3 có khả năng tổng hợp cả 3 loại curcuminoid bằng cách gắn
feruloyl-CoA hoặc p-coumaroyl-CoA với p-coumaroyldiketide-CoA hoặc
feruloyldiketide-CoA. Sự tồn tại của ba gen CURS cho thấy các dạng
curcuminoid được tổng hợp trong củ nghệ không chỉ phụ thuộc vào cơ chất như
p-coumaroyl-CoA và feruloyl-CoA mà còn phụ thuộc vào mức độ biểu hiện
của các gen này (Hình 1.3) [67].
Hình 1.3. Vai trò của các gen DCS và CURS trong tổng hợp curcuminoid ơ nghệ
vàng [67].
1.4.2.3. Gen mã hóa enzyme Curcuminoid synthase

33
Bên cạnh hệ thống enzyme sinh tổng hợp curcuminoid là DCS/CURS
trong củ nghệ vàng, hiện nay các nhà khoa học đã phát hiện thêm một enzyme
khác thuộc nhóm polyketide synthase type III là curcuminoid synthase (CUS)
ơ cây lúa (Oryza sativa) cũng có chức năng tương tự. Enzyme này xúc tác quá
trình tổng hợp curcuminoid như sau: đầu tiên, p-coumaroyl-CoA và malonyl-
CoA kết hợp tạo thành sản phẩm trung gian diketide-CoA. Sau đó, sự kết hợp
giữa diketide-CoA và phân tử p-coumaroyl-CoA khác tạo nên
bisdemethoxycurcumin. Bản thân enzyme CUS xúc tác cho cả 2 bước được
thực hiện riêng rẽ bơi DCS và CURS, vì sử dụng enzyme CUS sẽ đơn giản hơn
sử dụng hệ thống DCS/CURS nên có nhiều ưu thế hơn khi xây dựng hệ thống
tổng hợp curcuminoid trong vi sinh vật [68].
Mặc dù đều có khả năng tổng hợp curcuminoid nhưng hoạt động của hai
hệ enzyme CUS và DCS/CURS có một số điểm khác nhau. CUS tổng hợp
curcuminoid kèm theo sự hình thành của triketide pyrone. Trong khi đó, hệ
enzyme DCS/CURS chỉ tạo thành một lượng rất ít dehydrozingerone là sản
phẩm phụ (dạng vết), có nguồn gốc từ sự thủy phân không có sự tham gia của
enzyme (nonenzymatic hydrolysis) và sự decarboxyl hóa feruloyldiketide-
CoA. Do đó, hệ enzyme DCS/CURS chỉ tổng hợp curcumin và các chất trung
gian là feruloyldiketide-CoA. Hệ thống tổng hợp curcumin trong nghệ vàng
loại bỏ sản phẩm phụ triketide pyrone bằng cách chia quá trình tổng hợp thành
hai phần: phần sử dụng DCS và phần sử dụng CURS. Cơ chất để bắt đầu quá
trình tổng hợp của hai hệ enzyme cũng khác nhau, CUS sử dụng p-coumaroyl-
CoA trong khi cả DCS và CURS sử dụng feruloyl-CoA làm cơ chất khơi đầu
[66].
1.4.2.4. Gen mã hóa enzyme Chalcone synthase
Chalcone synthase (CHS) là một enzyme thuộc nhóm polyketide synthase
type III, xúc tác cho sự hình thành các hợp chất thứ cấp chính của thực vật như

34
acridone, bibenzyl, biphenyl, benzophenone, chalcone, chromone,
curcuminoid, phloroglucinol, pyrone, resorcinol và stilbene. Các phản ứng
được xúc tác bơi CHS được bắt đầu bằng cách chuyển nhóm acyl từ p-
coumaroyl-CoA lên trung tâm xúc tác cysteine của CHS. Sau đó, kết hợp với 3
phân tử malonyl-CoA đã bị decarboxyl để tạo nên chất trung gian tetraketide.
Phản ứng này được xúc tác bơi trung tâm hoạt động gồm Cys-Asn-His.
Tetraketide sau đó được đóng vòng thơm để tạo thành chalcone [66]. Gần đây,
các nhà khoa học đã tìm thấy gen CHSI (chalcone synthase like gene) và gen
CHSII ơ cây nghệ C. caesia, các gen này đã được tạo dòng, phân tích chức năng
và đăng ký ơ ngân hàng gen [23]. Trong quá trình sinh tổng hợp curcuminoid,
hoạt động mạnh của gen CHS sẽ làm giảm lượng curcuminoid tạo thành do sử
dụng các hợp chất trung gian là diketide-CoA và malonyl-CoA để tổng hợp nên
chalcone hay triketide pyrone [66].
Resmi và Soniya (2012) đã tìm ra hai gen mới mã hóa các enzyme thuộc
nhóm polyketide synthase type III là ClPKS9 và ClPKS10 từ nghệ vàng. Gen
ClPKS9 có trình tự tương đồng với gen CHS trong khi gen ClPKS10 tương
đồng với gen CUS. Hai gen này có sự biểu hiện khác nhau ơ các mô khác nhau,
ClPKS9 biểu hiện mạnh ơ chồi và củ hơn ơ lá, trong khi đó gen ClPKS10 biểu
hiện mạnh trong lá nhưng rất ít trong rễ và củ [125].
1.4.2.5. Các gen khác
Bên cạnh các gen đã kể trên, con đường phenylpropanoid tổng hợp nhóm
chất curcuminoid còn có sự tham gia của một số gen khác (Hình 1.2).
Gen PAL mã hóa phenylalanine ammonia lyase, enzyme tham gia phản
ứng chuyển hóa đầu tiên trong chu trình phenylpropanoid, khơi đầu của quá
trình sinh tổng hợp curcuminoid. Enzyme này xúc tác phản ứng loại gốc amine
không oxy hóa của L-Phe tạo thành trans-cinnamic acid và NH4+ [124].
Gen C4H mã hóa cinnamate 4-hydroxylase là monooxygenase phụ thuộc

35
Cyt P450 đầu tiên, tham gia ơ bước thứ 2 trong con đường phenylpropanoid.
Enzyme này xúc tác chuyển cinnamic acid thành p-coumaric acid. Hiện nay,
có hơn 20 gen C4H đã được phân lập [138].
Gen 4CL mã hóa 4-coumarate-CoA ligase, enzyme xúc tác phản ứng
chuyển hydroxycinnamic acid (p-coumaric acid, caffeic acid và ferulic acid)
sang dạng CoA esters (p-coumaroyl-CoA, caffeoyl-CoA và feruloyl-CoA)
[128].
Gen CST mã hóa p-coumaroyl shikimate transferase, enzyme tham gia
vào quá trình sản xuất methoxylate phenylpropanoid trong thực vật. Trong các
mô sản xuất curcuminoid, nếu thiếu enzyme này hoặc enzyme này ít hoạt động
thì curcumin sẽ được tổng hợp thông qua bisdemethoxycurcumin, trong đó tất
cả các chất trung gian trong con đường phenylpropanoid đều có nguồn gốc từ
p-coumaroyl-CoA chứ không phải từ feruloyl-CoA. Tuy nhiên, một số nghiên
cứu khác cho rằng enzyme này tham gia vào quá trình tạo ra khung cấu trúc
của curcuminoid và sử dụng cả p-coumaroyl-CoA và feruloyl-CoA làm cơ chất
[124].
1.4.3. Tổng hợp curcuminoid theo phương thức tái tổ hợp
Với sự hiểu biết về các gen cũng như vai trò của chúng trong con đường
sinh tổng hợp curcuminoid ơ thực vật nói chung và nghệ vàng nói riêng,
Katsuyama và cs (2008) đã tạo ra hệ thống tổng hợp curcuminoid trong tế bào
E. coli. Trong hệ thống này, gen PAL có nguồn gốc từ nấm men Rhodotorula
rubra, gen 4CL từ cây L. erythrorhizon, gen ACC (acetyl-CoA carboxylase)
từ vi khuẩn Corynebacterium glutamicum và gen CUS từ lúa. Tế bào E. coli
tái tổ hợp được nuôi trong môi trường có tyrosine hoặc phenylalanine, hoặc
cả hai để tổng hợp nên bisdemethoxycurcumin, dicinnamoylmethane và
cinnamoyl-p-coumaroylmethane. Một hệ thống khác chứa hai gen 4CL và
CUS đã tổng hợp được lượng lớn curcuminoid từ các thành phần

36
phenylpropanoid acid bổ sung từ bên ngoài như p-coumaric acid, cinnamic
acid và ferulic acid. Hàm lượng curcuminoid thu được khoảng 100 mg/L
[68], [65].
Rogrigues và cs (2015) đã xây dựng hệ thống tổng hợp curcuminoid nhân
tạo trong E. coli thông qua caffeic acid. Các tác giả đã sử dụng gen tổng hợp
4CL từ Arabidopsis thaliana, gen DCS và CURS1 từ nghệ vàng và đã thu được
70 mg/L curcumin với cơ chất là ferulic acid. Bisdemethoxycurcumin và
demethoxycurcumin cũng được tạo ra nhưng với hàm lượng thấp khi sử dụng
p-coumaric acid hoặc hỗn hợp p-coumaric acid và ferulic acid. Curcuminoid
cũng được tổng hợp từ tyrosine thông qua con đường caffeic acid. Để tạo ra
caffeic acid, gen TAL mã hóa hóa tyrosine ammonia lyase từ Rhodotorula
glutinis, gen C3H mã hóa 4-coumarate 3-hydroxylase từ Saccharothrix
espanaensis đã được sử dụng. Gen COMT mã hóa catechol-O-
methyltransferase từ cỏ linh lăng (Medicago sativa) được sử dụng để chuyển
caffeoyl-CoA thành feruloyl-CoA. Sử dụng caffeic acid, p-coumaric acid hoặc
tyrosine làm cơ chất, các tác giả đã thu được hàm lượng curcumin tương ứng
lần lượt là 3,9, 0,3, và 0,2 mg/L. Đây là nghiên cứu đầu tiên sử dụng gen DCS
và CURS1 để tổng hợp curcuminoid trong điều kiện in vitro và curcumin được
sản xuất từ tyrosine [128].
Wang và cs (2015) đã sử dụng 7 enzyme khác nhau từ thực vật và vi khuẩn
để tổng hợp nên các hợp chất có giá trị trong E. coli như 4 loại phenylpropanoid
acid (cinnamic acid, p-coumaric acid, caffeic acid và ferulic acid), 3 chất thuộc
nhóm stilbenoid (resveratrol, piceatannol và pinosylvin), và 3 chất thuộc nhóm
curcuminoid (curcumin, bisdemethoxycurcumin và dicinnamoylmethane).
Trong đó, các gen PAL có nguồn gốc từ Trifolium pretense, 4CL từ A. thaliana,
CUS từ lúa, TAL và C3H từ Saccharothrix espanaensis, COMT từ cỏ linh lăng
và STS mã hóa stilbene synthase từ cây lạc (Arachis hypogaea). Ban đầu, các
tác giả đã biểu hiện các gen PAL, 4CL và CUS trong E. coli để tổng hợp

37
dicinnamoylmethane. Sau đó, họ sử dụng TAL thay cho PAL, biểu hiện đồng
thời TAL, 4CL và CUS trong E. coli đã tạo ra sản phẩm là
bisdemethoxycurcumin (28,9 mg/L). Các tác giả này cũng nhận thấy có thể
tổng hợp curcumin từ L-tyrosine khi biểu hiện đồng thời 4CL và CUS trong E.
coli [160].
1.4.4. Cải thiện sự biểu hiện gen bằng xử lý elicitor
Những nghiên cứu theo hướng này còn chưa nhiều. Park và cs (2016) đã
xử lý YE và AgNO3 để tăng cường mức độ biểu hiện của các gen trong chu
trình phenylpropanoid và sự tích lũy của rosmarinic acid khi nuôi cấy tế bào
cây Agastache rugosa. Kết quả cho thấy bổ sung YE 0,5 g/L và AgNO3 30
mg/L ơ các thời điểm khác nhau đều kích hoạt sự biểu hiện của các các gen và
sự tích lũy của rosmarinic acid. Mức độ biểu hiện của các gen RAS mã hóa
rosmarinic acid synthase và HPPR mã hóa hydroxyl phenylpyruvate
reductase tăng lên cao nhất là 1,84, 1,97 và 2,86 lần khi được xử lý YE trong
các thời gian khác nhau 3, 6 và 12 giờ. Đối với gen PAL, mức độ phiên mã tăng
lên tới 52,31 lần khi xử lý bằng AgNO3 sau 24 giờ. Xử lý YE cũng làm tăng
mức độ tích lũy rosmarinic acid lên 4,98 mg/g, trong khi xử lý AgNO3
rosmarinic acid chỉ đạt 0,65 mg/g. Như vậy, có sự tương đồng cao trong việc
tăng mức độ biểu hiện của các gen trong chu trình phenylpropanoid và sự tích
lũy của rosmarinic acid khi được xử lý bằng YE [115].
Loc và cs (2016) đã nghiên cứu tăng cường mức độ biểu hiện của các gen
CaSQS, CabAS và CaCYS trong con đường chuyển hóa phytosterol và
triterpene ơ cây rau má (Centella asiatica (L.) Urban) được xử lý bơi salicilic
acid (50-200 µM). Phân tích PR phiên mã ngược (RT-PCR) và Northern blot
cho thấy các gen CaSQS, CabAS và CaCYS biểu hiện ơ cả trong cây tự nhiên
và trong tế bào nuôi cấy (có và không có xử lý elicitor). Trong tế bào được xử
lý elicitor, mức độ biểu hiện của các gen CaSQS, CabAS và CaCYS phụ thuộc
rất lớn vào nồng độ và thời gian xử lý salicilic acid. Mức độ biểu hiện cao nhất

38
được quan sát thấy khi xử lý salicilic acid 100 µM vào ngày thứ 10 của quá
trình nuôi cấy. Salicilic acid nồng độ từ 50-200 µM làm giảm mức độ biểu hiện
của các gen CaCYS và CaSQS trong tế bào được xử lý elicitor so với đối chứng
không xử lý elicitor [88].
Sarkate và cs (2007) đã sử dụng YE để tăng cường sản xuất các hợp chất
thứ cấp nhóm phenol và chất chống oxy hóa từ tế bào cây táo (Malus domestica
‘florina’). Ở các công thức xử lý YE, hàm lượng các hợp chất nhóm phenol và
chất chống oxy hóa đều tăng lên đáng kể, thông qua sự tăng cường hoạt động
của enzyme PAL. Dịch chiết methanol được phân tích HPLC cho thấy các hợp
chất phenol là chlorogenic acid (112 µg/g), 4-coumaric acid (122 µg/g), ferulic
acid (212 µg/g), benzoic acid (244 µg/g) và rutin (348 µg/g) đều tăng lên sau
khi tế bào được xử lý YE. Bên cạnh đó, hoạt tính của các chất chống oxy hóa
như DPPH và FRAP cũng đều tăng lên khi được xử lý YE, tương ứng với sự
tăng tích lũy các hợp chất phenol [136]./.

39
Chương 2
ĐỐI TƯỢNG VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. ĐỐI TƯỢNG NGHIÊN CỨU
Đối tượng nghiên cứu là sự biểu hiện của các gen tham gia vào quá trình
sinh tổng hợp curcuminoid trong tế bào cây nghệ đen (Curcuma zedoaria
Roscoe). Cây nghệ đen có nguồn gốc từ tỉnh Đắk Lắk, được trồng tại thành phố
Huế trước khi được sử dụng làm nguyên liệu nghiên cứu (Hình 2.1).
Hình 2.1. Cây nghệ đen.
2.2. PHƯƠNG PHÁP NGHIÊN CỨU
Quá trình nghiên cứu được thực hiện theo sơ đồ thí nghiệm ơ hình 2.2 và
sử dụng các phương pháp sau:

40
2.2.1. Nuôi cấy in vitro cây nghệ đen
2.2.1.1. Khử trùng mẫu vật
Chồi mầm tách từ củ nghệ được rửa sạch bằng nước xà phòng loãng và
nước máy nhiều lần trước khi khử trùng. Chồi được ngâm 30 giây trong cồn
70% và sau đó 20 phút trong dung dịch AgNO3 1% [101]. Sau cùng rửa lại
bằng nước cất vô trùng 5 lần trước khi cấy lên môi trường dinh dưỡng thích
hợp.
Cây nghệ đen
Chồi mầm của củ nghệ Phân lập các gen tổng hợp
curcuminoid
Tái sinh chồi in vitro
Nhân cụm chồi
Tạo rễ
Nhân giống cây in vitro
Củ nghệ
Nuôi cấy callus Phân tích biểu hiện
(RT-PCR và HPLC)
Nuôi cấy tế bào Xử lý elicitor Phân tích biểu hiện
(RT-PCR và HPLC)
Hình 2.2. Sơ đồ thí nghiệm.

41
2.2.1.2. Tái sinh chồi và tạo rễ in vitro
Chồi mầm sau khi khử trùng (1-1,5 cm) được cấy lên môi trường cơ bản
MS có sucrose 20 g/L, agar 8 g/L, bổ sung BAP 3 mg/L, IBA 0,5 mg/L và nước
dừa 20% (v/v) để tái sinh cụm chồi [79]. Chồi in vitro tái sinh (khoảng 2 cm)
nguyên vẹn hoặc chẻ đôi được chuyển lên môi trường tương tự như trên nhưng
thêm AgNO3 từ 0,5-2,5 mg/L để thăm dò khả năng nhân cụm chồi. Chồi chẻ
đôi là các chồi mang đỉnh sinh trương được chẻ đôi theo chiều dọc bằng giao
cấy, trong đó đỉnh sinh trương cũng được chẻ đôi. Các chồi in vitro (3-4 cm)
tách từ cụm chồi trên môi trường nhân được nuôi trên môi trường MS có
sucrose 20 g/L, agar 8 g/L, nước dừa 20% (v/v) bổ sung NAA 2 mg/L [79] và
AgNO3 từ 0,5-2,5 mg/L để thăm dò khả năng tạo rễ. Thí nghiệm được đánh giá
sau 4 tuần nuôi cấy.
2.2.1.3. Nuôi cấy callus
Cây nghệ đen in vitro 4 tuần tuổi sinh trương trên môi trường tạo rễ trong
lần cấy chuyển đầu tiên được sử dụng làm nguyên liệu tạo callus. Gốc lá (leaf-
base) (0,2×1 cm) được nuôi trên môi trường MS có sucrose 20 g/L, agar 8 g/L,
bổ sung 2,4-D 3 mg/L và BAP 3 mg/L để tạo callus. Callus sơ cấp sau đó được
cắt thành các khối nhỏ (đường kính ~2 mm) rồi cấy chuyển lên môi trường mới
có 2,4-D 1 mg/L và KIN hoặc NAA từ 0,5-2 mg/L hoặc AgNO3 từ 0,5-2,5 mg/L
để tăng sinh khối mô. Thí nghiệm được đánh giá sau 4 tuần nuôi cấy. Các callus
sinh trương trên môi trường thích hợp sẽ được cấy chuyển định kỳ 2 tuần/lần
để duy trì nguồn nguyên liệu cho nuôi cấy tế bào.
2.2.1.4. Nuôi cấy tế bào
Callus vàng rời, 2 tuần tuổi sinh trương trên môi trường tốt nhất được sử
dụng làm nguyên liệu trong nuôi cấy tế bào huyền phù. Chuyển 3 g callus vào
bình tam giác 250 mL chứa 50 mL môi trường MS lỏng có sucrose 20 g/L, bổ
sung 2,4-D 3 mg/L và BAP 3 mg/L, khuấy trộn cho đến khi xuất hiện các tế

42
bào tự do (môi trường bị đục và quan sát thấy tế bào dưới kính hiển vi). Sau đó,
các tế bào này được chuyển sang môi trường mới có thành phần tương tự và
nuôi lắc 150 vòng/phút trong 18 ngày để thu sinh khối [82].
Tế bào được thu hoạch 2 ngày/lần (thu nguyên bình), dịch môi trường
được lọc bằng giấy lọc và cân để xác định sinh khối tươi. Sinh khối tươi sau đó
được sấy ơ 50oC cho đến khi khối lượng không đổi để xác định khối lượng khô.
Các môi trường nuôi cấy đều được điều chỉnh pH đến 5,8 và khử trùng ơ
121oC trong 20 phút. Các thí nghiệm nuôi cấy được duy trì ơ 25±2ºC, cường
độ chiếu sáng 2.000-3.000 lux và thời gian chiếu sáng 12 giờ/ngày, ngoại trừ
nuôi cấy tế bào là 500 lux.
2.2.2. Phân lập các gen tổng hợp curcuminoid
2.2.2.1. Tách chiết DNA tổng số
DNA tổng số từ lá nghệ đen được tách chiết bằng phương pháp CTAB
theo Babu và cs. (2014) có cải tiến [21]. Nghiền 1 g lá trong nitrogen lỏng cho
đến khi thu được bột mịn sau đó chuyển vào tube Eppendorf, bổ sung 1 mL
đệm CTAB (Tris.HCl 100 mM pH 8, EDTA 20 mM pH 8, CTAB 2%, NaCl
1,4 M và β-mercaptoethanol 0,2%) đã được làm ấm từ trước ơ 65ºC và vortex
mạnh trong 5 phút. Ủ mẫu ơ 65ºC trong 1 giờ, vortex 15 phút/lần trong 10 giây,
sau đó ly tâm 13.000 vòng/phút trong 15 phút ơ 4ºC để thu dịch nổi. Bổ sung
một thể tích tương đương dung dịch phenol/chloroform/isoamylalcohol (tỷ lệ:
25/24/1), vortex và ủ ơ nhiệt độ phòng trong 10 phút. Ly tâm 13.000 vòng/phút
trong 15 phút ơ 4ºC để thu dịch nổi. Tiếp tục bổ sung một thể tích tương đương
isopropanol, vortex và ủ ơ -20ºC trong 1 giờ. Ly tâm 13.000 vòng/phút trong
20 phút ơ 4ºC để thu kết tủa DNA. Rửa kết tủa bằng 500 µL ethanol 70% lạnh,
ly tâm 12.000 vòng/phút trong 5 phút ơ 4ºC, loại bỏ dịch nổi. Để mẫu DNA
khô tự nhiên, sau đó hòa tan trong 20 µL 1×TE (Tris.HCl 10 mM pH 8 và
EDTA 1 mM). DNA tổng số được điện di kiểm tra trên gel agarose 1% và

43
nhuộm bằng RedSafeTM (Intron Biotechnology, Hàn Quốc). Hình ảnh điện di
được quan sát dưới ánh sáng tử ngoại (UV transilluminator, Wealtec). DNA
tổng số được bảo quản ơ -20ºC để sử dụng cho các thí nghiệm tiếp theo.
2.2.2.2. Khuếch đại PCR
DNA tổng số của nghệ đen được sử dụng làm khuôn mẫu để khuếch đại
gen mã hóa enzyme diketide-CoA synthase (CzDCS) và các gen mã hóa
enzyme curcumin synthase (CzCURS1, CzCURS2 và CzCURS3) dựa vào các
cặp primer đặc hiệu của chúng (Bảng 2.1). Các primer được thiết kế dựa trên
các vùng 5’ và 3’ không dịch mã ơ các gen DCS, CURS1, CURS2 và CURS3
của nghệ vàng (Mã số GenBank: AB495006.1, AB495007.1, AB506762.1 và
AB506763.1) để đảm bảo tính đặc hiệu của các sản phẩm PCR do trình tự
nucleotide của các gen này có độ tương đồng rất cao [66].
Để có được vùng bên cạnh (flanking region) trình tự CDS của các gen
tham gia tổng hợp curcuminoid của nghệ vàng, trình tự mRNA của các gen đã
được BLAST với cơ sơ dữ liệu EST của loài này, các đoạn có giá trị E-value
cao sẽ được lựa chọn để sử dụng với trình tự CDS tương ứng tạo ra các mRNA
liên tục hoàn chỉnh. Chuỗi mRNA hoàn chỉnh giả định của các gen DCS,
CURS1, CURS2 và CURS3 ơ nghệ vàng được dùng làm khuôn mẫu để thiết kế
primer khuếch đại các gen tương ứng ơ nghệ đen.
Thành phần PCR bao gồm: 50 ng DNA tổng số, 10 pmol primer mỗi loại,
6 µL 2×Go Taq® Green Master Mix (Promega, Mỹ), bổ sung nước cất vô trùng
vừa đủ 12 µL. Khuếch đại PCR được thực hiện trong máy luân nhiệt Veriti 96
Well Thermal Cycler (ABI, Mỹ) theo chu trình: 95ºC/15 phút; 30 chu kỳ:
95ºC/30 giây, 53ºC/30 giây, và 72ºC/1 phút; cuối cùng 72ºC/10 phút.
2.2.2.3. Tạo dòng và chú giải gen
Sản phẩm PCR được tinh sạch bằng GeneJET Gel Extraction Kit (Thermo

44
Scientific, Mỹ) theo hướng dẫn của nhà sản xuất. Sau khi tinh sạch chúng được
gắn vào vector pGEM®T-Easy (Hình 2.3) bằng T4 DNA ligase (Promega, Mỹ)
với thành phần phản ứng (trong 10 µL) gồm có: 3 unit T4 DNA ligase, 1 µL
10× T4 DNA ligase buffer, 50 ng pGEM®-T Easy, 70 ng sản phẩm PCR của
mỗi gen. Hỗn hợp được trộn đều và ủ ơ 4ºC qua đêm. Vector pGEM®-T Easy
mang đoạn chèn được biến nạp vào tế bào E. coli TOP10 bằng phương pháp
sốc nhiệt theo Sambrook và Russell (2001) [133]. Thể biến nạp được chọn lọc
trên môi trường LB rắn (trypton 1%, YE 0,5%, NaCl 1% và agar 1,5%) có bổ
sung 50 μg/mL ampicillin. Kiểm tra sự có mặt của đoạn chèn trong vector
pGEM®T-Easy tái tổ hợp bằng phương pháp PCR khuẩn lạc với các cặp primer
đặc hiệu. DNA plasmid sau đó được tách chiết theo phương pháp alkaline lysis
[133] và trình tự nucleotide của sản phẩm PCR được xác định bằng phương
pháp dideoxy terminator của Sanger trên máy ABI PRSIM 3100 (Mỹ). Các gen
(sản phẩm PCR) sau khi giải trình tự được so sánh với mRNA của nghệ vàng
bằng công cụ blastn, blastx và dự đoán cấu trúc của nó bằng phần mềm
FGENES.
Hình 2.3. Vector pGEM®-T Easy (Promega, Mỹ).
Bảng 2.1. Trình tự các primer được dùng để khuếch đại vùng CDS của các gen tổng

45
hợp curcuminoid ơ nghệ đen.
Gen Primer Trình tự nucleotide (5’- 3’) Chiều dài ước
tính của sản
phẩm PCR
(bp)
CzDCS DCS-F GTCGTTTCTGTGACCTTCTC
~ 1400 DCS-R CTTTTGGATGCAGACTGGAACA
CzCURS1 CURS1-F CTGCGACTGCGAGAAGAAGC
~ 1250 CURS1-R CAGATAGACAGCCATACAAACC
CzCURS2 CURS2-F GCACGCGTTTTCTTGCTAATC
~ 1300 CURS2-R GATCGTGTTCATAATTCACTGG
CzCURS3 CURS3-F CTAGCTAGCTGCAATTCGTT
~ 1250 CURS3-R GTGCTAGCTTAGCTTGACGTA
2.2.2.4. Xây dựng cây phả hệ
Phân tích mối quan hệ di truyền dựa trên các trình tự nucleotide thu được
sẽ giúp đánh giá chính xác hơn các gen mới tìm thấy ơ nghệ đen. Vùng CDS
của các gen DCS, CURS1, CURS2, CURS3 của cây nghệ đen, DCS1 và DCS2
(Mã số GenBank: KM880191.1 và AB535216.1) của nghệ vàng, gen type III
PKS (polyketide synthase) của cây chuối (Musa accuminata) (Mã số GenBank:
GU724611.1), và 4 gen mới thu được từ nghệ đen được sử dụng để xây dựng
cây phả hệ thông qua phần mềm MEGA7. Gen PKS từ nấm A. alternate (Mã
số GenBank: JX103640.1) được sử dụng làm nhóm ngoại. Cây phả hệ được
xây dựng dựa trên phương pháp Maximum Likelihood có độ lặp lại (bootstrap)
là 2.000 lần.

46
2.2.3. Xác định sự biểu hiện của các gen tổng hợp curcuminoid
2.2.3.1. Xử lý elicitor
Các thí nghiệm nghiên cứu ảnh hương của elicitor lên khả năng sinh
trương và tích lũy curcumin được thiết kế dựa trên kết quả nghiên cứu quá trình
nuôi cấy huyền phù tế bào cây nghệ đen và các kết quả đạt được trước đây của
nhóm nghiêm cứu trên các đối tượng khác. Các loại elicitor được sử dụng là
MeJA, SA và YE, đây là các loại elicitor đã được chứng minh là có hiệu quả
trên tế bào cây rau má, cà gai leo,… Khoảng nồng độ elicitor, thời gian xử lý
elicitor sử dụng trong nghiên cứu này cũng được kế thừa từ các nghiên cứu
trước đây của nhóm nghiên cứu [87], [88].
Chuyển 3 g sinh khối tế bào nghệ đen 10 ngày tuổi vào môi trường lỏng
(môi trường MS cơ bản có sucrose 20 g/L, bổ sung 2,4-D 3 mg/L và BAP 3
mg/L) để tiến hành xử lý với 3 loại elicitor khác nhau. Elicitor được bổ sung
vào môi trường nuôi cấy ơ các nồng độ khác nhau (MeJA 25-150 µM, SA 50-
150 µM và YE 0,1-1,5 g/L) và tại các thời điểm khác nhau (thời điểm ban đầu
hoặc ngày thứ 5 của quá trình nuôi cấy huyền phù). Sau 14 ngày nuôi cấy trong
bình tam giác, thu sinh khối tế bào để xác định khối lượng tươi và khối lượng
khô, mức độ biểu hiện của các gen và mức độ tích lũy curcumin [80], [82].
2.2.3.2. Phân tích RT-PCR
Mức độ biểu hiện của các gen tổng hợp curcuminoid trong các mẫu khác
nhau được phân tích bằng kỹ thuật PCR phiên mã ngược (RT-PCR) với các
primer được thiết kế cho các đoạn chỉ thị của các gen này (Bảng 2.2). Primer
được thiết kế dựa trên trình tự mRNA dự đoán của các gen CzDCS, CzCURS1,
CzCURS2 và CzCURS3 bằng công cụ Primer BLAST. Trong quá trình thiết kế,
các vùng có sự khác nhau lớn giữa các gen được lựa chọn để đảm bảo tính đặc
hiệu của đoạn chỉ thị cho toàn bộ gen (Hình 2.4).
Mẫu vật (200 mg callus/tế bào hoặc 100 mg củ nghệ đen 1 năm tuổi) được
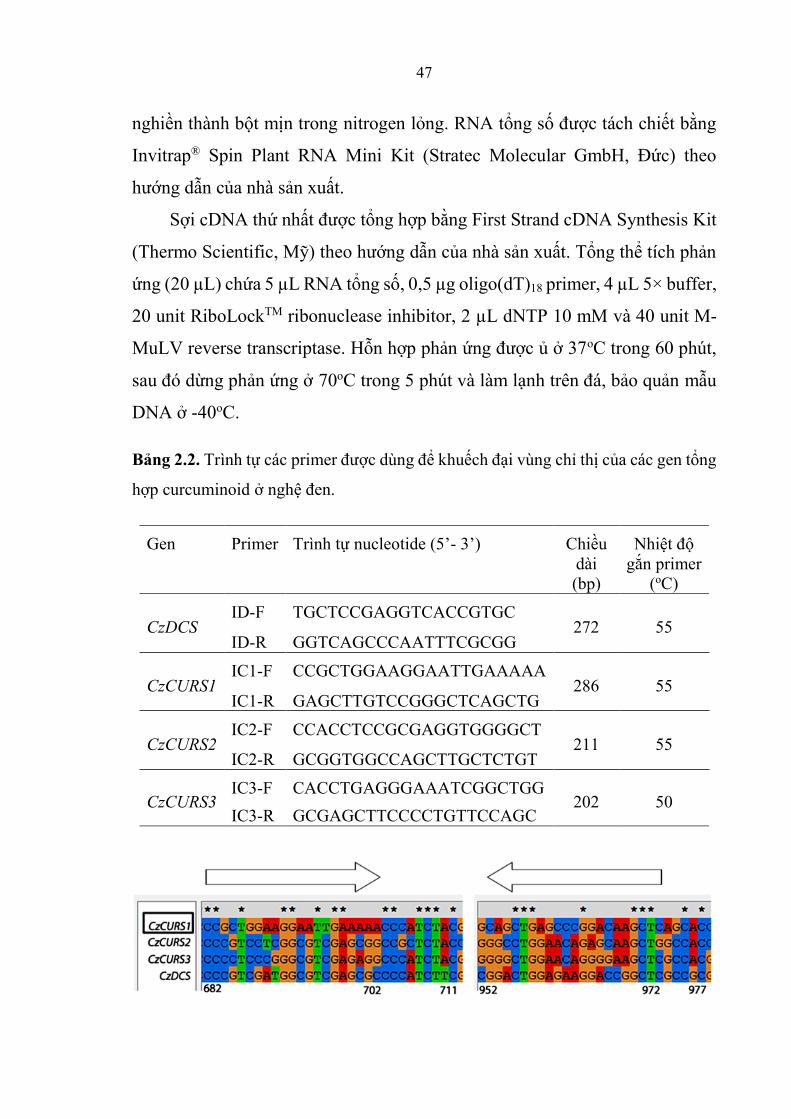
47
nghiền thành bột mịn trong nitrogen lỏng. RNA tổng số được tách chiết bằng
Invitrap® Spin Plant RNA Mini Kit (Stratec Molecular GmbH, Đức) theo
hướng dẫn của nhà sản xuất.
Sợi cDNA thứ nhất được tổng hợp bằng First Strand cDNA Synthesis Kit
(Thermo Scientific, Mỹ) theo hướng dẫn của nhà sản xuất. Tổng thể tích phản
ứng (20 µL) chứa 5 µL RNA tổng số, 0,5 µg oligo(dT)18 primer, 4 µL 5× buffer,
20 unit RiboLockTM ribonuclease inhibitor, 2 µL dNTP 10 mM và 40 unit M-
MuLV reverse transcriptase. Hỗn hợp phản ứng được ủ ơ 37oC trong 60 phút,
sau đó dừng phản ứng ơ 70oC trong 5 phút và làm lạnh trên đá, bảo quản mẫu
DNA ơ -40oC.
Bảng 2.2. Trình tự các primer được dùng để khuếch đại vùng chỉ thị của các gen tổng
hợp curcuminoid ơ nghệ đen.
Gen Primer Trình tự nucleotide (5’- 3’) Chiều
dài
(bp)
Nhiệt độ
gắn primer
(oC)
CzDCS ID-F TGCTCCGAGGTCACCGTGC
272 55 ID-R GGTCAGCCCAATTTCGCGG
CzCURS1 IC1-F CCGCTGGAAGGAATTGAAAAA
286 55 IC1-R GAGCTTGTCCGGGCTCAGCTG
CzCURS2 IC2-F CCACCTCCGCGAGGTGGGGCT
211 55 IC2-R GCGGTGGCCAGCTTGCTCTGT
CzCURS3 IC3-F CACCTGAGGGAAATCGGCTGG
202 50 IC3-R GCGAGCTTCCCCTGTTCCAGC

48
Hình 2.4. Vị trí của primer xuôi và ngược trên gen CzCURS1. Mô hình này cũng
được sử dụng để thiết kế primer cho các gen CzDCS, CzCURS2 và CzCURS3 của
nghệ đen.
Thành phần PCR bao gồm 50 ng cDNA, 6 µL 2× GoTaq® Green Master
Mix (Promega, Mỹ), 10 pmol primer mỗi loại, nước cất khử trùng được bổ sung
đến thể tích cuối cùng là 12 µL. Khuếch đại PCR được thực hiện trong máy
luân nhiệt Veriti 96 Well Thermal Cycler (ABI, Mỹ) theo chu trình: 95ºC/2
phút; 20-30 chu kỳ: 95ºC/30 giây, 50-55ºC/30 giây và 72ºC/30 giây; cuối cùng
là 72ºC/10 phút. 3 µL sản phẩm PCR được sử dụng để điện di trên gel agarose
1% (w/v) và xác định cường độ của các băng DNA bằng chương trình ImageJ
thông qua đo mật độ các điểm ảnh (ver. k1.45).
2.2.3.3. Phân tích HPLC
Tách chiết curcumin được tiến hành theo phương pháp của Paramapojn và
Gritsanapan (2009) [114]. Tế bào hoặc củ nghệ đen 10 tháng tuổi được sấy khô
ơ 50°C đến khối lượng không đổi sau đó nghiền thành bột mịn. Hòa 2 g bột
mịn trong 20 mL ethanol 70%. Hỗn hợp được xử lý bằng sóng siêu âm trên
máy T 700/H (Elma, Đức) trong 30 phút ơ 30oC. Dịch chiết được lọc bằng giấy
Whatman (No. 1), bã được chiết lại với cùng dung môi cho tới khi hết hoàn
toàn (dịch chiết cuối cùng là không màu). Dịch chiết nhiều lần được trộn lại và
cô ơ 70oC bằng máy cô quay chân không (Heidolph, Đức). Mẫu sau khi cô được
hòa tan trơ lại với ethanol 70% đến 10 mL, lọc bằng màng Millipore 0,25 μm
(Sartorius, Đức) và pha loãng dịch chiết 2 lần, 10 μL dịch chiết được sử dụng
để chạy HPLC.
Điều kiện HPLC: cột Hypersil™ MOS C8 LC Column (5 µm, 4,6×150
mm), tốc độ chạy 1 mL/phút, thời gian chạy 15-30 phút, pha tĩnh là silica gel
(pha ngược) và pha động là acetonitrile/acetic acid 2% (35/65, v/v). Dữ liệu
phân tích được thu nhận bằng đầu dò PDA (photodiode array) ơ bước sóng 420

49
nm trên hệ thống Shimazu 20AD (Nhật Bản).
Tất cả dung môi đạt tiêu chuẩn phân tích được mua từ các Hãng Sigma
(Mỹ) và Merck (Đức). Curcumin của hãng Sigma được dùng làm chất chuẩn
để xác định hàm lượng curcumin. Hàm lượng curcumin được xác định dựa theo
đường chuẩn được xây dựng từ diện tích peak của các nồng độ curcumin chuẩn
khác nhau trong phân tích HPLC, phương trình đường chuẩn là y = 0,213x,
trong đó y là hàm lượng curcumin, x là diện ticvhs peak, R2 = 0,983.
2.2.4. Xử lý thông kê
Các thí nghiệm được bố trí ngẫu nhiên, mỗi thí nghiệm được lặp lại 3 lần.
Đối với các thí nghiệm nuôi cấy in vitro mỗi lần lặp ít nhất 10 mẫu.
Đối với các thí nghiệm phân tích (RT-PCR và HPLC) mỗi lần lặp lại 3
mẫu.
Kết quả thí nghiệm được tính trung bình và phân tích ANOVA bằng
Duncan’s test với p<0,05 sử dụng phần mềm SPSS ver 19.0./.

50
Chương 3
KẾT QUẢ NGHIÊN CỨU
3.1. THIẾT LẬP NUÔI CẤY TẾ BÀO
Chồi mầm của củ nghệ đen được nuôi cấy để nhân giống cây in vitro. Sau
đó, dùng gốc lá của cây in vitro để tạo callus và thiết lập nuôi cấy tế bào.
3.1.1. Nhân giông cây nghệ đen in vitro
Trước tiên, cây in vitro được nhân giống để tạo nguồn nguyên liệu dùng
cho nuôi cấy callus. Trong các nghiên cứu được công bố trước đây, Loc và cs.
(2005) [79], Stanly và Keng (2007) [146], Shahinozzaman và cs. (2013) [142]
đã nhân giống được cây nghệ đen in vitro. Sau đó, Loc và cs. (2006, 2008 và
2009) cũng đã nuôi cấy thành công tế bào nghệ đen [81], [80], [82]. Theo một
số nghiên cứu, AgNO3 được bổ sung vào môi trường dinh dưỡng đã có tác dụng
ức chế hoạt động của ethylene nội sinh, nhờ đó kích thích được quá trình sinh
trương của tế bào [73], [134]. Trong nghiên cứu này, chúng tôi thử cải thiện
hiệu suất nuôi cấy bằng cách thăm dò ảnh hương của AgNO3 ơ các nồng độ
khác nhau lên các quá trình sinh trương in vitro của nghệ đen.
3.1.1.1. Tái sinh chồi
Sử dụng chồi in vitro nguyên vẹn. Kết quả nghiên cứu cho thấy tất cả
môi trường được thăm dò đều có khả năng tái sinh chồi (Bảng 3.1). Ở môi
trường đối chứng (ĐC), chồi đỉnh bắt đầu kéo dài từ ngày thứ 3 và sau đó xuất
hiện các chồi mới từ ngày thứ 5. Trên các môi trường có bổ sung AgNO3, chồi
đỉnh bắt đầu kéo dài và hình thành các chồi mới muộn hơn 1 ngày. Tuy nhiên,
các môi trường có bổ sung AgNO3 đều cho tỷ lệ tạo chồi mới cao hơn hoặc
tương đương ĐC. Số lượng chồi tăng dần (4,8-6,4 chồi/mẫu) và chiều cao chồi
khá đồng đều (6,6-7,1 cm, p> 0,05) ơ các nồng độ AgNO3 từ 0,5-1,5 mg/L. Khi

51
tăng AgNO3 lên 2 mg/L, số lượng và chiều cao chồi có xu hướng giảm (5,1
chồi/ 6,2 cm). Trên tất cả môi trường, các chồi mới hình thành có hiện tượng
tạo rễ nhưng không mạnh. Chồi hình thành trên môi trường bổ sung AgNO3 1,5
mg/L có màu xanh đậm hơn ĐC; ơ các môi trường khác chồi có màu xanh
tương đương hoặc nhạt hơn.
Bảng 3.1. Ảnh hương của AgNO3 lên khả năng tái sinh chồi mới của chồi in vitro
nguyên vẹn.
AgNO3 (mg/L) Tỷ lệ tạo chồi
(%) Sô chồi/mẫu
Chiều cao chồi
(cm)
ĐC 100 5,4ab 6,3ab
0,5 100 5,6ab 7,1a
1,0 100 6,1ab 6,9ab
1,5 100 6,4a 6,6ab
2,0 100 5,1b 6,2b
ĐC: đối chứng không bổ sung AgNO3, chú thích này dùng chung cho các bảng 3.2 và
3.3. Các chữ cái khác nhau trên cùng một cột chỉ sự sai khác có ý nghĩa thống kê với p<
0,05 (Duncan’s test), chú thích này dùng chung cho tất cả các bảng (ngoại trừ bảng 3.7).
Sử dụng chồi in vitro chẻ đôi. Số liệu ơ bảng 3.2 cho thấy tất cả môi
trường được thăm dò đều có khả năng tái sinh chồi tương tự như thí nghiệm
chồi in vitro nguyên vẹn. Ở môi trường ĐC, chồi mới xuất hiện từ ngày thứ 5,
còn trên các môi trường có AgNO3 chồi mới hình thành sớm hơn 2 ngày. Các
môi trường có bổ sung AgNO3 đều cho tỷ lệ tạo chồi mới cao hơn ĐC. Số lượng
chồi tăng dần (5,9-7,5 chồi/mẫu), nhưng chiều cao của chúng thì sai khác không
nhiều (3,6-3,9 cm) khi AgNO3 được bổ sung từ 0,5-1,5 mg/L. Nếu tăng AgNO3
lên 2 mg/L thì số lượng và chiều cao chồi bắt đầu giảm (6,1 chồi/ 3,4 cm). Hiện
tượng tạo rễ của các chồi mới tái sinh cũng xảy ra ơ tất cả môi trường nhưng
không đáng kể. Các chồi ơ môi trường có 1,5 mg/L AgNO3 có màu xanh đậm

52
hơn ĐC, còn ơ các môi trường khác chồi có màu xanh nhạt hơn. Nhìn chung,
so với chồi nguyên vẹn thì chồi chẻ đôi có hiệu quả tái sinh chồi mới cao
(7,5/6,4 chồi).
Bảng 3.2. Ảnh hương của AgNO3 lên khả năng tái sinh chồi mới của chồi in vitro đã
được chẻ đôi.
AgNO3 (mg/L) Tỷ lệ tạo chồi (%) Sô chồi/mẫu Chiều cao chồi
(cm)
ĐC 100 5,6c 3,4c
0,5 100 5,9bc 3,6bc
1,0 100 6,6b 3,9a
1,5 100 7,5a 3,7b
2,0 100 6,1bc 3,6bc
3.1.1.2. Tạo rễ in vitro
Trong quy trình nhân giống cây in vitro, tạo rễ là giai đoạn quan trọng để
tạo ra cây hoàn chỉnh, việc cải thiện khả năng tạo rễ của cây nghệ đen dựa trên
việc bổ sung AgNO3 vào môi trường sẽ góp phần hoàn chỉnh quy trình nhân
giống cây này. Để nghiên cứu ảnh hương của AgNO3 lên khả năng tạo rễ của
chồi nghệ đen in vitro, môi trường có bổ sung NAA 2 mg/L kết hợp với AgNO3
từ 0,5-2,5 mg/L đã được sử dụng và kết quả được trình bày ơ bảng 3.3.
Khi kết hợp NAA với AgNO3, tất cả môi trường thăm dò đều có khả năng
tạo rễ. Trên môi trường ĐC rễ xuất hiện muộn ơ ngày thứ 5, còn ơ các môi
trường có AgNO3 rễ hình thành sớm hơn, chỉ sau 3 ngày nuôi cấy và số lượng
cũng nhiều hơn.

53
Bảng 3.3. Ảnh hương của AgNO3 lên khả năng tạo rễ của chồi in vitro.
AgNO3 (mg/L) Tỷ lệ tạo rễ (%) Sô rễ/chồi Chiều dai rễ
(cm)
ĐC 100 18,3c 0,6c
0,5 100 20,0bc 0,7ab
1,0 100 22,5b 0,7ab
1,5 100 27,0a 0,8a
2,0 100 20,8bc 0,8a
Hình 3.1. Cây nghệ đen in vitro 2 tháng tuổi trên môi trường chứa NAA 2 mg/L và
AgNO3 1,5 mg/L.
Số lượng rễ tăng từ 20-27 rễ và chiều dài của chúng trong khoảng 0,7-0,8
cm (p> 0,05) ơ môi trường có AgNO3 từ 0,5-1,5 mg/L, khi tăng AgNO3 lên 2-

54
2,5 mg/L rễ giảm số lượng rõ rệt (18,8-20,8 rễ) và tương đương với ĐC (18,3
rễ) nhưng chiều dài không thay đổi đáng kể (0,6-0,8 cm và ĐC là 0,6 cm). Nhìn
chung, môi trường có bổ sung AgNO3, ngoài tác dụng tạo rễ còn có thể hình
thành một vài chồi con.
3.1.2. Nuôi cấy callus
Một số nghiên cứu trước đây cho thấy callus tế bào nghệ đen sinh trương
tốt trên môi trường có bổ sung 2,4-D và BAP, trong đó 2,4-D đóng vai trò chính
(Loc và cs. 2006, 2008 và 2009) [81], [80], [82]. Trong nghiên cứu này, chúng
tôi thay BAP bằng một chất kích thích sinh trương khác cùng nhóm cytokinine
(KIN), auxin (NAA) hoặc AgNO3 để tiếp tục cải thiện khả năng sinh trương
của callus nghệ đen, đây là các chất đã được nhiều tác giả sử dụng trên nhiều
loài nghệ khác nhau [102], [120], [168].
3.1.2.1. Ảnh hương của 2,4-D và KIN
Kết quả trình bày ơ bảng 3.4 cho thấy 2,4-D và KIN ảnh hương khá rõ rệt
lên sinh trương của callus. Nhìn chung, ơ môi trường có 2,4-D 1 mg/L và KIN
từ 1-1,5 mg/L callus sinh trương mạnh hơn ĐC; đường kính trung bình của
chúng khoảng 1,53-1,56 cm, khối lượng tươi và khối lượng khô tương ứng từ
0,84-0,9 g và 83,15-84,11 mg. Ở các công thức khác, đường kính callus tương
đương ĐC (p<0,05) nhưng khối lượng tươi và khô vẫn cao hơn. Callus ơ môi
trường có 2,4-D 1 mg/L và KIN 1,5 mg/L có màu vàng hơn ĐC, ơ các môi
trường khác callus có màu vàng tương đương hoặc nhạt hơn (Hình 3.2).

55
Bảng 3.4. Ảnh hương của KIN từ 0,5-2 mg/L và 2,4-D 1 mg/L lên sinh trương của
callus nghệ đen.
KIN (mg/L) Đường kính (cm) Khôi lượng
Tươi (g) Khô (mg)
ĐC 1,35c 0,68c 61,24c
0,5 1,48abc 0,79b 79,03b
1,0 1,53ab 0,84ab 83,15a
1,5 1,56a 0,90a 84,11a
2,0 1,38bc 0,85ab 80,37b
ĐC: đối chứng là môi trường có 2,4-D 3 mg/L và BA 3 mg/L. Chú thích này dùng
chung cho các bảng 3.5 và 3.6.
3.1.2.2. Ảnh hương của 2,4-D và NAA
Kết quả trình bày ơ bảng 3.5 cho thấy hiệu quả kích thích sinh trương
callus của NAA dường như không bằng KIN. Ở môi trường thích hợp nhất (2,4-
D 1 mg/L và NAA từ 1-1,5 mg/L), đường kính callus khoảng 1,5-1,56 cm ((p>
0,05), khối lượng tươi và khô tương ứng là 0,76-0,79 g và 73,91-80,16 mg. Ở
các môi trường khác sinh trương của callus tương đương ĐC (p> 0,05). Callus
trên môi trường chứa 2,4-D 1 mg/L và NAA 1 mg/L có màu vàng hơi xanh hơn
ĐC, ơ các môi trường khác callus có màu vàng nhạt hơn.
3.1.2.3. Ảnh hương của 2,4-D và AgNO3
Tương tự 2 thí nghiệm trên, kết quả nghiên cứu cho thấy ơ tất cả môi
trường được thăm dò, sinh trương của callus đều tăng so với ĐC. Mặc dù ảnh
hương của các nồng độ AgNO3 từ 0,5-2 mg/L lên đường kính và khối lượng
tươi của mô không khác nhau đáng kể (p> 0,05) nhưng ơ môi trường có AgNO3
1,5 mg/L callus có khối lượng khô cao nhất (62,43 mg) (Bảng 3.6).
Callus trên môi trường được bổ sung AgNO3 1,5 mg/L có màu vàng đậm
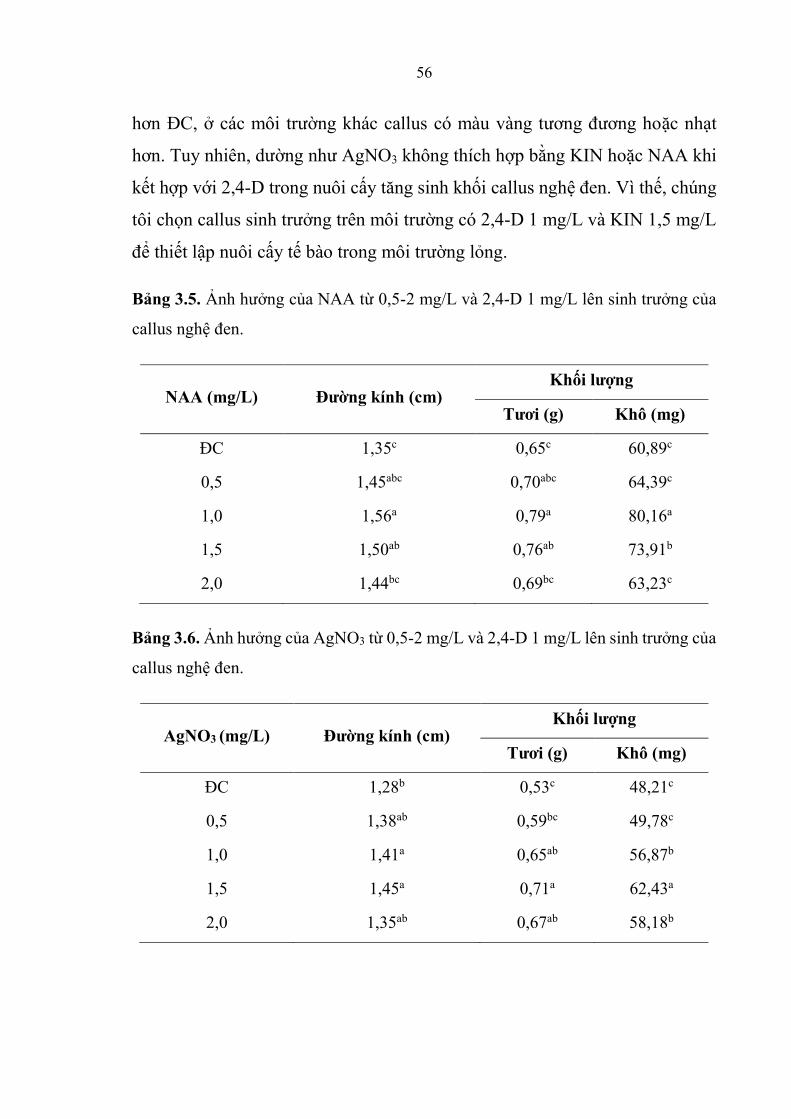
56
hơn ĐC, ơ các môi trường khác callus có màu vàng tương đương hoặc nhạt
hơn. Tuy nhiên, dường như AgNO3 không thích hợp bằng KIN hoặc NAA khi
kết hợp với 2,4-D trong nuôi cấy tăng sinh khối callus nghệ đen. Vì thế, chúng
tôi chọn callus sinh trương trên môi trường có 2,4-D 1 mg/L và KIN 1,5 mg/L
để thiết lập nuôi cấy tế bào trong môi trường lỏng.
Bảng 3.5. Ảnh hương của NAA từ 0,5-2 mg/L và 2,4-D 1 mg/L lên sinh trương của
callus nghệ đen.
NAA (mg/L) Đường kính (cm) Khôi lượng
Tươi (g) Khô (mg)
ĐC 1,35c 0,65c 60,89c
0,5 1,45abc 0,70abc 64,39c
1,0 1,56a 0,79a 80,16a
1,5 1,50ab 0,76ab 73,91b
2,0 1,44bc 0,69bc 63,23c
Bảng 3.6. Ảnh hương của AgNO3 từ 0,5-2 mg/L và 2,4-D 1 mg/L lên sinh trương của
callus nghệ đen.
AgNO3 (mg/L) Đường kính (cm) Khôi lượng
Tươi (g) Khô (mg)
ĐC 1,28b 0,53c 48,21c
0,5 1,38ab 0,59bc 49,78c
1,0 1,41a 0,65ab 56,87b
1,5 1,45a 0,71a 62,43a
2,0 1,35ab 0,67ab 58,18b

57
Hình 3.2. Callus nghệ đen 2 tuần tuổi sinh trương trên môi trường có 2,4-D 1 mg/L
kết hợp với KIN 1,5 mg/L (A) và đối chứng sinh trương trên môi trường có 2,4-D 3
mg/L kết hợp với BAP 3 mg/L (B).
3.1.3. Nuôi cấy tế bao nghệ đen
Kỹ thuật nuôi cấy tế bào thực vật có nhiều triển vọng và ứng dụng lâu dài
trong việc sản xuất các hợp chất tự nhiên, đặc biệt là các chất dùng trong y học.
Hướng nghiên cứu này sẽ dẫn đến sự ổn định về mặt chất lượng và số lượng
sản phẩm, ít phụ thuộc vào tự nhiên. Đồng thời, là nguồn nguyên liệu cho những
thí nghiệm sinh lý, hóa sinh và ứng dụng để tách chiết các hợp chất thứ cấp
khác nhau [5]. Các nghiên cứu cho thấy rằng, nuôi cấy tế bào thực vật có khả
năng sản xuất các sản phẩm thứ cấp với hàm lượng lớn hơn so với các chất đó
được chiết từ cây ngoài tự nhiên [83]. Ưu điểm của chúng là có thể cung cấp
sản phẩm một cách liên tục và đáng tin cậy. Trong sản xuất curcumin, nuôi cấy
tế bào các loài nghệ là một giải pháp được nhiều tác giả thực hiện.
Trong nghiên cứu này, chúng tôi đã nuôi cấy tế bào cây nghệ đen trong
A
B

58
môi trường lỏng nhằm tạo nguồn nguyên liệu cho quá trình nghiên cứu sản xuất
curcumin sau này. Để xác định thời điểm sinh trương cực đại của tế bào nghệ
đen khi nuôi cấy huyền phù, chúng tôi đã nuôi 3 g callus 2 tuần tuổi trong bình
tam giác 250 mL chứa 50 mL môi trường lỏng, sinh khối tế bào được thu hoạch
2 ngày/lần để xây dựng đường cong sinh trương, thời gian theo dõi là 18 ngày.
Kết quả trình bày ơ hình 3.3 cho thấy pha thích nghi của tế bào khá ngắn,
tế bào sớm chuyển qua pha tăng trương và tăng sinh liên tục từ 2 đến 14 ngày
nuôi cấy sau đó giảm dần. Sinh khối tươi đạt cực đại là gần 170 g/L, tương
đương với khối lượng khô khoảng 15 g/L (Hình 3.4 và 3.5). Các nghiên cứu
trước đây khối lượng khô chỉ đạt cao nhất khoảng 13,2 g/L trong một điều kiện
tương đương (Tuan và cs. 2011) [156]. Đây là cơ sơ để chúng tôi thiết kế các
thí nghiệm nghiên cứu ảnh hương của elicitor sau này.
Hình 3.3. Đường cong sinh trương của tế bào nghệ đen.
Khối lượng tương
Khối lượng khô
0.00
0.15
0.30
0.45
0.60
0.75
0.90
0.00
1.50
3.00
4.50
6.00
7.50
9.00
2 4 6 8 10 12 14 16 18
Khối l
ượ
ng
khô (
g/b
ình)
Khối l
ượ
ng
tươ
i (g/b
ình)
Thời gian nuôi cấy (ngày)
Sinh khối tươi
Sinh khối khô

59
Hình 3.4. Tế bào nghệ đen nuôi trong bình tam giác chứa môi trường MS bổ sung
2,4-D 3 mg/L và BAP 3 mg/L.
Hình 3.5. Sinh khối tươi (A) và khô (B) của tế bào nghệ đen.
3.2. NHẬN DẠNG CÁC GEN TỔNG HỢP CURCUMINOID
3.2.1. Phân lập gen
Kết quả khuếch đại PCR từ DNA tổng số của nghệ đen với các cặp primer
như đã trình bày ơ bảng 2.1 cho thấy sự hiện diện của các băng DNA đặc hiệu
có kích thước phù hợp với tính toán trên điện di đồ. Kích thước đầy đủ của các
A B

60
gen DCS, CURS1, CURS2 và CURS3 được giả định tương ứng khoảng 1,4 kb;
1,25 kb; 1,3 kb và 1,25 kb (Hình 3.6). Có thể nhận thấy 4 gen trên đều có mặt
trong cây nghệ đen và các primer được thiết kế là đặc hiệu cho các gen này.
Các sản phẩm PCR được tinh sạch và tạo dòng trong vector pGEM T-Easy
để giải trình tự nucleotide của chúng. Kết quả chỉ ra các gen DCS, CURS1,
CURS2 và CURS3 của nghệ đen có chiều dài lần lượt là 1382 bp, 1240 bp, 1288
bp và 1265 bp. Các gen này được đặt tên tương ứng là CzDCS, CzCURS1,
CzCURS2 và CzCURS3 và đã đăng ký trên GenBank với các mã số sau:
MF663785, MF402846, MF402847 và MF987835.
Để xác định các vùng intron/exon trên các gen CzDCS, CzCURS1,
CzCURS2 và CzCURS3 có nguồn gốc từ DNA tổng số, chúng tôi đã sử dụng
các phương pháp khác nhau. Trước tiên, so sánh trình tự của các gen này với
vùng mã hóa của các gen tương ứng ơ nghệ vàng đã được công bố trên
GenBank, các vùng không bắt cặp sẽ là vùng 5’ và 3’ không dịch mã cùng với
các intron. Thứ hai, chúng tôi sử dụng công cụ blastx và FGENES để xác định
lại kết quả của phương pháp trên. Bên cạnh đó, các lỗi gặp phải trong khi giải
trình tự các gen có thể ảnh hương đến việc dự đoán intron/exon sẽ được xử lý
để có trình tự chính xác. Kết quả cuối cùng cho thấy các gen CzCURS1,
CzCURS2 và CzCURS3 chỉ có 1 vùng intron trong khi gen CzDCS có 2 vùng
intron. Vùng intron/exon giả định của 4 gen được minh họa ơ hình 3.7.
Sau khi loại bỏ các intron, 2 gen CzDCS và CzCURS1 có chiều dài đoạn
cDNA là 1170 bp, gen CzCURS2 là 1176 bp và gen CzCURS3 là 1173 bp. Cả
4 gen này đều tương đồng 99% với các gen tương ứng ơ nghệ vàng (Hình 3.8-
3.11). So sánh trình tự amino acid suy diễn của các gen ơ nghệ đen và nghệ
vàng cho thấy CzDCS/DCS có tỷ lệ tương đồng là 387/389, CzCURS1/CURS1
là 387/389, CzCURS2/CURS2 là 387/391 và CzCURS3/CURS3 là 387/390
(Hình 3.12-3.15).

61
Hình 3.6. Sản phẩm PCR của các gen tổng hợp curcuminoid khuếch đại từ DNA tổng
số của nghệ đen. M1 và M2: DNA size marker (100 bp BioRad và 1 kb DNA Ladder
Geneaid), 1: CzDCS, 2: CzCURS1, 3: CzCURS2, 4: CzCURS3.
Hình 3.7. Sơ đồ sắp xếp của các intron/exon trên 4 gen tổng hợp curcuminoid ơ nghệ
đen. Trong đó, intron (đường thẳng), exon (hình hộp) và vị trí của nó. Số 1 là
nucleotide đầu tiên trong mã mơ đầu. Cấu trúc chung của chuỗi protein mã hóa được
thể hiện bên dưới (màu cam). Vị trí các vùng được xác định bằng công cụ
InterProScan.

62
CzDCS 1 ATGGAAGCGAACGGCTACCGCATAACTCACAGCGCCGACGGGCCGGCGACGATCTTGGCC 60
DCS 1 ATGGAAGCGAACGGCTACCGCATAACTCACAGCGCCGACGGGCCGGCGACGATCTTGGCC 60
CzDCS 61 ATCGGCACCGCCAACCCCACCAACGTCGTCGACCAGAACGCTTATCCCGACTTCTATTTC 120
DCS 61 ATCGGCACCGCCAACCCCACCAACGTCGTCGATCAGAACGCTTATCCCGACTTCTATTTC 120
CzDCS 121 CGGGTCACCAACTCCGAGCATCTGCAGGAACTCAAAGCCAAGTTTAGGCGCATCTGTGAG 180
DCS 121 CGGGTCACCAACTCCGAGTATCTGCAGGAACTCAAAGCCAAGTTTAGGCGCATCTGTGAG 180
CzDCS 181 AAAGCGGCGATCAGGAAGAGGCACTTGTACTTGACTGAGGAGATTTTGCGGGAGAATCCT 240
DCS 181 AAAGCGGCCATCAGGAAGAGGCACTTGTACTTGACTGAGGAGATTTTGCGGGAGAATCCT 240
CzDCS 241 AGCTTGCTGGCTCCCATGGCGCCGTCGTTCGACGCGCGGCAGGCGATCGTGGTGGAGGCG 300
DCS 241 AGCTTGCTGGCTCCCATGGCGCCGTCGTTCGACGCGCGGCAGGCGATCGTGGTGGAGGCG 300
CzDCS 301 GTGCCGAAGCTAGCGAAGGAGGCGGCGGAGAAGGCGATCAAGGAGTGGGGTCGCCCCAAA 360
DCS 301 GTGCCGAAGCTGGCGAAGGAGGCGGCGGAGAAGGCGATCAAGGAGTGGGGCCGCCCCAAA 360
CzDCS 361 TCGGACATCACGCACCTCGTCTTCTGCTCCGCGAGCGGAATCGACATGCCCGGCTCCGAC 420
DCS 361 TCGGACATCACGCACCTCGTCTTCTGCTCCGCGAGCGGAATCGACATGCCCGGCTCCGAC 420
CzDCS 421 CTGCAGCTTCTCAAGCTGCTCGGGCTCCCGCCGAGCGTCAATCGCGTCATGCTCTACAAC 480
DCS 421 CTGCAGCTTCTCAAGCTGCTCGGGCTCCCGCCGAGCGTCAATCGCGTCATGCTCTACAAC 480
CzDCS 481 GTCGGGTGCCACGCCGGTGGCACCGCCCTCCGCGTCGCCAAGGACCTCGCGGAGAACAAC 540
DCS 481 GTCGGGTGCCACGCCGGTGGCACCGCCCTCCGCGTCGCCAAGGACCTCGCGGAGAACAAC 540
CzDCS 541 CGCGGCGCGCGGGTGCTCGCCGTCTGCTCCGAGGTCACCGTGCTCTCCTACCGCGGCCCC 600
DCS 541 CGCGGCGCGCGGGTGCTCGCCGTCTGCTCCGAGGTCACCGTGCTCTCCTACCGCGGCCCC 600
CzDCS 601 CACCCCGCCCACATCGAGAGCCTCTTCGTCCAAGCTCTGTTTGGCGACGGCGCTGCCGCG 660
DCS 601 CACCCCGCCCACATCGAGAGCCTCTTCGTCCAAGCTCTGTTTGGCGACGGCGCCGCCGCG 660
CzDCS 661 CTCGTGGTCGGGTCCGACCCCGTCGATGGCGTCGAGCGCCCCATCTTCGAAATCGCCTCG 720
DCS 661 CTCGTGGTCGGGTCCGACCCCGTCGATGGCGTCGAGCGCCCCATCTTCGAAATCGCCTCG 720
CzDCS 721 GCATCCCAAGTGATGCTTCCGGAGAGCGAAGAGGCGGTAGGCGGCCACCTCCGCGAAATT 780
DCS 721 GCATCCCAAGTGATGCTTCCGGAGAGCGCAGAGGCGGTGGGCGGCCACCTCCGCGAAATT 780
CzDCS 781 GGGCTGACCTTCCACCTCAAGAGCCAGCTTCCGTCGATCATCGCGAGCAACATCGAGCAG 840
DCS 781 GGGCTGACCTTCCACCTCAAGAGCCAGCTTCCGTCGATCATCGCGAGCAACATCGAGCAG 840
CzDCS 841 AGCCTGACGACTGCGTGCTCGCCGCTGGGGCTGTCGGACTGGAACCAGCTGTTCTGGGCG 900
DCS 841 AGCCTGACGACTGCGTGCTCGCCGCTGGGGCTGTCGGACTGGAACCAGCTGTTCTGGGCG 900
CzDCS 901 GTTCACCCCGGCGGCCGAGCGATCCTGGACCAGGTGGAGGCGCGGCTCGGACTGGAGAAG 960
DCS 901 GTTCACCCCGGCGGCCGAGCGATCCTGGACCAGGTGGAGGCGCGGCTCGGACTGGAGAAG 960
CzDCS 961 GACCGGCTCGCCGCGACGCGGCACGTACTCAGCGAGTACGGCAACATGCAGAGCGCCACG 1020
DCS 961 GACCGGCTCGCCGCGACGCGGCACGTACTCAGCGAGTACGGCAACATGCAGAGCGCCACG 1020
CzDCS 1021 GTGCTGTTCATCCTGGACGAGATGCGGAACCGCTCGGCTGCGGAGGGCCACGCCACCACC 1080
DCS 1021 GTGCTGTTCATCCTGGACGAGATGCGGAACCGCTCGGCTGCGGAGGGCCACGCCACCACC 1080
CzDCS 1081 GGCGAGGGGCTCGACTGGGGCGTGCTTTTGGGCTTCGGCCCGGGACTCTCCATCGAGACC 1140
DCS 1081 GGCGAGGGGCTCGACTGGGGCGTGCTGTTGGGCTTCGGCCCGGGACTCTCCATCGAGACC 1140
CzDCS 1141 GTCGTCCTCCATAGTTGCAGACTGAACTAG 1170
DCS 1141 GTCGTCCTCCATAGTTGCAGACTGAACTAG 1170
Hình 3.8. So sánh trình tự nucleotide (nu) vùng CDS của gen CzDCS (MF663785) ơ
nghệ đen và DCS ơ nghệ vàng (AB495006.1). Số nu tương đồng: 1161/1170 (99%).
Hộp màu đen chỉ các nu khác nhau và đoạn chỉ thị của gen được đóng khung, chú
thích này dùng chung cho các hình 3.9-3.11.

63
CzCURS1 1 ATGGCCAACCTCCACGCGTTGCGCAGGGAGCAGAGGGCTCAAGGTCCCGCCACCATCATG 60
CURS1 1 ATGGCCAACCTCCACGCGTTGCGCAGGGAGCAGAGGGCTCAAGGTCCTGCCACCATCATG 60
CzCURS1 61 GCCATCGGGACGGCCACCCCCCCCAACCTCTACGAGCAGAGCACCTTCCCGGACTTCTAC 120
CURS1 61 GCCATCGGGACCGCCACCCCTCCCAACCTCTACGAGCAGAGCACCTTCCCGGACTTCTAC 120
CzCURS1 121 TTCCGCGTCACCAACTCCGACGACAAGCAGGAGCTCAAGAAAAAGTTCCGCCGCATTTGC 180
CURS1 121 TTCCGCGTCACCAACTCCGACGACAAGCAGGAGCTCAAGAAAAAGTTCCGCCGCATGTGC 180
CzCURS1 181 GATAAGACGATGGTGAAGAAGCGGTACCTGCACTTGACCGAGGAGATCCTGAAGGAGAGG 240
CURS1 181 GAGAAGACGATGGTGAAGAAGCGGTACCTGCACTTGACCGAGGAGATCCTGAAGGAGAGG 240
CzCURS1 241 CCCAAGCTCTGCTCCTACAAGGAGGCGTCGTTCGACGACCGGCAGGACATCGTGGTGGAG 300
CURS1 241 CCCAAGCTCTGCTCCTACAAGGAGGCGTCGTTCGACGACCGGCAGGACATCGTGGTGGAG 300
CzCURS1 301 GAGATACCGAGATTGGCTAAAGAAGCGGCGGAGAAGGCCATCAAGGAGTGGGGGCGGCCC 360
CURS1 301 GAGATACCGAGATTGGCTAAGGAAGCGGCGGAGAAGGCCATCAAGGAGTGGGGGCGGCCC 360
CzCURS1 361 AAATCGGAGATCACCCACCTGGTCTTCTGTTCCATCAGCGGGATCGACATGCCCGGCGCC 420
CURS1 361 AAATCGGAGATCACCCACCTGGTCTTCTGCTCCATCAGCGGGATCGACATGCCCGGCGCC 420
CzCURS1 421 GACTACCGCCTCGCCACGCTCCTCGGCCTCCCTCTCACCGTCAACCGCCTCATGATCTAC 480
CURS1 421 GACTACCGCCTCGCCACGCTCCTCGGGCTCCCTCTCACCGTCAACCGCCTCATGATCTAC 480
CzCURS1 481 AGCCAGGCCTGCCACATGGGCGCCGCCATGCTCCGCATCGCCAAGGACCTCGCCGAGAAC 540
CURS1 481 AGCCAGGCCTGCCACATGGGCGCCGCCATGCTCCGCATCGCCAAGGACCTCGCCGAGAAC 540
CzCURS1 541 AACAGGGGCGCGCGCGTGCTGGTGGTCGCCTGCGAGATCACCGTGCTCAGCTTCCGCGGC 600
CURS1 541 AACAGGGGCGCGCGCGTGCTGGTGGTCGCCTGCGAGATCACCGTGCTCAGCTTCCGCGGC 600
CzCURS1 601 CCGAACGAGGGCGACTTCGAGGCGCTCGCGGGGCAGGCCGGCTTCGGCGACGGCGCGGGG 660
CURS1 601 CCGAACGAGGGCGACTTCGAGGCGCTCGCGGGGCAGGCCGGCTTCGGCGACGGCGCGGGG 660
CzCURS1 661 GCCGTCGTCGTCGGGGCCGACCCGCTGGAAGGAATTGAAAAACCCATCTACGAGATCGCG 720
CURS1 661 GCCGTCGTCGTCGGGGCCGACCCGCTGGAAGGAATTGAAAAACCCATCTACGAGATCGCG 720
CzCURS1 721 GCGGCGATGCAGGAGACGGTGGCGGAGAGCCAGGGGGCGGTGGGCGGCCACCTGCGGGCG 780
CURS1 721 GCGGCGATGCAGGAGACGGTGGCGGAGAGCCAGGGGGCGGTGGGCGGCCACCTGCGGGCC 780
CzCURS1 781 TTCGGCTGGACGTTCTACTTCCTGAACCAGCTGCCGGCGATCATCGCCGACAACCTCGGG 840
CURS1 781 TTCGGCTGGACGTTCTACTTCCTGAACCAGCTGCCGGCGATCATCGCCGACAACCTCGGG 840
CzCURS1 841 AGGAGCCTGGAGCGGGCGTTGGCGCCGCTGGGGGTGACGGAGTGGAACGACGTCTTCTGG 900
CURS1 841 AGGAGCCTGGAGCGGGCGTTGGCGCCGCTGGGGGTGAGGGAGTGGAACGACGTCTTCTGG 900
CzCURS1 901 GTGGCGCACCCGGGCAACTGGGCCATCATGGACGCCATCGAAGCCAAGCTGCAGCTGAGC 960
CURS1 901 GTGGCGCACCCGGGCAACTGGGCCATCATTGACGCCATCGAAGCCAAGCTGCAGCTGAGC 960
CzCURS1 961 CCGGACAAGCTCAGCACCGCCCGCCACGTCTTCACAGAGTACGGCAACATGCAGAGCGCC 1020
CURS1 961 CCGGACAAGCTCAGCACCGCCCGCCACGTCTTCACAGAGTACGGCAACATGCAGAGCGCC 1020
CzCURS1 1021 ACCGTGTACTTCGTGATGGATGAGCTGAGGAAGCGGTCGGCGGTGGAGGGGCGGAGCACC 1080
CURS1 1021 ACCGTGTACTTCGTGATGGATGAGCTGAGGAAGCGGTCGGCGGTGGAGGGGCGGAGCACC 1080
CzCURS1 1081 ACCGGAGACGGCTTGCAGTGGGGAGTTCTCCTCGGTTTTGGGCCGGGCCTCAGCATCGAG 1140
CURS1 1081 ACCGGCGACGGCTTGCAGTGGGGAGTTCTCCTCGGTTTTGGGCCGGGCCTCAGCATCGAG 1140
CzCURS1 1141 ACCGTTGTACTGCGCAGTATGCCACTGTAG 1170
CURS1 1141 ACCGTTGTACTGCGCAGTATGCCACTGTAG 1170
Hình 3.9. So sánh trình tự nucleotide (nu) vùng CDS của gen CzCURS1 (MF402846)
ơ nghệ đen và CURS1 ơ nghệ vàng (AB495007.1). Số nu tương đồng: 1158/1170
(99%).

64
CzCURS2 1 ATGGCGATGATCAGCTTGCAGGCGATGCGCAAGGCGCAGAGAGCTCAAGGTCCGGCCACC 60
CURS2 1 ATGGCGATGATCAGCTTGCAGGCGATGCGCAAGGCGCAGAGAGCTCAAGGTCCGGCCACC 60
CzCURS2 61 ATCTTGGCCGTCGGCACCGCCAACCCGCCCAATCTCTACGAGCAGGACACGTATCCCGAC 120
CURS2 61 ATCTTGGCCGTCGGCACCGCCAACCCGCCCAATCTCTACGAGCAGGACACGTATCCCGAC 120
CzCURS2 121 TACTACTTCCGCGTCACCAACTCCGAGCACAGGCAGGAGCTCAAGAACAAGTTCCGCCGC 180
CURS2 121 TACTACTTCCGCGTCACCAACTCCGAGCACAAGCAGGAGCTCAAGAACAAGTTCCGCCTC 180
CzCURS2 181 ATGTGCGAGAAGACGATGGTGAAGAGGCGGTATCTTTACCTGACGCCGGAGATCCTGAAG 240
CURS2 181 ATGTGCGAGAAGACGATGGTGAAGAGGCGGTATCTTTACCTGACGCCGGAGATCCTGAAG 240
CzCURS2 241 GAGCGGCCGAAGCTGTGCTCGTACATGGAGCCGTCGTTCGACGACCGGCAGGACATCGTG 300
CURS2 241 GAGCGGCCGAAGCTGTGCTCGTACATGGAGCCGTCGTTCGACGACCGGCAGGACATCGTG 300
CzCURS2 301 GTGGAGGAGGTGCCGAAGCTGGCCGCGGAGGCAGCGGAGAAGGCCATCAAGGAGTGGGGC 360
CURS2 301 GTGGAGGAGGTGCCGAAGCTGGCCGCGGAGGCAGCGGAGAACGCCATCAAGGAGTGGGGC 360
CzCURS2 361 GGCGAGAAGTCGGCCATCACCCACCTGGTCTTCTGCTCCATCAGCGGCATCGACATGCCC 420
CURS2 361 GGCGACAAGTCCGCCATCACCCACCTGGTCTTCTGCTCCATCAGCGGCATCGACATGCCC 420
CzCURS2 421 GGAGCTGACTACCGCCTCGCCAAGCTCCTCGGGCTCCCGCTCGCCGTCAACCGCCTGATG 480
CURS2 421 GGAGCTGACTACCGCCTCGCCCAGCTCCTCGGACTCCCGCTCGCCGTCAACCGCCTGATG 480
CzCURS2 481 CTCTACAGCCAGGCCTGCCACATGGGCGCCGCCATGCTGCGCATAGCCAAGGACATCGCC 540
CURS2 481 CTCTACAGCCAGGCCTGCCACATGGGCGCCGCCATGCTGCGCATAGCCAAGGACATCGCC 540
CzCURS2 541 GAGAACAACCGCTCCGCTCGCGTCCTCGTCGTCGCCTGCGAGATCACCGTGCTCAGCTTC 600
CURS2 541 GAGAACAACCGCTCCGCGCGCGTCCTCGTCGTCGCCTGCGAGATCACCGTGCTCAGCTTC 600
CzCURS2 601 CGCGGCCCGGACGAGCGCGACTTCCAGGCGCTGGCCGGCCAGGCCGGCTTCGGGGACGGC 660
CURS2 601 CGCGGCCCGGACGAGCGCGACTTCCAGGCGCTGGCCGGCCAGGCCGGCTTCGGGGACGGC 660
CzCURS2 661 GCCGGCGCGATGATCGTCGGGGCCGACCCCGTCCTCGGCGTCGAGCGGCCGCTCTACCAC 720
CURS2 661 GCCGGCGCGATGATCGTCGGGGCCGACCCCGTCCTCGGCGTCGAGCGGCCGCTCTACCAC 720
CzCURS2 721 ATCATGTCGGCGACTCAGACGACGGTGCCGGAGAGCGAGAAGGCGGTGGGGGGCCACCTC 780
CURS2 721 ATCATGTCGGCGACTCAGACGACGGTACCGGAGAGCGAGAAGGCGGTGGGGGGCCACCTC 780
CzCURS2 781 CGCGAGGTGGGGCTGACCTTCCACTTCTTCAACCAGCTGCCGGCGATCATCGCCGACAAC 840
CURS2 781 CGCGAGGTGGGGCTGACCTTCCACTTCTTCAACCAGCTGCCGGCGATCATCGCCGACAAC 840
CzCURS2 841 GTGGGGAACAGCCTGGCGGAGGCGTTCGAACCGATCGGGATCAAGGACTGGAACAACATC 900
CURS2 841 GTGGGGAACAGCCTGGCGGAGGCGTTCGAACCGATCGGGATCAAGGACTGGAACAACATC 900
CzCURS2 901 TTCTGGGTGGCGCACCCGGGCAACTGGGCCATCATGGACGCCATCGAGACCAAGCTGGGC 960
CURS2 901 TTCTGGGTGGCGCACCCGGGCAACTGGGCCATCATGGACGCCATCGAGACCAAGCTGGGC 960
CzCURS2 961 CTGGAACAGAGCAAGCTGGCCACCGCACGCCACGTCTTCTCCGAGTTCGGCAACATGCAG 1020
CURS2 961 CTGGAACAGAGCAAGCTGGCCACCGCACGCCACGTCTTCTCCGAGTTCGGCAACATGCAG 1020
CzCURS2 1021 AGCGCCACCGTCTACTTCGTGATGGACGAGCTCAGGAAACGGTCGGCGGCGGAGAACCGG 1080
CURS2 1021 AGCGCCACCGTCTACTTCGTGATGGACGAGCTCAGGAAACGGTCGGCGGCGGAGAACCGG 1080
CzCURS2 1081 GCGACCACCGGCGACGGGCTCCGGTGGGGCGTGCTCTTCGGCTTCGGCCCCGGCATCAGC 1140
CURS2 1081 GCGACCACCGGCGACGGGCTCCGGTGGGGCGTGCTCTTCGGCTTCGGCCCGGGCATCAGC 1140
CzCURS2 1141 ATCGAAACCGTCGTGCTCCAAAGCGTGCCGCTTTAG 1176
CURS2 1141 ATCGAAACCGTCGTGCTCCAAAGCGTGCCGCTTTAG 1176
Hình 3.10. So sánh trình tự nucleotide (nu) vùng CDS của gen CzCURS2
(MF402846) ơ nghệ đen và CURS2 ơ nghệ vàng (AB506762.1). Số nu tương đồng:
1166/1176 (99%).

65
CzCURS3 1 ATGGGCAGCCTGCAGGCGATTCGCAGAGCACAGCGGGCTCAAGGCCCGGCCACCATCATG 60
CURS3 1 ATGGGCAGCCTGCAGGCGATGCGCAGGGCACAGCGGGCTCAAGGCCCGGCCACCATCATG 60
CzCURS3 61 GCTGTCGGCACCTCCAACCCGCCTAACCTCTACGAGCAGACCTCCTACCCCGACTTCTAT 120
CURS3 61 GCTGTCGGCACCTCCAACCCGCCTAACCTCTACGAGCAGACCTCCTACCCCGACTTCTAT 120
CzCURS3 121 TTCCGCGTCACCAACTCCGACCACAAGCACGAGCTCAAGAACAAATTCCGTGTTATCTGT 180
CURS3 121 TTCCGCGTCACCAACTCCGACCACAAGCACGCGCTCAAGAACAAATTCCGTGTTATCTGT 180
CzCURS3 181 GAGAAGACGAAGGTGAAAAGACGGTACTTGCACTTGACGGAGGAGATCCTGAAGCAGAGG 240
CURS3 181 GAGAAGACGAAGGTGAAAAGACGGTACTTGCACTTGACGGAGGAGATCCTGAAGCAGAGG 240
CzCURS3 241 CCCAAGCTCTGCTCCTACATGGAGCCCTCCTTCGACGACCGGCAGGACATCGTGGTGGAG 300
CURS3 241 CCCAAGCTCTGCTCCTACATGGAGCCCTCCTTCGACGATCGGCAGGACATCGTGGTGGAG 300
CzCURS3 301 GAGATACCGAAGCTGGCGAAGGAGGCGGCGGAAAAGGCGATCAAGGAATGGGGCCGCCCC 360
CURS3 301 GAGATACCAAAGCTGGCGAAGGAGGCGGCGGAGAAGGCGATCAAGGAGTGGGGCCGCCCC 360
CzCURS3 361 AAGTCGGAGATCACCCACTTGGTGTTCTGCTCCATCAGCGGYATCGACATGCCCGGCGCC 420
CURS3 361 AAGTCGGAGATCACCCACTTGGTGTTCTGCTCCATCAGCGGTATCGACATGCCCGGCGCC 420
CzCURS3 421 GATTACCGCCTCGCCACGCTCCTCGGCCTCCCCCTGTCCGTCAACCGCCTCATGCTCTAC 480
CURS3 421 GATTACCGCCTCGCCACCCTCCTCGGCCTCCCCCTGTCCGTCAACCGCCTCATGCTCTAC 480
CzCURS3 481 AGCCAGGCCTGCCACATGGGCGCGCAGATGCTGCGCATAGCCAAGGACCTCGCGGAGAAC 540
CURS3 481 AGCCAGGCCTGCCACATGGGCGCGCAGATGCTGCGCATAGCCAAGGACCTCGCGGAGAAC 540
CzCURS3 541 AACCGGGGCGCGCGCGTCCTGGCCGTCTCCTGCGAAATCACCGTGTTCAGCTTCCGCGGT 600
CURS3 541 AACCGGGGCGCGCGCGTCCTGGCCGTCTCCTGCGAAATCACCGTGCTCAGCTTCCGCGGT 600
CzCURS3 601 CCCGACGCGGGCGACTTCGAGGCCCTCGCGTGTCAGGCCGGCTTCGGCGACGGTGCCGCT 660
CURS3 601 CCGGACGCGGGCGACTTCGAGGCCCTCGCGTGTCAGGCCGGCTTCGGCGACGGTGCCGCT 660
CzCURS3 661 GCCGTCGTCGTCGGGGCCGACCCCCTCCCGGGCGTCGAGAGGCCCATCTACGAGATCGCG 720
CURS3 661 GCCGTCGTCGTCGGGGCCGACCCCCTCCCGGGCGTCGAGAGGCCCATCTACGAGATCGCG 720
CzCURS3 721 GCGGCGATGCAGGAAACGGTGCCGGAGAGCGAGAGGGCGGTGGGGGGCCACCTGAGGGAA 780
CURS3 721 GCGGCGATGCAGGAAACGGTGCCGGAGAGCGAGAGGGCGGTGGGGGGCCACCTGAGGGAA 780
CzCURS3 781 ATCGGCTGGACCTTCCACTTCTTCAACCAGCTGCCGAAGCTGATCGCGGAAAACATCGAG 840
CURS3 781 ATCGGCTGGACCTTCCACTTCTTCAACCAGCTTCCGAAGCTGATCGCGGAAAACATCGAG 840
CzCURS3 841 GGCAGCCTGGCGCGGGCGTTCAAGCCGCTGGGGATCAGCGAGTGGAACGACGTGTTCTGG 900
CURS3 841 GGCAGCCTGGCGCGGGCGTTCAAGCCGCTGGGGATCAGCGAGTGGAACGACGTGTTCTGG 900
CzCURS3 901 GTGGCGCACCCGGGGAATTGGGGCATCATGGACGCCATCGAGACCAAGCTGGGGCTGGAA 960
CURS3 901 GTGGCGCACCCGGGGAACTGGGGCATCATGGACGCCATCGAGACCAAGCTGGGGCTGGAA 960
CzCURS3 961 CAGGGGAAGCTCGCCACGGCGCGCCACGTCTTCAGCGAGTACGGAAACATGCAGAGCGCC 1020
CURS3 961 CAGGGGAAGCTCGCCACGGCGCGCCACGTCTTCAGCGAGTACGGAAACATGCAGAGCGCC 1020
CzCURS3 1021 ACCGTGTACTTCGTGATGGACGAGGTGAGGAAGCGGTCGGCGGCGGAGGGGCGGGCCACC 1080
CURS3 1021 ACCGTGTACTTCGTGATGGACGAGGTGAGGAAGCGGTCGGCGGCGGAGGGGCGGGCCACC 1080
CzCURS3 1081 ACCGGCGAAGGCCTGGAGTGGGGAGTGCTGTTTGGGTTTGGCCCAGGCCTCACCATAGAA 1140
CURS3 1081 ACCGGCGAAGGCCTGGAGTGGGGAGTGCTGTTTGGGTTTGGCCCAGGCCTCACCATAGAA 1140
CzCURS3 1141 ACTGTCGTGCTACGCAGTGTACCATTACCGTAG 1173
CURS3 1141 ACTGTCGTGCTACGCAGTGTACCATTACCGTAG 1173
Hình 3.11. So sánh trình tự nucleotide (nu) vùng CDS của gen CzCURS3 (NCBI:
MF987835) ơ nghệ đen và CURS3 ơ nghệ vàng (NCBI: AB506763.1). Số nu tương
đồng: 1160/1173 (99%).

66
CzDCS 1 MEANGYRITHSADGPATILAIGTANPTNVVDQNAYPDFYFRVTNSEHLQELKAKFRRICE 60
DCS 1 MEANGYRITHSADGPATILAIGTANPTNVVDQNAYPDFYFRVTNSEYLQELKAKFRRICE 60
CzDCS 61 KAAIRKRHLYLTEEILRENPSLLAPMAPSFDARQAIVVEAVPKLAKEAAEKAIKEWGRPK 120
DCS 61 KAAIRKRHLYLTEEILRENPSLLAPMAPSFDARQAIVVEAVPKLAKEAAEKAIKEWGRPK 120
CzDCS 121 SDITHLVFCSASGIDMPGSDLQLLKLLGLPPSVNRVMLYNVGCHAGGTALRVAKDLAENN 180
DCS 121 SDITHLVFCSASGIDMPGSDLQLLKLLGLPPSVNRVMLYNVGCHAGGTALRVAKDLAENN 180
CzDCS 181 RGARVLAVCSEVTVLSYRGPHPAHIESLFVQALFGDGAAALVVGSDPVDGVERPIFEIAS 240
DCS 181 RGARVLAVCSEVTVLSYRGPHPAHIESLFVQALFGDGAAALVVGSDPVDGVERPIFEIAS 240
CzDCS 241 ASQVMLPESEEAVGGHLREIGLTFHLKSQLPSIIASNIEQSLTTACSPLGLSDWNQLFWA 300
DCS 241 ASQVMLPESAEAVGGHLREIGLTFHLKSQLPSIIASNIEQSLTTACSPLGLSDWNQLFWA 300
CzDCS 301 VHPGGRAILDQVEARLGLEKDRLAATRHVLSEYGNMQSATVLFILDEMRNRSAAEGHATT 360
DCS 301 VHPGGRAILDQVEARLGLEKDRLAATRHVLSEYGNMQSATVLFILDEMRNRSAAEGHATT 360
CzDCS 361 GEGLDWGVLLGFGPGLSIETVVLHSCRLN 389
DCS 361 GEGLDWGVLLGFGPGLSIETVVLHSCRLN 389
Hình 3.12. So sánh trình tự amino acid suy diễn của 2 gen CzDCS và DCS
(C0SVZ5.1). Hộp màu đen chỉ các amino acid khác nhau, chú thích này dùng chung
cho các hình 3.13-3.15.
CzCURS1 1 MANLHALRREQRAQGPATIMAIGTATPPNLYEQSTFPDFYFRVTNSDDKQELKKKFRRMC 60
CURS1 1 MANLHALRREQRAQGPATIMAIGTATPPNLYEQSTFPDFYFRVTNSDDKQELKKKFRRMC 60
CzCURS1 61 DKTMVKKRYLHLTEEILKERPKLCSYKEASFDDRQDIVVEEIPRLAKEAAEKAIKEWGRP 120
CURS1 61 EKTMVKKRYLHLTEEILKERPKLCSYKEASFDDRQDIVVEEIPRLAKEAAEKAIKEWGRP 120
CzCURS1 121 KSEITHLVFCSISGIDMPGADYRLATLLGLPLTVNRLMIYSQACHMGAAMLRIAKDLAEN 180
CURS1 121 KSEITHLVFCSISGIDMPGADYRLATLLGLPLTVNRLMIYSQACHMGAAMLRIAKDLAEN 180
CzCURS1 181 NRGARVLVVACEITVLSFRGPNEGDFEALAGQAGFGDGAGAVVVGADPLEGIEKPIYEIA 240
CURS1 181 NRGARVLVVACEITVLSFRGPNEGDFEALAGQAGFGDGAGAVVVGADPLEGIEKPIYEIA 240
CzCURS1 241 AAMQETVAESQGAVGGHLRAFGWTFYFLNQLPAIIADNLGRSLERALAPLGVTEWNDVFW 300
CURS1 241 AAMQETVAESQGAVGGHLRAFGWTFYFLNQLPAIIADNLGRSLERALAPLGVREWNDVFW 300
CzCURS1 301 VAHPGNWAIMDAIEAKLQLSPDKLSTARHVFTEYGNMQSATVYFVMDELRKRSAVEGRST 360
CURS1 301 VAHPGNWAIMDAIEAKLQLSPDKLSTARHVFTEYGNMQSATVYFVMDELRKRSAVEGRST 360
CzCURS1 361 TGDGLQWGVLLGFGPGLSIETVVLRSMPL 389
CURS1 361 TGDGLQWGVLLGFGPGLSIETVVLRSMPL 389
Hình 3.13. So sánh trình tự amino acid suy diễn của 2 gen CzCURS1 và CURS1
(AJF45913.1).

67
CzCURS2 1 MAMISLQAMRKAQRAQGPATILAVGTANPPNLYEQDTYPDYYFRVTNSEHRQELKNKFRR 60
CURS2 1 MAMISLQAMRKAQRAQGPATILAVGTANPPNLYEQDTYPDYYFRVTNSEHKQELKNKFRR 60
CzCURS2 61 MCEKTMVKRRYLYLTPEILKERPKLCSYMEPSFDDRQDIVVEEVPKLAAEAAEKAIKEWG 120
CURS2 61 MCEKTMVKRRYLYLTPEILKERPKLCSYMEPSFDDRQDIVVEEVPKLAAEAAEKAIKEWG 120
CzCURS2 121 GEKSAITHLVFCSISGIDMPGADYRLAKLLGLPLAVNRLMLYSQACHMGAAMLRIAKDIA 180
CURS2 121 GDKSAITHLVFCSISGIDMPGADYRLAKLLGLPLAVNHLMLYSQACHMGAAMLRIAKDIA 180
CzCURS2 181 ENNRSARVLVVACEITVLSFRGPDERDFQALAGQAGFGDGAGAMIVGADPVLGVERPLYH 240
CURS2 181 ENNRSARVLVVACEITVLSFRGPDERDFQALAGQAGFGDGAGAMIAGADPVLGVERPLYH 240
CzCURS2 241 IMSATQTTVPESEKAVGGHLREVGLTFHFFNQLPAIIADNVGNSLAEAFEPIGIKDWNNI 300
CURS2 241 IMSATQTTVPESEKAVGGHLREVGLTFHFFNQLPAIIADNVGNSLAEAFEPIGIKDWNNI 300
CzCURS2 301 FWVAHPGNWAIMDAIETKLGLEQSKLATARHVFSEFGNMQSATVYFVMDELRKRSAAENR 360
CURS2 301 FWVAHPGNWAIMDAIETKLGLEQSKLATARHVFSEFGNMQSATVYFVMDELRKRSAAENR 360
CzCURS2 361 ATTGDGLRWGVLFGFGPGISIETVVLQSVPL 391
CURS2 361 ATTGDGLRWGVLFGFGPGISIETVVLQSVPL 391
Hình 3.14. So sánh trình tự amino acid suy diễn của 2 gen CzCURS2 và CURS2
(BAW81545.1).
CzCURS3 1 MGSLQAIRRAQRAQGPATIMAVGTSNPPNLYEQTSYPDFYFRVTNSDHKHELKNKFRVIC 60
CURS3 1 MGSLQAMRRAQRAQGPATIMAVGTSNPPNLYEQTSYPDFYFRVTNSDHKHELKNKFRVIC 60
CzCURS3 61 EKTKVKRRYLHLTEEILKQRPKLCSYMEPSFDDRQDIVVEEIPKLAKEAAEKAIKEWGRP 120
CURS3 61 EKTKVKRRYLHLTEEILKQRPKLCSYMEPSFDDRQDIVVEEIPKLAKEAAEKAIKEWGRP 120
CzCURS3 121 KSEITHLVFCSISXIDMPGADYRLATLLGLPLSVNRLMLYSQACHMGAQMLRIAKDLAEN 180
CURS3 121 KSEITHLVFCSISGIDMPGADYRLATLLGLPLSVNRLMLYSQACHMGAQMLRIAKDLAEN 180
CzCURS3 181 NRGARVLAVSCEITVFSFRGPDAGDFEALACQAGFGDGAAAVVVGADPLPGVERPIYEIA 240
CURS3 181 NRGARVLAVSCEITVLSFRGPDAGDFEALACQAGFGDGAAAVVVGADPLPGVERPIYEIA 240
CzCURS3 241 AAMQETVPESERAVGGHLREIGWTFHFFNQLPKLIAENIEGSLARAFKPLGISEWNDVFW 300
CURS3 241 AAMQETVPESERAVGGHLREIGWTFHFFNQLPKLIAENIEGSLARAFKPLGISEWNDVFW 300
CzCURS3 301 VAHPGNWGIMDAIETKLGLEQGKLATARHVFSEYGNMQSATVYFVMDEVRKRSAAEGRAT 360
CURS3 301 VAHPGNWGIMDAIETKLGLEQGKLATARHVFSEYGNMQSATVYFVMDEVRKRSAAEGRAT 360
CzCURS3 361 TGEGLEWGVLFGFGPGLTIETVVLRSVPLP 390
CURS3 361 TGEGLEWGVLFGFGPGLTIETVVLRSVPLP 390
Hình 3.15. So sánh trình tự amino acid suy diễn của 2 gen CzCURS3 và CURS3
(AJF45914.1).
Do có sự tương đồng cao giữa các gen tổng hợp curcuminoid của nghệ
đen và nghệ vàng, chúng tôi đã xây dựng cây phả hệ cho các gen này. Vùng
CDS của các gen DCS, CURS1, CURS2 và CURS3 từ nghệ đen và nghệ vàng,
gen mã hóa enzyme type III polyketide synthase từ M. accuminata và gen mã

68
hóa enzyme polyketide synthase từ nấm A. alternate (nhóm ngoại) cũng được
sử dụng. Việc xây dựng cây phả hệ được thực hiện bằng phần mềm MEGA7
theo phương pháp Maximum Likelihood, độ lặp lại (bootstrap) là 2.000 lần.
Kết quả trình bày ơ hình 3.16 cho thấy nhóm gen nghiên cứu có giá trị bootstrap
rất cao, thể hiện đặc tính cùng nguồn gốc và chức năng (orthologs and paralogs)
của các gen này giữa 2 loài nghệ. Trên cây phả hệ, nhóm các gen CURS tạo
thành 1 nhóm với độ tương đồng rất cao, mức độ sai khác giữa các gen chỉ từ
0,1-0,2. Các gen DCS cũng có độ tương đồng khá cao và tạo thành một nhóm
trên cây phả hệ. Kết quả so sánh với gen PKS type III từ cây chuối (nhóm nội)
cũng cho thấy có sự tương đồng cao với 2 nhóm gen trên, sự khác biệt giữa các
gen này chưa tới 0,5. Trong khi đó, so sánh với gen PKS của nấm A. alternate
(nhóm ngoại) cho thấy chúng có sự khác biệt lớn hơn, mức độ sai khác là
khoảng 1,5, thể hiện nguồn gốc khác nhau của chúng. Kết quả so sánh này cho
thấy dù là các gen có cùng tính năng, nhưng sự sai khác thể hiện rất rõ ơ các
loài có họ hàng xa nhau, trong khi các loài gần nhau thì sự tương đồng rất cao.
Hình 3.16. Cây phả hệ của các gen tổng hợp curcuminoid.
3.2.2. Phân tích biểu hiện của các gen tổng hợp curcuminoid

69
Để nghiên cứu sự biểu hiện của các gen tổng hợp curcuminoid trong 2 cấu
trúc khác nhau của nghệ đen (củ và callus), chúng tôi sử dụng phương pháp
RT-PCR sau đó xác định độ sáng của băng DNA trên hình ảnh điện di (PCR
định tính: non-quantitative RT-PCR). Do các primer được thiết kế để khuếch
đại các gen hoàn chỉnh (full-length) từ genome không cho kết quả tốt nhất trong
trường hợp này, chúng tôi đã thiết kế primer để khuếch đại các đoạn DNA ngắn
hơn (vùng đặc hiệu cho gen) (Hình 3.8-3.11). Các primer được thiết kế dựa trên
trình tự nucleotide của vùng có sự tương đồng thấp giữa các gen để đảm bảo
tính đặc hiệu và dễ dàng phân tích kết quả.
Phân tích dữ liệu RT-PCR của 4 gen CzDCS, CzCURS1, CzCURS2 và
CzCURS3 ơ củ và callus cho thấy tất cả chúng đều có sản phẩm khuếch đại,
chứng tỏ các gen này đều biểu hiện ơ cả 2 loại mô nói trên (Hình 3.17). Kết quả
này còn minh chứng cho nhận định cơ chế sinh tổng hợp curcuminoid của nghệ
đen cũng xuất hiện trong quá trình nuôi cấy in vitro, và callus là nguyên liệu
thích hợp để phát triển hệ thống nuôi cấy tế bào nhằm mục đích sản xuất
curcumin. Hình ảnh điện di cho thấy sản phẩm PCR tương ứng với các gen
CzDCS, CzCURS1, CzCURS2 và CzCURS3 có kích thước lần lượt khoảng 272
bp, 286 bp, 211 bp và 202 bp phù hợp với tính toán (Bảng 2.2 và Hình 3.8-
3.11). Xem xét mức độ biểu hiện của các gen này trong 2 loại mô khác nhau,
chúng tôi nhận thấy mức độ biểu hiện của các gen trong củ đều cao hơn trong
callus, ngoại trừ gen CURS1.
Để củng cố nhận định trên, chúng tôi đã phân tích hàm lượng curcumin,
một thành phần chính của nhóm curcuminoid, ơ các mẫu trên bằng HPLC. Kết
quả cho thấy cả hai dịch chiết từ củ và callus đều có chứa peak curcumin với
thời gian lưu tương tự như curcumin chuẩn (Hình 3.18-3.20). Tuy nhiên, diện
tích và số lượng peak xuất hiện trong các hình 3.19 và 3.20 cho thấy đã có sự
chuyển hóa sinh học diễn ra trong quá trình nuôi cấy callus phù hợp với nhận
định ơ những nghiên cứu đã công bố trước đây [82], [132]. Curcumin được tìm

70
thấy trong callus có thể liên quan đến sự biểu hiện của các gen tổng hợp
curcuminoid ơ nghệ đen như trong phân tích RT-PCR (Hình 3.17). Nhận định
này sẽ được củng cố thêm ơ phần sau (mục 3.3).
Hình 3.17. Phân tích RT-PCR vùng đặc hiệu của các gen CzDCS, CzCURS1,
CzCURS2 và CzCURS3 trong 2 loại mô khác nhau của nghệ đen. M: DNA size
marker (1 kb), 1-3-5-7: củ, 2-4-6-8: callus, A: CzDCS, B: CzCURS1, C: CzCURS2,
và D: CzCURS3.
Bảng 3.7. Mật độ băng DNA từ phân tích RT-PCR vùng đặc hiệu của các gen CzDCS,
CzCURS1, CzCURS2 và CzCURS3 trong 2 loại mô khác nhau của nghệ đen
Gen Mẫu Đường
chạy
Mật độ Tỷ lệ giữa củ/callus
CzDCS Củ 1 4061,3 1,8
Callus 2 2231,9
CzCURS1 Củ 3 2288,1 0,6
Callus 4 3946,7
CzCURS2 Củ 5 5018,3 1,9
Callus 6 2641,9
CzCURS3 Củ 7 4684,5 1,4
Callus 8 3281,8
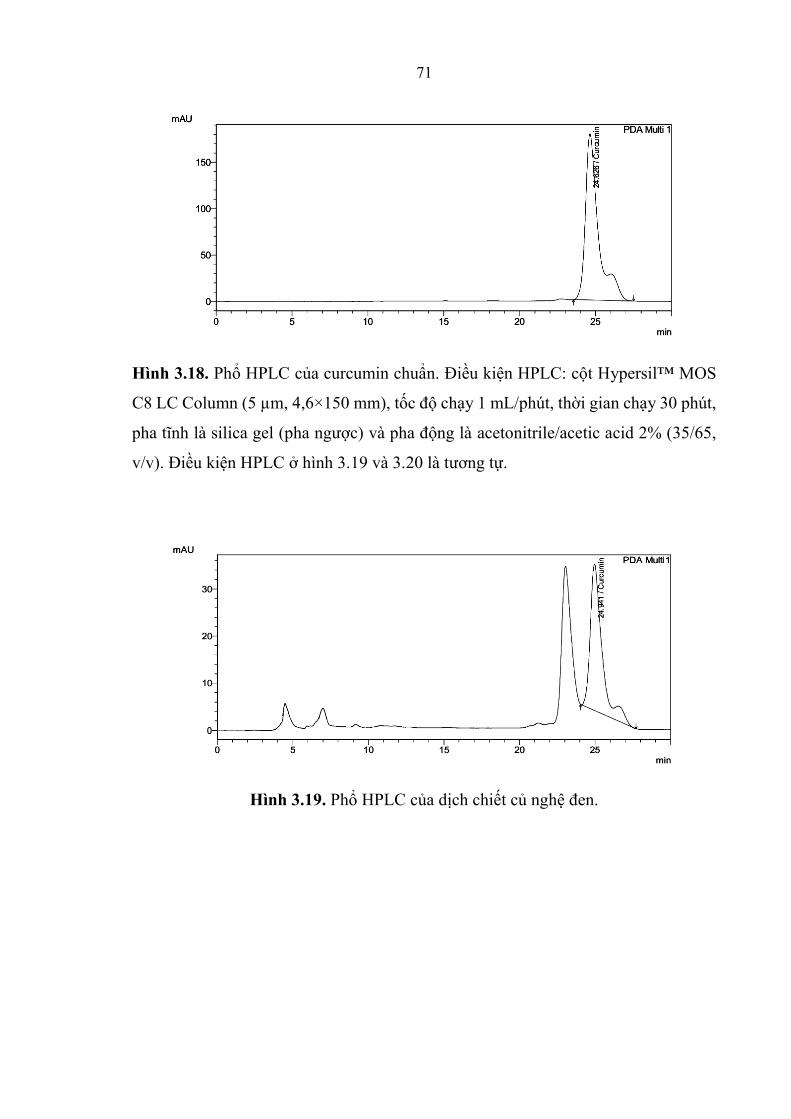
71
Hình 3.18. Phổ HPLC của curcumin chuẩn. Điều kiện HPLC: cột Hypersil™ MOS
C8 LC Column (5 µm, 4,6×150 mm), tốc độ chạy 1 mL/phút, thời gian chạy 30 phút,
pha tĩnh là silica gel (pha ngược) và pha động là acetonitrile/acetic acid 2% (35/65,
v/v). Điều kiện HPLC ơ hình 3.19 và 3.20 là tương tự.
Hình 3.19. Phổ HPLC của dịch chiết củ nghệ đen.

72
Hình 3.20. Phổ HPLC của dịch chiết callus nghệ đen.
3.3. ẢNH HƯỞNG CỦA ELICITOR LÊN QUÁ TRÌNH SINH
TỔNG HỢP CURCUMINOID
3.3.1. Thăm dò ảnh hưởng của các elicitor lên biểu hiện của các gen tổng
hợp curcuminoid
Trong các gen CURS tham gia vào con đường sinh tổng hợp curcuminoid,
gen CURS1 đóng vai trò chính trong quá trình sinh tổng hợp curcumin trong
khi gen CURS3 có vai trò lớn hơn trong quá trình tổng hợp
bisdemethoxycurcumin (Hình 1.3). Vì vậy, để giảm bớt số mẫu cần phân tích,
chúng tôi đã phân tích sơ bộ mức độ biểu hiện của 2 gen này khi được xử lý
các loại elicitor khác nhau với các nồng độ khác nhau. Tế bào nghệ đen 10 ngày
tuổi ơ vào thời điểm gần kết thúc pha sinh trương (pha logarithm) (Hình 3.3)
được chuyển sang nuôi cấy trên môi trường mới có thành phần tương tự nhưng
có bổ sung các elicitor khác nhau (SA, MeJA và YE) với các nồng độ khác
nhau, sau đó thu hoạch sinh khối vào ngày thứ 14 (kết thúc pha sinh trương) để
đánh giá mức độ biểu hiện của các gen tổng hợp curcuminoid và hàm lượng
curcumin đã được tích lũy.
Kết quả phân tích RT-PCR cho thấy phần lớn các công thức xử lý elicitor
mức độ biểu hiện của 2 gen CURS1 và CURS3 đều có thay đổi so với đối chứng
không xử lý. Khi xử lý MeJA (25-150 mM), mức độ biểu hiện của gen CURS1

73
gần như không thay đổi so với đối chứng (Hình 3.22A) trong khi CURS3 có
chiều hướng tăng lên, tuy nhiên mức độ tăng lên không nhiều (Hình 3.22B).
Khi xử lý salicilic acid (50-150 mM), mức độ biểu hiện của 2 gen này đều tăng
so với đối chứng, trong đó gen CURS1 tăng mạnh nhất khi xử lý ơ nồng độ 100
mM trong khi gen CURS3 mạnh nhất ơ nồng độ 150 mM. Trong khi đó, khi xử
lý bằng dịch chiết nấm men, mức độ biểu hiện của các gen chỉ tăng ơ nồng độ
0,7-1,0 g/L, các công thức xử lý còn lại các gen đều có mức độ biểu hiện thấp
hơn so với đối chứng.
Từ các kết quả sơ bộ, chúng tôi nhận thấy MeJA ít có ảnh hương lên sự
biểu hiện của các gen CURS, trong khi đó salicilic acid và dịch chiết nấm men
có ảnh hương lớn đến mức độ biểu hiện của các gen này. Xét trên vai trò của
các gen, chúng tôi nhận thấy CURS1 có vai trò lớn hơn các gen khác trong quá
trình sinh tổng hợp curcuminoid, vì vậy chúng tôi lựa chọn các công thức xử lý
cho mức độ biểu hiện của gen CURS1 cao nhất. Các nồng độ xử lý elicitor được
lựa chọn là salicilic acid 100 mM và dịch chiết nấm men 1 g/L, các công thức
xử lý này sẽ được sử dụng để đánh giá mức độ biểu hiện của tất cả các gen ơ
các thời điểm xử lý khác nhau về sau.
Hình 3.21. RNA tổng số của các mẫu tế bào sau khi được xử lý elicitor. M: DNA
size marker (1 kb), MeJA. Methyl jasmonate, SA. Salicilic acid, YE. Dịch chiết nấm
men. ĐC. Tế bào không xử lý elicitor.
-------MeJA-------- ------------SA------------- ----------YE---------------
M ĐC 25 50 100 150 50 70 100 150 0,1 0,5 0,7 1,0 M

74
Hình 3.22. Phân tích RT-PCR vùng đặc hiệu của các gen CzCURS1 (A) và CzCURS3
(B) sau khi được xử lý elicitor. M: DNA size marker (1 kb), MeJA. Methyl jasmonate,
SA. Salicilic acid, YE. Dịch chiết nấm men. ĐC. Tế bào không xử lý elicitor. PC. Đối
chứng dương tính (gen CzCURS1 và CzCURS3)
3.3.2. Biểu hiện của gen tổng hợp curcuminoid
Kết quả nghiên cứu đường cong sinh trương cho thấy tế bào nghệ đen đạt
sinh trương cực đại sau 14 ngày nuôi cấy, vì vậy, các thời điểm được chúng tôi
lựa chọn để bổ sung elicitor vào môi trường là lúc bắt đầu nuôi cấy và thời điểm
5 ngày, là thời điểm tế bào bào đầu sinh trương mạnh ơ đầu pha sinh trương
(pha log). Trong nghiên cứu này, YE 1 g/L và SA 100 µM và MeJA được bổ
sung vào môi trường tại 2 thời điểm trên. Kết quả phân tích hình ảnh điện di
các sản phẩm RT-PCR cho thấy sự biểu hiện của các gen ơ tế bào nghệ đen đã
được xử lý elicitor mạnh hơn đối chứng là tế bào không xử lý (Hình 3.23). Mức
độ biểu hiện của các gen này được đánh giá thông qua độ sáng của các băng
DNA đã cho thấy các tế bào có xử lý cao hơn khoảng 1,14-3,64 lần so với
không xử lý (Bảng 3.8).
-------MeJA-------- ------------SA------------- ----------YE---------------
M ĐC 25 50 100 150 50 70 100 150 0,1 0,5 0,7 1,0 1,5 PC
A
B

75
Nhìn chung, YE có tác dụng kích thích tốt hơn so với SA, mật độ tổng số
của các băng DNA đạt 71.528 so với 31.625 (hơn khoảng 2,3 lần). Số liệu ơ
bảng 3.8 còn cho thấy SA được bổ sung vào ngày thứ 5 đã tăng khả năng biểu
hiện của tất cả các gen so với việc bổ sung vào thời điểm ban đầu. Trong khi
đó, xử lý YE cho kết quả khác, các gen CzCURS1 và CzCURS3 biểu hiện mạnh
khi xử lý ơ thời điểm ban đầu, trong khi các gen CzDCS và CzCURS2 thì ngược
lại, biểu hiện mạnh khi xử lý ơ thời điểm 5 ngày. Xử lý YE ơ thời điểm ban đầu
của nuôi cấy thu được mật độ tổng số của cả 4 gen cao hơn khoảng 1,3 lần so
với khi bổ sung vào ngày thứ 5.
Hình 3.23. Sản phẩm RT-PCR vùng đặc hiệu của các gen tổng hợp curcuminoid ơ
nghệ đen. A: CzDCS, B-D: CzCURS1-3, E: RNA tổng số. 1: đối chứng (tế bào không
xử lý elicitor), 2 và 4: tế bào được xử lý SA và YE tại thời điểm ban đầu. 3 và 5: tế
bào được xử lý SA và YE vào ngày thứ 5 của quá trình nuôi cấy. M: DNA size marker
(1 kb DNA Ladder).

76
Bảng 3.8. Mật độ các băng DNA của vùng đặc hiệu ơ các gen tổng hợp curcuminoid.
Gen Đôi
chứng
SA (100 µM) YE (1 g/L)
0 5 0 5
CzDCS 4706 5312 6146 5113 13425
CzCURS1 6881 7810 10766 32576 13524
CzCURS2 4646 6186 8510 12139 17783
CzCURS3 4414 4146 6203 21700 12664
Tổng số 20647 23455 31625 71528 57397
Tăng so với ĐC 1,00 1,14 1,53 3,46 2,78
0 và 5 là ký hiệu elicitor được bổ sung vào môi trường ơ thời điểm ban đầu hoặc ngày
thứ 5 của quá trình nuôi cấy.
3.3.3. Tích lũy curcumin
Kết quả trình bày ơ bảng 3.9 cho thấy sinh trương của tế bào nghệ đen ơ
tất cả các công thức được xử lý elicitor đều bị ức chế đáng kể, sinh khối tươi
thu được cao nhất chỉ đạt 105,20 g khối lượng tươi/L (9,80 g khối lượng khô)
so với 154,20 g khối lượng tươi/L (12,80 g khối lượng khô) ơ đối chứng. Tuy
nhiên, có sự tương quan giữa mức độ biểu hiện của các gen khi phân tích RT-
PCR với sự tích lũy curcumin khi phân tích bằng HPLC. Ảnh hương của YE
lên khả năng sản xuất curcumin cao hơn so với khi xử lý bằng SA và đối chứng.
Hàm lượng curcumin thu được khi xử lý SA và YE vào thời điểm 5 ngày nuôi
cấy cao hơn khoảng 1,2 đến 1,6 lần so với khi bổ sung vào thời điểm ban đầu,
tương ứng đạt 1,32 với 1,14 mg/g khối lượng khô khi xử lý SA và 1,44 với 0,92
mg/g khối lượng khô khi xử lý YE. Ở trường hợp xử lý bằng YE, hàm lượng
curcumin thu được có sự sai khác so với mức độ biểu hiện của các gen. Cường
độ tổng số của các băng DNA trong phân tích RT-PCR tại thời điểm bổ sung
ban đầu cao hơn khi bổ sung vào ngày thứ 5 (Bảng 3.8), trong khi đó hàm lượng
curcumin khi bổ sung YE vào ngày thứ 5 lại cao hơn (Bảng 3.9). Nguyên nhân
có thể do các gen này ngoài việc tham gia quá trình sinh tổng hợp curcumin

77
còn tham gia tổng hợp demethoxycurcumin và bisdemethoxycurcumin.
Bảng 3.9. Ảnh hương của các elicitor lên sinh trương và tích lũy curcumin trong tế
bào nghệ đen
Elicitor
Sinh trưởng tế bao Curcumin (mg/g
khô) Khôi lượng tươi
(g/L)
Khôi lượng khô
(g/L)
YE(1) 105,20b 9,80b 0,92d
YE(2) 97,6c 8,40c 1,44a
SA(1) 69,00d 6,20d 1,14c
SA(2) 63,20d 6,00d 1,32b
Đối chứng (I) 154,2a 12,80a 0,57f
Đối chứng (II) - - 0,78e
Elicitor được bổ sung vào môi trường nuôi cấy ơ thời điểm ban đầu (1) hoặc sau 5
ngày (2). Đối chứng (I): tế bào không xử lý elicitor. Đối chứng (II): củ nghệ đen 10
tháng tuổi.
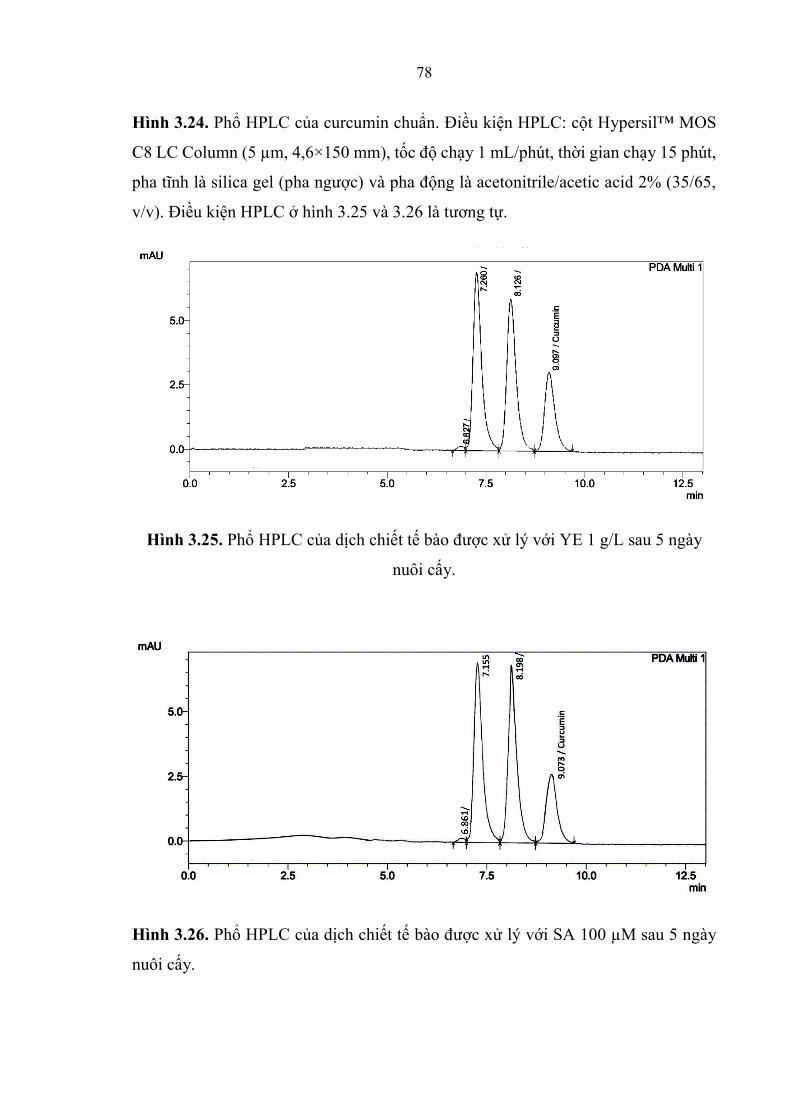
78
Hình 3.24. Phổ HPLC của curcumin chuẩn. Điều kiện HPLC: cột Hypersil™ MOS
C8 LC Column (5 µm, 4,6×150 mm), tốc độ chạy 1 mL/phút, thời gian chạy 15 phút,
pha tĩnh là silica gel (pha ngược) và pha động là acetonitrile/acetic acid 2% (35/65,
v/v). Điều kiện HPLC ơ hình 3.25 và 3.26 là tương tự.
Hình 3.25. Phổ HPLC của dịch chiết tế bào được xử lý với YE 1 g/L sau 5 ngày
nuôi cấy.
Hình 3.26. Phổ HPLC của dịch chiết tế bào được xử lý với SA 100 µM sau 5 ngày
nuôi cấy.

79
Chương 4
BÀN LUẬN
4.1. THIẾT LẬP NUÔI CẤY TẾ BÀO
Trong tự nhiên, nghệ đen là loài nhân giống bằng củ (thân rễ, rhizome),
do phải mất một thời gian dài để tạo củ nên hệ số nhân kém; năng suất thu
hoạch thường thấp, đặc biệt là phụ thuộc rất lớn vào điều kiện đất đai, khí hậu
và mùa vụ nên khó có đủ nguồn nguyên liệu để sản xuất một lượng lớn các hoạt
chất sinh học quý dùng trong bào chế dược phẩm [100]. Nuôi cấy tế bào huyền
phù thực vật từ lâu đã được xem là một phương thức thay thế có nhiều tiềm
năng trong sản xuất các hợp chất chuyển hóa thứ cấp có giá trị cao nhờ tính
đồng nhất của quần thể tế bào in vitro, sự sẵn có một lượng lớn nguyên liệu, và
tốc độ sinh trương nhanh của chúng [106]. Nghiên cứu cải thiện hiệu suất nhân
giống cũng như sự sinh trương của callus, tế bào có sử dụng AgNO3 sẽ làm
tăng hiệu quả của quá trình sản xuất curcumin từ tế bào huyền phù cây nghệ
đen.
4.1.1. Nhân giông cây nghệ đen in vitro
Nghệ đen là loài nhân giống bằng củ, nhưng phải mất thời gian dài để
tạo củ, nên hệ số nhân thấp. Mặt khác, các cây thuộc họ Gừng thường bị bị xâm
nhiễm bơi các mầm bệnh, chẳng hạn như bệnh thối củ gây ra bơi các loài
Pithium và bệnh đốm lá do các loài Coleotrichum [31]. Do đó, sử dụng kỹ thuật
in vitro trong nhân giống nghệ đen, tạo nguồn dược liệu bền vững là hướng
nghiên cứu đang thu hút sự quan tâm của nhiều nhà khoa học. Marcia và cs.
(2000) đã nghiên cứu nhân giống cây nghệ đen ơ Malaysia và xây dựng công
thức nhằm đánh giá số lượng cây giống in vitro [92]. Bharalee và cs. (2005)
cũng đã nghiên cứu nhân giống bằng chồi củ của cây nghệ đen. Khả năng hình
thành chồi tốt nhất trên môi trường MS bổ sung 1 mg/L BAP và 0,5 mg/L NAA,
đạt 4,5 chồi/mẫu. Số rễ cao nhất 8,9 rễ/mẫu khi nuôi cấy chồi trên môi trường

80
MS bổ sung 0,5 mg/L IAA. Các cây tạo rễ có thể đưa ra ngoài đất [26].
Anisuzzaman và cs (2008) đã nghiên cứu tạo củ in vitro cây nghệ đen ơ
Bangladesh. Chồi in vitro khoảng 10-12 tuần tuổi được nuôi cấy trên môi
trường MS với nồng độ khác nhau của BA và NAA và các nguồn carbon khác
nhau. Củ hình thành tốt nhất trên môi trường có 4,0 mg/L BA và 6% sucrose
sau 7-9 tuần nuôi cấy. Củ được nảy chồi trên môi trường MS không có bổ sung
chất kích thích sinh trương [15]. Stanly và cs (2010) nghiên cứu nhân giống in
vitro cây nghệ đen ơ Malaysia thông qua nhân nhanh chồi in vitro trên môi
trường rắn, nuôi cấy lắc trong bình tam giác và nuôi cấy ngập chìm tạm thời.
Tất cả các cây con in vitro sản xuất từ 3 hệ thống này đều sống và sinh trương
bình thường, không có sự thay đổi hình thái. Nghiên cứu cũng cho thấy hệ
thống nuôi cấy ngập chìm tạm thời là sự lựa chọn tốt nhất trong kỹ thuật nhân
giống in vitro để sản xuất cây con [147]. Tuy nhiên, các tác giả đã chưa sử dụng
AgNO3 trong quá trình nuôi cấy. Theo Kumar và cs (2009) và Sandra và Maira
(2013), AgNO3 khi được bổ sung vào môi trường nuôi cấy mô thực vật có tác
dụng ức chế hoạt động của ethylene nội sinh, vì thế quá trình sinh trương của
tế bào sẽ được thuận lợi hơn [73], [134].
Đối với quá trình tái sinh chồi, nghiên cứu của Loc và cs (2005) cho thấy
môi trường có bổ sung 3 mg/L BAP cho hiệu quả tốt nhất, số chồi/mẫu đạt 2,3
và chiều cao chồi đạt 3,42 cm [79]. Trong nghiên cứu này, chúng tôi sử dụng
công thức môi trường này để tiến hành nghiên cứu ảnh hương của AgNO3. Kết
quả nghiên cứu cho thấy số chồi tạo thành cao hơn (3,3 so với 2,3 chồi) và
chiều dài chồi cũng cao hơn (4,1 cm so với 3,42 cm). Như vậy, AgNO3 đã có
tác dụng trong việc nâng cao hiệu quả của quá trình tái sinh chồi. AgNO3 khi
được bổ sung vào môi trường nuôi cấy mô thực vật có tác dụng ức chế hoạt
động của ethylene nội sinh; do đó quá trình sinh trương của tế bào vì thế sẽ
được thuận lợi hơn [73], [134]. Các kết quả thu được của chúng tôi cũng phù
hợp với nhận định trên.

81
Môi trường tốt nhất để tạo rễ theo Loc và cs (2005) là 2 mg/L NAA, khi
bổ sung AgNO3 vào môi trường này, chúng tôi nhận thấy 1,5 mg/L AgNO3 cho
hiệu quả tốt nhất. So với nghiên cứu trước đây của Loc và cs (2005) thì kết quả
thu được của chúng tôi tốt hơn, số rễ/chồi nhiều hơn (27,0 so với 18,5), rễ tạo
thành cũng dài hơn (0,80 cm so với 0,51 cm) [79].
Qua các kết quả nghiên cứu về nhân giống, chúng tôi nhận thấy bổ sung
AgNO3 vào môi trường nuôi cấy sẽ nâng cao hiệu suất nhân giống, các chỉ tiêu
nghiên cứu đều tăng lên từ 25-50%.
4.1.2. Nuôi cấy callus va tế bao cây nghệ đen
Qua một số nghiên cứu trước đây cho thấy, đa số trường hợp nuôi cấy
callus hoặc tế bào thực vật đều cho hàm lượng các chất thứ cấp cao hơn nhiều
lần so với trong điều kiện tự nhiên. Chẳng hạn, hàm lượng anthocyanin ơ callus
của cây hoa cúc (Rudbeckia hirta) chiếm đến 4,97% trong khi hoa của cây này
trong tự nhiên, nơi tích lũy chủ yếu anthocyanin, chỉ có 0,28% [90]. Nguyễn
Hoàng Lộc và cs (2010, 2011) đã nghiên cứu sản xuất solasodine từ callus cây
cà gai leo in vitro và thu được kết quả cho thấy, hàm lượng solasodine tích lũy
trong callus cao hơn khoảng 8,5 lần so với tách chiết từ rễ cây tự nhiên 1 năm
tuổi [84]. Tiếp tục nghiên cứu nuôi cấy tế bào cà gai leo trong chai tam giác,
các tác giả nhận thấy solasodine tích lũy trong tế bào huyền phù cao hơn trong
callus [85].
Sản xuất các hợp chất thứ cấp có giá trị từ callus hoặc tế bào huyền phù
là hướng nghiên cứu đã được triển khai trong vài thập kỷ qua và mang lại nhiều
thành quả đáng khích lệ. Ngoài các nghiên cứu nhân giống in vitro cây nghệ
đen, các nghiên cứu nuôi cấy callus và tế bào huyền phù cây nghệ đen cũng đã
được công bố. Mello và cs. (2001a,b) đã tạo callus từ đoạn rễ của cây nghệ đen
in vitro trên môi trường MS có 3% sucrose, 13,4 μM NAA và 2,2 μM BAP
trong điều kiện tối. Các tác giả cũng đã nghiên cứu nuôi cấy tế bào huyền phù

82
cây nghệ đen. Kết quả cho thấy với 0,5 g tế bào trong bình tam giác 125 ml
chứa 10 ml môi trường MS có 3% sucrose, 13,4 μM NAA và 2,2 μM BAP, tốc
độ lắc 60 vòng/phút đã thu được khoảng 6 g sinh khối tươi của tế bào sau 35
ngày nuôi cấy. Nhóm tác giả này cũng đã tiến hành khảo sát ảnh hương của các
nguồn carbon khác nhau lên sinh trương của tế bào huyền phù nghệ đen. Kết
quả cho thấy sucrose là nguồn carbon thích hợp cho sinh trương của tế bào nghệ
đen, các nguồn carbon khác như galactose, glycerol và sorbitol không có vai
trò đáng kể trong kích thích sinh trương của tế bào [97], [98].
Theo Miachir và cs. (2004), callus không hình thành từ mô lá và rễ của
cây nghệ đen in vitro trên môi trường có 2,4-D. Callus hình thành tốt nhất từ
đoạn rễ trên môi trường MS bổ sung 1 mg/L NAA và nuôi cấy trong tối. Tác
giả cũng đã thiết lập nuôi cấy tế bào huyền phù cây nghệ trong bình tam giác
250 ml trên máy lắc. Nuôi cấy 1 g tế bào trong 75 ml môi trường MS có bổ
sung 1 mg/L NAA đã thu được sinh khối tế bào cao nhất là 8 g sau hơn 20 ngày
nuôi cấy ơ tốc độ lắc 100 vòng/phút [100].
Loc và cs. (2006, 2008) đã nghiên cứu sản xuất sinh khối tế bào cây nghệ
đen thông qua nuôi cấy callus, callus đã được tạo thành khi nuôi cấy mảnh lá
trên môi trường có chứa 3 mg/L BAP và 3 mg/L 2,4-D. Nuôi cấy tế bào huyền
phù được thiết lập bằng cách nuôi 3 g callus tươi trong bình tam giác 250 ml
chứa 50 ml cùng loại môi trường. Tế bào huyền phù sau đó được nuôi trên hệ
lên men 5 L, sinh khối tế bào đạt cao nhất sau 14 ngày nuôi cấy (sinh khối tươi
đạt 338,15 g, sinh khối khô đạt 38,85 g) với tốc độ khuấy 150 vòng/phút, tốc
độ sục khí 2,5 L/phút, tỷ lệ tiếp mẫu ban đầu là 50 g. Trong điều kiện này, hàm
lượng tinh dầu thu được là 1,78% trong khi hàm lượng curcumin là 9,69% [81],
[80].
Trong nghiên cứu của chúng tôi, việc bổ sung AgNO3 vào môi trường
nuôi cấy đã làm tăng khả năng sinh trương của tế bào. Sau 14 ngày nuôi cấy,
sinh khối tươi đạt cực đại là gần 8,5 g/bình, tương đương với khối lượng khô

83
khoảng 0,75 g/bình, cao hơn kết quả nghiên cứu trước đây của Tuan và cs.
(2011) (khoảng 0,66 g khô/bình) [156] hay Miachir và cs. (2004) (quy đổi
tương đương 5,33 g tươi/bình) [100]. Bên cạnh đó, thời gian đạt sinh trương
cực đại của chúng tôi (14 ngày) cũng ngắn hơn rất nhiều so với các nghiên cứu
đã công bố như trong nghiên cứu của Mello và cs. (2001) là 35 ngày [97] hay
Miachir và cs. (2004) là 20 ngày [100]. Thời gian nuôi cấy ngắn sẽ góp phần
làm tăng hiệu quả của quá trình sản xuất curcumin. So với các đối tượng khác,
thời gian sinh trương của tế bào huyền phù cây nghệ đen ngắn hơn, ví dụ sinh
khối tế bào huyền phù rau má đạt cực đại sau 24 ngày [86], cà gai leo đạt cực
đại sau 28 ngày [87].
Khi nuôi cấy callus nghệ đen trên môi trường được bổ sung AgNO3 1,5
mg/L, các khối callus có màu vàng đậm hơn đối chứng, ơ các môi trường khác
callus có màu vàng tương đương hoặc nhạt hơn. Tuy nhiên, kết quả cũng cho
thấy AgNO3 không thích hợp bằng KIN hoặc NAA khi kết hợp với 2,4-D trong
nuôi cấy tăng sinh khối callus nghệ đen. Trong các nồng độ AgNO3 được sử
dụng, sinh trương cao nhất của callus chỉ đạt 0,71 g tươi, tương ứng với 62,43
mg khô, thấp hơn rất nhiều so với công thức kết hợp 2,4-D 1 mg/L và KIN từ
1,5 mg/L, sinh khối tươi thu được 0,90 g với sinh khối khô là 84,11 mg.
4.2. PHÂN LẬP CÁC GEN TỔNG HỢP CURCUMINOID
4.2.1. Phân lập gen tổng hợp curcuminoid
Các kết quả nghiên cứu đã công bố cho thấy có sự tương đồng lớn giữa
các gen của nghệ đen với nghệ vàng (xấp xỉ 99%). Các phân tích trình tự cũng
cho thấy các gen thuộc tất cả loài của trong chi Curcuma có sự tương đồng rất
lớn, thậm chí ngay ơ các vùng không mã hóa. Ví dụ, gen mã hóa 5S RNAs
ribosome của nghệ vàng (LT853920.1) và nghệ đen (KU936068.1) có sự tương
đồng 92%. Chính sự tương đồng cao trong trình tự của các gen này dẫn đến sẽ
có sự tương đồng cao trong các trình tự amino acid của các protein được mã

84
hóa. Đây là cơ sơ để chúng tôi thiết kế các mồi để phân lập các gen sinh tổng
hợp curcuminoid từ nghệ đen.
Trong quá trình thiết kế mồi, chúng tôi gặp nhiều khó khăn khi các gen
trong mỗi loài thuộc chi Curcuma đều có sự đặc hiệu cao, đặc biệt là ơ vị trí
mơ đầu và kết thúc của vùng mã hóa (CDS). Do không có bất kỳ trình tự mRNA
đầy đủ nào của các gen này từ chi Curcuma được công bố trên GenBank, chúng
tôi phải dựa vào các trình tự sẵn có của các gen này trên cây nghệ vàng để
nghiên cứu, tuy nhiên, tất cả các gen đã được công bố đều chỉ có vùng mã hóa.
Để xác định các vùng 5’ và 3’ không dịch mã (UTR), chúng tôi sử dụng các
gen của nghệ vàng, tìm kiếm cơ sơ dữ liệu EST (expressed sequence tag) của
cây nghệ vàng nhận thấy các EST có sự ổn định cao ơ các gen này. Các EST
này được sử dụng cùng với các vùng CDS để tạo ra các chuỗi các đoạn DNA
được nhân bản nằm liền kề nhau (contig) dài hơn. Vì vùng không dịch mã của
các gen này có sự khác biệt lớn, các mồi cho vùng CDS được thiết kế dựa trên
các vùng này để đảm bảo tính đặc hiệu cho phản ứng PCR. Kết quả nghiên cứu
của chúng tôi cho thấy các sản phẩm PCR đặc hiệu cho tất cả các gen, điều này
chứng tỏ cây nghệ đen chứa tất cả các gen này.
Mặc dù các gen DCS và CURS trên cây nghệ đen chưa được nghiên cứu
nhiều, tuy nhiên các gen này trên cây nghệ vàng đã được nhiều tác giả nghiên
cứu và công bố. So sánh kết quả phân lập của chúng tôi với các kết quả đã công
bố trên cây nghệ vàng có thể thấy các gen giữa hai loài này có độ tương đồng
rất cao, lên đến 99%. Ngoài ra, nghiên cứu trên loài họ hàng cho thấy, M.
accuminata chứa 3 bản sao của gen tương tự CURS3 (LOC103968790,
LOC103968789 và LOC103968788), tất cả đều nằm trên nhiễm sắc thể số 10,
và 5 bản sao của gen tương tự CURS2 (LOC103976634, LOC103976805,
LOC103986053, LOC103987346, và LOC103968787) trên các nhiễm sắc thể
số 2, 5, 6 và 10 (https://banana-genome-hub.southgreen.fr/).

85
4.2.2. Sự biểu hiện của các gen tổng hợp curcuminoid
Curcumin là thành phần chính của nhóm curcuminoid, được tìm thấy
trong callus và liên quan đến sự biểu hiện của các gen tổng hợp curcuminoid
như trong phân tích RT-PCR (Hình 3.17). Tuy nhiên, diện tích peak thể hiện
trong các hình 3.18-3.20 cho thấy đã có sự chuyển hóa sinh học diễn ra trong
quá trình nuôi cấy callus. Các nghiên cứu trước đây của Sakui và cs (1992)
[132] và Loc và cs (2009) [82] cũng cho thấy có sự chuyển hóa sinh học của
các sesquiterpenoid trong quá trình nuôi cấy huyền phù tế bào nghệ đen có
nguồn gốc từ callus. Theo Muffler và cs (2011), quá trình chuyển hóa sinh học
là bắt buộc để thu các hoạt chất sinh học từ các tiền chất trong quá trình sản
xuất triterpene từ nuôi cấy mô thực vật [107]. Nghiên cứu của Dvořáková và cs
(2007) cũng cho thấy quá trình chuyển hóa sinh học của hỗn hợp monoterpene
trong nuôi cấy huyền phù tế bào 2 loài Picea abies và Taxus baccata [42].
Trên các đối tượng nghệ, mức độ biểu hiện của các gen DCS và CURS
cũng đã được nhiều tác giả nghiên cứu. Katsuyama và cs (2009 a, b) đã xác
định sự biểu hiện của các gen DCS, CURS1, CURS2 và CURS3 trong củ và lá
của cây nghệ vàng bằng kỹ thuật qPCR (quantitative PCR) [66], [67]. Behar và
cs (2016) cũng nhận thấy sự phiên mã của các gen DCS, CURS1, CURS2 và
CURS3 trong củ và lá cây C. caesia, trong đó mức độ phiên mã của gen CURS
trong củ 5 tháng tuổi cao hơn 4 lần so với củ 10 tháng tuổi [24].
4.3. NGHIÊN CỨU ẢNH HƯỞNG CỦA ELICITOR LÊN SỰ BIỂU
HIỆN CỦA CÁC GEN TỔNG HỢP CURCUMINOID
4.3.1. Đặc điểm của các elicitor sử dụng trong nghiên cứu
4.3.1.1. Dịch chiết nấm men

86
Dịch chiết nấm men (YE) là một elicitor sinh học, được sử dụng trong
nuôi cấy mô và tế bào thực vật để tăng khả năng kích thích bộ máy bảo vệ, từ
đó gia tăng sản xuất các hợp chất thứ cấp [53]. Pitta-Alvarez và cs (2000)
nghiên cứu sử dụng YE 0,8 mg/mL trên rễ tơ cây Brugmansia candida, sau 48
giờ nuôi cấy các tác giả nhận thấy hàm lượng scopolamine và hyoscyamine
tăng khoảng 200% so với đối chứng (không có YE). Ngoài ra, kết quả cũng cho
thấy scopolamine cao hơn khoảng 600% so với đối chứng sau 24 giờ nuôi cấy.
Điều này có thể được lý giải bơi trong thành phần của YE được cho là có chứa
một số cation như Zn2+, Ca2+ và Co2+ có vai trò như là các elicitor phi sinh học.
Bên cạnh đó YE cũng là một hợp chất bao gồm nhiều thành phần khác nhau,
ngoài các amino acid, vitamin và khoáng chất, nó còn có thể chứa các thành
phần elicitor khác chưa được xác nhận [118].
Cơ chế tác động chính xác của YE lên tế bào thực vật hiện nay vẫn chưa
được làm rõ. Tuy nhiên, sự truyền tín hiệu cảm ứng được xem là một hệ thống
nhiều thành phần có chức năng phức tạp trong các phản ứng bảo vệ khác nhau
của tế bào thực vật, giống như: các kênh ion, sự hoạt hóa tác nhân phân bào
khơi động protein kinase hoặc con đường jasmonic acid hoặc SA [127], [143].
Các nghiên cứu sử dụng YE để tăng mức độ tích lũy các hợp chất thứ
cấp đã được một số tác giả công bố. Chẳng hạn, nuôi cấy rễ tơ cây rau sam đã
được Pirian và cs (2013) thực hiện để sản xuất noradrenaline. Các tác giả đã sử
dụng YE ơ các nồng độ 0,125-1 g/L để cảm ứng làm tăng quá trình sản xuất
noradrenaline, thời gian cảm ứng 2 ngày. Các tác giả nhận thấy liều lượng 0,25
và 0,5 g/L YE là thích hợp nhất, hàm lượng noradrenaline tăng 3-4 lần so với
đối chứng [117]. Khi bổ sung dịch chiết nấm men, Aspergillus niger, Fusarium
oxysporum vào môi trường nuôi cấy Hypericum triquetrifolium Turra ơ các
nồng độ từ 0,1-0,75 mg/L, Azeez và cs (2013) nhận thấy các elicitor này có vai
trò khác nhau trong việc sản xuất các hợp chất thứ cấp. Đối với nấm men (0,5
mg/L), hàm lượng p-OH-benzoic acid và chlorgenic acid tăng lên đáng kể so

87
với đối chứng. Rutin, hypersoid và quercitin tăng lên đáng kể so với đối chứng
khi được xử lý kích kháng ơ tất cả các công thức [20]. Pitta-Alvarez và cs
(2000) nghiên cứu sử dụng YE 0,8 g/L cảm ứng rễ tơ Brugmansia candida sau
48 giờ nuôi cấy nhận thấy, hàm lượng scopolamine và hyoscyamine trong rễ
tăng khoảng 200% so với đối chứng. Ngoài ra, kết quả cho thấy scopolamine
cao hơn khoảng 600% so với đối chứng (không có YE) sau 24 giờ nuôi cấy
[118].
4.3.1.2. Salicylic acid
Salicylic acid là một hợp chất tín hiệu thực vật quan trọng trong việc kích
hoạt các gen liên quan đến cơ chế bảo vệ. Khi sử dụng làm elicitor, SA ảnh
hương đến khả năng tích lũy các hợp chất liên quan đến cơ chế bảo vệ. SA được
biết như là tác nhân chống chịu tương tác thực vật gây bệnh, nhưng không phải
là chất cảm ứng vạn năng trong sản xuất các chất chuyển hóa bảo vệ thực vật
[29]. Theo Taguchi và cs (2001), SA tích lũy ơ các vị trí mầm bệnh để tấn công
và kích thích hệ thống bảo vệ thực vật và việc sản xuất các hợp chất thứ cấp
[150].
Việc sử dụng SA làm elicitor để tăng cường tổng hợp các hợp chất thứ
cấp từ nuôi cấy tế bào thực vật rất phổ biến. Yu và cs (2005) đã nghiên cứu sự
tích lũy jaceosidin và siringin trong nuôi cấy tế bào của sen tuyết với elicitor là
SA, kết quả thu được jaceosidin (95 mg/L) cao gấp 2,5 lần và siringin (631
mg/L) cao gấp 2,7 lần so với mẫu không bổ sung SA [167]. Dong và cs (2010)
nghiên cứu vai trò của SA đến sự tích lũy của phenolic acid và tăng hoạt động
các enzyme chống oxy hóa trong nuôi cấy tế bào của cây đan sâm. Phenolic
acid gồm có hai thành phần salvianolic acid B và caffeic acid. Khi bổ sung 6,25
mg/L SA vào môi trường nuôi cấy sau 8 giờ, hàm lượng salvianolic acid B và
caffeic acid thu được cao nhất, gấp 2 lần so với mẫu không bổ sung SA [39].
Pitta-Alvarez và cs (2000) nghiên cứu ảnh hương của SA đến khả năng tích lũy

88
scopolamine và hyoscyamine trong nuôi cấy rễ tơ Brugmansia candida nhận
thấy cả hai loại alkaloid này đều tăng sau 24 giờ cảm ứng. Ở tất cả các nồng độ
SA khảo sát (0,01-1,00 mM), hyoscyamine và scopolamine tích lũy cực đại lần
lượt sau 48 và 72 giờ nuôi cấy (gấp khoảng 6 và 10 lần so với đối chứng) [118].
4.3.1.3. Methyl jasmonate
MeJA là một hợp chất tạo hương dễ bay hơi lần đầu tiên được tìm thấy
trong hoa của cây Jasminum grandiflorum, sau đó được chứng minh là phân bố
rộng rãi trong giới thực vật. MeJA được ghi nhận như một chất điều hòa cần
thiết cho sự sống tế bào, có vai trò trung gian trong quá trình phát triển khác
nhau (nảy mầm, tạo rễ, sinh sản, chín quả, già yếu) và các phản ứng bảo vệ tế
bào chống lại các stress sinh học và phi sinh học (tổn thương do côn trùng gây
ra, điều kiện bất lợi của môi trường như hạn hán, nhiệt độ thấp, độ mặn) [33].
MeJA và jasmonic acid là những hợp chất điều hoà sinh trương thực vật
có vai trò trong sự lão suy của thực vật, tham gia vào con đường truyền tín hiệu
ngoại sinh (stress, vết thương, sinh vật gây bệnh) và cảm ứng hoạt động của hệ
thống bảo vệ ơ thực vật. MeJA khơi động sự phiên mã mRNA de novo của các
gen PAL dẫn tới sự tạo phenylalanine ammonia-lyase, enzyme đầu tiên của con
đường phenylpropanoid liên quan chặt chẽ đến sự sản xuất các HCTC ơ thực
vật (Brincat và cs, 2002) (dẫn theo Lê Thị Thủy Tiên và cs, 2010) [8]. MeJA
đã được nghiên cứu và sử dụng rộng rãi làm elicitor trong sản xuất các hợp chất
thứ cấp từ nuôi cấy tế bào thực vật [154].
Jasmonate ngoại sinh khi ứng dụng vào thực vật đã cho thấy có ảnh hương
đến sự biểu hiện về hình thái và sinh lý. Jasmonate kết hợp với sự tích lũy của
các hợp chất thứ cấp đã thể hiện sự đáp ứng bảo vệ ơ thực vật. MeJA làm tăng
hàm lượng shikonin và dẫn xuất của nó trong nuôi cấy tế bào cây Onosma
paniculatum, MeJA còn tăng cường sinh tổng hợp IAA nội sinh ơ thực vật.
Ứng dụng jasmonate ngoại sinh đã kích thích mạnh sự sinh tổng hợp nhiều hợp

89
chất thứ cấp trong nuôi cấy huyền phù tế bào và nuôi cấy cây in vitro. MeJA
cảm ứng tích lũy anthocynin đã được báo cáo ơ cây Arabidopsis thaliana,
Vaccinium pahalae và Vitis vinifera. Hơn nữa, MeJA và salicylic acid cảm ứng
sản xuất anthocyanine trong nuôi cấy callus cây D. carota [148].
MeJA ức chế sinh trương tế bào và tăng cường sản xuất hợp chất thứ cấp
trong nuôi cấy rễ cây Bupleurum falcatum L., Taxus spp. và lúa. Ảnh hương
của MeJ ngoại sinh lên hàm lượng của các amine sinh học bao gồm putrescine,
spermidine, tyramine, cadaverine và 2-phenylethylamine trong cây kiều mạch
(Fagopyrum esculentum) cũng đã được nghiên cứu [13].
4.3.2. Ảnh hưởng của elicitor lên sự sinh trưởng của tế bao
Trong hầu hết các công trình nghiên cứu được công bố, việc xử lý elicitor
sẽ ức chế sự sinh trương của tế bào giống như trong nghiên cứu của chúng tôi.
Đối với tế bào cây nghệ đen, xử lý SA sinh khối tươi của tế bào giảm còn 27,24-
44,75% trong khi xử lý YE sinh khối tươi giảm còn 63,29- 68,22% (Bảng 3.9).
Như vậy, tác dụng ức chế của SA lên sinh trương của tế bào nghệ đen gần gấp
đôi của YE.
Các nghiên cứu trước đây cũng cho kết quả tương tự. Yu và cs (2001) đã
nghiên cứu ảnh hương của SA và chất tách chiết từ nấm (F5) ơ các nồng độ
riêng rẽ và phối hợp lên khả năng sản xuất taxol trong nuôi cấy huyền phù tế
bào cây thông đỏ lá ngắn. Các tác giả nhận thấy khi bổ sung vào môi trường
nuôi cấy 50 mg/L F5 hoặc 50 mg/L SA hoặc phối hợp 50 mg/L F5 và 50 mg/L
SA đều ức chế sinh trương của tế bào, sinh khối thấp nhất khi sử dụng phối hợp
50 mg/L F5 và 50 mg/L SA [166]. Nuôi cấy rễ tơ cây Brugmansia candida bổ
sung SA (0,01-1,00 mM) ơ thời điểm sau 18 ngày, Pitta-Alvarez và cs (2000)
nhận thấy sinh trương tế bào bị ảnh hương, sinh khối tươi chỉ đạt 40% (480 mg)
so với đối chứng sau 72 giờ cảm ứng [118]. Nghiên cứu của Loc và cs (2012)
cũng có kết quả tương tự, sinh khối tế bào rau má sau 24 ngày nuôi cấy giảm

90
so với đối chứng ơ tất cả các nồng độ SA (50-200 µM) bổ sung tại các thời
điểm khác nhau (5-15 ngày) [86].
4.3.3. Ảnh hưởng của elicitor lên mức độ biểu hiện gen
Curcumin là một trong 3 thành phần của nhóm curcuminoid được tìm
thấy trong các loài thuộc chi Curcuma, bao gồm thêm demethoxycurcumin và
bisdemethoxycurcumin. Trong con đường sinh tổng hợp curcuminoid ơ cây
nghệ vàng, sự khơi đầu bắt nguồn từ phenylalanine, gen DCS xúc tác phản ứng
kết hợp feruloyl-CoA và p-coumaroyl-CoA tạo thành feruloyldiketide-CoA,
sau đó các gen CURS1-3 chuyển feruloyldiketide-CoA thành curcumin. Bên
cạnh đó, CURS 3 có thể tạo thành demethoxylcurcumin từ feruloyldiketide-
CoA. Đồng thời, gen DCS cũng tham gia chuyển hóa p-coumaroyl-CoA thành
p-coumaroyldiketide-CoA, sau đó các gen CURS1-3 hoặc chỉ gen CURS3
chuyển hóa chất này thành demethoxycurcumin và bisdemethoxycurcumin
[67].
Sự tích lũy của các sản phẩm thứ cấp chính trong nuôi cấy tế bào thực
vật tăng lên có liên quan đến mức độ biểu hiện của các gen cũng như tăng lên
dưới tác dụng của các elicitor. Trong nghiên cứu của chúng tôi, khi xử lý elicitor
ơ các nồng độ và thời điểm khác nhau sẽ tăng lên ơ các mức độ khác nhau. Đối
với gen CzDCS, mức độ biểu hiện tăng cao nhất khi xử lý YE vào thời điểm 5
ngày sau khi nuôi cấy, mức biểu hiện gấp 2,85 lần so với đối chứng (cường độ
băng DNA đạt 13.425 so với 4.706 của đối chứng). Đối với các gen CURS, gen
CzCURS1 mức biểu hiện cao nhất ghi nhận được tại công thức xử lý YE vào
thời điểm ban đầu (nồng độ 32.576), gen CzCURS2 là khi xử lý YE ơ thời điểm
5 ngày (cường độ 17.778) còn gen CzCURS3 cao nhất khi xử lý YE ơ thời điểm
ban đầu (cường độ 21.700) (Bảng 3.8). Như vậy, trong tất cả các công thức xử
lý, mức độ biểu hiện của các gen khi xử lý bằng YE đều cao hơn khi xử lý bằng
SA, điều này chứng tỏ YE phù hợp hơn SA trong việc tăng cường mức độ biểu

91
hiện của các gen trong chu trình phenylpropanoid ơ cây nghệ đen.
Xét về thời điểm bổ sung elicitor, bổ sung vào thời điểm ban đầu cho
hiệu quả tốt hơn vào thời điểm 5 ngày sau nuôi cấy, mức độ biểu hiện của tất
cả các gen khi bổ sung YE vào thời điểm ban đầu là 71.528 (gấp 3,46 lần đối
chứng) trong khi tại thời điểm 5 ngày là 57.397 (gấp 2,78 lần so với đối chứng).
Thông qua quá trình xử lý elicitor, vai trò của các gen cũng đã được thể
hiện. Khi xử lý YE ơ thời điểm ban đầu, mật độ băng DNA của gen CzDCS là
5.133 trong khi tổng mật độ của 3 gen CzCURS là 66.415 (Bảng 3.8) cho hàm
lượng curcumin là 0,92 mg/g khô (Bảng 3.9). Trong khi đó bổ sung elicitor tại
thời điểm 5 ngày, mật độ của gen CzDCS là 13.425 (tăng hơn 2 lần) và tổng
mật độ của các gen CzCURS là 43.971 (giảm hơn 30% so với bổ sung thời điểm
ban đầu) nhưng hàm lượng curcumin đã tăng lên là 1,44 mg/g khô (gấp 1,5
lần).
Xử lý SA cũng cho kết quả tương tự nhưng không rõ bằng YE. Khi xử
lý SA ơ thời điểm ban đầu, mật độ băng DNA của gen CzDCS là 5.312 trong
khi tổng mật độ của 3 gen CzCURS là 18.142 (Bảng 3.8) cho hàm lượng
curcumin là 1,14 mg/g khô (Bảng 3.9). Trong khi đó bổ sung elicitor tại thời
điểm 5 ngày, mật độ của gen CzDCS là 6.146 (tăng khoảng 1,2 lần) và tổng mật
độ của các gen CzCURS là 25.479 (tăng 1,4 lần) thu được hàm lượng curcumin
là 1,32 mg/g khô (gấp 1,15 lần). Như vậy, việc tăng mức độ biểu hiện của gen
CzDCS đã làm tăng khả năng sinh tổng hợp curcumin trong tế bào, mức độ biểu
hiện của các gen CzCURS ít tác động đến khả năng sinh tổng hợp curcumin. So
sánh khả năng sinh tổng hợp curcumin khi xử lý SA và YE cho thấy, mức độ
biểu hiện của gen CzDCS cao hơn sẽ thu được hàm lượng curcumin cao hơn.
Đối với các hợp chất được sinh tổng hợp theo chu trình phenylpropanoid,
các nghiên cứu cải thiện mức độ biểu hiện của các gen tham gia chu trình cũng
đã được nghiên cứu. Chẳng hạn, các gen trong con đường phenylpropanoid
được kích hoạt biểu hiện và mức độ tích lũy furanocoumarin trong nuôi cấy tế

92
bào cây rau mùi tây (Petroselinum crispum) [44].
Nghiên cứu tăng cường biểu hiện của các gen trong chu trình
phenylpropanoid bằng elicitor cũng đã được nhiều tác giả thực hiện. Park và cs
(2016) đã nghiên cứu sử dụng YE và AgNO3 để tăng cường mức độ biểu hiện
của các gen trong chu trình phenylpropanoid trong nuôi cấy tế bào cây hoắc
hương núi (Agastache rugosa). Kết quả cho thấy xử lý YE 0,5 g/L và AgNO3
30 mg/L ơ các thời điểm khác nhau đều làm tăng biểu hiện của các các gen
trong chu trình phenylpropanoid và sự tích lũy của rosmarinic acid. Mức độ
biểu hiện của các gen RAS và HPPR tăng lên cao nhất là 1,84, 1,97 và 2,86 lần
khi được xử lý YE ơ các mốc thời gian 3, 6 và 12 giờ. Đối với gen PAL, mức
độ phiên mã tăng lên tới 52,31 lần khi xử lý bằng AgNO3 sau 24 giờ [115].
Một số nghiên cứu trước đây cũng cho thấy sự tăng biểu hiện của các
gen khi được xử lý elicitor, ví dụ gen OsWRKY53 trong tế bào cây lúa khi xử
lý cerebroside từ nấm (Chujo và cs. 2007) [34], gen bảo vệ ơ tế bào rau mùi tây
khi xử lý bằng MeJA (Ellard-Ivey và Douglas. 1996) [44], hay gen bảo vệ ơ tế
bào 2 loài nho (Vitis rupestris và V. vinifera) được xử lý bằng Harpin (Qiao và
cs 2010) [121]. Trong một nghiên cứu khác, SA có tác dụng làm tăng biểu hiện
các gen tham gia vào con đường tổng hợp phytosterol và triterpene trong tế bào
cây rau má (Centella asiatica) [88].
4.3.4. Ảnh hưởng của các elicitor lên khả năng sản xuất curcumin
Trong nghiên cứu của chúng tôi trên cây nghệ đen, xử lý YE vào thời
điểm 5 ngày sau quá trình nuôi cấy cho kết quả tốt nhất, hàm lượng curcumin
đạt 1,44 mg/g, gấp 1,53 lần so với đối chứng không xử lý elicitor (Bảng 3.9).
Các công thức xử lý khác cũng đều thu được hàm lượng curcumin cao hơn
đối chứng, xử lý bằng SA vào thời điểm 5 ngày cũng cho kết quả rất tốt, hàm
lượng curcumin cao gấp 2,32 lần so với đối chứng. Đối chiếu với kết quả
phân tích mức độ biểu hiện của các gen trong chu trình, sự tích lũy curcumin

93
trong tế bào có sự khác biệt. Đối với YE, mức độ biểu hiện của các gen
CURS tại thời điểm ban đầu cao hơn thời điểm 5 ngày, trong khi hàm lượng
curcumin tại thời điểm 5 ngày lại cao hơn, điều này có thể thấy vai trò quan
trọng của gen DCS trong quá trình sinh tổng hợp curcumin, việc tăng mức
độ biểu hiện của gen DCS tại thời điểm 5 ngày có ảnh hương quyết định đến
việc tăng khả năng tổng hợp curcumin trong tế bào. Trong chu trình này, gen
DCS xúc tác phản ứng kết hợp feruloyl-CoA và p-coumaroyl-CoA tạo thành
feruloyldiketide-CoA, đây là nguyên liệu để tổng hợp nên các chất thuộc
nhóm curcuminoid, các gen CURS1-3 sẽ tham gia xúc tác tổng hợp curcumin
và các dẫn xuất của nó, việc biểu hiện các gen này ơ các mức độ khác nhau
có thể dẫn đến việc tổng hợp curcumin và các dẫn xuất với mức độ khác
nhau. Việc xử lý SA sẽ làm tăng đều mức độ biểu hiện của các gen nhưng ơ
mức độ thấp hơn khi xử lý bằng YE.
Các nghiên cứu đã công bố trước đây cho thấy mỗi elicitor cho hiệu quả
cải thiện hiệu suất của các hợp chất thứ cấp khác nhau đối với từng loài thực
vật. Vì vậy, việc lựa chọn elicitor phù hợp với mỗi đối tượng nghiên cứu là rất
quan trọng. Chẳng hạn MeJA tăng cường tích lũy azadirachtin trong nuôi cấy
tế bào Azadirachta indica tốt hơn các elicitor khác như SA, pectinase,
pectolyase, AgNO3, CuSO4 và dịch chiết nấm [123].
Tương tự nghiên cứu của chúng tôi, một số công trình nghiên cứu cho
rằng kích kháng bằng các tác nhân sinh học tốt hơn các tác nhân phi sinh học.
Goyal và cs (2008) tăng cường tích lũy isoflavonoid trong nuôi cấy tế bào của
cây Pueraria tuberose có bổ sung các elicitor như YE, MeJA và SA. Hàm
lượng isoflavonoid thu được ơ mẫu có bổ sung 200 µg/L YE đạt cao nhất gấp
12,4 lần so với mẫu không bổ sung YE. Trong khi các mẫu có bổ sung SA hoặc
MeJA, hàm lượng isoflavonoid thấp hơn mẫu bổ sung YE nhưng vẫn cao hơn
mẫu đối chứng từ 9,53-10,1 lần [51].

94

95
KẾT LUẬN VÀ KIẾN NGHỊ
KẾT LUẬN
1. Đã xây dựng thành công hệ thống nuôi cấy cây nghệ đen có sử dụng
AgNO3, bổ sung AgNO3 1,5 mg/L vào các môi trường nuôi cấy tạo cây in vitro
và nuôi cấy callus đều cho hiệu quả nuôi cấy tốt nhất.
2. Đã phân lập thành công 4 gen tham gia vào quá trình sinh tổng hợp
curcuminoid, các gen này được đặt tên tương ứng là CzDCS, CzCURS1,
CzCURS2 và CzCURS3; có chiều dài lần lượt là 1382 bp, 1240 bp, 1288 bp và
1265 bp; tương đồng 99% so với các gen tương ứng ơ cây nghệ vàng và đã
được đăng ký trên ngân hàng gen với các mã số lần lượt là MF663785,
MF402846, MF402847 và MF987835. Các gen này đều có sự biểu hiện ơ trong
củ và callus của cây nghệ đen.
3. Các elicitor (dịch chiết nấm men và salicilic acid) đều có tác dụng tăng
cường mức độ biểu hiện của các gen cũng như sự tổng hợp curcumin. Giá trị
tốt nhất thu được khi xử lý dịch chiết nấm men (1 g/L) sau 5 ngày nuôi cấy,
mức độ biểu hiện của các gen cao hơn 2,78 lần và hàm lượng curmin tích lũy
cao hơn 2,5 lần so với đối chứng không xử lý.
4. Gen DCS có vai trò lớn nhất trong các gen tham gia vào quá trình sinh
tổng hợp curcumin ơ cây nghệ đen, mức độ biểu hiện của gen này quyết định
trực tiếp đến hàm lượng curcumin thu được.
KIẾN NGHỊ
Đề tài cần được tiếp tục nghiên cứu theo hướng ứng dụng, mơ rộng ơ
quy mô pilot, thăm dò thêm các elicitor khác, từ đó hoàn thiện quy trình sản
xuất để có thể áp dụng vào sản xuất ơ quy mô công nghiệp.

96
DANH MỤC CÁC CÔNG BỐ LIÊN QUAN ĐẾN LUẬN ÁN
1. Trương Thị Phương Lan, Lê Thị Anh Thư, Ngô Thị Sen (2017) Ảnh hương
của AgNO3 đến quá trình tái sinh in vitro cây nghệ đen (Curcuma zedoaria
Roscoe). Tạp chí Khoa học Đại học Huế: Nông nghiệp và Phát triển nông
thôn 126(3D): 5-15.
2. Trương Thị Phương Lan, Lê Thị Anh Thư, Nguyễn Thị Hà Ngân (2017)
Thăm dò ảnh hương của môi trường nuôi cấy đến sinh trương của callus
nghệ đen (Curcuma zedoaria Roscoe). Tạp chí Khoa học và Công nghệ,
Trường đại học Khoa học, Đại học Huế: Chuyên san Hóa-Sinh-Khoa học
Trái đất 12(2): 63-74.
3. Truong Thi Phuong Lan, Nguyen Duc Huy, Nguyen Ngoc Luong, Nguyen
Van Nghi, Trinh Huu Tan, Le Viet Quan, Nguyen Hoang Loc (2018)
Identification and characterization of genes in curcuminoid pathway of
Curcuma zedoaria Roscoe. Current Pharmaceutical Biotechnology. DOI:
10.2174/1389201019666181008112244.
4. Truong Thi Phuong Lan, Nguyen Duc Huy, Nguyen Ngoc Luong, Hoang
Tan Quang, Trinh Huu Tan, Le Thi Anh Thu, Nguyen Xuan Huy, Nguyen
Hoang Loc (2019) Expression of elicitor-induced curcuminoid genes in cells
of zedoary (Curcuma zedoaria Roscoe). Journal of Plant Biotechnology
(accepted).

97
TÀI LIỆU THAM KHẢO
TÀI LIỆU TIẾNG VIỆT
1. Lê Quý Bảo (2004), "Các cấu tử dễ bay hơi của thân rễ Nga truật (Curcuma
zedoaria Rosc.) trồng ơ tỉnh Nghệ An và Hà Tĩnh", Tạp chí Dược học, 343, tr.
9-11.
2. Đào Hùng Cường (2003), "Tách chiết curcumin từ củ nghệ bằng dung môi
thực phẩm", Tạp chí Hóa học và Ứng dụng, 11, tr. 36-40.
3. Phan Minh Giang, Văn Ngọc Hưng, Phan Tống Sơn (1997), "Đóng góp vào
việc nghiên cứu các sesquiterpenoid trong thân rễ nghệ đen (Curcuma
zedoaria)", Tạp chí Hóa học và Công nghiệp Hóa chất, 4, tr. 9-11.
4. Trần Thị Việt Hoa, Trần Thị Phương Thảo, Vũ Thị Thanh Tâm (2007),
"Thành phần hóa học và tính kháng oxy hóa của nghệ đen Curcuma zedoaria
Berg trồng ơ Việt Nam", Tạp chí Phát triển Khoa học và Công nghệ, 10(4), tr.
37-47.
5. Nguyễn Hoàng Lộc (2011), Nuôi cấy mô và tế bào thực vật-Các khái niệm
và ứng dụng, Nxb Đại học Huế.
6. Đỗ Tất Lợi (1995), Tinh dầu Việt Nam, Nxb Y học, TP. Hồ Chí Minh.
7. Đỗ Tất Lợi (2004), Những cây thuốc và vị thuốc Việt Nam, Nxb Y học, Hà
Nội.
8. Lê Thị Thủy Tiên, Bùi Trang Việt, Nguyễn Đức Lượng (2010), Khảo sát vài
yếu tố ảnh hương đến sự sinh tổng hợp taxol của các hệ thống tế bào Taxus
wallichiana Zucc. in vitro, Tạp chí Phát triển Khoa học và Công nghệ, 13(T3),
tr. 67-77.
TÀI LIỆU TIẾNG ANH
9. Abraham F., Bhatt A., Keng C.L., Indrayanto G., Sulaiman S.F. (2011),
Effect of yeast extract and chitosan on shoot proliferation, morphology and

98
antioxidant activity of Curcuma mangga in vitro plantlets, African Journal of
Biotechnology, 10(40), pp. 7787-7795.
10. Agrawal G.K., Rakwal R., Iwahashi H. (2002), Isolation of novel rice
(Oryza sativa L.) multiple stress responsive MAP kinase gene, OsMSRMK2,
whose mRNA accumulates rapidly in response to environmental cues,
Biochemical and Biophysical Research Communications, 294(5), pp. 1009-
1016.
11. Ahlawat S., Saxena P., Alam P., Wajid S., Abdin M.Z. (2014), Modulation
of artemisinin biosynthesis by elicitors, inhibitor, and precursor in hairy root
cultures of Artemisia annua L., Journal of Plant Interactions, 9(1), pp. 811-
824.
12. Ahmed S.A., Baig M.M.V. (2014), Biotic elicitor enhanced production of
psoralen in suspension cultures of Psoralea corylifolia L., Saudi Journal of
Biological Sciences, 21(5), pp. 499-504.
13. Akula R., Ravishankar G.A. (2011), Influence of abiotic stress signals on
secondary metabolites in plants, Plant Signaling & Behavior, 6(11), pp. 1720-
1731.
14. Amalraj A., Pius A., Gopi S., Gopi S. (2017), Biological activities of
curcuminoids, other biomolecules from turmeric and their derivatives – A
review, Journal of Traditional and Complementary Medicine, 7(2), pp. 205-
233.
15. Anisuzzaman M., Sharmin A., Mondal S.C., Sultana R., Khalekuzzaman
M., Alam I., Alam M.F. (2008), In vitro microrhizome in Curcuma zedoaria
(Christm.) Roscoe-a conservation prioritized medicinal plant, Journal of
Biological Sciences, 8(7), pp. 1216-1220.
16. Ansari M.H., Ahmad S. (1991), Screening of some medicinal plants for
antiamoebic action, Fitoterapia, 62, pp. 171-175.

99
17. Apostol I., Heinstein P.F., Low P.S. (1989), Rapid Stimulation of an
Oxidative Burst during Elicitation of Cultured Plant Cells : Role in Defense
and Signal Transduction, Plant Physiology, 90(1), pp. 109-116.
18. Arıcan E. (2016), Elicitation triterpene yield in Alstonia scholaris cell
cultures via synergetic organisms, Biotechnology & Biotechnological
Equipment, 30(5), pp. 915-920.
19. Armero J., Tena M. (2001), Possible role of plasma membrane H+-ATPase
in the elicitation of phytoalexin and related isoflavone root secretion in
chickpea (Cicer arietinum L.) seedlings, Plant Science, 161(4), pp. 791-798.
20. Azeez T.H.A., Ibrahim K.M. (2013), Effect of biotic elicitors on secondary
metabolite production in cell suspensions of Hypericum triquetrifolium,
Bulletin UASVM Horticulture, 70(1), pp. 26-33.
21. Babu K.N., Rajesh M.K., Samsudeen K., Minoo D., Suraby E.J., Anupama
K., Ritto P. (2014), Randomly Amplified Polymorphic DNA (RAPD) and
Derived Techniques. In Besse P, ed. Molecular Plant Taxonomy: Methods and
Protocols. Totowa, NJ, Humana Press: 191-209
22. Badrhadad A., Piri K., Ghiasvand T. (2013), Increasing of alpha-tocopherol
in cell suspension cultures Elaeagnus angustifolia L., International Journal of
Agriculture and Crop Sciences, 5, pp. 1328-1331.
23. Behar N., Tiwari K.L., Jadhav S.K. (2013), In silico survey of genes
involved in curcuminoid synthesis for expression studies in Curcuma caesia
Roxb. IJCA Proceedings on National Seminar on Application of Artificial
Intelligence in Life Sciences NSAAILS, pp: 37-39.
24. Behar N., Tiwari K.L., Jadhav S.K. (2016), Semi-quantitative expression
studies of genes involved in biosynthesis of curcuminoid in Curcuma caesia
Roxb., Indian Journal of Biotechnology, 15, pp. 491-494.
25. Benhamou N. (1996), Elicitor-induced plant defence pathways, Trends in
Plant Science, 1(7), pp. 233-240.

100
26. Bharalee R., Das A., Kalita M.C. (2005), In vitro clonal propagation of
Curcuma caesia Roxb and Curcuma zedoaria Rosc. from rhizome bud
explants, Journal of Plant Biochemistry and Biotechnology, 14, pp. 61-63.
27. Bota C., Deliu C. (2012), Effect of some biotic elicitors on flavonoids
production in Digitalis lanata cell cultures, Revista medico-chirurgicala a
Societatii de Medici si Naturalisti din Iasi, 116(2), pp. 624-629.
28. Brand S., Hölscher D., Schierhorn A., Svatoš A., Schröder J., Schneider B.
(2006), A type III polyketide synthase from Wachendorfia thyrsiflora and its
role in diarylheptanoid and phenylphenalenone biosynthesis, Planta, 224(2),
pp. 413-428.
29. Bulgakov V.P., Tchernoded G.K., Mischenko N.P., Khodakovskaya M.V.,
Glazunov V.P., Radchenko S.V., Zvereva E.V., Fedoreyev S.A., Zhuravlev
Y.N. (2002), Effect of salicylic acid, methyl jasmonate, ethephon and
cantharidin on anthraquinone production by Rubia cordifolia callus cultures
transformed with the rolB and rolC genes, Journal of Biotechnology, 97, pp.
213-221.
30. Champakaew D., Choochote W., Pongpaibul Y., Chaithong U., Jitpakdi A.,
Tuetun B., Pitasawat B. (2007), Larvicidal efficacy and biological stability of
a botanical natural product, zedoary oilimpregnated sand granules, against
Aedes aegypti, Parasitology Research, 100, pp. 729-737.
31. Chan L.K., Thong W.H. (2004), In vitro propagation of Zingiberaceae
species with medicinal properties, Journal of Plant Biotechnology, 6, pp. 181-
188.
32. Cheong J.J., Hahn M.G. (1991), A specific, high-affinity binding site for
the hepta-beta-glucoside elicitor exists in soybean membranes, The Plant Cell,
3(2), pp. 137.
33. Cheong J.J., Choi Y.D. (2003), Methyl jasmonate as a vital substance in
plants, TRENDS in Genetic, 19(7), pp. 409-413.

101
34. Chujo T., Takai R., Akimoto-Tomiyama C., Ando S., Minami E.,
Nagamura Y., Kaku H., Shibuya N., Yasuda M., Nakashita H., Umemura K.,
Okada A., Okada K., Nojiri H., Yamane H. (2007), Involvement of the elicitor-
induced gene OsWRKY53 in the expression of defense-related genes in rice,
Biochimica et Biophysica Acta - Gene Structure and Expression, 1769(7), pp.
497-505.
35. Chunfen Y., Xun S., Anchang L., Lin Z., Aihua Y., Yanwei X., Guangxi Z.
(2013), Advances in clinical study of curcumin, Current Pharmaceutical
Design, 19(11), pp. 1966-1973.
36. Cousins M.M., Adelberg J., Chen F., Rieck J. (2010), Secondary
metabolism-inducing treatments during in vitro development of turmeric
(Curcuma longa L.) rhizomes, Journal of Herbs, Spices & Medicinal Plants,
15(4), pp. 303-317.
37. Daduang S., Sattayasai N., Sattayasai J., Tophrom P., Thammathaworn A.,
Chaveerach A., Konkchaiyaphum M. (2005), Screening of plants containing
Naja naja siamensis cobra venom inhibitory activity using modified ELISA
technique, Analytical Biochemistry, 341, pp. 316-325.
38. De Navarro D.F., De Souza M.M., Neto R.A., Golin V., Niero R., Yunes
R.A., Delle M.F., Cechinel F.V. (2002), Phytochemical analysis and analgesic
properties of Curcuma zedoaria grown in Brazil, Phytomedicine, 9, pp. 427-
432.
39. Dong J., Wan G., Liang Z. (2010), Accumulation of salicylic acid-induced
phenolic compounds and raised activities of secondary metabolic and
antioxidative enzymes in Salvia miltiorrhiza cell culture, Journal of
Biotechnology, 148(2-3), pp. 99-104.
40. Droillard M.J., Thibivilliers S., Cazalé A.C., Barbier-Brygoo H., Laurière
C. (2000), Protein kinases induced by osmotic stresses and elicitor molecules

102
in tobacco cell suspensions: two crossroad MAP kinases and one
osmoregulation-specific protein kinase, FEBS Letters, 474(2), pp. 217-222.
41. Duke A.J., Bogenschutz G.M.J., Du Cellier J., Duke P.G.K. (2003), CRC
handbook of medicinal spices, CRC Press, Boca Raton, Florida.
42. Dvořáková M., Valterová I., Vaněk T. (2007), Biotransformation of a
monoterpene mixture by in vitro cultures of selected conifer species, Natural
Product Communications, 2, pp. 233-238.
43. Ebrahimi M., Zarinpanjeh N. (2015), Bio-elicitation of β-carboline
alkaloids in cell suspension culture of Peganum harmala L., Journal of
Medicinal Plants, 14, pp. 35-49.
44. Ellard-Ivey M., Douglas C.J. (1996), Role of jasmonates in the elicitor- and
wound-Inducible expression of defense genes in parsley and transgenic
tobacco, Plant Physiology, 112(1), pp. 183-192.
45. Estrada-Soto S., Vergara-Martínez V., Arellano J., C. Rivera-Leyva J.,
Castillo-España P., Flores Flores A., T. Cardoso-Taketa A., Perea-arango I.
(2018), Methyl jasmonate and salicylic acid enhanced the production of ursolic
and oleanolic acid in callus cultures of Lepechinia caulescens, Pharmacognosy
Magazine, 13, pp. 886-889.
46. Fan X., Zhang C., Liu D.B., Yan J., Liang H.P. (2013), The clinical
applications of curcumin: Current state and the future, Current Pharmaceutical
Design, 19(11), pp. 2011-2031.
47. Felix G., Grosskopf D.G., Regenass M., Boller T. (1991), Rapid changes
of protein phosphorylation are involved in transduction of the elicitor signal in
plant cells, Proceedings of the National Academy of Sciences, 88(19), pp. 8831-
8834.
48. Gelli A., Higgins V.J., Blumwald E. (1997), Activation of Plant Plasma
Membrane Ca2+-Permeable Channels by Race-Specific Fungal Elicitors, Plant
Physiology, 113(1), pp. 269-279.

103
49. Giang P.M., Huong V.N., Son P.T. (2000), Antimicrobial activity of
sesquitrepene contituents from some Curcuma species of Vietnam, Journal of
Chemistry, 38(1), pp. 91-94.
50. Gilani S.A., Kikuchi A., Shimazaki T., Wicaksana N., Wunna, Watanabe
K.N. (2015), Molecular genetic diversity of curcuminoid genes in Curcuma
amada: Curcuminoid variation, consideration on species boundary and
polyploidy, Biochemical Systematics and Ecology, 61, pp. 186-195.
51. Goyal S., Ramawat K.G. (2008), Increased isoflavonoids accumulation in
cell suspension cultures of Pueraria tuberosa by elicitors, Indian Journal of
Biotechnology, 7(3), pp. 378-382.
52. Gunawardena A.H., Greenwood J.S., Dengler N.G. (2004), Programmed
cell death remodels lace plant leaf shape during development, Plant Cell
Report, 16, pp. 60-73.
53. Hamza M.A. (2013), Effect of yeast extract (biotic elicitor) and incubation
periods on selection of Lupinus termis explant in vitro, Journal of Applied
Sciences Research, 9(7), pp. 4186-4192.
54. Hanania U., Avni A. (1997), High-affinity binding site for ethylene-
inducing xylanase elicitor on Nicotiana tabacum membranes, The Plant
Journal, 12(1), pp. 113-120.
55. Hanif R., Qiao L., Shiff S.J., Rigas B. (1997), Curcumin, a natural plant
phenolic food additive, inhibits cell proliferation and induces cell cycle changes
in colon adenocarcinoma cell lines by a prostaglandin-independent pathway,
Journal of Laboratory and Clinical Medicine, 130(6), pp. 576-584.
56. Harathi K., Naidu C.V. (2016), Influence of ethylene inhibitor silver nitrate
on direct shoot regeneration from in vitro raised shoot tip explants of
Sphaeranthus indicus Linn.-An important antijaundice medicinal plant,
American Journal of Plant Sciences, 7, pp. 525-532.

104
57. Hasanloo T., Eskandari S., Najafi F. (2014), Chitosan (middle-viscous) as
an effective elicitor for silymarin production in Silybum marianum hairy root
cultures, Research Journal of Pharmacognosy, 1, pp. 9-13.
58. Hayakawa H., Minaniya Y., Ito K., Yamamoto Y., Fukuda T. (2011),
Difference of curcumin content in Curcuma longa L. (Zingiberaceae) caused
by hybridization with other Curcuma species, American Journal of Plant
Sciences, 2, pp. 111-119.
59. Hong H.C., Noh M.S., Lee W.Y., Lee S.K. (2002), Inhibitory effects of
natural sesquiterpenoids isolated from the rhizome of Curcuma zedoaria on
prostaglandin E2 and nitric oxide production, Planta Medica, 68, pp. 545-547.
60. Huang M.T., Newmark H.L., Frenkel K. (1997), Inhibitory effects of
curcumin on tumorigenesis in mice, Journal of Cellular Biochemistry, 27, pp.
26-34.
61. Jang M.K., Sohn D.H., Ryu J.H. (2001), A curcuminoid and sesquiterpenes
as inhibitors of macrophage TNF-alpha release from Curcuma zedoaria, Planta
Medica, 67(6), pp. 550-552.
62. Jeong G.T., Park D.H. (2005), Enhancement of growth and secondary
metabolite biosynthesis: Effect of elicitors derived from plants and insects,
Biotechnology and Bioprocess Engineering, 10(1), pp. 73.
63. Jiang M.C., Yang Yen H.F., Yen J.J., Lin J.K. (1996), Curcumin induces
apoptosis in immortalized NIH 3T3 and malignant cancer cell lines, Nutrition
and Cancer, 26(1), pp. 111-120.
64. Joy P.P., Thomas J., Mathew S., Skari B.P. (1998), Medicinal Plants:
Tropical Horticulture, Naya Prakash, Calcutta, India.
65. Katsuyama Y., Matsuzawa M., Funa N., Horinouchi S. (2008), Production
of curcuminoids by Escherichia coli carrying an artificial biosynthesis
pathway, Microbiology, 154(Pt 9), pp. 2620-2628.

105
66. Katsuyama Y., Kita T., Funa N., Horinouchi S. (2009), Curcuminoid
biosynthesis by two type III polyketide synthases in the herb Curcuma longa,
Journal of Biological Chemistry, 284(17), pp. 11160-11170.
67. Katsuyama Y., Kita T., Horinouchi S. (2009), Identification and
characterization of multiple curcumin synthases from the herb Curcuma longa,
FEBS Letters, 583(17), pp. 2799-2803.
68. Katsuyama Y., Hirose Y., Funa N., Ohnishi Y., Horinouchi S. (2010),
Precursor-directed biosynthesis of curcumin analogs in Escherichia coli,
Bioscience, Biotechnology, and Biochemistry, 74(3), pp. 641-645.
69. Khojasteh A., Mirjalili M., Palazon J., Eibl R., Cusido R. (2016), Methyl
jasmonate-enhanced production of rosmarinic acid in cell cultures of Satureja
khuzistanica in a bioreactor, Engineering in Life Sciences, 16(8), pp. 740-749.
70. Kim D.I., Lee T.K., Jang T.H., Kim C.H. (2005), The inhibitory effect of a
Korean herbal medicine, Zedoariae rhizoma, on growth of cultured human
hepatic myofibroblast cells, Life Sciences, 77(8), pp. 890-906.
71. Kim Y.B., Reedb D.W., Covellob P.S. (2015), Production of triterpenoid
sapogenins in hairy root cultures of Silene vulgaris, Natural Product
Communications, 10(11), pp. 1919-1922.
72. Kita T., Imai S., Sawada H., Kumagai H., Seto H. (2008), The biosynthetic
pathway of curcuminoid in turmeric (Curcuma longa) as Revealed by 13C-
labeled precursors, Bioscience, Biotechnology, and Biochemistry, 72(7), pp.
1789-1798.
73. Kumar V., Parvatam G., Ravishankar G. (2009), AgNO3 - a potential
regulator of ethylene activity and plant growth modulator, Electronic Journal
of Biotechnology, 12(2), pp.
74. Lee J., Jung Y., Shin J.H., Kim H.K., Moon B.C., Ryu D.H., Hwang G.S.
(2014), Secondary metabolite profiling of Curcuma species grown at different

106
locations using GC/TOF and UPLC/Q-TOF MS, Molecules, 19(7), pp. 9535-
9551.
75. Lee J.K., Eom S.H., Hyun T.K. (2018), Enhanced biosynthesis of saponins
by coronatine in cell suspension culture of Kalopanax septemlobus, 3 Biotech,
8(1), pp. 59.
76. Leonel M., Sarmento S.B.S., Cereda M.P. (2003), New starches for the food
industry: Curcuma longa and Curcuma zedoaria, Carbohydrate Polymers, 54,
pp. 385-388.
77. Li P., Mao Z., Lou J., Li Y., Mou Y., Lu S., Peng Y., Zhou L. (2011),
Enhancement of diosgenin production in Dioscorea zingiberensis cell cultures
by oligosaccharides from its endophytic fungus Fusarium oxysporum Dzf17,
Molecules, 16(12), pp. 10631-10644.
78. Lobo R., Prabhu K.S., Shirwaikar A., Shirwaikar A. (2009), Curcuma
zedoaria Rosc. (white turmeric): a review of its chemical, pharmacological and
ethnomedicinal properties, Journal of Pharmacy and Pharmacology, 61(1), pp.
13-21.
79. Loc N.H., Duc D.T., Kwon T.H., Yang M.S. (2005), Micropropagation of
zedoary (Curcuma zedoaria Roscoe)-a valuable medicinal plant, Plant Cell,
Tissue and Organ Culture, 81, pp. 119-122.
80. Loc N.H., Ha T.T.T., Hirata Y. (2006), Effect of several factors on cell
biomass production of zedoary (Curcuma zedoaria Roscoe) in bioreactor,
Vietnamese Journal of Biotechnol, 4(2), pp. 213-220.
81. Loc N.H., Diem D.T.H., Binh D.H.N., Huong D.T., Kim T.G., Yang M.S.
(2008), Isolation and characterization of antioxidation enzymes from cells of
zedoary (Curcuma zedoaria Roscoe) cultured in a 5-l bioreactor, Molecular
Biotechnology, 38(1), pp. 81-87.
82. Loc N.H., Tuan V.C., Binh D.H.N., Phuong T.T.B., Kim T.G., Yang M.S.
(2009), Accumulation of secquiterpenes and polysaccharides in cells of

107
zedoary (Curcuma zedoaria Roscoe) cultured in a 10 L bioreactor,
Biotechnology and Bioprocess Engineering, 14, pp. 619-624.
83. Loc N.H., An N.T.T. (2010), Asiaticoside production from cell culture of
centella (Centella asiatica L. Urban), Biotechnology and Bioprocess
Engineering, 15, pp. 1065-1070.
84. Loc N.H., Anh N.H.T., Binh D.H.N., Yang M.S., Kim T.G. (2010),
Production of glycoalkaloids from callus cultures of Solanum hainanense
Hance, Journal of Plant Biotechnology, 37, pp. 96-101.
85. Loc N.H., Thanh L.T.H. (2011), Solasodine production from cell culture of
Solanum hainanense Hance, Biotechnology and Bioprocess Engineering,
16(3), pp. 581-586.
86. Loc N.H., Giang N.T. (2012), Effects of elicitors on the enhancement of
asiaticoside biosynthesis in cell cultures of centella (Centella asiatica L.
Urban), Chemical Papers, 66(7), pp. 642-648.
87. Loc N.H., Anh N.H.T., Khuyen L.T.M., An T.N.T. (2014), Effects of yeast
extract and methyl jasmonate on the enhancement of solasodine biosynthesis
in cell cultures of Solanum hainanense Hance, Journal of Plant Biochemistry
and Biotechnology, 3(1), pp. 1-6.
88. Loc N.H., Giang N.T., Huy N.D. (2016), Effect of salicylic acid on
expression level of genes related with isoprenoid pathway in centella (Centella
asiatica (L.) Urban) cells, 3 Biotech, 6(1), pp. 86-93.
89. Low P.S., Merida J.R. (1996), The oxidative burst in plant defense:
Function and signal transduction, Physiologia Plantarum, 96(3), pp. 533-542.
90. Luczkiewcz M., Cisowski W. (2001), Optimisation of the second phase of
a two phase growth system for anthocyanin accumulation in callus cultures of
Rudbeckia hirta, Plant Cell, Tissue and Organ Culture, 65(1), pp. 57-68.
91. Manivannan A., Soundararajan P., Park Y.G., Jeong B.R. (2016), Chemical
elicitor-induced modulation of antioxidant metabolism and enhancement of

108
secondary metabolite accumulation in cell suspension cultures of Scrophularia
kakudensis Franch, International Journal of Molecular Sciences, 17(3), pp.
399.
92. Marcia O.M., Antônio F.C.A., Murilo M. (2000), Quantifying the micro
propagation of Curcuma zedoaria Roscoe, Scientia Agricola, 57, pp. 703-707.
93. Martinoia E., Klein M., Geisler M., Bovet L., Forestier C., Kolukisaoglu
U., Muller-Rober B., Schulz B. (2002), Multifunctionality of plant ABC
transporters-more than just detoxifiers, Planta Med, 214, pp. 345-355.
94. Masuda T., Jitoe A., Isobe J., Nakatani N., Yonemori S. (1993),
Antioxidative and anti-inflammatory curcumin-related phenolics from
rhizomes of Curcuma domestica, Phytochemistry, 32, pp. 1557-1560.
95. Matsuda H., Ninomiya K., Morikawa T., Yoshikawa M. (1998), Inhibitory
effect and action mechanism of sesquiterpenes from zedoariae rhizoma on D-
galactosamine/lipopolysaccharide -induced liver injury, Bioorganic &
Medicinal Chemistry Letters, 8, pp. 339-344.
96. Matsuda H., Morikawa T., Toguchida I., Ninomiya K., Yoshikawa M.
(2001), Inhibitors of nitric oxide production and new sesquiterpenes,
zedoarofuran, 4-epicurcumenol, neocurcumenol, gajutsulactones A, and B,
zedoarolodes A and B, from zedoariae rhizoma, Chem Pharma Bull, 49, pp.
1558-1566.
97. Mello M.O., Dias C.T.S., Amaral A.F.C., Melo M. (2001), Growth of
Bauhinia forficata Link, Curcuma zedoaria Roscoe and Phaseolus vulgaris L.
cell suspension cultures with carbon sources, Scientia Agricola, 58, pp. 481-
485.
98. Mello M.O., M. M., Appezzato-da-Glória B. (2001), Histological analysis
of the callogenesis and organogenesis from root segments of Curcuma zedoaria
Roscoe, Brazilian Archives of Biology and Technology, 44(2), pp. 197-203.

109
99. Memelink J., Verpoorte R., Kijne J.W. (2001), ORCAnization of
jasmonate-responsive gene expression in alkaloid metabolism, Trends in Plant
Science, 6(5), pp. 212-219.
100. Miachir J.I., Romani V.L.M., De Campos A.A.F., Mello M.O., Crocomo
O.J., Melo M. (2004), Micropropagation and callogenesis of Curcuma zedoaria
Roscoe, Scientia Agricola, 61(4), pp. 427-432.
101. Mihaljević I., Dugalić K., Tomaš V., Viljevac M., Pranjić A., Čmelik Z.,
Puškar B., Jurković Z. (2013), In vitro sterilization procedures for
micropropagation of ‘Oblačinska’ sour cherry, Journal of Agricultural
Sciences, 58(2), pp. 117-126.
102. Mohanty S., Panda M.K., Subudhi E., Nayak S. (2008), Plant regeneration
from callus culture of Curcuma aromatica and in vitro detection of somaclonal
variation through cytophotometric analysis, Biologia Plantarum, 52(4), pp.
783-786.
103. Moharrami F., Hosseini B., Farjaminezhad M., Sharafi A. (2017), Effects
of yeast extract on antioxidant activity and tropane alkaloids production in
hairy roots cultures of Hyoscyamus reticulatus L., Scientific Journal
Management System, 33(3), pp. 466-480.
104. Mohiuddin A.K.M., Chowdhury M.K.U., Abdullah Z.C., Napis S. (1997),
Influence of silver nitrate (ethylene inhibitor) on cucumber in vitro shoot
regeneration, Plant Cell, Tissue and Organ Culture, 51(1), pp. 75-78.
105. Morikawa T., Matsuda H., Ninomiya K., Yoshikawa M. (2002), Medicinal
foodstuffs. XXIX, Potent protective effects of sesquiterpenes and curcumin
from zedoariae rhizoma on liver injury induced by Dgalactosamine/
lipopolysaccharide or tumour necrosis factor-α, Biological & Pharmaceutical
Bulletin, 25, pp. 627-631.

110
106. Moscatiello R., Baldan B., Navazio L. (2013), Plant Cell Suspension
Cultures. In Maathuis FJM, ed. Plant Mineral Nutrients: Methods and
Protocols. Totowa, NJ, Humana Press: 77-93
107. Muffler K., Leipold D., Scheller M.-C., Haas C., Steingroewer J., Bley T.,
Neuhaus H.E., Mirata M.A., Schrader J., Ulber R. (2011), Biotransformation
of triterpenes, Process Biochemistry, 46(1), pp. 1-15.
108. Mulabagal V. T.H.S. (2004), Plant cell cultures- an alternative and
efficient source for the production of biologically important secondary
metabolites, Int J Appl Sci Engin, 2, pp. 29-48.
109. Nadkarni K.M. (1999), Indian Materia Medica, Popular Prakashan Private
Ltd., Mumbai, India.
110. Namdeo A.G., Patil S., Fulzele D.P. (2002), Influence of fungal elicitors
on production of ajmalicine by cell cultures of Catharanthus roseus,
Biotechnology Progress, 18(1), pp. 159-162.
111. Namdeo A.G. (2007), Plant cell elicitation for production of secondary
metabolites: A review, Pharmacognosy Reviews, 1(1), pp. 69-79.
112. Neumann K.H., Kumar A., Imani J. (2009), Plant cell and tissue culture-
A Tool in biotechnology, Principles and Practice, Springer-Verlag, Berlin,
Heidelberg.
113. Paek K., Chakrabarty D., Hahn E.J. (2005), Application of bioreactor
systems for large scale production of horticultural and medicinal plants, Plant
Cell, Tissue and Organ Culture, 81, pp. 287-300.
114. Paramapojn S., Gritsanapan W. (2009), Free radical scavenging activity
determination and quantitative analysis of curcuminoids in Curcuma zedoaria,
Current Science, 97(7), pp. 1069-1073.
115. Park T.W., Arasu V.M., Al-Dhabi A.N., Yeo K.S., Jeon J., Park S.J., Lee
Y.S., Park U.S. (2016), Yeast extract and silver nitrate induce the expression

111
of phenylpropanoid biosynthetic genes and induce the accumulation of
rosmarinic acid in Agastache rugosa cell culture, Molecules, 21(4), pp. 426.
116. Pawar H., Karde M., Mundle N., Jadhav P., Mehra K. (2014),
Phytochemical evaluation and curcumin content determination of turmeric
rhizomes collected from Bhandara district of Maharashtra (India), Journal of
Medical Chemistry, 4, pp. 588-591.
117. Pirian K., Piri K. (2013), Influence of yeast extract as a biotic elicitor on
noradrenaline production in hairy root culture of Portulaca oleracea L.,
International Journal of Plant Production, 4(11), pp. 2960-2964.
118. Pitta-Alvarez S.I., Spollansky T.C., Guilietti A.M. (2000), The influence
of different biotic and abiotic elicitors on the production and profile of tropane
alkaloids in hairy root cultures of Brugmansia candida, Enzyme and Microbial
Technology, 26, pp. 252-258.
119. Plunkett G.M., Lowry P.P., Vu N.V. (2004), Phylogenetic relationships
among Polyscias (Araliaceae) and close relatives from the Western Indian
ocean sasin, International Journal of Plant Sciences, 165(5), pp. 861-873.
120. Prakash S., Elangomathavan R., Seshadri S., Kathiravan K., Ignacimuthu
S. (2004), Efficient Regeneration of Curcuma amada Roxb. Plantlets from
Rhizome and Leaf Sheath Explants, Plant Cell, Tissue and Organ Culture,
78(2), pp. 159-165.
121. Qiao F., Chang X.-L., Nick P. (2010), The cytoskeleton enhances gene
expression in the response to the Harpin elicitor in grapevine, Journal of
Experimental Botany, 61(14), pp. 4021-4031.
122. Raghuveer G.P.S., Ali M.M., Eranna D., Ramachandra S.S. (2003),
Evaluation of anti-ulcer effect of root of Curcuma zedoaria in rats, Indian
Journal of Traditional Knowledge, 2(4), pp. 375-377.
123. Ramawat K.G., Merillon J.M. (2008), Bioactive Molecules and Medicinal
Plants, Springer-Verlag, Berlin, Germany.

112
124. Ramirez-Ahumada M.d.C., Timmermann B.N., Gang D.R. (2006),
Biosynthesis of curcuminoids and gingerols in turmeric (Curcuma longa) and
ginger (Zingiber officinale): Identification of curcuminoid synthase and
hydroxycinnamoyl-CoA thioesterases, Phytochemistry, 67(18), pp. 2017-2029.
125. Resmi M.S., Soniya E.V. (2012), Molecular cloning and differential
expressions of two cDNA encoding Type III polyketide synthase in different
tissues of Curcuma longa L., Gene, 491(2), pp. 278-283.
126. Rezaei A., Ghanati F., Dehaghi M.A. (2011), Stimulation of taxol
production by combined salicylic acid elicitation and sonication in Taxus
baccata cell culture. International Conference on Life Science and Technology,
Singapore, pp: 193-197.
127. Rhee H.S., Cho H.-Y., Son S.Y., Yoon H.S.-Y., J.M. J.M.P. (2010),
Enhanced accumulation of decursin and decursinol angelate in root cultures and
intact roots of Angelica gigas Nakai following elicitation, Plant Cell, Tissue
and Organ Culture, 101, pp. 295-302.
128. Rodrigues J.L., Araújo R.G., Prather K.L.J., Kluskens L.D., Rodrigues
L.R. (2015), Production of curcuminoids from tyrosine by a metabolically
engineered Escherichia coli using caffeic acid as an intermediate,
Biotechnology Journal, 10(4), pp. 599-609.
129. Romeis T. (2001), Protein kinases in the plant defence response, Current
Opinion in Plant Biology, 4(5), pp. 407-414.
130. Roughley P.J., Whiting D.A. (1973), Experiments in the biosynthesis of
curcumin, Journal of the Chemical Society, Perkin Transactions 1, (0), pp.
2379-2388.
131. Saeed S., Ali H., Khan T., Kayani W., Khan M.A. (2017), Impacts of
methyl jasmonate and phenyl acetic acid on biomass accumulation and
antioxidant potential in adventitious roots of Ajuga bracteosa Wall ex Benth.,

113
a high valued endangered medicinal plant, Physiology and Molecular Biology
of Plants, 23(1), pp. 229-237.
132. Sakui N., Kuroyanagi M., Ishitobi Y., Sato M., Ueno A. (1992),
Biotransformation of sesquiterpenes by cultured cells of Curcuma zedoaria,
Phytochemistry, 31(1), pp. 143-147.
133. Sambrook J., Russell W.D. (2001), Molecular cloning: a laboratory
manual, 3rd edn, Cold Spring Harbor Laboratory Press, New York.
134. Sandra A.T., Maira O. (2013), Effect of culture medium consistence and
silver nitrate on micropropagation of two potato (Solanum tuberosum)
cultivars, Revista Colombiana de Biotecnología, 15, pp. 55-62.
135. Saralamp P., Chuakual W., Temsirikul R., Clayton T. (2000), Medicinal
plants in Thailand, Amerin Printing and Publishing Public Co., Ltd., Bangkok,
Thailand.
136. Sarkate A. (2017), Antioxidant and cytotoxic activity of bioactive
phenolic metabolites isolated from the yeast-extract treated cell culture of
apple, Plant Cell, Tissue and Organ Culture, 130(3), pp. 641-649.
137. Sarropoulou V., Dimassi-Theriou K., Therios I. (2016), Effect of the
ethylene inhibitors silver nitrate, silver sulfate, and cobalt chloride on
micropropagation and biochemical parameters in the cherry rootstocks CAB-
6P and Gisela 6, Turkish Journal of Biology, 40, pp. 670-683.
138. Schoch G.A., Nikov G.N., Alworth W.L., Werck-Reichhart D. (2002),
Chemical inactivation of the cinnamate 4-hydroxylase allows for the
accumulation of salicylic acid in elicited cells, Plant Physiology, 130(2), pp.
1022-1031.
139. Schopfer C.R., Kochs G., Lottspeich F., Ebel J. (1998), Molecular
characterization and functional expression of dihydroxypterocarpan 6a-
hydroxylase, an enzyme specific for pterocarpanoid phytoalexin biosynthesis
in soybean (Glycine max L.), FEBS Letters, 432(3), pp. 182-186.

114
140. Seo W.G., Hwang J.C., Kang S.K., Jin U.H., Suh S.J., Moon S.K. (2005),
Suppressive effect of Zedoariae rhizome on pulmonary metastasis of B16
melanoma cells, Journal of Ethnopharmacology, 101, pp. 249-257.
141. Sgamma T., Thomas B., Muleo R. (2015), Ethylene inhibitor silver nitrate
enhances regeneration and genetic transformation of Prunus avium (L.) cv
Stella, Plant Cell, Tissue and Organ Culture (PCTOC), 120(1), pp. 79-88.
142. Shahinozzaman M., Faruq M.O., Kalam Azad M.A., Amin M.N. (2013),
Studies on in vitro propagation of an important medicinal plant-Curcuma
zedoaria Roscoe using rhizome explants, Persian Gulf Crop Protection, 2(2),
pp. 1-6.
143. Sharma M., Sharma A., Ashwani K., Kumar B.S. (2011), Enhancement of
secandary metabolites in cultured plant cells thoungh stress stimulus, American
Journal of Plant Physiology, 6(2), pp. 50-71.
144. Shin K.H., Yoon K.Y., Cho T.S. (1994), Pharmacologycal activities of
sesquiterpenes from the rizhome of Curcuma zedoaria, Saengyak Hakhoechi,
25, pp. 221-225.
145. Srvidya A.R., Yadev A.K., Dhanbal S.P. (2009), Antioxidant and
antimicrobial activity of rhizome of Curcuma aromatica and Curcuma
zeodaria, leaves of Abutilon indicum, Archives of Pharmaceutical Science and
Research, 1(1), pp. 14-19.
146. Stanly C., Keng C.L. (2007), Micropropagation of Curcuma zedoaria
Roscoe and Zingiber zerumbet Smith, Biotechnology & Biotechnological
Equipment, 6(4), pp. 555-560.
147. Stanly C., Bhatt A., Keng C.L. (2010), A comparative study of Curcuma
zedoaria and Zingiber zerumbet plantlet production using different
micropropagation systems, African Journal of Biotechnology, 9(28), pp. 4326-
4333.

115
148. Sudha G., Ravishankar G.A. (2003), Influence of methyl jasmonate and
salicylic acid in the enhancement of capsaicin production in cell suspension
cultures of Capsicum frutescens Mill, Current Science, 85(8), pp. 1212-1217.
149. Syu W.J., Shen C.C., Don M.J., Ou J.C., Lee G.H., Sun C.M. (1998),
Cytotoxicity of curcuminoids and some novel compounds from Curcuma
zedoaria, Journal of Natural Products, 61, pp. 1531-1534.
150. Taguchi G., Yazawa T., Hayashida N., Okazaki M. (2001), Molecular
cloning and heterologous expression of novel glucosyltransferases from
tobacco cultured cells that have broad substrate specificity and are induced by
salicylic acid and auxin, Journal of Biochemistry, 268, pp. 4086-4094.
151. Tamimi S.M. (2015), Effects of ethylene inhibitors, silver nitrate
(AgNO3), cobalt chloride (CoCl2) and aminooxyacetic acid (AOA), on in vitro
shoot induction and rooting of banana (Musa acuminata L.), African Journal
of Biotechnology, 14(32), pp. 2510-2516.
152. Teixeira Da Silva J.A. (2013), The effect of ethylene inhibitors (AgNO3,
AVG), an ethylene-liberating compound (CEPA) and aeration on the formation
of protocorm-like bodies of hybrid Cymbidium (Orchidaceae), Frontiers in
Biology, 8(6), pp. 606-610.
153. Thach B.D., Le N.T.L., Diep T.C., Trinh T.B., Tran T.L.G., Nguyen
P.A.U., Ngo K.S. (2016), Protocol establishment for multiplication and
renegeration of Polycias fruticosa (L.) Harms. An inportant medicinal plant in
Viet Nam, European Journal of Biotechnology and Genetic Engineering, 3(1),
pp. 31-37.
154. Thanh N.T., Murthy H.N., Yu K.W., Hahn E.J., Paek K.Y. (2005), Methyl
jasmonate elicitation enhanced synthesis of ginsenoside by cell suspension
culture of Panax ginseng in 5L balloon type bubble bioreactor, Apply
Microbiology Biotechnology, 67, pp. 197-201.

116
155. Tohda C., Nakayama N., Hatanaka F., Komatsu K. (2006), Comparison
of antiinflammatory activities of six curcuma rhizomes: a possible curcuminoid
indepent pathway mediated by curcuma phaeocaulis extract, Evid Based
Complement Alternat Med, 3, pp. 255-260.
156. Tuan V.C., Hoang V.D., Loc N.H. (2011), Cell suspension culture of
zedoary (Curcuma zedoaria Roscoe), VNU Journal of Science, Natural
Sciences and Technology, 27, pp. 64-70.
157. Ullah H.M.A., Zaman S., Juhara F., Akter L., Tareq S.M., Masum E.H.,
Bhattacharjee R. (2014), Evaluation of antinociceptive, in-vivo & in-vitro anti-
inflammatory activity of ethanolic extract of Curcuma zedoaria rhizome, BMC
Complementary and Alternative Medicine, 14(1), pp. 346.
158. Walker T.S., Pal Bais H., Vivanco J.M. (2002), Jasmonic acid-induced
hypericin production in cell suspension cultures of Hypericum perforatum L.
(St. John's wort), Phytochemistry, 60(3), pp. 289-293.
159. Wang J., Qian J., Yao L., Lu Y. (2015), Enhanced production of
flavonoids by methyl jasmonate elicitation in cell suspension culture of
Hypericum perforatum, Bioresources and Bioprocessing, 2(1), pp. 5.
160. Wang S., Zhang S., Xiao A., Rasmussen M., Skidmore C., Zhan J. (2015),
Metabolic engineering of Escherichia coli for the biosynthesis of various
phenylpropanoid derivatives, Metabolic Engineering, 29, pp. 153-159.
161. Watanabe K., Shibata M., Yano S., Cai Y., Shibuya H., Kitagawa I.
(1986), Antiulcer activity of extracts and isolated compound from zedoary
(Gajustsu) cultivated in Yakushima (Japan), Yakugaku Zasshi, 106(12), pp.
1137-1142.
162. Wilson B., Abraham G., Manju V.S., Mathew M., Vimala B., Sundaresan
S., Nambisan B. (2005), Antibacterial activity of Curcuma zedoaria and
Curcuma malabarica tubers, Journal of Ethnopharmacology, 99, pp. 147-151.

117
163. Xie Z., Ma X., Gang D.R. (2009), Modules of co-regulated metabolites in
turmeric (Curcuma longa) rhizome suggest the existence of biosynthetic
modules in plant specialized metabolism, Journal of Experimental Botany,
60(1), pp. 87-97.
164. Xing Y. (1999), The chemical constituents of the essential oil from
Curcuma zedoaria (Christm) Rosc., Journal of essential oil-bearing plants
JEOP, 19(1), pp. 95-96.
165. Yeoman M.M., Meidzybrodzka M.B., Lindsey K., McLauchlan W.R.
(1980), The synthetic potential of cultured plant cells. Plant cell cultures:
results and perspectives. Amsterdam, Elsevier: 327-343
166. Yu L.J., Lan W.Z., Qin W.M., Xu H.B. (2001), Effects of salicylic acid
on fungal elicitor-induced membrane-lipid peroxidation and taxol production
in cell suspension cultures of Taxus chinensis, Process Biochemistry, 37(5), pp.
477-482.
167. Yu Z.Z., Fu C.X., Han Y.S., Li Y.X., Zhao D.X. (2005), Salicylic acid
enhances jaceosidin and siringin production in cell cultures of Saussurea
medusa, Biotechnology Letters, 28(13), pp. 1027-1031.
168. Zhang S., Liu N., Sheng A., Ma G., Wu G. (2011), In vitro plant
regeneration from organogenic callus of Curcuma kwangsiensis Lindl.
(Zingiberaceae), Plant Growth Regulation, 64(2), pp. 141-145.
169. Zhao J.L., Zhou L.G., Wu J.Y. (2010), Effects of biotic and abiotic
elicitors on cell growth and tanshinone accumulation in Salvia miltiorrhiza cell
cultures, Applied Microbiology and Biotechnology, 87(1), pp. 137-144.

a
PHỤ LỤC
Phụ lục 1 : Trình tự các gen trên ngân hang gen
LOCUS MF663785 1382 bp DNA linear PLN 22-JUL-2018
DEFINITION Curcuma zedoaria diketide-CoA synthase (DCS) gene, complete cds.
ACCESSION MF663785
VERSION MF663785.1
KEYWORDS .
SOURCE Curcuma zedoaria
ORGANISM Curcuma zedoaria
Eukaryota; Viridiplantae; Streptophyta; Embryophyta; Tracheophyta;
Spermatophyta; Magnoliophyta; Liliopsida; Zingiberales;
Zingiberaceae; Curcuma.
REFERENCE 1 (bases 1 to 1382)
AUTHORS Loc,N.H., Huy,N.D., Lan,T.T.P., Quan,L.V. and Nghi,N.V.
TITLE Identification and characterization of genes in curcuminoid pathway
of Curcuma zedoaria
JOURNAL Unpublished
REFERENCE 2 (bases 1 to 1382)
AUTHORS Loc,N.H., Huy,N.D., Lan,T.T.P., Quan,L.V. and Nghi,N.V.
TITLE Direct Submission
JOURNAL Submitted (12-AUG-2017) Institute of Bioactive Compounds and
Department of Biotechnology, College of Sciences, Hue University,
77 Nguyen Hue street, Hue, Thua Thien Hue 530000, Vietnam
COMMENT ##Assembly-Data-START##
Sequencing Technology :: Sanger dideoxy sequencing
##Assembly-Data-END##
FEATURES Location/Qualifiers
source 1..1382
/organism="Curcuma zedoaria"
/mol_type="genomic DNA"
/db_xref="taxon:136224"
gene <1..>1382
/gene="DCS"
/note="CzDCS"
mRNA join(<1..175,303..856,942..>1382)
/gene="DCS"
/product="diketide-CoA synthase"
CDS join(1..175,303..856,942..1382)
/gene="DCS"
/codon_start=1
/product="diketide-CoA synthase"
/protein_id="AXC59785.1"
/translation="MEANGYRITHSADGPATILAIGTANPTNVVDQNAYPDFYFRVTN
SEHLQELKAKFRRICEKAAIRKRHLYLTEEILRENPSLLAPMAPSFDARQAIVVEAVP
KLAKEAAEKAIKEWGRPKSDITHLVFCSASGIDMPGSDLQLLKLLGLPPSVNRVMLYN
VGCHAGGTALRVAKDLAENNRGARVLAVCSEVTVLSYRGPHPAHIESLFVQALFGDGA
AALVVGSDPVDGVERPIFEIASASQVMLPESEEAVGGHLREIGLTFHLKSQLPSIIAS
NIEQSLTTACSPLGLSDWNQLFWAVHPGGRAILDQVEARLGLEKDRLAATRHVLSEYG
NMQSATVLFILDEMRNRSAAEGHATTGEGLDWGVLLGFGPGLSIETVVLHSCRLN"
ORIGIN
1 atggaagcga acggctaccg cataactcac agcgccgacg ggccggcgac gatcttggcc
61 atcggcaccg ccaaccccac caacgtcgtc gaccagaacg cttatcccga cttctatttc
121 cgggtcacca actccgagca tctgcaggaa ctcaaagcca agtttaggcg catctgtaag

b
181 atttatgtag tttgattgaa tgatgggtcg taattcaaac agaatgatcg atgatcatgt
241 taggaattgg tggtttggtg gatttatgtg ggtgcgtgtg acctgttcga ttacatgtgt
301 aggtgagaaa gcggcgatca ggaagaggca cttgtacttg actgaggaga ttttgcggga
361 gaatcctagc ttgctggctc ccatggcgcc gtcgttcgac gcgcggcagg cgatcgtggt
421 ggaggcggtg ccgaagctag cgaaggaggc ggcggagaag gcgatcaagg agtggggtcg
481 ccccaaatcg gacatcacgc acctcgtctt ctgctccgcg agcggaatcg acatgcccgg
541 ctccgacctg cagcttctca agctgctcgg gctcccgccg agcgtcaatc gcgtcatgct
601 ctacaacgtc gggtgccacg ccggtggcac cgccctccgc gtcgccaagg acctcgcgga
661 gaacaaccgc ggcgcgcggg tgctcgccgt ctgctccgag gtcaccgtgc tctcctaccg
721 cggcccccac cccgcccaca tcgagagcct cttcgtccaa gctctgtttg gcgacggcgc
781 tgccgcgctc gtggtcgggt ccgaccccgt cgatggcgtc gagcgcccca tcttcgaaat
841 cgcctcggca tcccaagtaa tcgctgctcc agtcgaaatt aaccaaaatt cgatcttttg
901 atatcgaacg aactttaaaa atcaaatctt tggcaccgca ggtgatgctt ccggagagcg
961 aagaggcggt aggcggccac ctccgcgaaa ttgggctgac cttccacctc aagagccagc
1021 ttccgtcgat catcgcgagc aacatcgagc agagcctgac gactgcgtgc tcgccgctgg
1081 ggctgtcgga ctggaaccag ctgttctggg cggttcaccc cggcggccga gcgatcctgg
1141 accaggtgga ggcgcggctc ggactggaga aggaccggct cgccgcgacg cggcacgtac
1201 tcagcgagta cggcaacatg cagagcgcca cggtgctgtt catcctggac gagatgcgga
1261 accgctcggc tgcggagggc cacgccacca ccggcgaggg gctcgactgg ggcgtgcttt
1321 tgggcttcgg cccgggactc tccatcgaga ccgtcgtcct ccatagttgc agactgaact
1381 ag
//

c
LOCUS MF402846 1240 bp DNA linear PLN 23-MAY-2018
DEFINITION Curcuma zedoaria curcuminoid synthase 1 (CURS1) gene, complete cds.
ACCESSION MF402846
VERSION MF402846.1
KEYWORDS .
SOURCE Curcuma zedoaria
ORGANISM Curcuma zedoaria
Eukaryota; Viridiplantae; Streptophyta; Embryophyta; Tracheophyta;
Spermatophyta; Magnoliophyta; Liliopsida; Zingiberales;
Zingiberaceae; Curcuma.
REFERENCE 1 (bases 1 to 1240)
AUTHORS Loc,N.H., Huy,N.D., Lan,T.T.P., Quan,L.V. and Nghi,N.V.
TITLE Identification and characterization of genes in curcuminoid pathway
of Curcuma zedoaria Roscoe
JOURNAL Unpublished
REFERENCE 2 (bases 1 to 1240)
AUTHORS Loc,N.H., Huy,N.D., Lan,T.T.P., Quan,L.V. and Nghi,N.V.
TITLE Direct Submission
JOURNAL Submitted (26-JUN-2017) Institute of Bioactive Compounds and
Department of Biotechnology, College of Sciences, Hue University,
77 Nguyen Hue street, Hue, Thua Thien Hue 530000, Vietnam
FEATURES Location/Qualifiers
source 1..1240
/organism="Curcuma zedoaria"
/mol_type="genomic DNA"
/db_xref="taxon:136224"
gene <1..>1240
/gene="CURS1"
mRNA join(<1..178,249..>1240)
/gene="CURS1"
/product="curcuminoid synthase 1"
CDS join(1..178,249..1240)
/gene="CURS1"
/EC_number="2.3.1.217"
/codon_start=1
/product="curcuminoid synthase 1"
/protein_id="AWK77185.1"
/translation="MANLHALRREQRAQGPATIMAIGTATPPNLYEQSTFPDFYFRVT
NSDDKQELKKKFRRMCDKTMVKKRYLHLTEEILKERPKLCSYKEASFDDRQDIVVEEI
PRLAKEAAEKAIKEWGRPKSEITHLVFCSISGIDMPGADYRLATLLGLPLTVNRLMIY
SQACHMGAAMLRIAKDLAENNRGARVLVVACEITVLSFRGPNEGDFEALAGQAGFGDG
AGAVVVGADPLEGIEKPIYEIAAAMQETVAESQGAVGGHLRAFGWTFYFLNQLPAIIA
DNLGRSLERALAPLGVTEWNDVFWVAHPGNWAIMDAIEAKLQLSPDKLSTARHVFTEY
GNMQSATVYFVMDELRKRSAVEGRSTTGDGLQWGVLLGFGPGLSIETVVLRSMPL"
ORIGIN
1 atggccaacc tccacgcgtt gcgcagggag cagagggctc aaggtcccgc caccatcatg
61 gccatcggga cggccacccc ccccaacctc tacgagcaga gcaccttccc ggacttctac
121 ttccgcgtca ccaactccga cgacaagcag gagctcaaga aaaagttccg ccgcatgtgt
181 aagtaatcga tcgatgatcc gttcagttga tgtaattcga tcggtgattg atttggagaa
241 gtaaataggc gataagacga tggtgaagaa gcggtacctg cacttgaccg aggagatcct
301 gaaggagagg cccaagctct gctcctacaa ggaggcgtcg ttcgacgacc ggcaggacat
361 cgtggtggag gagataccga gattggctaa agaagcggcg gagaaggcca tcaaggagtg
421 ggggcggccc aaatcggaga tcacccacct ggtcttctgt tccatcagcg ggatcgacat
481 gcccggcgcc gactaccgcc tcgccacgct cctcggcctc cctctcaccg tcaaccgcct
541 catgatctac agccaggcct gccacatggg cgccgccatg ctccgcatcg ccaaggacct
601 cgccgagaac aacaggggcg cgcgcgtgct ggtggtcgcc tgcgagatca ccgtgctcag
661 cttccgcggc ccgaacgagg gcgacttcga ggcgctcgcg gggcaggccg gcttcggcga

d
721 cggcgcgggg gccgtcgtcg tcggggccga cccgctggaa ggaattgaaa aacccatcta
781 cgagatcgcg gcggcgatgc aggagacggt ggcggagagc cagggggcgg tgggcggcca
841 cctgcgggcg ttcggctgga cgttctactt cctgaaccag ctgccggcga tcatcgccga
901 caacctcggg aggagcctgg agcgggcgtt ggcgccgctg ggggtgacgg agtggaacga
961 cgtcttctgg gtggcgcacc cgggcaactg ggccatcatg gacgccatcg aagccaagct
1021 gcagctgagc ccggacaagc tcagcaccgc ccgccacgtc ttcacagagt acggcaacat
1081 gcagagcgcc accgtgtact tcgtgatgga tgagctgagg aagcggtcgg cggtggaggg
1141 gcggagcacc accggagacg gcttgcagtg gggagttctc ctcggttttg ggccgggcct
1201 cagcatcgag accgttgtac tgcgcagtat gccactgtag
//

e
LOCUS MF402847 1288 bp DNA linear PLN 23-MAY-2018
DEFINITION Curcuma zedoaria curcuminoid synthase 2 (CURS2) gene, complete cds.
ACCESSION MF402847
VERSION MF402847.1
KEYWORDS .
SOURCE Curcuma zedoaria
ORGANISM Curcuma zedoaria
Eukaryota; Viridiplantae; Streptophyta; Embryophyta; Tracheophyta;
Spermatophyta; Magnoliophyta; Liliopsida; Zingiberales;
Zingiberaceae; Curcuma.
REFERENCE 1 (bases 1 to 1288)
AUTHORS Loc,N.H., Huy,N.D., Lan,T.T.P., Quan,L.V. and Nghi,N.V.
TITLE Identification and characterization of genes in curcuminoid pathway
of Curcuma zedoaria Roscoe
JOURNAL Unpublished
REFERENCE 2 (bases 1 to 1288)
AUTHORS Loc,N.H., Huy,N.D., Lan,T.T.P., Quan,L.V. and Nghi,N.V.
TITLE Direct Submission
JOURNAL Submitted (26-JUN-2017) Institute of Bioactive Compounds and
Department of Biotechnology, College of Sciences, Hue University,
77 Nguyen Hue street, Hue, Thua Thien Hue 530000, Vietnam
COMMENT ##Assembly-Data-START##
Sequencing Technology :: Sanger dideoxy sequencing
##Assembly-Data-END##
FEATURES Location/Qualifiers
source 1..1288
/organism="Curcuma zedoaria"
/mol_type="genomic DNA"
/db_xref="taxon:136224"
gene <1..>1288
/gene="CURS2"
mRNA join(<1..184,297..>1288)
/gene="CURS2"
/product="curcuminoid synthase 2"
CDS join(1..184,297..1288)
/gene="CURS2"
/EC_number="2.3.1.217"
/codon_start=1
/product="curcuminoid synthase 2"
/protein_id="AWK77186.1"
/translation="MAMISLQAMRKAQRAQGPATILAVGTANPPNLYEQDTYPDYYFR
VTNSEHRQELKNKFRRMCEKTMVKRRYLYLTPEILKERPKLCSYMEPSFDDRQDIVVE
EVPKLAAEAAEKAIKEWGGEKSAITHLVFCSISGIDMPGADYRLAKLLGLPLAVNRLM
LYSQACHMGAAMLRIAKDIAENNRSARVLVVACEITVLSFRGPDERDFQALAGQAGFG
DGAGAMIVGADPVLGVERPLYHIMSATQTTVPESEKAVGGHLREVGLTFHFFNQLPAI
IADNVGNSLAEAFEPIGIKDWNNIFWVAHPGNWAIMDAIETKLGLEQSKLATARHVFS
EFGNMQSATVYFVMDELRKRSAAENRATTGDGLRWGVLFGFGPGISIETVVLQSVPL"
ORIGIN
1 atggcgatga tcagcttgca ggcgatgcgc aaggcgcaga gagctcaagg tccggccacc
61 atcttggccg tcggcaccgc caacccgccc aatctctacg agcaggacac gtatcccgac
121 tactacttcc gcgtcaccaa ctccgagcac aggcaggagc tcaagaacaa gttccgccgc
181 atgtgtaagt tcttcgatcg atcctttttt ggttttcgaa aaaagtgagt ttttaatggg
241 aaaagcagag gatcgaggtg gaacgggagg attgatttgg tttaatttga ttgcaggcga
301 gaagacgatg gtgaagaggc ggtatcttta cctgacgccg gagatcctga aggagcggcc
361 gaagctgtgc tcgtacatgg agccgtcgtt cgacgaccgg caggacatcg tggtggagga
421 ggtgccgaag ctggccgcgg aggcagcgga gaaggccatc aaggagtggg gcggcgagaa
481 gtcggccatc acccacctgg tcttctgctc catcagcggc atcgacatgc ccggagctga

f
541 ctaccgcctc gccaagctcc tcgggctccc gctcgccgtc aaccgcctga tgctctacag
601 ccaggcctgc cacatgggcg ccgccatgct gcgcatagcc aaggacatcg ccgagaacaa
661 ccgctccgct cgcgtcctcg tcgtcgcctg cgagatcacc gtgctcagct tccgcggccc
721 ggacgagcgc gacttccagg cgctggccgg ccaggccggc ttcggggacg gcgccggcgc
781 gatgatcgtc ggggccgacc ccgtcctcgg cgtcgagcgg ccgctctacc acatcatgtc
841 ggcgactcag acgacggtgc cggagagcga gaaggcggtg gggggccacc tccgcgaggt
901 ggggctgacc ttccacttct tcaaccagct gccggcgatc atcgccgaca acgtggggaa
961 cagcctggcg gaggcgttcg aaccgatcgg gatcaaggac tggaacaaca tcttctgggt
1021 ggcgcacccg ggcaactggg ccatcatgga cgccatcgag accaagctgg gcctggaaca
1081 gagcaagctg gccaccgcac gccacgtctt ctccgagttc ggcaacatgc agagcgccac
1141 cgtctacttc gtgatggacg agctcaggaa acggtcggcg gcggagaacc gggcgaccac
1201 cggcgacggg ctccggtggg gcgtgctctt cggcttcggc cccggcatca gcatcgaaac
1261 cgtcgtgctc caaagcgtgc cgctttag
//

g
LOCUS MF987835 1265 bp DNA linear PLN 01-SEP-2018
DEFINITION Curcuma zedoaria curcumin synthase 3 (CURS3) gene, complete cds.
ACCESSION MF987835
VERSION MF987835.1
KEYWORDS .
SOURCE Curcuma zedoaria
ORGANISM Curcuma zedoaria
Eukaryota; Viridiplantae; Streptophyta; Embryophyta; Tracheophyta;
Spermatophyta; Magnoliophyta; Liliopsida; Zingiberales;
Zingiberaceae; Curcuma.
REFERENCE 1 (bases 1 to 1265)
AUTHORS Loc,N.H., Huy,N.D., Truong,L.T.P., Luong,N.N., Tan,T.H., Quan,L.V.
and Nghi,N.V.
TITLE Identification and characterization of several genes in curcuminoid
pathway of Curcuma zedoaria
JOURNAL Unpublished
REFERENCE 2 (bases 1 to 1265)
AUTHORS Loc,N.H., Huy,N.D., Truong,L.T.P., Luong,N.N., Tan,T.H., Quan,L.V.
and Nghi,N.V.
TITLE Direct Submission
JOURNAL Submitted (19-SEP-2017) Institute of Bioactive Compounds and
Department of Biotechnology, College of Sciences, Hue University,
77 Nguyen Hue street, Hue, Thua Thien Hue 530000, Vietnam
COMMENT ##Assembly-Data-START##
Sequencing Technology :: Sanger dideoxy sequencing
##Assembly-Data-END##
FEATURES Location/Qualifiers
source 1..1265
/organism="Curcuma zedoaria"
/mol_type="genomic DNA"
/db_xref="taxon:136224"
gene <1..>1265
/gene="CURS3"
/note="CzCURS3"
mRNA join(<1..178,271..>1265)
/gene="CURS3"
/product="curcumin synthase 3"
CDS join(1..178,271..1265)
/gene="CURS3"
/EC_number="2.3.1.217"
/codon_start=1
/product="curcumin synthase 3"
/protein_id="AXQ39866.1"
/translation="MGSLQAIRRAQRAQGPATIMAVGTSNPPNLYEQTSYPDFYFRVT
NSDHKHELKNKFRVICEKTKVKRRYLHLTEEILKQRPKLCSYMEPSFDDRQDIVVEEI
PKLAKEAAEKAIKEWGRPKSEITHLVFCSISGIDMPGADYRLATLLGLPLSVNRLMLY
SQACHMGAQMLRIAKDLAENNRGARVLAVSCEITVFSFRGPDAGDFEALACQAGFGDG
AAAVVVGADPLPGVERPIYEIAAAMQETVPESERAVGGHLREIGWTFHFFNQLPKLIA
ENIEGSLARAFKPLGISEWNDVFWVAHPGNWGIMDAIETKLGLEQGKLATARHVFSEY
GNMQSATVYFVMDEVRKRSAAEGRATTGEGLEWGVLFGFGPGLTIETVVLRSVPLP"
ORIGIN
1 atgggcagcc tgcaggcgat tcgcagagca cagcgggctc aaggcccggc caccatcatg
61 gctgtcggca cctccaaccc gcctaacctc tacgagcaga cctcctaccc cgacttctat
121 ttccgcgtca ccaactccga ccacaagcac gagctcaaga acaaattccg tgttatctgg
181 aattaattcg cacccatctt ttggttattg atcagttcga tttaaattag ggagttagtt
241 aattaataat attattatta ttctaaatag gtgagaagac gaaggtgaaa agacggtact
301 tgcacttgac ggaggagatc ctgaagcaga ggcccaagct ctgctcctac atggagccct

h
361 ccttcgacga ccggcaggac atcgtggtgg aggagatacc gaagctggcg aaggaggcgg
421 cggaaaaggc gatcaaggaa tggggccgcc ccaagtcgga gatcacccac ttggtgttct
481 gctccatcag cggyatcgac atgcccggcg ccgattaccg cctcgccacg ctcctcggcc
541 tccccctgtc cgtcaaccgc ctcatgctct acagccaggc ctgccacatg ggcgcgcaga
601 tgctgcgcat agccaaggac ctcgcggaga acaaccgggg cgcgcgcgtc ctggccgtct
661 cctgcgaaat caccgtgttc agcttccgcg gtcccgacgc gggcgacttc gaggccctcg
721 cgtgtcaggc cggcttcggc gacggtgccg ctgccgtcgt cgtcggggcc gaccccctcc
781 cgggcgtcga gaggcccatc tacgagatcg cggcggcgat gcaggaaacg gtgccggaga
841 gcgagagggc ggtggggggc cacctgaggg aaatcggctg gaccttccac ttcttcaacc
901 agctgccgaa gctgatcgcg gaaaacatcg agggcagcct ggcgcgggcg ttcaagccgc
961 tggggatcag cgagtggaac gacgtgttct gggtggcgca cccggggaat tggggcatca
1021 tggacgccat cgagaccaag ctggggctgg aacaggggaa gctcgccacg gcgcgccacg
1081 tcttcagcga gtacggaaac atgcagagcg ccaccgtgta cttcgtgatg gacgaggtga
1141 ggaagcggtc ggcggcggag gggcgggcca ccaccggcga aggcctggag tggggagtgc
1201 tgtttgggtt tggcccaggc ctcaccatag aaactgtcgt gctacgcagt gtaccattac
1261 cgtag
//

Phụ lục 2. Đường chuẩn curcumin
Hình 1. Đường chuẩn curcumin dựa trên kết quả HPLC

Phụ lục 3. Hình ảnh thí nghiệm
Hình 2: Tế bao nghệ đen để lắng trong bình tam giác 250 ml
Hình 3: Cây nghệ đen in vitro

Hình 4: Tế bao Callus nghệ đen

Hình 5: Nuôi cấy tế bao nghệ đen trong bình tam giác 250ml đặt trên máy
lắc
Hình 6: Sinh khôi tươi tế bao nghệ đen sau 14 ngay nuôi cấy

Hình 7: Cây nghệ đen tự nhiên