nature, - pnas.org · that the as the of these are found 1
TRANSCRIPT

VOL. 53, 1965 ZOOLOGY: E. J. DUPRAW 161
21 Counce, S., P. Smith, R. Barth, and G. D. Snell, Ann. Surg., 144, 198 (1956).22Gorer, P. A., J. Pathol. Bacteriol., 47, 231 (1938).23 Davies, D. A. L., Transplantation, 1, 201 (1963).
THE ORGANIZATION OF NUCLEI AND CHROMOSOMESIN HONEYBEE EMBRYONIC CELLS*
BY E. J. DUPRAW
DEPARTMENT OF ZOOLOGY, UNIVERSITY OF CALIFORNIA (DAVIS)
Communicated by Carroll M. Williams, November 4, 1964
Electron microscopy of thin-sectioned material has provided little insight intothe macromolecular organization of nuclei and chromosomes. Recently a "wholemount" technique, originally used by Kleinschmidt' with bacteria and viruses, hasbeen applied by several authors2-4 to nucleated cells. In this technique, entire cellsare spread on the air-water interface of a Langmuir trough, picked up directly on acoated grid, dehydrated, and dried by the critical point method of Anderson.5Such preparations, which may or may not be fixed and stained, make possibledirect examination of the fine structure of entire metaphase chromosomes and ofwhole interphase nuclei.Examination of a wide range of material by this method2 4 has shown that both
nuclei and chromosomes contain a system of long, irregularly organized fibers,whose diameter according to most authors varies around 250 A; the occurrence ofthis approximate dimension across a range of both vertebrates and invertebrateshas been impressive. Similar fibers have recently been demonstrated in thin-sectioned calf thymus nuclei,6 and evidence is accumulating that the chromosomalfibers are nucleoprotein in nature, 6 that they are the sites of messenger RNAsynthesis,6 and that they are organized as a central core surrounded by a sheath ofdistinctly different physical and chemical properties.2' 4
In the work reported here, the Kleinschmidt technique has been applied to em-bryonic cells of the honeybee.7 This material is especially suitable for whole mountelectron microscopy, due to the remarkably small size of the chromosomes (whoselengths and diameters are less than those of most vertebrate mitochondria) and tothe correspondingly low concentration of chromatin in the interphase nuclei.
Materials and Methods.-Interphase nuclei were prepared by squashing 1-4 stage 6 ("gastrula-tion" stage) honeybee embryos on the tip of a glass microtool, which was then touched to thesurface of a Langmuir trough containing 0.25 M sucrose. The spread material was picked up bytouching the sucrose surface lightly with the coated side of a formvar carbon-coated grid. Gridswere then washed by floating them face-down on distilled water for 5 min or less, and were mountedin a plastic grid holder under 25% ethanol. Holder and grids were dehydrated through an ethanolseries (5 min each in 35, 50, 70, 95, 100, 100, 100%), and then passed through three baths of amylacetate (10 min in each). Finally, the grids were placed in the pressure chamber of the Andersoncritical point drying apparatus, in which the amyl acetate is replaced by liquid CO2 under pressureand at reduced temperature (an ice bath); drying occurs instantly and with minimum distortionwhen the temperature is raised above the critical point (31.10C).Chromosomes were obtained by injecting late stage 2 (cleavage stage) honeybee embryos with
10-3 M colchicine (N.B. Co.), incubating the embryos at 35.50C for one hr, and then squashing

162 ZOOLOGY: E. J. DUPRAW PROC. N. A. S.
them for surface spreading as above. Subsequent treatment was identical to that for interphasenuclei.Some interphase nuclei were partially digested with trypsin. In these experiments, spread ma-
terial was picked up on grids, which were then treated for 3-7 min with 0.001 % trypsin (Wor-thington, 1 X crystallized) in Tris buffer at pH 8.0. In other experiments, grids treated withtrypsin in this way were afterwards digested for 5-10 min in 0.01 % DNAase (Worthington, beefpancreas, 1 X crystallized) in 0.0025 M phosphate buffer at pH 7.32 and with 0.00025 M mag-nesium ion added. Following enzyme digestion, grids were mounted in the holder under 25%ethanol, dehydrated, and dried as above.
In some control preparations, cells were spread on glass-distilled water instead of on sucrosesolution; in other preparations, material was fixed, immediately after being picked up on the grid,in either 10% formalin or glutaraldehyde buffered at pH 7.0. Such variations in technique donot change the basic aspect of the interphase fibers, and in most preparations, fixation has beenomitted.
Preparations were viewed and photographed with the RCA EMU-3E; size measurements werestandardized against two carbon grating replicas having 28,800 and 54,800 lines to the inch, re-spectively. All dimensions were measured from the "in-focus" image of a through-focal series,ordinarily made at 0.25-/A intervals and photographed after careful normalization of the lenses.
Results.-On grids prepared by the above technique, interphase nuclei are abun-dant (20 or more per grid), readily recognizable, and, particularly in the case ofembryonic honeybee material, remarkably undistorted in their gross morphology.An entire nucleus is illustrated in Figure 1, complete with its nuclear envelope andchromatin contents, which have spilled out from the partially retracted envelope.The entire spectrum of spreading has been observed in different preparations, fromtotally unruptured nuclei to nuclei whose envelopes have been completely strippedaway and lost.The chromatin contents, when seen at higher magnification (Fig. 2), appear as
long, spaghetti-like fibers, which look more or less alike and which have distinc-tively "bumpy" profiles. The pattern and electron density of bumps are fre-
FIG. 1.-An entire interphase nucleus, prepared by the Kleinschmnidt technique. Arrowindicates chromatin contents, which have "spilled out" from the hemispherical nuclearenvelope. 3840 X.

VOL. 53, 1965 ZOOLOGY: E. J. DUPRAW 163
quently suggestive of a loose, irregular coil, and in some preparations, it is commonto find fibers which are twisted into distinct loops (including both left- and right-handed configurations); consequently, the "bumpiness" of the chromatin fibersis believed to represent an irregular helical twist in the fiber structure. Theapparent diameter of the fibers has a mode around 230-250 A, but it varies fromless than 180 A to over 400 A, depending on the degree of stretching and "bumpi-ness." If the wider regions are regarded as due to twists in an otherwise uniformfiber, the diameter of the "fundamental" fiber is very possibly less than 200 A.Approximately the same dimensions have been observed for drone (haploid) nuclei,worker (diploid) nuclei, nuclei from early and late embryonic stages, and for thefibers of metaphase chromosomes. Clear-cut examples of "paired" fibers, or of100 A substrands, as reported by Ris,3 have not been found. Occasional instancesof parallel fibers, such as those illustrated by Wolfe,4 have been seen, but theseare regarded as purely random, or accidental, fiber configurations.
After colchicine treatment of embryonic stages in near-synchronous mitosis,grids contain numerous entire chromosomes (2N = 32), which are frequently paired,usually occur in groups, and most often measure about 0.3 in diameter with lengthsvarying from less than 1 I to about 3 IA (Fig. 3). Here, too, all degrees of stretchinghave been found, from the most compact, typical metaphase pairs to highly "un-raveled" forms (Fig. 4). In all preparations, the chromosomes appear to be com-posed of fibers having the same dimension and general aspect as the interphasechromatin (Fig. 2), and in highly unraveled material it is apparent that:
1. The entire body of the chromosome consists of such fibers.2. The fibers are much longer than the chromosomes, so that a given chromosome
may contain very few.3. In gross appearance, each chromosome (or chromatid) seems to be a struc-
FIG. 2-hInterphase "chromatin" fibers, as seen at higher magnification. Note the"bumpiness" of fibers, which in some places is suggestive of a helical twist (arrow). 61,900X.

164 ZOOLOGY: E. J. DUPRAW PROC. N. A. S.
0~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.
FIG.3.-Anentiremitoticchromosome, fixed in 10% formalin. This chromosome measures~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.about2.4~~~~ainlength,by 0.6 s in diameter. Arrow indicates folding of the fibers, whichare~~~~~~~~~~~~~~~~~I_
FIG. 5.-Strentchedmtoin cl roenveopewt nui oeteboefiben1%frmln hscroendsoamte medguesoFIGt .46.Ainunroengtbatacmntodacrmatin. fibrtowidctheseodgegofa annhu (abrrow).c48,20
tuilra unitdime.,iontoisnotorgaiedchoainto oethbeneovisiblechronieemadiomesnot havebavismaibydoubl2s2hrmtd"srctr7n0niipto0ouurXiisos
4I. Withi eacho mtphschromosome,h ies arc inriatelye accdndtappasrentlyeirrgu
5.nrTheredisNotthiniationd ofrangorerlyo she cndryoroomstetarytera"olngth(asditinc
From foldingubofeathe achront fahoatinfibers. h deo nanuuarw.40
In gethnea easpcttheioieahschromosomes ofthersareihoneybeeatiapparn rear-

VOL. 53, 1965 ZOOLOGY: E. J. DUPRAW 165
to be compact, but more or less irregular "packages" of interphase fibers, whichhave "condensed" mainly by folding.The nuclear envelope is frequently present in interphase material as a stretched
and flattened membrane (Fig. 5). In these preparations, the annuli appear asdoughnut-shaped thickenings in the membrane, their diameters varying con-siderably to either side of 1000 A, and the membrane itself is usually continuousacross the center of the "doughnut" (except when it is seen to have been stretchedand broken). Thus, in well-preserved material, the annuli are clearly not holes or"pores," but a specialized substructure of the envelope. In unstretched en-velopes, the annuli average about 800 A in total diameter, while the center of the"doughnut" measures only about 150 A.
It has been observed repeatedly that the chromatin fibers of the interphasenucleus are attached to the nuclear envelope, typically at the edge of an annulus(Fig. 6). In preparations where the envelope has been stripped away from thechromatin, the broken ends of these fiber attachments are frequently found ad-hering to the annuli (Fig. 5). The presence of such broken fiber ends (which infact led to the search for unbroken attachments) constitites good evidence, boththat the fiber system represents the real structure of the interphase nucleus, andthat the attachments to the annuli are structural rather than accidental in nature.In some cases these attachments seem to be terminal (i.e., at the fiber end), butother configurations have been seen which suggest that the attachments can occuralong the length of a fiber as well.With brief digestion in low concentrations of trypsin, the interphase nuclei re-
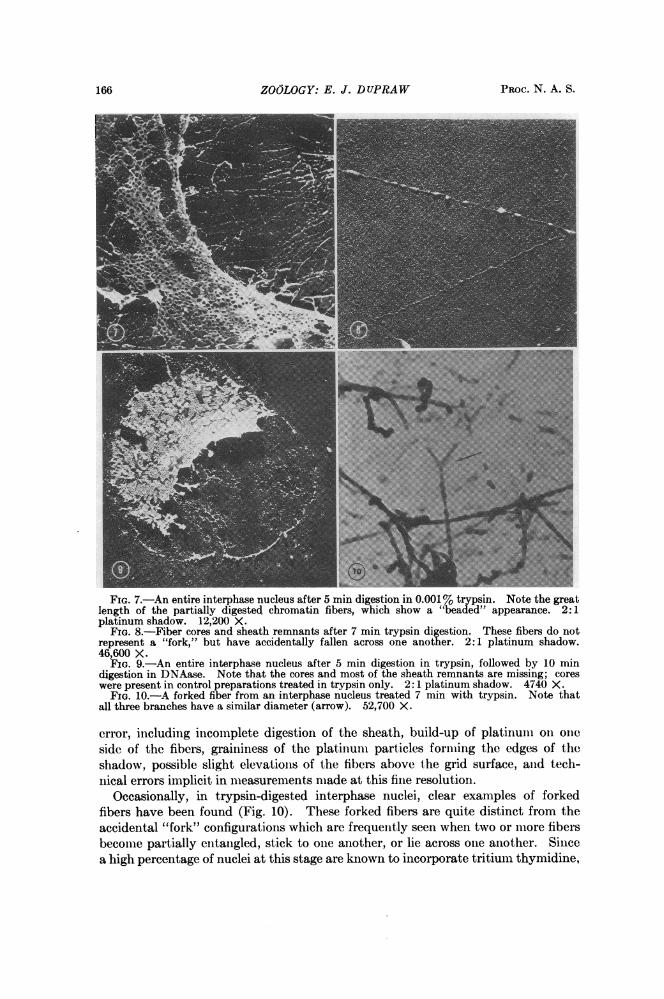
main morphologically recognizable (Fig. 7); however, the central membranes of theannuli are usually digested away, so that the annuli now appear as actual holes inthe nuclear envelope. The interphase fibers are also more or less diminished indiameter, many of them show a "beads on a string" appearance, and in geqeral, thefibers give the impression of having "sprung out," i.e., they have lost much of theirtangled appearance and can now be traced in relatively straight courses for distancesof several micra. The great length of the fibers in trypsin-digested nuclei confirmsthe evidence for very long fibers in chromosomes which have been unraveled bystretching (Fig. 4).
Individual trypsin-digested fibers, when seen at higher magnification (Fig. 8),show a long, trypsin-resistant core, to which adhere remnants of a partially digestedsheath (thereby giving rise to the "beaded" appearance). In preparations which arefirst treated with trypsin and then with DNAase, both the cores and most of thesheath remnants are removed, while recognizable nuclear envelopes remain (Fig. 9).It appears that the cores are digested by DNAase and that in the absence of coresthe sheaths lose their structural attachments to the envelope and wash off the grids.This interpretation is supported by the finding of some fiber remnants having theaspect of coreless sheaths.The diameters of the cores have been measured, both directly in the horizontal
dimension, and from 2:1 platinum shadows for the vertical dimension. In bothcases, diameters as low as 23-26 A are common; however, most cores measure30-50 A. These dimensions are consistent with an interpretation of the cores assingle Watson-Crick DNA molecules. Discrepancies between the expected diami-eter of 20 A and the observed diameters are accounted for by various sources of
166 ZOOLOGY: E. J. DUPRAW PROC. N. A. S.
FIG. 7.-An entire interphase nucleus after 5 min digestion in 0.001% trypsin. Note the greatlength of the partially digested chromatin fibers, which show a "beadted" appearance. 2:1platinum shadow. 12,200 X.
FIG. 8.-Fiber cores and sheath remnants after 7 min trypsin digestion. These fibers do notrepresent a "fork," but have accidentally fallen across one another. 2:1 platinum shadow.46,600 X.
FIG. 9.-An entire interphase nucleus after 5 min digestion in trypsin, followed by 10 mindigestion in DNAase. Note that the cores and most of the sheath remnants are missing; coreswere present in control preparations treated in trypsin only. 2: 1 platinum shadow. 4740 X.
FIG. 10.-A forked fiber from an interphase nucleus treated 7 min with trypsin. Note thatall three branches have a similar diameter (arrow). 52,700 X.
error, including incomplete digestion of the sheath, build-up of platinum oin omieside of the fibers, graininess of the platmuni particies forming the edges of theshadow, possible slight elevations of the fibers above the grid surface, aind tech-nical errors implicit in nieasurements made at this fine resolution.
Occasionally, in trypsin-digested interphase nuclei, clear examples of forkedfibers have been found (Fig. 10). These forked fibers are quite distinct from theaccidental "fork" configurations which are frequently seen when two or mnore fibersbecome partially entaiigled, stick to one another, or lie acioss one another. Sincea high percentage of nuclei at this stage are known to incorporate tritium thymidine,

VOL. 53, 1965 ZOOLOGY: E. J. DUPRAW 167
it is suggested that the forked fibers may represent sites of DNA replication and thateach fork may correspond to a "replication fork" in the DNA molecule at the coreof the fiber.Discussion.-The most striking aspect of nuclear and chromosomal organization,
as revealed by whole-mount electron microscopy, is the apparent irregularity of thefiber systems. In the earliest work, it was natural to suppose that this irregularitywas the result of preparation artifacts. However, the reality of the chromosomaland interphase fibers, as such, has now been well established: several laboratorieshave reported them,2-4' 6 they occur with or without fixation in formalin or glutar-aldehyde, and, as reported here, they leave their broken ends firmly attached tothe annuli of stripped-off nuclear envelopes. Moreover, the superficially irregulararrangement of fibers has been observed in the mitotic chromosomes of a widevariety of species.4 Although it is conceivable that chromosomal organization ismuch more orderly than it looks, the possibility remains that the genetic material isarranged in some manner which does not require a precise, three-dimensionalorientation.
This paper reports the discovery of very long, single, DNAase-sensitive, trypsin-resistant filaments (i.e., cores) in the nuclei of honeybee embryonic cells; the di-mensions of these filaments are consistent with the expected diameters of singleWatson-Crick DNA molecules. Experimental evidence for the existence of sin-gle long DNA molecules has also been found by Gall,8 Callan,' Swift,'0 Taylor,"and others, using dipteran giant salivary chromosomes and amphibian oocyte lamp-brush chromosomes. If the filaments in honeybee embryonic cells are indeed sin-gle DNA molecules, the organization of DNA in these cells would be fundamentallysimilar to the DNA of viruses and bacteria, which has been shown by Cairns,'2Kleinschmidt, 1 and others to consist of single molecules tens or hundreds of micronslong.
In microorganisms these long, single DNA molecules act as independent func-tional units, each one replicating sequentially along its length at a "replicationfork" (as postulated by Watson and Crick'3). If this also proves to be true inhigher organisms (cf. Fig. 10), it would be consistent with the mass of genetic andbiochemical data to suppose that the folding of the molecules in the mitotic chro-mosome is indeed irregular, bearing only a casual relationship to the linear organiza-tion of the genetic material. In this respect, mitotic chromosomes might be ex-pected to differ in organization from meiotic chromosomes (which are charac-terized by the presence of a "synaptinemal complex") and from atypical "synthesiz-ing" chromosomes (such as the giant salivary and lampbrush chromosomes).
In view of the observations reported here, the most likely interpretation of theinterphase and chromosomal fibers is that they are single DNA molecules, eachenclosed in a proteinaceous sheath. Presumably the sheath contains histones,which are known to be associated quantitatively with DNA, and it may also con-tain DNA and RNA polymerase, as well as the relatively insoluble, acid proteinfraction of cell nuclei. i\Iazia'4 has shown that, at certain stages in the develop-ment of the sea urchin, DNA polymerase activity is specifically bound to thenuclei (including fragmented nuclei), and doubles when the chromosome numberis doubled in artificially induced tetraploids; DNA polymerase has also beenfound closely associated with nuclei or with DNA itself in several other systems.'5

168 ZOOLOGY: E. J. DUPRAW PROC. N. A. S.
The possibility exists that histones, which apparently form complexes with acidproteins as well as with DNA,16 function as a structural matrix in the sheath,while control of DNA and RNA synthesis is effected through the organization ofpolymerases and other enzymes into functional molecular arrays.'4 Parts of thesheath lacking in RNA polymerase, for instance, could account for "geneticallyinert" regions on a cross-over map, and might act as stop signals for the readingof genetic information in differentiated cells.The sheath may also contain part or all of the mechanism responsible for folding
the chromatin fibers into the metaphase configuration; regions of "contractile"protein, located at characteristic intervals and capable of shortening characteristicamounts, could account for reproducible patterns of chromatin condensation withina species (including the occurrence of "heterochromatic" chromosomes and chro-mosomal regions). This seems the more likely since no mechanism is known bywhich DNA itself would be able to expend energy for the massive mechanical fold-ing which takes place.
This work has been greatly facilitated by the competent technical assistance of Mr. RichardJahn, and the generous professional advice of Dr. Stephen Wolfe; their help is gratefully ac-knowledged.
* Supported in part by NSF grant GB-1924.'Kleinschmidt, A. K., D. Lang, D. Jacherts, and R. K. Zahn, Biochim. Biophys. Acta, 61,
857 (1962).2 Gall, J., Science, 139, 120 (1963).3 Ris, H., and B. L. Chandler, in Synthesis and Structure of Macromolecules, Cold Spring Harbor
Symposia on Quantitative Biology, vol. 28 (1963), p. 1.4 Wolfe, S., Exptl. Cell Res., in press; J. Ultrastruct. Res., in press.5 Anderson, T., Trans. N. Y. Acad. Sci., Ser. II, 13, 130 (1951).6 Littau, V. C., V. G. Allfrey, J. H. Frenster, and A. E. Mirsky, these PROCEEDINGS, 52, 93
(1964).7 DuPraw, E. J., Analysis of Embryonic Development in the Honeybee I, 16-mm sound film,
available from The Library of Congress, Washington, D. C. (1963).8 Gall, J., in Cytodifferentiation and Macromolecular Synthesis, ed. Michael Locke (New York:
Academic Press, 1963), p. 122.9 Callan, H. G., Intern. Rev. Cytol., 15, 1 (1963).10 Swift, H., in The Molecular Control of Cellular Activity, ed. John Allen (New York: McGraw-
Hill, 1962), p. 84.11 Taylor, J. H., Genetics, 43, 515 (1958).12 Cairns, J., J. Mol. Biol., 6, 208 (1963); in Synthesis and Structure of Macromolecules, Cold
Spring Harbor Symposia on Quantitative Biology, vol. 28 (1963), p. 43.13 Watson, J. D., and F. H. C. Crick, in Viruses, Cold Spring Harbor Symposia on Quantitative
Biology, vol. 18 (1953), p. 123.14 Mazia, D., J. Cell. Comp. Physiol., 62 (supp.), 123 (1963).16 Billen, D., Biochem. Biophys. Res. Commun., 7, 179 (1962); Keir, H. M., R. M. S. Smellie,
and G. Siebert, Nature, 196, 752 (1962); Littlefield, J. W., A. P. McGovern, and K. B. Margeson,these PROCEEDINGS, 49, 102 (1963).
16 Busch, H., W. J. Steele. L. S. Hnilica, C. W. Taylor, and H. Mavioglu, J. Cell. Comp. Physiol..62 (supp.), 95 (1963).