nanoscale optoregulation of neural stem cell
TRANSCRIPT

FULL PAPERwww.afm-journal.de
© 2017 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim1701420 (1 of 10)
Nanoscale Optoregulation of Neural Stem Cell Differentiation by Intracellular Alteration of Redox Balance
Sara Hassanpour-Tamrin, Hossein Taheri, Mohammad Mahdi Hasani-Sadrabadi, S. Hamed Shams Mousavi, Erfan Dashtimoghadam, Mahdi Tondar, Ali Adibi, Alireza Moshaverinia,* Amir Sanati Nezhad,* and Karl I. Jacob*
Regulation of stem cell (SC) fate, a decision between self-renewal and differ-entiation, is of immense importance in regenerative medicine and has been proven to be a powerful stimulus regulating many cell functions influencing the SC fate. This study uses triphenylphosphonium-functionalized gold nanoparti-cles (TPP-AuNPs) to explore the interplay of intracellular electromagnetic (EM) exposure and the SC fate. Localized EM waves are generated inside neural stem cells (NSCs) to stimulate TPP-AuNPs (AuNPs), targeting the mitochon-dria through inducing reactive oxygen species and differentiating these cells into neurons. Following laser irradiation of TPP-AuNPs-transfected NSCs, their differentiation to neurons is monitored by tracing the relevant markers both at the genetic and protein levels. The electrophysiology technique is further used to examine the functionality of neurons. The results confirm that TPP-AuNPs subjected to electromotive forces have the potential to regulate cellular fate, although further investigations are still required to shed light on the mecha-nisms underlying the interaction of EM-stimulated TPP-AuNPs on cellular fate to design highly adjustable cell differentiation and reprogramming methods.
DOI: 10.1002/adfm.201701420
1. Introduction
Stem cells (SCs) are unspecialized cells known for their self-renewal capacity and pluripotency. These cells can proliferate without being differentiated into other cellular types, and are
Stem Cells
S. Hassanpour-Tamrin, Prof. A. Sanati NezhadDepartment of Mechanical and Manufacturing EngineeringCentre for Bioengineering Research and Education (CBRE)University of CalgaryCalgary, Alberta T2N 1N4, CanadaE-mail: [email protected]. Hassanpour-TamrinDepartment of Biomedical EngineeringAmirkabir University of TechnologyTehran 15916-34311, IranDr. H. Taheri, S. H. Shams Mousavi, Prof. A. AdibiSchool of Electrical and Computer EngineeringGeorgia Institute of TechnologyAtlanta, GA 30332, USADr. M. M. Hasani-Sadrabadi, Prof. K. I. JacobParker H. Petit Institute for Bioengineering and BioscienceG.W. Woodruff School of Mechanical EngineeringSchool of Materials Science and EngineeringGeorgia Institute of TechnologyAtlanta, GA 30332, USAE-mail: [email protected]
Dr. M. M. Hasani-Sadrabadi, Prof. A. MoshaveriniaWeintraub Center for Reconstructive BiotechnologyDivision of Advanced ProsthodonticsSchool of DentistryUniversity of CaliforniaLos Angeles, CA 900951668, USAE-mail: [email protected]. M. M. Hasani-SadrabadiCalifornia NanoSystems InstituteUniversity of California570 Westwood Plaza, Los Angeles, CA 90095-7227, USADr. E. DashtimoghadamMarquette University School of DentistryMilwaukee, WI, USAM. TondarDepartment of Biomedical SciencesBoard of Governors Regenerative Medicine InstituteCedars Sinai Medical CenterLos Angeles, CA, USA
The ORCID identification number(s) for the author(s) of this article can be found under https://doi.org/10.1002/adfm.201701420.
capable of differentiating into a variety of cell lineages.[1] Therefore, SCs are key players in numerous vital processes such as morphogenesis and organ repair.[2] Neural stem cells (NSCs), from three spe-cific niches (the subventricular zone, the subgranular zone of the dentate gyrus, and the external germinal layer of the cer-ebellum),[3] are multipotent adult SCs with both self-renewing and differentiating properties in development of fetal and adult central nervous system.[3] NSCs can keep their stem cell identity and renew themselves, or can differentiate into other types of cells such as neurons, astrocytes, and oligodendrocytes.[4] Therefore, they have tremendous promise for a variety of cell-based treatments for neural dis-eases and injuries.[5] However, therapeutic applications of NSCs have been met with only modest success due to the challenge of defining a transplantable NSC popula-
tion and directing NSC differentiation. Appropriate numbers of NSCs in the nervous system depend on a fine balance between self-renewal and differentiation, something that highlights the importance of controlling the NSC fate as a choice between self-renewal and differentiation.
Adv. Funct. Mater. 2017, 1701420

www.afm-journal.dewww.advancedsciencenews.com
1701420 (2 of 10) © 2017 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
The balance between self-renewal and differentiation is an important factor for determining NSC fate. Cells use dif-ferent ways of communication to harmonize their activities. In fact, many cellular functions in SCs rely on both intrinsic and extrinsic signals. Therefore, SC fate can be controlled by inter-actions between extrinsic signals, derived from the microenvi-ronment in which cells are located (their niche),[6] and intrinsic signals (like transcriptional factors and cell cycle regulators).[7] Electromagnetic fields (EMFs), in particular, influence intracel-lular communications via the transport of energy across cells and their microenvironment. EMFs, characterized based on their wavelength and energy,[8,9] are known as low-level radia-tions (nonionizing radiations) among the electromagnetic (EM) waves, and do not break atomic bonds.[10] EM stimulations as extrinsic signals have a potential to direct the fate of SCs.[10] According to several reports, the EM nature of biological sys-tems makes the transmission of EM signals possible for the control and regulation of cellular processes.[10] In fact, living cells consisting of biological polar units such as ions are capable of producing EMFs in the form of ultraweak photons.[9–11] Pre-vious studies have demonstrated the influence of EMFs on some intracellular mechanisms and gene expressions.[12–14]
Moreover, many studies have reported the influence of externally applied EMFs on cells.[10] The observed effects (like differentiation) are strongly correlated to the cellular or intra-cellular physiological states and parameters of applied fields such as frequency, intensity, and exposure time.[10,15] Based on several studies on NSCs, EMFs can promote NSC differentia-tion into functional cells, for example the differentiating effect of extremely low-frequency electromagnetic fields (ELF-EMFs) on NSCs in rats[16] and embryonic NSCs in mice.[17] Although these studies reported that EMFs can induce differentiation in NSCs, the mechanisms underlying these effects are not well understood. One possible mechanism for the activation of cell differentiation pathways through EM stimulations can be linked to the generation of free radical reactive oxygen species (ROS) and activation of pathways related to ROS.[13,18] ROS are highly reactive molecules and one site for their generation is mitochondria where they are a byproduct of oxidative phos-phorylation.[19,20] Mitochondrial activities, as a cellular power source,[20,21] play a regulatory role in cell functions.[21,22] There-fore, it is reasonable to explore the effects of mitochondria on differentiation of SCs. The aim of this study is to investigate how localized generation of EMFs around mitochondria can regulate the fate of NSC, and explore the feasibility of NSC dif-ferentiation through localized EMF.
2. Results and Discussion
A localized EMF is generated around the gold nanoparticles (AuNPs) in the target mitochondria site to examine how EMFs influence the differentiation behavior of NSCs (Figure 1a).[23,24] Biocompatibility, chemical stability, and adjustable optical properties of AuNPs make them versatile for many biomedical applications.[24–28] Besides, the possibility of surface modifica-tion on AuNPs allow to target specific regions within the cells, which makes the use of these particles a promising candidate for cellular investigations. AuNPs have also been successfully
delivered into cells without affecting cell viability,[29] and internalized in mitochondria with no reported apoptosis or necrosis.[30,31]
The interaction of nanoparticles (NPs) with cells starts with the contact with plasma membrane followed by subsequent entry into cells via endocytosis or other means such as pores and channels on the plasma membrane.[28,31–33] This complex interaction is highly dependent on the size, shape, and sur-face property of NPs. After internalization, it is necessary to direct NPs to their specific target within cells. Conjugation of cationic groups to the surface of AuNPs results in water-dispersed NPs.[34] Triphenylphosphonium (TPP) is a lipophilic phosphonium cation with a high affinity to mitochondria. TPP cations have been used for studying mitochondrial bioener-getics and free radicals.[27,31,35] In this study, the AuNPs were functionalized with the phosphonium groups (TPP cations) for mitochondrial localization. The attachment of AuNPs to the TPP cations produced a mechanism targeting the mitochondria followed by their accumulation within the cells. This system enables studying the intracellular effects of localized EMFs by laser irradiation.
AuNPs were synthesized as detailed in the Supporting Infor-mation, functionalized with TPP, and analyzed with UV–vis spectrophotometry, high resolution transmission-electron microscopy (HR-TEM), and dynamic light scattering (DLS). The HR-TEM analysis of the AuNP-TPP (Figure 1b) shows spherical and monodispersed functionalized NPs, corroborated with the size distribution analysis of particles using DLS, presenting an average particle size of ≈10 nm (Figure 1c). Particles with the size of 10 nm are selected in this study given their acceptable level of cellular uptake[31,32] and the ease of tracing them inside the cells.[36] The UV–vis absorption measurement of AuNPs (Figure 1d) illustrates the maximum absorption wavelength at 529 nm corresponding to the localized surface plasmon reso-nance of AuNPs,[37] shifted to 520 nm after TPP conjugation. The refractive index around the particles changes after TPP binding, which consequently causes the shift in the wavelength of absorption peak, confirming the strong binding of TPP to the surface of AuNPs (Figure 1d). The successful conjugation of TPP on the surface of AuNP-Amine is also confirmed using 1H NMR (Figure S1, Supporting Information). It was demon-strated that the surface density of TPP on the surface of AuNPs can be controlled by changing the concentration of TPP in the reactant mixture.
The absorption cross-section, σabs, of the light-stimulated AuNPs is the equivalent surface area required to absorb the same amount of power in the absence of NPs. NPs typi-cally have absorption cross-sections larger than their dimen-sions because of the localized surface plasmon polariton resonances.[38] Increasing the size of nanospheres from 10 to 14 nm does not change the resonance wavelength considerably (lres ≈ 527 nm) but increases σabs significantly (Figure 1e). As an example, a NP with a radius of 5 nm has a σabs of ≈71.84 nm2 at 540 nm. Assuming the exposure of cells to 1 mW of laser power over a spot size diameter of 2 µm, the power of ≈22.87 pW is absorbed per NP (Figure 1f–h).
The intracellular localization and distribution of NPs were investigated by confocal laser scanning microscopy (CLSM). Figure 2a–d illustrates the fluorescent images of incubated
Adv. Funct. Mater. 2017, 1701420

www.afm-journal.dewww.advancedsciencenews.com
1701420 (3 of 10) © 2017 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
NSCs with nonfunctionalized AuNPs (Figure 2a,c) and func-tionalized AuNPs (Figure 2b,d) for 4 h at 37 °C. The green fluorescence image is related to the fluorescein isothiocyanate isomer labeled NPs (panel (i)), while the red fluorescence image indicates MitoTracker (panel (ii)). Panel (iii) with yellow spots shows the merged images of green and red, demon-strating the mitochondrial localization of NPs. The degree of colocalization is also demonstrated in Figure 2b,d for cells incubated with AuNPs and TPP-AuNPs, respectively. The con-focal images reveals the high level of NPs internalization in NSCs and the successful colocalization of AuNP-TPP in mito-chondria, concluded from yellow fluorescence signals in panel (iii). The higher association between the red and green fluo-rescence channels, represented in intensity-based 3D recon-struction of fluorescent images (Figure 2d), also confirms a successful colocalization with TPP-AuNPs in comparison with unmodified AuNPs.
Additional spectrometric assays were performed to con-firm and quantify the presence of TPP-AuNPs inside the cells. Cellular internalization of AuNPs and TPP-AuNPs were measured by the fluorescence activated cell sorting method (Figure 2e(i)) and inductively coupled plasma mass spectrom-etry (Figure 2e(ii)). Although surface functionalization with TPP results in a slight decrease in cellular uptake compared to nonfunctionalized NPs, a high number of internalized NPs are observed in both functionalized and nonfunctionalized NPs. However, mitochondrial uptake, as quantified by mitochon-dria isolation, confirms a higher targeting efficiency of 75% for AuNP-TPP compared to 30% for AuNPs after 4 h coculturing.
The examination of TPP-AuNPs cytotoxicity as one crit-ical step after internalization was performed by the MTT (3-(4,5-dimethylthiazol-2-Yl)-2,5-diphenyltetrazolium bromide) assay. The viability of cells in the presence of AuNPs and TPP-AuNPs with and without irradiation (see the Supporting
Adv. Funct. Mater. 2017, 1701420
Figure 1. Physical characteristics of triphenylphosphonium-functionalized gold nanoparticles (AuNP-TPP); a) The schematic representation of mitochondrial-targeted gold nanoparticles (AuNPs) to localize electromagnetic field (EMF) for neural differentiation of neural stem cells (NSCs). b) Transmission electron micrographs of functionalized AuNP-TPP (scale bar is 50 nm). c) The hydrodynamic diameter of AuNP and AuNP-TPP as measured via dynamic light scattering method. d) UV–vis absorption spectra of AuNP and AuNP-TPP. e) Distribution of electric field (E-field) and magnetic field (H-field) around the nanosphere with the diameter of 10 nm. The excitation field is a plane-wave propagating in direction of kexc. f) The absorption cross-section of the nanosphere for diameters of 10, 11, 12, 13, and 14 nm. g) E-field enhancement factor (|E|/|Eexc|) in the xy-plane crossing the center of the nanosphere. h) E-field enhancement factor (|E|/|Eexc|) versus the radial distance from the center of AuNP of 10 nm diameter.

www.afm-journal.dewww.advancedsciencenews.com
1701420 (4 of 10) © 2017 WILEY-VCH Verlag GmbH & Co. KGaA, WeinheimAdv. Funct. Mater. 2017, 1701420
Figure 2. Mitochondrial targeting efficiencies. Confocal laser scanning microscopy images on mitochondrial colocalization of a,c) AuNP and b,d) AuNP-TPP. Panels (i)–(iii) indicate the mitochondria stained with MitoTracker (red), AuNPs labeled by fluorescein isothiocyanate (FITC) (green), and the overlay image of them (yellow) (scale bar is 20 µm). e) Cellular uptake of fluorescently labeled AuNP and AuNP-TPP using fluorescence-activated cell sorting (FACS) (left panel). Cellular uptake evaluated using inductively coupled plasma-mass spectrometry (ICP-MS) spectroscopy. Mitochondrial uptake portions are indicated within each bar. *p < 0.05 and **p < 0.01 between cellular/ mitochondrial uptake after 4 and 1 h for AuNP and AuNP-TPP. ##p < 0.01 between mitochondrial uptake for AuNP-TPP and AuNP. f) Cellular viability of NSC after 72 h incubation with AuNP and AuNP-TPP w/o irradiation. *p < 0.05 between each treatment group and cells treated with AuNP without irradiation (control (−)). g) In vial generation of reactive oxygen species (ROS) after laser irradiation. h) Monitoring of intracellular ROS generation during 120 min of treatment with laser in the presence and absence of AuNPs. The empty triangles indicate the irradiation points. *p < 0.05 and **p < 0.01 between the sample with and without irradiation (negative controls).

www.afm-journal.dewww.advancedsciencenews.com
1701420 (5 of 10) © 2017 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
Information for details) was measured, and statistically com-pared among various concentrations of NPs from 0.25 to 40 µg mL−1 (Figure 2f). It seems that the viability of all treated cells (AuNPs-TPP and AuNPs-Amine with/-out irradiation) are quite high (>95%) after the incubation with low concentra-tions of NPs (up to 1 µg mL−1). The results also indicate that the surface functionalization of NPs has a trivial impact on the toxicity of NPs. The increase in the concentration of AuNPs from 0.25 to 40 µg mL−1 causes a significant cellular toxicity, especially in the presence of irradiation. The irradiated cells in both groups, incubated with AuNPs and TPP-AuNPs, show a lower cell viability compared to nonirradiated cells. A plausible explanation for the reduction in the cell viability is the involve-ment of a higher production rate of ROS. The excessive concen-tration of ROS can lead to oxidative stress and cell death.[39,40] The results indicate that the fraction of dead cells increases for cells loaded with high concentrations of NPs (above 5 µg mL−1) while the cell viability is above 90% at low concentrations of NPs (up to 5 µg mL−1). The dose-dependent results confirm that the cellular toxicity of NPs is trivial even after IR irradia-tions at concentrations below 2 µg mL−1. It should be noted that the optimal cell viability (>95%) was achieved by setting the concentration of NPs to 1 µg mL−1, even in the presence of irra-diation selected for further experiments.
Despite the destructive effects of ROS, these active species can act as the second messenger in a wide variety of biological processes.[39–41] A growing body of evidence indicates that ROS have mediatory roles in cell growth, proliferation, and differen-tiation.[39,42] The implication of ROS as the second-messengers in maintenance and growth of cells has been demonstrated for NSCs.[43] Interestingly, it has been reported that ROS can influ-ence NSC differentiation and regulate neurogenesis.[41,44] EMFs affect ROS generation as a factor influencing the SC fate. The stimulation of NADH (nicotinamide adenine dinucleotide—hydrogen (reduced))-oxidase pathway and ROS production in mouse bone marrow-derived cells after applying extremely low frequency magnetic field (ELF-MF) (50 Hz, 1 mT); enhanced production of ROS in murine L929 fibrosarcoma cells exposed to the radiofrequency EMFs (at 900 MHz); increased ROS level in the rat lymphocytes under treatment with a combination of radiofrequency radiation and iron ions; and elevated ROS level and enhanced neural differentiation of human MSCs exposed to ELF-EMF (50 Hz, 1 mT), are some evidence of the EMF impact on ROS generation. It should be noted that EMFs can also influence the activation of antioxidant enzymes and pre-vent ROS generation. In rat’s liver mitochondria, for example, the generation of free radicals was prevented upon the acti-vation of antioxidant enzymes through EM pulse exposure (60 kV m−1 strength). An increase in the activity of SOD (super-oxide dismutase) enzymes after the exposure of mice and rat brains, respectively, to the ELF-magnetic field of 60 and 50 Hz are other indicators of the effect of EMF on antioxidant activities.[10] In a study on embryonic NSC, the messenger ribo-nucleic acid (mRNA) level of genes related to neuronal differ-entiation was affected after the treatment of cells with 50 Hz ELF-EMF (2 mT for 3 d).[45]
On the other hand, mitochondrial activity and the amount of ROS generation are closely related.[10] The rate of mitochon-drial activity is higher in undifferentiated NSCs in contrast to
differentiated neurons and glial cells while the rate of ROS generation is less.[46] It is, therefore, suggested that the intra-cellular redox balance may be influenced after laser irradiation, and ROS may serve as a regulating factor in differentiation of NSCs.
To examine the effect of laser irradiation on intracellular redox state, the cellular ROS value was measured after laser irradiation. To detect ROS formation, the fluorescence intensity of dihydrorhodamine 123 (DHR123), as a ROS tracking agent, was measured using fluorescence spectrophotometry. Figure 2g shows the increase in the fluorescence intensity with irradiation time, indicating that the ROS concentration can be adjusted by tuning the irradiation time. Also, Figure 2h shows the increase in the intracellular ROS concentrations within the cells after laser irradiation. The detected decline in the plateau level of the ROS intensity occurring after each peak in the graph can be attributed to the short lifetime of ROS and the activity of var-ious redox systems inside the cells (like antioxidants[47] having a suppressive effect on the ROS), which balances the cellular oxidative stress.[26]
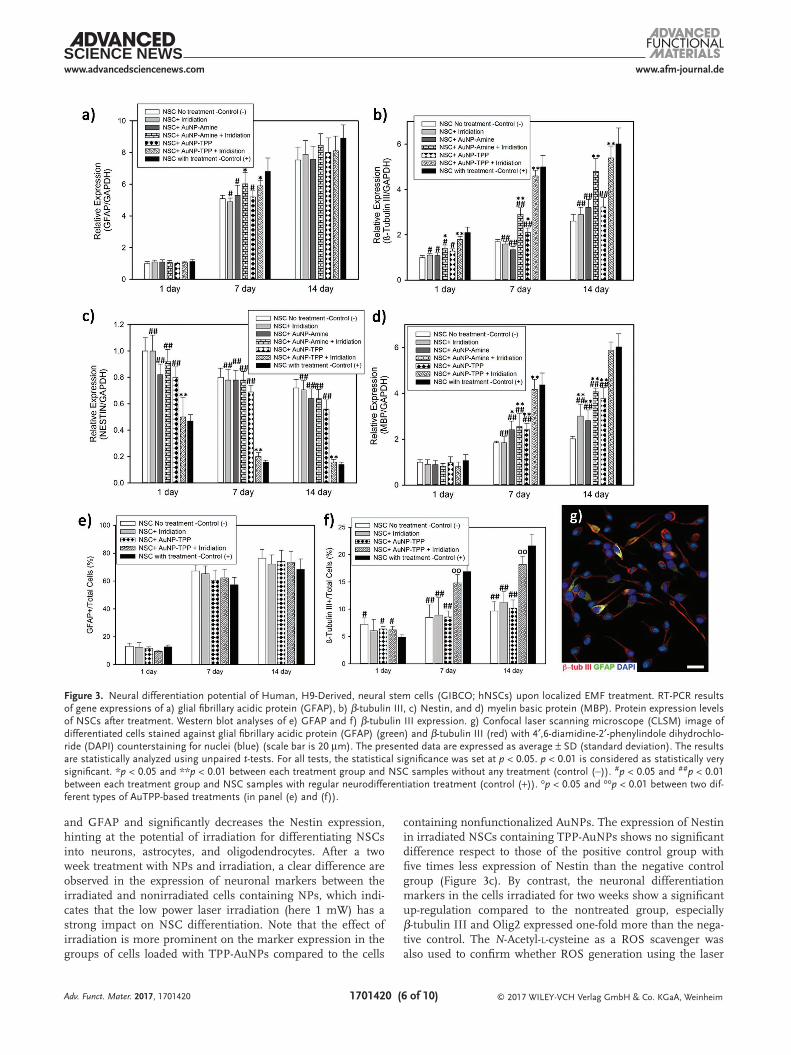
The reverse transcription polymerase chain reaction (RT-PCR) technique was further employed to investigate neural differentiation of NSCs after treatment with NPs followed by laser irradiation. The RT-PCR data shows that NPs and laser irradiation induced differentiation of NSCs via altering the expression levels of Nestin, β-tubulin III, myelin basic pro-tein (MBP), Olig2, and Glial fibrillary acidic protein (GFAP), as representative marker genes for neural stem/precursor cells, neuron, oligodendrocyte, and astrocytes, respectively. It is well-known that upon differentiation of NSCs, the expres-sion of Nestin, a marker for undifferentiated NSCs, decreases progressively and the level of neuronal differentiation markers increases,[48] implying the production of more mature cells, in agreement with RT-PCR data (Figure 3).
At three different stages (day 1, 7, and 14), cells from dif-ferent groups (treated with or without laser irradiation and TPP-AuNPs/AuNPs) were processed for gene expression anal-yses. The expression of each gene was plotted in Figure 3a–d and Figure S4 (Supporting Information) after normalizing to the expression of glyceraldehyde-3-phosphate dehydrogenase mRNA. For negative control group, the cells were cultured in normal culture medium without any treatment with NPs and irradiation. For positive control group, the cells were cultured in differentiation medium containing neurobasal medium with B-27 serum-Free supplement and GlutaMAX-I to induce neural differentiation. Under the negative control condition, NSCs express a high level of Nestin while neuronal differen-tiation markers (β-tubulin III, MBP, Olig2, and GFAP) stay at their lowest levels. As determined by RT-PCR, the expression of β-tubulin III, MBP, Olig2, and GFAP is up-regulated within two weeks of differentiation, and the expression of Nestin decreases in the differentiation groups compared to negative control.
The effect of laser irradiation on up-regulation of the differ-entiation markers like β-tubulin III, MBP, and GFAP and down-regulation of the Nestin level are shown in Figure 3a–d. More importantly, the results indicate that changes in the expression of markers are most conspicuous when NSCs are treated with both NPs and laser irradiation. The exposure to laser irradia-tion increases the gene expression of β-tubulin III, MBP, Olig2,
Adv. Funct. Mater. 2017, 1701420
www.afm-journal.dewww.advancedsciencenews.com
1701420 (6 of 10) © 2017 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
and GFAP and significantly decreases the Nestin expression, hinting at the potential of irradiation for differentiating NSCs into neurons, astrocytes, and oligodendrocytes. After a two week treatment with NPs and irradiation, a clear difference are observed in the expression of neuronal markers between the irradiated and nonirradiated cells containing NPs, which indi-cates that the low power laser irradiation (here 1 mW) has a strong impact on NSC differentiation. Note that the effect of irradiation is more prominent on the marker expression in the groups of cells loaded with TPP-AuNPs compared to the cells
containing nonfunctionalized AuNPs. The expression of Nestin in irradiated NSCs containing TPP-AuNPs shows no significant difference respect to those of the positive control group with five times less expression of Nestin than the negative control group (Figure 3c). By contrast, the neuronal differentiation markers in the cells irradiated for two weeks show a significant up-regulation compared to the nontreated group, especially β-tubulin III and Olig2 expressed one-fold more than the nega-tive control. The N-Acetyl-l-cysteine as a ROS scavenger was also used to confirm whether ROS generation using the laser
Adv. Funct. Mater. 2017, 1701420
Figure 3. Neural differentiation potential of Human, H9-Derived, neural stem cells (GIBCO; hNSCs) upon localized EMF treatment. RT-PCR results of gene expressions of a) glial fibrillary acidic protein (GFAP), b) β-tubulin III, c) Nestin, and d) myelin basic protein (MBP). Protein expression levels of NSCs after treatment. Western blot analyses of e) GFAP and f) β-tubulin III expression. g) Confocal laser scanning microscope (CLSM) image of differentiated cells stained against glial fibrillary acidic protein (GFAP) (green) and β-tubulin III (red) with 4′,6-diamidine-2′-phenylindole dihydrochlo-ride (DAPI) counterstaining for nuclei (blue) (scale bar is 20 µm). The presented data are expressed as average ± SD (standard deviation). The results are statistically analyzed using unpaired t-tests. For all tests, the statistical significance was set at p < 0.05. p < 0.01 is considered as statistically very significant. *p < 0.05 and **p < 0.01 between each treatment group and NSC samples without any treatment (control (−)). #p < 0.05 and ##p < 0.01 between each treatment group and NSC samples with regular neurodifferentiation treatment (control (+)). op < 0.05 and oop < 0.01 between two dif-ferent types of AuTPP-based treatments (in panel (e) and (f)).

www.afm-journal.dewww.advancedsciencenews.com
1701420 (7 of 10) © 2017 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
irradiation is the cause of the proposed biological effects. As shown in Figure S2 in the Supporting Information, the ROS scavenger can eliminate ROS signals in vial. Evaluating the RT-PCR results of β-tubulin III and Nestin expression com-pared to housekeeping gene (GFAP), it is found that the inhi-bition of ROS generation can prevent neural differentiation as expected (Figure S3, Supporting Information).
Western blotting assay and CLSM were used to verify the laser irradiation effect on NSC differentiation in the presence of TPP-AuNPs. In agreement with the RT-PCR results, the protein expression assay demonstrates the neural differentia-tion potential of NSCs containing NPs after the irradiation in which the cells are labeled for β-tubulin III and GFAP. The protein expression plot for β-tubulin III shows the increasing number of β-tubulin III positive cells within 14 d after treat-ment with TPP-AuNPs in both groups of irradiated and nonir-radiated, like what detected for the control group with normal neurodifferentiation treatment. As depicted in Figure 3f, the number of β-tubulin III positive cells from the irradiated group is approximately twice the number of positive cells in the nonir-radiated group. After 14 d, the number of β-tubulin III positive cells reaches about 20% of the total cells in the irradiated group while that is close to the control group with normal neural dif-ferentiation treatment (See Supporting Information for more details). A similar trend is observed within 14 d for the GFAP expression. However, there is no significant difference between the irradiated, nonirradiated, and control groups after 14 d. Overall, the protein expression plots show increased percent-ages of GFAP positive and β-tubulin III positive cells after irra-diation of NSCs containing NPs, confirming the differentiation of NSCs into neurons and astrocytes. Notably, NSCs containing NPs show more tendencies toward glial differentiation with or without irradiation. Also the incidence of neuronal differentia-tion is also supported by the CLSM image of NSCs containing TPP-AuNPs after laser irradiation at 1 mW (Figure 3g). The cells are stained with antibodies against GFAP (green) and β-tubulin III (red) while the nuclei are counterstained with 4′,6-diamidine-2′-phenylindole dihydrochloride (blue). Both GFAP and β-tubulin III are detected in CLSM images, con-firming the presence of GFAP+ and β-tubulin III+ cells, and the differentiation of cells at day 14.
To assess the functionality of differentiated cells, their neural activity was examined by measuring their electrophysiological properties, including the membrane currents and Ca2+ concen-tration. First, the presence of the voltage-gated channels was studied after stimulation of the differentiated cells with the depolarizing current injection using patch clamp whole-cell recording as described before.[49] In voltage-dependent currents, recorded in the 7 and 21 d (Figure 4a,b), the holding potential was set at −70 mV, and 20 ms depolarizing pulses were applied from −60 to +60 mV in 20 mV steps to evoke channel opening. The current–voltage relationship of sodium (Na+) currents in cells undergoing 7 and 21 d terminal differentiation show remarkable inward Na+ current with a mean peak of approxi-mately −870 ± 50 pA at −20 mV for the second stage of matura-tion (day 21) (Figure 4a). The I–V relationship of the potassium (K+) current at two stages of maturation demonstrates an out-ward current (Figure 4b). These voltage-dependent Na+ and K+ currents in the stimulated cells indicate that voltage-sensitive
ion channels exist in the differentiated cells, confirming the presence of physiologically active neuronal cells. However, NSCs treated with AuNP-TPP without irradiation fail to show the same level of voltage-dependent Na+ and K+ currents (Figure S5, Supporting Information).
Next, calcium imaging experiments were performed to monitor the change of Ca2+ level as a physiological marker of differentiation inside the differentiated cells. To verify intracel-lular Ca2+ concentration, the cells were electrically stimulated as described before.[50] The calcium imaging represents a tran-sient calcium increase in the stimulated cells (Figure 4c), which is quantified in Figure 4d. Based on the plot of the relative change (ΔF/F) of the cell fluorescence intensity versus the stim-ulation period, over 60% increase in the fluorescence intensity is observed after the stimulation, corroborating an increased intracellular Ca2+ concentration. This result implies the appear-ance of voltage-sensitive Ca2+ channels in the differentiated cells, which is a basic neuronal membrane characteristic. Thus, the voltage pulse stimuli open the Ca2+ channels and increase the Ca2+ concentration in the differentiated cells.[51] Overall, this study show that laser irradiation significantly increases the expression of neuronal differentiation genes in the presence of NPs, emphasizing the potential of EM exposure to enhance neuronal differentiation of NSCs.
Although researchers are struggling to bring up the mecha-nisms underlying EM exposure for SC differentiation, some reports have suggested the probable relationship between ROS production and underlying mechanisms of EM exposure for differentiation. Additionally, the relationship between ROS generation and mitochondrial activity of differentiated and undifferentiated NSCs suggests the control of SC fate through controlling ROS generation. Our results also show that ROS production increases in NSCs after the exposure to laser irra-diation. It seems reasonable to suggest that ROS production is involved in the mechanisms underlying the interaction of EMF and NSCs.
It should be highlighted that EMF may cause differen-tiation in NSCs through the activation of ionic channels and alteration in ion concentration,[52] changes in gene expres-sion (transcriptional factors and pluripotency markers),[53] and influencing intracellular molecular mechanisms such as MAPK (mitogen-activated protein kinase) pathway.[14] How-ever, the underlying mechanisms of EM exposure responsible for neural differentiation of NSCs is still unclear and need to be further clarified. Undoubtedly, further experiments are needed to better understand the redox-dependent mechanisms for NSC differentiation in response to laser irradiation. In our experiments, although EM exposure induced differentiation in NSCs in the presence of NPs, the differentiation was non-selective. One major challenge would be the differentiation of NSCs into specific neuronal and glial cell lines. There is a need to investigate whether the combination of NPs and laser irra-diation treatments could guide the NSCs differentiation toward specific cell types, and whether ROS-related mechanisms work differently in astrocyte, neuron and oligodendrocyte differ-entiation to present a selective protocol for the differentiation of NSCs. However, our approach has been utilized as a tech-nique (nanoparticles and stimulatory EM generation) to pro-mote the differentiation of NSCs in the absence of soluble cues
Adv. Funct. Mater. 2017, 1701420

www.afm-journal.dewww.advancedsciencenews.com
1701420 (8 of 10) © 2017 WILEY-VCH Verlag GmbH & Co. KGaA, WeinheimAdv. Funct. Mater. 2017, 1701420
(like exogenous proteins). This methodology can also poten-tially be helpful to investigate how localized generation of EMFs in a targeted site (mitochondria or cell membrane) can influ-ence cell responses to explore the feasibility of NSC differentia-tion through localized EMF. Furthermore, our proposed system can be used as a research tool to study the intracellular effects of localized EMFs within the cells and identify genes and sign-aling pathways effective in regulation of NSC fate. This system can also be translated into better understanding how NSCs bal-ance self-renewal versus differentiation to address an optimal treatment for neurodegenerative diseases like Alzheimer's and Parkinson's. Although we focused on NSCs differentiation, this technique could be extended to study and control the fate of other stem cells, from a variety of types, stages of differentia-tion and sources.
Supporting InformationSupporting Information is available from the Wiley Online Library or from the author.
AcknowledgementsS.H.-T, H.T., and M.M.H.-S contributed equally to this work. The manuscript was written through contributions of all authors. All authors have given approval to the final version of the manuscript. The authors
highly appreciate critical reading of the manuscript by Mohsen Janmaleki at the University of Calgary. This research was supported by the Natural Sciences and Engineering Research Council of Canada and ALS Canada/Brain Canada funding. This research was performed in the framework of Biologically Inspired Developing Advanced Research (BiDAR) group.
Conflict of InterestThe authors declare no conflict of interest.
Keywordselectromagnetic fields, nanoparticles, neural stem cells, optoregulation, stem cell fate
Received: March 17, 2017Revised: June 27, 2017
Published online:
[1] S. Yamanaka, J. Li, G. Kania, S. Elliott, R. P. Wersto, J. Van Eyk, A. M. Wobus, K. R. Boheler, Cell Tissue Res. 2008, 331, 5.
[2] a) S. N. B. Bernhard, O. Palsson, Tissue Engineering, Pearson Pren-tice Hall, University of California, San Diego, 2004; b) B. Mead, A. Logan, M. Berry, W. Leadbeater, B. A. Scheven, Stem Cells 2017, 35, 61; c) I. Lombaert, M. M. Movahednia, C. Adine, J. N. Ferreira, Stem Cells 2017, 35, 97.
Figure 4. Functional evaluation of neurons; current–voltage relationship for a) Na+ and b) K+ currents as a function of test potential for the differenti-ated cells. c) Ca2+ imaging, d) the change of relative fluorescence intensity for the differentiated cells (scale bar is 150 µm). *p < 0.05 and **p < 0.01 between treated groups at day 21 and day 7.

www.afm-journal.dewww.advancedsciencenews.com
1701420 (9 of 10) © 2017 WILEY-VCH Verlag GmbH & Co. KGaA, WeinheimAdv. Funct. Mater. 2017, 1701420
[3] K. Reekmans, J. Praet, J. Daans, V. Reumers, P. Pauwels, A. Van der Linden, Z. N. Berneman, P. Ponsaerts, Stem Cell Rev. Rep. 2012, 8, 262.
[4] A. Chojnacki, S. Weiss, Nat. Protoc. 2008, 3, 935.[5] a) G. Martino, S. Pluchino, Nat. Rev. Neurosci. 2006, 7, 395;
b) B. J. Cummings, N. Uchida, S. J. Tamaki, D. L. Salazar, M. Hooshmand, R. Summers, F. H. Gage, A. J. Anderson, Proc. Natl. Acad. Sci. USA 2005, 102, 14069.
[6] a) F. Gattazzo, A. Urciuolo, P. Bonaldo, Biochim. Biophys. Acta, Gen. Subj. 2014, 1840, 2506; b) F. M. Watt, W. T. Huck, Nat. Rev. Mol. Cell Biol. 2013, 14, 467; c) C. Wang, J. Chen, P. Wen, P. Sun, R. Xi, in Regenerative Medicine-from Protocol to Patient, Springer, Cham, Switzerland, 2016, pp. 57–85.
[7] a) J. A. Knoblich, Cell 2008, 132, 583; b) H. Toledano, D. L. Jones, in StemBook, Cambridge, MA, 2009.
[8] A. Markkanen, Kuopio Univ. Environ. Sci. 2009, 253, 1.[9] C. Furse, D. A. Christensen, C. H. Durney, Basic Introduction to Bio-
electromagnetics, CRC Press, Boca Raton, FL, 2009.[10] S. H. Tamrin, F. S. Majedi, M. Tondar, A. Sanati-Nezhad,
M. M. Hasani-Sadrabadi, in Reviews of Physiology, Biochemistry and Pharmacology, Vol. 171 (Eds: B. Nilius, P. de Tombe, T. Gudermann, R. Jahn, R. Lill, O. H. Petersen), Springer International Publishing, Cham, Switzerland 2016, p. 63.
[11] M. Rahnama, J. A. Tuszynski, I. Bokkon, M. Cifra, P. Sardar, V. Salari, J. Integr. Neurosci. 2011, 10, 65.
[12] a) J. Zimmerman, M. Pennison, I. Brezovich, N. Yi, C. Yang, R. Ramaker, D. Absher, R. Myers, N. Kuster, F. Costa, Br. J. Cancer 2012, 106, 307; b) H. Sauer, G. Rahimi, J. Hescheler, M. Wartenberg, J. Cell. Biochem. 1999, 75, 710; c) J. Friedman, S. Kraus, Y. Hauptman, Y. Schiff, R. Seger, Biochem. J. 2007, 405, 559; d) M. Blank, R. Goodman, Pathophysiology 2009, 16, 71.
[13] S. Falone, M. R. Grossi, B. Cinque, B. D’Angelo, E. Tettamanti, A. Cimini, C. Di Ilio, F. Amicarelli, Int. J. Biochem. Cell Biol. 2007, 39, 2093.
[14] a) J. Arthur, Biochem. J. 2007, 405, e5; b) H.-J. Li, L.-M. Guo, L.-L. Yang, Y.-C. Zhou, Y.-J. Zhang, J. Guo, X.-J. Xie, G.-Z. Guo, Toxicol. Lett. 2013, 220, 35; c) D. Leszczynski, S. Joenväärä, J. Reivinen, R. Kuokka, Differentiation 2002, 70, 120.
[15] a) K. S. Kang, J. M. Hong, J. A. Kang, J.-W. Rhie, Y. H. Jeong, D.-W. Cho, Exp. Mol. Med. 2013, 45, e6; b) C. Consales, C. Merla, C. Marino, B. Benassi, Int. J. Cell Biol. 2012, 2012, 683897.
[16] Q. Gao, Ph.D. Thesis, The Hong Kong Polytechnic University, 2016.
[17] Q. Ma, C. Chen, P. Deng, G. Zhu, M. Lin, L. Zhang, S. Xu, M. He, Y. Lu, W. Duan, PLoS One 2016, 11, e0150923.
[18] a) J.-E. Park, Y.-K. Seo, H.-H. Yoon, C.-W. Kim, J.-K. Park, S. Jeon, Neurochem. Int. 2013, 62, 418; b) J. C. St. John, J. Ramalho-Santos, H. L. Gray, P. Petrosko, V. Y. Rawe, C. S. Navara, C. R. Simerly, G. P. Schatten, Cloning Stem Cells 2005, 7, 141.
[19] A. Y. Andreyev, Y. E. Kushnareva, A. Starkov, Biochemistry 2005, 70, 200.
[20] V. Adam-Vizi, C. Chinopoulos, Trends Pharmacol. Sci. 2006, 27, 639.
[21] R. Loureiro, K. A. Mesquita, P. J. Oliveira, I. Vega-Naredo, Recent Pat. Endocr., Metab. Immune Drug Discovery 2013, 7, 102.
[22] a) G. C. Parker, G. Acsadi, C. A. Brenner, Stem Cells Dev. 2009, 18, 803; b) X. Q. Ye, Q. Li, G. H. Wang, F. F. Sun, G. J. Huang, X. W. Bian, S. C. Yu, G. S. Qian, Int. J. Cancer 2011, 129, 820; c) M. B. de Moura, L. S. dos Santos, B. Van Houten, Environ. Mol. Mutagen. 2010, 51, 391.
[23] L. Bergamini, V. Voliani, V. Cappello, R. Nifosì, S. Corni, Nanoscale 2015, 7, 13345.
[24] X. Huang, M. A. El-Sayed, J. Adv. Res. 2010, 1, 13.
[25] V. Raji, J. Kumar, C. Rejiya, M. Vibin, V. N. Shenoi, A. Abraham, Exp. Cell Res. 2011, 317, 2052.
[26] M. K. Khaing Oo, Y. Yang, Y. Hu, M. Gomez, H. Du, H. Wang, ACS Nano 2012, 6, 1939.
[27] Y. Li, A. M. Gobin, G. W. Dryden, X. Kang, D. Xiao, S. P. Li, G. Zhang, R. C. Martin, Int. J. Nanomed. 2013, 8, 2153.
[28] Ö. F. Karatas, E. Sezgin, Ö. Aydın, M. Çulha, Colloids Surf., B 2009, 71, 315.
[29] D. N. Grant, M. J. Cozad, D. A. Grant, R. A. White, S. A. Grant, J. Biomed. Mater. Res., Part B 2015, 103, 1532.
[30] R. R. Sathuluri, H. Yoshikawa, E. Shimizu, M. Saito, E. Tamiya, PloS One 2011, 6, e22802.
[31] M. Kodiha, Y. M. Wang, E. Hutter, D. Maysinger, U. Stochaj, Theranostics 2015, 5, 357.
[32] A. Christofidou, Ph.D. Thesis, University of Southampton, 2013.[33] S. Soleimani, M. M. Hasani-Sadrabadi, F. S. Majedi,
E. Dashtimoghadam, M. Tondar, K. I. Jacob, Colloids Surf., B 2016, 145, 802.
[34] a) N. Lalwani, Y.-S. Chen, G. Brooke, N. A. Cross, D. W. Allen, A. Reynolds, J. Ojeda, G. J. Tizzard, S. J. Coles, N. Bricklebank, Chem. Commun. 2015, 51, 4109; b) Y. Ju-Nam, Y.-S. Chen, J. J. Ojeda, D. W. Allen, N. A. Cross, P. H. Gardiner, N. Bricklebank, RSC Adv. 2012, 2, 10345.
[35] M. Ross, G. Kelso, F. Blaikie, A. James, H. Cocheme, A. Filipovska, T. Da Ros, T. Hurd, R. Smith, M. Murphy, Biochemistry 2005, 70, 222.
[36] B. D. Chithrani, A. A. Ghazani, W. C. Chan, Nano Lett. 2006, 6, 662.[37] T. El-Brolossy, T. Abdallah, M. B. Mohamed, S. Abdallah, K. Easawi,
S. Negm, H. Talaat, Eur. Phys. J.: Spec. Top. 2008, 153, 361.[38] a) C. E. Talley, J. B. Jackson, C. Oubre, N. K. Grady, C. W. Hollars,
S. M. Lane, T. R. Huser, P. Nordlander, N. J. Halas, Nano Lett. 2005, 5, 1569; b) R. Bharadwaj, S. Mukherji, S. Mukherji, Plasmonics 2016, 11, 753; c) S. H. Shams Mousavi, A. A. Eftekhar, A. H. Atabaki, A. Adibi, ACS Photonics, 2015, 2, 1546.
[39] T. Rharass, H. Lemcke, M. Lantow, S. A. Kuznetsov, D. G. Weiss, D. Panáková, J. Biol. Chem. 2014, 289, 27937.
[40] Z. Krpetic, P. Nativo, V. Sée, I. A. Prior, M. Brust, M. Volk, Nano Lett. 2010, 10, 4549.
[41] B. C. Dickinson, C. J. Chang, Nat. Chem. Biol. 2011, 7, 504.[42] a) M. Schieber, N. S. Chandel, Curr. Biol. 2014, 24, R453;
b) W. Dröge, Physiol. Rev. 2002, 82, 47; c) H. Sauer, M. Wartenberg, J. Hescheler, Cell. Physiol. Biochem. 2001, 11, 173.
[43] a) Z. Radak, K. Suzuki, M. Higuchi, L. Balogh, I. Boldogh, E. Koltai, Free Radical Biol. Med. 2016, 98, 187; b) M. Yoneyama, K. Kawada, Y. Gotoh, T. Shiba, K. Ogita, Neurochem. Int. 2010, 56, 740; c) J. E. Le Belle, N. M. Orozco, A. A. Paucar, J. P. Saxe, J. Mottahedeh, A. D. Pyle, H. Wu, H. I. Kornblum, Cell Stem Cell 2011, 8, 59.
[44] a) M. Tsatmali, E. C. Walcott, H. Makarenkova, K. L. Crossin, Mol. Cell. Neurosci. 2006, 33, 345; b) K. A. Kennedy, S. D. Sandiford, I. S. Skerjanc, S. S. C. Li, Cell. Mol. Life Sci. 2012, 69, 215; c) H. L. Vieira, P. M. Alves, A. Vercelli, Prog. Neurobiol. 2011, 93, 444.
[45] Q. Ma, P. Deng, G. Zhu, C. Liu, L. Zhang, Z. Zhou, X. Luo, M. Li, M. Zhong, Z. Yu, PloS One 2014, 9, e90041.
[46] a) V. A. Rafalski, A. Brunet, Prog. Neurobiol. 2011, 93, 182; b) J. Zhang, I. Khvorostov, J. S. Hong, Y. Oktay, L. Vergnes, E. Nuebel, P. N. Wahjudi, K. Setoguchi, G. Wang, A. Do, H. J. Jung, J. M. McCaffery, I. J. Kurland, K. Reue, W. N. P. Lee, C. M. Koehler, M. A. Teitell, EMBO J. 2011, 30, 4860.
[47] a) L. Gallego-Villar, B. Pérez, M. Ugarte, L. R. Desviat, E. Richard, Biochem. Biophys. Res. Commun. 2014, 452, 457; b) W. Van den Ende, D. Peshev, L. De Gara, Trends Food Sci. Technol. 2011, 22, 689; c) K. Das, A. Roychoudhury, Front. Environ. Sci. 2014, 2, 53.

www.afm-journal.dewww.advancedsciencenews.com
1701420 (10 of 10) © 2017 WILEY-VCH Verlag GmbH & Co. KGaA, WeinheimAdv. Funct. Mater. 2017, 1701420
[48] H. Cho, Y. K. Seo, H. H. Yoon, S. C. Kim, S. M. Kim, K. Y. Song, J. K. Park, Biotechnol. Prog. 2012, 28, 1329.
[49] O. P. Hamill, A. Marty, E. Neher, B. Sakmann, F. Sigworth, Pfluegers Arch. 1981, 391, 85.
[50] C. Y. Pan, L. S. Kao, J. Neurochem. 1997, 69, 1085.[51] S. Y. Park, J. Park, S. H. Sim, M. G. Sung, K. S. Kim, B. H. Hong,
S. Hong, Adv. Mater. 2011, 23, H263.[52] a) C. Grassi, M. D'Ascenzo, A. Torsello, G. Martinotti,
F. Wolf, A. Cittadini, G. B. Azzena, Cell Calcium 2004, 35, 307;
b) C. B. Koch, M. Sommarin, B. Persson, L. Salford, J. Eberhardt, Bioelectromagnetics 2003, 24, 395; c) R. Piacentini, C. Ripoli, D. Mezzogori, G. B. Azzena, C. Grassi, J. Cell. Physiol. 2008, 215, 129.
[53] a) M. Maioli, S. Rinaldi, S. Santaniello, A. Castagna, G. Pigliaru, S. Gualini, C. Cavallini, V. Fontani, C. Ventura, Cell Transplant. 2013, 22, 1227; b) M. Maioli, S. Rinaldi, S. Santaniello, A. Castagna, G. Pigliaru, S. Gualini, V. Fontani, C. Ventura, Cell Transplant. 2012, 21, 1225.

本文献由“学霸图书馆-文献云下载”收集自网络,仅供学习交流使用。
学霸图书馆(www.xuebalib.com)是一个“整合众多图书馆数据库资源,
提供一站式文献检索和下载服务”的24 小时在线不限IP
图书馆。
图书馆致力于便利、促进学习与科研,提供最强文献下载服务。
图书馆导航:
图书馆首页 文献云下载 图书馆入口 外文数据库大全 疑难文献辅助工具