mutations in polycombeotic, a drosophila polycomb-group ... · mutations in polycombeotic, a...
TRANSCRIPT

Copyright 0 1990 by the Genetics Society of America
Mutations in polycombeotic, a Drosophila Polycomb-Group Gene, Cause a Wide Range of Maternal and Zygotic Phenotypes
Mark D. Phillips and Allen Shearn Department of Biology, The Johns Hopkins University, Baltimore, Maryland 21218
Manuscript received May 1 1, 1989 Accepted for publication January 18, 1990
ABSTRACT The polycomb-group genes, a set of genes characterized by mutations that cause similar phenotypes
and dosage-dependent interactions, are required for the normal expression of segment-specific homeotic loci. .Here we report that polycombeotic (formerly 1(3)1902), originally identified by a lethal mutation that causes a small-disc phenotype, is also a member of this group of essential genes. Adults homozygous for temperature-sensitive pco alleles that were exposed to the restrictive temperature during larval life display the second and third leg to first leg transformation characteristic of polycomb- group mutants. Adult females homozygous for temperature-sensitive alleles exposed to the restrictive temperature during oogenesis produce embryos that show anterior segments with structures normally unique to the eighth abdominal segment, another transformation characteristic of polycomb-group mutants. Mutations in the polycombeotic gene also cause defects not reported for mutations in other polycomb-group genes. Females homozygous for the most extreme temperature-sensitive allele are sterile, and larvae homozygous for null alleles have small imaginal discs and reduced frequencies of mitotic figures in the brain. Dominant mutations originally identified as enhancers or suppressors of zeste are gain-of-function alleles of polycombeotic. The type and variety of defects displayed by different mutations in this gene indicate that the product might be involved in chromosome structure and/or function.
T HE polycomb-group of genes has become the subject of intense study recently, because of the
variety of homeotic transformations caused by muta- tions in these genes, and because of their role in the maintenance of appropriate expression of the bithorax and Antennapedia gene complexes (BX-C and ANT- C, respectively). These different loci are considered a ‘group’ because mutations in each of these genes cause similar phenotypes, because loss-of-function muta- tions in a gene can enhance the phenotype of loss-of- function mutations in another gene of this set (JUR- GENS 1985; KENNISON and TAMKUN 1988), and be- cause duplications of a gene can suppress the pheno- type of loss-of-function mutations in another gene of this set (KENNISON and RUSSELL 1987). The polycomb- group is thought to contain as many as 40 different genes (JURGENS 1985), including Polycomb (PC) (DE- NELL 1978; DUNCAN and LEWIS 1982), Polycomb-like ( P c l ) (DUNCAN 1982), Addit ional sex combs (Asx) ( JUR- GENS 1985), Posterior sex combs (Psc) ( JURGENS 1985), Sex combs on midleg (Scm) (JURGENS 1985), super sex combs (sxc) (INGHAM 1984), extra sex combs (esc) (STRUHL 1981), polyhomeotic ( p h ) (DURA, BROCK and SANTAMARIA 1985), Sex comb extra (&e) (BREEN and DUNCAN 1986), pleiohomeotic (pho) (DENELL, HUMMELS and GIRTON 1988), and extra dent ic les (exd) (WIE- SCHAUS, NUSSLEIN-VOLHARD andJuRGENs 1984). Mu-
tants of this group display a transformation of the second and third legs to the first leg in adults and a transformation of the ventral setal belt patterns of thoracic and abdominal segments to patterns of more posterior segments in larvae. Such transformations, as well as the altered expression of RNAs and proteins from BX-C and ANT-C as shown by in s i tu hybridi- zation to tissue sections and antibody staining of the CNS and imaginal discs, suggest that these complex loci are being inappropriately expressed in some tis- sues (BEACHY, HELFAND and HOGNESS 1985; STRUHL and WHITE 1985; WEDEEN, HARDING and LEVINE 1986; GLICKSMAN and BROWER 1988). Recent exper- iments have suggested that homeotic transformations caused by poly-comb-group mutations may be due to an instability in the segment-specific expression of ANT-C and BX-C (STRUHL and AKAM 1985; DURA et al. 1987).
Another subject of intense study has been the zeste gene. zeste was discovered due to the effect of a gain- of-function allele, z’, on the expression of the white locus (GANS 1953). This effect was shown to be de- pendent on the number and proximity of copies of the white locus. Later it was determined that zeste also had an influence on the complex loci bithorax (BX-C) and decapentaplegic (DPP-C) (KAUFMAN, TAKAZA and SUZUKI 1973; GELBART and Wu 1982). A common feature of these complex loci is the influence of close
Genetics 125: 91-101 (May, 1990)

92 M. D. Phillips and A. Shearn
proximity of gene copies on expression, an effect called transvection (LEWIS 1955).
Our studies on the polycombeotic gene, and those of WU et al. (1989), suggest that the functions of the polycomb-group of genes and the zeste gene may be related at some fundamental level. polycombeotic (pco) , originally identified as letha1(3)1902 (1(3)1902) (SHEARN et al. 1972, 1978; SHEARN and GAREN 1974; SHEARN 1977; SHEARN, HERSPERGER and HERSPER- GER 1978), displays a wide range of phenotypes, de- pending on the type of mutation observed. Leaky alleles, such as the temperature-sensitive alleles
formations similar to those seen in other polycomb- group genes. Null alleles, such as cause a small disc phenotype and block mitosis. In addition, alleles of the Enhancer of zeste gene ( E ( z ) ) (KALISCH and RASMUSON 1974), which can either enhance or sup- press the zeste-white interaction, have been identified as gain-of-function alleles of polycombeotic.
The wide range of phenotypes caused by different alleles suggest that the primary role of polycombeotic is in a basic cellular process, such as chromosome struc- ture or function. This gene may be one of many that are a link between cell proliferation and determina- tion.
PCOPC025hs ' PCOox73hhs , and p ~ ~ ~ y ~ ~ ~ ~ ~ cause homeotic trans-
MATERIALS AND METHODS
Fly strains and culture: All strains were maintained on standard culture medium at 20°, unless stated otherwise. The pco alleles used in this study were induced by N-methyl- A"-nitro-N-nitrosoguanidine (pco""') or by ethylmethane- sulfonate ( p ~ ~ ~ y ~ ~ ~ ~ ~ , pconhoUh, pcoox73hhs , and pcop'""") (SHEARN, et al. 1971, 1978; SHEARN, HERSPERCER and HERSPERCER 1978). The E(z) ' mutation was induced by ethylmethane- sulfonate (KALISCH and RASMUSON 1974), and the Su(z)?OZ mutation was induced by X-rays (W. GELBART, unpublished results); both the E ( z ) and Su(z)?OZ stocks were kindly provided to us by RICK JONES in the laboratory of WILLIAM GELBART. For the other stocks used, the Df(?L)vin2 stock was provided by the Mid-America Drosophila Stock Center, Bowling Green, Ohio, the Tp(3)PZO stock was provided by E. B. LEWIS, the Dp(?;3)S2a? stock was provided by MINX FULLER, and the Df(3L)lxd' and D f ( ? L ) l ~ d ' ~ stocks were provided by VICTORIA FINNERTY. The breakpoints associ- ated with the above rearrangement stocks are listed in Table 1. Other stocks used are described in LINDSLEY and GRELL (1 968) or LINDSLEY and ZIMM (1 985, 1986, 1987).
Adult phenotype analysis: Adult legs were examined at 10-50X magnification using a stereomicroscope.
T o examine the adult homeotic phenotype, homozygous
TM? or mwh ~ C O " ) " ~ red eITM3 males and their eggs were collectedat 20" for 24 hr, shifted to 20°, 24", or 27", and allowed to develop into adults. Viability of male progeny was calculated as the percent of mutant males eclosing relative to the number 0fpcoJ"~'~~~~red/TM3 control brothers. The "extra sex combs" phenotype was analyzed in two ways. Some males were observed live under a stereomicroscope and the total number of legs displaying sex comb teeth was scored. Other males were dissected and their ventral thor- aces were mounted on slides. The number of sex comb teeth
PCOPC"25hs red females were crossed to either p ~ 0 @ ~ ' ~ ~ ~ ~ red/
TABLE 1
Breakpoints of rearrangement chromosomes used
-
Df(3L)lxd" 67E; 68C10-15
DJ(3L)PZO" 67F: 68E3-4
DJ(3L)uin2 67F2-3: 68D6
Dp(3;3)SZa? 67D9-1 1: 68.41-2 TP(3)PZO 67F; 68E3-4: 89E
D J ( ~ L ) P c ( D ~ ( ~ L ) A S C ) 78D1-2: 79A4-Cl
Dp(3;3)PZO" 67F; 68E3-4; 89E
Reference
SCHOTT, BALDWIN and FINNERTY ( 1 986)
SCHOTT, BALDWIN and FINNERTY (1 986)
LEWIS (1980) HAYNIE (1983) AKAM ( 1 978) LEWIS ( 1 980) CRAYMER (1 984) LEWIS (1 980) ~ ~~~~~ ~~ ~
Derived from the Tp(3)PZO chromosome by recombination, ~~
on each leg of these males was then scored using a compound microscope at 1 OOX magnification.
Oogenesis and maternal-effect phenotype anal sis: To examine the effect of p ~ # ~ ' ~ ~ ~ ~ , ~ c o ~ ~ ~ ~ ~ ~ ' , and pcomYy Y yhs hom- ozygosis on oogenesis, 20 4-day old 99, derived from het- erozygous parents raised continuously at the permissive temperature of 20° , were mated to 20 88 homozygous for the marker mutation red. All of the progeny of this cross were heterozygous for a pco temperature-sensitive allele. Eggs were collected from these crosses for two 4-hr periods daily. Eggs laid during each 16-hr overnight period were discarded. The parents were maintained for three days at 20" to determine control values for fecundity and viability. For that three day period each pcoox736h5 female laid on average 1.7 eggs/hr and 66% of those eggs gave rise to viable progeny; each COP^^'^^^^ female laid on average 1.4 eggs/hr and 37% of those eggs gave rise to viable progen P c ~ ~ ~ ~ ~ ~ ~ ' females laid no eggs. The ~ c o ~ ~ ~ ~ ~ ~ ~ ' and pcop"" 7 ; hr
parents were then shifted to a restrictive temperature of 29" for 5 days and then returned to 20" for an additional 10 days. After each 4-hr egg collection the number of eggs was counted and those eggs were incubated at a permissive temperature of 20". Viability was calculated as the fraction of the eggs which gave rise to viable progeny. As a control, honlozygous red 99 were mated to homozygous ~ c o ~ ~ ~ ~ ~ ~ ~ red 66.
T o examine the cuticular phenotype of embryos derived from COP^""^' homozygous females, COP'^'^^^^ red 99 were raised at 20" and mated to COP^(^^^^^ red or mwh red e 66. they were then shifted to 30", eggs were collected at 4-hr intervals and allowed to develop at 20" for about 24 hr. The eggs were dechorionated, the embryos were mounted in Hoyer's medium and examined using the method de- scribed by VAN DER MEER (1977) and modified by WIE- SCHAUS and NUSSLEIN-VOLHARD (1 986). Hoyer's medium was prepared by mixing 30 g Gum Arabic (Acacia) and 50 nil distilled water overnight in a tightly sealed container, adding 200 g chloral hydrate (a few grams at a time while stirring), and 20 g glycerol. The resulting solution was filtered through several layers of cheesecloth and stored in a moisture-tight container.
T o examine the female sterility phenoty e, homozygous ~ c o ~ ~ ~ ~ ~ ' ~ ' females and heterozygous pcomyy3phs/TM3 females were mated to heterozygousp~o"~~~'~"/TM3 males and placed at 29 O . Following 4 and 8 days of exposure to the restrictive temperature, females were dissected and their ovaries in- cubated for 5 min in 1 pg/ml Hoechst 33342 in Ringer's solution, washed in Ringer's solution, fixed for 5 min in acetone at -20", mounted in 90% glycerol, and examined under epifluorescence illumination.

pco Is a Polycomb-Group Gene 93
I
I1
Dp(3;3)S2a3
TM6, e h 9 9 % mwh red Pr Le mwh red Pr Le C M
sc z l w is . H z ) + Dp(3;3)SZa3 99% - * TM3, Sb e Ser ‘ mwhredPrLe rn
s c z l w i s ’
s c z w A 9 9 l i s i
I11 mwh red Pr Le Dp(3;3)S2a3 E(z) I E(z) TM3, Sb e Ser TM3, Sb e Ser mwh red Pr Le Dp(3;3)S2a3 sc z lw is
T (33
Gene Dosage: (+I+) (+I+/+)
Interaction analysis: To examine the effect of three doses of pco+ on the phenotype caused by a deficiency of Polycomb, Dp(3;3)SZa3/TMl 99 were mated to Df(3L)Pcl TM3 dd at 27 ’. The “extra sex combs” phenotype of Dp(3;3)SZa3/Df(3L)Pc dd was compared to Df(3L)PcITMI control 88. The significance of the differences was evaluated using the t-test.
Brain squashes: Brain lobes and ventral ganglia were dissected out of larvae in Ringer’s solution. The brains were then incubated in 1 % sodium citrate for 10 minutes, fixed in a 1 : 1 mixture of acetic acid:methyl alcohol for 3 min, and squashed in a drop of a filtered solution of 2% natural orcein dissolved in 1 : 1 glacial acetic acid:lactic acid on a precleaned slide and sealed with clear nail polish.
Allele activity test: The activity of the E(z)’ and Su(z)301 mutations were assayed according to the following cross (see also Figure 1): five sc z’ W”/SC z’ w“; E@)’ [or Su(z)301]/ TM3, Sb e Ser virgin 99 were mated to five +/Y; Dp(3;3)SZa3/mwh red e PrL W . The eye color of the progeny was scored to determine the relative enhancement or suppression of the interaction between zeste and white. The eye color of the non-Pr“,, non-Sb progeny (genotype: sc z’ w’”/+; E(z)’ [or Su(z)3Ol]/Dp(3;3)SZa3 and sc z’ w’”/Y; E@)’ [or Su(z)3OI]/Dp(3;3)SZa3) relative to the eye color of the Pr‘,, non-S6, progeny (genotype: sc z’ w”/+; E(z)’ [or Sz~(z)301]/mwh red e Pr’ and sc z’ w’’/Y; E(z)’ [or Su(z)301] /mwh red e Pr“) allowed us to determine if there was any change in the effect of the dominant mutation when two wild-type copies of the gene were present, instead of one wild-type copy, in addition to the mutant copy of the gene.
In the case of E(z) ’ , if the non-Pr”, nonSb (E(z)/+/+) class had a lighter eye color ( i e . , more like the lemon-yellow of z’/z’ 99) relative to the Pr‘. non-Sb (E( z ) /+) class, then this would indicate that the extra wild-type copy of the gene had enhanced the zeste-enhancement phenotype of E(z)’. If the non-Pr’,, non-Sb (E(z)/+/+) class had a darker eye color (?.e., less like the lemon-yellow of z’/z’ 99) relative to the P r ‘ , non-Sb (E(z) /+) class, then this would indicate that the extra wild-type copy of the gene had suppressed the zeste- enhancement phenotype of E(z)’. If the non-PrL, non-Sb (E(z)/+/+) class had the same eye color as the PrL, non-Sb (E( z ) /+) class, then this would indicate that the extra wild- type copy of the gene had no effect on the zeste-enhancement phenotype of E(z) ’ .
In the case of Su(z)301, if the non-Prl, non-Sb (Su(z)/+/ +) class had a lighter eye color (i.e., more like the lemon-
FIGURE 1 .--Scheme for establish- ing E(r)’ and Su(r)3Ul activity. The progeny of generation 111 were ex- amined and compared for their eye phenotype. The relative dosage of mutant and wild-type copies for each genotype is shown. The Su(r)301 crosses were identical except the E(z)’ chromosome was replaced by the Su(z)3UZ chromosome.
yellow of z’/z’ 99) relative to the PrL, non-Sb (Su(z)/+) class, then this would indicate that the extra wild-type copy of the gene had suppressed the zeste-suppression phenotype of Su(z)301. If the non-Ps, non-Sb (Su(z)/+/+) class had a darker eye color (i.e., less like the lemon-yellow of z’/z’ S) relative to the PrL, non-Sb (Su(z)/+) class, then this would indicate that the extra wild-type copy of the gene had enhanced the zeste-suppression phenotype of Su(z)301. I f the non-P8, nonS6 (Su(z)/+/+) class had the same eye color as the Ps , non-Sb (Su(z)/+) class, then this would indicate that the extra wild-type copy of the gene had no effect on the zeste-suppression phenotype of Su(z)301.
RESULTS
Null alleles are zygotic lethals and display a mi- totic defect: In order to determine the null phenotype of pco, we first had to identify a deficiency for the gene. Earlier genetic mapping experiments indicated that the pco gene is located at approximately 30 on the third chromosome genetic map (SHEARN et al. 1978). We attempted to further localize the gene and examine the effects of a complete loss of function by finding deficiencies that uncover the pco gene. Several deficiencies in the area were tested in trans with a chromosome carrying the allele pco’yu2. Three defi- ciencies Df(?L)lxd6, DF(3L)lxdJ5, and Df(3L)P20, do not complement the lethality of P C O ~ ~ ’ ~ , while the deficiency Df(3L)vin’ does complement ~ C O ’ ~ ~ ~ . These data indicate that pco is within the 67E-F chromosomal region (see Table 1 and Figure 2).
The discovery of deficiencies uncovering pco al- lowed us to test the activity of PcoIyo2, a homozygous lethal allele which displays the most extreme pheno- type of all the pco alleles examined: homozygous pco1902/pcoJ902 larvae have small imaginal discs and die shortly after puparium formation (SHEARN et al. 1978). Larvae homozygous for pco1yu2 and larvae trans-heterozygous for pcoIYu2 and the deficiency Df(?L)Zxd‘ were examined to determine their effec- tive lethal phase. In both cases, the mutant larvae died

94 M. D. Phillips and A. Shearn
n
68 I69
~ ~ W R K 2,"Map of deficiencies near to or uncovering pco. Thc deficiencies Df(3L)lxd" Df(3L)lxdJ3, and Df(3L)P20 do not comple- ment the n1ut;Ition proJ"'", but the deficiency Df(3L)uin2 does com- plement. These data indicate the pco gene is located with the 6 7 F F region. The bars indicate the extent of each deficiencv. with the open portions indicating uncertainty in the breakpoint locations.
shortly after puparium formation (see Table 2). This shows that ~ C O ' ~ ' ) ~ homozygotes have the same phe- notype as ~ C O ' ~ * ~ hemizygotes, indicating that this al- lele is effectively null.
The small disc phenotype of pco null mutations led us and others to suspect that pco mutations could affect the process of mitosis. We compared squashes of brain tissue from control and mutant larvae at several times in development. The results are shown in Table 3 and Figure 3. The mutant squashes show virtually no discernible mitotic figures, whereas they are fre- quently seen in control larvae. Other workers have reported similar mitotic defects in ~ C O ' ~ " ~ mutant lar- vae (GATTI et al. 1983; GATTI and BAKER 1989).
Weak alleles cause adult homeotic phenotypes: In an earlier study it was reported that adult males and females homozygous for the temperature-sensitive al- lele P c o ~ ~ ~ ~ ~ ~ ~ , derived from heterozygous parents, had transformations of their second and third legs to first legs if they had been exposed to a restrictive temper- ature for one or two days during larval development (SHEARN, HERSPERCER and HERSPERGER 1978). Adults homozygous for ~ c P ~ ~ ~ ~ express similar trans- formations. In order to quantify this phenotype, males homozygous for the temperature-sensitive pco allele p ~ ~ ~ ~ ~ ~ ~ ~ ~ ' or trans-heterozygous for ~ c O P C ~ ) ~ ~ ~ ~ and the null allele, ~ c o ' " ' ~ ~ , were raised continuously at 20", 24", and 27", and the frequency of sex comb teeth on the first, second, and third legs were scored. As shown in Table 4, homozygous ~cOPC'"~~ ' males raised at 20" had, relative to wild type, reduced viability, down to 5 1 %, and a slight increase in the number of sex combs, usually having sex comb teeth on one or both second legs. Homozygous ~ C O P C ' " ~ " ~ males raised at 24" had lower viability (43%) and a greater number of sex combs, always having sex comb teeth on both second legs, and usually having sex comb teeth on one or both third legs. ~ C O P C ' ~ ~ ~ ~ ' males raised continuously
TABLE 2
Lethal phenotype in homozygous and hemizygous f ~ e o ' ~ ~ ' larvae
TABLE 3
Average number of mitotic figures in brains of wild-type and peo mutant larvae
Y 4
125*8 2 2 1 I55 f 18 2 + 1
5 1.51 + 27 4 + 1
s:mulhpro J""2/y+ T.1.13 females were crossed to y:Df(3L)lxd"/y+ TJi3 males. pro '"'''/Df(3L)lxdh mutant larvae were distinguished from prol"""/~+ T.M3 and I)f(3L)lxd"/y+ T.\f3 ("wild type") rontrols by their lighter-colored (yrllour) mouth hooks.
l , ' t ( ; t .R+. :<.-\ 'ct~~t.al plllgliott I)llcm)t\lx', (;I) S~II;ISII 01 ;I \+ild- t y x larval brain. (1)) s q u a d t of a pro'"'"/~f(3t.)/sr~" larva1 brain. Arro\vlte;~tls in (e) indicate nornwlly condensed chromosomes, which are rarely seen i n mutant brains.
at 27" showed no adult viability, but pharate adults had sex comb teeth on all legs. At all temperatures, ~ c o ' ~ ' ~ ~ / P c o P ~ ' ~ ~ ~ ~ ~ ~ trans-heterozygotes had lower viabil- ity and higher numbers of extra sex combs than pcOPCoZshs homozygotes. Legs from selected males are shown in Figure 4. Two other alleles also express this homeotic phenotype. Homozygotes of ~ ~ 0 " ' ~ ~ ~ ~ ~ ~ ~ are even more sensitive to high temperature than either
or COP"'^^^'. Exposure to 27" for as short as 8 hr during the third larval instar can cause transfor- mations of the second and third legs to first legs (data not shown). Homozygotes of Pconhooh are not temper- ature-sensitive. They are lethal at the pharate adult stage and express leg transformations regardless of
P C O ~ ~ 7 3 h h s

pco Is a Polvcomb-Group Gene 95
TABLE 4
Viability and extent of transformation of peo temperature-sensitive mutant adults
20" ( I 00%) 2.00 f 0.00 266 51% 2.83 f 0.79 x2 40% 5.03 f 0.74 51 24 ( I 00%) 2.00 f 0.00 361 43% 3.47 f 1.07 90 32% .5.75 * 0.44 67 27" ( I 00%) 2.00 f 0.00 237 0% 6.00 f 0.00" 24 0%
FIGURE 4.--Adult homeotic phe- notype. (From left to right) first, sec- ond, and third legs of: (a) wild-type adult male raised at 24". (b)p~op'"~'"/ ~ C O " ' " ~ male raised at 20", and (c) p~OP~'~'*'/pcoI~'~ male raised at 24'.
temperature. Weak alleles cause female sterility and/or mater-
nal-effect lethality: In an earlier study it was found that ovaries transplanted from larvae homozygous for the null allele, pco'"''*, are sterile and that homozygous
thality (SHEARN, HERSPERGER and HERSPERGER 1978a). In order to further characterize these two oogenesis defects, homozygous ~ c o ~ J ~ ~ ~ ' ~ ~ ' , p ~ $ r ~ ' ~ ~ ~ ~ ~ , and
females were mated to red males and incu- bated at 20" for 3 days to collect control eggs. These sets of parents were then shifted to 29" for 5 days to examine the effect of restrictive temperature and then returned to 20" for an additional 10 days to monitor recovery. All of the eggs collected, regardless of the temperature at which they were collected, were incu- bated at 20". As a control, homozygous red females were mated to homozygous ~ c o ~ ~ ~ ~ ~ ~ ' males and treated as above.
causes nonconditional female sterility. No eggs at all were recovered from homozygous females even when maintained continuously at a permissive temperature. In order to ensure that little or no pco+ factor was produced within mutant ovaries, homozy- gous P c o ~ ~ " ~ ' ) ~ ~ females were raised at 29". Examina- tion of ovaries dissected from females following 4 days at the restrictive temperature revealed a normal num- ber of ovarioles and apparently normal stage egg
PCOox73hh" adult females expressed maternal-effect le-
p C O o x i 3 h h . t
pCOm+/3Yh.~
chambers up to stage three or four, using the staging of KING (1970). However, as shown in Figure 5c, there was degeneration of the nurse cells, and little growth in size of egg chambers beyond the earliest stages. Further exposure to the restrictive tempera- ture increases the severity of the mutant phenotype- ovarioles contain almost exclusively small egg cham- bers and rarely are identifiable nurse cells found (Fig. 5d). Identically treated heterozygous sisters displayed normal ovarioles and egg chambers, as shown in Fig- ure 5, a and b.
fect lethality. Upon shifting to 29" there is no signif- icant loss of fecundity (Figure 6), however, the viabil- ity of the eggs laid, all of which are heterozygous for p ~ $ r ~ ' ~ ~ ~ ~ ' drops from 39% to near zero in just four hours (Figure 6). The viability of the eggs collected from these mothers remains low as long as the parents are incubated at 20". However, within one day after returning these parents to a permissive temperature, the viability of their progeny returns to preshift levels. The rapid loss and rapid return of viability indicates that the eggs of homozygous COP^^)^^^^' mothers must only be at a pernlissive temperature either just before or just after egg-laying for the progeny to be viable. I n other words the tenlperature-sensitive period for maternal-effect lethality is late in oogenesis and/or early in embryogenesis.
PCOPc025hr causes temperature-sensitive maternal-ef-
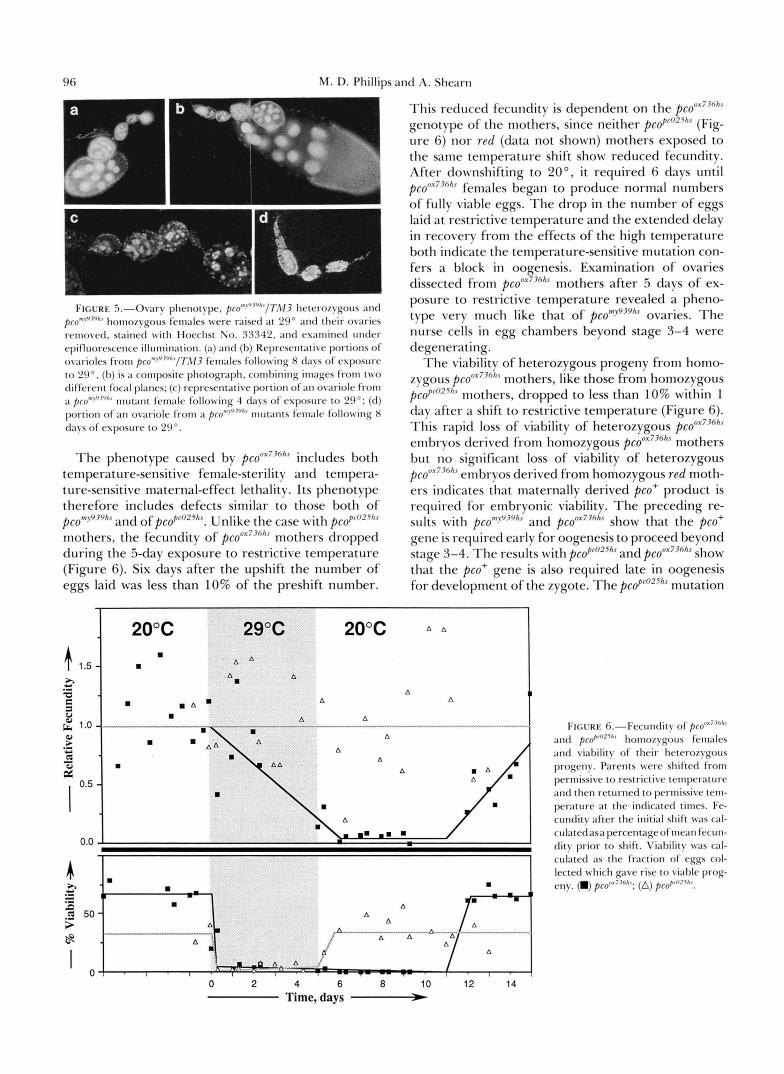
96 M. D. Phillips and A. Shearn
l:l(;[xF . - ) . - o \ : l l . \ p l l e l l o t \ ~ ~ < ~ . / ,~~~" '~ ' ' ; ' ' / , , /~ ' . \ /~ l l ~ l < ~ l ~ o / \ ~ o ~ l \ ;111d
prom>""'"' honlozygous fi~nmles were raised a t ?!I" and t lwir o\.;wies ren1ovetl. stained wi th tloechst So. 333411, and examined under epifluorescetlre illunlination. (a) m r l (b) Representative portions of ov;lrioIes fro111 ~ c o ~ + ' ~ ~ ~ ' / T . ~ I J females following 8 chys of exposure to 29". (b) is a composite photograph, conlbining inlages from two different focal planes; (c) representative portion of an ovariole from
portion of a n ovariole from :I ~ c o ~ ~ ~ ~ " ~ ~ n1ut;mts fen1ale following X clays of exposure to 29".
;I p r O m t ' 1 7 ' ~ h c mutant female following 4 days of exposure to 29"; (d)
The phenotype caused by P c o " ~ ' ~ ~ ~ ' includes both temperature-sensitive female-sterility and tempera- ture-sensitive maternal-effect lethality. Its phenotype therefore includes defects similar to those both of pc~"')~''~" and of p ~ 8 ' " ~ ~ ~ ' . Unlike the case with P C ~ " ~ " ' ~ ' mothers, the fecundity of ~ ~ 0 ~ ~ ~ ~ ~ ' ' ' mothers dropped during the 5-day exposure to restrictive temperature (Figure 6). Six days after the upshift the number of eggs laid was less than 10% of the preshift number.
This reduced fecundity is dependent on the P c o ~ ~ ~ ~ ~ ~ ~ ' genotype of the mothers, since neither PCOPC"'~~~' (Fig- ure 6) nor red (data not shown) mothers exposed to the same temperature shift show reduced fecundity. After downshifting to 2 0 ° , it required 6 days until
of fully viable eggs. The drop in the number of eggs laid at restrictive temperature and the extended delay in recovery from the effects of the high temperature both indicate the temperature-sensitive mutation con- fers a block in oogenesis. Examination of ovaries dissected from ~ c o ~ ~ ~ ~ ~ ~ ~ mothers after 5 days of ex- posure to restrictive temperature revealed a pheno- type very much like that of ~ c o ~ ~ ~ ~ ~ ~ ~ ovaries. The nurse cells in egg chambers beyond stage 3-4 were degenerating.
The viability of heterozygous progeny from homo- zvgous P c o " ~ ~ ~ ~ ~ ' ~ ' mothers, like those from homozygous &#''25"r mothers, dropped to less than 10% within 1 dav after a shift to restrictive temperature (Figure 6). This rapid loss of viability of heterozygous ~ c o ~ ~ ~ ~ ~ ~ ~ ~ embryos derived from homozygous P c o ~ ~ ~ ~ ~ ~ ~ mothers but no significant loss of viability of heterozygous
ers indicates that maternally derived pco+ product is required for embryonic viability. The preceding re- sults with p ~ 0 ~ ~ " ~ ~ ~ ~ and ~ c o ~ ~ ~ ~ ~ ~ . ' show that the pco+ gene is required early for oogenesis to proceed beyond stage 3-4. The results with p ~ ~ P ~ ~ ~ ~ ~ ~ ~ ' a n d P c o ~ ~ ~ ~ ~ ~ ~ show that the pco+ gene is also required late in oogenesis for development of the zygote. The p ~ 8 ~ ~ ~ ~ ~ mutation
PC0""736hr females began to produce normal numbers
PCOox736hs embryos derived from homozygous red moth-
FIGURE fi.-Fecunclity of pr0ox77hh' and prOP'"2'h' homozygous fenlales ;und vi;lbility of their heterozygous progeny. Parents were shifted lion1 permissive to restrictive tenlperature and then returned to permissive tern- perature at the indicated times. Fe- cundity after the initial shift \vas cal- culated;~sapercentageofme;lnfecull- dity prior to shift. I'iability w a s cal- culated a s the fraction of eggs col- lected which gave rise to vi;111le prog- eny. (W) pro"x"hh'; (A) PrOP'"-"h'.

pco Is a Polycomb-Group Gene 97
TABLE 5
Cuticle phenotypes of progeny from p c ~ ~ ~ * ~ ~ ' females shifted to 30"
Hollrs ;Ifrrr upsllifi Tl/T2 (%) A I / , \ ? (%) Egg viability (%)
0-4 1 10 50 4-x 7 14 I X x-12 X 46 5
I'crcent of l a r v ; ~ tlispl;lying ;I tr;msfornlation of denticle belts of citller the first or second tlloracic segments (Tl/T2) toward a I X I I I ~ I - I I resembling the eighth abdominal segment ;Inti those dis- playing :I tl-;lllsfo~rn;lrio~~ of denticle belts of either the first or scrontl ;Ibtlominal segnlents (A 1 /A2) toward a pattern resembling the eighth ;Il)dominal segment.
FICCRE 7.-l.;tt-\;II l l o l l l ( . o l i ( ~ ) I I ( ~ I I I ~ I \ ~ ) ~ ~ . (;I) ( i l r i c l e of a nornml embryo from an egg h i d 1)). ;I proP"'""'/+ fktn;~lc 8- 12 hr following a shift t o X)". (b) cuticle of a partially transfortnctl embryo from an egg Iaid b y ;I prOP"'~'hL/pCoP"'Z'h' female 4-X hr fbIIo\ving :I shift t o 30". (c) ;Ind (d) cuticles of strongly transfornvxl embryos from an cgg laid by :I p~oP""rh'/pcOpr""h~ fell1;de 4-8 hr following a shift t o 3 0 " .
only affects the late requirement. We examined the cuticle of embryos derived from homozygous p ~ O p r ~ ) ~ ~ ~ ~ mutant mothers in order to examine the maternal- effect phenotvpe. Females homozygous for p ~ O p r ~ ) ~ ~ ~ . '
were raised at 20" and mated to males also homozy- gous for pcOProZShs. The females were then shifted to SO" and eggs were collected at 4-hr intervals and allowed to develop at 20" for about 24-hr. The eggs were dechorionated and the embryos examined. The resulting phenotypes are listed in Table 5 and shown in Figure 7. After 8-1 2 hr of exposure of homozygous ~COPC"~~".' mothers to the restrictive temperature, a large proportion of the homozygous p ~ $ c ~ ) ~ ~ ~ ~ ' larvae show transformations of the thoracic and/or abdomi- nal segments toward a denticle pattern resembling the eighth abdominal segment. A smaller-scale test of the effect of the zygotic genome on the cuticle phenotype, in which homozygous P c O ~ ~ ' ' ' ~ ~ ~ females were mated to homozygous mwh red e males, shifted to 30", and their eggs collected and examined as in the experiment above, indicated that few, if any, heterozygous p ~ O p r ~ ) ~ ~ ~ ~ larvae of homozygous COP'^)'^^' mothers have transformations of cuticle denticle belt patterns (data not shown). So while the cuticle phenotype is rescued by the wild-type copy of pco, the extremely low viabil- ity of the heterozygous progeny of homozygous tem- perature-sensitive mothers (Figure 6) demonstrates the maternal-effect lethality is not rescued by the wild- type copy of pco.
A duplication of pco interacts with a deficiency of Polycomb: The maternal-effect larval and zygotic adult homeotic transformations in weak pco mutants suggest that pco is a polycomb-group gene. In order to further determine the relationship of pco to other members of this group, we examined the interaction of pco with Polycomb. The deficiency mapping of pco indicated that the tandem duplication Dp(3;3 )S2a3 , which duplicates the 67E-F region, included pco. We examined the phenotype of adult males containing this duplication in trans with Df(3L)Pc (also known as Df(3L)ASC), a deficiency for the Polycomb gene. KEN- NEON and TAMKUN 1988) previously used this type of interaction as a screen for other polycomb-group genes. The results are summarized in Table 6. The Df(3L)Pc/7"1 control males, which have only one copy of the PC+ gene and two copies of the pco+ gene, showed sex comb teeth on all legs with an average of over 4 teeth per leg on the second leg and over 2 teeth per leg on the third leg. In contrast, the Df(3L)Pc/Dp(3;3)S2a3 males, which have one copy of the PC+ gene and three copies of the pco+ gene,
TABLE 6
Interaction of a duplication for polycombeotic with a deficiency for Polycomb
,\vcKlg<- 110. of kg. Average 110. of scs COIIIII reeth/leg
(;clloi\pc \vith Sex c - o n ~ l ,
.Y Ic.rtll h' 1.1 1.2 1.3
I ) / (31.)I'r/T.\l I 4.5 6 .0 f 0 . 0 54 I 1.26 * 1 ..53 4..52 f 2.91 2.49 * 2.4x I?/ (3I.)f'r/Dp(3: 3)S2a3 48 5. I * 0.x" 90 12.6 I f 1 .04 2.27 f I .X7" 0.79 f 1.13"
" l n d i c a t c h signilic:lnrly diffiwnt li.0111 the l ) f ( 3 f , ) P r / T . ~ / / control ;Iccortling t o the t-test (I' < 0 . 0 0 1 ) .

98 M . D. Phillips and A. Shearn
have on average, fewer legs with sex comb teeth and fewer teeth per leg on the second and third legs. Thus, a duplication carrying an extra copy of the wild- type pco gene partially suppresses the Polycomb mutant phenotype.
Gain-of-function alleles enhance or suppress the reste-white interaction: Similar crossover map loca- tions have been reported for the Enhancer of zeste gene (KALISCH and RASMUSON 1974) and the polycombeotic gene (SHEARN and GAREN 1974). In addition, other workers reported that E(%)’ alleles display a Polycomb- type phenotype and have a small-disc lethal phenotype (WU et al. 1989; R. JONES, personal communication). This led us to test whether E(%)’ and pco were muta- tions in the same gene. E(z)’ and ~ C O ’ ~ ’ ~ , both homo- zygous lethal mutations, were tested in trans with the deficiency D f ( 3 L ) P 2 0 and in trans with the transposi- tion chromosome Tp(3)P20, which contains the defi- ciency D f ( 3 L ) P 2 0 and the duplication D p ( 3 ; 3 ) P 2 0 . Both E(z)’ and P C O ’ ~ ~ ) ~ were lethal in trans with the deficiency, but viable in trans with the deficiency and the duplication. These data indicate that the defi- ciency lacks the sequences for both E ( z ) and pco and that the duplication D p ( 3 ; 3 ) P 2 0 contains sequences for both E ( z ) and pco, and that the genes are located very close to each other. We then tested E(z)’ in trans with P C O ’ ~ ~ ’ ~ . E ( ~ ) ‘ / ” O ’ ’ ’ ~ trans-heterozygotes did not survive, indicating that they were both alleles of the same gene. Another mutation, S u ( z ) 3 0 1 , a recessive lethal and dominant suppressor of zeste allelic to E(z)’ (WU et al. 1989), was also tested and it as well did not complement ~ c o ‘ ’ ) ~ ) ~ for lethality.
The D p ( 3 ; 3 ) S 2 a 3 tandem duplication containing pco provided insight into the activity of E(%)’ and Su(z)301. By placing E(z)’ and S u ( z ) 3 0 1 in trans with Dp(?;3)S2a?, and comparing the phenotype of these flies to siblings which have E@)’ and S u ( z ) 3 0 1 in trans with a normal third chromosome, we could examine the effect of an extra wild-type copy of the pco gene on the mutant zeste-modifying phenotypes of E@)’ and Su(z)30l. In the cross involving E(z) ’ , 348 prog- eny were scored, and the 54 E(z)/+/+ females were observed to have an identical dark-orange eye color with respect to their 66 E(z)/+ sisters; the 25 E(z) /+/ + males were observed to have an identical light- orange eye color with respect to their 5? E(z ) /+ brothers. Therefore, the extra wild-type copy of the gene had no visible effect on the E(z)’ eye color phenotype. Using the terminology of MULLER ( 1 932), since the mutation’s phenotype was unaffected by the addition of wild-type copies, the mutation was neo- morphic in nature (causing a change in the nature, or a new activity), with respect to the effect on the zeste- white interaction. In the cross involving S u ( z ) 3 0 l , 3 12 total progeny were scored, and the 50 Su(z) /+/+ males were observed to have a consistent, distinctly orange eye color while their 51 Su(z) /+ brothers had
a virtually wild-type red-brown eye color. The extra copy of the wild-type gene had suppressed the effect of the Su(z)301 mutation on the eye color. Using the terminology of MULLER ( 1 932), since the addition of an extra wild-type copy had lessened its phenotype, the S u ( z ) 3 0 I mutation was antimorphic in nature (causing an opposite nature, or a competing activity), with respect to the effect on the zeste-white interaction.
In contrast to the dominant effects of E(z)’ and Su(z)30l, flies heterozygous for the pco null allele Pco’~‘~ show no detectable change in the interaction of zeste and white (data not shown).
Because of the data indicating the allelism between E(z) and pco, we refer to the E(%) alleles as alleles of pco (i.e., PCO”(~)’ and pco~su(z)sol).
DISCUSSION
Loss of function alleles: The viability of pco null mutant larvae is comparable to wild type (Table 2). This was originally interpreted to mean that the pco+ gene product was required by imaginal cells but not larval cells (SHEARN 1977, 1980). Null mutant larvae have small imaginal discs and show a dramatic reduc- tion in brain mitotic figures (Table 3 and Figure 3). GATTI and BAKER (1989) have reported mitotic de- fects in larvae homozygous for mutations in many genes whose phenotypes include late lethality and poorly developed imaginal discs. They reported that third-instar ~ C O ’ ~ ’ ~ mutant brains show very low mi- totic indices and frequent chromosome fragmentation and concluded that the effects of the mutant were sufficiently specific on a central aspect of mitosis, that the pco locus was likely to encode an essential cell- cycle function. The pco+ product may function as a structural element of the nucleus or of chromosomes.
If this interpretation were correct, then one would expect that the pco+ product would also be required during embryonic development and oogenesis (and spermatogenesis). Analyses of temperature-sensitive alleles have demonstrated that the pco+ product is indeed required during those stages. The normal embryonic development of null mutant larvae derived from heterozygous mothers is apparently due to the presence in such homozygotes of enough pco’ product of maternal origin to support embryonic develop- ment. This interpretation is supported by tempera- ture-shift experiments performed with mothers homozygous for the temperature-sensitive alleles
to restrictive temperature, heterozygous embryos, de- rived from the cross of homozygous mothers and nonmutant fathers, lose the ability to hatch into larvae (Figure 6). This initial affect of the temperature up- shift is on oocytes or eggs that had completed nearly all of their development at a permissive temperature. It represents an effect on the mutant PCO product
PCOPC025hs and ~ c o ~ ~ ~ ~ ~ ~ ~ . Within the first day of a shift

pco Is a Polycomb-Group Gene 99
which had been deposited in the oocyte to support embryonic development. The same experiment also revealed a requirement for pco+ product during oogenesis itself. After 5-6 days of exposure to restric- tive temperature, homozygous ~ c o ~ ~ ~ ~ ~ ~ ~ mothers stopped laying eggs. This effect is reversible; the fecundity and fertility of such females eventually re- covered when they were returned to permissive tem- perature. So, there are at least two distinct require- ments for pco+ product during oogenesis. The egg chamber requires pco" for its own development and also the oocyte requires pco+ for the embryonic devel- opment of the next generation. The hypothesis of a requirement for pco+ product during oogenesis (and spermatogenesis) itself is supported further by the observations that ovaries of females homozygous for
lethal allele that is also an unconditional female sterile, contain no egg chambers that develop beyond stage 4 (Figure 5); that transplanted ovaries from null mutant larvae do not produce eggs (SHEARN, HERSPERCER and HERSPERCER 1978); and that another tempera- ture-sensitive allele, ~ c o ~ ~ " ) ~ ~ ~ , judged to be more ex- treme than ~ c o ~ ~ ~ ~ ~ ~ ~ or COP^^^^^^^ based on its zygotic phenotype, is both female and male sterile even at permissive temperatures (A. SHEARN, unpublished ob- servations).
The hypomorphic temperature-sensitive alleles of polycombeotic, when exposed to restrictive tempera- tures, can display phenotypes characteristic of the polycomb-group genes. The transformation of the second and third legs to a first leg pattern in adults which occurs when mutant zygotes are exposed during larval development is a zygotic effect (Table 4 and Figure 4). The transformation of the thoracic and abdominal segments toward an eighth abdominal seg- ment pattern in larvae, which is also recognized as characteristic phenotype of mutations in polycomb- group genes, is a maternal-effect (Table 5 and Figure 7). It occurs in homozygous PcOP""~' embryos derived from homozygous temperature-sensitive PCOP"~"
mothers which have been briefly exposed to restrictive temperature. Heterozygous embryos from homozy- gous temperature-sensitive females do not display a strong (if any) cuticle transformation phenotype, but are still unable to survive to adulthood (Figures 6 and 7), indicating that the homeotic larval transformations but not the lethality of the embryos can be paternally rescued. This attenuation, but not elimination, of maternal effects on the embryo by paternally contrib- uted wild-type copies of the gene is similar to that seen in the polycomb-group gene Sce (BREEN and DUNCAN 1986).
The indication that a duplication of pco+ lessens the severity of the Polycomb phenotype is important but must be interpreted with caution, as the 'extra sex comb' phenotype of Polycomb and related mutants is
PCOrnYYPYhS , an extreme temperature-sensitive zygotic
known to be influenced by genetic background and other factors (HANNAH-ALAVA 1964; KENNISON and RUSSELL 1987). However, the relatively small size of the duplication used in our experiments, (based on the cytology of CRAYMER (1984), D p ( 3 ; 3 ) S 2 a 3 com- prises approximately 15 bands), makes any significant effect due to the extra chromosomal material unlikely. The duplications utilized in the study of KENNISON and RUSSELL (1 987), were much larger by comparison, ranging from approximately 50 to 250 bands. Fur- ther, our work with D p ( 3 ; 3 ) S 2 a 3 has shown that there is no significant developmental delay effect caused by this duplication (A. SHEARN, unpublished observa- tions).
The larval and adult homeotic transformations, and the suppression of the phenotype of a Polycomb defi- ciency by a duplication containing the pco gene lead us to place pco within the polycomb-group of genes. ZINK and PARO (1989) identified the 67F region, which contains pco, as one of many binding sites for the Polycomb protein. If the polycomb-group of genes acts as a group of cooperating or interacting proteins or if the group consists of a regulatory network, such binding of one group member's product to the gene of another member might be expected.
Gain of function alleles: The Enhancer of zeste enhancer ( E ( z ) ' ) and suppressor ( S u ( z ) 3 0 I ) mutations are gain-of-function alleles of pco. They both alter the interaction of the zeste and white loci. The interaction of zeste with white, BX-C, and DPP-C, is correlated with the influence on mutant expression by the phys- ical proximity of copies of these loci to each other. Although the exact role of zeste is still unclear, the zeste product has been identified as a transcription factor (BIGGIN et al. 1988, and reviewed by Wu and GOLDBERG 1989). Such a regulatory role, and the role of zeste in the chromosome pairing-dependent trans- vection effect raises the possibility that zeste is involved in or a link between nuclear structure and gene reg- ulation ( JACK and JUDD 1979).
The existence of alleles of polycombeotic that affect the zeste-white interaction raises the possibility that the products of pco and z are involved in similar or related processes. Since these zeste-white interacting alleles are gain-of-function alleles o f p c o , it is difficult to speculate on the nature of the pco gene product based on their phenotypes. However, the interaction of pco alleles with a transcription factor indicates that pco may have a general role in nuclear structure, gene regulation, or both as well. In addition to the dominant alleles of pco discussed here, WU et al. (1 989) identified domi- nant zeste-interacting alleles of two other polycomb- group genes: S u ( z ) 3 0 2 , is an allele of Scm; and Su(z)2', S u ( z ) z 4 , S U ( Z ) ~ ~ , a n d P s c , are either alleles of a gene or an interacting group of genes in the S u ( z ) 2 region.
While the exact functions of any of the polycomb- group genes have yet to be defined, the requirement

100 M. D. Phillips and A. Shearn
of this set of genes for proper gene expression has been clearly established. LEWIS (1978) showed that PC‘ caused expression in thoracic segments of struc- tures normally seen only on the eighth abdominal segment (A8), and that this phenotype was dependent on the state of the bithorax complex genes. DENELL (1973), identified a complex interaction between PC’ and revertants of AntpN5. From these data and the phenotypes of the Pcl, esc, and 1(4)29 (now pho) genes, DUNCAN and LEWIS (1 982) concluded that these genes functioned to regulate the positioning on the body axis of BX-C and ANT-C gene activity. More recent molecular studies correlate with LEWIS’ conclusions. WEDEEN, HARDING and LEVINE (1986) detected an increase in the level of BX-C and Antp transcripts in PC’ mutants. ZINK and PARO (1989) found that anti- bodies to the Polycomb product bind to the sites of ANT-C, BX-C as well as over 50 other sites on salivary gland polytene chromosomes, including many sites of other polycomb-group genes.
While DURA et al. (1987, 1988) concluded that the type and variety of effects of mutant alleles of the polycomb-group gene polyhomeotic are difficult to cor- relate with p h being simply a negative regulator of BX-C and ANT-C, the mutant phenotypes ofph alleles are consistent with a broader role for the p h product in gene regulation. The phenotypes of pco mutants are also not supportive of polycombeotic being exclu- sively a direct regulator of segment-specific homeotic loci and that in general, the range of phenotypes displayed by the large number of polycomb-group genes indicates that this is a class of factors that must have a broad role in development in addition to attenuating the expression of BX-C and ANT-C. While there are sufficient phenotypic similarities caused by mutations in different polycomb-group genes to warrant classifying them as a group, not all polycomb-group genes have identical functions. For example, homozygous extra sex combs null mutants are viable as long as their mother was heterozygous wild type ( e x / + ) (STRUHL 1983), whereas homozygous Po- lycomb and polycombeotic mutants are lethal, regardless of the genotype of the mother.
While we do not expect that the polycomb-group genes are a homogeneous class, with all genes provid- ing identical factors, the similarity of their phenotypes and the mutual enhancement of members’ phenotypes indicate to us that these genes work together in cel- lular or determinative processes, and the insights that we gain from the study of pco may also shed light on the function of other polycomb-group genes.
This work was supported by a grant from the National Institutes of Health. We would like to thank EVELYN HERSPERCER for help in examining mutant ovaries, CHARLES HO for mounting and ex- amining legs for the PC-pco interaction analysis, AMANDA SIMCOX, JOSEPH BIGGS, NICHOLAS TRIPOULAS, TRUDI SCHUPBACH, and two ;Inonynlous reviewers for critical reading of the manuscript and
many IvApful buggestions, KICK JONES for providing many stocks and prepublication results, and CHAO-TING WU for sharing her prepublication manuscript and ideas.
LITERATURE CITED
AKAM, M . E., U. B. ROBERTS, G. P. RICHARDS and M . ASHBURNER, 1978 Drosophila: the genetics of two major larval proteins. Cell 13: 21 5-255 .
BEACHY, 1’. A , , S. L. HELFAND and D. S. HOCNESS, 1985 Segtnental distribution of bithorax complex protein during Dro- sophila development. Nature 313: 545-551.
BIGGIN, M . D., S. BICKEL, M. BENSON, V . PIRROTTA and R. TJIAN, 1988 zeste encodes a sequence-specitic transcription Factor that activates the Ultra-bithorax promoter i n vitro. Cell 53: 7 13- 722.
BREEN, T . R.. and I. M. DUNCAN, 1986 Maternal expression of genes t h a t regulate the bithorax complex of Drosophila melano- gaster. Dev. Biol. 118: 442-456.
CRAYMER, L., 1984 New mutants. Drosophila Inform. Serv. 60: 234-236.
DINELL, K. E., 1973 Homeosis in Drosophila. I. Complementation studies with revertants of Nasobemia. Genetics 75: 279-297.
DENEI.1.. K. E., 1978 Homeosis in Drosophila. 11. A genetic analysis of Polycomb. Genetics 90: 277-289.
DENELL, R. E., K . R. HUMMELS and J . R. GIRI‘ON, I988 Pleiohomeotic: mutations of Drosophila melanogaster causing honleosis and cell lethality. Genetics (in press).
DUNCAN, I . , 1982 Polycomblike: a gene that appears to be required for the n o r n d expression of the bithroax and Antennapedia gene complexes of Drosophila melanogaster. Genetics 102: 49- 70.
DUNCAN, I . , and E. B. LEWIS, I982 Genetic control of body segment differentiation in Drosophila, pp. 533-554 in Deuel- opmental Order: Its Origin and Regulation. edited by S. SUB- TEL.NY. AILIII R. Liss, New York.
DURA, J.-M., H . W. BRocKand P. SANTAMARIA, 1985 polyhomeotic: a gene of Drosophila melanogaster required for correct expres- sion of segmental identity. Mol. Gen. Genet. 198: 213-220.
DURA, J.-M., N. B. KANDSHOLT, J . DEATRICK, 1. ERK, P. SANTA- MARIA, J. D. FREEMAN, S. J. FREEMAN, D. WEDDELL and H. W. BROCK, 1987 A complex genetic locus, polyhomeotic, is re- quired for segmental specification and epidermal development i n I ) . melanogaster. Cell 51: 829-839.
DURA, J.-M., J. DEATRICK, N. B. RANDSHOLT, 1. ERK, H. W. BROCK and P. SANTAMARIA, 1988 Maternal and zygotic requirement for the polyhomeotic complex genetic locus in Drosophila. Roux’s Arch. Dev. Biol. 197: 239-246.
GANS, M . , 1953 Etude ginktique et physiologique du mutant z de Drosophila melanogaster. Bulletin Biol. France Belg. (Suppl.) 38: 1-90.
GArTI, M . , and B. S. BAKER, 1989 Genes controlling essential cell-cycle functions in Drosophila melanogaster. Genes Dev. 3: 438-453.
GATTI, M. S., PIMPINELLI, C. BOVE, B. S . BAKER, D. A. SMITH, A. T. C. CARPENTER and P. RIPOLL, 1983 Genetic control of mitotic cell division in Drosophila melanogaster, pp. 193-204 in Genetirs: New Frontiers. Proceedings of the Fifteenth International Congress of Genetics., Vol. 3 . Oxford and IBH Publishing C o . . New Delhi.
GELRART, W., and C.-T. Wu, 1982 Interactions of zeste mutations with loci exhibiting transvection effects in Drosophila melano- gaster. Genetics 102: 179-189.
GLICKSMAN, M. A , , and D. L. BROWER, 1988 Misregulation of honleotic gene expression i n Drosophila larvae resulting from mutations at the extra sex combs locus. Dev. Bid. 126: 2 19-227.
HANNAH-ALAVA, A,, 1964 Interaction ofnon-allelic loci in expres- sion of the extra-sexconlb phenotype in Drosophila melano-

pco Is a Polycomb-Group Gene 101
gaster. 1. Vererbungslehre. 95: 1-9. HAYNIE, J. I>., 1983 The maternal and zygotic roles of the gene
Polyomb in embryonic determination in Drosophila melano- gastrr. Dev. Biol. 100: 399-41 1.
INGHAM, P., 1984 A gene that regulates the bithorax complex ctifferentially it1 larval and adult cells of Drosophila. Cell 37: 8 15-823.
JACK, J , W., and B. H. JUDD, 1979 Allelic pairing and gene t.egul;ltion: a nlodel for the reste-white interaction in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 76: 1368-1372.
JURGENS, G , , 1985 A group of genes controlling the spatial expres- sion o f ' the bithorax complex in Drosophila. Nature 316: 153- 155.
KALISCH. W.-E., and B. KASMUSON, 1974 Changes in zeste phe- notype induced by autosomal mutations in Drosophila melano- gaster. Hereditas 78: 97- 104.
KAUFMAN, T. C., S. E. TAKASA and D. T. SUZUKI, 1973 The it1ter;lction of two complex loci, zeste and bithorax in Drosophila melanogaster. Genetics 75: 299-32 1.
KENNISON, J. A,, and M. A. RUSSELL, 1987 Dosage dependent modifiers of homoeotic mutations in Drosophila melanogaster. Genetics 116: 75-86.
KENNISON, J. A., and J. W. TAMKUN, 1988 Dosage dependent modifiers of Polycomb and Antennapedia mutations in Drosoph- ila. Proc. Natl. Acad. Sci. USA 85: 8136-8140.
KING, R. C., 1970 Ovarian Development in Drosophila melanogaster.
LEWIS, E. B.. 1955 Some aspects of position pseudoallelism. Am.
LEWIS, E. B., 1978 A gene complex controlling segmentation in Drosophzla. Nature 276: 565-570.
LEWIS, E. B., 1980 New mutants. Drosophila Information Service 55: 205.
LINDSLEY, D. L., and E. H. GRELL, 1968 Genetic Variations of Drosophila melanogaster. Carnegie Inst. Wash. Publ. 627.
LINDSLEY, I)., and G. ZIMM, 1985 The Genome of Drosophila mrlanogaster (progress report). Part 1 : genes A-K. Drosophila Inform. Serv. 62.
LINDSLEY, D., and G. ZIMM, 1986 The Genome of Drosophila melanogaster (progress report). Part 2: lethals; cytogenetic map. Drosophila Inform. Serv. 64.
LINDSLEY, D., and G. ZIMM, 1987 The Genome of Drosophila melanogaster (progress report). Part 3: rearrangements. Dro- sophila Inform. Serv. 65.
MULLER, H. J., 1983 Further studies on the nature and causes of gene mutations, pp. 21 3-255 in Proceedings of the Sixth Inter- national Congress ofGenetics, Vol. 1. edited by D. F. JONES.
NUSSLEIN-VOLHARD, C., E. WIESCHAUS and H. KLUDING, 1984 Mutations affecting the pattern of the larval cuticle in Drosophila melanogaster. I . Zygotic loci on the second chromo- some. Koux's Arch. Dev. Biol. 193: 267-282.
SCHOTT, D. R., M. C. BALDWIN and V. FINNERTY, 1986 Molybdenum hydroxylases in Drosophila. 111. Further charac- teriz~tion of the low xanthine dehydrogenase gene. Biochem.
Academic Press, New York.
Natl. 89: 73-89.
Getlet. 24: 509-527. SHEARN, A , , 1977 Mutational dissection of imaginal disc devel-
opment i n Drosophila melanogaster. Am. Zool. 17: 585-594. SHEARN, A. , 1980 What is the normal function of genes which
give rise to homeotic mutations? pp. 155-162 in Development and Neurobiology of Drosophila, edited by 0. SIDDIQI, P. BABU, I,. M . HALL, and J. C . HALL. Plenum Publishing, New York.
SHEARN, A , , and A. GAREN, 1974 Genetic control of imaginal disc dwelopnlent in Drosophila. Proc. Natl. Acad. Sci. USA 71: 1393- 1397.
SHEARN, >4., G. HERSPERGER and E. HERSPERGER, 1978 Genetic ;In;llysis of two allelic, temperature-sensitive mutants of Dro- sophila melanogaster, both of which are zygotic and maternal- effect lethals. Genetics 89: 341-353.
SHEARN, A , , T. RICE, A. GAREN and W. GEHRING, 1971 Imaginal disc abt1ornralities in lethal mutants of Drosophila. Proc. Natl. Acad. Sci. USA 68: 2594-2598.
SHEARN, A, , G. HERSPERGER, E. HERSPERGER, E. S. PENTZ and P. DENKER, 1978 Multiple allele approach to the study of genes i n Drosophila melanogaster that are involved in imaginal disc development. Genetics 89: 355-370.
STRUHL, G., 1981 A gene product required for correct initiation of segtnental determination in Drosophila. Nature 293: 36-4 1.
STRUHI., G., 1983 Role of the est+ gene product in ensuring the selective expression of segment-specific homeotic genes in Dro- sophila. J. Enlbryol. Expt. Morphol. 7 6 297-331.
STRUHL, G . . and M. AKAM, 1985 Altered distributions of Ultra- bithorax transcripts in extra sex combs mutant embryos of Dro-
STRC'HL, G., and R. A. H. WHITE, 1985 Regulation of the Ultra- bithorax gene of Drosophila by other bithorax complex genes. Cell 43: 507-519.
VAN DER MEER, J., 1977 Optically clean and permanent whole tnount preparations for phase contrast microscopy of cuticular structures of insect larvae. Drosophila Inform. Serv. 52: 160.
WEDEEN, C., I(. HARDING and M. LEVINE, 1986 Regulation of Antennapedia and bithorax gene expression by the Polycomb locus in Drosophila. Cell 44: 739-748.
WIESCHAUS, E., and C. NUSSLEIN-VOLHARD, 1986 Looking at embryos, pp. 199-227 in Drosophila: A Practical Approach, edited by D. B. ROBERTS. IRL Press, Oxford.
WIESCHAUS, E., C. NUSSLEIN-VOLHARD and G. JURGENS, 1984 Mutations affecting the pattern of the larval cuticle in Drosoph- ila melanogaster. 111. Zygotic loci on the X-chromosome and fourth chromosome. Roux's Arch. Dev. Biol. 193: 296-307.
Wu, C.-T., and M. L. GOLDBERG, 1989 The Drosophila zeste gene ;tnd transvection. Trends Genet. 5: 189-194.
WU, C.-T., R. S. JONES, P. F. LASKO and W. M. GELBART, 1989 Homeosis ;md the interaction of reste and white in Dro- sophila. Mol. Gen. Genet. 218: 559-564.
ZINK, B., and R. PARO, 1989 I n vivo binding of a trans-regulator of homoeotic genes in Drosophila melanogaster. Nature 337: 468-47 1.
Communicating editor: T. SCHUPBACH
sophila. EMBO J. 4: 3259-3264.