multiple representations of body movements in mesial area 6 and the adjacent cingulate cortex: an...
TRANSCRIPT

THE JOURNAL OF COMPARATIVE NEUROLOGY 311:463-482 (1991)
Multiple Representations of Body Movements in Mesial Area 6 and the
Adjacent Cingulate Cortex: An Intracortical Microstimulation Study in
the Macaque Monkey
G. LUPPINO, M. MATELLI, R.M. CAMARDA, V. GALLESE, AND G. RIZZOLATTI Istituto di Fisiologia Umana, Universita di Parma, 43100 Parma, Italy
ABSTRACT The mesial agranular frontal cortex that lies rostral to area 4 (Fl) is formed by two distinct
cytoarchitectonic areas: F3, located caudally, and F6, located rostrally. In the present experiments we investigated the organization of F3 and F6 by observing the motor responses evoked by their intracortical electrical microstimulation. Our main purpose was to find out whether the cytoarchitectonic subdivision of the mesial agranular frontal cortex into two areas has a physiological counterpart.
The result showed that F3 (the caudal area) contains a complete motor representation with hindlimb movements located caudally, forelimb movements located centrally, and orofacial movements located rostrally. The great majority of limb movements involved proximal joints. With respect to F1, F3 showed the following functional characteristics: (1) lack of segregation between proximal and distal movements, (2) larger percentage of complex movements, and (3) higher excitability threshold.
Movements were more difficult to elicit from F6 (the rostral area) than from F3. However, by using a longer stimulus train duration (100 ms) 39.3% of tested sites produced body movements. This percentage increased (50.5%) when the electrical stimulation was applied during monkey natural movements instead of when the monkey was still in its chair. Most of the evoked movements concerned the forelimb. More rarely, neck and upper face movements were observed. Unlike F1 and F3 where most movements were fast, slow movements were frequently observed with stimulation of F6. Many of them mimicked natural movements of the animal. Eye movements were evoked from F7 (superior area 6) but not from F6.
An additional motor representation was found in the dorsocaudal part of area 24 (24d). This area is topographically organized with a forelimb representation located caudally and ventrally and a hindlimb representation located rostrally and dorsally. The excitability threshold of area 24d is higher than that of F1 and F3. Evoked movements were occasionally observed also after stimulation of area 24c.
In conclusion, on the mesial cortical wall rostral to F1, there are at least three independent motor representations. On the basis of somatotopic organization and excitability properties, we propose that the term supplementary motor area (SMA-proper) should be reserved to F3.
Key words: premotor areas, supplementary motor area, area 24, motor representation, primates
Since the classical studies of Penfield and Welch ('51) and Woolsey and coworkers ('52), it is well established that mesial area 6 contains a motor representation independent of that of the precentral cortex. According to Woolsey et al. ('521, this second motor representation, usually referred to as the supplementary motor area (SMA), is somatotopically organized and, in the monkey, occupies the whole mesial
Although Woolsey's picture of the SMA organization has been questioned in recent years (e.g., Orgogozo and Larsen, '79; Eccles, '82; Macpherson et al., '82a,b; Wiesendanger, '86), the notion that the SMA is a single functional area
Accepted May 31, 1991, aspect Of area 6, rostrally extending anterior end of the superior arcuate sulcus.
the level Of the Address reprint requests to G. Rizzolatti, Istituto di Fisiologia Umana, Universita di Parma, via Gramsci 14,43100 Parma, Italy.
O 1991 WILEY-LISS, INC.

464 G. LUPPINO ET AL.
basically co-extensive with mesial area 6 was generally accepted (see Wiesendanger, ’86). The results, however, of recent studies in which the SMA functional organization was investigated with intracortical electrical microstimula- tion appear to contradict this notion.
In a series of experiments in which the SMA was stimu- lated with short trains of pulses, Wiesendanger and his coworkers (Macpherson et al., ’82a,b; Hummelsheim et al., ’86) found that only the caudal part of the SMA gave motor responses. No movements were obtained with stimulation of the SMA rostral part, where, according to the Woolsey’s maps, oro-facial movements are represented. However, since oro-facial movements were not evoked at all, the possibility remained that the rostral SMA indeed contained an oro-facial representation, but that, for some reasons, the stimulation procedure employed failed to demonstrate it.
By using a modified stimulation technique and a longer stimulus train duration, Mitz and Wise (’87) obtained a picture of the SMA somatotopy more detailed than that of Wiesendanger (Macpherson et al., ’82a,b; Hummelsheim et al., ’86). According to their findings, the SMA contains a hindlimb, a forelimb and an oro-facial representation, and these representations are located in a caudo-rostral progres- sion. The oro-facial representation, however, is small and lies few millimeters rostral to a line tangent to the border of the postarcuate sulcus. The rostralmost part of mesial area 6 was found virtually unexcitable.
Prompted by these observations and by our findings (Matelli et al., ’91) that the mesial area 6 is cytoarchitecton- ically formed by two distinct areas (F3 and F6), we decided to reinvestigate the problem of the motor representation in mesial area 6 by using intracortical electrical microstimula- tion and referring the stimulation results to the new cytoarchitectonic maps. Our working hypothesis was that the posterior cytoarchitectonic area (F3) contains a com- plete somatotopically organized motor representation and that the anterior cytoarchitectonic area (F6) is a function- ally independent area, possibly with properties more com- plex than the former (see Rizzolatti, ’89). The experiments fully confirmed this hypothesis. They showed that in F3 there is a complete motor representation with a somato- topic organization analogous to that described by Mitz and Wise (’87) and that in F6 there is a separate motor representation essentially related to arm movements. Addi- tional motor representations were found in area 24d and 24c. In the former area, leg and arm movements are represented, whereas in the latter, mainly arm movements were observed. The multiplicity of motor representations in mesial area 6 appears to be an important cue for clarifying the controversial issue of somatotopic organization of this region as well as that of the role of the so called “SMA” in “motor” and “supramotor” functions (Eccles, ’82; see Wiesendanger, ’86). Some of the results of the present study have been reported previously in a preliminary account (Matelli et al., ’90).
METHODS Three monkeys, one Macaca nemestrina (MK5) and two
Macaca fascicularis (MK6 and MK7), were used in the experiments. Extensive microstimulation of the mesial motor areas was carried out in three hemispheres, one for
each animal (MK5, MK6, and MK7). Procedures that facilitate the occurrence of evoked movements in F6 (see below) were employed in MK7. In this animal the func- tional organization of F6 was studied in detail in both hemispheres.
Animal preparation and stimulation procedure All monkeys were trained to sit quietly in the primate
chair and to be touched by the experimenters. After 2-3 weeks of training, the animals were ready for the experi- ments. A large craniotomy over the posterior part of the frontal lobe was then performed under general anesthesia (Ketamine hydrochloride, 15 mgkg i.m., supplemented every 30 min) and strict aseptic conditions. A chamber was positioned over the opening and cemented to the skull. In addition, a device that allowed a rigid painless fixation of the head was implanted. For details on the recording- stimulation set up, see Gentilucci et al. (’88).
The experiments were carried out 5 days a week for about 7 hours each day. In order to render the restrained condi- tion pleasant and to keep the animal relaxed, food and fruit juice were given to the animal throughout the experiments. In the rare cases in which the animal showed nervousness, the experiment was stopped. Relaxed mood and posture were found to be a necessary prerequisite to obtain con- stant, reliable responses. When the animal was stiff or restless, responses were difficult to elicit and to detect especially in areas rostral to F1.
Penetrations, perpendicular to the cortical surface, were made using stereotaxic coordinates. Penetrations were spaced at 1,000-pm intervals in the rostrocaudal direction. In the three hemispheres more extensively explored, they formed long rows roughly parallel to the interhemispheric fissure extending from F1 to the rostral end of F6. In addition, penetrations outside the rows were performed in all monkeys in order to clarify andor to confirm data obtained during the initial systematic exploration of the cortical areas.
Microstimulation was performed using tungsten micro- electrodes (impedance 0.5-1.5 MSZ, measured at 1 Khz frequency). The same electrodes were also used for multi- spike and single neuron recordings. In each penetration the first stimulation was carried out at the site at which the first action potentials (multispike activity) were recorded. The subsequent stimulations were made every 500 km until the penetration end. Occasionally, stimulations were carried out every 250 pm. Histological reconstructions (see below) showed that most penetrations crossed the gray matter in the deep part of layers 111, V, and VI. This was true for F1, F3, or F6. Thus all areas were stimulated at similar depths.
The applied stimuli were trains of cathodal pulses generated by a constant current stimulator. The following parameters were used: train duration = 50 ms, pulse duration = 0.2 ms, frequency = 330 Hz, current intensity 3 to 40 +A. The current strength was controlled on an oscilloscope measuring the voltage drop across a 10 KSZ resistor in series with the stimulating electrode. All micro- electrodes were examined microscopically before and after use.
The standard stimulation procedure consisted of an initial stimulation with a current of 40 FA, followed by successive stimulations with progressively decreasing cur-

MOTOR MAPS OF MESIAL AREA 6 AND AREA 24 465
a
C A S P A
i i
Fig. 1. Reconstruction procedure of the mesial cortical surface. (a) Dorsal view of a monkey brain; arrows and dotted lines indicate the level of the genu of the arcuate sulcus (PA) and the rostral end of the superior arcuate sulcus (AS); C = central sulcus. (b) Frontal section in
rent intensities. Two observers scanned the entire animal for evoked movements. Movements evoked with intracorti- cal stimulation were recorded only if they were clearly identified by two observers, observers agreed on the type of movements, and movements were evoked repeatedly from the same site. Once movements at a given site were identified, the threshold for each movement was deter- mined. Movement threshold was defined as the current at which the movement was evoked on 50% of the trials. Single neuron recording before and after microstimulation showed that currents used in the present experiments did not cause any appreciable modification of neuronal activity (for similar observations, see Donoghue and Wise, '82).
The standard stimulation procedure described above was routinely used to map the mesial motor areas. However, when no movement was evoked with this procedure two variations were introduced. First, the stimulus train dura- tion was increased to 100 ms (see Mitz and Wise, '87). Second, if this procedure was ineffective, movements were elicited with natural stimuli (reaching for food, pushing away annoying stimuli) and the effect of 50 and 100 ms duration currents was assessed during the execution of these movements (activation procedure). Current intensi- ties, pulse duration, and repetition rate of individual pulses remained the same as during standard stimulation.
b
Dorsal Surface
Mesial Surface
Dorsal B a n k of Cingulate Sulcus
Ventral Bank of Cingulate Sulcus
Cingulate G y r u s
which the dotted lines separate the cortical surfaces reported in the unfolded reconstruction shown in c. (c) Unfolded view of the mesial surface of the hemisphere; caudal is on the right,
Histology and reconstruction of stimulation sites
About 1 week before sacrificing the animal, a series of electrolytic lesions (10 p A cathodal current for 10 s) equally spaced one from another were made at the lateral border of the studied area. The lesions formed a row parallel to the rows of experimental penetrations. In addition, lesions were made in the depth of the mesial cortical surface in F1 in order to have a marker for reconstructing the penetra- tion depth.
The animals were sacrificed with a lethal dose of sodium thiopental and perfused through the heart with a buffered saline followed by fixative. The animal was then placed in the stereotaxic apparatus, the dura was removed, and the stereotaxic coordinates of the arcuate and central sulci were assessed. The brain was cut coronally on a stereotaxic frame. It was then removed, photographed, and sectioned on a freezing microtome at 60 pm coronally. The sections were stained with thionin and processed for cytochrome oxidase histochemistry (Wong-Riley, '79; see also Matelli et al., '85). The reconstructed trajectory of each penetration was plotted on drawings of sections taken at 120-km intervals. The reconstruction was based on the recognition of the penetration traces, recording coordinates, recording
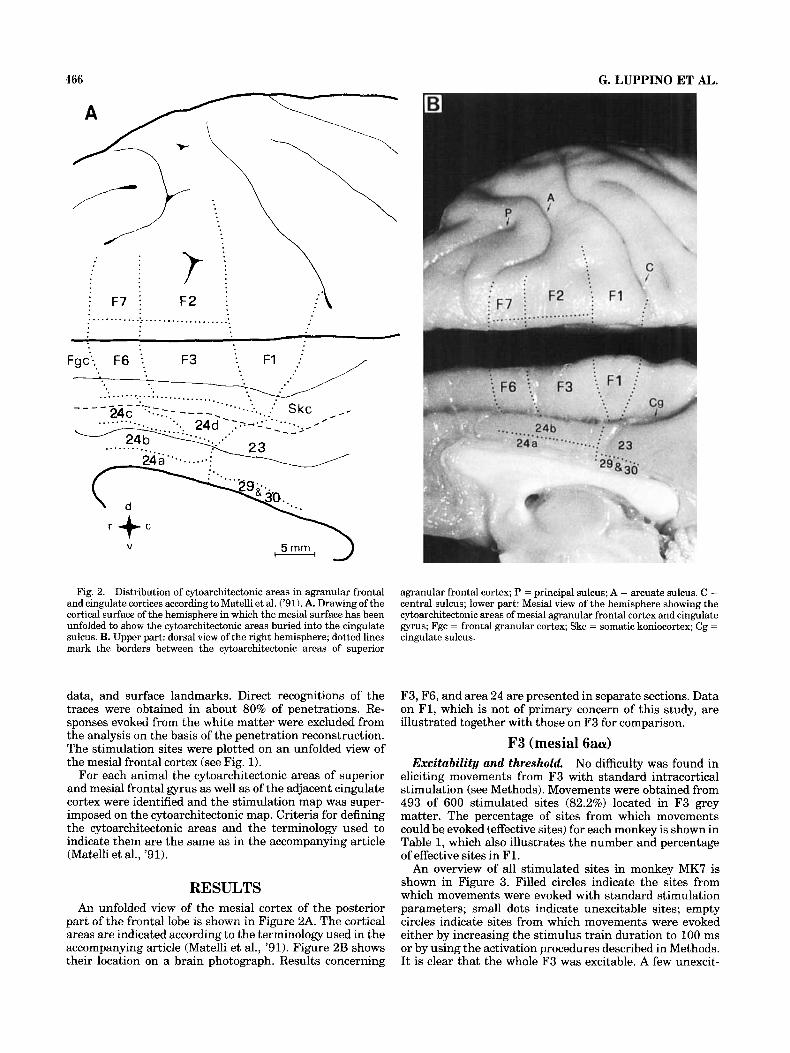
466 G. LUPPINO ET AL.
.................................. I /
FgG: F6' . . F3
...........................
V 5 rnrn
Fig. 2. Distribution of cytoarchitectonic areas in agranular frontal and cingulate cortices according to Matelli et al. ('91). A. Drawing of the cortical surface of the hemisphere in which the mesial surface has been unfolded to show the cytoarchitectonic areas buried into the cingulate sulcus. B. Upper part: dorsal view of the right hemisphere; dotted lines mark the borders between the cytoarchitectonic areas of superior
data, and surface landmarks. Direct recognitions of the traces were obtained in about 80% of penetrations. Re- sponses evoked from the white matter were excluded from the analysis on the basis of the penetration reconstruction. The stimulation sites were plotted on an unfolded view of the mesial frontal cortex (see Fig. 1).
For each animal the cytoarchitectonic areas of superior and mesial frontal gyrus as well as of the adjacent cingulate cortex were identified and the stimulation map was super- imposed on the cytoarchitectonic map. Criteria for defining the cytoarchitectonic areas and the terminology used to indicate them are the same as in the accompanying article (Matelli et al., '91).
RESULTS An unfolded view of the mesial cortex of the posterior
part of the frontal lobe is shown in Figure 2A. The cortical areas are indicated according to the terminology used in the accompanying article (Matelli et al., '91). Figure 2B shows their location on a brain photograph. Results concerning
agranular frontal cortex; P = principal sulcus; A = arcuate sulcus. C = central sulcus; lower part: Mesial view of the hemisphere showing the cytoarchitectonic areas of mesial agranular frontal cortex and cingulate gyrus; Fgc = frontal granular cortex; Skc = somatic koniocortex; Cg = cingulate sulcus.
F3, F6, and area 24 are presented in separate sections. Data on F1, which is not of primary concern of this study, are illustrated together with those on F3 for comparison.
F3 (mesial 6aa) Excitability and threshold. No difficulty was found in
eliciting movements from F3 with standard intracortical stimulation (see Methods). Movements were obtained from 493 of 600 stimulated sites (82.2%) located in F3 grey matter. The percentage of sites from which movements could be evoked (effective sites) for each monkey is shown in Table 1, which also illustrates the number and percentage of effective sites in F1.
An overview of all stimulated sites in monkey MK7 is shown in Figure 3. Filled circles indicate the sites from which movements were evoked with standard stimulation parameters; small dots indicate unexcitable sites; empty circles indicate sites from which movements were evoked either by increasing the stimulus train duration to 100 ms or by using the activation procedures described in Methods. I t is clear that the whole F3 was excitable. A few unexcit-
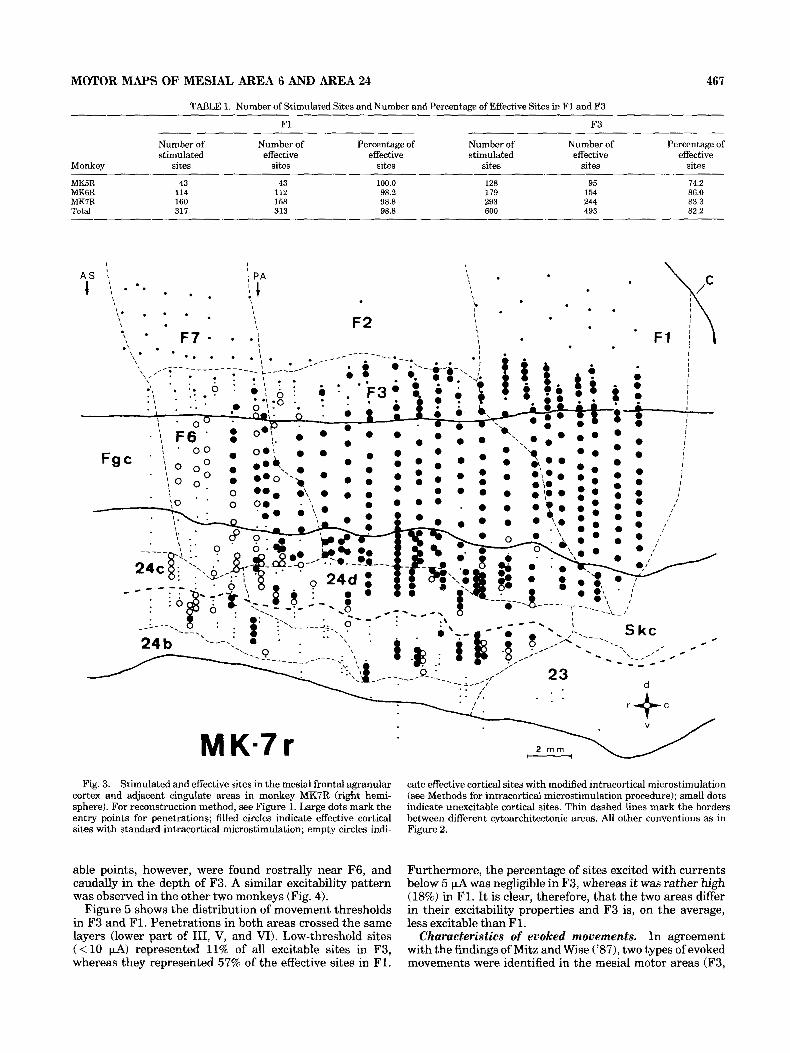
MOTOR MAPS OF MESIAL AREA 6 AND AREA 24 467
TABLE 1. Number of Stimulated Sites and Number and Percentage of Effective Sites in F1 and F3
P1 F3 _ - Number of Number of Percentage of Number of Number of Percentage of stimulated effective effective stimulated effective effective
Monkey sites sites sites sites sites sites
43 43 100.0 128 95 74.2 86.0
MK5R MKGR 114 112 98.2 179 154
160 158 98.8 293 244 83.3 MK7R 317 313 98.8 600 493 82.2 Total
---I- 2 rnrn M K.7 r Fig. 3. Stimulated and effective sites in the mesial frontal agranular
cortex and adjacent cingulate areas in monkey MK7R (right hemi- sphere). For reconstruction method, see Figure 1. Large dots mark the entry points for penetrations; filled circles indicate effective cortical sites with standard intracortical microstimulation; empty circles indi-
able points, however, were found rostrally near F6, and caudally in the depth of F3. A similar excitability pattern was observed in the other two monkeys (Fig. 4).
Figure 5 shows the distribution of movement thresholds in F3 and F1. Penetrations in both areas crossed the same layers (lower part of 111, V, and VI). Low-threshold sites ( < l o FA) represented 11% of all excitable sites in F3, whereas they represented 57% of the effective sites in F1.
cate effective cortical sites with modified intracortical microstimulation (see Methods for intracortical microstimulation procedure); small dots indicate unexcitable cortical sites. Thin dashed lines mark the borders between different cytoarchitectonic areas. All other conventions as in Figure 2.
Furthermore, the percentage of sites excited with currents below 5 pA was negligible in F3, whereas it was rather high (18%) in F1. It is clear, therefore, that the two areas differ in their excitability properties and F3 is, on the average, less excitable than F1.
Characteristics of evoked movements. In agreement with the findings of Mitz and Wise ('87), two types of evoked movements were identified in the mesial motor areas (F3,

468 G. LUPPINO ET AL.
MK.5 r 2 m m -
, 23 : -4 b . .
MK.6 r Fig. 4. Stimulated and effective sites in the mesial frontal agranular cortex and adjacent cingulate areas
in monkeys MK5R and MKGR (right hemispheres). All conventions 8s in Figure 3.
MOTOR MAPS OF MESIAL AREA 6 AND AREA 24
MK-5r MK-Gr
F3 20.
10-
0 -
469
% 40.
Area 30.
2 4 d 20.
M K.7r Total
Area % 30 L11 N 95 1 N =154 N = 244
N=16
i N=34
l 0 1 0 , I I , ! I 7 1 1
0 5 10 15 2 0 2 5 3 0 35 40 0 5 1015 2025 3035 40 PA PA
N = 4 8 1 N = 98
0 5 10 15 20 25 30 35 40 0 5 10 15 20 25 30 35 40
P A P A
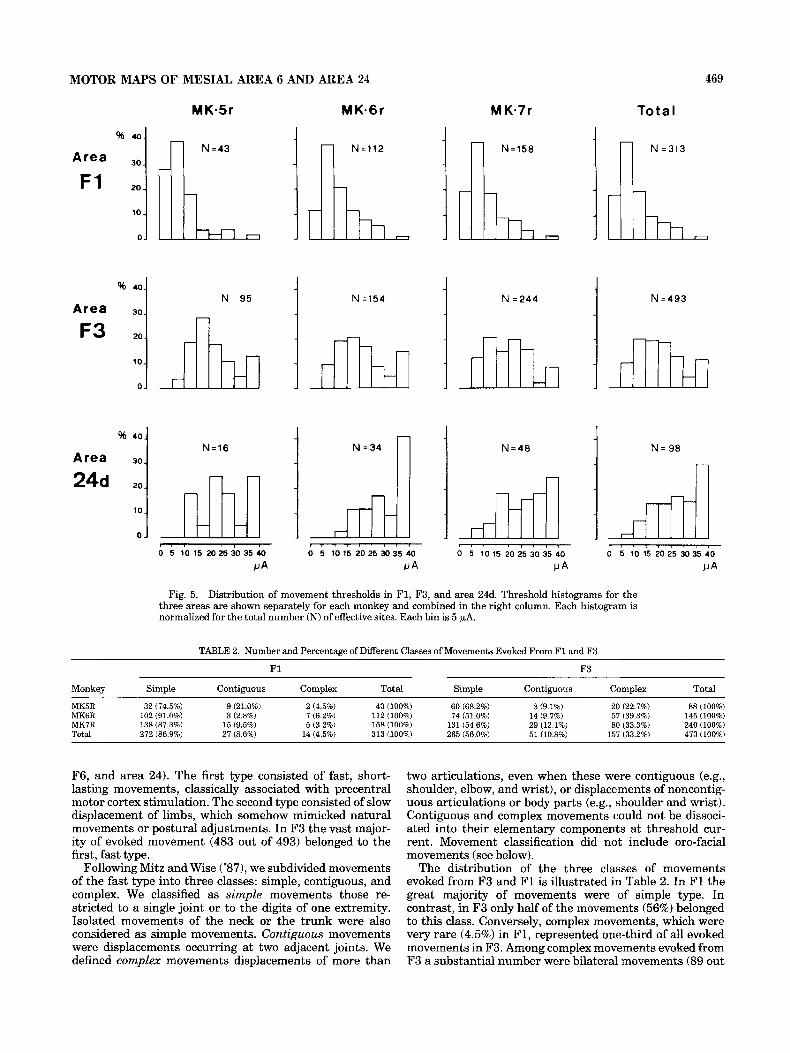
Fig. 5. Distribution of movement thresholds in F1, F3, and area 24d. Threshold histograms for the three areas are shown separately for each monkey and combined in the right column. Each histogram is normalized for the total number (N) of effective sites. Each bin is 5 wA.
TABLE 2. Number and Percentage of Different Classes of Movements Evoked From F1 and F3
F1 F3
Monkey Simple Contiguous Complex Total Simple Contiguous Complex Total
MK5R 32 (74.5%) 9 (21.0%) 2 (4.5%) 43 (100%) 60 (68.2%) 8 (9.1%) 20 (22.7%) 88 (100%) MK6R 102 (91.0%) 3 (2.8%) 7 (6.2%) 112 (100%) 74 (51.0%) 14 (9.7%) 57 (39.3%) 145 (100%) MK7R 138 (87.3%) 15 (9.5%) 5 (3.2%) 158 (100%) 131 (54.6%) 29 (12.1%) 80 (33.3%) 240 (100%) Total 272 (86.9%) 27 (8.6%) 14 (4.5%) 313 (100%) 265 (56.0%) 51 (10.8%) 157 (33.2%) 473 (100%)
F6, and area 24). The first type consisted of fast, short- lasting movements, classically associated with precentral motor cortex stimulation. The second type consisted of slow displacement of limbs, which somehow mimicked natural movements or postural adjustments. In F3 the vast major- ity of evoked movement (483 out of 493) belonged to the first, fast type.
Following Mitz and Wise ('87), we subdivided movements of the fast type into three classes: simple, contiguous, and complex. We classified as simple movements those re- stricted to a single joint or to the digits of one extremity. Isolated movements of the neck or the trunk were also considered as simple movements. Contiguous movements were displacements occurring at two adjacent joints. We defined complex movements displacements of more than
two articulations, even when these were contiguous (e.g., shoulder, elbow, and wrist), or displacements of noncontig- uous articulations or body parts (e.g., shoulder and wrist). Contiguous and complex movements could not be dissoci- ated into their elementary components at threshold cur- rent. Movement classification did not include oro-facial movements (see below).
The distribution of the three classes of movements evoked from F3 and F1 is illustrated in Table 2. In F1 the great majority of movements were of simple type. In contrast, in F3 only half of the movements (56%) belonged to this class. Conversely, complex movements, which were very rare (4.5%) in F1, represented one-third of all evoked movements in F3. Among complex movements evoked from F3 a substantial number were bilateral movements (89 out
470 G. LUPPINO ET AL.
% 40
30
20
10
0
Area F3
N =I57 (493 1
;hn I I I I I I I I 1
0 5 10 15 20 25 30 35 40 JJA
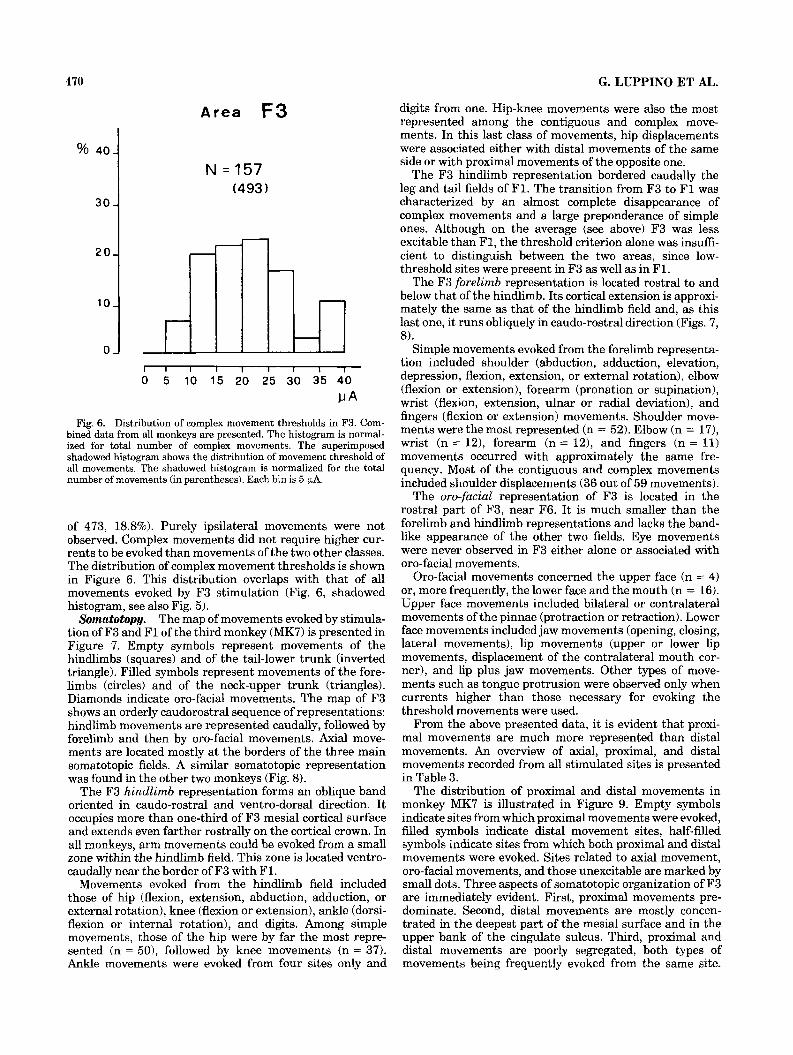
Fig. 6. Distribution of complex movement thresholds in F3. Com- bined data from all monkeys are presented. The histogram is normal- ized for total number of complex movements. The superimposed shadowed histogram shows the distribution of movement threshold of all movements. The shadowed histogram is normalized for the total number of movements (in parentheses). Each bin is 5 PA.
of 473, 18.8%). Purely ipsilateral movements were not observed. Complex movements did not require higher cur- rents to be evoked than movements of the two other classes. The distribution of complex movement thresholds is shown in Figure 6. This distribution overlaps with that of all movements evoked by F3 stimulation (Fig. 6, shadowed histogram, see also Fig. 5).
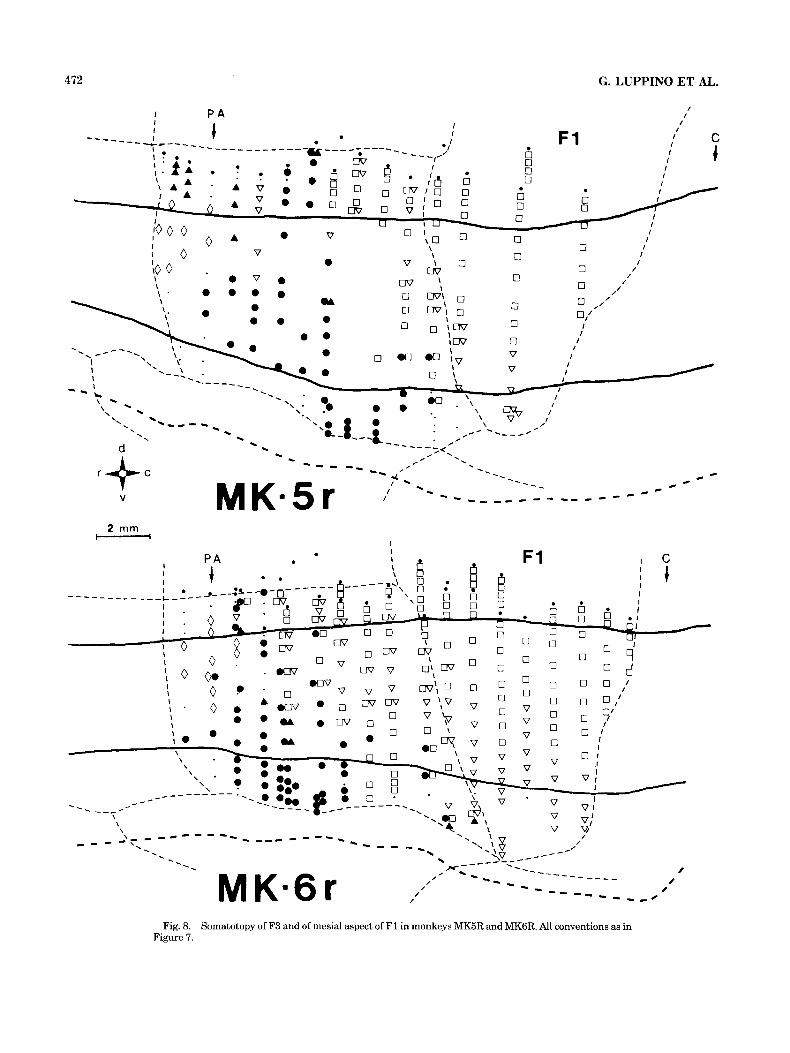
SomatotopH. The map of movements evoked by stimula- tion of F3 and F1 of the third monkey (MK7) is presented in Figure 7. Empty symbols represent movements of the hindlimbs (squares) and of the tail-lower trunk (inverted triangle). Filled symbols represent movements of the fore- limbs (circles) and of the neck-upper trunk (triangles). Diamonds indicate oro-facial movements. The map of F3 shows an orderly caudorostral sequence of representations: hindlimb movements are represented caudally, followed by forelimb and then by oro-facial movements. Axial move- ments are located mostly at the borders of the three main somatotopic fields. A similar somatotopic representation was found in the other two monkeys (Fig. 8).
The F3 hindlimb representation forms an oblique band oriented in caudo-rostral and ventro-dorsal direction. I t occupies more than one-third of F3 mesial cortical surface and extends even farther rostrally on the cortical crown. In all monkeys, arm movements could be evoked from a small zone within the hindlimb field. This zone is located ventro- caudally near the border of F3 with Fl.
Movements evoked from the hindlimb field included those of hip (flexion, extension, abduction, adduction, or external rotation), knee (flexion or extension), ankle (dorsi- flexion or internal rotation), and digits. Among simple movements, those of the hip were by far the most repre- sented (n = 501, followed by knee movements (n = 37). Ankle movements were evoked from four sites only and
digits from one. Hip-knee movements were also the most represented among the contiguous and complex move- ments. In this last class of movements, hip displacements were associated either with distal movements of the same side or with proximal movements of the opposite one.
The F3 hindlimb representation bordered caudally the leg and tail fields of F1. The transition from F3 to F1 was characterized by an almost complete disappearance of complex movements and a large preponderance of simple ones. Although on the average (see above) F3 was less excitable than F1, the threshold criterion alone was insuffi- cient to distinguish between the two areas, since low- threshold sites were present in F3 as well as in F1.
The F3 forelimb representation is located rostral to and below that of the hindlimb. Its cortical extension is approxi- mately the same as that of the hindlimb field and, as this last one, it runs obliquely in caudo-rostra1 direction (Figs. 7, 8).
Simple movements evoked from the forelimb representa- tion included shoulder (abduction, adduction, elevation, depression, flexion, extension, or external rotation), elbow (flexion or extension), forearm (pronation or supination), wrist (flexion, extension, ulnar or radial deviation), and fingers (flexion or extension) movements. Shoulder move- ments were the most represented (n = 52). Elbow (n = 17), wrist (n = 121, forearm (n = 12), and fingers (n = 11) movements occurred with approximately the same fre- quency. Most of the contiguous and complex movements included shoulder displacements (36 out of 59 movements).
The oro-facial representation of F3 is located in the rostral part of F3, near F6. It is much smaller than the forelimb and hindlimb representations and lacks the band- like appearance of the other two fields. Eye movements were never observed in F3 either alone or associated with oro-facial movements.
Oro-facial movements concerned the upper face (n = 4) or, more frequently, the lower face and the mouth (n = 16). Upper face movements included bilateral or contralateral movements of the pinnae (protraction or retraction). Lower face movements included jaw movements (opening, closing, lateral movements), lip movements (upper or lower lip movements, displacement of the contralateral mouth cor- ner), and lip plus jaw movements. Other types of move- ments such as tongue protrusion were observed only when currents higher than those necessary for evoking the threshold movements were used.
From the above presented data, it is evident that proxi- mal movements are much more represented than distal movements. An overview of axial, proximal, and distal movements recorded from all stimulated sites is presented in Table 3.
The distribution of proximal and distal movements in monkey MK7 is illustrated in Figure 9. Empty symbols indicate sites from which proximal movements were evoked, filled symbols indicate distal movement sites, half-filled symbols indicate sites from which both proximal and distal movements were evoked. Sites related to axial movement, oro-facial movements, and those unexcitable are marked by small dots. Three aspects of somatotopic organization of F3 are immediately evident. First, proximal movements pre- dominate. Second, distal movements are mostly concen- trated in the deepest part of the mesial surface and in the upper bank of the cingulate sulcus. Third, proximal and distal movements are poorly segregated, both types of movements being frequently evoked from the same site.

Fig. 7. Somatotopy of '$3 and of mesial aspect of F1 in monkey MK7R. Movements symbols: diamonds, oro-facial; filled triangles, neck-upper trunk; filled circles, forelimb; empty squares, hindlimb; empty triangles, lower trunk-tail. Other conventions as in Figure 3.
Note the markedly different organization of the adjacent hindlimb field of F1. In this area both distal and proximal movements are richly represented with a minimal overlap between the two representations. Distal movements are represented mostly on the cortical crown, proximal move- ments (including those of the tail) occupy the mesial frontal cortex and a small sector of the upper bank of the cingulate sulcus.
Monkey MK6 showed an organization of distal-proximal movements identical to that of monkey MK7, whereas a more marked segregation of proximal from distal move- ments was observed in F3 of monkey MK5 (Fig. 10). This difference may be due to individual variations, to a sample bias, or to a species difference since, in contrast to MK6 and MK7, which were Java monkeys, MK5 was a pig-tail monkey.
F6 (mesial 6ap) Excitability and threshold. Experiments carried out on
the first two monkeys showed that, occasionally, body movements could be obtained from sites rostral to F3 mouth field. Histological reconstruction of the electrode traces demonstrated that these sites were located in F6. A detailed study of the excitability o f this area (not limited to its caudal end) was then carried out in the third monkey (MK7). In studying this animal, in addition to the standard
stimulation, long stimulus train durations (100 ms) and activation procedures (see Methods) were employed. Both hemispheres were explored and, in total, 214 sites were stimulated. Of them, 50 (23.4%) gave motor responses with standard stimulation. When longer stimulus train duration and/or activation procedures were employed, the number of the effective sites increased. The results are shown in Table 4.
The distribution of movement thresholds in F6 is pre- sented in Table 5. The movement threshold was much higher in F6 than in F1 and F3. No site could be excited with currents below 10 PA. The location of excitable sites of MK7 right hemisphere is shown in Figure 3. Similar results were obtained for the left hemisphere.
Somatotopy and characteristics of evoked movements. The map of movements evoked by stimulation of F6 is shown in Figure 11. For the construction of this map, movements evoked with standard stimulation, 100 ms stimulus train duration, and activation procedures were used. Arm movements (filled circles) were by far the most represented. They were evoked from most of the mesial cortical surface and from the upper bank of the cingulate sulcus. Exception to this were two sites (empty square) near area 24 whose stimulation produced leg movements. Neck and upper face movements (generally movements of the ears) were found on the cortical crown and in the upper-
472
I P A
G. LUPPINO ET
I I
AL.
C t
1
I
- -- -------- \ \ \
# # # -- _ _ _ . A
_ - - - - '. -I---
- _-- - - - - - \ - - - - - - - - 0 -----_ _ _ _ _
Fig. 8. Somatotopy of F3 and of mesial aspect of F1 in monkeys MK5R and MKGR. All conventions as in Figure 7.

MOTOR MAPS OF MESIAL AREA 6 AND AREA 24 473
TABLE 3. Number and Percentage of Axial, Proximal, and Distal Movements Evoked From F3 in All Monkeys
Number of movements
Axial Proximal Distal Ax-Prox Prox-Dist Ax-Prox-Dist Total
Forelimb 102 33 8 26 169 (35.7%) Hindlimb 134 7 48 19 6 214 (45.2%) Forelimb +
hindlimb 11 7 6 24 (5.1%) Axial 66 66 (14.0%) Total 66 (14.0%) 247 (52.2%) 40 (8.5%) 63 (13.3%) 51 (10.8%) 6 (1.2%) 473 (100%)
I I P A I F1 C
Fig. 9. Distribution of proximal, distal and tail movements in F3 and in the mesial aspect of F1 in monkey MK7R. Movement symbols: filled circles, distal forelimb (fingers, wrist); empty circles, proximal forelimb (forearm, elbow, shoulder); filled squares, distal hindlimb
(toes, ankle); empty squares, proximal hindlimb (knee, hip); empty triangles, tail; half-filled symbols, coexistence of distal and proximal movements. Other conventions as in Figure 3.
most part of the mesial cortical surface (filled diamonds). From few sites lower face (empty diamonds) movements were elicited. Eye movements were not observed. Eye movements, however, could be evoked from a zone located on the dorsal cortical surface (Schlag and Schlag-Rey, '85, '87; Huerta and Kaas, '90). Histological control showed that this zone was located in F7.
An overview of the movements recorded from all stimu- lated sites of F6 is shown in Table 6. Movements evoked with standard procedure and modified procedures (100 ms stimulus train duration and/or activation procedures) are presented separately. Note the overwhelming majority of arm movements. Note also that the percentage of different types of movements remained the same both with or without the modified procedures.
Unlike F1 and F3, where practically the totality of limb movements were of the fast type, many F6 movements were classified as slow. Out of 41 arm movements evoked with standard procedure, 29 (59%) belonged to the fast type, whereas 12 (41%) belonged to the slow type. When also arm movements elicited with 100 ms stimulus train duration were included (n = 68), the percentage of the two types of movements remained basically the same (fast movements 58%, slow movements 42%). It is important to stress that, in the absence of an objective measurement of the move- ment, we classified as slow only those movements that clearly differed in their appearance (see below) from the classical fast movements observed with stimulation of F1. Those movements that appeared somehow slower than F1 movements, but in other aspects were similar to them, were

474 G. LUPPINO ET AL.
I P A / /
2 rnm - I
uv v 0; uv - . o w I 0 I .
I n , c, *
Fig. 10. Distribution of proximal, distal and tail movements in F3 and in the mesial aspect of F1 in monkeys MKBR and MKGR. All conventions as in Figure 9.
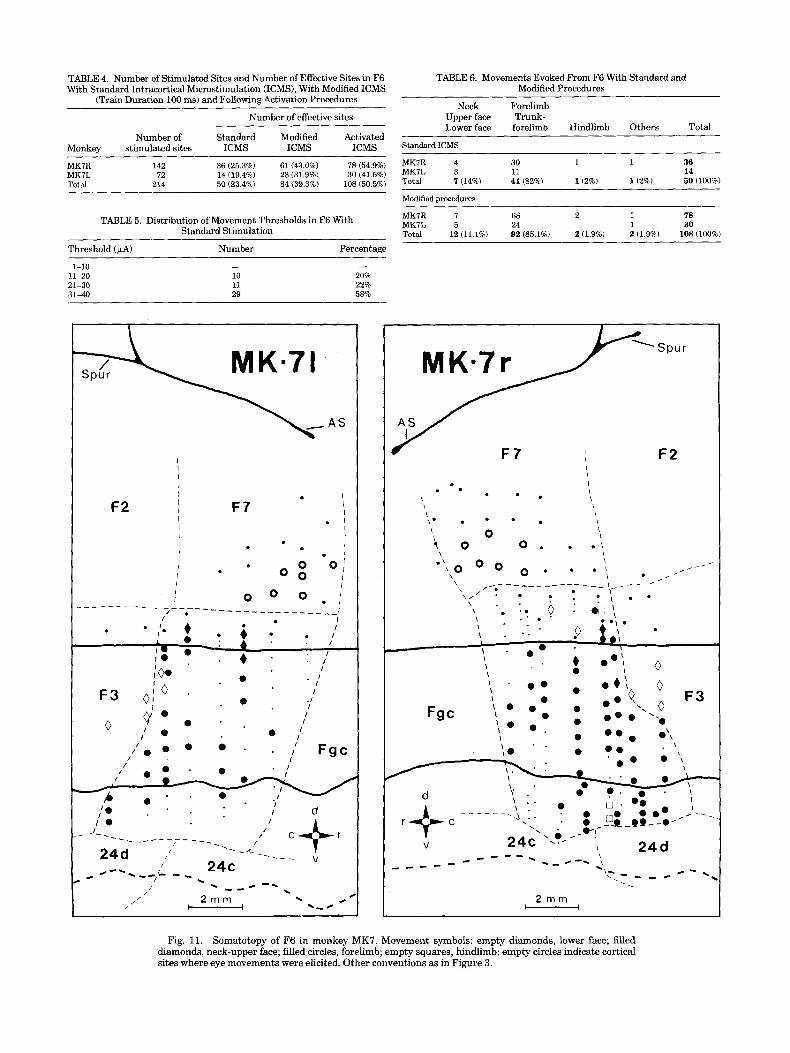
TABLE 4. Number of Stimulated Sites and Number of Effective Sites in F6 With Standard Intracortical Microstimulation (ICMS), With Modified ICMS
(Train Duration 100 ms) and Following Activation Procedures
Number of effective sites
Number of Standard Modified Activated Monkey stimulated sites ICMS ICMS ICMS
MKIR 142 36 (25.3%) 61 (43.0%) 78 (54.9%) MK7L 72 14 (19.4%) 23 (31.9%) 30 (41.6%) Total 214 50 (23.4%) 84 (39.3%) 108 (50.5%)
TABLE 6. Movements Evoked From F6 With Standard and Modified Procedures
Neck Forelimb Upper face Trunk- Lower face forelimb Hindlimb Others Total
Standard ICMS
MKIR 4 30 1 1 36 MKIL 3 11 14 Total 7 (14%) 41 (82%) 1(2%j 1(2%) 50 (100%)
Modified procedures
TABLE 5. Distribution of Movement Thresholds in F6 With Standard Stimulation
MKIR I 68 2 1 78 MKIL 5 24 1 30 Total 12 (11.1%) 92 (85.1%) 2 (1.9%) 2 (1.9%) 108 (100%)
Threshold (4) Number Percentage
1-10 11-20 21-30 31-40
- 10 11 29
- 20% 22% 58%
F2 I I
i I
I I
. I I I
I F7
I , I I I I
I
. I
* . I . 0 0 ; I 0 0 I
I I o o 0 . :
/ I O 0 0 . . I‘ Fgc /
+ I V
MKm7r
I I . I . . 1 \ \ i I \ I
\ 0 .
\ \ \ * . * a . \ \ 0 t o 0 . . . I
\ \ ,-*- - - _ _ _ ------ .--
* I
* \ j O 0 0 0 . . . :\ I . --‘I-----’
\,’ . . .
2 rnm )--------I
Fig. 11. Somatotopy of F6 in monkey MK7. Movement symbols: empty diamonds, lower face; filled diamonds, neck-upper face; filled circles, forelimb; empty squares, hindlimb; empty circles indicate cortical sites where eye movements were elicited. Other conventions as in Figure 3.

476 G. LUPPINO ET AL.
MK=7r
,500 ?? Ta d e v + K n f l e x
!' F2 Ta f l ex +Hi I e C a d d + K n f l e x
Hi1 a d d + K n f l e x a
K n I s C f i e x m
l - a e x t + H i I & C e x t
HI a d d
.T5 I f l ex
, p r o n
Y-J 9 0 0 0 4 n r
Fig. 12. Reconstruction of two microelectrode tracks on outlines of coronal sections at two different antero-posterior levels. Section on the left is more caudal. The dashes along the electrode penetrations indicate the stimulation sites. On the left of the dashes the depth in microns from the cortical surface is indicated. On the right of the dashes the movements evoked and the current thresholds are indicated. Asterisks indicate movements evoked with a stimulus train duration of
classified as fast. Thus it is possible that a bias was introduced against slow movements and that, in reality, their percentage is higher than that here reported.
Out of 41 arm fast movements evoked with standard procedure, 28 were proximal, 12 were proximal and distal, and only 1 was purely distal. By taking into consideration also fast movements evoked with longer stimulus train duration (100 ms), 34 movements turned out to be proxi- mal, 29 were mixed, and 5 exclusively distal. Proximal movements were, therefore, much more frequent than distal movements in F6.
As far as slow movements are concerned, most of them were characterized by a displacement of the arm from an initial position to a new one. This type of displacement is radically different from that of fast movements, which, even in the case of movement involving several articulations, consisted in a short-lasting displacement terminating at the same point from which it started. From some sites, slow movements were evoked with the monkey still in its chair, from others they were evoked exclusively during the mon- key's movements. Typically, the stimulation was effective toward the end of a movement. The stimulation terminated the animal natural movement and brought the arm to a new position, different from that which it would have reached without stimulation. The slow movements evoked during natural movements were highly repeatable and had the appearance of spontaneous movements. In some cases,
Fm p r o n
El e x t + Fm p r o n
H I f l e x + A n f l e x + l
70004 n r
'1 a d d
100 ms. Abbreviations: TI-5, toes; An, ankle; Kn, knee; Hi, hip; Ta, tail; F1-5, fingers; Fm, forearm; El, elbow; S, shoulder; n.r., non responsive; ext, extension; flex, flexion; abd, abduction; add, adduction; dev, deviation; retr, retraction; elev, elevation; ext rot; external rotation; pron, pronation; I, ipsilateral movement; I & C, bilateral movement. Dotted lines on the outlines indicate borders between cytoarchitectonic areas. Dots represent large pyramidal cells.
they brought the arm consistently to the same final posi- tion, regardless of where the arm was located during stimulation. The interpretation of slow movements is not simple. Their high frequency, however, in F6 and their virtual absence in F3 is a strong evidence for a functional independence of the two cytoarchitectonic areas.
Area 24d (dorso-caudal cingulate cortex) Excitability and threshold. While mapping F3, we no-
ticed that in the depth of the cortex there was frequently an abrupt change of somatotopy and a modification in the characteristics of the evoked movements. Two penetrations showing these changes are presented in Figure 12. Penetra- tion 43 (left side) crossed F1 (leg field), the underneath located white matter and F3. In F3 the stimulation pro- duced tail and leg movements. At 6,500 pm, however, leg movements were substituted by shoulder movements, which were subsequently observed for about 2 mm. Shoulder movements were of simple type. Penetration 38 (right side) was more rostral. It crossed F3 leg field and then F3 arm field. At the depth of 5,250 pm, leg movements appeared. They concerned the ankle, toes, and hip. Histological examination showed that the changes in somatotopy in the depth of the penetrations coincided with the boundary between F3 and area 24d.
MOTOR MAPS OF MESIAL AREA 6 AND AREA 24 477
TABLE 7. Number of Stimulated Sites and Number of Effective Sites in Area 24d With Standard ICMS or With Modified ICMS
(Train Duration 100 ms)
Number of effective sites
~ ~~
Number of Standard Modified Monkey stimulated sites ICMS ICMS MK5R MK6R MK7R Total
48 72
110 230
16 (33.3%) 20 (41.6%) 34 (47.2%) 45 (62.5%) 48 (43.6%) 62 (56.4%) 98 (42.6%) 127 (55.2%)
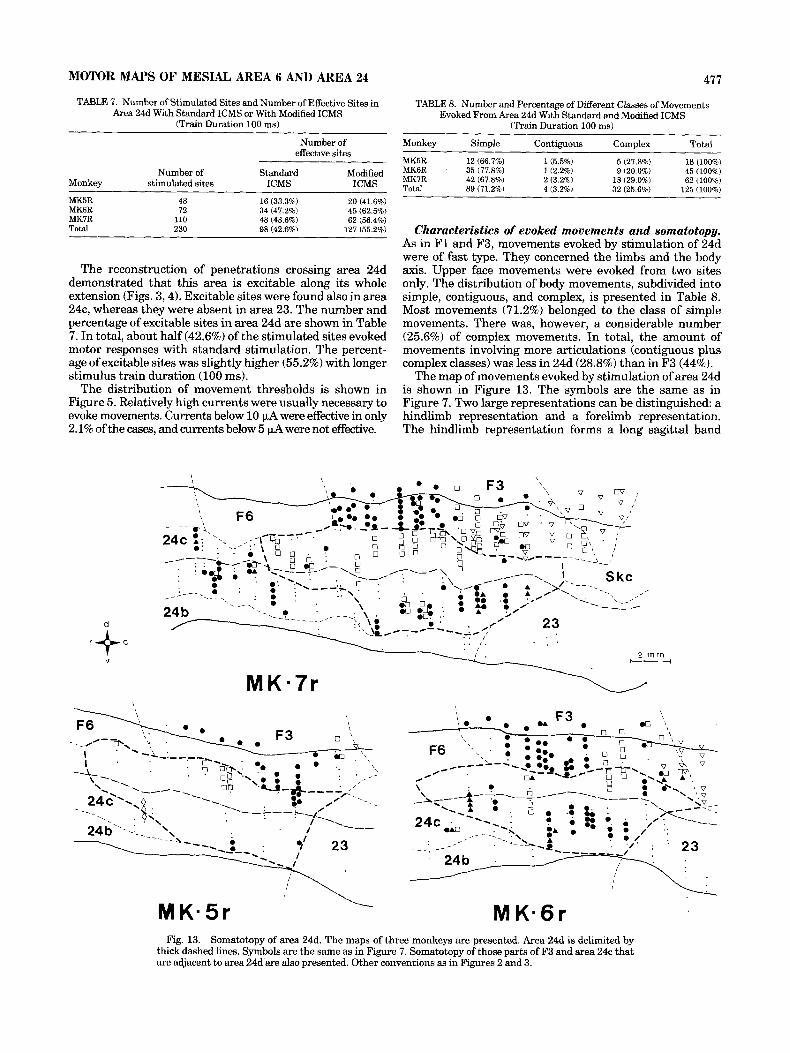
The reconstruction of penetrations crossing area 24d demonstrated that this area is excitable along its whole extension (Figs. 3,4). Excitable sites were found also in area 24c, whereas they were absent in area 23. The number and percentage of excitable sites in area 24d are shown in Table 7. In total, about half (42.6%) of the stimulated sites evoked motor responses with standard stimulation. The percent- age of excitable sites was slightly higher (55.2%) with longer stimulus train duration (100 ms).
The distribution of movement thresholds is shown in Figure 5. Relatively high currents were usually necessary to evoke movements. Currents below 10 p4 were effective in only 2.1% of the cases, and currents below 5 p A were not effective.
TABLE 8. Number and Percentage of Different Classes of Movements Evoked From Area 24d With Standard and Modified ICMS
(Train Duration 100 ms)
Monkey Simple Contiguous Complex Total
MK5R 12 (66.7%) 1 6 5 % ) 5 (27.8%) 18 (100%) MK6R 35 (77.8%) l(2.290) 45 (100%) MK7R 42 (67.8%) 2 (3.2%) 18 (29.0%) 62 (100%) Total 89 (71.2%) 4 (3.2%) 32 (25 6 6 ) 125 (100%)
9 (20.0%)
Characteristics of evoked movements and somatotopy. As in Fl and F3, movements evoked by stimulation of 24d were of fast type. They concerned the limbs and the body axis. Upper face movements were evoked from two sites only. The distribution of body movements, subdivided into simple, contiguous, and complex, is presented in Table 8. Most movements (71.2%) belonged to the class of simple movements. There was, however, a considerable number (25.6%) of complex movemeuts. In total, the amount of movements involving more articulations (contiguous plus complex classes) was less in 24d (28.8%) than in F3 (44%).
The map of movements evoked by stimulation of area 24d is shown in Figure 13. The symbols are the same as in Figure 7. Two large representations can be distinguished: a hindlimb representation and a forelimb representation. The hindlimb representation forms a long sagittal band
V 2 m m
r+c - MKl7r
r o h
--.
Fig. 13. Somatotopy of area 24d. The maps of three monkeys are presented. Area 24d is delimited by thick dashed lines. Symbols are the same as in Figure 7. Somatotopy of those parts of F3 and area 24c that are adjacent to area 24d are also presented. Other conventions as in Figures 2 and 3.

478 G. LUPPINO ET AL.
TABLE 9. Number and Percentage of Axial, Proximal and Distal Movements Evoked From Area 24d With Standard and Modified ICMS
(Train Duration 100 ms) ~~ ~
Number of movements ~
Axial Proximal Distal Prox.-Dist. Total
Forelimb Hindlimb Forelimb
Hindlimb +
35 10 9 54 (43.2%) 14 26 14 54 (43.2%)
3 3 6 (4.8%)
Axial 11 11 (8.8%) Total 11 (8.8%) 52 (41.6%) 36 (28.8%) 26 (20.8%) 125 (100%)
mostly located in the upper bank and fundus of the cingulate sulcus. For most of its extension it lies below the arm representation of F3. Area 24d forelimb representation is roughly parallel to that of the hindlimb and occupies essentially the lower bank of the cingulate sulcus. Caudally, the forelimb representation tends to have a more dorsal position surrounding the caudal end of the hindlimb field. The two sites from which upper face movements (blinking) were evoked were located in the rostral part of area 24d, on the border with 24c (monkey MK5).
Movements evoked from the hindlimb field included distal movements (n = 261, proximal movements (n = 141, and mixed, proximo-distal movements (n = 14). Distal move- ments consisted of movements of the allux, toes, and ankle. Proximal movements consisted in displacements of hip and knee. Mixed hindlimb and forelimb movements were evoked from 6 sites. There was no spatial segregation between distal and proximal movements.
Movements evoked from the forelimb field also included distal movements (n = lo), proximal movements (n = 3 3 , and mixed proximo-distal movements (n = 9). Distal and proximal movements were not segregated. Shoulder move- ments were the most represented among the proximal movement (n = 35), whereas finger movements were the most represented (n = 8) among the distal movements. Wrist movements in isolation (n = 4) or associated with finger movements (n = 1) were also observed. An overview of axial, proximal, and distal movements recorded from all stimulated sites is presented in Table 9.
DISCUSSION The present study shows that the mesial cortical surface
rostral to the precentral motor area (Fl) is formed by a mosaic of areas that differ in their physiological properties and somatotopic organization. Of these areas two lie in the mesial agranular frontal cortex. The first, adjacent to F1, contains a complete movement representation. This area, which appears to possess most of the characteristics classi- cally attributed to the supplementary motor area (SMA), is co-extensive with cytoarchitectonic F3. A second area, rostral to the first, contains essentially a forelimb represen- tation. It is much less excitable with intracortical stimula- tion than the former area and, cytoarchitectonically, corre- sponds to F6. Two additional motor areas are located in the cingulate agranular cortex. The first is co-extensive with area 24d and contains a forelimb and a hindlimb representa- tion. The second one, which was only marginally studied in the present experiments, is located in area 24c. I t contains an arm representation. Although complex, this picture of the fronto-cingulate region clarifies several debated points about the somatotopic organization of this region and its
role in motor and "supramotor" functions (Eccles, '82; Wiesendanger , '86).
F3 (SMA-proper) Somatotopic organization. Early stimulation studies
revealed that on the mesial cortical wall, just in front of the precentral motor area, there is another motor area (supple- mentary motor area, SMA; Penfield and Welch, '51; Wool- sey et al., '52). Woolsey and coworkers ('52) showed also that this area is somatotopically organized with a rostro- caudal sequence of oro-facial, forelimb, and hindlimb fields. In recent years two series of arguments have been advanced against the classical Woolsey's picture of the SMA. First, regional blood flow studies (e.g., Orgogozo and Larsen, '79) and intracortical stimulation in neurosurgical patients (Talairach and Bancaud, '66) failed to reveal a rostro- caudal somatotopic pattern. Second, the discovery that the mesial cortex rostral to the precentral motor area becomes active during movement preparation (e.g., Kornhuber and Deecke, '65; Roland et al., '80) led some reviewers to favor the notion of SMA as a nontopographically organized "supramotor" center (Eccles, '82; Eccles and Robinson, '84; see also Wiesendanger, '86).
Neurophysiological data obtained in monkeys do not support the nontopographical idea of the SMA. Single neuron recordings in behaving monkeys showed that neu- rons active before hindlimb movements were located cau- dally in respect with those active before forelimb move- ments (Brinkman and Porter, '79; Tanji and Kurata, '82). Furthermore, there was a segregation between forelimb neurons related to proximal and to distal movements (Tanji and Kurata, '79). A clear, convincing rostro-caudal progres- sion of oro-facial, forelimb, and hindlimb representation was reported in a recent intracortical microstimulation study in which a relatively long stimulus train duration (100 ms) was used (Mitz and Wise, '87).
Skepticism about a somatotopic organization of the SMA was expressed, however, by Wiesendanger and his cowork- ers (Macpherson et al., '82a,b; Hummelsheim et al., '86) on the basis of their microstimulation studies. Although they also observed that hindlimb representation was more cau- dal than the forelimb representation, they raised several criticisms against the classical Woolsey's scheme. Their objections can be summarized as follow: (1) the SMA leg representation is small and intermingled with the forelimb field. Rostrally, ventral to forelimb field, there is an exten- sion of the leg representation. This anterior leg representa- tion, observed also by Penfield and Welch ('51) and Tanji and Kurata ('82), is obviously at odds with the classical SMA somatotopy; (2) the excitable sites of the SMA are confined to its caudal half; no movements could be evoked from the SMA rostral part (Hummelsheim et al., '86); and (3) no face representation is present in the SMA.
Although the data of Wiesendanger were obtained using parameters of intracortical microstimulation (12 stimuli at 330 Hz, train duration = 36 msec), which are not the best for exciting the SMA (see also Smith, '79; Wise and Tanji, '81; Palmer et al., '82; and the methodological discussion on this point in Mitz and Wise, '871, their objections cannot be simply dismissed as due to this methodological weakness. Furthermore, their objections are reinforced by some incon- sistencies on the SMA somatotopy that emerge if one compares single neuron data (Brinkman and Porter, '79;

MOTOR MAPS OF MESIAL AREA 6 AND AREA 24 479
Tanji and Kurata, '82) with stimulation findings (Mitz and Wise, '87). In fact, the arm representation described in recording experiments not only overlap with the oro-facial representation of stimulation experiments but even ex- tends farther rostrally (see, e.g., Tanji and Kurata, '82, who present a precise overview of their penetration sites).
The findings of the present studies appear to give a satisfactory answer to all the above mentioned difficulties and inconsistencies. They support the view that the SMA of the macaque is somatotopically organized, provided, how- ever, that the SMA, is considered limited to the cytoarchitec- tonical F3. Let us now examine the various objections of Wiesendanger one by one.
There are two problems here: (1) the apparent small extension of this field, and (2) the existence of a leg field located rostrally below the forelimb representation. The findings of the present and the accom- panying article indicate that the apparent smallness of the hindlimb representation is due to an erroneous setting of the boundary between area 4 and area 6 with attribution to area 4 of the caudal part of area 6 (see a discussion of this point in Matelli et al., '91). Physiologically, F1 (area 4) is characterized by a clear segregation of distal and proximal movements. Such a segregation is very limited in F3. Complex and contiguous types of movements are very common in F3, whereas they are rare or absent in F1. Finally, on the average, thresholds are higher in F3 than in F1. These functional properties are found also in the caudal sector of F3, classically attributed to area 4. When this sector of agranular cortex is given to F3, the leg field of this area becomes as large as its arm field.
A leg field located rostrally, below the arm field, was constantly found in the present experiments. Transition from F3 forelimb representation to rostral leg field was accompanied, however, by an increase in threshold and by a decrease of complex movements. These changes in physio- logical properties coincided with the anatomical transition from F3 to area 24d. Thus the rostral leg field is not an evidence for a intermingled somatotopy of F3 (Macpherson et al., '82a,b). This field simply belongs to another motor area.
Oro-facial field covers a lim- ited amount of F3. Its location is in the approximate correspondence of a line passing at the level of the posterior end of the arcuate sulcus (see also Mitz and Wise, '87). This location and its limited extent is in conflict with the oro-facial representation of Woolsey's map, which is large and co-extensive with mesial 6ap (F6). Since F6 is electri- cally little excitable, it is possible that a spread of current from F6 to rostral F3 led Woolsey and his coworkers ('52) to attribute the properties of rostral F3 to F6.
Curiously enough, one of the apparently strongest evi- dence in favor of a somatotopic organization of the SMA, i.e., that coming from single neuron recordings (Brinkman and Porter, '79; Tanji and Kurata, '82), is, in reality, irrelevant to the issue. The rostral border of F3 (SMA- proper) extends about 2.5 mm rostral to the postarcuate line (Matelli et al., '91). Recordings showing forearm repre- sentation in the "SMA" (and the distal representation in particular) extended up to 6 mm in front of the postarcuate line (Brinkman and Porter, '79; Tanji and Kurata, '82). These recordings, therefore, were located outside F3. In other words, in agreement with the present findings, what they show is that in mesial 6ap (F6) there is a further arm
Hindlimb representation.
Oro-facial representation.
movement representation. They do not say anything about the organization of the SMA-proper.
Summing up, the available data indicate that on the mesial cortical wall, rostral to precentral motor area, there is a somatotopically organized motor area. This area is co-extensive with F3 and extends for about 9 mm in rostro-caudal direction. It is electrically excitable and has direct connections with the spinal cord (Biber et al., '78; Murray and Coulter, '81; Macpherson et al., '82a; Toyoshima and Sakai, '82; Hutchins et al., '88; Keizer and Kuypers, '89; Nudo and Masterton, '90). This area, which we propose to call SMA-proper, does not include either the mesial aspect of area 6ap (F6), or the motor areas of the cingulate cortex. The inclusion of these areas into the SMA led to confusion about the somatotopy of the region and, inevita- bly, about its functions. Although electrical cortical stimula- tion tells little about the function of stimulated areas (for discussion of this issue, see Phillips and Porter, '771, a few points emerging from the present data on SMA-proper are of interest for a better understanding of the possible role of this area. They are discussed in the next section.
Functional characteristics. A first feature that distin- guishes SMA-proper from the precentral motor area is the frequency with which movements involving two or more articulations are evoked from it at threshold current (com- plex and contiguous classes of movements). Results qualita- tively in agreement with ours were reported in previous microstimulation studies (Macpherson et al., '82a; Hum- melsheim et al., '86; Mitz and Wise, '87). Movements involving several joints at threshold stimulation were re- ported also in inferior area 6 (F4) (Gentilucci et al., '88). Taken together, these data confirm early observations based on surface cortical stimulation (see ref. in Humphrey, '79) that the motor areas rostral to area 4 control the motor activity in a more global way than the precentral cortex.
A second feature that characterizes SMA-proper is the strong predominance of proximal movements over distal movements. These findings are in contrast with the data of Mitz and Wise ('87) who reported a rather rich distal representation. They are, however, consistent with early stimulation experiments (Penfield and Welch, '5 1; Macpher- son et al., '82a) as well as with the observation that following ablation of the precentral cortex, SMA stimula- tion effects are limited to proximal and axial movements (Penfield and Welch, '51; Wiesendanger et al., '73).
A rich distal forearm representation in the mesial wall of the agranular frontal cortex was observed in studies based on the examination of neuronal discharge during move- ments (Brinkman and Porter, '79; Tanji and Kurata, '82). Inspection of the penetration locations shows, however, that in these studies the distal representation was mostly outside the SMA-proper (see above). If the same data are now re-examined discarding penetrations outside SMA- proper, a completely different picture emerges.
For example, Tanji and Kurata ('79) reported that out of 225 neurons that changed their activity with forelimb movements, 60 were related to distal movements, 150 to proximal movements, and 15 to both. If, however, the rostral penetrations (from 2 mm from the postarcuate line upward) are not counted, the proportion proximal vs. distal becomes 5 to 1 (150 proximal units to 32 distal units). Thus data on neuronal activity if limited to SMA-proper, support rather than disprove the notion that the SMA-proper controls mostly proximal movements.

480 G. LUPPINO ET AL.
According to Woolsey et al. ('52), the site of distal representation is primary located on the dorsal convexity and on the adjacent mesial cortical surface. In contrast, Macpherson et al. ('82a), reported that distal movements are generally evoked from sites located deep on the mesial surface of the hemispheres. Finally, Mitz and Wise ('87) did not find any preferential location for distal movements.
Distal movements were little represented in our experi- ments and, because of the frequent occurrence of complex movements, they were only partially segregated from proxi- mal movements. However, a limited pure distal movement representation was found in all monkeys. This representa- tion was located in the lowest part of the mesial surface and in the upper bank of the cingulate sulcus.
F6 (mesial 6ap) The present findings clearly indicate that the rostral part
of the mesial agranular frontal cortex (F6, mesial 6ap) differs functionally from its caudal part. F6 is poorly excitable with standard electrical intracortical stimulation. Furthermore, when appropriate activation procedures are applied, its stimulation frequently produces slow, global movements virtually never observed in the SMA-proper. A large part of F6 is related to arm movements. Its forelimb representation, however, is separated from that of F3 by the oro-facial field of this last area. A reappearance of the representation of the same body part in another cytoarchi- tectonic area is considered a strong argument for separat- ing functionally cortical areas.
The poor excitability of F6 is consistent with its cortico- descending and cortico-cortical connections. Unlike SMA- proper, 6ap (of which F6 is a part) does not send fibers to the spinal cord. I ts subcortical projections are directed to putamen and caudate nucleus, to the pons, and to those brainstem zones that have been associated with the control of proximal and axial movements (Kuypers and Lawrence, '67; Kunzle, '78; Kuypers, '81; Keizer and Kuypers, '89). Cortical connections of F6 as derived from experiments in which connections of precentral cortex were studied (Mat- sumura and Kubota, '79; Muakkassa and Strick, '79; Godschalk et al., '84; Matelli et al., '86; Ghosh et al., '87) or from experiments specifically devoted to F6 (Luppino et al., '90) clearly indicate that this area has no direct connections with F1. F6 is connected with the other premotor areas and especially with F5 (inferior area 61, the prefrontal cortex, and with cingulate areas. Thus the lack of cortico-spinal pathways, as well as the absence of a direct access to the precentral motor cortex, most likely explains the difficulty to evoke movements from this area.
I t is not easy to give a satisfactory explanation of slow movements that the stimulation of F6 evokes during rest and, especially, during natural animal's movements. These slow movements in many aspects are similar to movements spontaneously executed by the animal. Our impression was that the stimulation of F6 was somehow able to trigger arm motor programs and that, when the animal was already moving, the probability of triggering these programs in- creased. It is impossible to indicate which pathways may be responsible for the evoked slow movements. The most parsimonious explanation seems to be that brainstem centers, connected with F6, control various arm motor programs and that the stimulation of F6 triggers them, especially if they are already activated by natural animal's movements. Alternative explanations, i.e., that basal- ganglia, cerebellar, or cortico-cortical circuits are responsi-
ble for slow movements, although possible, appear to be less likely because of the length of these circuits and the consequent low probability of their activation.
A puzzling aspect of F6 functional organization concerns the type of movements represented in this area. Rizzolatti et al. ('90) recorded single neuron activity from F6 and correlated it with natural movements triggered by food or by threatening stimuli. In agreement with the present findings, they found that orienting movements were repre- sented on the dorso-medial convexity of F6 and in a narrow strip adjacent to it on the mesial convexity. Neurons recorded from the remaining part of F6 were mostly active before and during arm movements. Among arm-related neurons, some fired every time the animal moved its arm, regardless of its purpose, others were specifically related to reaching-grasping movements. Rizzolatti et al. ('90) stressed the fact that, in contrast to inferior area 6, where neurons become selectively active during motor acts involving proxi- mal or distal movements (Gentilucci et al., '88; Rizzolatti et al., '881, in F6 this relation was absent. They suggested that F6 is involved in a global control of arm movements and, specifically, in the arm movement initiation.
Although this theorization on the role of F6 in movement control is consistent with the anatomical connections of this area, it is in sharp contrast with the findings that in the rostral part of the mesial agranular cortex (presumably F6, see above) there is a representation of distal forelimb movements (Brinkman and Porter, '79; Tanji and Kurata, '79, '82). Our impression is that this discrepancy is mostly related to the different assumptions that underlie the two sets of experiments. The assumption on the basis of the experiments in which neurons related to distal movements were recorded (Brinkman and Porter, '79; Tanji and Ku- rata, '79, '82) was that elementary movements (wrist flexion, extension, pronation, finger movements, etc.) are represented in F6 as they are in F1. The aim of the investigators was to determine which elementary move- ment was represented (see especially Brinkman and Porter, '79). Rizzolatti et al. ('90) studied F6 after their discovery that in inferior area 6 the goal of an action was as important in triggering the neurons as the peripheral movement manifestations. Thus they examined first whether F6 neu- rons become active during particular actions and then whether, within this action, there was a specificity for distal or proximal motor acts.
The difference in the assumptions underlying the two sets of experiments may lead to different results. An example of this is represented by the experiments of Tanji and Kurata ('79). In these experiments the monkey's arm was blocked in a cast and fixed to the sitting chair in such a way that only rotations of the shoulder joint could be executed. A key was attached to the distal end of the cast, which could be pressed by flexion of the fingers. The monkey was required to move the cast to a correct position and then to press the key in response to a light stimulus. Neurons firing with displacements of the shoulder were classified as proximal, neurons firing with finger flexion were classified as distal. The examined shoulder and finger movements differed, however, not only in the moved joints, as stated by the authors, but also in their purpose. Shoulder movements changed the animal posture, whereas finger flexion were a kind of modified grasping movement, whose aim was to take an object. Thus it may be that, as suggested by Tanji and Kurata ('79), the presence of neurons firing with finger flexion indicates a distal motor representation.

MOTOR MAPS OF MESIAL AREA 6 AND AREA 24 481
It may be equally well that what is really represented in F6 is a global reaching-grasping action, but since proximal goal-directed movements were not tested, the global aspect of the motor representation passed unnoticed.
Supplementary eye field (SEF) The existence of an electrically excitable eye movement
representation in the rostral part of area 6 was first described by Schlag and Schlag-Rey ('85, '87) and, subse- quently, confirmed by many authors (Gould et al., '86; Huerta et al., '87; Mitz and Wise, '87; Mann et al., '88; Huerta and Kaas, '90). This eye field is generally referred to as the supplementary eye field (SEF).
In the present study, eye movements could be evoked with low threshold currents from a cortical region, extend- ing for about 2 x 3 mm in the rostral and medial part of superior area 6. The location of this region coincides with that of the SEF as indicated by Schlag and Schlag-Rey ('85, '87) and corresponds to the medial part of cytoarchitectonic F7 (Matelli et al., '91). Our study showed also that no eye movements could be evoked from mesial area 6. The location of eye representation in a cytoarchitectonic area, rostral to F2 and not to F3 (SMA-proper) and lateral to F6, suggests that the SEF is not a part of the supplementary motor area, but it represents a different functional area. It is important to note that the SEF markedly differs from F6 for its cortico-cortical connections (Luppino et al., '90; Huerta and Kaas, '90). F7 is connected with the frontal eye field (area 8), whereas F6 is connected with the arm representations in the premotor cortex. Furthermore, F6 and F7 receive afferents from different prefrontal and cingulate regions.
Recently, Mitz and Godschalk ('89) reported that eye movements can be evoked with intracortical microstimula- tion from a broad cortical region, located on the dorsal convexity and extending rostrally into the mesial cortex. Unfortunately, these authors did not provide information on the cytoarchitectonics of the stimulated areas. However, judging from the location of their penetrations, which were well in front to the end of superior arcuate sulcus, it is very likely that their stimulation sites were located in the prefrontal cortex. Thus considering its completely different anatomical location, the eye field reported by Mitz and Godschalk ('89) should represent a further eye movement representation, independent of the SEF.
Cingulate motor areas There are several observations that surface electrical
stimulation of the cingulate cortex produces somatic motor responses (Smith, '45; Ward, '48; Kaada, '51; Showers, '59; Van Buren and Fedio, '76). No systematic studies, however, have been carried out of this region with intracortical microstimulation, although occasionally motor responses have been obtained with this technique (Macpherson et al., '82a,b; Mitz and Wise, '87). It is interesting to note that there is a good evidence that cingulate cortex (or, at least, part of it) sends projections to the spinal cord (Biber et al., '78; Murray and Coulter, '81; Macpherson et al., '82a; Toyoshima and Sakai, '82; Hutchins et al., '88; Keizer and Kuypers, '89; Nudo and Masterton, '90) and to the precen- tral motor cortex (Muakkassa and Strick, '79; Godschalk et al., '84; Leichnetz, '86; Morecraft and Van Hoesen, '88).
The present study demonstrated that, in the cingulate cortex, there are two subareas from which motor responses can be evoked with intracortical microstimulation. The first
subarea corresponds to the cytoarchitectonic area 24d (Matelli et al., '91). This area is topographically organized, with a forelimb representation located caudally and ven- trally and a hindlimb representation located rostrally and dorsally. From two cortical sites, oro-facial movements were also elicited. Thus an oro-facial representation most likely exists in area 24d. This representation, however, appears to be very small. Although correlations between anatomical and physiological data derived from different sources are difficult to draw, our finding that area 24d is topographically organized fits well with anatomical data showing that both area 24 projections to the spinal cord (Hutchins et al., '88) and connections with area 4 (Muak- kassa and Strick, '79; Morecraft and Van Hoesen, '88) are also topographically organized.
The second subarea, whose stimulation produced body movements, is area 24c (Matelli et al., '91). Although this region has not been extensively studied in the present experiments, its lower electrical excitability and less clear topographic organization suggest that area 24c is an area functionally independent of area 24d. In favor of this idea are anatomical data showing that the dorso-caudal part of area 24 (24d) has richer cortico-spinal projections than the ventro-rostral part (24c) (Hutchins et al., '88) and that, whereas area 24d is connected with F1 (area 4) and F3 (Muakkassa and Strick, '79; Luppino et al., '901, area 24c is not.
In conclusion, the physiological data of the present study, together with previous anatomical observations, strongly support the notion (Muakkassa and Strick, '79) that area 24d should be considered as a further motor area; its role, however, in motor control remains to be ascertained.
ACKNOWLEDGMENTS The work was supported by EEC contract no SCl*-
0177-C and by grants from CNR and MPI to G.R.
LITERATURE CITED Biber, M.P., L.W. Kneisley, and J.H. Lavail(1978) Cortical neurons project-
ing to the cervical and lumbar enlargements of the spinal cord in young and adult rhesus monkeys. Exp. Neurol. 59:492-508.
Brinkman, C., and R. Porter (1979) Supplementary motor area in the monkey: activity of neurons duringperformance of a learned motor task. J. Neurophysiol. 42:681-709.
Donoghue, J.P., and S.P. Wise (1982) The motor cortex of the rat: Cytoarchi- tecture and microstimulation mapping. J. Comp. Neurol. 212%-88.
Eccles, J.C. (1982) The initiation of voluntary movements by the supplemen- tary motor area. Arch. Psychiat. Nervenkr. 231:423-431.
Eccles, J.C., and D.N. Robinson (1984) The Wonder of Being Human. Our Brain and Our Mind. New York: Free Press, Macmillan.
Gentilucci, M., L. Fogassi, G. Luppino, M. Matelli, R. Camarda, and G. Rizzolatti (1988) Functional organization of inferior area 6 in the macaque monkey: I. Somatotopy and the control of proximal move- ments. Exp. Brain Res. 71:475490.
Ghosh, S., C. Brinkman, and R. Porter (1987) A quantitative study of the distribution of neurons projecting to the precentral motor cortex in the monkey (Macaca fascicularis). J. Comp. Neurol. 259:424444.
Godschalk, M., R.N. Lemon, H.G.J.M. Kuypers, and H.K. Ronday (1984) Cortical afferents and efferents of monkey postarcuate area: An anatom- ical and electrophysiological study. Exp. Brain Res. 56:410-424.
Gould, H.J. 111, C.G. Cusick, T.P. Pons, and J.H. Kaas (1986) The relation- ship of corpus callosum connections to electrical stimulation maps of motor, supplementary motor, and the frontal eye fields in owl monkeys. J. Comp. Neurol. 247r297-325.
Huerta, M.F., L.A. Krubitzer, and J.H. Kaas (1987) Frontal eye field as defined by intracortical microstimulation in squirrel monkeys, owl

482 G. LUPPINO ET AL.
monkeys, and macaque monkeys 11. Cortical connections. 3. Comp. Neurol. 265332-36 1.
Huerta, M.F., and J.H. Kaas (1990) Supplementary eye field as defined by intracortical microstimulation: Connections in Macaques. J. Comp. Neurol. 293:299-330.
Hummelsheim, H., M. Wiesendanger, M. Bianchetti, R. Wiesendanger, and J. Macpherson (1986) Further investigations of the efferent linkage of the supplementary motor area (SMA) with the spinal cord in the monkey. Exp. Brain Res. 6535-82.
Humphrey, D.R. (1979) On the cortical control of visually directed reaching: contributions by nonprecentral motor areas. In R.E. Talbott and D.R. Humphrey (eds): Posture and Movement. New York Raven Press, pp. 51-112.
Hutchins, K.D., A.M. Martino, and P.L. Strick (1988) Corticospinal projec- tions from the medial wall of the hemisphere. Exp. Brain Res. 71:667472.
Kaada, B.R. (1951) Somato-motor, autonomic and elettrocorticographic responses to electrical stimulation of “rhinencephalic” and other struc- tures in primates, cat and dog. ActaPhysiol. Scand. 24(Suppl. 83): 1-285.
Keizer, K., and H.G.J.M. Kuypers (1989) Distribution of corticospinal neurons with collaterals to the lower brain stem reticular formation in monkey (Macaca fascicularis). Exp. Brain Res. 74:311-318.
Kornhuber, H.H., and L. Deecke (1965) Hirnpotentialandenmgen by Willkar- bewegungen und passiven Bewegungen des Menschen: Bereitschaftpoten- tial und reafferente Potentiale. Pfluegers Arch. Ges. Physiol. 284:l-17.
Kunzle, H. (1978) An autoradiographic analysis of the efferent connections from premotor and adjacent prefrontal regions (area 6 and 9) in Macaca fascicularis. Brain Behav. Evol. 15:185-234.
Kuypers, H.G.J.M. (1981) Anatomy of the descending pathways. In Brooks V.B. (ed): Handbook of Physiology, Vol. 11. Motor Control, Part 1. Bethesda: American Physiological Society, pp. 597-666.
Kuypers, H.G.J.M., and D.G. Lawrence (1967) Cortical projections to the red nucleus and the brain stem in the rhesus monkey. Brain Res. 41151-188.
Leichnetz, G.R. (1986) Afferent and efferent connections of the dorsolateral precentral gyrus (area 4, handiarm region) in the macaque monkey, with comparison to area 8. J. Comp. Neurol. 254:460-492.
Luppino, G., M. Matelli, and G. Rizzolatti (1990) Cortico-cortical connec- tions of two electrophysiologically identified arm representations in the mesial agranular frontal cortex. Exp. Brain Res. 82:214-218.
Macpherson, J.M., C. Marangoz, T.S. Miles, and M. Wiesendanger (1982a) Microstimulation of the supplementary motor area (SMA) in the awake monkey. Exp. Brain Res. 45410-416.
Macpherson, J.M., M. Wiesendanger, C. Marangoz, and T.S. Miles (1982b) Corticospinal neurones of the supplementary motor area of monkeys. A single unit study. Exp. Brain Res. 483-88.
Mann, S.E., R. Thau, and P.H. SchiUer (1988) Conditional task-related responses in monkey dorsomedial frontal cortex. Exp. Brain Res. 69:46&468.
Matelli, M., G. Luppino, and G. Rizzolatti (1985) Patterns of cytochrome oxidase activity in the frontal agranular cortex of macaque monkey. Behav. Brain Res. 18:125-137.
Matelli, M., G. Luppino, and G. Rizzolatti (1991) Architecture of superior and mesial area 6 and the adjacent cingulate cortex in the macaque monkey. J. Comp. Neurol. 311:445462.
Matelli, M., R. Camarda, M. Glickstein, and G. Rizzolatti (1986) Afferent and efferent projections of the inferior area 6 in the macaque monkey. J. Comp. Neurol. 251t281-298.
Matelli, M., G. Luppino, R. Camarda, and G. Rizzolatti (1990) Mesial area 6 in monkey is formed by two different cytoarchitectonic and physiological areas. SOC. Neurosci. Abstr. 16/1):106.8.
Matsumura, M., and K. Kubota (1979) Cortical projection of hand-arm motor area from postarcuate area in macaque monkey: a histological study of retrograde transport of horseradish peroxidase. Neurosci. Lett. 11 241-246.
Mitz, A.R., and S.P. Wise (1987) The somatotopic organization of the supplementary motor area: intracortical microstimulation mapping. J. Neurosci. 7:lOlO-1021.
Mitz, A.R., and M. Godschalk (1989) Eye-movement representation in the frontal lobe of rhesus monkeys. Neurosci. Lett. 106:157-162.
Morecraft, R.J., and G.W. Van Hoesen (1988) Somatotopical organization of cingulate projections to the primary and supplementary motor cortices in the old-world monkey. SOC. Neurosci. Abstr. 14:329.5.
Muakkassa, K.F., and P.L. Strick (1979) Frontal lobe inputs to primate motor cortex: evidence for four somatotopically organized “premotor” areas. Brain Res. 177:176-182.
Murray, E., and J.D. Coulter (1981) Organization of corticospinal neurons in the monkey. J. Comp. Neurol. 195:339-365.
Nudo, R.J., and R.B. Masterton (1990) Descending pathways to the spinal cord, 111: Sites of origin of the corticospinal tract. 3. Comp. Neurol. 296:559-583.
Orgogozo, J.M., and 8. Larsen (1979) Activation ofthe supplementary motor area during voluntary movements suggests it works as a supramotor area. Science 206:847-850.
Palmer, C., E.M. Schmidt, and J.S. McIntosh (1981) Corticospinal and corticorubral projections from the supplementary motor area in the monkey. Brain Res. 209:305-314.
Penfield, W., and K. Welch (1951) The supplementary motor area of the cerebral cortex. Arch. Neurol. Psychiatry 66.289-317.
Phillips, C.G., and R. Porter (1977) Corticospinal Neurones: Their Role in Movement. London: Academic Press (Monogr. Physiol. SOC. 341, pp. 158-161.
Rizzolatti, G. (1989) Functional properties of a mesial premotor area (area F6) in the macaque monkey. Proc. XXXI Int. Congress Physiological Sciences, S 5045.
Rizzolatti, G., R. Camarda, M. Fogassi, M. Gentilucci, G. Luppino, and M. Matelli (1988) Functional organization of inferior area 6 in the macaque monkey: 11. Area F5 and the control of distal movements. Exp. Brain Res. 71:491-507.
Rizzolatti, G., M. Gentilucci, R. Camarda, V. Gallese, G. Luppino, and M. Matelli (1990) Neurons related to reaching-grasping arm movements in the rostral part of area 6 (area 6ap). Exp. Brain Res. 82337-350.
Roland, P.E., N.A. Larsen, and E. Shinhoj (1980) Supplementary motor area and other cortical areas in organization of voluntary movements in man. J. Neurophysiol. 43:118-136.
Schlag, J., and M. Schlag-Rey (1985) Unit activity related to spontaneous saccades in frontal dorsomedial cortex of monkey. Exp. Brain Res. 58:208-2 11.
Schlag, J., and M. Schlag-Rey (1987) Evidence for a supplementary eye field. J. Neurophysiol. 57:179-200.
Showers, M.J.C. (1959) The cingulate gyrus: additional motor areas and cortical autonomic regulator. J. Comp. Neurol. 112:231-287.
Smith, A.M. (1979) The activity of supplementary motor area during a mantained precision grip. Brain Res. 172:315-327.
Smith, W.K. (1945) The functional significance of the rostral cingular cortex as revealed by its responses to electrical excitation. J. Neurophysiol. 8.241-255.
Talairach, J., and J. Bancaud (1966) The supplementary motor area in man. Int. J. Neurol. 5:330-347.
Tanji, J., and K. Kurata (1979) Neuronal activity in the cortical supplemen- tary motor area related with distal and proximal forelimb movements. Neurosci. Lett. 12:201-206.
Tanji, J., and K. Kurata (1982) Comparison of movement-related activity in two cortical motor areas of primates. J. Neurophysiol. 48:633453.
Toyoshima, K., and H. Sakai (1982) Exact cortical extent of the origin of corticospinal tract (CST) and the quantitative contribution to the CST in different cytoarchitectonic areas. A study with horseradish peroxidase in monkey. J. Hirnforsch. 23:257-269.
Van Buren, J.M., and P. Fedio (1976) Functional representation on the medial aspect of the frontal lobe in man. J. Neurosurg. 44t275-289.
Ward, A.A. (1948) The cingular gyrus: area 24. J. Neurophysiol. 11:13-23. Wiesendanger, M. (1986) Recent developments in studies of the supplemen-
tary motor area of primates. Rev. Physiol. 103:l-59. Wiesendanger, M., J.J. Seguin, and H. Kunzle (1973) The supplementary
motor area-a control system for posture? In R.B. Stein, K.C. Pearson, R.S. Smith, and J.B. Redford (eds): Control of Posture and Locomotion. New York, Plenum, pp. 331-346.
Wise, S.P., and J. Tanji (1981) Supplementary and precentral motor cortex: contrast in responsiveness to peripheral input to the hindlimb area of the unanesthetized monkey. J. Comp. Neurol. 195:433451.
Wong-Riley, M. (1979) Changes in the visual system of monocularly sutured or enucleated cats demonstrable with cytochrome oxidase histochemis- try. Brain Res. 171r11-28.
Woolsey, C.N., P.H. Settlage, D.R. Meyer, W. Sencer, T. Pinto Hamuy, and A.M. Travis (1952) Patterns of localization in precentral and “supplementary” motor areas and their relation to the concept of a premotor area. Res. Publ. Assoc. Nerv. Ment. Dis. 30,238-264.