mps587 - advanced plant biochemistry course fall...
TRANSCRIPT

MPS587 - Advanced Plant Biochemistry Course
Fall Semester 2011
Lecture 8
Phenylpropanoids IV
1. Flavonoids and related phenylpropanoids - overview
2. Flavonoids and anthocyanins
3. Phlobaphenes and proanthocyanidins
4. Isoflavonoids
5. Focus: O-methyltransferases
6. Focus: glycosyltransferases

Flavonoids and related phenylpropanoid-acetate metabolites –biosynthetic overview
(Winkel-Shirley (2001) Plant Physiology 126: 485)
2

Flavonoid nomenclature
3

4
Functional roles of flavonoids(Pollastri & Tattini (2011) Annals of Botany doi: 10.1093/aob/mcr234)
Quercetin derivatives in thenano- to micromolar range mayregulate both the cellular redoxhomeostasis and developmentalprocesses. They may inhibit thephosphorylation of auxin effluxfacilitator proteins located atboth the endoplasmic reticulum(ER) and the plasma membrane(PM). The presence of the wholeset of genes for quercetinbiosynthesis, coupled with theoccurrence of ‘short’ PINproteins at the ER (the site offlavonoid biosynthesis) detectedin liverworts and mosses,suggests ancestral functions forflavonols as developmentalregulators. Quercetin derivativeshave also been shown to tightlycontrol the oxidative stress-induced MAPK activities inanimals, but conclusive evidencefor this functional role in plantsis still lacking (dotted arrows atthe bottom).

5
Benzalacetone in rhubarb(Morita et al. (2010) Proceedings of the National Academy of Sciences in the USA 107: 669)
Flavonoids as developmental regulators
(Taylor & Grotewold (2005) Current Opinion in Plant Biology 8: 317)
5

Core flavonoid/anthocyanidin pathway(Grotewold (2006) Annual Reviews in Plant Biology 57: 761-780)
6
Schematic representation of thebiosynthetic pathway of the mostabundant anthocyanin pigments.The names of the compounds areindicated. The enzyme names, inblack boxes, are CHS, chalconesynthase; CHI, chalconeisomerase; F3H, flavanone 3-hydroxylase; F3′H, flavanone 3′-hydroxylase; F3′5′H, flavanone3′,5′-hydroxylase; DFR,dihydroflavonol 4-reductase;LDOX/ANS, leucoanthocyanidindioxygenase/anthocyanidinsynthase. The A-, B-, and C-ringswith the carbon numbers areindicated in the structurecorresponding to the flavanonenaringenin.

7
Chalcone isomerase(Jez & Noel (2002) Journal of Biological Chemistry 277: 1361)
CHI-catalyzed reaction and active sitearchitecture.
A, overall reaction catalyzed by CHI.
B, view of the active site hydrogen bondnetwork in the CHI·naringenin complex(17). Hydrogen bond interactions (smallspheres) occur within a network centeredon two water molecules (red spheres) thatcontact the flavanone ketone oxygen andthrough interactions of the 7-hydroxylmoiety of the flavanone product withAsn113 and Thr190.
C, proposed cyclization reaction catalyzedby CHI. After nucleophilic attack of the 2′-oxyanion on the α,β-unsaturated doublebond, a water molecule acts as a generalacid to stabilize the enolate, resulting information of a flav-3-en-4-ol intermediatethat tautomerizes into the expectedreaction product.

8
Flavanone 3β-hydroxylase and dihydroflavonol reductase(Springob et al. (2003) Natural Products Reports 20: 288)
Flavanones are converted to dihydroflavonols by ahydroxylation in position 3 catalyzed by flavanone 3β-hydroxylase (F3H). This enzyme is classified as a soluble 2-oxoglutarate-dependent dioxygenase according to itsrequirement of the co-factors 2-oxoglutarate, molecularoxygen, ferrous iron (Fe(II)) and ascorbate. F3H catalyzes thestereospecific hydroxylation of (2S)-naringenin and (2S)-eriodictyol to form (2R,3R)-dihydrokaempferol and (2R,3R)-dihydroquercetin, respectively.
DFR catalyzes the stereospecific conversion of (2R,3R)-dihydroflavonols to (2R,3S,4S)-leucoanthocyanidins DFRrequires NADPH as reducing cofactor and catalyzes thetransfer of the pro-S hydrogen of NADPH to the re-face ofthe 4-keto group of dihydroflavonol.

Anthocyanidin synthase is a oxoglutarate-dependent dioxygenase(Wilmouth et al. (2002) Structure 10: 93)
Proposed mechanism for anthocyanidin synthasecatalyzed C-3 hydroxylation, with the oxidation ofa substrate analogue, dihydroquercetin (DHQ), toquercetin shown as an example. The scheme ismeant to indicate preferred pathways only. It isproposed that the hydroxylation step duringoxidation of the leucoanthocyanidins follows asimilar catalytic scheme. The DHQ substrate isshown in blue, 2OG is in green, and the oxygenatoms from dioxygen are in purple.
9
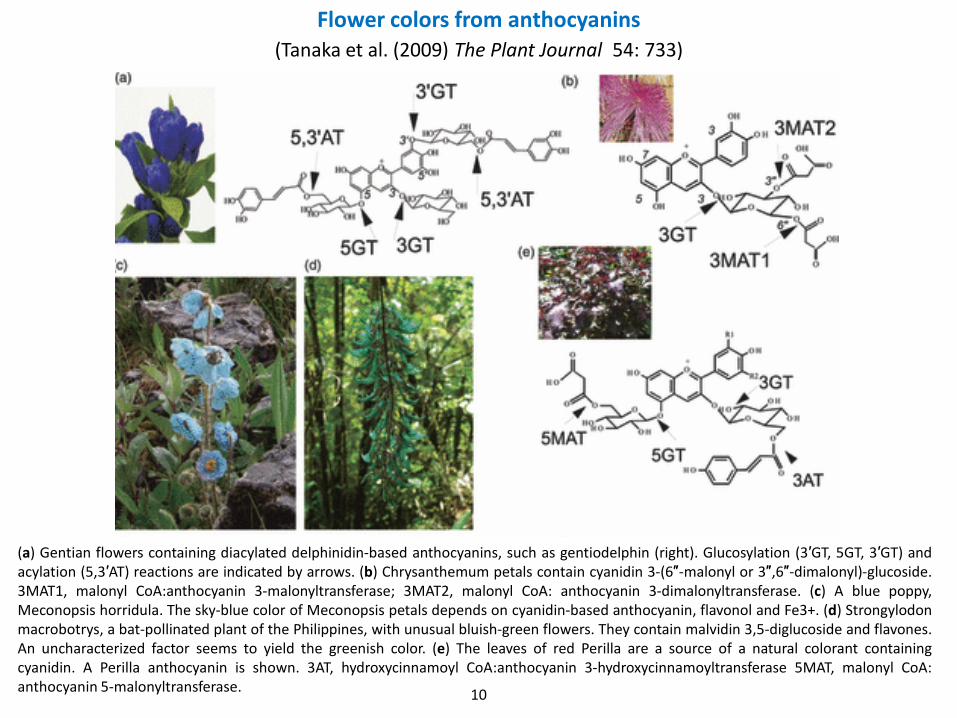
Flower colors from anthocyanins(Tanaka et al. (2009) The Plant Journal 54: 733)
(a) Gentian flowers containing diacylated delphinidin-based anthocyanins, such as gentiodelphin (right). Glucosylation (3′GT, 5GT, 3′GT) andacylation (5,3′AT) reactions are indicated by arrows. (b) Chrysanthemum petals contain cyanidin 3-(6″-malonyl or 3″,6″-dimalonyl)-glucoside.3MAT1, malonyl CoA:anthocyanin 3-malonyltransferase; 3MAT2, malonyl CoA: anthocyanin 3-dimalonyltransferase. (c) A blue poppy,Meconopsis horridula. The sky-blue color of Meconopsis petals depends on cyanidin-based anthocyanin, flavonol and Fe3+. (d) Strongylodonmacrobotrys, a bat-pollinated plant of the Philippines, with unusual bluish-green flowers. They contain malvidin 3,5-diglucoside and flavones.An uncharacterized factor seems to yield the greenish color. (e) The leaves of red Perilla are a source of a natural colorant containingcyanidin. A Perilla anthocyanin is shown. 3AT, hydroxycinnamoyl CoA:anthocyanin 3-hydroxycinnamoyltransferase 5MAT, malonyl CoA:anthocyanin 5-malonyltransferase.
10

Proanthocyanidins - chemistry
11
(Ferreira & Slade (2002) Natural Products Reports 19: 517)

12
(a) The flavonoid biosynthetic enzymesprovide naringenin for catechin production(green). A series of enzyme-catalyzed reactions(purple) leads to the generation of catechinand epicatechins. The stereochemistry of thehydroxyl group in red is determined by thepreceding enzymatic reactions.
(b) Chemical structures of catechin epimers(i.e. (+)-catechin and (−)-epicatechin) andcatechin derivatives (EGC and EGCG).
Proanthocyanidins(Jez & Yu (year) The Plant Journal Vol: Page)

13
Leucoanthocyanidin reductase
Ordering of the VvLAR1 171–175 fragment in the vicinity of the binding site.
(a) Solvent-accessibility surface of the substrate binding site as observed in the binary complex I structure. Red spheresrepresent water molecules. The NADPH molecule is represented using a stick model (carbon atoms are colored green).
(b) Helix 3b interactions with product and coenzyme. The mFo − DFc electron density omit map is contoured at 2.5 σ levelwith NADPH, (+)-catechin, and residues 171–175 excluded from the model. The map contouring residues 171 to 175,which could only be modeled in the ternary complex structure, is colored dark blue. The (+)-catechin molecule isrepresented using a stick model (carbon atoms are colored pink).
(c) Solvent-accessibility surface in the ternary complex structure [same orientation as in (a) and same color code as in (a)and (b)].
(Maugé et al. (2010) Journal of Molecular Biology 397: 1079)

Hydrogen-bonding network in VvLAR1active site and proposed reactionmechanism. (a) VvLAR1 active site and itsinteraction with NADPH and (+)-catechin.Hydrogen bonds are shown in brokenlines; NADPH is represented in green and(+)-catechin is represented in pink. (b)Catalytic mechanism proposed forVvLAR1. B stands for Lys140, whichpromotes the substrate OH7deprotonation via Wat1. A stands forHis122, which is hydrogen-bonded toWat2.
(Maugé et al. (2010) Journal of Molecular Biology 397: 1079)
Leucoanthocyanidin reductase
14

Regulation of flavonoid/pro-anthocyanidin biosynthesis
Broun (2005) Current Opinion in Plant Biology 8: 272-279
15

16
Genetically mappted TT genes involved in proanthocyanidin biosynthesis
tt mutants and their encodingenzymes are as follows: tt4,chalcone synthase; tt5,chalcone isomerase; tt6,flavanone 3-hydroxylase; tt7,flavonoid 3′ hydroxylase; tt3,dihydroflavonol 4-reductase;ban, ANR; tt18 (at the samelocus as tt11), leucocyanidindioxygenase; tt10, laccase-likepolyphenol oxidase; tt12,MATE antiporter; tt15, UDP-Glc:sterol glycosyltransferase;tt19 (at the same locus astt14), GST; aha10, P-type H+-ATPase. tt mutants encodingregulatory proteins are asfollows: tt1, WIK-type zincfinger transcription factor; ttg1(for transparent testa glabra1),WD40 repeat transcriptionfactor; tt2, R2R3 Mybtranscription factor; tt8, basichelix-loop-helix transcriptionfactor; tt16, MADS domaintranscription factor.
(Zhao et al. (2010) Plant Physiology 153: 437)

PA regulatory transcription factors (TT factors)activate PA biosynthesis structural genes (TTs) in thenuclei of seed coat endothelial cells underappropriate conditions.
PA pathway proteins are translocated to thecytosolic side of the ER for synthesis of epicatechin(white circles) and anthocyanins (red circles).
Epicatechin and anthocyanins are readilyglycosylated, and the conjugates are transportedinto the vacuole by MATE (TT12) transporters. Theycould also be loaded into the ER membrane systemor derived membrane vesicles, which aretransported to the central vacuole throughprevacuole compartment (PVC)-dependent vesicletrafficking, or else they could be bound to the TT19GST, which facilitates their transport into the ER,vacuole, or other compartments.
The acidic vacuolar conditions may facilitatenonenzymatic condensation of PA units, or the unitsmay undergo enzymatic condensation catalyzed byTT10 or by yet unidentified proteins (red cylinders).
TT10 could be sorted and targeted within membranevesicles where is may catalyze the condensation ofPA units into oligomers. In the vacuole PA chainelongation could be further catalyzed by TT10 usingepicatechin glucoside and PA oligomers assubstrates.
PAs can also be transported through membranevesicles or other mechanisms to the apoplasticspace, where they are subjected to oxidativepolymerization and further cross-linked with othercell wall components, catalyzed by apoplastic TT10-like polyphenol oxidases.
Proanthocyanidin transport and polymerization
Zhao et al. (2010) Plant Physiology 153: 437
17

18
Phlobaphenes
In maize, at least two flavonoidbiosynthetic pathways have beencharacterized that are regulatedindependently. One pathway results in 3-hydroxy flavonoids such as anthocyaninspurple pigment, whereas the otherpathway produces 3-deoxy flavonoids suchas the phlobaphene red pigmentaccumulated in kernel pericarp, silks andcob.
Whereas the anthocyanin pathway and itsphysiological functions in plant are welldocumented, little is known aboutphlobaphenes. One of the reasons is thatthis family of flavonoids is not synthesizedin Arabidopsis thaliana, the model plant forgenetic and molecular biology. Moreover,breeders selected maize withoutphlobaphenes for food industry (sweetcorn), because it alters the taste of thekernels for consumers. Phlobaphenebiosynthesis results from the oxidation ofcolorless flavan-4-ol monomers orpolymers.
http://www.biosci.ohio-state.edu/pcmb/osu_pcmb/faculty_sites/Erich/projects_phlobaphene.html

19
Isoflavonoids
(Dixon & Ferreira (2002) Phytochemistry 60, 205-211)
http://en.wikipedia.org/wiki/Image:Tofu-beijingchina.jpg
http://en.wikipedia.org/wiki/Image:Lucerne_flowers.jpg
The Soy Health Claim In October 1999, FDA approved a health claim that can be used on labels of soy-based foods to tout their heart-healthy benefits. The agency reviewed research from 27 studies that showed soy protein's value in lowering levels of total cholesterol and low-density lipoprotein (LDL, or "bad" cholesterol).Food marketers can now use the following claim, or a reasonable variation, on their products: "Diets low in saturated fat and cholesterol that include 25 grams of soy protein a day may reduce the risk of heart disease. One serving of (name of food) provides __ grams of soy protein." To qualify for the claim foods must contain per serving: 6.25 grams of soy protein low fat (less than 3 grams) low saturated fat (less than 1 gram) low cholesterol (less than 20 milligrams) sodium value of less than 480 milligrams for individual foods, less than 720 milligrams if considered a main dish, and less than 960 milligrams if considered a meal.

Isoflavonoids - chemistry
20
(Veitch (2009) Natural Products Reports 26: 776)

21
The flavonoid biosynthetic enzymes providenaringenin (or liquiritigenin) for isoflavonoidproduction (green).
An enzyme-catalyzed aryl migration reactiongenerates the core isoflavonoid structure(orange).
Additional modifications, indicated in blue,lead to a variety of isoflavonoid compounds invarious plants (yellow).
Isoflavonoid biosynthesis
(Yu & Jez (2008) The Plant Journal 54: 750)

22
(Steele et al. (1999) Archives of Biochemistry and Biophysics 367: 146)
Isoflavone synthase is a cytochrome P450-dep. monooxygenase catalyzing an aryl migration
Proposed mechanism of aryl migration by CYP93C2. Route a is the aryl migration of (2S)-flavanone to form the mainproduct, 2-hydroxyispflavanone. Route b is the hydroxylation of (2S)-flavanone at C-3 to form the by-product, 3-hydroxyflavanone.

(A) Stereo view of the electron density associated with the 2S,3R-stereoisomer of 2,7,4′-trihydroxyisoflavanone observed crystallographicallyin the HI4′OMT complex.
(B) Stereo view of the electron density associated with 6aR,11aR-6a-hydroxymaackiain observed crystallographically in the HI4′OMT complex.
(C) Close-up view of the HI4′OMT substrate/product binding site with SAHand 2S,3R-2,7,4′-trihydroxyisoflavanone shown. The putative hydrogenbonds are depicted as green spheres.
(D) Close-up view as in (C) illustrating the conformation and location ofbound 6aR,11aR-6a-hydroxymaackiain. For clarity, the active site residuesAsp-269, Phe-328, and Lys-337 that only participate in a hydrogen-bondingnetwork in the HI4′OMT-SAH-6a-hydroxymaackiain complex shown in (D)were omitted in (C).
Complexes of HI4′OMT with 2,7,4′-Trihydroxyisoflavanone and 6aR,11aR-6a-Hydroxymaackiain.
(Liu et al. (2006) The Plant Cell 18: 3656)
23

24
Glycosyltransferases of the flavonoid/isoflavonoid pathway can be promiscuous
Isoflavonoids are often modified byglycosylation with various sugars.Crystal structures of five plant UGTshave been determined.
Among these five UGTs, M. truncatulaUGT85H2 is an (iso)flavonoidglycosyltransferase (Li et al. 2007); M.truncatula UGT78G1 and grape (Vitisvinifera) VvGT1 also recognize someisoflavonoid compounds although theyare able to glycosylate cyanidin to yieldcyanidin 3-O-glycoside and may beinvolved in the biosynthesis ofanthocyanins in vivo; M. truncatulaUGT71G1 also recognizes(iso)flavonoids in spite of its potentialrole in saponin biosynthesis.
(a) Crystal structure of M. truncatulaUGT78G1 with bound UDP; (b) sugardonor binding site and interactionbetween the donor molecule UDP-glucose and the enzyme; (c) acceptorbinding pocket with UDP-2-fluoroglucose (upper left) and acceptorkaempferol (lower right).
(Wang (2010) Functional and Integrative Genomics 11: 13)

Chemical structures of plant metabolites that are commonly glycosylated
(Bowles et al. (2006) Annual Reviews in Plant Biology 57: 567)
Glycosyltransferases of small molecules transfer sugars to a wide range ofacceptors, from hormones and secondary metabolites to biotic and abioticchemicals and toxins in the environment.
The enzymes are encoded by large multigene families and can be identified bya signature motif in their primary sequence, which classifies them as a subsetof Family 1 glycosyltransferases.
The transfer of a sugar onto a lipophilic acceptor changes its chemicalproperties, alters its bioactivity, and enables access to membrane transportersystems.
In vitro studies have shown that a single gene product can glycosylate multiplesubstrates of diverse origins; multiple enzymes can also glycosylate the samesubstrate.
These features suggest that in a cellular context, substrate availability is adetermining factor in enzyme function, and redundancy depends on theextent of coordinate gene regulation.
25