morphology and physiology of local auditory interneurons in the prothoracic ganglion of the...
TRANSCRIPT

THE JOURNAL OF EXPERIMENTAL ZOOLOGY 279:43–53 (1997)
© 1997 WILEY-LISS, INC.
JEZ 824
Morphology and Physiology of Local AuditoryInterneurons in the Prothoracic Ganglion ofthe Cricket Acheta domesticus
OLIVER STIEDL,* ANDREAS STUMPNER, DAVID N. MBUNGU,GORDON ATKINS, AND JOHN F. STOUTDepartment of Biology, Andrews University, Berrien Springs,Michigan 49104
ABSTRACT The omega-neuron 2 (ON2) is an auditory interneuron in Acheta domesticus thatcould play an important role in prothoracic auditory information processing. ON2 neurons wereintracellularly recorded and iontophoretically stained with lucifer-yellow.
The morphology of ON2 is similar to that described in other cricket species. Wholemounts andtransverse sections of prothoracic ganglia show that ON2 branches cover the same regions as theomega-neuron 1 (ON1) branches except that ON2 has crossover branches in the anterior ring tractfrom both the ipsilateral and contralateral side. Auditory responses of ON2 are measured withspiking thresholds of about 60 dB SPL at 16 kHz (courtship song frequency) and about 80 dB SPLat 5 kHz (calling song frequency) under open field and closed field (monaural) stimulation. Openand closed field stimulations show almost identical threshold curves and response magnitudes tosoma-ipsilateral and soma-contralateral stimulation. These data do not support a function of ON2for directional hearing.
In addition to excitation, ON2 receives inhibitory inputs from both ears at 5 and 16 kHz. Sub-threshold inhibition is obvious in responses with open and closed field stimulation at 5 kHz. Re-sponses show mixed excitation and inhibition at intensities above 80 dB SPL at 5 and 16 kHz.These data confirm previous immunohistochemical results showing GABAergic input to ON2 inGryllus bimaculatus. Two newly described local neurons (“LN1’’, “LN2”), which are probably non-spiking and have low 5-kHz thresholds, could be candidates for the inhibitory input to ON2. J.Exp. Zool. 279:43–53, 1997. © 1997 Wiley-Liss, Inc.
The omega-neuron 2 (ON2) is a prothoracic au-ditory interneuron interconnecting both hemigan-glia. While the morphology of the ON2 had beendescribed in Gryllus campestris (Wohlers and Hu-ber, ’82, ’85), little has been published about itsresponse characteristics (Wohlers and Huber, ’82;Schmitz, ’89; Lewis, ’92). For Teleogryllus oceani-cus, Lewis (’92) described ON2 as a high-frequencyneuron with higher sensitivity to 16 and 30 kHzthan to 5 kHz. In comparison, ON1 was most sen-sitive to 5 kHz in G. campestris (Wohlers and Hu-ber, ’78, ’82), G. bimaculatus (Wiese and Eilts, ’85),Teleogryllus commodus (Atkins and Pollack, ’86),and Acheta domesticus (Stumpner et al., ’95).
The ON2 neuron was only shown in crickets.Although a neuron with ON2-like morphology wasstained together with ON1 in Cyphoderris mon-strosa (Haglidae), a “primitive” ensiferan withboth gryllid and tettigoniid features (Mason, ’91;Mason and Schildberger, ’93), there was no infor-mation on its response properties.
Investigations of phonotaxis in crickets revealed
a central role of the prothoracic ganglion in audi-tory information processing (e.g., Atkins et al., ’84,’92; Schildberger and Hörner, ’88; Stout et al., ’91;Pires and Hoy, ’92). Inactivation of a single ON2caused occasional changes in phonotactic orienta-tion by female A. domesticus (Atkins et al., ’84),while Schildberger and Hörner (’88) could not see
Contract grant sponsor: Feodor-Lynen Fellowship of the Alexandervon Humboldt Foundation; Contract Grant sponsor: the National Sci-ence Foundation; Contract Grant number: IBN92-22129; ContractGrant sponsor: Andrews University Faculty Research.
Both the first and second authors contributed equally to this pub-lication.
Oliver Stiedl’s present address is Max-Planck-Institut for Experi-mental Medicine, Department of Molecular Neuroendocrinology, Her-mann-Rein-Str. 3, 37075 Goettingen, FRG.
Andreas Stumpner’s present address is I. Zoological Institute, Uni-versity Goettingen, Berliner Str. 28, 37073 Goettingen, FRG.
David N. Mbungu’s present address is Biology Department, Uni-versity of East Africa, P.O. Box 2500, Eldoret, Kenya.
*Correspondence to: Oliver Stiedl, Max-Planck-Institut for Ex-perimental Medicine, Department of Molecular Neuroendocrinol-ogy, Hermann-Rein-Str. 3, D-37075 Göttingen, FRG. E-mail:[email protected]
Received 15 November 1996; Revision accepted 24 April 1997

44 O. STIEDL ET AL.
a directional effect on phonotaxis during hyper-polarization of one ON2 in G. campestris.
In order to develop a better understanding ofthe possible roles of ON2 within the prothoracicauditory network, the goals of this investigationwere (1) to determine the intraganglionic morphol-ogy of the ON2 neuron, and (2) to describe its neu-ronal response characteristics in order to (3) inferits possible function in prothoracic auditory in-formation processing in A. domesticus.
MATERIALS AND METHODSAnimal Care
Crickets (A. domesticus L.) were obtained fromFluker’s Cricket Farm, Baton Rouge, LA. Theywere raised in the laboratory in large plastic con-tainers at 22°C on a 12:12 h light:dark cycle. Adultfemales were isolated on the day of their imagi-nal moult and kept in 1.5 l plastic containers. Allcrickets were provided with shelter (egg cartons),cricket chow (Purina, St. Louis, MO), and mois-ture (water or fresh potatoes).
Acoustic stimuliResponse characteristics of the ON2 were in-
vestigated using pure tone stimuli resembling thethree syllable pattern (chirp) of the conspecificcalling song with a syllable duration of 25 ms anda syllable period of 50 ms replayed at a rate of1.5 Hz. Intensity calibration (±2 dB) was carriedout with a real time spectrum analyzer (HeathAD-1308, St. Joseph, MI). Frequency/intensity pa-rameters were controlled by computer (DEC PDP11) in the range of 2–17 kHz and 45–95 dB SPLin 5-dB increments. Special attention was drawnto responses to 5 kHz (calling song carrier fre-quency) and 16 kHz (courtship song carrier fre-quency and the frequency which can elicit negativephonotaxis during flight). Neuronal responseswere investigated under two acoustic conditions:(1) For open field stimulation song models wereplayed by piezo loudspeakers (Realistic 40-1379,Radio Shack, Ft. Worth, TX) at a distance of 37cm from the animal in a sound damped cham-ber. (2) For closed field stimulation sound wasdelivered to individual ears with legphones(Atkins et al., ’89; Stumpner et al., ’95) madeof adapted earphones (Realistic 33-977) with aportion of the medial crossover branch of theacoustic trachea removed. The dimensions ofthe chamber allowed the use of frequencies upto at least 20 kHz (see Stumpner et al., ’95;compare Kleindienst et al., ’81). Acoustic/elec-
tric separation of the two legphones was about36 dB SPL.
Neurophysiological experimentsand data analysis
Females (2–28 days following their imaginalmoult) were mounted ventral side up on a waxblock using a beeswax/resin (3/1) mixture. Wings,middle legs, hindlegs, and gut were removed. Theprothoracic ganglion was exposed by removing aportion of the ventral cuticle between the fore-legs. Exposed tissue was covered with saline(Fielden, ’60). The ganglion was supported by asmall spoon and some neck (55a and b) and coxalmuscles were cut to add stability. Intracellular re-cordings were made in the prothoracic ganglionusing borosilicate electrodes (outer/inner diameterin mm: 1.0/0.5 or 1.0/0.78) with a resistance of80–220 MΩ. A silver wire placed in the hemolymphserved as an indifferent electrode. Neuronal re-sponses were amplified using conventional elec-trode amplifiers, monitored on an oscilloscope(Tektronix R5103N, Beaverton, OR), and storedon a DAT recorder (Vetter, 4-channel, Reebers-burg, PA). Recordings were digitized (MacAdiosBoard, GW Instruments, Somerville, MA) andanalyzed (Superscope, GW Instruments, Somer-ville, MA) on a computer (Macintosh, Apple,Cuppertino, CA). The auditory threshold was de-fined by a response of 3 action potentials (APs)per chirp for ON2 and 3 EPSPs/IPSPs for the lo-cal neurons. Latencies were measured fromstimulus onset to the first AP or the beginning ofan EPSP. Lucifer Yellow-filled ON2 neurons weremorphologically identified using epifluorescencemicroscopy (Leitz Laborlux, Wetzlar, Germany).
MorphologyProthoracic ganglia were excised and fixed in
4% buffered formaldehyde (pH 7.2) for 10 minand in 4% formaldehyde in absolute methanolfor 1 hr. The ganglia were then dehydrated inabsolute ethanol for 10–15 min before they werecleared in methyl salicylate for 10 min. Neu-rons were iontophoretically stained (hyperpolar-ization up to –4 nA DC) with 3% Lucifer YellowCH (Sigma, St. Louis, MO, or Aldrich, Milwau-kee, WI) in 0.5 M LiCl. Their morphology wasevaluated in wholemount drawings, transversesections, and by image processing (NIH Image1.47). Ganglia were embedded in Spurr’s me-dium (Spurr, ’69) and sectioned transversally(20 µm) to determine the intraganglionic mor-phology. The nomenclature used to describe the
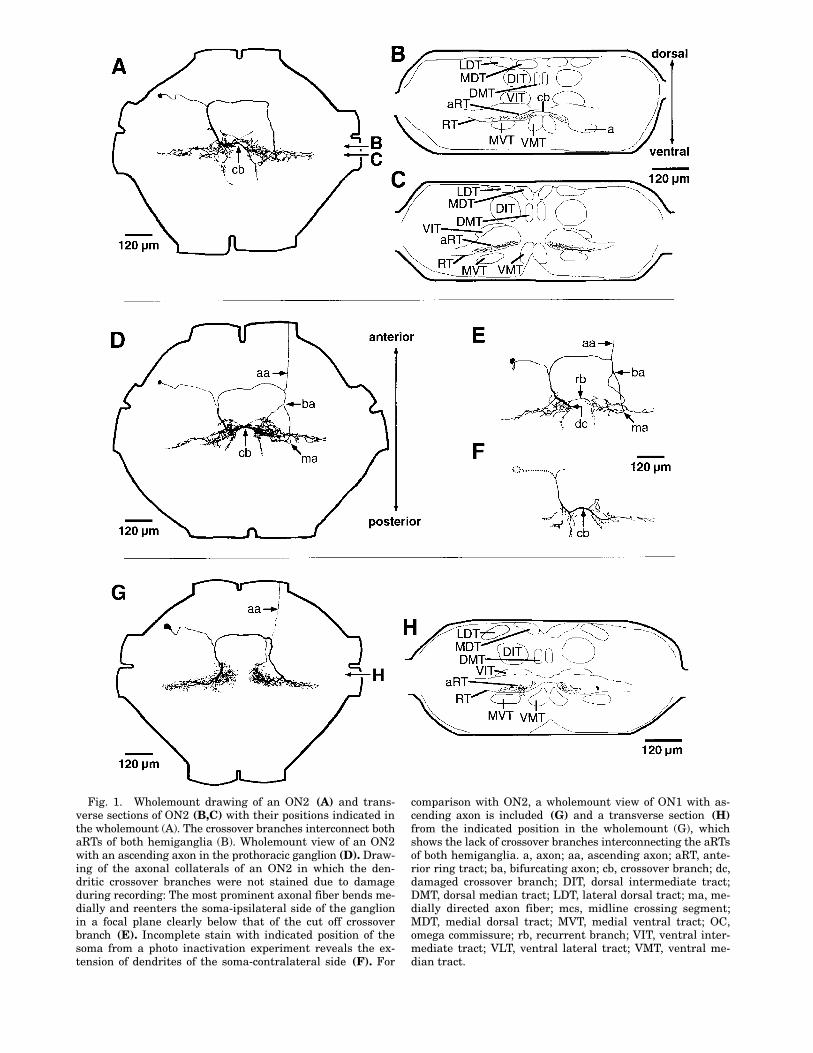
AUDITORY INTERNEURONS IN ACHETA 45
Fig. 1. Wholemount drawing of an ON2 (A) and trans-verse sections of ON2 (B,C) with their positions indicated inthe wholemount (A). The crossover branches interconnect bothaRTs of both hemiganglia (B). Wholemount view of an ON2with an ascending axon in the prothoracic ganglion (D). Draw-ing of the axonal collaterals of an ON2 in which the den-dritic crossover branches were not stained due to damageduring recording: The most prominent axonal fiber bends me-dially and reenters the soma-ipsilateral side of the ganglionin a focal plane clearly below that of the cut off crossoverbranch (E). Incomplete stain with indicated position of thesoma from a photo inactivation experiment reveals the ex-tension of dendrites of the soma-contralateral side (F). For
comparison with ON2, a wholemount view of ON1 with as-cending axon is included (G) and a transverse section (H)from the indicated position in the wholemount (G), whichshows the lack of crossover branches interconnecting the aRTsof both hemiganglia. a, axon; aa, ascending axon; aRT, ante-rior ring tract; ba, bifurcating axon; cb, crossover branch; dc,damaged crossover branch; DIT, dorsal intermediate tract;DMT, dorsal median tract; LDT, lateral dorsal tract; ma, me-dially directed axon fiber; mcs, midline crossing segment;MDT, medial dorsal tract; MVT, medial ventral tract; OC,omega commissure; rb, recurrent branch; VIT, ventral inter-mediate tract; VLT, ventral lateral tract; VMT, ventral me-dian tract.

46 O. STIEDL ET AL.
prominent structures in the sections follows Ty-rer and Gregory (’82).
RESULTSMorphology
ON2 is an omega-shaped neuron, usually con-fined to the prothoracic ganglion (Fig. 1). ON2arborizes in the auditory neuropils in both hemi-ganglia (Fig. 1B,C). The intraganglionic morphol-ogy is very similar to that described for ON1 (Fig.1G,H). Only in ON2 do characteristic arborizingcrossover branches exist, which originated fromthe soma-ipsilateral side and cross in the ante-rior portion of the aRT to the soma-contralateralaRT (Fig. 1A,B,D,F). At least the main crossoverbranch (Fig. 1F) forms a connection of both den-dritic areas (Fig. 1B,F). This connection is notpresent in ON1 (Fig. 1H). The most conspicuousvariability in ON2 occurred in the soma-contralat-eral axon, which bifurcated in 16 (33%) out of 49stained neurons before it entered the aRT (Fig.1D,E). The soma-contralateral axon or one of itsbranches always bent medially towards the mid-line of the ganglion (Fig. 1D,E). In one case, inwhich the main crossover branch, which origi-nated from the soma-ipsilateral side, was notstained due to damage during recording, the soma-contralateral axonal collaterals could be identifiedas bending medially and projecting back into thesoma-ipsilateral hemiganglion (Fig. 1F). In allother staining, the soma-ipsilateral projections ofthe axonal collaterals were masked by the den-dritic arborizations.
An ascending axon was present in 6 out of 41complete stainings of ON2 (Fig. 1D,E). The as-cending axon, which in each case had a very smalldiameter, originated from the anterolateral partof the main axon on the soma-contralateral sideand projected into the neck connective. All ascend-ing axons occurred in 1–7-day-old females. In 12females age 8–28 days no ascending axon wasfound in ON2. Ascending axons have also beenfound in ON1 (Fig. 1G).
NeurophysiologyRecordings from ON2 neurons were usually
made on the soma-ipsilateral side close to the mid-line of the ganglion (Fig. 2). Responses of the ON2recorded at these sites exhibited excitatory andinhibitory postsynaptic potentials (EPSPs and IP-SPs) with attenuated APs. Soma-ipsilateral re-cordings obtained more anterior and lateral fromthe previous site revealed larger AP amplitudesin relation to the EPSP amplitudes (Fig. 2).
ON2 showed excitatory responses to soma-ipsi-lateral and soma-contralateral open field stimu-lation with about 20 dB lower threshold to highfrequency sound (16 kHz) than to low frequencysound (5 kHz) (Fig. 3). The thresholds for soma-ipsilateral and soma-contralateral stimulation didnot differ (Fig. 3B), strongly suggesting binauralexcitation.
Auditory stimulation with legphones revealeda threshold curve similar to open field stimula-tion (Fig. 3A). Again, the thresholds with soma-ipsilateral and soma-contralateral stimulation didnot differ significantly (Fig. 3B), clearly demon-
Fig. 2. Typical ON2 responses obtained from the two in-dicated recording sites of two preparations. All responses wererecorded with soma-ipsilateral open field stimulation of 95dB SPL.

AUDITORY INTERNEURONS IN ACHETA 47
strating excitatory input from both ears. Soma-contralateral input was also confirmed by one ex-periment in which the soma-ipsilateral input wascut and soma-contralateral stimulation producedunchanged response characteristics. Simultaneousstimulation of both ears at intensities abovethreshold revealed in some experiments a slightincrease of the response magnitude with bothstimulation conditions.
The intensity response curve for 5 kHz reached
Fig. 3. Mean threshold curves of the ON2 neuron withsoma-ipsilateral stimulation under open field and closed fieldcondition (A). Threshold values were obtained during differ-
ent stimulation conditions with soma-ipsilateral (ip) andsoma-contralateral (co) stimulation at 5 and 16 kHz (B). Er-ror bars represent standard deviation.
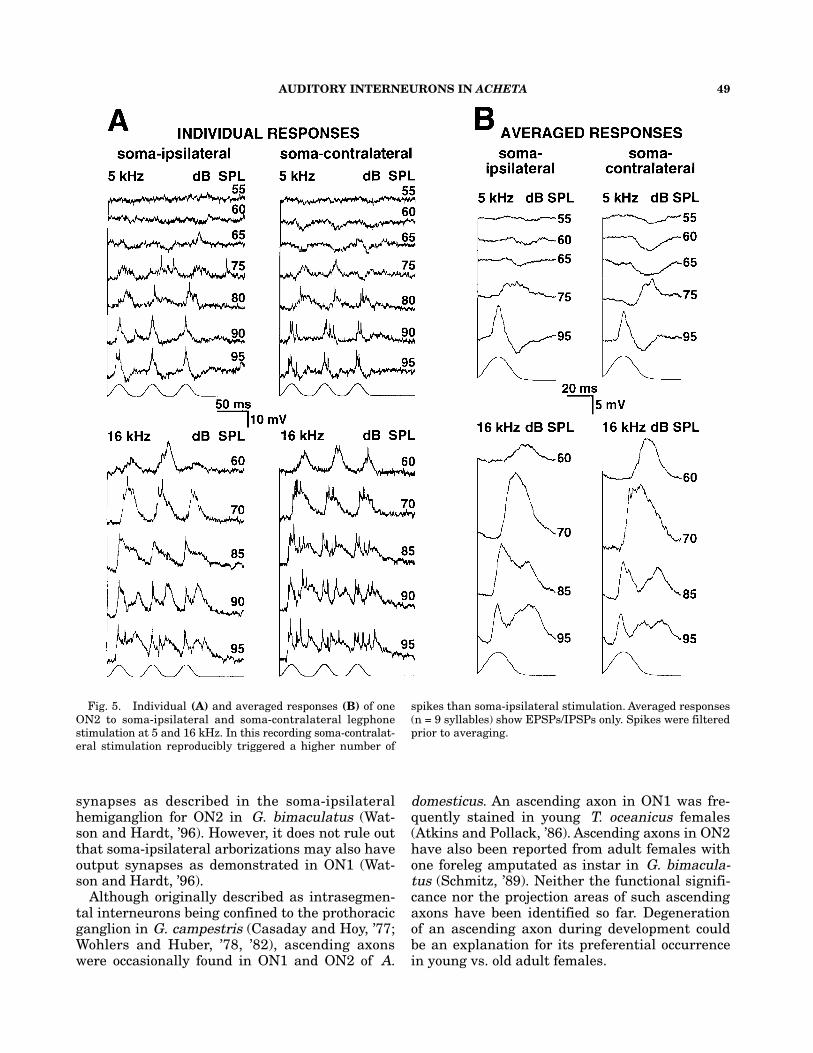
15 APs/chirp at maximum, and at 16 kHz did notexceed 18 APs/chirp at 95 dB SPL (Fig. 4A). Re-sponse latencies of the ON2 did not fall below 15ms (Fig. 4B) and were slightly shorter with 16kHz soma-ipsilateral stimulation than with otherstimuli. At a syllable period of 50 ms, each of thethree syllables forming a chirp was encoded sepa-rately (Figs. 2, 5) except at high intensities at 16kHz. Above 85 dB SPL EPSPs partly merged(Figs. 2, 5A).

48 O. STIEDL ET AL.
In dendritic recordings inhibitory postsynap-tic potentials (IPSPs) were visible. At 5 kHz,IPSPs occurred at subthreshold intensitiesstarting around 55 dB SPL (Fig. 5). EPSPs fol-lowed by IPSPs were visible at 95 dB SPL. In-hibition appeared to be stronger with auditorystimulation delivered from the soma-contralat-eral side (Fig. 5). At 16 kHz, IPSPs were vis-ible at intensities of 80–95 dB SPL, whichseparated excitation into two parts (Fig. 5B).IPSPs occurred with soma-ipsilateral and soma-contralateral stimulation revealing inhibitoryinputs from both ears (Fig. 5).
Local neuronsThe first, so far undescribed local neuron “LN1”)
was recorded under open field conditions andstained two times. It had a large arborization,primarily on the soma-ipsilateral side, whichextended into the soma-contralateral hemi-ganglion (Fig. 6A). “LN1” seemed to be non-spiking. This interneuron received verysensitive 5 kHz excitation with a threshold of55 dB SPL in both recordings and encoded thesyllable structure of calling song models (Fig.6A). At 16 kHz it received strong inhibition witha threshold of 60/70 dB SPL and an IPSP du-ration of more than 20 ms per syllable. EPSPsfollowed by IPSPs were measured at high in-tensities (95 dB SPL, Fig. 6A). At 5 kHz/95 dBSPL response latencies ranged from 9–11.5 ms.“LN1” showed similar responses to both soma-ipsilateral and soma-contralateral stimulation.
The second so far undescribed local neuron(“LN2) was recorded and stained three timesunder open field conditions. It arborized exclu-sively in the soma-ipsilateral hemiganglion andhad a typical “loop-like” branch in the caudalarborization area (Fig. 6B). Action potentialscould not be evoked with sound and/or positiveor negative current injection, thus it seemed tobe a non-spiking interneuron (Fig. 6B). It re-ceived prominent 5 kHz excitation with athreshold of 60 dB SPL in all three recordings.The 5 kHz threshold was identical to that ofthe ON1-neuron in each individual animal.At 16 kHz “LN2” received excitation only at in-tensities from 85–95 dB SPL (Fig. 6B). Theshortest latencies measured at the highest in-tensities ranged from 7.5–11 ms. With increas-ing depolarizing currents, sound activatedEPSPs became smaller and were inverted aboveabout 1 nA current. In addition, “LN2” was excitedby wind stimuli and inhibited by vibration (datanot shown).
DISCUSSIONMorphology
ON2 has one of the most complex morphologiesamong prothoracic auditory interneurons knownfrom crickets so far. The intraganglionic morphol-ogy is identical to that described in G. campestris(Wohlers and Huber, ’85; Watson and Hardt, ’96).Partial stainings revealed that both dendriticbranches and axonal collaterals cross the midlineand arborize in both hemiganglia (Fig. 1E). Thisstructure could explain the presence of output
Fig. 4. Effect of increasing intensity on response magni-tude (A) and response latency (B) of two ON2 recordings.Response magnitudes (n = 3) were measured during open fieldstimulation and response latencies (n = 3) were measuredduring closed field stimulation at 5 and 16 kHz. Data aregiven as mean values ± standard deviation. ipsi, soma-ipsi-lateral stimulation; contra, soma-contralateral stimulation.
AUDITORY INTERNEURONS IN ACHETA 49
synapses as described in the soma-ipsilateralhemiganglion for ON2 in G. bimaculatus (Wat-son and Hardt, ’96). However, it does not rule outthat soma-ipsilateral arborizations may also haveoutput synapses as demonstrated in ON1 (Wat-son and Hardt, ’96).
Although originally described as intrasegmen-tal interneurons being confined to the prothoracicganglion in G. campestris (Casaday and Hoy, ’77;Wohlers and Huber, ’78, ’82), ascending axonswere occasionally found in ON1 and ON2 of A.
domesticus. An ascending axon in ON1 was fre-quently stained in young T. oceanicus females(Atkins and Pollack, ’86). Ascending axons in ON2have also been reported from adult females withone foreleg amputated as instar in G. bimacula-tus (Schmitz, ’89). Neither the functional signifi-cance nor the projection areas of such ascendingaxons have been identified so far. Degenerationof an ascending axon during development couldbe an explanation for its preferential occurrencein young vs. old adult females.
Fig. 5. Individual (A) and averaged responses (B) of oneON2 to soma-ipsilateral and soma-contralateral legphonestimulation at 5 and 16 kHz. In this recording soma-contralat-eral stimulation reproducibly triggered a higher number of
spikes than soma-ipsilateral stimulation. Averaged responses(n = 9 syllables) show EPSPs/IPSPs only. Spikes were filteredprior to averaging.

50 O. STIEDL ET AL.
Neurophysiology
The small diameter of ON2 and the resultingdifficulty to obtain long-lasting intracellular re-cordings (Lewis and Harrison, ’87; Lewis, ’92) isthe most probable reason for the small amount ofphysiological data available for ON2 and had prob-ably led to the hypothesis that ON2 might be anon-spiking neuron (Lewis and Harrison, ’87).Our recordings (Fig. 3) showed that ON2 of A.domesticus clearly is a spiking neuron. APs weremore attenuated when recorded in the crossoverbranch close to the midline of the prothoracic gan-glion as was also suggested by Lewis (’92) for T.oceanicus. A recording in ON2 made on the soma-contralateral side in T. commodus (Atkins, unpub-lished data) revealed APs without underlyingEPSPs as shown in ON1 in G. bimaculatus(Wohlers and Huber, ’78).
In G. campestris ON2 was described as a rela-
Fig. 6. Wholemount views and responses of the local pro-thoracic neurons “LN1” (A) and “LN2” (B) recorded with openfield stimulation. contra, soma-contralateral stimulation;
depol, depolarization; hyperpol, hyperpolarization; ipsi, soma-ipsilateral stimulation; L, “loop-like” branch.
tively frequency-unselective interneuron about 10dB more sensitive to 4–5 kHz than to 16 kHz(Wohlers and Huber, ’82). This result was con-firmed in monaural animals of G. bimaculatus(Schmitz, ’89) and in intact crickets (Watson andHardt, ’96). In contrast to these data, in T.oceanicus ON2 was much more sensitive to highfrequency sound (16 kHz) than to 5 kHz (Lewis,’92). We can clearly show that in A. domesticusON2 is about 20 dB more sensitive to 16 kHz thanto 5 kHz. A slightly stronger response to soma-ipsilateral stimulation was described in G. bi-maculatus (Schmitz, ’89) with intensity responsedynamics comparable to the ON2 responses in A.domesticus described here. Binaural input to ON2was already shown in G. bimaculatus by Wohlersand Huber (’82) and confirmed by Schmitz (’89)in animals with monaural input. Simultaneousstimulation from both sides frequently resultedin an increase of the EPSP amplitude in A.

AUDITORY INTERNEURONS IN ACHETA 51
domesticus as also found in G. campestris (Wohlersand Huber, ’82). In G. bimaculatus ON2 showeda less precise coding (merged EPSPs) of the syl-lable structure of chirps eliciting phonotaxis thanON1 (Wohlers and Huber, ’82; Schmitz, ’89). InA. domesticus the syllables of a chirp elicitingphonotaxis with a syllable period of 50 ms werecoded separately. Only at 95 dB SPL/16 kHzEPSPs of the syllables started to merge. Unfortu-nately, there is no information on binaural inputsand temporal pattern coding in T. commodus.
In contrast to all previous neurophysiological in-vestigations on ON2 (Wohlers and Huber, ’82;Schmitz, ’89; Lewis, ’92) our physiological dataclearly demonstrated inhibitory input to ON2 ofA. domesticus (Fig. 5). Inhibitory input was mostprominent at low frequencies with soma-contralat-eral stimulation. As ON2 receives inhibitory andexcitatory input from both sides, at least two in-puts must converge on each auditory input sideof ON2.
It has been demonstrated in G. bimaculatus byimmunohistochemical means that ON2 receivedsoma-ipsilateral and soma-contralateral GABAer-gic inputs and also formed output synapses onthe soma-ipsilateral and soma-contralateral side(Watson and Hardt, ’96). In A. domesticus, the au-ditory interneurons L1 (homologous to AN1 inGryllus: Stout et al., ’88; Stumpner et al., ’95) andON1 are the only identified auditory neurons sofar that could be candidates providing inhibitoryinput to ON2 starting at 55 dB SPL at 5 kHz.Electrophysiological results for inhibitory process-ing of auditory information in crickets exist onlyfor the contralateral inhibition by ON1 (Selver-sten et al., ’85; Wiese and Eilts, ’85; Stumpner etal., ’95). Data obtained by electron microscopy inG. bimaculatus revealed output synapses on thesoma-ipsilateral side of ON1 (Watson and Hardt,’96) suggesting it could provide this 5-kHz sub-threshold inhibition. GABA-immunoreactivitycould not be demonstrated in ON1 (in Gryllus:Spörhase-Eichmann et al., ’89; Watson and Hardt,’96; in Acheta: Lang and Stumpner, unpublisheddata) but there is evidence for histamine as theinhibitory transmitter in ON1 of G. bimaculatus(Skiebe et al., ’90). In addition, no prothoracic out-put synapses were found in AN1 (Hardt and Wat-son, ’94). Other known auditory interneurons(DN1, DN4, TN1, and TN5; in T. oceanicus: At-kins and Pollack, ’87a) with processes partly over-lapping with those of ON2 in the aRT (DN1, TN1,in T. oceanicus: Atkins and Pollack, ’87b) or soundsensitive DUM neurons (in G. bimaculatus: Gras
et al., ’90) were not candidates for this inhibition.They either had a relatively high 5-kHz thresh-old, weak and/or habituating response character-istics, and/or extremely long latencies. The localinterneurons (Stumpner, ’95) here described as“LN1” and “LN2” have been included in this study,because they indicate that other auditory inter-neurons exist in the prothoracic ganglion. In ad-dition, such neurons are necessary in order toexplain the complex response characteristics de-scribed for ON2 in A. domesticus (Fig. 5). The lo-cal neurons “LN1” and “LN2” could provide the5-kHz subthreshold inhibition as demonstrated inON2 (Fig. 5), as their 5-kHz sensitivity matchedthe threshold of ON1 (Fig. 6) and they might beGABAergic. These two local neurons were de-scribed only in a very preliminary way as indi-cated by their names in quotation marks. It willrequire further experiments to more completelycharacterize these neurons and their contributionto prothoracic auditory information processing inA. domesticus.
Response latencies in ON2 seldom fell below 15ms. ON1 and L1 (considered to be first order in-terneurons) responded to auditory stimulationwith latencies as short as 11 ms when stimulatedat 95dB SPL (Stumpner et al., ’95). A possible ex-planation for the slightly increased latencies inON2 could be its inhibitory inputs. Recordings inG. bimaculatus showed response latencies in ON2and ON1 that are as short as in AN1 and AN2 at4.5 kHz measured at approximately 15 dB abovethe threshold of each neuron (Wohlers and Hu-ber, ’82).
Possible roles of ON2We have demonstrated that ON2 receives exci-
tatory and inhibitory inputs from both ears. Themorphological basis for the physiological datamight be the extension of dendritic arborizationsinto both hemiganglia. Axonal output branchesmost likely occur in both hemiganglia as well. Thealmost identical threshold values obtained withsoma-ipsilateral and soma-contralateral open andclosed field stimulation in A. domesticus (Fig. 4)and G. bimaculatus do not support a function ofON2 for directional hearing as discussed by Horse-man and Huber (’94).
Previous investigations revealed that the syl-lable period within a chirp determined the pho-notactic responsiveness of female A. domesticus(Stout and McGhee, ’88) with the L3-interneuron(probably homologous to AN2) possibly involved(Atkins et al., ’89; Henley et al., ’92). ON2 may

52 O. STIEDL ET AL.
play a role in the neuronal circuit that regulatessyllable pattern processing. We suggest that ON2,“LN1”, and “LN2” are involved in inhibitory pat-terning of the responses of ascending auditory neu-rons. However, ON2 neither showed GABAergic(Watson and Hardt, ’96) nor serotonergic immunore-activity (Hardt et al., ’94) in G. bimaculatus.
Another possible role of ON2 might be to func-tion as a local “high-frequency neuron” mediatingfrequency specific inhibition or excitation. Soma-contralateral high-frequency (16 kHz) inhibitionfrom 65 dB SPL on was described in the L1(Stumpner et al., ’95). Due to its threshold, ON2is a likely candidate for this high-frequency inhi-bition and might also contribute to high-intensityinhibition at 5 kHz. With regard to other high-frequency functions, ON2 is a less likely candi-date for being involved in bat avoidance behavior(Moiseff et al., ’78; Nolen and Hoy, ’86) due to itslack of directionality. However, ON2 could be in-volved in courtship song processing (Lewis, ’92)in which directional information is not requiredbecause courtship song is emitted at close rangeonly (Alexander, ’61).
ACKNOWLEDGMENTSWe thank John Henley and James Ramseier for
supplying some threshold values, and WolfgangRössler and Martin Hardt for helpful discussions.This work was supported by a Feodor-Lynen Fel-lowship of the Alexander von Humboldt-Founda-tion to O.S. and a National Science FoundationPostdoctoral Fellowship to A.S., a National Sci-ence Foundation grant (IBN92-22129), and An-drews University Faculty Research grants to G.A.and J.S. The experiments were performed accord-ing to the “Principles of Animal Care,” publica-tion No. 86-23, revised 1985, of the NationalInstitute of Health, and according to the currentlaws of Michigan.
LITERATURE CITEDAlexander, R.D. (1961) Aggressiveness, territoriality and
sexual behaviour in field crickets (Orthoptera: Gryllidae).Behaviour, 17:130–223.
Atkins, G., and G.S. Pollack (1986) Age-dependent occurrenceof an ascending axon on the omega neuron of the cricket,Teleogryllus oceanicus. J. Comp. Neurol., 243:527–534.
Atkins, G., and G.S. Pollack (1987a) Response properties ofprothoracic, interganglionic, sound-activated interneuronsin the cricket Teleogryllus oceanicus. J. Comp. Physiol. A,161:681–693.
Atkins, G., and G.S. Pollack (1987b) Correlations betweenstructure, topographic arrangement, and spectral sensitiv-ity of sound-sensitive interneurons in crickets. J. Comp.Neurol., 266:398–412.
Atkins, G., S. Ligman, F. Burghardt, and J.F. Stout (1984)Changes in phonotaxis by the female cricket Achetadomesticus L. after killing identified acoustic interneurons.J. Comp. Physiol. A, 154:795–804.
Atkins, S., G. Atkins, M. Rhodes, and J.F. Stout (1989) Influ-ence of syllable period on song encoding properties of anascending auditory interneuron in the cricket Achetadomesticus. J. Comp. Physiol. A, 165:827–836.
Atkins, G., J. Henley, R. Handysides, and J. Stout (1992)Evaluation of the behavioral roles of ascending auditory in-terneurons in calling song phonotaxis by the female cricket(Acheta domesticus). J. Comp. Physiol. A, 170:363–372.
Casaday, G.B., and R.R. Hoy (1977) Auditory interneurons inthe cricket Teleogryllus oceanicus: Physiological and ana-tomical properties. J. Comp. Physiol. A, 121:1–13.
Fielden, A. (1960) Transmission through the last abdominalganglion of the dragonfly nymph, Anax imperator. J. Comp.Exp. Biol., 37:832–844.
Gras, H., M. Hörner, L. Runge, and F.-W. Schürmann (1990)Prothoracic DUM neurons of the cricket Gryllus bimacu-latus: Responses to natural stimuli and activity in walk-ing behavior. J. Comp. Physiol. A, 166:901–914.
Hardt, M., and A.H.D. Watson (1994) Distribution of synapseson two ascending interneurones carrying frequency-specificinformation in the auditory system of the cricket: Evidencefor GABAergic input. J. Comp. Neurol., 345:481–495.
Hardt, M., U. Spörhase-Eichmann, and H. Agricola (1994) Se-rotonin immunoreactivity of identified sensory interneuronsof the cricket. Soc. Neurosci. Abstr. 20:913. (Abstr. 378.4).
Henley, J., J. Greenwood, J. Stout, and G. Atkins (1992) Age-correlated changes and juvenile hormone III regulation ofthe syllable period specific responses of the L3 auditory in-terneurons in the cricket, Acheta domesticus. J. Comp.Physiol. A, 170:373–378.
Horseman, G., and F. Huber (1994) Sound localisation in crick-ets. II. Modelling the role of a simple neural network inthe prothoracic ganglion. J. Comp. Physiol. A, 175:399–413.
Kleindienst, H.U., U.T. Koch, and D.W. Wohlers (1981) Analy-sis of the cricket auditory system using a closed sound field.J. Comp. Physiol. A, 141:283–296.
Lewis, B. (1992) Processing of auditory signals in the CNS ofOrthoptera. In: The Evolutionary Biology of Hearing. D.B.Webster, R.R. Fay and A.N. Popper, eds. Springer, Heidel-berg, pp. 95–114.
Lewis, B., and L. Harrison (1987) Responses of central audi-tory neurones in crickets. Proc. 6th Int. Meet. Insect SoundVibration. Gamle Avernæs (Abstr. 27).
Mason, A.C. (1991) Hearing in a primitive ensiferan: The au-ditory system of Cyphoderris monstrosa (Orthoptera:Haglidae). J. Comp. Physiol. A, 168:351–363.
Mason, A.C., and K. Schildberger (1993) Auditory interneu-rons in Cyphoderris monstrosa (Orthoptera: Haglidae). J.Comp. Physiol. A, 171:749–757.
Moiseff, A., G.S. Pollack, and R.R. Hoy (1978) Steering re-sponses of flying crickets to sound and ultrasound: Mateattraction and predator avoidance. Proc. Natl. Acad. Sci.U.S.A., 75:4052–4056.
Nolen, T.G., and R.R. Hoy (1986) Phonotaxis in flying crick-ets. 1. Attraction to the calling song and avoidance of bat-like ultrasound are discrete behaviors. J. Comp. Physiol A,159:423–439.
Pires, A., and R.R. Hoy (1992) Temperature coupling in cricketacoustic communication. II. Localization of temperature ef-fects on song production and recognition networks in Gryl-lus firmus. J. Comp. Physiol. A, 171:79–92.

AUDITORY INTERNEURONS IN ACHETA 53
Schildberger, K., and M. Hörner (1988) The function of audi-tory neurons in cricket phonotaxis. I. Influence of hyperpo-larization of identified neurons on sound localization. J.Comp. Physiol. A, 163:621–631.
Schmitz, B. (1989) Neuroplasticity and phonotaxis in mon-aural adult female crickets (Gryllus bimaculatus de Geer).J. Comp. Physiol. A, 164:343–358.
Selversten, A., H.-U. Kleindienst, and F. Huber (1985) Syn-aptic connectivity between cricket auditory interneuronsas studied by selective photoinactivation. J. Neurosci.,5:1283–1292.
Skiebe, P., B.J. Corrette, and K. Wiese (1990) Evidence thathistamine is the inhibitory transmitter of the auditory in-terneuron ON1 of crickets. Neurosci. Lett., 116:361–366.
Spörhase-Eichmann, U., M. Hanssen, and F.-W. Schürmann(1989) GABA-immunoreactive neurones in the prothoracicganglion of the cricket Gryllus bimaculatus. In: Proceed-ings of the 17th Göttingen Neurobiology Conference. N.Elsner and W. Singer, eds. Thieme, Stuttgart, Germany(Abstr. 55).
Spurr, A.R. (1969) A low-viscosity epoxy-resin embedding me-dium for electron microscopy. J. Ultrastruct. Res., 26:31–43.
Stout, J.F., and R.W. McGhee (1988) Attractiveness of themale Acheta domesticus calling song to females. II. The rela-tive importance of syllable period, intensity and chirp rate.J. Comp. Physiol. A, 164:277–287.
Stout, J.F., C.H. DeHann, J.C. Hall, and M. Rhodes (1988)Processing of calling songs by an L-shaped neuron in theprothoracic ganglion of the female cricket, Acheta domes-ticus. Physiol. Entomol., 13:89–101.
Stout, J.F., G. Atkins, and D. Zacharias (1991) Regulation ofcricket phonotaxis through hormonal control of the thresh-old of an identified auditory neuron. J. Comp. Physiol. A,169:765–772.
Stumpner, A. (1995) Some new local and descending audi-tory neurons in the prothoracic ganglion of crickets andbushcrickets. In: Proceedings of the 23rd Göttingen Neuro-biology Conference. N. Elsner and M. Heisenberg, eds.Thieme, Stuttgart, Germany (Abstr. 274).
Stumpner, A., G. Atkins, and J.F. Stout (1995) Processing ofunilateral and bilateral auditory inputs by the ON1 andL1 interneurons of the cricket Acheta domesticus and com-parison to other cricket species. J. Comp. Physiol. A,177:379–388.
Tyrer, N.M., and G.E. Gregory (1982) A guide to the neu-roanatomy of locust subesophageal and thoracic ganglia.Phil. Trans. R. Soc. London B, 297:91–123.
Watson, A.H.D., and M. Hardt (1996) Distribution of syn-apses on two local auditory interneurones, ON1 and ON2,in the prothoracic ganglion of the cricket: Relations withGABA-immunoreactive neurones. Cell Tissue Res., 283:231–246.
Wiese, K., and K. Eilts (1985) Evidence for the matched fre-quency dependence of bilateral inhibition in the auditorypathway of Gryllus bimaculatus. Zool. Jb. Physiol.,89:181–201.
Wohlers, D., and F. Huber (1978) Intracellular recording andstaining of cricket auditory interneurons (Gryllus campestrisL., Gryllus bimaculatus DeGeer). J. Comp. Physiol. A,127:11–28.
Wohlers, D., and F. Huber (1982) Processing of sound sig-nals by six types of neurons in the prothoracic ganglion ofthe cricket, Gryllus campestris L. J. Comp. Physiol. A,146:161–173.
Wohlers, D., and F. Huber (1985) Topographical organiza-tion of the auditory pathway within the prothoracic gan-glion of the cricket Gryllus campestris L. Cell Tissue Res.,239:555–565.