morpho-genetic variation analysis and in vitro …
TRANSCRIPT

MORPHO-GENETIC VARIATION ANALYSIS AND IN VITRO PROPAGATION OF ROSA CENTIFOLIA
By
GULZAR AKHTAR M. Sc. (Hons.) Horticulture
2003-ag-1849
A THESIS SUBMITTED IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF
DOCTOR OF PHILOSOPHY IN
HORTICULTURE
INSTITUTE OF HORTICULTURAL SCIENCES,
FACULTY OF AGRICULTURE,
UNIVERSITY OF AGRICULTURE, FAISALABAD
PAKISTAN
2014

In The Name Of
The Most Beneficent The Most Merciful

Declaration
I hereby declare that the contents of the thesis “Morpho-Genetic Variation analysis and in
vitro propagation of Rosa centifolia” are the product of my own research and no part has
been occupied from any published sources (except the references, standard mathematical or
genetic model/ equations/formula/ protocol etc). I further declare that this work has not been
submitted for the award of any other diploma/degree. The university may take action if the
information provided is found inaccurate at any stage. In case of any default, the scholar will
be proceeded against as per HEC plagiarism policy.
________________
GULZAR AKHTAR
2003-ag-1849
M.Sc. (Hons) Horticulture

To
The Controller of Examinations, University of Agriculture, Faisalabad.
“We, the members of supervisory committee, certify that the contents and form of thesis submitted by Mr. Gulzar Akhtar, 2003-ag-1849, have been found satisfactory and recommend that it be processed for evaluation, by the External Examiner(s) for the award of degree”
Supervisory Committee
1. Chairman _______________________ (Dr. Muhammad Aslam Khan)
2. Member _______________________
(Dr. Muhammad Jafar Jaskani)
3. Member ________________________ (Dr. Muhammad Ashfaq)

This humble effort is
Dedicated To
My Father and Mother
By virtue of whose prayers I have been able to
reach at this position

ACKNOWLEDGEMENTS
All humble praise and thanks for ALMIGHTY ALLAH who is the most kindly Merciful and Durood e Pak on the Holy Prophet, Muhammad (Peace Be Upon Him) ( ھ ھ آ و علی سلمّ و لل .who is a Mercy and source of guidance for all humanity ( ا یص I have been indebted in the research work and preparation of this thesis to my supervisor, Dr. Muhammad Aslam Khan, Professor, Institute of Horticultural Sciences (IHS), whose kindness and academic experience have been invaluable to me. I am extremely grateful to Dr. Muhammad Jafar Jaskani, Professor IHS and Dr. Muhammad Ashfaq, Professor, Department of Plant Breeding and Genetics who always kept an eye on my work as member of my supervisory committee. I would like to thank Dr. Adnan Younis, Dr. Atif Riaz and Dr. Iftikhar Ahmad Assistant Professor HIS, for their help and guidance throughout my research work and thesis write-up. I also need to express my deeply-felt thanks to Dr. Muhammad Aslam Pervez, Director IHS and Dr. M. Qasim, Professor IHS, Professor IHS, for their warm encouragement and thoughtful guidance. How can I forget my Co-supervisor, Prof. Dr. David H. Byrne and Natalie Anderson (Research Associate) for their guidance and cooperation during my work on Microsatellites at Department of Horticultural Sciences, Texas A & M University, Texas, USA. I also acknowledge with thanks financial support from The Higher Education Commission (HEC) of Pakistan for giving me opportunity of Indigenous 5000 Fellowship scholarship and funds for International Research Support Initiative Program (IRSIP) to conduct research at Texas A & M University, Texas, USA as a visiting scientist. The informal support and encouragement of many friends has been indispensable, and I would like particularly to acknowledge the contribution of Mr. Ahsan Akram, Dr. Muhammad Nadeem, Mr. Yasir Sajad, Mr. Asim Mehmood, Dr. Abdul Kareem, Mr. Faheem Nawaz, Mr. Manan Shah, Mr. Adnan Bukhari, Mr. Muhammad Rashid Abbasi, Mr. Imran Muaavia, Mr. Muhammad Shafique, Mr. Ockert, Tim Hartman, Suheb Mohammed, Qianni Dong, Mr. Jake, Sabeeh Ahmad and (Texas A&M University, College Station, TX). My father, mother and Taya jaan have been a constant source of support, emotional, moral and of course financial and this thesis would certainly not have existed without them. It is thanks to my uncle who encouraged and supported me to get into PhD but he left me forever when I was doing my research work in Texas A & M University, USA. May Allah keeps his soul in peace (Ameen). I pay my respect to my beloved brothers Shahzad Akhtar, Shahbaz Akhtar, Waqar Akhtar, Sadam Hussian my cusion Muhammad Nadeem, Muhammad Waseem, Muhammad Kaleem my loving, sincere and well wisher sister for the sacred prayers and endless kind behavior. May Allah bless my parents and family members with good health and prosperity.
Gulzar Akhtar

C O N T E N T S
Chapter Title Page
1 INTRODUCTION 1
2 REVIEW OF LITERATURE 7
2.1 Morphological Analysis 7
2.2 Molecular Analysis 9
2.3 In vitro propagation 11
3 MATERIALS AND METHODS 18
3.1 Phenotypic plasticity among the genotypes of Rosa centifolia
18
3.1.1 Climate of Pakistan 18
3.1.2 Areas selected for morphological study 18
3.1.3 Layout of experiment 21
3.1.3.1 Genotypes of Rosa centifolia from Pakistan 21
3.1.3.2 Genotypes of Rosa centifolia from USA 21
3.1.4 Morphological Parameters 21
3.1.4.1 Plant parameters 22
3.1.4.2 Leaf parameters 23
3.1.4.3 Flower parameters 25
3.1.5 Statistical Analysis 26
3.2 Genetic diversity among genotypes of Rosa centifolia 27
3.2.1 Plant material from Pakistan 27
3.2.2 Plant material from USA 27
3.2.3 Rosa damascena samples 27
3.2.4 Primers 29
3.2.5 DNA extraction protocol 29
3.2.6 DNA Quantification 30
3.2.7 PCR (Polymerase Chain Reaction) Amplification 30
3.2.8 Gel electrophoresis 30

3.2.9 Data analysis 31
3.3 IN VITRO PROPAGATION OF ROSA CENTIFOLIA 31
3.3.1 Plant Material 31
3.3.2 Media 31
3.3.3 Preparation of stock solutions 31
3.3.4 Stock solutions for growth regulator 32
3.3.5 Preparation of media 32
3.3.6 Sterilization of explant 33
3.3.7 Inoculation 33
3.3.8 Sub culture 34
3.3.9 Hardening 34
3.3.10 Collection of Data 34
3.3.10.1 Number of days for shoot initiation 34
3.3.10.2 Number of shoots 34
3.3.10.3 Number of leaves per shoot 34
3.3.10.4 Shoot length 34
3.3.10.5 Number of days to initiate roots 35
3.3.10.6 Number of roots 35
3.3.10.7 Root length 35
3.3.11 Data analysis 35
4 RESULTS 36
4.1 Phenotypic plasticity among the genotypes of Rosa centifolia
36
4.1.1 Cluster analysis for different morphological characters of genotypes of Rosa centifolia from Pakistan
36
4.1.2 Principle component analysis for different morphological characters of genotypes of Rosa centifolia
36
4.1.3 Correlation between morphological parameters of Rosa centifolia genotypes from Pakistan
36
4.1.4 Cluster analysis for different morphological characters of different varieties of Rosa centifolia from USA
40
4.1.5 Principle component analysis for different morphological characters of different varieties of Rosa centifolia from USA
40
4.1.6 Correlation between morphological parameters of Rosa centifolia varieties from USA
40

4.1.7 Cluster analysis for different morphological characters of different varieties of Rosa centifolia from Pakistan and USA
44
4.2 Genetic diversity among genotypes of Rosa centifolia 48
4.2.1 Genetic analysis of Rosa centifolia from Pakistan 48
4.2.2 Genetic analysis of Rosa centifolia from Pakistan and USA 53
4.2.3 Genetic analysis of Rosa centifolia and Rosa damascena from Pakistan, USA and Iran
59
4.3 In vitro propagation of Rosa centifolia through axillary buds
64
4.3.1 Number of days to initiate shoots 64
4.3.2 Number of shoots 64
4.3.3 Shoot length 65
4.3.4 Number of leaves 65
4.3.5 Number of days to induce roots 65
4.3.6 Number of roots 65
4.3.7 Root length 65
5 DISCUSSION 75
5.1 Phenotypic plasticity among the genotypes of Rosa centifolia
75
5.2 Genetic diversity among genotypes of Rosa centifolia 77
5.2.1 Genetic analysis of different genotypes of Rosa centifolia from Pakistan
77
5.2.2 Genetic analysis of different genotypes of Rosa centifolia from Pakistan and USA
78
5.2.3 Genetic analysis of different genotypes of Rosa centifolia and Rosa damascena from Pakistan, USA and Iran
79
5.3 In vitro propagation of Rosa centifolia through axillary buds
80
5.3.1 In vitro shoot formation of Rosa centifolia 80
5.3.2 In vitro root induction of Rosa centifolia 82
SUMMARY 84
LITARATURE CITED 87

List of Tables
No. Title Page
3.1 Different site of data collection Punjab, Pakistan 19
3.2 Analysis of water samples collected from different sites of Punjab, Pakistan
19
3.3 Analysis of soil samples collected from different sites of Punjab, Pakistan
19
3.4 Roses genotypes used for SSR analysis 28
3.5 List of primers 29
3.6 Ingredients of MS media 32
3.7 Treatments for shoot formation 33
3.8 Treatments for root formation 33
4.1 Correlation between morphological parameters of Rosa centifolia genotypes from Pakistan
39
4.2 Correlation between morphological parameters of Rosa centifolia genotypes from USA
43
4.3 Grouping of 24 genotypes of Rosa centifolia on the bases of microsatellite markers
53
4.4 Grouping of 10 genotypes of Rosa centifolia on the bases of microsatellite markers
59
4.5 Grouping of 14 genotypes of Rosa centifolia and Rosa damascena on the bases of microsatellite markers
63

List of Figures
No. Title Page
3.1 Figure 3.1 Map of Pakistan showing the production areas of Rosa centifolia in Punjab province
20
4.1 Cluster analysis of genotypes of Rosa centifolia from Pakistan 37
4.2 Principle component analysis of genotypes of Rosa centifolia 38
4.3 Cluster analysis of different varieties of Rosa centifolia from USA 41
4.4 Principle component analysis of different varieties of Rosa centifolia from USA
42
4.5 Cluster analysis of 8 genotypes of Rosa centifolia from Pakistan and 8 varieties from USA
44
4.6 Undulation of leaf margins of different genotypes of Rosa Centifolia; a. week undulation of leaf margins, b. strong undulation of leaf margins
45
4.7 Terminal leaflet of different genotypes of Rosa Centifolia; a. leaflet with greater width, b. leaflet with greater length
45
4.8 Terminal leaflet shape of base of different genotypes of Rosa Centifolia; a. leaflet with round base, b leaflet with heart shape base
46
4.9 Number of thorns of different genotypes of Rosa Centifolia; a. stem with more number of thorns, b. stem with less number of thorns
46
4.10 Number of petals per flower of different genotypes of Rosa Centifolia; a. flower with greater number of petals, b. flower with less number petals
47
4.11 Dendrogram explaining diversity among 8 genotypes of Rosa centifolia from Pakistan
52
4.12 Dendrogram explaining diversity among 8 genotypes of Rosa centifolia from USA and 2 from Pakistan
58
4.13 Dendrogram explaining diversity among 8 genotypes of Rosa centifolia from USA, 2 from Pakistan along with 4 genotypes of Rosa damascena, 2 from Pakistan and 2 from Iran
63
4.14 Number of days taken to produce shoots from nodal explants of Rosa centifolia under in vitro conditions.
66
4.15 Number of shoots produced from nodal explants of Rosa centifolia under in vitro conditions.
67
4.16 Length of shoots produced from nodal explants of Rosa centifolia under in vitro conditions.
68
4.17 Number of leaves produced from nodal explants of Rosa centifolia under in vitro conditions.
69
4.18 Shoot proliferation in the MS media supplemented with BAP(A= control, B= 0.2 µM BAP, C= Callus and shoot both produced in MS media supplemented with 0.6 µM BAP along with 0.2 µM Kinetin)
70
4.19 Number of days taken to induce roots from in vitro grown shoots of 71

Rosa centifolia under in vitro conditions. 4.20 Number of roots produced from in vitro grown shoots of Rosa
centifolia under in vitro conditions. 72
4.21 Length of roots produced from in vitro grown shoots of Rosa centifolia under in vitro conditions.
73
4.22 Root induction in MS media supplemented with IBA 74
4.23 Acclimatization of plantlets (A= Plantlet covered with polythene bags for hardening, B= The acclimatized plantlets ready to shift in greenhouse)
74

List of Plates
No. Title Page
4.1 Primer (RW3N19) with the DNA samples of Rosa centifolia from different districts of Punjab, Pakistan
49
4.2 Primer (RW14H21) with the DNA samples of Rosa centifolia from different districts of Punjab, Pakistan
50
4.3 Primer (RW55C6) with the DNA samples of Rosa centifolia from different districts of Punjab, Pakistan
51
4.4 Primer (RW10M24) with the 2 DNA samples of Rosa centifolia of Pakistan and eight from USA
54
4.5 Primer (RW3N19) with the 2 DNA samples of Rosa centifolia of Pakistan and eight from USA
55
4.6 Primer (RW18N19) with the 2 DNA samples of Rosa centifolia of Pakistan and eight from USA
56
4.7 Primer (RW55C6) with the 2 DNA samples of Rosa centifolia of Pakistan and eight from USA
57
4.8 Primer (RW3k19) with the 8 DNA samples of different centifolia from USA and 2 from Pakistan along with 4 genotypes of Rosa damascena, 2 from Pakistan and 2 from Iran
60
4.9 Primer (RW3k19) with the 8 DNA samples of different centifolia from USA and 2 from Pakistan along with 4 genotypes of Rosa damascena, 2 from Pakistan and 2 from Iran
61
4.10 Primer (RW18N19) with the 8 DNA samples of different centifolia from USA and 2 from Pakistan along with 4 genotypes of Rosa damascena, 2 from Pakistan and 2 from Iran
62

Abstract
Rosa centifolia is one of the famous oil producing species of roses, used to extract oil and
other products. The present study was performed at Institute of Horticultural Sciences,
University of Agriculture Faisalabad (Pakistan) to evaluate the morphological and genetic
diversity among different genotypes of Rosa centifolia taken from Pakistan and USA and to
develop an in vitro propagation protocol for mass production of healthy plants round the
year. For study morphological diversity 24 plant, leaf and flower parameters were selected
from (UPOV) plant descriptor. Data of these parameters from 8 genotypes of Rosa centifolia
found in 8 districts of Punjab, Pakistan and 8 genotypes found in USA was collected
according to the descriptor. Young unopened leaves of these genotypes were collected for
DNA extraction through CTAB method, which was used for genetic analysis by SSR
markers. For in vitro propagation of Rosa centifolia young axillary buds were used as
explant. Different concentrations of BAP (0.2, 0.4, 0.6 and 0.8 µM) and Kinetin (0.2, 0.4, 0.6
and 0.8 µM) alone and in combination were used for shoot formation. For root induction
various concentrations of IBA (0.2, 0.4 and 0.6 µM) and NAA (0.2, 0.4 and 0.6 µM) were
used. Statistical analysis was performed on the data by using SAS statistical analysis
(Version 9.3) software. Principle component analysis was used to indicate the morphological
diversity among 8 genotypes of Rosa centifolia in Pakistan and 8 from USA. It showed the
highest phenotypic diversity between Sargodha and Sheikhupura genotypes while maximum
similarity was observed between Pattoki and Faisalabad genotypes. Among 8 genotypes of
Rosa centifolia from USA, maximum phenotypic association was noticed within Fantin-
Latour and Rosa de Meaux while maximum diversity was between Cabbage rose 2 and Paul
Ricault. During studying genetic diversity by using SSR markers genetic similarity was
observed among the genotypes of one district but genotype of different districts expressed
genetic diversity, which was maximum between Kahror Pakka and Pattoki genotypes but
minimum between Faisalabad and Sheikhupura genotypes from Pakistan. Rosa centifolia of
Pakistan and Cabbage rose 2 of USA were in the same group, which reflect genetic
relationship between them. During comparison of Rosa centifolia with Rosa damascena,
Pakistani genotypes of Rosa centifolia were genetically more closer to Rosa damascena
genotype from Isfahan (Iran) and Pattoki (Pakistan). Rosa damascena Faisalabad was in

separate group than other Rosa damascena genotypes and showed closer genetic association
with Rose de Meaux and Gros Choux d’Hollande. During in vitro propagation of Rosa
centifolia BAP at 0.4 µM along with 0.2 µM of Kinetin produced shoots with minimum
number of days (10.20). BAP at concentration of 0.2 µM proved as prominent by producing
maximum number of shoots (1.93) while 0.2 µM BAP in combination with 0.2 µM Kinetin
produced shoots with maximum shoot length (5.55 cm). MS media supplemented with higher
concentration (0.4 µM and 0.6 µM) of BAP along with 0.2 µM Kinetin first produced callus
on the explants and after few days it also produced the shoots. In case of root induction 0.4
µM IBA proved excellent and produced roots with minimum number of days (10.07),
maximum number of roots (2.47) and maximum root length (4.21 cm).

1
Chapter-1
INTRODUCTION
Rose, an economically very important floriculture crop in the world (Castilon et al., 2006;
Senapati and Rout, 2008; Heinrichs, 2008), belongs to family Rosaceae, which is consisted
of about 125 genera and 3500 species (Landrein et al., 2009) and genus Rosa made up about
200 species and 20000 cultivars (Cuizhi et al., 2003; Ritz et al., 2005). The genus Rosa is
native to temperate regions of northern hemisphere (Krussmann, 1981), which includes
North America, Europe, Asia and Middle East, North China is known for the great diversity
of Rosa species (Phillips and Rix, 1988; Ertter, 2001). The history of rose is as old as the
history of human civilization. Rose fossils from Asia, Europe and America have been dated
as 30 million years old (Chakraborty, 2005). There is clear evidence of roses in the Egyptian
pyramids (Anonymous, 2011), in the gardens of Semiramis, queen of Assyria (Lehner and
Lehner, 1960) and in the Zoroastrian text Bundehesh, which described that hundred-petalled
rose and dog rose develop thorns when evil came in the world (Joret, 1892). A common
concept among Turks and the Persians is that roses were produced from the drops of
Mohammed's sweat (Joret, 1892). It is king of flowers (Peter Bealis, 1990) with great
diversity in the world and is used beyond for ornamental purposes, in the food, cosmetics
(Gustavsson, 1998; Kaur et al., 2007) and the medicinal industry (Basim and Basim, 2003;
Ozkan et al., 2004; Achuthan et al., 2003; Mahmood et al., 1996; Bown, 2002) since the
dawn of civilization (Randhawa and Mukhopadyay, 1994). Now the roses have proved the
most popular garden plant and cut flower in the world (Horst, 1983). It is the leading cut
flower because it is used to increase decoration and charm in different celebrations like
marriage ceremonies, birthday, Valentine’s Day etc. Due to its extensive use in the landscape
no garden can be completed without roses. It is used to grow in Netherland, Colombia, Israel,
Italy, Bulgaria and in many other countries. There is estimated area of 8,000 ha of roses
under protected production to produce 15-18 billion rose stems annually in the world (Blom
and Tsujita, 2003). In total, an area of 16,000 ha in greenhouse space and 3,000 ha in the
open field space is used throughout the world to produce cut roses and potted rose plants
(Brichet, 2003). The annual value of cut flower, potted roses and garden roses is about $ 10
billion (Evans, 2009).

2
Some roses have pleasant fragrance in their petals and are used for the extraction of essential
oils (Marchant et al., 1996; Lu et al., 2003). Unfortunately most cut flowers do not have
fragrance (Zuker et al., 1998) and are used only for ornamental purposes. Rose oil was first
discovered by Geronimo Rossi in 1574 when he was working with rose water and saw an
upper layer of fragrant oil (Parry, 1925; Gordon, 1953). Now rose oil is one of the most
expensive oils in the world (Baydar and Baydar, 2005) due to the presence of very little
amount of oil in rose petals as compared to other oil producing crops Thus large number of
flowers are needed to extract oil from rose petals, about 3000 kg of rose petals are needed to
produce one kg of rose oil (Baser, 1992). But rose oil is very unique and important for
making perfumes (Krussman, 1981; Widrlechner et al., 1981). It contains more than 500
volatiles (Knudsen et al., 2006) and high percentage of monoterpene alcohols including
citronellol, nerol, geraniol, linalool, and phenylethyl alcohol, which are very significant for
perfume (Anaç, 1984; Kovats, 1987; Lawrence, 1991; Baser, 1992; Bayrak and Akgül,
1994). It has also been proved that rose oil has antibacterial, antimicrobial, anti HIV and
antioxidant effect (Achuthan et al., 2003; Basim and Basim, 2003; Buckle, 2003; Ozkan et
al., 2004), is useful as an asthma remedy (Berkowsky and Bruce, 2000), for creating a feeling
of vitality and well being (Sheppard-Hanger, 1995) and is used in various food products.
Annual rose oil production in the world is 4.25 tons and is increasing every year (Narayan
and Kumar, 2003). Famous rose oil producing species in the world are R. damascena Mill.,
Rosa gallica L., R. centifolia L. and R. alba L. (Topalov, 1978; Tucker and Maciarello, 1988;
Rusanov et al., 2005; Tabaei-Aghdaei et al., 2007). The main rose oil producing countries
are Bulgaria, Turkey and Iran with lesser amounts produced in India, China and Northern
Africa (Rusanov et al., 2005; Weiss, 1997). Rose hips are also an excellent source of vitamin
C in the world (Marchant et al., 1996) as researchers have found that hips may contain 400%
more vitamin C than oranges. These are also used to make jam, marmalade, fruit juice (Uggla
et al., 2003) tea from dried hips (Kurt and Yamankaradeniz, 1983; Ercisli and Guleryuz,
2005) and antibacterial and antimicrobial products (Ozkan et al., 2004).
Rosa centifolia is famous among oil producing species of roses (Lawrence, 1991) in the
world. It has pink fragrant flowers with many petals. It was originally cultivated in Bulgaria.
Then distributed in Turkey (Baydar et al., 2004) and other parts of the world. Now it is
grown in Bulgaria, Israel, Italy, United States, Japan, Turkey, Iran, India, Pakistan and

3
France but it is more common in Morocco, France and Egypt (Nasir et al., 2007). In Pakistan
it is a very high value and demanded nontraditional crop and popular among small and large
growers. The Rosa project at the Institute of Horticultural Sciences, University of Agriculture
Faisalabad has been growing this species for many years and has used it for different
purposes with the most important product being rose oil and rose water. Volatile oils are
extracted by supercritical fluid extraction, solvent extraction, steam distillation, cold
pressing, and hot pressing methods (Lis-Balchin and Deans, 1997; Younis et al., 2009). The
Rosa project has extracted oil from the flowers of R. centifolia by steam distillation (Tuan
and Iiangantileke, 1997; Simandi et al., 1999), soxhlet extractor and recently by the super
critical fluid extraction method.
The oil of R. centifolia contains chemical constituents which are very useful in the
production of food products, in medicine and perfumes on commercial scale. The products
are very common and in high demand in Pakistan as people use rose water to clean different
sacred places and the body especially the face as it has a fresh pleasant smell, as antibacterial
and eye protection properties (Jalali-Heravi et al., 2008). Its yellowish brown colored oil
(Khan and Rehman, 2005) is antiseptic, antidepressant, astringent, digestive, anti-
inflammatory, aphrodisiac (Allardice, 1994) and anti-tussive with citronellol, geraniol, nerol,
linalool, methyl eugenol, eugenol and rose oxide as main chemical components (Shiva et al.,
2007). In Pakistan it is mostly used for perfume making. As rose jam containing petals is
considered good for digestive system and eye drops are considered very effective for eyes.
This is also a medicinally important therapeutic agent (Jitendra et al., 2012). Now mostly the
growers from different districts of Punjab are also growing this specie and selling its flowers
as fresh or making different by- products. More and more farmers are entering in the
business. An acre of 5,000 plants can produce 5000 kg flowers, which yield almost 1.0 kg of
oil. This sells for much more than input cost. Another advantage in this species is that it can
produce flowers throughout the year as compared to other rose species which are not able to
produce flowers during the hot summer conditions.
There are variations among the plants of R. centifolia present in different districts of Punjab,
Pakistan. This has given rise to selection of several promising landraces in different parts of
the country. These different landraces of R. centifolia with different characters are preferred
by farmers. Farmers entrust in landraces rather than formal breeding systems (Almekinders et

4
al., 1994) because landraces contain convincing phenotypic variations, can develop
resistance against biotic and abiotic stresses (Eagles and Lothrop, 1994), have historical
origin, noticeable identity, genetic diversity, accepted and are preferred by local farmers and
absent from formal crop improvements (Camacho Villa et al., 2005). Standardization of
different landraces is difficult because they are different by environmental condition or
genetic variations (Adugna et al., 2006). Due to growing demand of Rosa centifolia, the need
of its standardization by morpho genetic analysis is also increasing, which was a part of this
study.
The usual method of propagation of roses is asexual by rooted cuttings or grafting. (Zieslin,
1996). Seeds are also used for the propagation of species and rootstock (Horn et al., 1992).
These conventional methods are facing various obstacles like the limitation in stock plant,
long production time (Skirvin et al., 1990), low multiplication rate and dependence on season
(Pati et al., 2006), low ability of root formation (Collet and Le, 1987, Podwyszyfiska and
Hempel ,1987; Rogers and Smith, 1992). Now there is a great need to develop new effective
ways for rapid mass propagation of roses (Shabbir et al., 2009). Plant tissue culture is
becoming more common as an alternative way for plant propagation (Khosravi et al., 2007)
especially for rapid propagation by its various techniques (Khosh-Khoi and Sink, 1982,
Ishioka and Tanimato, 1990; Kornova and Michailova, 1994; Kumar et al., 2001; Pati et al.,
2001; Kapchina-Toteva et al., 2002). It has helped the nursery business (Pierik, 1991) as
400,000 plants can be produced in a year by this method (Martin, 1985). It could produce
healthy, disease free plants throughout the year (Dhawan and Bhojwani, 1986; Razavizadeh
and Ehsanpour, 2008). In vitro grown plants produce good quality planting material (Paek et
al., 1998), better branched, compact plants, more flower yields (Reist, 1985; Pati et al., 2006)
and greater root formation (Pati et al., 2004). It is also a good substitute for conventional rose
propagation (Khosravi et al., 2007). For in vitro propagation any plant part can be used
because every plant part has the ability of totipotency (Khan and Shaw, 1988). Different
explants including in vitro grown leaves (Dohm et al., 2001; Zakizadeh et al., 2008)
immature leaves and internodal stem segments (Li et al., 2002), in vivo grown leaves
(Kintzios et al., 1999), immature seeds (Kunitake et al., 1993), in vitro derived petioles and
roots (Marchant et al. 1996), adventitious roots (Van der Salm et al., 1996), petals (Murali et
al., 1996), filaments (Noriega and Söndah, 1991) and different flower parts (Arene et al.,

5
1993), which are transferred on MS medium containing different levels of plant growth
regulators. In vitro regeneration methods have facilitated the mass propagation of roses and
boosted the rose floriculture industry (Dohare et al., 1991; Short and Roberts, 1991). The
main focus in rose in vitro culture is micropropagation (Jacobs et al., 1969; Hasegawa, 1979;
Bressan et al., 1982) and mostly axillary buds are used for mass propagation by this
technique (Elliott, 1970; Davies, 1980; Bressan et al., 1982; Barve et al., 1984; Douglas et
al., 1989; Ishioka and Tanimoto, 1990; Yan et al., 1996; Ara et al., 1997; Carelli and
Echeverrigaray, 2002; Rout and Jain, 2004). This has been proved the best for shoot
proliferation of roses (Pati et al., 2006; Xing et al., 2010).
There are production and market related risks to agricultural production in the world. These
risks are increasing with the passage of time, as planet is becoming warmer, rainfall pattern
changing and other incidents like flood, drought and forest fire are also becoming common
(Zoellick, 2009). These climate changes may result in poor and unpredictable yield, more
pest and disease problems, more weeds, unstable quality and poor farmer returns herbivores
(Zvereva and Kozlov, 2006). Growing R. centifolia is not very common in Pakistan, but the
demand of R. centifolia is increasing due to its use in a variety of products and high returns to
producers. The liberalization and globalization of the economy have diminished the
worldwide boundaries and magnify the competition in the world market, which has also
increased the opportunities for Rosa centifolia both domestically and globally. The opening
up of the world flower market has also built new challenges in terms of product quality,
safety and price in order to fulfill the global competition as well as WTO standards. At a
global level, the morphological and genetic characterization is very important to meet the
WTO standards and intellectual rights.
Morphological traits are the earlier traditional markers used for the identification and
management of different germplasms but these are not reliable markers because of less
polymorphism, less heritability, late expression of traits and susceptibility to environmental
factors (Smith and Smith, 1992). These phenotypic markers may be changed with the
cultivation techniques and growing environments so identification could be confusing. To
characterize Rosa centifolia types in systematic way, specific morphological and genetic
markers have been developed such as restriction fragment length polymorphism (AFLP)
(Rajapakse, 2003; Baydar et al., 2004; Pirseyedi et al., 2005), Random amplified

6
polymorphism DNA (RAPD) (Agaoglu et al., 2000; Martin et al., 2001; Fredrick et al.,
2002; Kiani et al., 2007) and Simple Sequence Repeats (SSR) (Esselink et al., 2003; Nybom
et al., 2004; Rusanov et al., 2005; Zhang et al., 2006; Babaei et al., 2007). Dominant
polymerase chain reaction (PCR)-based marker technologies such as random amplified
polymorphic DNA (RAPD) were more commonly used and proved as suitable tool for the
study of genetic variation in the fragrant roses (Prasad et al., 2006; Tabaei et al., 2006; Kiani
et al., 2007) because they are simple, require lower cost and infrastructure and are easy to
perform. Random amplified polymorphism DNA (RAPD) have some drawbacks (Jones et
al., 1997) and do not provide complete information for the analysis of chromosome
distribution. Amplified fragment length polymorphism (AFLP) analysis is comparatively
cheap, easy, fast, reliable and highly reproducible for screening of a large number of
molecular markers from the genome (Vos et al., 1995; Vos and Kuiper, 1997). They can be
used to determine the phylogenetic relationship on the basis of genetic distance (Heun, 1997;
Beismann, 1997; Semblat, 1998; Huys, 1996; Keim, 1997; Tohme, 1996) but they are
difficult to use to identify homologous markers so these are not suitable for heterozygosity
analysis. AFLPs have recently been used in a set of 88 rose genotypes to detect a clear
distinction between cultivars and wild species (Leus et al., 2004).
Simple sequence repeats (SSRs) are neutral, co-dominant (unlike RAPDs and AFLPs), and
produce more polymorphic markers (Tautz, 1989; Koreth et al., 1996) which are informative,
easily transferable (Rajapakse, 2003) and show high level of heterozygosity in Rosaceae
family (Guilford et al., 1997; Hokanson et al., 1998; Sosinski et al., 2000; Esselink et al.,
2003). They were developed to improve the availability of molecular tools for genetic studies
and further marker assisted breeding in R. centifolia and its closely related species. Hence,
the study of the genotype at the genetic level along with phenotypic characterization will be
an important step towards efficient conservation, maintenance and utilization of the existing
genetic diversity.
Thus considering the problems related with R. centifolia, present research work was
performed with two objectives. 1. To document the genetic diversity of R. centifolia through
morphological and molecular markers. 2. To develop an in vitro propagation protocol for
year round mass propagation of disease free plants of R. centifolia.

7
Chapter-II
REVIEW OF LITERATURE
Roses are most popular, enjoyable and high demanded flowering plants in the whole world
due to its major traits like flower color, fragrance, recurrent blooming, vase life, stem length,
hardiness etc. Now more research is going on in the inheritance of important rose traits.
(Debener, 1999; Zykov and Klimenko, 1999; Gudin, 2000; Rajapakse et al., 2001)
2.1 Morphological Analysis
Variations among different individuals of species or population may be due to genetic,
influence of environment or other factors. In individual populations some traits are affected
greater by environment than other more reliable trait and used in morphological studies
(Debener, 2002). Many scientists in the world have studied the variation among rose plants
on morphological basis. As Shafiq et al. (2006) studied correlation in four important Rosa
species, Rosa centifolia, Rosa damascena, Rosa chinensis and Rosa bourboniana by using
five morphological characters under Faisalabad climatic conditions in Pakistan.
Morphological characters like plant height, fresh flower weight, dry flower weight, number
of flowers per plant and number of branches per plant were evaluated. Correlation was
significant between fresh flower weight and other morphological characters and positive
correlation was observed between the number of flowers per plant and dry flower weight.
Correlation between the number of flowers per plant and other morphological characters
showed significant and positive with fresh flower weight and dry flower weight. Negative
correlation was observed between the number of flowers per plant and dry flower weight. R.
centifolia and R. damascena are two most important oil producing species of roses. Nawaz et
al. (2011) showed diversity in the leaf anatomy of different roses commonly present in
Faisalabad and adjoining area. According to Nawaz et al. (2011) lot of anatomical
modifications in leaves made R. damascena ecological successful to different environmental
conditions along with other species (R. bourboniana ‘Gruss-an-Teplitz).
Flower is a main part in roses and great diversity in flowers of different genotypes under
various climatic conditions has been studied by many scientists. Zeinali et al. (2009) studied
the flower yield and numerous factors that affect R. damascene, most famous oil bearing
specie of rose found in various regions of Iran. After studying twelve yield affecting

8
parameters, they concluded that except one trait (fresh weight of petals per flower) all other
were similar. Cluster analysis explained rose from Khuzestan and Shiraz were very similar
but western and eastern Azerbaijan were different genotypes. They also suggested that most
important parameter for flower yield is the number of flowers. R. centifolia produces
yellowish brown color oil, which has greater refractive index and all chemical constituents
except phenyl ethyl alcohol than yellowish color R. damascena oil (Khan and Rehman,
2005). Morphological and oil based diversity among eight landraces of R. damascena in
Punjab was found by Farooq et al. (2013). They explained negative correlation of flower
weight with peduncle length but highly positive with rest of parameters. Landrace of Choha
Syedan Shah produces maximum oil contents and Dendrogram did not show any relation of
genetic variation. Positive correlation of flower yield with number of flowers and branches
per plant of twenty different genotypes of damask rose in India by Singh and Kayiyar,
(2001). Khan et al. (2011) studied the effect of waste stabilization ponds (WSP) effluent on
growth of R. damascene and also compared it with Hoagland solution. Both treatments were
better for number of flower, size of flower and the number of petals of a flower but plant
height and flower fresh weight enhances only by Hoagland solution. They proved that using
such treated effluent can reduce the need of fresh water and fertilizer for roses. Riaz et al.
(2007) morphologically characterized wild rose plants of Rosa species (R. webbiana and R.
brunonii), which were collected from five different sites of Northern areas of Pakistan. R.
webbiana, which was collected from Nathia gali and Murree, showed 83% similarity with all
other rose genotypes. R. brunonii collected from Sunny and Ayyubia also showed 80%
similarity with other genotypes. Different genotypes from different geological regions
showed very little difference. Small difference was observed only in environmentally
dependent characters like leaf length, plant height and fruit length. Kaul et al. (2009)
evaluated the F1 hybrids that were produced by crossing two cultivars (Jwala and Himroz) of
R. damascena with R. bourboniana by using morphological, random amplified polymorphic
DNA (RAPD) and microsatellite (SSR) markers. Twelve morphological characters were
studied, five characters out of 12 (i.e. plant height, number of primary branches, leaf length,
length of leaflets, width of leaflets) were different to the interspecific hybrids. The other
characters (the number of internodes per branch and the number of prickles in top 10
internodes and the number of buds per bunch) were similar to those of R. bourboniana and

9
the number of petals was closer with R. damascena than R. bourboniana. Twenty-two RAPD
and three SSR primers were used for the identification of the hybrids. Random amplified
polymorphic DNA (RAPD) and SSR markers were classified into seven types according to
the presence and absence of the bands. Bands that were specific with the pollen parent and
present in the hybrids were good markers to identify the hybrids. Bands absent in the parents
were very effective in distinguishing the hybrids from each other.
2.2 Molecular Analysis
Molecular markers are one of the major tools in rose breeding. Genetic diversity increases
knowledge about genetics and relationship, which is significant in development of
commercial type varieties (Debener, 2002). (Simple sequence repeat) markers are very
effective for genetic studies of genus Rosa. As Samiei et al. (2010) found genetic variation
among various accessions of wild roses collected from different areas of Iran by using 10
SSR primers. Kiani et al. (2010) suggested Iran as a region of great diversity of R.
damascena by studying genetic diversity among 41 genotypes of R. damascena used to
grown in different areas of Iran and one from Bulgaria with 37 SSR primers. With
unweighted paired-group of arithmetic mean (UPGMA) cluster analysis they observed most
of them were tetraploids and there was one triploid and two hexaploids. The species of one
province were more related with each other than the other provinces. Emadpour et al. (2009)
studied genetic diversity in fifteen commercial cultivars of rose by using ten RAPD primers.
All the primers showed polymorphism among the cultivars. Primer E and F produced the
highest but primer H produced the lowest number of bands, which ranged from 37% to 81%
with an average of 63.9%. The average numbers of polymorphic bands produced were 7.3
per primer. Results showed that RAPD markers are a useful tool for the study of genetic
diversity among different Rosa species. Babaei et al. (2007) studied 40 accessions of Rosa
damascena from major and minor rose oil producing areas of Iran by using nine polymorphic
microsatellite markers. All microsatellite markers produced a high level of polymorphism (5-
15 alleles per microsatellite marker, with an average of 9.11 alleles per locus). Microsatellite
markers identified nine different genotypes and the most common genotypes (27/40
accessions) were from the major Isfahan province. Other genotypes (each represented by 1-4
accessions) were collected from minor production areas in several provinces, mainly from
the mountainous Northwest of Iran. Baydar et al. (2004) studied genetic relationship among

10
morphological different plants of R. damascene grown in different regions of Turkey by
using microsatellite and AFLP markers. Twenty-three AFLP and nine microsatellite primer
pairs were used. There was no polymorphism detected among the plants of different regions,
as the marker patterns obtained from morphologically different plants were similar. This
study proved that morphological variations in the plants were due to the point mutations,
which cannot be detected by molecular markers.
Rusanov et al. (2005) used microsatellite markers for genetic diversity of twenty-six oil-
bearing R. damascena Mill. accessions and 13 garden Damask roses that are grown in
different countries of Europe and Asia for rose oil production. Eleven SSR markers showed
that old European Damask rose varieties Quatre Saisons, York, and Lancaster, and R.
damascena accessions from Bulgaria, Iran, and India used for rose oil production had
identical microsatellite profiles, suggesting a common ancestors and origin. The results also
showed that oil rose cultivation are based on narrow genepool and there are many genetically
identical accessions in rose oil collections. Kiani et al. (2008) studied genetic relationships
among 41 Rosa damascena accessions collected from different areas of Iran and one
accession from Bulgaria by using 31 RAPD primers. PCR amplifiable DNA was isolated
using the CTAB method and each primer produced 3–12 banding patterns for a total of 343
scorable and 184 polymorphic bands. The frequency of banding pattern was different in each
primer, which was from 0.02 to 1. The discriminating powers of primer were from 0.263
(BE11) to 0.702 (BE5), with a mean value of 0.521. ANOVA results indicated that majority
of variations were due to differences within the provinces (65.7%) and only 34.3% were due
to differences among provinces. The genetic distance between the 5 provinces ranged from
0.01 between Fars and Isfahan to 0.51 between East Azerbaijan and Kerman.
Damask rose is famous in varietal development in roses. It has triparental origin from R.
moschata, R. gallica and R. fedschenkoana grown for years by vegetative means, was proved
by Iwata et al. (2002) by using 24 RAPD primers. Azeem et al. (2012) documented the role
of RAPD markers in roses. They use 25 RAPD markers to study the heterogeneity among the
4 species and 30 accessions of rose collected from floricultural area of the Institute of
Horticulture University of Agriculture, Faisalabad. Ten out of twenty markers showed 100%
polymorphism among the marker GLD-20 produced highest number of 10 bands and GLD-
02 produced least number of 2 bands. Kaul et al. (2009) made crosses between R.

11
damascena Mill (scented rose) and R. bourboniana L. (recurrent flower and easy to
propagate rose). The F1 hybrids were checked by morphological, RAPD and microsatellite
(SSR) markers and were divided into two groups. They proved the usefulness of these two
dominant and co dominant markers for identification of hybrids at seeding level to remove
the doubtful plants, which saves the space and other inputs. Yan et al. (2005) used 520
molecular markers of AFLP, SSR, RFLP, SCAR to construct genetic linkage map of 88
individuals of a population. They made seven linkage map of one parent and integrated map
by consolidating the parental maps which is most advanced and informative map for future
roses research. Basaki et al. (2009) used AFLP technique to study the genetic diversity
among 128 accessions of R. persica from Iranian. 171 polymorphic fragments were produced
and the numbers of polymorphic loci were in the range of 101 to 147. The polymorphism
information content (PIC) was in the range of 0.289 to 0.073 with an average value of 0.16.
Cluster analysis was done by the use of UPGMA method, which grouped all accession in six
clusters. According to molecular variance analysis 48% of the genetic variation of R. persica
was within population and 52% was among populations. Sequenced tagged microsatellite site
(STMS) markers are very effective tool for determining sources of varieties and illegitimate
propagation. As 24 Sequenced tagged microsatellite site (STMS) markers were used for
characterization of 46 hybrid tea and 30 rootstock varieties, found clones and flower color
mutants identified by Esselink et al. (2003). Scariot et al. (2006) proves that DNA analysis
can help in botanical classes of roses by constructing genetic profile of 65 accessions of old
garden roses by using six polymorphic sequence tagged microsatellite sites (STEM). Results
showed 82 alleles at six loci with average of 13.5 alleles at one locus. They made seven
clusters for grouping of genotypes by using cluster analysis.
2.3 In vitro propagation
There are many reports about direct in vitro propagation of roses through different techniques
by using different explants. In vitro regeneration of R. centifolia from the node is scarce.
Baig et al. (2011) established a system for in vitro propagation of two very famous species of
roses in Pakistan, Rosa gruss an teplitz and R. centifolia by using shoot tips as explant.
Different concentrations of benzylaminopurine (BAP) were used in the MS media for shoot
formation and different concentrations of indole-3-butyric acid (IBA) in half strength MS

12
media for root formation. 1.0 mg L-1 BAP was proved the best treatment for in vitro shooting
and 0.50 mg L-1 was the best for rooting. In vitro regeneration of rose from axillary nodes
was reported by Chakrabarty et al. (2000). Razavizadeh and Ehsanpour (2008) developed a
protocol for micropropagation of R. hybrida through auxiliary buds. The explants were
cultured on MS medium supplemented with benzyl adenin (BA) (0-12 mgL-1) and agar (4.4
and 5.6g L-1). BA significantly affected the number and height of produced shoots. The
highest proliferation rate (4.78) was achieved in MS medium containing 8 mgL-1 BA and
5.6gL-1 agar while the effect of agar was significant only on the height of produced shoots.
The effect of different concentrations of IAA (0-6 mgL-1) and MS salt concentration (1/4, ½
and ¾) were also examined for optimal rooting. The result shows that the root number was
significantly increased in higher concentration of indole acetic acid (IAA) but salt had a litter
effect on root parameters. Root plantlets were transferred to 3 different mixture of sterile soil
consisting of peat mass + sand 1:1 (v/v) and perlite + vermiculite 1:1 (v/v) and after 4 weeks
acclimatization, were placed in the greenhouse. Mohan et al. (2000) developed rapid and
efficient protocols for in vitro propagation of essential oil-producing species of roses, R.
damascena and R. chinensis by testing 12 various media combinations. Murashige and Skoog
MS medium was supplemented with different concentrations of BAP and IAA, to induce
adventitious shoots regeneration from nodal explants of R. damascena and R. chinensis.
Singh and Syamal (2000) Studied that shoots of rose cultivars Super Star and Sonia were
multiplied for ten subcultures at 4-week intervals on solidified MS medium supplemented
with 22.19 µM BAP + 1.07 µM napthalene acetic acid (NAA) + 0.05 µM gibberellic acid
(GA). Addition of anti-auxins 2,3,5-triiodobenzoic acid (TIBA; 2.0 µM) and 2,4,6-
trichlorophenoxy-acetic acid (2,4,6-T; 0.39 µM) into proliferation medium increased the
number of shoots per explant and the length of shoots in both cultivars. Treatment with TIBA
also increased the number of leaves per shoot and leaf chlorophyll content. Misra et al.
(2009) developed protocol for clonal propagation of R. clinophylla, through in vitro axillary
bud culture by using cytokinins alone and in combination of GA3. Cytokinins alone were
able to induce shoot buds, but their proper growth and number could be increased only when
they were used in combination with GA3. Shoot tip necrosis and leaf fall were observed in
the proliferated shoots, which was controlled by AgNO3 at 58.85 µM. Activated charcoal
(AC) at 250 mg L-1 was also found effective at all the stages of shoot multiplication as well

13
as rooting. Ninety percent rooting could be achieved in 1/2 MS medium supplemented with
4.92 µM IBA and 250 mg L-1AC and rooted plantlets were hardened and transferred to the
field successfully with 80% survival rate. Ozel et al. (2006) selected shoot tip segments from
2 years old plants of Heritage rose to culture on MS medium supplemented with various
levels of thidiazuron (TDZ), kinetin and NAA. The effect of full, ½ and ¼ strength of MS
macro, micro elements and vitamins was also observed on root induction. The results showed
highest shoot proliferation on MS medium containing 0.05 mg L-1 TDZ, 0.2 mg L-1 kinetin
and 0.1 mg L-1 NAA while reduction in shoot formation was observed on media without
NAA or with increased level of TDZ. The results also showed that MS medium with half
concentration of macro, micro salts and vitamins was most suitable for in vitro root
induction. Pawlowska, (2011) used five Rosa species from Poland to culture on MS media
containing BA alone and in combination with GA3 under 16 hours photoperiod, 23/25°C
temperature 80% humidity. R. canina and R. rubiginosa and R. dumalis showed the highest
number of shoots with 1 μM BA and 1.5 μM GA3. Shabbir et al. (2009) cultured 4-5 mm
long shoot tip as explant on MS media containing BA alone and in combination with NAA to
investigate best growth regulator and media for in vitro propagation of R. indica. Media
containing 1.5 mg L-1 BAP showed excellent results for shoot formation. Jaskani et al.
(2005) reported an extensive work for in vitro propagation of rose cvs. Queen Elizabeth and
Angel Face. Khalifah et al. (2005) studied the effect of sucrose concentration and genotype
on growth of axillary buds of five different hybrid-tea rose (Rosa hybrida L.) cultivars.
Shoots were produced in MS medium supplemented with 4.0 mg L-1 BA, 0.2 mg L-1 kinetin
1.0 mg L-1 GA3 and 30 mg L-1 sucrose and then sub cultured on medium with 3.0 mg L-1 BA,
0.2 mg L-1 Kinetin and three different concentrations of sucrose (30, 40 and 50 g L-1).
Different numbers of shoots along with main shoot were produced in all concentrations of
sucrose. Best rooting was in the medium containing1.0 mg L-1of IBA and 40 g L-1of sucrose
than 30 and 50 g L-1. Genotype difference was also very clear because of difference in values
of shoots produced and elongations of shoots produced. Kermani et al. (2010) investigated
the in vitro propagation of R. persica and concluded VS media with 8 μM of BAP superior
from MS and QL media. According to the results agar was effective for controlling
vitrification and no significant difference among different treatments for rooting.

14
Nodal explants with an axillary bud are very persuasive explants used by different scientists.
Roy et al. (2004) used both Nodal segments and shoot tips as explant and BAP, NAA, zeatin,
IAA and IBA as growth regulators in MS media for in vitro propagation of roses. Nodal
explant yielded excellent results with 1.5 mg L-1 BAP + 0.25 mg L-1 zeatin in MS media for
shoot formation as compared with shoot tip. For root formation from shoots half strength MS
media supplemented with 1.0 mg L-1 IBA and 0.5 mg L-1 IAA proved the best. Nodal
explants were proved better than shoot tip for producing in vitro shoots of 'hybrid tea' roses
with 3 mg L-1 BAP and IAA and 0.5 mg L-1 IBA for root formation by Iqbal et al. (2003).
Hegde et al. (2011) resulted the nodal explants better shoot multiplication than shoot tip in
MS media containing 2 mg L-1 BAP + 0.2 mg L-1 kinetin and 25 mg L-1 adenine sulphate.
Mamaghani et al., (2010) propagated damask rose by axillary buds on MS and WPM
medium containing Thidiauzuron (TDZ) 6-benzylaminopurine (BAP) alone and in
combination for shooting and NAA for rooting. They observed that MS media was inimitable
with 5 mg L-1 BAP and 0.1 mg L-1 TDZ for shooting and with 0.1 and 0.2 mg L-1 NAA for
rooting. Noodezh et al. (2012) developed a protocol for in vitro propagation of damask rose
by culturing nodal explants on new media A19 (MS media with higher quantity of nitrates,
calcium, and iron along with 4 mg L-1 BAP and 0.25 mg L-1 IAA). After shoot initiation
explants should be subcultured on same media with addition of 0.2 mg L-1 GA, which
resulted in improvement in shoot growth, length and number. For root formation shoots were
transferred in liquid ½A19 medium-A containing 0.1 mg L-1 IBA after 7 days dark treatment
without growth regulator. BAP and NAA are very promising for producing healthy axillary
shoots from the axillary buds of R. hybrid cv baccara and 2 mg L-1 IBA for rooting of shoots
(Salekjalali et al., 2011). A system for micropropagation of rose ‘Pareo’ was developed using
nodal segments as explants, MS media with 1.5 mg L-1 BAP for shooting and half MS media
with 1.0 mg L-1IBA for rooting by Mukhambetzhanov et al. (2011). Nak-Udom et al. (2009)
used nodal explants micropropagation of R. hybrida L. cv. ‘Perfume Delight’ on MS media
containing different growth regulators. They resulted 3 mg L-1 BA and 0.003 mg L-1 NAA
effective for shoot formation and 3 mg L-1 BA and 0.003 mg L-1 NAA for rooting. Nikbakht
et al. (2005) cultured two cultivars of Iranian Damask rose on MS media by using different
growth regulators. Xing et al. (2010) succeeded in propagation of Rosa rugosa Thunb. by
culturing nodal segments on MS media containing BA 2.2 μM, NAA 0.054 μM, GA3 2.0 μM

15
and 3% sucrose for shoots and IBA (2.5-5.0 μM) in half MS media for roots. Asadi et al.
(2009) cultured the axillary buds of rose cv. Morrasia to study the effect of different
concentration of NAA and BAP to produce in vitro shoot and 3 mg L-1 BAP produces the
maximum number of shoots. For root formation from the in vitro shoots they used different
concentrations and combinations of IAA and NAA in the MS medium and IAA in third level
of NAA formed maximum number of roots. Wilson and Nayar, (1998) standardized the
sterilization, stage of explant and MS media for in vitro shoot formation of rose ‘folklore’
from axillary buds. They resulted that 0.08% mercuric chloride for 12 min. was best for the
sterilization of explants, 1.0 cm length of axillary buds of and BAP at 2.5 mg L-1, 2, 4-D at
0.5 mg L-1 were best for shoot formation. Extensive work on shoot formation from nodal
explants of R. hybrida ‘Heirloom’ was performed by Kanchanapoom et al. (2010).
Nodal explants can be used for flower formation under in vitro conditions as Kanchanapoom
et al. (2009) reported the in vitro shoot and flower formation from the nodal explants of R.
hybrida L. ‘red masterpiece’ on MS medium supplemented with different concentrations of
BA and kinetin. Maximum 5 shoots were produced on MS medium having 3 mg L-1 BA and
1 mg L-1 kinetin. Flowers were induced in the MS medium having 2 mg L-1 BA for 9 weeks.
Nikbakht et al. (2005) studied the in vitro propagation of two cultivars of Damask rose
‘Azaran’ and ‘Ghamsar’ by using nodal segments having lateral buds. Explants were
disinfected by using antibiotics for each cultivar and cultured on 12 different Media among
them liquid MS medium (with eliminated Cl- and reduced NH4+ ions) produces best shoots
without senescence. Different growth regulators BA (0, 1, 2 and 3 mg L-1), GA3 (0, 0.1, 0.25
and 0.5 mg L-1) and NAA (0 and 1 mg L-1) were also used in the medium among these BA
(1-2 mg L-1), GA3 (0.1 mg L-1) and NAA (0-0.1 mg L-1) yielded good results for ‘Azaran’
and same concentration of BA and GA3 without NAA showed maximum results for
"Ghamsar". Maximum rooting was obtained by placing shoots in liquid MS medium after
treating with 2000 ppm IBA solution.
There are different factors effecting in vitro propagation. Mahmoud et al. (2011) developed a
protocol for in vitro propagation of Eiffel Tower (R. hybrida, L.) by studying its various
factors. Nodal explants were very efficient if taken from two year old stock plants.
Murashige and Skoog media with 2% sucrose, half strength macronutrients and with AC are
good for shoot formation. In case of growth regulators IAA was better than IBA and BA.

16
Culturing of explants in petri dishes under 25°C temperature 60% humidity and 16h light
gave more reliable results than test tubes and jars under the same conditions. Sucrose is a
known factor affecting the floral morphogenesis (Vu et al., 2006), the number of shoots
(Kumar et al., 2010), development of longer and the number of roots of in vitro propagated
plantlets (Hyndman et al., 1982). Capelladesl et al. (1991) used shootlets of R. multiflora to
culture on MS media containing four different concentrations of sucrose. Results showed that
plantlets grown in media without sucrose contain plastoglobuli in chloroplast except starch
granules and media with 5% sucrose produced the maximum amount of starch, which is
helpful during acclimatization period.
Ramanayake et al. (2001) Studied axillary shoot proliferation and in vitro flowering using
single node explants from a mature field clump of Dendrocalamus giganteus (giant bamboo).
The shoots proliferated in a basal (MS) medium with 6 mgl−1 (26.6 μM) BA and 2% sucrose
and the rate of shoot proliferation gradually increased to over three-fold before in vitro
flowering took place. In vitro flowering was not the expression of a species-specific
mechanism believed to occur during gregarious flowering, as the mother clump did not
flower. The rate of shoot proliferation decreased at flowering, accompanied by reversion of
flowering. The development of axillary meristems into vegetative or generative shoots
depended on the level of BA. The possible role of BA, changes in the rate of shoot
proliferation decreased at flowering, accompanied by reversion of flowering. The
development of axillary meristems into vegetative or generative shoots depended on the level
of BA. Priyakumari and Sheela (2005) studied micropropagation of Gladiolus grandiflorus
L. cv. ‘Peach Blossom’ through axillary buds. The cultures were established using intact
cormels in MS medium containing different concentrations of growth regulators. Shoot
proliferation was maximum in MS medium fortified with BA 4 mg L-1+NAA 0.5 mg L-1 but
low concentrations of BA (1 or 2 mgL-1) was suitable for further shoot multiplication. In case
of root induction IBA (2 mgL-1) produced the earliest rooting (7 days) and the longest roots
(5 cm) and in vitro raised plantlets were successfully planted in sterile media consisting of
sand: soil (2:1) in plastic pots. Gonzalez et al. (2008) studied the micropropagation of
blueberry (Vaccinium corymbosum L. ‘Berkeley’), blackberry (Rubus sp. ‘Smoothstem’) and
raspberry (R. idaeus L. ‘Gradina’) from nodal segments of hardwood and softwood cuttings
of mature field-grown plants. The cultures successfully established were softwood from all

17
three species, and hardwood only from blueberry. Shoot-bud establishment from blueberry
was achieved by culturing explants in WPM salts with MS vitamins for 15 days, and then 30
days in the same medium with 18 mM zeatin. The best results of multiplication were
obtained in the same medium with 25 mM 2iP. For blackberry, shoot-bud establishment was
achieved by culturing explants in MS medium for 15 days, and then in the same medium
with 4 mM BA and 0.25 mM IBA while in raspberry explants were cultured in MS medium
for 15 days and then transferred to MS medium supplemented with 4 μm BA and 0.25 mM
IBA. After 30 days of culture, only 'Gradina' explants survived, from which shoot-bud
establishment was obtained in a modified MS medium with the same growth regulators.

18
Chapter-3
MATERIALS AND METHODS
Present study was conducted to find the morphological and genetic variation among and
within the genotypes of R. centifolia from Pakistan and USA. All three experiments of
morphological and genetic variation and in vitro propagation of R. centifolia were carried out
from 2009 to 2013. The research work was completed in Institute of Horticultural Sciences,
Centre of Agricultural Biochemistry and Biotechnology, University of Agriculture,
Faisalabad and Department of Horticultural Sciences, Texas A&M University, USA. Detail
of three experiments is discussed below.
3.1 Phenotypic plasticity among the genotypes of Rosa centifolia
3.1.1 Climate of Pakistan
Pakistan occupies an area of 7,96,096 square between the latitudes of 24° and 37° North and
longitudes of 61° to 75° East. A variety of climates, namely tropical, temperate, arid, and
coastal areas, exists in Pakistan because of its geography. The average high temperature is
23.9°C and an annual rainfall of 489 mm with almost 70% of precipitation occurring in the
Monsoon season (July-September).
3.1.2 Areas selected for morphological study
Punjab is heavily populated and a flourishing province of Pakistan located at 31.33° North
and 74.21° East with temperatures ranging between -2° and 45°C, but can reach 53°C in
summer and -5°C in winter. Many districts in Punjab have been growing R. centifolia for
many years and are increasing their acreage with time. Eight districts (Sargodha, Sialkot,
Lahore, Pattoki, Pakpattan, Kahror Pakka, Faisalabad and Sheikhupura) of Punjab were
selected for morphological and genetic study after observing high morphological diversity
among the landraces of R. centifolia in these districts (Table 3.1). Soil (Table 3.2) and water
(Table 3.3) samples used for growing these landraces were also collected and tested by the
Soil and Water Testing Laboratory, Ayub Agriculture Research Institute, Faisalabad,
Pakistan.

19
Table 3.1: The geographical position of selected study area in Punjab, Pakistan Code Collection site Latitude
(N) Longitude (E)
Altitude (m)
Rainfall (mm)
D1 Sargodha 32°33 72°73 193 441
D2 Sialkot 32°33 74°75 256 981 D3 Lahore 31°32 74°75 213 588
D4 Pattuki 31°32 73°74 186 413
D5 Pakpattan 30°31 73°74 155 256
D6 Kahror Pakka 29°30 71°72 123 167
D7 Faisalabad 31°32 73°74 184 256
D8 Shaikupura 31°32 74°75 236 588
(The New Oxford Atlas for Pakistan, 2006). Table 3.2: Analysis of water samples collected from study area of Punjab, Pakistan
Code Collection site Electrical Conductivity (EC µs/cm)
Sodium Absorption Ratio (SAR)
Residual Sodium Carbonate (RSC)
D1 Sargodha 367 0.38 Nil D2 Sialkot 201 0.03 0.02 D3 Lahore 1567 4.05 3.88 D4 Pattuki 976 0.35 Nil D5 Pakpattan 707 1.83 1.20 D6 Kahror Pakka 283 0.09 Nil D7 Faisalabad 1146 6.30 5.07 D8 Shaikupura 384 0.89 Nil
Table 3.3: Analysis of soil samples collected from different sites of Punjab, Pakistan
Code Collection site Electrical Conductivity (EC ms/cm)
Soil pH Organic Matter
Saturation (% age)
D1 Sargodha 1.99 7.7 0.36 38 D2 Sialkot 1.04 7.5 0.36 37 D3 Lahore 1.07 7.2 0.52 38 D4 Pattuki 1.89 7.5 0.47 40 D5 Pakpattan 1.16 7.4 0.26 40 D6 Kahror Pakka 1.15 7.4 0.88 42 D7 Faisalabad 1.09 7.3 0.36 40 D8 Sheikhupura 28.50 6.0 0.36 100

20
Figure 3.1 Map of Pakistan showing the production areas of Rosa centifolia in Punjab Province

21
3.1.3 Layout of experiment
Plants of similar maturity were selected from all the areas mentioned below and the same
type of pruning and other cultural practices were conducted. All the plants in selected areas
were propagated through cuttings.
3.1.3.1 Genotypes of Rosa centifolia from Pakistan Landraces were given the name on the basis of areas D1 Sargodha D2 Sialkot D3 Lahore D4 Pattoki D5 Pakpattan D6 Kahror pakka D7 Faisalabad D8 Sheikhupura
3.1.3.2 Genotypes of Rosa centifolia from USA G1 ‘Fantin-Latour’ G2 ‘Gros Choux d’ Hollande’ G3 ‘Centifolia Varigata’ G4 ‘De Meaux’ G5 ‘Cabbage Rose’1 G6 ‘Cabbage Rose’2 G7 ‘Pompon de Bourgogne’ G8 ‘Paul Ricault’ Cuttings of USA genotypes collected from Rogue Valley Roses, Oregon, USA grown in a
greenhouse at the Department of Horticultural Sciences, Texas A&M University, USA
(Table 4). Growing media used for these cuttings was Metro-Mix Professional Growing Mix
#900 composed of pine bark, vermiculite, Canadian sphagnum moss, perlite and dolomitic
limestone. Reverse osmosis water was used for irrigation
3.1.4 Morphological Parameters
Data on 24 different morphological parameters was collected from all genotypes according to
the plant descriptor (UPOV). Ten plants were randomly selected from each location and data
was registered from nine observations from each plant. In USA all plants were grown on one
site.

22
3.1.4.1 Plant parameters
1. Growth habit
a. miniature
b. dwarf
c. broad shrub
d. bed
e. climber
dwarf shrub broad bed climber
2. Plant height
a. short
b. medium
c. tall
3. Plant width
a. Shallow
b. Medium
c. Broad
4. Young shoot anthocyanin
a. absent or very light
b. light
c. medium
d. dark
e. very dark
5. Prickles
a. absent
b. present
6. Prickles shape of lower side
a. deep concave

23
b. concave
c. flat
d. convex
e. strong convex
strong convex flat concave strong concave
7. Short prickles number
a. absent or very few
b. few
c. medium
d. many
e. too many
8. Long prickles number
a. Absent or very few
b. Few
c. Medium
d. Many
e. Too many
3.1.4.2 Leaf parameters
1. Green colour
a. Very weak
b. Weak
c. Medium
d. Strong
e. Very strong
2. Glossiness of upper side
a. Very weak
b. Weak

24
c. Medium
d. Strong
e. Very strong
3. Leaflet cross section
a. Concave
b. Slightly concave
c. Flat
d. Slightly convex
e. Convex
4. Leaf undulation of margin
a. Very weak
b. Weak
c. Medium
d. Strong
e. Very strong
5. Terminal leaflet length of blade
a. Short
b. Medium
c. Long
6. Terminal leaflet width of blade
a. shallow
b. medium
c. broad
7. Terminal leaflet shape of base
a. type v
b. obtuse
c. round
d. heart shape

25
Type V Obtuse Round Heart shape
3.1.4.3 Flower parameters
1. Number of flowers on flowering shoot
a. Very few
b. Few
c. Medium
d. Many
e. Too many
2. Flower type
a. Single
b. Semi - single
c. Double
3. Number of petals
a. Very few
b. Few
c. Medium
d. Many
e. Too many
4. Flower diameter
a. Very small
b. Small
c. Medium
d. Big
e. Very big
5. Flower view from above

26
a. Round
b. Roundish
c. Star shape
6. Flower side view of upper part
a. Flat
b. Flat - convex
c. Convex
7. Flower side view of lower part
a. Concave
b. Flat
c. Flat - convex
d. Convex
8. Flower fragrance
a. Absent or very weak
b. Weak
c. Medium
d. Strong
e. Very strong
9. Petal size
a. Very small
b. Small
c. Medium
d. Big
e. Very big
10. Time of beginning of flower
a. Very early
b. Early
c. Medium
d. Late
e. Very late

27
3.1.5 Statistical Analysis
Data was subjected to Principle Component Analysis (PCA) in order to identify
morphological variations. Data analysis was performed using SAS statistical software
(Version 9.3, SAS, 2011 Institute Inc NC USA)
3.2 Genetic diversity among genotypes of Rosa centifolia
3.2.1 Plant material from Pakistan
Eight districts of Punjab were selected for this study. Three samples of young leaves of R.
centifolia were collected from each district and a total of 24 genotypes were selected from
eight different districts of Punjab, Pakistan. These leave samples were brought to Centre of
Agricultural Biochemistry and Biotechnology (CABB), University of Agriculture Faisalabad,
Pakistan for DNA extraction.
3.2.2 Plant material from USA
Eight genotypes of Rosa centifolia were collected from Rogue Valley Roses, Oregon, USA
and grown under greenhouse in Department of Horticulture, Texas A&M University, USA.
The cuttings were grown on Metro-Mix Professional Growing Mix #900 which was
composed of pine bark, vermiculite, Canadian sphagnum moss, perlite and dolomitic
limestone and reverse osmosis water was used for irrigation. At the time of producing leaves
young partially opened leaves were collected and stored at -20°C for DNA extraction in the
laboratory.
3.2.3 Rosa damascena samples
After comparing all samples of R. centifolia from Pakistan and USA, four samples of R.
damascena previously brought by from Isfahan and East Azerbaijan districts of Iran (Kiani et
al., 2010) and (Farooq et al., 2013) from Faisalabad and Pattoki districts of Pakistan were
brought Department of Horticultural Sciences, Texas A&M University and added in the
study.
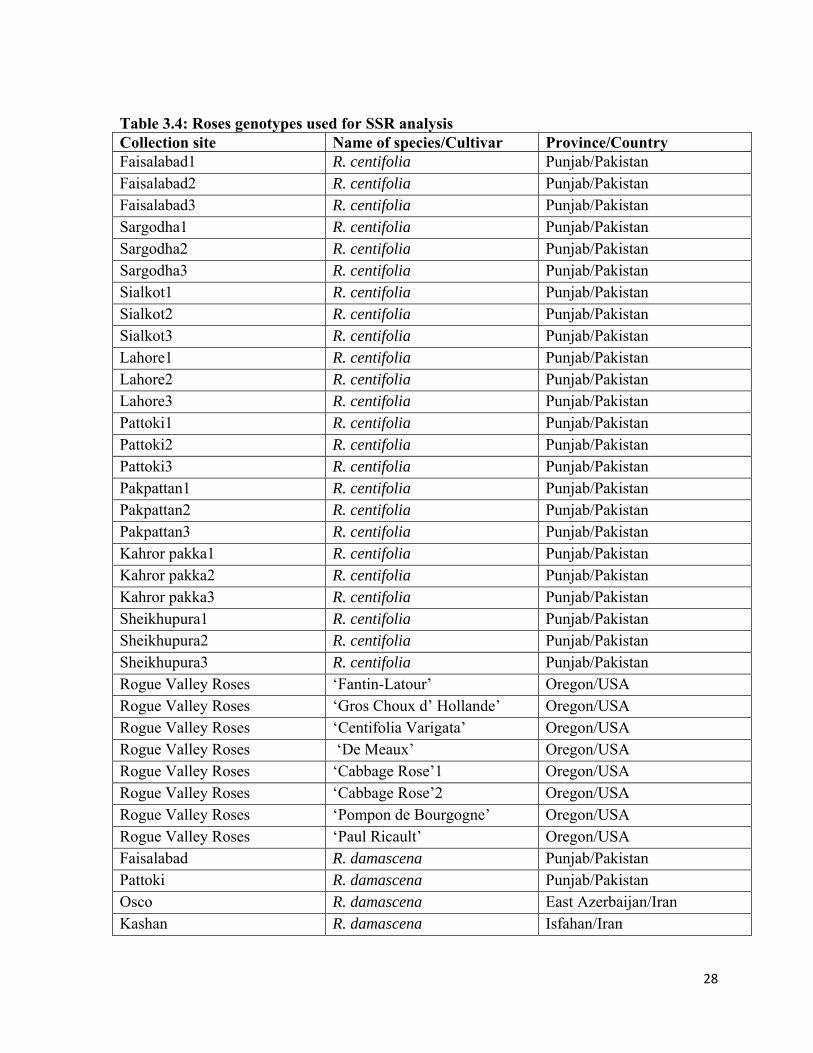
28
Table 3.4: Roses genotypes used for SSR analysis Collection site Name of species/Cultivar Province/CountryFaisalabad1 R. centifolia Punjab/Pakistan Faisalabad2 R. centifolia Punjab/Pakistan Faisalabad3 R. centifolia Punjab/Pakistan Sargodha1 R. centifolia Punjab/Pakistan Sargodha2 R. centifolia Punjab/Pakistan Sargodha3 R. centifolia Punjab/Pakistan Sialkot1 R. centifolia Punjab/Pakistan Sialkot2 R. centifolia Punjab/Pakistan Sialkot3 R. centifolia Punjab/Pakistan Lahore1 R. centifolia Punjab/Pakistan Lahore2 R. centifolia Punjab/Pakistan Lahore3 R. centifolia Punjab/Pakistan Pattoki1 R. centifolia Punjab/Pakistan Pattoki2 R. centifolia Punjab/Pakistan Pattoki3 R. centifolia Punjab/Pakistan Pakpattan1 R. centifolia Punjab/Pakistan Pakpattan2 R. centifolia Punjab/Pakistan Pakpattan3 R. centifolia Punjab/Pakistan Kahror pakka1 R. centifolia Punjab/Pakistan Kahror pakka2 R. centifolia Punjab/Pakistan Kahror pakka3 R. centifolia Punjab/Pakistan Sheikhupura1 R. centifolia Punjab/Pakistan Sheikhupura2 R. centifolia Punjab/Pakistan Sheikhupura3 R. centifolia Punjab/Pakistan Rogue Valley Roses ‘Fantin-Latour’ Oregon/USA Rogue Valley Roses ‘Gros Choux d’ Hollande’ Oregon/USA Rogue Valley Roses ‘Centifolia Varigata’ Oregon/USA Rogue Valley Roses ‘De Meaux’ Oregon/USA Rogue Valley Roses ‘Cabbage Rose’1 Oregon/USA Rogue Valley Roses ‘Cabbage Rose’2 Oregon/USA Rogue Valley Roses ‘Pompon de Bourgogne’ Oregon/USA Rogue Valley Roses ‘Paul Ricault’ Oregon/USA Faisalabad R. damascena Punjab/Pakistan Pattoki R. damascena Punjab/Pakistan Osco R. damascena East Azerbaijan/Iran Kashan R. damascena Isfahan/Iran

29
3.2.4 Primers
Thirteen Primers were selected from Kiani et al. (2010) and Hibrand-Saint Oyant et al.
(2008) work.
Table 3.5: List of primers
# Primer code Primer sequence Expected size (bp)
Annealing Temp. °C
1 RW3K19 F: GCCATCACTAACGCCACTAAA R: GCGTCGTTCGCTTTGTTT
418 56
2 RW3N19 F: CTGGCTGGTTCTCTTTCTG R:ATGGGTCGTCGTCGATATG
125 54
3 RW14H21 F: ATCATGTGCAGTCTCCTGGT R:AATTGTGGGCTGGAAATATG
145 55
4 RW1717 F: CAGGTAATTTGCGGATGAAG R: GATCCGCCGTTTCCAGT
216 58
5 RW18N19 F: CCCGAGAAAGAGACAGTAAA R: ATCGAGAGAGACACCGACTC
250 54
6 RW22A3 F: AGAGAATTGAAAAGGGCAAG R: GAGCAAGCAAGACACTGTAA
150 53
7 RW22B6 F: ACAGTGAGTTGTTCGCTTCT R: TTCATTGCTAGGAAGCAGTA
158 54
8 RW32D19 F: GAAGTCCAGAGCCAATTCCA R: AGGGTCCTCATCCACCACTT
540 54
9 RW55C6 F: GTGGATTTTCAGAGATACGC R: TCACAGACAGGACCACCTAT
265 50
10 RW55D22 F: GATCCGTTTAAGTAACCTTT R: CCACAAGGATTCTGATTTAT
201 51
11 H22CO1 F: TCATAACCAACCATCTCCATCA R: AGGATTTCACCCAGAACACG
228 51
12 H24D11 F: CCTCCTCAGCTTTCCTCCTT R: CAGCAACCATCTCTTCGTGA
168 55
13 RW10M24 F: TTAATCCAAGGTCAAAGCTG R: TCTCTTTCCCTCCTCACTCT
252 53
(All primers were obtained from Applied Biosystems Inc., Foster City, CA, USA)
3.2.5 DNA extraction protocol
DNA was extracted by CTAB methods of Doyle and Doyle, (1987), Boonprakob, (1996) and
Jan, (1996) with some modifications. Approximately 50 mg of frozen leaf tissue was put into
a 1.5 mL microtube and cooled by adding liquid nitrogen. A handheld, battery operated
power drill was used to grind the tissue with a chilled plastic pestle, sized to fit the tubes,
used in place of a drill bit. 700 μL of 2X CTAB were added and thoroughly mixed in the
tubes. Samples were incubated in a 65°C water bath for 2.5 h and then cooled to room

30
temperature. Next, 700 μL of chloroform: iso-amyl alcohol, 24:1 were mixed with the
samples. Centrifugation was done at 13,200 gn for 10 min, and repeated if the upper layer of
29 sample will not yet clear and colorless. The upper layer of the samples was then moved to
a new 1.5 mL tube and combined with another 700 μL of CIA. The above centrifugation
steps were repeated. The upper layer was again shifted to a new tube, and then mixed with
chilled isopropanol. After inverting the tubes several times, the samples were placed in a -
20°C freezer overnight. The next day, samples were centrifuged at 6,000 gn for 10 min. The
supernatant was discarded, taking care not to dislodge the pellet at the bottom of the
microtube. Samples were allowed to dry briefly, followed by a 70% ethanol rinse, which was
repeated once, with a short centrifugation in between to ensure that the pellet would be
retained in the tube. After the last ethanol rinse was poured off, samples were allowed to dry
completely at room temperature. To resuspend the samples, 50-200 μL of TE buffer/sample
was used. Samples were vortexed for about 10 min, or until the pellet was completely
dissolved.
3.2.6 DNA Quantification
DNA quantification of all samples was done by using Hoefer DQ 300 fluorometer. DNA
dilutions of 5 ng/μl were made by using nuclease free water.
3.2.7 PCR (Polymerase Chain Reaction) Amplification
PCR reactions were performed in Eppendorf PCR machine with a total volume of 10μl of all
PCR reagents, which includes 8.0μL Go Taq Master Mix (New England BioLabs, Inc.),
0.5+0.5 μl forward and reverse primers and 1 μl DNA. All the reaction mixture and reagents
were kept on ice during and after making PCR mixture. For PCR reaction the following
conditions were used: heating started at 105°C, denaturation at 98°C for 10 sec and 30 cycles
of 98°C for 1 s : 55°C for 5 s : 72°C for 1 min, with final extension of 72°C for 1 min and a
final hold at 4°C. PCR products were stored at -20°C.
3.2.8 Gel electrophoresis (GE)
1% agarose gel was used for GE. 1μl ethidium bromide and 3μl of dye were mixed in all
PCR products by centrifuging them for few seconds. PCR products were loaded on the gel
and run at 155 mA and 81 volt for 45 min. DNA GE was photographed under UV
transilluminator.

31
3.2.9 Data analysis
GeneMapper v. 4.0, and Peak Scanner v. 1.0 (Applied Biosystems, Inc.) were used for
fragment data analysis. NTSYS-pc v. 2.2 (Numerical Taxonomy and Multivariate Analysis
System, Exeter Software) was used for cluster analysis of the accessions to help visualize
relationships. A data set will be created first from the SSR data in Microsoft Excel (Microsoft
Corp.) with the alleles as the rows, and the accessions as the columns, using “1” for presence
of an allele and “0” for absence. Dice coefficient was used to calculate similarity coefficients
(Nei and Li, 1979) and UPGMA clustering was used to create a dendrogram.
3.3 IN VITRO PROPAGATION OF ROSA CENTIFOLIA
The present investigation on in vitro propagation of Rosa centifolia was carried out in the
Plant Tissue Culture Cell Institute of Horticultural Sciences, University of Agriculture,
Faisalabad during the period of 2009-2012.
3.3.1 Plant Material
For in vitro propagation, healthy plants of Rosa centifolia present in the Rosa Project,
Institute of Horticultural Sciences, University of Agriculture, Faisalabad were used as source
of explant. Young well developed axillary buds were used as explant (Ishioka and Tanimoto,
1990), which were collected from healthy, disease free and recently grown young shoots of
R. centifolia in the field.
3.3.2 Media
Murashige and Skoog, (1962) media was used as basal media in this experiment. As it is the
most used media for propagation of roses through plant tissue culture (Skirvin et al., 1990;
Vijaya and Satrayana, 1991). The only modifications in the basal MS media were made by
adding various concentrations of different growth regulators.
3.3.3 Preparation of stock solutions
Murashige and Skoog (MS) medium was used in all the treatments. Stock solutions of its
major components were made with distilled water by the following way and then stored in
refrigerator at 4°C.
Macro nutrients – 1000 ml
Micro nutrients – 1000 ml
Vitamin – 100 ml
Iron – 100 ml

32
3.3.4 Stock solutions for growth regulator
Different types of cytokinins (kinetin and 6-benzylamino purine) and auxin (indol-3-butyric
acid and naphthalene acetic acid) of different concentrations were used for shoot and root
formation. These were first completely dissolved in few drops of 1N NaOH and required
volumes were made by adding distilled water.
3.3.5 Preparation of media
MS (Murashige and Skoog, 1962) media for 1000 ml was prepared by the following way
Table 3.6: Ingredients of MS media (Stock solution)
Macronutrients 100ml/l
Micronutrients 1ml/l
Vitamins 10ml/l
Iron 10ml/l
Sugar 30g/l
Agar 8 g/l
First macro, micro, vitamins, iron and growth regulators were added in a beaker. The pH was
measured with a help of digital pH meter after dissolving sugar in it as a carbon source, and
adjusted at 5.7 by using either 0.1 N HCl or NaOH. On the other hand agar was boiled in
separate beaker, mixed with the solution and the final volume was consisted to 1000ml. Then
10ml of this warm MS media was dispensed in each test tube, which was covered with
polythene and tied with a rubber band. All the solutions were made in sterile distilled water.
To sterilize the media, all test tubes with media were placed in the autoclave at 121°C and
15psi pressure for 20 minutes. After sterilization media was allowed to cool and placed in the
growth room until further use. The MS media was also supplemented with different levels of
growth regulators for root induction and shoot formation (Table 3.7)

33
Table 3.7: Treatments for shoot formation
T1 MS T2 MS + BAP 0.2 µM T3 MS + BAP 0.4 µM T4 MS + BAP 0.6 µM T5 MS + BAP 0.8 µM T6 MS + KIN 0.2 µM T7 MS + KIN 0.4 µM T8 MS + KIN 0.6 µM T9 MS + KIN 0.8 µM T10 MS + BAP 0.2 µM + KIN 0.2 µM T11 MS + BAP 0.4 µM + KIN 0.2 µM T12 MS + BAP 0.6 µM + KIN 0.2 µM T13 MS + KIN 0.8 µM + BAP 0.2 µM T14 MS + KIN 0.2 µM + BAP 0.2 µM T15 MS + KIN 0.4 µM + BAP 0.2 µM T16 MS + KIN 0.6 µM + BAP 0.2 µM T17 MS + KIN 0.8 µM + BAP 0.2 µM
Table 3.8: Treatments for root formation
T1 MS T2 MS + IBA 0.2 µM T3 MS + IBA 0.4 µM T4 MS + IBA 0.6 µM T5 MS + NAA 0.2 µM T6 MS + NAA 0.4 µM T7 MS + NAA 0.6 µM
3.3.6 Sterilization of explant
Selected explants were brought in the lab, with leaves removed and placed under the running
water for 30 minutes. For surface sterilization explants were first immersed in 70% ethanol
for 5 minutes and washed three times with sterile autoclave water to remove the residues of
ethanol. After this preliminary ethanol treatment explants were sterilized with 5% sodium
hypochlorite (Bleach) for 3 minutes and again rinsed with sterile autoclaved water to remove
the residues of bleach.
3.3.7 Inoculation
All operation of inoculation was performed in laminar air flow cabinet, which was sterilized
with 100% ethanol before transferring all required materials like media, spirit lamp, lighter,
glassware etc. Inoculation room was sterilized by turning on UV light for half an hour. Now

34
sterilized explants were brought into the laminar air flow cabinet and cut into proper size by a
slanting cut. The cut portions of explants were inserted in the medium. After inoculation test
tubes were again closed with polythene, tied with rubber band and placed in the growth room
at 25±2°C, 16 hours photoperiod and 3000 Lux cool white inflorescent light.
3.3.8 Sub culture
Shoots produced from the explants were separated and cultured on the half strength rooting
MS media under sterile conditions of laminar air flow cabinet with sterile dissecting scissor
and forceps. Explants were also subcultured in the same fresh MS media when dryness or
browning at the lower end was observed.
3.3.9 Hardening
Young rooted plantlets were taken from the test tubes and washed thoroughly with distilled
water to ensure removal of traces of agar from their roots. Before being transplanted in the
small plastic pots containing compost as growing media, which was first autoclaved at 121°C
for 20 minutes. Pots with rooted plantlets were covered with polythene to increase humidity
and irrigated. After 4 or 5 days one corner of polythene was lifted, then another corner and at
the end the whole polythene cover was removed. After that pots with plantlets were in a
greenhouse.
3.3.10 Collection of Data
3.3.10.1 Number of days for shoot initiation
The number of days taken from the date of inoculation to first shoot initiation were recorded
and expressed as the number of days to initiate shoot.
3.3.10.2 Number of shoots
The number of shoots were counted at time of transferring the explants in rooting media.
3.3.10.3 Number of leaves per shoot
The number of leaves were counted from each explant before transferring the shoots in
rooting media.
3.3.10.4 Shoot length
Shoot length was measured from tip of shoot to the end of shoot when it was transferred in
rooting media with a scale.

35
3.3.10.5 Number of days to initiate roots
Days from transferring of shoots into rooting media to initiation of roots were counted and
expressed as number of days to initiation of root.
3.3.10.6 Number of roots
The number of roots was counted when plantlets were transferred into small pots.
3.3.10.7 Root length
From each shoot the length of the longest roots was measured from the collar region to the
highest root tip as a root length and expressed in cm.
3.3.11 Data analysis
Data was analyzed statistically by using SAS statistical analysis (Version 9.3, SAS, 2011
Institute Inc NC USA) and differences among treatments were compared by using Duncan’s
Multiple Range (DMR) test.

36
Chapter-4
RESULTS
4.1 Phenotypic plasticity within and among the genotypes of Rosa centifolia
of Pakistan and USA
4.1.1 Cluster analysis for different morphological characters of genotypes of Rosa
centifolia of Pakistan
Diversity among eight different genotypes of R. centifolia found in Punjab (Pakistan) was
done by constructing a dendrogram consisting of eight genotypes on the basis of
morphological characters by using Statistica 7 software and delineated in figure 4.1. All the
genotypes are divided into three groups at a linkage distance of 1.0. There are two genotypes
in group A, namely Sheikhupura and Pakpattan. Group B was the largest group consisting of
Kahror Pakka, Faisalabad, Pattoki, and Lahore. Group C consists of genotypes Sialkot and
Sargodha. Group B, the largest of the dendrogram, was again divided into two groups at a
linkage distance of 0.6. First subgroup was made up of D6 (Kahror Pakka) and other with
Faisalabad, Pattoki and Lahore genotypes. Genotypes of Faisalabad were at the closest to
Pattoki and at the greatest distance to Sialkot.
4.1.2 Principle component analysis for different morphological characters of genotypes
of Rosa centifolia
The results of different characters of genotypes of R. centifolia were explained by a plot in
Figure 4.2. Principle component analysis showed 35.74% and 34.29% of total variation was
due to factor 1 and factor 2, respectively. All genotypes of R. centifolia collected from
different the districts were in the same group. The genotypes were distributed into groups in
the plot, which were in accordance with the results of cluster analysis in Figure 4.2.
4.1.3 Correlation between morphological parameters of Rosa centifolia genotypes from
Pakistan
Nine important morphological parameters were selected to estimate their correlation (Table
4.1). Number of small prickles was negatively correlated with the number long prickle, leaf
size, number of prickles on flower pedicel, number of flowers, number of petals per flower,
flower fragrance (-0.314, -0.087, -0.090, -0.087, -0.040, and -0.027, respectively). Number of
small prickles was positively correlated with all other parameters. Longer prickles number

37
was positively correlated with all other parameters except number of prickles on flower
pedicel. The leaf size also had negative association with number of prickles on flower
pedicel, number of flowers, and number of petals per flower and a positive correlation with
rest of the parameters included in the study. Leaf green color and undulation of leaf margin
showed positive and strong association with all parameters. Negative association of number
of prickles on flower pedicel was found with all the parameters except for flower diameter.
Flower number, petal number, and flower diameter showed positive correlation with rest of
parameters. Flower fragrance was positively correlated with all parameters except number of
small prickles and number of prickles on flower pedicel.
D1 = Sargodha D2 = Sialkot D3 = Lahore D4 = Pattoki D5 = Pakpattan D6 = Kahror Pakka D7 = Faisalabad D8 = Sheikhupura Figure 4.1: Cluster analysis of genotypes of Rosa centifolia from Pakistan

38
D1 = Sargodha D2 = Sialkot D3 = Lahore D4 = Pattoki D5 = Pakpattan D6 = Kahror Pakka D7 = Faisalabad D8 = Sheikhupura Figure 4.2: Principle component analysis of genotypes of Rosa centifolia
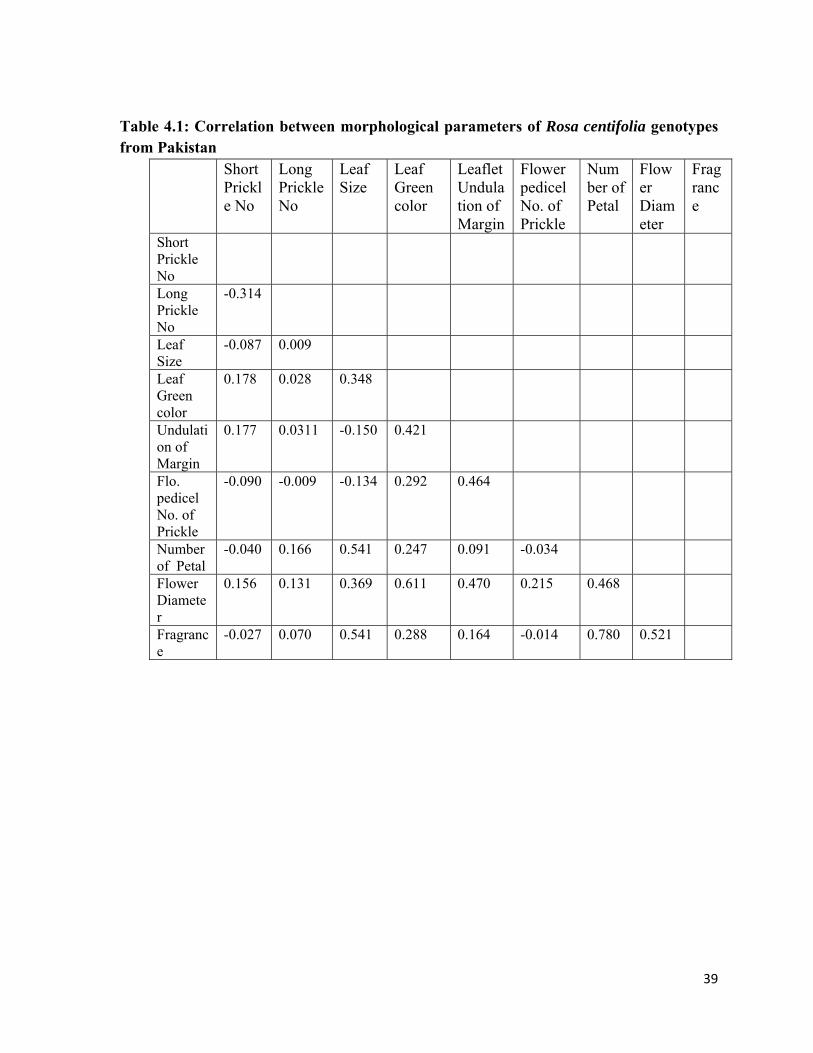
39
Table 4.1: Correlation between morphological parameters of Rosa centifolia genotypes from Pakistan
Short Prickle No
Long Prickle No
Leaf Size
Leaf Green color
Leaflet Undulation of Margin
Flower pedicel No. of Prickle
Number of Petal
Flower Diameter
Fragrance
Short Prickle No
Long Prickle No
-0.314
Leaf Size
-0.087 0.009
Leaf Green color
0.178 0.028 0.348
Undulation of Margin
0.177 0.0311 -0.150 0.421
Flo. pedicel No. of Prickle
-0.090 -0.009 -0.134 0.292 0.464
Number of Petal
-0.040 0.166 0.541 0.247 0.091 -0.034
Flower Diameter
0.156 0.131 0.369 0.611 0.470 0.215 0.468
Fragrance
-0.027 0.070 0.541 0.288 0.164 -0.014 0.780 0.521

40
4.1.4 Cluster analysis for different morphological characters of different varieties of
Rosa centifolia of USA
Cluster analysis Figure 4.3 showed the diversity among eight different varieties of Rosa
centifolia grown in the USA. This diversity was determined through 26 morphological
characters. In the dendrogram all eight cultivar of R. centifolia are differentiated into three
main groups at the linkage distance of 1.00. Among these three groups group A and C are the
shortest groups containing only genotypes ‘Fantin-Latour’ and ‘Rosa de Meaux’,
respectively. These two varieties were maximum apart and variant from each other. Group B
is the second largest group in the dendrogram, containing the remaining 6 varieties and again
divided into 4 subgroups at a linkage distance of 0.76. First three subgroups of group B were
consisted of ‘Centifolia Varigata’, ‘Cabbage Rose’1 and ‘Pompon de Bourgogne’,
respectively. The fourth subgroup combines ‘Paul Ricault’, ‘Cabbage Rose’ 2 and ‘Gros
Choux d’ Hollande’ together.
4.1.5 Principle component analysis for different morphological characters of different
varieties of R. centifolia of the USA
Principle component analysis was conducted on various morphological parameters of
different varieties of centifolia found in the USA and a total variation of 47.78% and 39.75%
was documented by factor 1 and factor 2, respectively (Figure 4.4). All genotypes were
grouped in one block in the plot. All different types of R. centifolia were divided into three
groups. Two varieties with the greatest distance, G1 and G6 were in two separate groups,
while all the remaining ones found in the same group. These results of principle component
analysis are according to the preceding results of results of cluster analysis.
4.1.6 Correlation between morphological parameters of R. centifolia varieties of the
USA
Eleven important morphological parameters were selected to study correlation among them
(Table 4.2). There was a negative correlation of the number of small prickles with undulation
of leaf margin (-0.071) but a positive one with all other characteristics. Number of long
prickles had negative association with all characters except for the numbers of prickles on
flower pedicel and with the number petal. Leaf size was correlated positively with all
parameters except the number of prickles on flower pedicel. Leaf green color was negatively
correlated with the number of prickles on flower pedicel. Undulation of leaf margin had
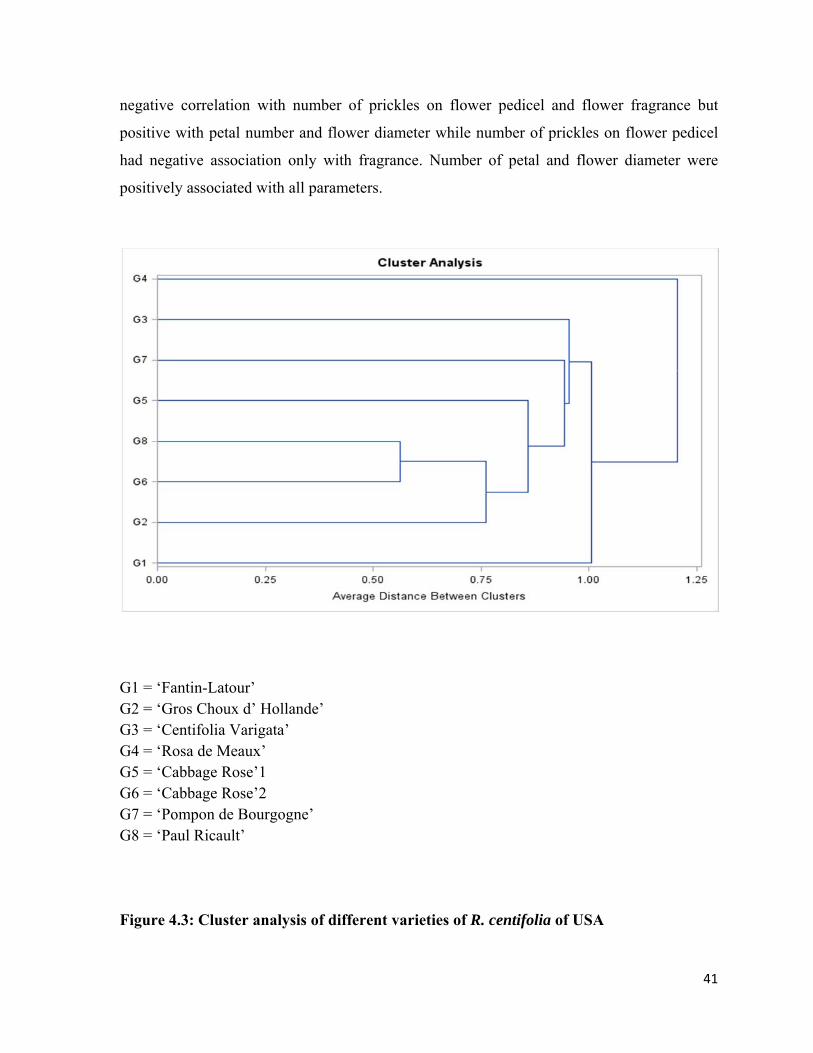
41
negative correlation with number of prickles on flower pedicel and flower fragrance but
positive with petal number and flower diameter while number of prickles on flower pedicel
had negative association only with fragrance. Number of petal and flower diameter were
positively associated with all parameters.
G1 = ‘Fantin-Latour’ G2 = ‘Gros Choux d’ Hollande’ G3 = ‘Centifolia Varigata’ G4 = ‘Rosa de Meaux’ G5 = ‘Cabbage Rose’1 G6 = ‘Cabbage Rose’2 G7 = ‘Pompon de Bourgogne’ G8 = ‘Paul Ricault’ Figure 4.3: Cluster analysis of different varieties of R. centifolia of USA

42
G1 = ‘Fantin-Latour’ G2 = ‘Gros Choux d’ Hollande’ G3 = ‘Centifolia Varigata’ G4 = ‘Rosa de Meaux’ G5 = ‘Cabbage Rose’1 G6 = ‘Cabbage Rose’2 G7 = ‘Pompon de Bourgogne’ G8 = ‘Paul Ricault’ Figure 4.4: Principle component analysis of different varieties of Rosa centifolia of USA

43
Table 4.2: Correlation between morphological parameters of Rosa centifolia genotypes of USA
No. of Short Prickle
No. of Long Prickle
Leaf Size
Leaf Green Color
Leaflet Undulation of Margin
No. of Prickles on Flower pedicel
No. of Petal
Flower Diameter
Fragrance
No. Short Prickle
No. Long Prickle
0.404
Leaf Size
0.298 -0.193
Leaf Green Color
0.086 -0.371 0.455
Undulation of Margin
-0.071 -0.410
0.756 0.401
No. of Prickles on Flower pedicel
0.795 0.404 -0.027 -0.374 -0.071
No. of Petal
0.324 0.093 0.342 0.072 0.422 0.324
Flower Diameter
0.444 -0.023 0.746 0.571 0.688 0.181 0.706
Fragrance
0.082 -0.259 0.156 0.460 -0.137 -0.312 0.180 0.347

44
4.1.7 Cluster analysis for different morphological characters of different varieties of R.
centifolia of Pakistan and USA
After separate analysis of genotypes of R. centifolia from Pakistan and USA, a cluster
analysis was made to determine morphological relationships among all genotypes together
(Figure 4.5). All genotypes were divided into 3 groups at a linkage distance of 1.13. In first
group, only ‘De Meaux’ genotype was present. The second group was made up of ‘Cabbage
Rose’1 and ‘Pompon de Bourgogne’ genotypes. The third group was the largest which was
further divided into two sub groups, one made up of ‘Fantin-Latour’ and the other subgroup
containing all other remaining genotypes from USA and Pakistani genotypes.
D1 = Sargodha G1 = ‘Fantin-Latour’ D2 = Sialkot G2 = ‘Gros Choux d’ Hollande’ D3 = Lahore G3 = ‘Centifolia Varigata’ D4 = Pattoki G4 = ‘De Meaux’ D5 = Pakpattan G5 = ‘Cabbage Rose’1 D6 = Kahror Pakka G6 = ‘Cabbage Rose’2 D7 = Faisalabad G7 = ‘Pompon de Bourgogne’ D8 = Sheikhupura G8 = ‘Paul Ricault’
Figure 4.5: Cluster analysis of 8 genotypes of R. centifolia from Pakistan and 8 varieties from USA

45
Figure 4.6: Undulation of leaf margins of different genotypes of R. Centifolia; a. week undulation of leaf margins, b. strong undulation of leaf margins
Figure 4.7: Terminal leaflet of different genotypes of Rosa Centifolia; a. leaflet with greater width, b. leaflet with greater length
A B
A B

46
Figure 4.8: Terminal leaflet shape of base of different genotypes of Rosa Centifolia; a. leaflet with round base, b leaflet with heart shape base
Figure 4.9: Number of thorns of different genotypes of Rosa Centifolia; a. stem with more number of thorns, b. stem with less number of thorns
A
A B
B

47
Figure 4.10: Number of petals per flower of different genotypes of Rosa Centifolia; a. flower with greater number of petals, b. flower with less number petals
A B

48
4.2 Genetic diversity among genotypes of Rosa centifolia
4.2.1 Genetic analysis of Rosa centifolia of Pakistan
All the primers produced scorable bands and displayed polymorphism among genotypes.
These thirteen primers produced 251 bands out of which 54 were polymorphic.
Unweighted paired-group of arithmatic mean (UPGMA) analysis of Dice similarity
distributed all genotypes of Rosa centifolia into four groups at 0.68 similarity coefficient
(Figure 4.6). Group-A contained all three genotypes of Kahror pakka, closer to the Sargodha
genotypes of group-B which was composed of all three samples from district Sargodha and
1st and 2nd samples of district Sialkot. Group-C was the largest group and had all genotypes
from district Faisalabad, Sheikhupura and Lahore with 3rd genotype of district Sialkot. All
three genotypes of the district Pakpattan and Pattoki were in the separate group-D. Group-B
was further divided into two subgroups at similarity coefficient of 0.78. One contained three
genotypes of Sargodha and the other one containing the first two genotypes of district
Sialkot. At same 0.78 value of similarity coefficient group-C was also divided into two
subgroups. The first consisted of three genotypes of Faisalabad along with 3rd genotype of
Sargodha district. Second group contained all three genotypes of district Lahore, which were
also closer to Pakpattan genotypes of group-D. Genotypes of district Pattoki were genetically
less similar to genotypes of district Kahror pakka of group-A. Genotypes from Faisalabad
district were genetically more similar to Sheikhupura and Sialkot genotypes but genetically
diverse from genotypes of Kahror Pakka and Pattoki of group A and D, respectively.

49
1. Sargodha1 13. Pakpattan1 2. Sargodha2 14. Pakpattan2 3. Sargodha3 15. Pakpattan3 4. Siakot1 16. Kahror pakka1 5. Siakot2 17. Kahror pakka2 6. Siakot3 18. Kahror pakka3 7. Lahore1 19. Faisalabad1 8. Lahore2 20. Faisalabad2 9. Lahore3 21. Faisalabad3 10. Pattoki1 22. Sheikhupura1 11. Pattoki2 23. Sheikhupura2 12. Pattoki3 24. Sheikhupura3
Plate 4.1: Primer (RW3N19) with the DNA samples of Rosa centifolia from different districts of Punjab, Pakistan
1 2 3 4 5 6 7 8 9 10 11 12
1 4 15 16 17 18 19 20 21 22 23 24
RW3N19

50
1. Sargodha1 13. Pakpattan1 2. Sargodha2 14. Pakpattan2 3. Sargodha3 15. Pakpattan3 4. Siakot1 16. Kahror pakka1 5. Siakot2 17. Kahror pakka2 6. Siakot3 18. Kahror pakka3 7. Lahore1 19. Faisalabad1 8. Lahore2 20. Faisalabad2 9. Lahore3 21. Faisalabad3 10. Pattoki1 22. Sheikhupura1 11. Pattoki2 23. Sheikhupura2 12. Pattoki3 24. Sheikhupura3
Plate 4.2: Primer (RW14H21) with the DNA samples of Rosa centifolia from different districts of Punjab, Pakistan
RW22A3
1 2 3 4 5 6 7 8 9 10 11 12 13
1 4 15 16 17 18 19 20 21 22 23 24

51
1. Sargodha1 13. Pakpattan1 2. Sargodha2 14. Pakpattan2 3. Sargodha3 15. Pakpattan3 4. Siakot1 16. Kahror pakka1 5. Siakot2 17. Kahror pakka2 6. Siakot3 18. Kahror pakka3 7. Lahore1 19. Faisalabad1 8. Lahore2 20. Faisalabad2 9. Lahore3 21. Faisalabad3 10. Pattoki1 22. Sheikhupura1 11. Pattoki2 23. Sheikhupura2 12. Pattoki3 24. Sheikhupura3
Plate 4.3: Primer (RW55C6) with the DNA samples of Rosa centifolia from different districts of Punjab, Pakistan
RW55C6
1 2 3 4 5 6 7 8 9 10 11 12 13
1 4 15 16 17 18 19 20 21 22 23 24

52
Figure 4.11: Dendrogram of microsatellite diversity among 8 genotypes of Rosa centifolia of Pakistan.

53
Table 4.3: Grouping of 24 genotypes of Rosa centifolia on the basis of microsatellite
markers
Groups Number of genotypes Name of genotypes
A 3 Kahror pakka1, Kahror pakka2, Kahror pakka3
B 5 Sargodha1, Sargodha2, Sargodha3, Sialkot1 Sialkot2
C 10 Sialkot3, Faisalabad1, Faisalabad2, Faisalabad3, Sheikhupura1, Sheikhupura2, Sheikhupura3, Lahore1, Lahore2, Lahore3
D 6 Pakpattan1, Pakpattan2, Pakpattan3, Pattoki1, Pattoki2, Pattoki3
4.2.2 Genetic analysis of Rosa centifolia of Pakistan and USA
Eight genotypes (‘Fantin-Latour’, ‘Gros Choux d’Hollande’, ‘Centifolia Varigata’, ‘De
Meaux, ‘Cabbage Rose’ 1, ‘Cabbage Rose’ 2, ‘Pompon de Bourgogne’, ‘Paul Ricault’) R.
centifolia used to grown in USA were bought from Oregon state, USA and grown in the
Department of Horticulture, Texas A&M University, USA. Same thirteen microsatellite
markers were previous used. Two genotypes of R. centifolia from district Faisalabad and
Sargodha of Pakistan were also compared with these eight different centifolia of USA to
investigate genetic linkage among genotypes of R. centifolia from Pakistan and USA.
All thirteen SSR primers run with 10 different centifolia (8 USA and 2 Pakistan). Total 169
bright DNA fragments were produced from 10 genotypes by using thirteen SSR primers out
of these 169, 13 loci were found polymorphic. Unweighted paired-group of Arithmatic
Means (UPGMA) analysis of Dice similarity made four groups of all genotypes at similarity
coefficient of 0.82. Two Pakistani genotypes (R. centifolia Faisalabad and R. centifolia
Sargodha) were in the group-C along with ‘Cabbage rose’ 2. ‘Centifolia Varigata’, ‘Cabbage
Rose’1 and ‘Pompon de Bourgogne’ were in group-B, which was further divided into two
subgroups. One of them contained ‘Cabbage Rose’1 and ‘Pompon de Bourgogne’ while
other had ‘Centifolia Variegata’. Group-A was made up of three genotypes ‘Gros Choux
d’Hollande’, ‘De Meaux’ and ‘Fantin-Latour’. At 0.88 similarity coefficient Group-A was
further divided into two subgroups. First subgroup was occupied ‘Gros Choux d’Hollande’
and ‘De Meaux’ and the second had ‘Fantin-Latour’. Genotype ‘Fantin-Latour’ of group-A
was at maximum genetic distance with Pakistani genotypes. Both Pakistani genotypes were

54
closer to each other and were in same group with ‘Cabbage Rose’ 2. The Results of thirteen
SSR markers showed that Pakistani genotypes were also genetically closer to genotype and
genetically more divergent from genotype ‘Fantin Latour’ of group-A. ‘Paul Ricault’ was in
separate group-D. This showed that ‘Paul Ricault’ is genetically different from all genotypes
of R. centifolia.
G1. ‘Fantin-Latour’ G2. ‘Gros Choux d’ Hollande’ G3. ‘Centifolia Varigata’ G4. ‘De Meaux’ G5. ‘Cabbage Rose’1 G6. ‘Cabbage Rose’2 G7. ‘Pompon de Bourgogne’ G8. Rosa centifolia Faisalabad G9. Rosa centifolia Sargodha G10. ‘Paul Ricault’
Plate 4.4: Primer (RW10M24) with the 2 DNA samples of Rosa centifolia of Pakistan and eight from USA
RW10M24
10 9 8 7 6 5 4 3 2 1

55
G1. ‘Fantin-Latour’ G2. ‘Gros Choux d’ Hollande’ G3. ‘Centifolia Varigata’ G4. ‘De Meaux’ G5. ‘Cabbage Rose’1 G6. ‘Cabbage Rose’2 G7. ‘Pompon de Bourgogne’ G8. Rosa centifolia Faisalabad G9. Rosa centifolia Sargodha G10. ‘Paul Ricault’
Plate 4.5: Primer (RW3N19) with the 2 DNA samples of Rosa centifolia of Pakistan and eight from USA
RW3N19
10 9 8 7 6 5 4 3 2 1

56
G1. ‘Fantin-Latour’ G2. ‘Gros Choux d’ Hollande’ G3. ‘Centifolia Varigata’ G4. ‘De Meaux’ G5. ‘Cabbage Rose’1 G6. ‘Cabbage Rose’2 G7. ‘Pompon de Bourgogne’ G8. Rosa centifolia Faisalabad G9. Rosa centifolia Sargodha G10. ‘Paul Ricault’
Plate 4.6: Primer (RW18N19) with the 2 DNA samples of Rosa centifolia of Pakistan and eight from USA
RW18N19
10 9 8 7 6 5 4 3 2 1
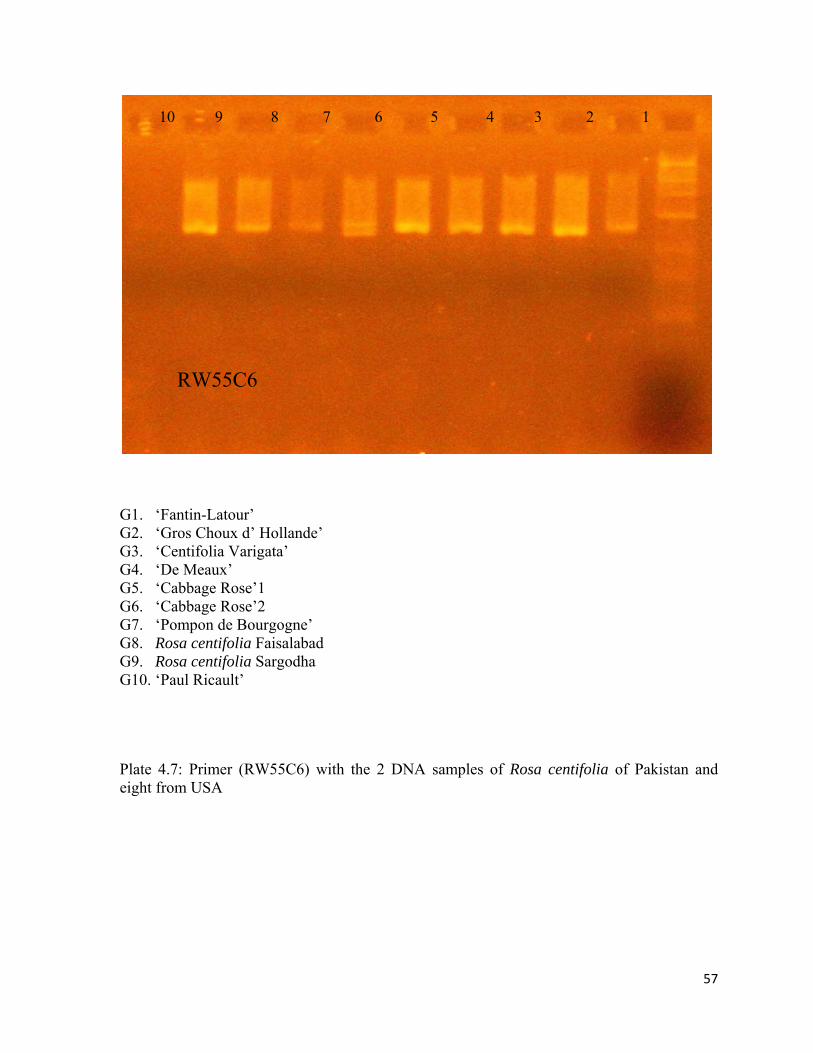
57
G1. ‘Fantin-Latour’ G2. ‘Gros Choux d’ Hollande’ G3. ‘Centifolia Varigata’ G4. ‘De Meaux’ G5. ‘Cabbage Rose’1 G6. ‘Cabbage Rose’2 G7. ‘Pompon de Bourgogne’ G8. Rosa centifolia Faisalabad G9. Rosa centifolia Sargodha G10. ‘Paul Ricault’
Plate 4.7: Primer (RW55C6) with the 2 DNA samples of Rosa centifolia of Pakistan and eight from USA
10 9 8 7 6 5 4 3 2 1
RW55C6

58
Figure 4.12: Dendrogram of microsatellite diversity among 8 genotypes of Rosa centifolia from USA and 2 from Pakistan

59
Table 4.4: Grouping of 10 genotypes of Rosa centifolia on the bases of microsatellite
markers
Groups Number of genotypes Name of genotypes A 3 ‘Fantin-Latour’, ‘Gros Choux d’Hollande’, ‘De
Meaux’B 3 ‘Centifolia Varigata’, ‘Cabbage Rose’1,
‘Pompon de Bourgogne’C 3 ‘Cabbage Rose’2, R. centifolia Faisalabad, R.
centifolia SargodhaD 1 'Paul Ricault’
4.2.3 Genetic analysis of Rosa centifolia and Rosa damascena of Pakistan, USA and Iran
After comparing different genotypes of R. centifolia, four genotypes of R. damascena were
also added in this study to investigate their genetic relationship with genotypes of R.
centifolia. Two of these genotypes of R. damascena were selected from Iran, previously
collected by Kiani et al. (2010) which showed genetic diversity among other. Two more
genotypes of R. damascena were selected from Pakistan, previously collected by Farooq et
al. (2013) and added with ten R. centifolia for comparison. The Same thirteen microsatellite
markers were used to investigate diversity among all fourteen genotypes (10 R. centifolia and
4 R. damascena) and experiment was done in Department of Horticultural Sciences, Texas
A&M University, USA.
According to the results presented in the figure 4.8, all thirteen markers produced 190
scorable bands from 14 different genotypes. Out of these 190 bands 15 were polymorphic.
The genotypes were divided into 5 groups with a similarity coefficient of 0.81. Group-A was
further divided into two subgroups with a similarity coefficient of 0.87. One was consisted of
genotype ‘Fantin-Latour’, ‘Gros Choux d’Hollande’ and ‘De Meaux’ and the other on R.
damascena Faisalabad. Group-B was divided to two subgroups. One of them contained
genotype ‘Centifolia Varigata’, while the other contained genotypes of ‘Cabbage Rose’1,
‘Pompon de Bourgogne’, R. damascena East Azerbaijan. R. damascena Pattoki and R.
damascena Isfahan were combined in group-C. Group-D had genotypes ‘Cabbage Rose’ 2,
R. centifolia Faisalabad and R. centifolia Sargodha. Genotype ‘Paul Ricault’ was alone in the
group E. Two genotypes of R. centifolia from Pakistan were closer to R. damascena Isfahan,
R. damascena Pattoki and R. damascena East Azerbaijan of group-C and group-B,
respectively than R. damascena Faisalabad in group-A.

60
G1. ‘Fantin-Latour’ G2. ‘Gros Choux d’Hollande’ G3. ‘Centifolia Varigata’ G4. ‘De Meaux’ G5. ‘Cabbage Rose’ 1 G6. ‘Cabbage Rose’ 2 G7. ‘Pompon de Bourgogne. G8. Rosa Centifolia Faisalabad G9. Rosa Centifolia Sargodha G10. ‘Paul Ricault’ G11. Rosa damascena Faisalabad G12. Rosa damascena Pattoki G13. Rosa damascena Isfahan G14. Rosa damascena East Azerbaijan
Plate 4.8: Primer (RW3k19) with the 8 DNA samples of different centifolia from USA and 2 from Pakistan along with 4 genotypes of Rosa damascena, 2 from Pakistan and 2 from Iran
14 13 12 11 10 9 8 7 6 5 4 3 2 1
RW3K19

61
G1. ‘Fantin-Latour’ G2. ‘Gros Choux d’Hollande’ G3. ‘Centifolia Varigata’ G4. ‘De Meaux’ G5. ‘Cabbage Rose’ 1 G6. ‘Cabbage Rose’ 2 G7. ‘Pompon de Bourgogne. G8. Rosa Centifolia Faisalabad G9. Rosa Centifolia Sargodha G10. ‘Paul Ricault’ G11. Rosa damascena Faisalabad G12. Rosa damascena Pattoki G13. Rosa damascena Isfahan G14. Rosa damascena East Azerbaijan
Plate 4.9: Primer (RW3k19) with the 8 DNA samples of different centifolia from USA and 2 from Pakistan along with 4 genotypes of Rosa damascena, 2 from Pakistan and 2 from Iran
RW10M24
14 13 12 11 10 9 8 7 6 5 4 3 2 1

62
G1. ‘Fantin-Latour’ G2. ‘Gros Choux d’Hollande’ G3. ‘Centifolia Varigata’ G4. ‘De Meaux’ G5. ‘Cabbage Rose’ 1 G6. ‘Cabbage Rose’ 2 G7. ‘Pompon de Bourgogne. G8. Rosa Centifolia Faisalabad G9. Rosa Centifolia Sargodha G10. ‘Paul Ricault’ G11. Rosa damascena Faisalabad G12. Rosa damascena Pattoki G13. Rosa damascena Isfahan G14. Rosa damascena East Azerbaijan
Plate 4.10: Primer (RW18N19) with the 8 DNA samples of different centifolia from USA and 2 from Pakistan along with 4 genotypes of Rosa damascena, 2 from Pakistan and 2 from Iran
RW18N19
14 13 12 11 10 9 8 7 6 5 4 3 2 1

63
Figure 4.13: Dendrogram of microsatellite diversity among 8 genotypes of Rosa centifolia from USA, 2 from Pakistan along with 4 genotypes of Rosa damascena, 2 from Pakistan and 2 from Iran.
Table 4.5: Grouping of 14 genotypes of Rosa centifolia and Rosa damascena on the basis of microsatellite markers
Groups Number of genotypes Name of genotypes A 4 ‘Fantin-Latour’, ‘Gros Choux d’Hollande’,
Rose ‘De Meaux’, R. damascena Faisalabad,B 4 ‘Centifolia Varigata’, ‘Cabbage rose’1,
‘Pompon de Bourgogne’, R. damascena East Azarbaijan
C 2 R. damascena Pattoki, R. damascena IsfahanD 3 ‘Cabbage Rose’2, R. centifolia Faisalabad, R.
centifolia SargodhaE 1 ‘Paul Ricault’

64
4.3 In vitro propagation of Rosa centifolia through axillary buds
This experiment was performed to perceive the most suitable concentration of cytokinins and
auxin for shoot proliferation and formation of healthy disease-free plants of R. centifolia
present in Rosa project, University of Agriculture Faisalabad, Pakistan. As lots of work has
been done on in vitro regeneration of roses and we tried to develop an in vitro protocol for
propagation of R. centifolia.
Explants with one axillary bud were collected from new flushes of Rosa centifolia grown in
the field and cultured on MS media comprising of various components with different
concentrations of growth regulators. After 3 to 4 weeks explants started producing shoots and
data (The number of days to initiate shoot, the number of shoots, the number of leaves per
shoot and shoot length) were recorded before transferring them into the rooting media.
4.3.1 Number of days to initiate shoots
Data regarding to the number of days to initiate shoots was presented in Figure 4.14.
MS media supplemented with 0.4 µM BAP in combination with 0.2 µM KIN took minimum
number of days (10.20) to initiate of shoots and proved as the best combination for producing
shoots in short number of days. 0.2 µM and 0.6 µM BAP in the MS media also showed
satisfactory results as compared to kinetin and control. MS media without growth regulators
took maximum number of days (21.93) for shoots initiation.
4.3.2 Number of shoots
The number of shoots was counted from each explant and data was presented in Figure 4.15.
The maximum number of shoots (1.93) was produced in MS media with 0.2 µM BAP. While
0.4 µM BAP alone and in combination with 0.2 µM of kinetin produced 1.87 and 1.80 shoots
respectively. But MS media without growth regulators produced minimum number of shoots
(1.0). 0.6 µM and 0.8 µM of kinetin proved as least important for proliferation of R.
centifolia under in vitro condition. It was observed that MS media supplemented with
relatively high concentration BAP (0.4 µM and 0.6 µM) along with 0.2 µM kinetin produced
shoots and callus both at the same time.
4.3.3 Shoot length
Shoot length was measured and presented in Figure 4.16.
MS media supplemented with 0.2 µM BAP along with 0.2 µM KIN produced the longest
shoot length (5.57cm) followed by 0.4 µM and 0.6 µM BAP in combination with 0.2 µM

65
kinetin, which produced shoot length 5.44 cm and 5.13 cm, respectively. Minimum shoot
length (3.17 cm) was observed in the MS media supplemented with 0.8 µM of BAP along
with 0.2 µM Kinetin and MS media without growth regulator produced shoots with shoot
length (3.67).
4.3.4 Number of leaves:
According to the data presented in the Fig. 4.17 0.4 µM BAP produced shoots with the
maximum number of leaves (2.53) followed by 0.4 µM BAP along with 0.2 µM kinetin. But
0.4 µM kinetin produced the minimum number of leaves (1.07) while control produced
number of leaves (1.13).
4.3.5 Number of days to induce roots
Results regarding to the number of days to induce roots are presented in Figure 4.19.
Half strength MS media supplemented with IBA (0.4 µM) was the most suitable for
producing roots in minimum number of days (10.07). 0.4 µM NAA and 0.6 µM IBA were
also showed satisfactory results by producing roots in 12.9 and 13.5 days, respectively. MS
media without growth regulators produced roots in a maximum number of days 19.8 and
proved as least effective in case of producing roots of R. centifolia.
4.3.6 Number of roots
The data on number of roots was recorded and presenting in Figure 4.20.
Maximum number of roots (2.47) were produced in half strength MS media containing 0.4
µM IBA followed by 2.13 in 0.4 µM concentration of NAA. While minimum numbers of
roots (1.40) were observed in half strength MS media without any growth regulator.
4.3.7 Root length
The results of root length as affected by various concentrations of growth regulators are in
shown in Figure 4.22.
Half strength MS media supplemented with 0.4 µM IBA produced roots with maximum root
length (4.21cm) followed by 0.6 µM IBA and 0.4 µM NAA which produced 3.90 cm and
3.87 cm, respectively. Minimum root length (3.30 cm) was observed in control followed by
3.50 cm and 3.67 at 0.2 µM of NAA and 0.6 µM NAA, respectively.

66
T1 – MS T2 – MS + BAP 0.2 µM T3 – MS + BAP 0.4 µM T4 – MS + BAP 0.6 µM T5 – MS + BAP 0.8 µM T6 – MS + KIN 0.2 µM T7 – MS + KIN 0.4 µM T8 – MS + KIN 0.6 µM T9 – MS + KIN 0.8 µM T10 – MS + BAP 0.2 µM + KIN 0.2 µM T11 – MS + BAP 0.4 µM + KIN 0.2 µM T12 – MS + BAP 0.6 µM + KIN 0.2 µM T14 – MS + KIN 0.2 µM + BAP 0.2 µM T15 – MS + KIN 0.4 µM + BAP 0.2 µM T16 – MS + KIN 0.6 µM + BAP 0.2 µM T17 – MS + KIN 0.8 µM + BAP 0.2 µM
Figure 4.14: Number of days taken to produce shoots from nodal explants of Rosa centifolia under in vitro conditions.
0
5
10
15
20
25
1 17 14 8 16 7 9 15 13 5 6 12 10 4 3 2 11
Day
Treatment

67
T1 – MS
T2 – MS + BAP 0.2 µM
T3 – MS + BAP 0.4 µM T4 – MS + BAP 0.6 µM T5 – MS + BAP 0.8 µM T6 – MS + KIN 0.2 µM T7 – MS + KIN 0.4 µM T8 – MS + KIN 0.6 µM T9 – MS + KIN 0.8 µM T10 – MS + BAP 0.2 µM + KIN 0.2 µM T11 – MS + BAP 0.4 µM + KIN 0.2 µM T12 – MS + BAP 0.6 µM + KIN 0.2 µM T14 – MS + KIN 0.2 µM + BAP 0.2 µM T15 – MS + KIN 0.4 µM + BAP 0.2 µM T16 – MS + KIN 0.6 µM + BAP 0.2 µM T17 – MS + KIN 0.8 µM + BAP 0.2 µM
Figure 4.15: Number of shoots produced from nodal explants of Rosa centifolia under in vitro conditions.
0
0.5
1
1.5
2
2.5
2 3 11 10 12 13 4 6 16 17 9 15 5 14 7 8 1
Shoot
Treatment

68
T1 – MS T2 – MS + BAP 0.2 µM T3 – MS + BAP 0.4 µM T4 – MS + BAP 0.6 µM T5 – MS + BAP 0.8 µM T6 – MS + KIN 0.2 µM T7 – MS + KIN 0.4 µM T8 – MS + KIN 0.6 µM T9 – MS + KIN 0.8 µM T10 – MS + BAP 0.2 µM + KIN 0.2 µM T11 – MS + BAP 0.4 µM + KIN 0.2 µM T12 – MS + BAP 0.6 µM + KIN 0.2 µM T14 – MS + KIN 0.2 µM + BAP 0.2 µM T15 – MS + KIN 0.4 µM + BAP 0.2 µM T16 – MS + KIN 0.6 µM + BAP 0.2 µM T17 – MS + KIN 0.8 µM + BAP 0.2 µM
Figure 4.16: Length of shoots produced from nodal explants of Rosa centifolia under in vitro conditions.
0
1
2
3
4
5
6
10 11 12 3 2 4 5 6 7 1 16 15 17 14 8 9 13
cm
Treatment
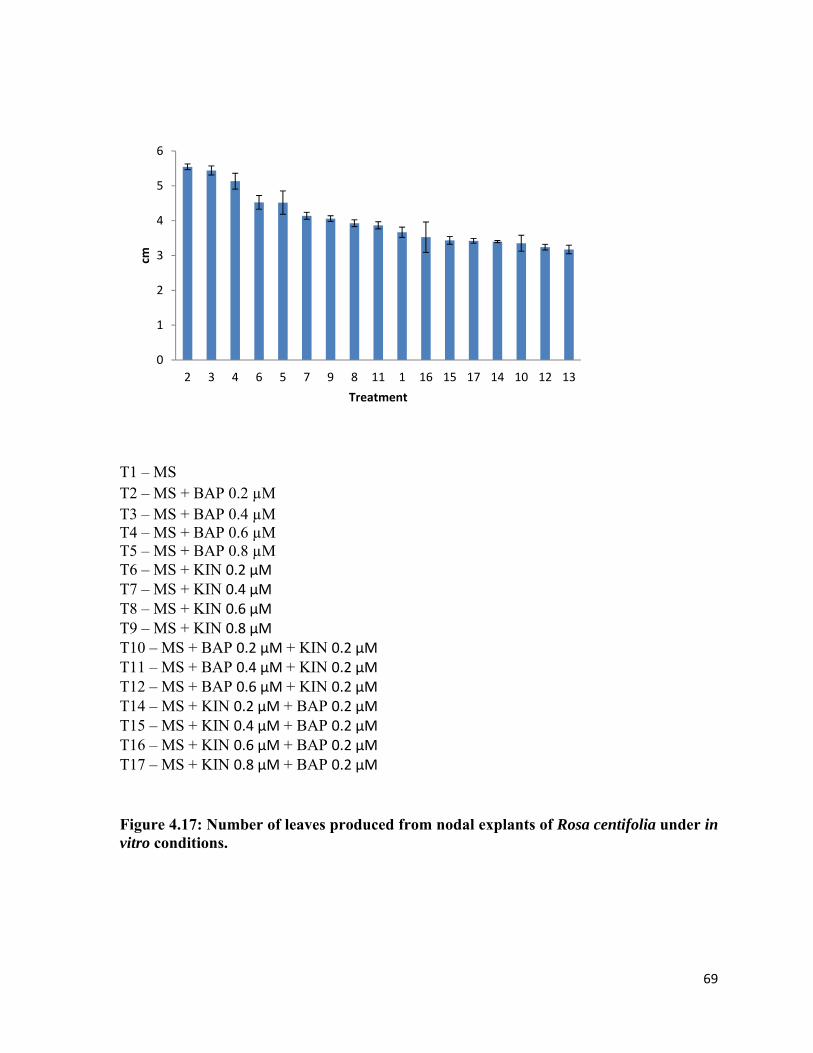
69
T1 – MS T2 – MS + BAP 0.2 µM T3 – MS + BAP 0.4 µM T4 – MS + BAP 0.6 µM T5 – MS + BAP 0.8 µM T6 – MS + KIN 0.2 µM T7 – MS + KIN 0.4 µM T8 – MS + KIN 0.6 µM T9 – MS + KIN 0.8 µM T10 – MS + BAP 0.2 µM + KIN 0.2 µM T11 – MS + BAP 0.4 µM + KIN 0.2 µM T12 – MS + BAP 0.6 µM + KIN 0.2 µM T14 – MS + KIN 0.2 µM + BAP 0.2 µM T15 – MS + KIN 0.4 µM + BAP 0.2 µM T16 – MS + KIN 0.6 µM + BAP 0.2 µM T17 – MS + KIN 0.8 µM + BAP 0.2 µM
Figure 4.17: Number of leaves produced from nodal explants of Rosa centifolia under in vitro conditions.
0
1
2
3
4
5
6
2 3 4 6 5 7 9 8 11 1 16 15 17 14 10 12 13
cm
Treatment

70
Figure 4.18: Shoot proliferation in the MS media supplemented with BAP (A= control,
B= 0.2 µM BAP, C= Callus and shoot both produced in MS media supplemented with
0.6 µM BAP along with 0.2 µM Kinetin)
A B C

71
T1 – MS T2 – MS + IBA 0.2 µM T3 – MS + IBA 0.4 µM T4 – MS + IBA 0.6 µM T5 – MS + NAA 0.2 µM T6 – MS + NAA 0.4 µM T7 – MS + NAA 0.6 µM
Figure 4.19: Number of days taken to induce roots from in vitro grown shoots of Rosa centifolia under in vitro conditions.
0
5
10
15
20
25
1 5 7 2 4 6 3
Day
Treatment

72
T1 – MS T2 – MS + IBA 0.2 µM T3 – MS + IBA 0.4 µM T4 – MS + IBA 0.6 µM T5 – MS + NAA 0.2 µM T6 – MS + NAA 0.4 µM T7 – MS + NAA 0.6 µM
Figure 4.20: Number of roots produced from in vitro grown shoots of Rosa centifolia under in vitro conditions.
0
0.5
1
1.5
2
2.5
3
3 6 7 4 5 2 1
Root
Treatment

73
T1 – MS T2 – MS + IBA 0.2 µM T3 – MS + IBA 0.4 µM T4 – MS + IBA 0.6 µM T5 – MS + NAA 0.2 µM T6 – MS + NAA 0.4 µM T7 – MS + NAA 0.6 µM
Figure 4.21: Length of roots produced from in vitro grown shoots of Rosa centifolia under in vitro conditions.
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5
3 6 4 2 7 5 1
cm
Treatment

74
Figure 4.22: Root induction in MS media supplemented with IBA
A B
Figure 4.23: Fig 3. Acclimatization of plantlets (A= Plantlet covered with polythene bags for hardening, B= The acclimatized plantlets ready to shift in greenhouse)
A B

75
Chapter-5
DISCUSSION
5.1 Phenotypic plasticity among the genotypes of Rosa centifolia
Interaction of plants with their environment and their phenotypic plasticity allows for better
adjustment and survival in a climate (Bradshaw, 1965; McNaughton et al., 1974). Diversity
in ecology supports the genetic variations and survival of a variety of plants (Nilsson, 1972).
Morphological differences can be used to differentiate closely related species and to study
variations within or among species (Nybom et al., 1996, 1997). Morphologically, Rosa
centifolia from Sargodha and Sialkot districts were identical. Similar attributes were found
with Pakpattan and Sheikhupura genotypes and with Faisalabad and Pattoki genotypes.
Maximum morphological diversity was observed between Sargodha and Sheikhupura
genotype. Riaz et al. (2007) reported a higher morphological similarity level among the
genotypes of R. webbiana and R. brunonii found in the hilly areas of Pakistan.
Soil is ultimate storage of water and effective nutrients necessary for healthy plant growth. If
soil is not properly drained, it can affect roots and nutrient availability to plants (Karlik et al.,
2003). Nutrients affect the quality and growth of rose plants (Savvas, 2002). Flower,
production also depends on adequate nutrient supply. Water and soil samples of these sites
carry differences which develop morphological variations in Rosa centifolia growing in the
different sites of Punjab. Younis et al. (2006) studied the effect of rainfall and temperature on
the yield of four Rosa species (R. damascena, R. centifolia, R. borboniana and R. ‘Gruss an
Teplitz’) and found positive correlation among rainfall and temperature. Griffing and
Langridge (1963) documented more phenotypic stability of Arabidopsis thaliana with a
range of temperature. Edwards and Richardson (2004) noticed the effect of climate on
different types of organisms found in various climates.
Different number of small and large prickles found in roses are also called thorns (Rosu et
al., 1995). The number of prickles decreases from bottom to top (Andre, 2003) and is
controlled by one dominant gene (Debener, 1999; Debener, 2003; Rajapakse et al., 2001).
The number of short thorns adversely affected leaf size, petal number and flower fragrance
but the number of long thorns does not correlate with these findings.

76
Plant leaves are also sources of variation and are used considered to distinguish between the
species (Olsson et al., 2000; Mclellan, 2000) as well as vegetatively propagated plant
(Persson and Gustavsson, 2001). Leaf size positively affected the number of petals per
flower, flower diameter and fragrance, which are very influential parameters for rose plant
yield. Nawaz et al. (2011) found environmentally important anatomical variations in the
leaves of Rosa damascena plants collected from different sites. Leaf green color at time of
first flowering had supportive influence on all morphological parameters but flower diameter
had strong positive correlation with it. The number of petals per flower had strong positive
effect on the flower diameter and fragrance. Debener (2003) and Rajapakse et al. (2001)
described the effect of environment and genes on the production of number of petals per
flower. Fragrance is also positively associated by flower diameter. In roses, the number of
petals, flower diameter, and fragrance are the main product liked by farmers and used to
make different products like rose jam, oil etc. Farooq et al. (2013) noticed significant
correlation between flower diameter and the number of petals with oil content. Positive
correlation among number of petals, flower diameter and fragrance showed the linkage of
these traits with one common gene or some linked genes.
Among different varieties of R. centifolia present in the USA, ‘De Meaux’ showed maximum
morphological diversity compared to the others. ‘Fantin-Latour’ also showed morphological
plasticity while ‘Cabbage Rose’ 2 and ‘Paul Ricault’ appeared highly similar in
morphological features.
Crespel et al. (2002) observed that diversity among the density of prickles is controlled by
various QTL (Qualitative trait loci). Number of short prickles was positively correlated with
all characters but had negative association with undulation of leaf margins only. Leaf size
was positively associated with the number of petals, flower diameter, and fragrance. Leaf
green color at the time of first flowering also had positive relationship with all parameters
except the number of prickles on flower pedicel. Undulation of leaf margins and the number
of prickles on flower pedicel both had positive relationship with the number of petals and
flower diameter but adversely affected the flower fragrance for oil extraction, a significant
parameter for selection of rose plants. The number of petals, flower diameter and flower
fragrance are important characteristics of R. centifolia. Variation in morphological data
collected from Pakistan and the USA may be due to differences in genetic makeup or

77
environment. Riaz et al. (2011) found 83% similarity of R. webbiana through morphological
markers and 72% similarity of R. brunonii through genetic markers during his diversity study
of four wild Rosa species from hilly areas of Pakistan. Younis et al. (2006) documented that
R. centifolia found in Pakistan was a different strain because it showed recurrent flowering at
high temperature.
All genotypes of R. centifolia from Pakistan and the USA produced double flowers with
many petals. In roses, this character is controlled by one dominant gene (Debener, 1999,
2003; Crespel et al., 2002). Recurrent blooming as a prominent and important character was
observed in all genotypes of R. centifolia from Pakistan, while all genotypes from the USA
were nonrecurrent. De Vries and Dubois (1978) and Semeniuk (1971b) described that the
character of recurrent blooming present in many roses is controlled by recessive alleles. R.
centifolia found in Pakistan can produce fragrant flowers at high temperature (Younis et al.,
2006) but different genotypes of Rosa centifolia found in the USA are used for ornamental
purposes and cannot produce flowers at high temperature.
Pakpattan and Sheikhupura genotypes of Pakistan were morphologically closer to the
‘Centifolia Varigata’ genotypes of R. centifolia from the USA. ‘Gros Choux d’ Hollande’,
‘Cabbage Rose’ 2 and ‘Paul Ricault’ genotypes of USA were phenotypically closer to the
Pakistani genotypes.
5.2 Genetic diversity among genotypes of Rosa centifolia
5.2.1 Genetic diversity among different genotypes of Rosa centifolia from Pakistan
In family Rosaceae mostly morphological and genetic markers are used to check the diversity
and classification (Jan et al., 1999; Hancock et al., 2004; Chang et al., 2007; Evans et al.,
2007). For characterization of roses mostly morphological parameters like growth habit,
flower and leaf parameters are used (Mohapatra and Rout, 2005). These are affected by
environmental factors. On the other hand molecular markers, which are not affected by the
environment used for studying genetic diversity (Roder et al., 1998). Among these markers
microsatellite markers have been used with a large number of vegetatively propagated crops
and species (Hokanson et al., 1998; Hvarleva et al., 2004). Microsatellite markers can be
applied for studying the genetic connection between closely associated species and cultivars
(Alvarez et al., 2001) and have also been proved effective for diversity in oil bearing Rosa
species (Farooq et al., 2013; Kiani et al., 2010; Babaei et al., 2007; Rusanov et al., 2005).

78
R. centifolia is important among oil producing species of roses (Lawrence, 1991) which was
originally cultivated in Bulgaria, then distributed in Turkey (Baydar et al., 2004) and other
parts of the world. In this study 13 Microsatellite markers were used to determine the genetic
diversity among 24 genotypes R. centifolia found in 8 districts of Punjab, Pakistan. All
genotypes were divided into 4 groups by the using UPGMA analysis, which showed the
genetic variation and divided genotypes into groups and varieties in accordance with rose
history (Wylie, 1954; Maia and Venard, 1976). All the genotypes from one district were in
the same groups which showed high level of homology within the district. As oil bearing
roses are mostly propagated by vegetative means (Robert and Schum, 2003) and in all these
districts of Punjab cutting is used as a common method of propagation, which is an evident
that plants in a district are distributed from a common source. The genetic difference among
the collection sites indicates variability among the R. centifolia clones used commercially in
Pakistan. The genotype of Faisalabad commonly used for the production of rose oil, water,
jam is genetically closer to genotypes of Sheikhupura, Lahore, Sialkot and Sargodha districts.
These genetically closer genotypes of R. centifolia were sampled from five closely located
districts in the province of Punjab, Pakistan. Samiei et al. (2010) also reported that
accessions within a site were closely related than those from different sites by using 10 SSR
markers. Genotypes closely related indicated that they may have common parents. It is also
expected that these to be seedling from the same source plant or sport of one another. Kiani
et al. (2008) experienced higher genetic diversity in plant from different provinces than same
province. Farooq et al. (2013) observed the genetic diversity among R. damascena and other
garden roses from Pakistan and compared them with genotypes from Iran and USA by using
10 simple sequence repeat (SSR) markers. Kiani et al. (2010) documented a high level of
genetic diversification among various accessions of R. damascena collected from different
areas in Iran using 37 microsatellite markers.
5.2.2 Genetic diversity among different genotypes of Rosa centifolia from Pakistan and
USA
Different varieties of R. centifolia are also found in USA. Eight of these varieties of R.
centifolia were selected and compared with two genotypes of R. centifolia from Pakistan to
investigate diversity among them. The same 13 microsatellite markers were used and the
same UPGMA analysis was used, which divided all 10 genotypes into four groups. Two

79
Pakistani genotypes were in the same group along with ‘Cabbage Rose’2. This shows the
maximum genetic similarity of Pakistani centifolia with ‘Cabbage Rose’2. This also
suggested that they have originated and distributed from some common sources. These were
also genetically closer to the ‘Pompon de Bourgogne’ of next group. ‘Fantin-Latour’, ‘Gros
Choux d’Hollande’ and ‘De Meaux’ were at maximum distance from Pakistani genotypes
which proved genetic distances among them and different sources of origin. Scariot et al.
(2006) found genetic similarities and same allelic profile between R. centifolia Rubra and
‘Crested Moss’ during studying the relation in wild species and old garden roses with
microsatellite markers. Some scientists also worked on R. centifolia Rubra and ‘Crested
Moss’ and documented ‘Crested Moss’ as a seedling of R. centifolia (Beales et al., 1998).
Cairns (2000) also considered this ‘Crested Moss’ is as variety of R. centifolia.
5.2.3 Genetic diversity among different genotypes of Rosa centifolia and Rosa
damascena from Pakistan, USA and Iran
Rosa damascena is very valuable species of rose used for oil extraction in India and Pakistan
along with R. centifolia and R. bourboniana (Kaul, et al., 2009; Younis, 2006) and as garden
plant from many years (Rusanov et al., 2005). Rose oil is very effective for its antioxidant,
antibacterial and antimicrobial activities (Achuthan et al., 2003; Ozkan et al., 2004). In this
experiment two genotypes of R. damascena from Pakistan and two from Iran were compared
with different R. centifolia genotypes from Pakistan and USA to find genetic relationship
among them. This study was extension of previous works of Kiani et al. (2011) and Farooq et
al. (2013) who compared the genotypes of R. damascena from Iran and Pakistan with one
sample of R. centifolia. Same 13 previously used Microsatellite markers and UPGMA
analysis were used for this experiment. R. damascena Isfahan and R. damascena Pattoki were
in same group which shows the genetic similarity between them. This conformed the
previous finding of Farooq et al. (2013). R. damascena Isfahan was also genetically closer to
R. damascena East Azerbaijan according to the previous reports Kiani et al. (2010) and
Farooq et al. (2013). These three R. damascena genotypes were also reflected genetic
similarity with genotypes of ‘Pompon de Bourgogne’, ‘Cabbage Rose’1 and ‘Centifolia
Varigata’ of the same group and with ‘Cabbage Rose’2 of the next group. R. damascena
Faisalabad was in separate group from other R. damascena genotypes and had genetically
more relation with ‘De Meaux’ and ‘Gros Choux d’Hollande’ which showed relatively

80
genetic relatedness among them. The genetic distance between R. damascena Faisalabad and
other damask genotypes also confirms the results of Farooq et al. (2013). These results
showed the existence of genetic diversity among genotypes of R. damascena (Babaei et al.,
2007; Mohapatre and Rout, 2005; Pirseyedi et al., 2005; Kiani et al., 2010; Kiani et al.,
2008) and R. centifolia.
5.3 In vitro propagation of Rosa centifolia through axillary buds
5.3.1 In vitro shoot formation of Rosa centifolia
In this experiment nodal explants were collected from Rosa centifolia plant grown in the field
of Rosa Project, Institute of Horticultural Sciences, University of Agriculture, Faisalabad,
Pakistan and brought in the plant tissue culture lab of this institute to culture the explants on
MS media supplemented with various concentrations of different growth regulators. Explants
were first inoculated on MS media containing different concentrations of cytokinins (BAP,
kinetin) alone and in combination with a fix amount of other to produce shoots. Different
parameters studied for shooting were number of days to shoots, the number of shoots, shoot
length and the number of leaves.
MS media supplemented with 0.4 µM of BAP along with 0.2 µM of kinetin produced shoots
in minimum number of days (10.2). The higher concentration of BAP in combination with
lower concentration of Kinetin decreases the number of days to produce shoots. These results
are similar to the findings of Hameed et al. (2006) who observed in R. indica L. that higher
concentration of BAP along with constant lower concentration of kinetin was effective to
produce shoots in decreased number of days. As this plant growth regulator (BAP) is
effective in bud break and help in the opening of buds to form shoots (Kapchina et al., 2000).
Senapati and Rout (2008) also found BAP an effective growth regulator in MS medium for
getting bud break in early days. Maximum shoot proliferation (1.93) was deemed in 0.2 µM
of BAP which was previously suggested very effective for shoot proliferation for most
species and cultivars of roses by Roberts and Schum (2003). BAP is substantial type of
cytokinin causing shoot proliferation (Noodezh et al., 2012; Xing et al., 2010; Nikbakht et
al., 2005; Pratpakumar et al., 2001) increases axillary shoot buds and decreases shoot
elongation (Hameed et al., 2006) because all the nutrients were utilized for lateral growth of
shoot (Yakimova et al., 2000). The number of shoots were decreased with the increase of
BAP concentration alone or with kinetin. This showed that lower concentration of BAP in

81
MS medium is preeminent for shoot proliferation. These results were in accordance with the
findings of Kim et al. (2003) who observed positive effect of lower concentration of BAP on
shoot proliferation of six rose cultivars and adverse effect of higher concentration. Baig et al.
(2011) concluded after culturing Rosa gruss an teplitz and R. centifolia on MS media with
different growth regulators that lower concentration of BAP was effective for shoot
formation and higher concentration could decrease the number of shoot. Hegde et al. (2011)
propagated field grown plants of rose by using shoot tip and nodes as explant and suggested
lower concentration of BAP in the MS media because higher concentration can cause
undesirable shoot multiplication. Detrimental effect of higher concentration of BAP was also
noticed on shoot proliferation by Hameed et al. (2006) and Shabbir et al. (2009). Vijaya et al.
(1991) observed BAP as very influential for producing maximum number of roots in rose.
BAP is effective growth regulator for shoot proliferation as it was observed to cause bud
break and help in shoot proliferation of Rosa hybrida (Hasegawa, 1980; Wulster and Sacalis,
1980).
MS media supplemented with high concentration (0.4 µM and 0.6 µM) of BAP in
combination with 0.2 µM kinetin produced callus and shoots in this research. Iqbal et al.
(2003) and Baig et al. (2011) also notice callus on in vitro shoot under higher cytokinin
concentrations. Akram and Aftab (2012) observed callus formation from nodal explants of
Morus macroura with increasing level of BA and IBA. Vantu (2011) observed callogenesis
in MS media supplemented with excess of kinetin and BAP. This observation was different
from findings of Nak-Udom et al. (2009) who produced shoots in R. hybrida L. cv.
‘Perfume Delight’ by using BA and NAA as growth regulator without callus formation.
Many scientists have also produced callus by using different explants in roses like leaf (Rout
et al., 1991; Dohm et al., 2001; Kim et a., 2003c; Kim et al., 2004a), stem (Kintzios et al.,
1999), root (Marchant et al., 1996) and seed (Kunitake et al., 1993; Kim et al., 2003b). But
they did not find both callus and shoot on the same culture and media. This different finding
may be due to different species of rose or different growth regulator concentration.
Maximum shoot length (5.55cm) was delineated at 0.2 µM BAP in combination with 0.2 µM
kinetin followed by 5.44cm and 5.13cm. BAP and kinetin in same concentration produced
maximum shoot length. Allahverdi et al., (2010) also observed effective role of same
concentration of BAP and kinetin on shoot length. Hameed et al., (2006) also documented

82
better results of shooting in MS media supplemented with BAP and KIN. Shoot length was
decreased with increased concentration of BAP. Baig et al. (2011) and Carelli and
Echeverrigary (2002) also documented decrease in shoot length with increase of BAP
concentration in roses. As increased concentration of cytokinins can produce thicker, shorter
and distorted shoot (Hussey, 1976).
5.3.2 In vitro root induction of Rosa centifolia
Shoot were transferred into half strength MS media fortified with various concentrations of
auxin (IBA and NAA) for better root induction, which was considered necessary for success
of plantlets in the field (Pati et al., 2005). Root formation is very essential step for
development of protocols for in vitro propagation and genetic engineering as it had been
observed arduous especially for woody plant species (Bais et al., 2001).
Roots were produced in minimum number of days (10.07) in half strength MS media
supplemented with 0.4 µM IBA followed by 0.4 µM NAA in 12.93 number of days.
Torkashvand and Shadparvar (2012) also found delay in rooting of Hibiscus rosa- sinensis
with the increase in IBA (Indole butyric acid) concentration. IBA was an effective auxin at
concentration of 0.4 µM to induce roots in R. centifolia. It is considered as the most preferred
growth regulator for root induction from cuttings of variety of plant species (Pati et al.,
2004). It enhanced the root initiation because of its more stability, slow transport and
changing into IAA characteristics (Epstein and Lavee, 1984).
0.4 µM IBA induced the maximum number of roots (4.22) and NAA produced 3.9 roots at
the same concentration. The number of roots decreased with decrease or increase of IBA
concentration. IBA level increased the redistribution of effective nutrients during the process
of root induction, which caused the production of useful nucleic acid and proteins essential
for better rooting (Blazich, 1988). Iqbal et al. (2003) documented the alteration in RNA and
protein after the application of IBA.
Maximum root length (4.21 cm) was observed in half strength MS media supplemented with
0.2 µM IBA but MS media without growth regulators produced root with least root length
(3.3). IBA had tested as the best auxin in solid half strength MS media for in vitro rooting of
R. centifolia. Similar results were observed on increase in number of root and root length
with IBA in very important specie of rose, R. damascena Pati et al. (2004). Kumar et al.
(2010) obtained better rooting of Jatropha strain DARL-2 in medium with IBA than without

83
growth regulator treatment. Auxins are very influential for root formation and their specific
quantity is effective for better root formation (Sulusoglu and Cavusoglu, 2010). As Auxin is
one of important factors for root formation from the shoots (Lambardi and Rugini, 2003) and
used for this purpose from long period of time (Weaver, 1972). Auxin causes root initiation,
which converted into root meristem (De Klerk, 2002) and root primordia formation is also
reported from auxin application (Carboni et al., 1997), indirectly causing the cell
enlargement by effecting different factor like properties of cell wall, turgor and osmotic
pressure and water permeability (Taiz and Zeiger, 2006). But at higher concentrations it can
inhibit the cell expansion (Barbier-Brygoo et al., 1991; Evans et al., 1994) because it helps to
instigate the enzymes to produce ethylene, which inhibits the cell expansion, cell division
(Apelbaum and Burg, 1972; Burg, 1973) and transport of auxin (Goldsmith, 1977; Suttle,
1988). Among different types of auxin, IBA is the most frequently and commonly used to
enhance root formation because it is less toxic and more stable (Weisman et al., 1988;
Hartmann et al., 2007). Zimmerman and Wilcoxon, (1935) discovered IBA along with
various other synthetic auxins. It is useful for better root induction in different types of plants
and used for this purpose for a long period of time (Hartmann et al., 1990). Priyakumari and
Sheela (2005) found the earliest root induction and maximum root length in MS media
supplemented with IBA during micropropagation of Gladiolus ‘Peach Blossom’. Kesari et al.
(2010) observed the influence of different levels of auxin on root initiation, number of roots
and root length of Pongamia pinnata. Ebrahim and Ibrahim (2000) reckoned better shoot
formation and root development in the solid MS media (agar as solidifying agent) than liquid
media.
It was also observed that healthy shoot with more number of leave produced better root. As
Hartmann et al. (2007) also noticed the production of some substances from the leaves,
which helps root elongation after moving down the stem.

84
SUMMARY
Rose is famous floricultural crop in the world with 200 species and 18,000 cultivars. Many of
its species are used to extract oil and to make valuable products. In Pakistan R. centifolia
found in University of Agriculture, Faisalabad is famous because it produces year-round
recurrent flowering including during the hot season of the country. This study was planned to
investigate the morphological and genetic diversity among the genotypes of Rosa centifolia
found in eight districts of the Punjab, Pakistan. Eight varieties of R. centifolia were also
selected from USA and compared with the two of its famous genotypes from Pakistan. The
protocol of in vitro propagation of this species was developed by using the explants of its
most common and famous Faisalabad genotype from Pakistan.
For morphological analysis eight genotypes of R. centifolia were selected from eight districts
of Punjab, Pakistan. Data of 24 parameters was recorded and analyzed by principle
component analysis (PCA) which showed morphological parameters like number of short
prickles, the number of long prickles, leaf size, leaf green color at the time of first flowering,
undulation of leaf margin, the number of prickles on flower pedicels, the number of prickles,
number of petals per flower, flower diameter and flower fragrance as important phenotypic
traits for studying morphological diversity of R. centifolia.
A dendrogram of eight Pakistani genotypes of R. centifolia was constructed by PCA, which
divided all genotypes into three groups. Genotypes Sheikhupura and Pakpattan were in first
group A, Kahror pakka, Faisalabad, Pattoki and Lahore in largest group B while Sialkot and
Sargodha were in the group C. Correlation was also determined among nine important
morphological traits. Number of small prickles was negatively correlated with longer prickle
number, leaf size, the number of prickles on flower pedicels, the number of flowers, the
number of petals flower-1, flower fragrance and positively correlated with all other
parameters. While number of long prickles was positively correlated with all the characters
except flower pedicel number of prickles. Leaf size also showed negative association with
the number of prickles on flower pedicels, number of flowers, the number of petals per
flower but positive with rest. Leaf green color and undulation of leaf margin showed strong
positive association with all parameters. Negative association was observed between the
numbers of prickles on flower pedicels and all other parameters except flower diameter.
Numbers of flowers plant-1, petal numbers flower-1 and flower diameter showed positive

85
correlation with rest of parameters. Morphological diversity was also measured among eight
varieties (‘Fantin-Latour’, ‘Gros Choux d’ Hollande’, ‘Centifolia Varigata’, ‘De Meaux’,
‘Cabbage rose’1, ‘Cabbage Rose’2, ‘Pompon de Bourgogne’, ‘Paul Ricault’) of R. centifolia
found in USA. Dendrogram showed maximum morphological diversity among genotypes of
‘Fantin-Latour’ and ‘De Meaux’ while maximum similarity between ‘Paul Ricault’ and
‘Cabbage Rose’2. During studying the correlation nine selected characters negative
correlation was observed between the number of small prickles and undulation of leaf margin
while leaf size, the number of petals and flower diameter showed positive association with all
selected parameters.
Information about genetic diversity is very useful in molecular genetics and cluster analysis
further described the genetic linkage among the genotype. Maximum genetically diversity
was observed among the genotypes of R. centifolia from in districts of Kahror pakka and
Pattoki. Genotypes from one district had closer genetic relationship but they showed
diversity with genotypes of other districts. Two Pakistani genotypes (Faisalabad and
Sargodha) were compared with 8 genotypes of R. centifolia found in USA to investigate
genetic relationship among them. Dendrogram combined both Pakistani genotypes and
‘Cabbage Rose’2 in same group and proved genetic closeness of Pakistani genotypes with
this USA genotype of R. centifolia which confirmed the common parentage of these
genotypes. All these R. centifolia genotypes were also genetically compared with 4
genotypes of R. damascena (2 from Pakistan and 2 two from Iran). R. damascena Isfahan
(Iran) and R. damascena Pattoki (Pakistan) were in same group and showed genetic
similarity between them. Three genotypes of R. damascena (2 from Iran and 1 from Pakistan)
showed genetic similarity with genotypes of ‘Pompon de Bourgogne’, ‘Cabbage Rose’1 and
‘Centifolia Varigata’ of same group and with ‘Cabbage Rose’2. Pakistani genotypes of R.
centifolia were genetically closer to genotype of R. damascena from Isfahan (Iran) and
Pattoki (Pakistan).
BAP, kinetin and IBA were proved as effective plant growth regulators for in vitro
propagation of R. centifolia from nodal segments. As maximum number of shoots (1.93)
were produced by 0.2 µM BAP and maximum shoots length (5.55cm) at 0.2 µM of BAP
along with 0.2 µM kinetin. But the minimum numbers of days was to induce shoots were
observed in treatment 0.4 µM BAP in combination with 0.2 µM kinetin. For root induction

86
IBA at 0.4 µM was observed as best treatment among various levels of IBA and NAA. As it
produced roots with the minimum number of days (10.07), the maximum number of roots
(2.47) and the maximum root length (4.21). MS media supplemented with comparatively
high concentration of BAP (0.4 µM and 0.6 µM) along with 0.2 µM kinetin also produced
callus and shoots from nodal explant R. centifolia.
Conclusions
Present study has unveiled the morphological and genetic diversity among genotypes of R.
centifolia from Pakistan and USA. During morphological study of R. centifolia genotypes
from Pakistan the maximum diversity was observed between Sargodha and Sheikhupura
genotypes. During studying the genotype from USA, ‘Fantin-Latour’ and ‘De Meaux’ were
noticed morphologically maximum similar among 8 genotypes. Genetic analysis by using
SSR markers has documented the genetic similarity among the genotypes from one district
and diversity among genotypes of different districts. Genotypes of R. centifolia from Pakistan
showed the highest genetic closeness with ‘Cabbage Rose’2 genotype R. centifolia from
USA. Pakistani genotypes of R. centifolia also showed genetic relationship with Isfahan and
Pattoki genotypes of R. damascena from Iran and Pakistan, respectively. This study showed
the morphological and genetic diversity among genotypes of R/ centifolia which will be
helpful for breeding programmes. For in vitro shoot formation 0.2 µM BAP produced the
maximum number of shoots and 0.2 µM BAP in combination with 0.2 µM KIN produced
maximum shoots length. But maximum number of leaves were produced by 0.4 µM BAP.
0.4 µM IBA was the best concentration to produce roots with the minimum number of days,
the maximum number of roots and the maximum root length. In vitro propagation is proved
an effective tool for year-round production of healthy plant of R. centifolia.

87
LITERATURE CITED
Achuthan, C.R., B.H. Babu and J. Padikkala. 2003. Antioxidant and hepatoprotective effects
of Rosa damascene. Pharmaceutical Biology oil. Fitoterapia, 74: 394-396.
Adugna, W., M.T. Labuschagne and C.D. Viljoen. 2006. The use of morphological and
AFLP markers in diversity analysis of linseed. Biodivers. Conserv. 15: 3193-3205.
Agaoglu, Y., A. Ergul and N. Baydar. 2000. Molecular analyses of genetic diversity of oil
rose (Rosa damascena Mill.) grown in Isparta (Turkey) region. Biotechnol. Biotechnol.
Equip. 14:16-18.
Akram, M. and F. Aftab. 2012 Efficient micropropagation and rooting of king white
Mulberry (morus macroura miq.) Var. Laevigata from nodal explants of mature tree.
Pak. J. Bot., 44: 285-289.
Allahverdi Mamaghani, B., M. Ghorbanli, M.H. Assareh and A. Ghamari Zare. 2010. In vitro
propagation of three Damask Roses accessions. Iran. J. Plant Physiol. 1 (2): 85-94.
Allardice, P. 1994. The art of Aromatherapy. Random House Value publishing, New York,
pp. 65-67.
Almekinders, C.J.M., N.P. Louwaars and G.H. De Bruijn. 1994. Local seed systems and their
importance for an improved seed supply in developing countries. Euphytica. 78: 207-216.
Alvarez, A.E., C.C.M. van de Wiel, M.J.M. Smulders and B. Vosman. 2001. Use of
microsatellites to evaluate genetic diversity and species relationships in the genus
Lycopersicon. Theor Appl Genet 103: 1283-1292.
Anaç, O. 1984. Gas chromatographic analysis on Turkish rose oil. Perfum. Flavor 9: 1-14.
Andre, J.P. 2003. Morphology and anatomy: Shoots and stems. A.V. Roberts, T. Debener,
and S. Gudin (Eds.). Encyclopedia of rose science. no. 1. Elsevier, Oxford. pp. 491-497.
Anonymous, 2011. Roses evolved much before man. Times City, Times News Net work,
Sunday Times of India, Lucknow, January23, PageNo.2.
Apelbaum, A. and S.P. Burg. 1972. Effect of ethylene on cell division and t DNA synthesis
in Pisium sativumn. Plant Phvsiol. 50: 117-125.
Ara, K.A., M.M. Hossain, M.A. Quasim, M. Ali, and J.U. Ahmed. 1997. Micropropagation
of rose (Rosa sp. cv). Plant Tissue Culture 7: 135-42.

88
Arene, L., C. Pellegrino and S. Gudin. 1993. A comparison of the somaclonal variation level
of Rosa hybrida L. cv. ‘Meirutral’ plants regenerated from callus or direct induction from
different vegetative and embryonic tissues. Euphytica. 71: 83-90.
Asadi, A.A., C. Vedadi, M. Rahimi and B. Naserian. 2009. Effect of plant growth hormones
on root and shoot regeneration in Rose (Morrasia) under in-vitro conditions, Bioscience
Research. 6(1): 40-45.
Azeem, S., A.I. Khan, F.S. Awan, A. Riaz and S. Bahadur. 2012 Genetic diversity of rose
germplasm in Pakistan characterized by random amplified polymorphic DNA (RAPD)
markers. Afri. J. Biotech. 11: 10650-10654
Babaei, A., S.R. Tabaei-Aghdaei, M. Khosh-Khui, R. Omidbaigi, M.R. Naghavi, G.D.
Esselink and M.J.M. Smulders. 2007. Microsatellite analysis of Damask rose (Rosa
damascena Mill.) accessions from various regions in Iran reveals multiple genotypes.
BMC Plant Biol. 10: 7-12.
Baig, M.M.Q., A.H. Ishfaq, H. Azhar, A. Touqeer and A.A Nadeem. 2011. An efficient
protocol for in vitro propagation of Rosa gruss an teplitz and Rosa centifolia. Afri. J.
Biotech. 10(22): 4564-4573.
Bais, H.P., G. Sudha and G.A. Ravishankar. 2001. Influence of putrescence, silver nitrate and
polyamine on morphogenetic response in untransformed and transformed tissue of
Cichorium intybus and their regenerates. Plant Cell Report. 20: 547-555
Barbier-Brygoo, H., G. Ephritikhine, D. Klambt, C. Maurel, K. Palme, J. Schell and J. Guern.
1991. Perception of the auxinsignal at the plasma membrane of tobacco mesophyll
protoplasts. Plant J. 1: 83-93.
Barve, D.M., R.S. Iyenr, S. Kendurkar and A.F. Mascarenhas. 1984. An efficient method for
rapid propagation of some budded rose varieties. Indian J. Hortic. 41: 1-7.
Basaki,T., M. Mardi, K.M. Jafarkhani, S.M. Pirseyedi, M.R. Ghaffari, A. Haghnazari, S.P.
Salehi, and P. Koobaz. 2009. Assessing Rosa persica genetic diversity using amplified
fragment length polymorphisms analysis. Scientia Horticulturae, 120: 538-543.
Baser, K.H.C. 1992. Turkish rose oil. Perfum. Flavor 17: 45-52.
Basim., E. and H. Basim.2003. Antibacterial activity of Rosa damascene essential oil.
Elsevier Science, Fitoterapia 74: 394-396.

89
Baydar, G.N., H. Baydar and T. Debener. 2004. Analysis of genetic relationships among
Rosa damascena plants grown in Turkey by using AFLP and microsatellite markers. J
Biotech. 111: 263-267.
Baydar, H. and N.G. Baydar. 2005. The effects of harvest date, fermentation duration and
Tween 20 treatment on essential oil content and composition of industrial oil rose (Rosa
damascena Mill.). Ind. Crop. Prod. 21:251-255.
Bayrak, A. and A. Akgül. 1994. Volatile oil composition of Turkish rose (Rosa damascena).
J. Sci. Food Agric. 64: 441-448.
Beales, P., T. Cairns, W. Duncan, G. Fagan, W. Grant, K. Grapes, P. Harkness, K. Hughes, J.
Mattock, D. Ruston, P. Sutherland and T. Williams. 1998. Botanica's roses. The
encyclopedia of roses. Random House, Australia.
Beismann, H. 1997. AFLP analysis sheds light on distribution of two Salix species and their
hybrid along a natural gradient, Mol. Ecol. 6: 989-993
Berkowsky, P. and J. Bruce. 2000. Importance of rose oil in asthma remedy. Academic
Press, NewYork.
Blazich, F.A. 1988. Mineral nutrition and adventitious rooting. In: Adventitious root
formation in cuttings. (Eds.): T.D. Davis, B.E. Haissig and N. Sankhla. Dioscorides
Press. Portland, Oregon: p. 150-152.
Blom, T.J. and M.J. Tsujita. 2003. Cut rose production. In: Roberts, A.V., T. Debener and S.
Gudin. Encyclopedia of Rose Science, Volume 2. Elsevier Academic Press. pp 595-600.
Boonprakob, U. 1996. Texas A&M University. RAPD polymorphisms in diploid plums:
Genetic relationships and genetic linkage maps. Doctoral Thesis, Texas A&M University.
Bown, D. 2002. The royal horticultural society. New encyclopedia of herbs and their uses,
Dorling Kindersley Limited, Penguin Company, London, 346-347
Bradshaw, A.D. 1965. Evolutionary significance of phenotypic plasticity in plants. Advances
in Genetics. 13: 115-155.
Bressan, R.H., Y.J. Kim, S.E. Hyndman, P.M. Hasegawa and R.A. Bressan. 1982. Factors
affecting in vitro propagation of rose. J. Amer. Soc. Hort. Sci., 107: 979-990.
Brichet H., 2003. Distribution and ecology. In: Roberts A.V., Debener T., Gudin S. (eds),
Encyclopedia of Rose Science. London, Elsevier Academic Press: 199-227.

90
Buckle, J. 2003. Clinical aromatherapy: Essential oils in practice, 2nd Ed. Edinburgh:
Churchill Livingtone.
Burg, S.P. 1973. Ethylene in plant growth. Proc Natl Acad Sci USA 70: 591-597
Cairns, T. 2000. Modern Roses XI, The World Encyclopedia of Roses. Academic Press
Camacho Villa, T.C., N. Maxted, M. Scholten and B. Ford-Lloyd. 2005. Defining and
identifying crop landraces. Plant Genetic Resources. 3: 373-384.
Capelladesl, M., R. Lemeur and P. Debergh. 1991. Effects of sucrose on starch accumulation
and rate of photosynthesis in Rosa cultured in vitro. Plant Cell Tiss. Org. Cult. 25: 21-26.
Carboni, E., M.G. Tonelli, P. Lauri, P. Lacovacci, C. Kervers, C. Damiano and T. Gaspar.
1997. Biochemical aspects of almond micro-cuttings related to in vitro rooting and their
requirements. Plant Physiol. Biochem. 28: 631-638.
Carelli, B.P. and S. Echeverrigaray. 2002. An improved system for the in vitro propagation
of rose cultivars. Sci. Hortic. 92: 69 -74.
Castilon, J., B. Jones and K. Kamo. 2006. Efficient regeneration of rose plants from somatic
embryos of three genetically diverse cultivars. Floral Nursery Plants Research Unit, US
National Arboretum, Beltsville.
Chakrabarty, D., A.K. Mandal and S.K. Data. 2000. In vitro propagation of rose cultivars.
Indian j. plant physi. 5(2): 189-192.
Chakraborty, K. 2005. Knowing Roses. The Rose Bulletin pp 4- 5.
Chang, K.S., C.S. Chang, T.Y. Park and M.S. Roh. 2007. Reconsideration of the Prunus
serrulata complex (Rosaceae) and related taxa in eastern Asia. Bot. J. Linn. Soc., 154:
35-54.
Collet, G.F. and C.L. Le. 1987. Effect of auxin during in vitro rhizogenesis of rose and apple
trees. Acta Hort., 212:273-280.
Crespel, L., M. Chirollet, C.E. Durel, D. Zhang, J. Meynet, and S. Gudin. 2002. Mapping of
qualitative and quantitative phenotypic traits in Rosa using AFLP markers. Theor. Appl.
Genet. 105:1207-1214
Cuizhi, G. and K.R. Robertson, 2003. Rosa Linnaeus, Sp. Flora of China, 9: 339-381.
Davies, D.R. 1980. Rapid propagation of roses in vitro. Sci. Hortic. 13: 385- 9.
De klerk, G. 2002. Rooting of microcuttings: theory and practice. In vitro cell. Dev. Biol.
Plant, 38: 415-422.

91
De Vries, D.P. and A.M. Dubois. 1978. On the transmission of the yellow flower colour
form Rosa foetida to recurrent flowering hybrid tea-roses. Euphytica 27:205-210.
Debener, T. 1999. Genetic analysis of horticulturally important morphological and
physiological characters in diploid roses. Gartenbauwissenschaft 64(1):14-20.
Debener, T. 2002. Molecular markers as a tool for analyses of genetic relatedness and
selection in ornamentals. p.329-345. In: A. Vainstein (ed.) Breeding for ornamentals:
Classical and molecular approaches. Kluwer Academic Publishers, Dordrecht, The
Netherlands.
Debener, T. 2003. Genetics: Inheritance of characteristics. In A.V. Roberts, T. Debener, and
S. Gudin (Eds.). Encyclopedia of rose science. no. 1. Elsevier, Oxford. pp. 286-292.
Dhawani, V. and S.S. Bhojwani. 1986. Micropropagation in crop plants. Glimpses in plants
Res., 7: 1-75.
Dohare, S.R., M. Shafi and U.S. Kaicker. 1991. Micropropagation in rose. Acta Hortic. 289 :
107-108.
Dohm, A., C. Ludwig, K. Nehring and T. Debener. 2001. Somatic embryogenesis in roses.
Acta Hort. 547:341-7.
Douglas, G.C., C.B. Rutledge, A.D. Casey and D.H.S. Richardson. 1989. Micropropagation
of Floribunda, ground cover and miniature roses. Plant Cell Tiss. Org. 19: 55-64.
Doyle, J.J. and J.L. Doyle. 1987. A rapid DNA isolation procedure for small quantities of
fresh leaf tissue. Phytochem. Bull. 19(1):11-15.
Eagles, H.A. and J.E. Lothrop. 1994. Highland maize from central Mexico – its origin,
characteristics, and use in breeding programs. Crop Science. 34:11–19.
Ebrahim, M.K.H. and A.I. Ibrahim. 2000. Influence of medium solidification and pH value
on in vitro propagation of Maranta leuconeura cv Kerchoviana. Sci Hortic. 86: 211-21.
Edwards, M. and A.J. Richardson. 2004. Impact of climatic change on marine pelagic
phenology and trophic mismatch. Nature. 4 (30):881-884.
Elliott, R.F. 1970. Axenic culture of meristem tips of Rosa multiflora. Planta 95: 183-186.
Emadpour, M., R. Hosseini and E. Beiramizadeh. 2009. A high genetic diversity revealed
between commercial rose cultivars by RAPD-PCR technique. Inter. J. Plant Produc. 3(4):
61-66.

92
Epstein, E. and S. Lavee. 1984. Conversion of Indole-3-butyric Acid to Indole-3-acetic Acid
by Cuttings of Grapevine (Vitis vinifera) and Olive (Olea europea). Plant Cell Physiol.
25(5): 697-703
Ercisli, S. and M. Guleryuz. 2005. Rose hip utilization in Turkey. Acta Horticulturae, 490:
77-83.
Ertter, B. 2001. Native California Roses. An on-line Monograph published through the
University of California, available online at http://
ucjeps.Berkeley.edu/ina/roses/roses:html.
Esselink, G.D., M.J.M. Smulders and B. Vosman. 2003. Identification of cut-rose (Rosa
hybrida) and rootstock varieties using robust Sequence Tagged Microsatellite markers.
Theor. Appl. Genet. 106: 277-286.
Evans, A. 2009. Rose imports. Floraculture International . Feb. 2009. pp. 42-43.
Evans, K.J., D.E. Symon, M.A. Whalen, J.R. Hosking, R.M. Barker and J.A. Oliver. 2007.
Systematics of the Rubus fruticosus aggregate (Rosaceae) and other exotic Rubus taxa in
Australia. Aust. Syst. Bot., 20: 187-251.
Evans, M.L., H. Ishikawa, M.A. Estelle. 1994. Responses of Arabidopsis roots to auxin
studied with high temporal resolution: Comparison of wild type and auxin-response
mutants. Planta 194: 215-222.
Farooq, A., M. Kiani, M.A. Khan, A. Riaz, A.A. Khan, N. Anderson and D.H. Byrne. 2013.
Microsatellite Analysis of Rosa damascena from Pakistan and Iran. Hort. Environ.
Biotechnol. 54(2): 141-147.
Fredrick, C., A. Waganer and N. Morvillo. 2002. Randomly amplified polymorphic DNA
(RAPD) analysis of musk rose (Rosa moschata). Pro Fla State Hort Soc. 115: 117-119.
Goldsmith, M.H.M. 1977. The polar transport of auxin. Annu. Rev. Plant Physiol. 28: 439-
478.
Gonzalez, V.M., M. Lopez, A.E. Valdes and R.J. Ordas. 2008. Micropropagation of three
berry fruit species using nodal segments from field-grown plants. Ann. Appli. Bio. 137:
73-78
Gordon, J. 1953. Pageant of the Rose. Studio Publications, New York, in association with
Thomas Y. Crowell.

93
Griffing, B., and J. Langridge. 1963. Phenotypic stability of growth in the self-fertilized
species, Arabidopsis thaliana. In Statistical Genetics in Plant Breeding. Nat. Acad. Sci.-
Nat. Res. Council Publ. 982:368-394.
Gudin, S. 2000. Rose: Genetics and breeding, p. 159-189. In: J. Janick (ed.). Plant breeding
reviews. vol. 17. Wiley, New York.
Guilford, P., S. Prakash, J.M. Zhu, E. Rikkerink, S. Gardiner and H. Bassett. 1997.
Microsatellites in Malus_domestica (apple): abundance, polymorphism and cultivar
identification. Theor Appl Genet 94: 249–254.
Gustavsson, L.A. 1998. Rosor for nordiska tradgardar. Natur och Kultur, Stockholm.
Hameed N, A. Shabbir, A. Ali and R. Bajwa. 2006. In vitro micropropagation of disease free
rose (Rosa indica L.). Mycopath. 4(2): 35-38.
Hancock, J. F., S. Serce, C.M. Portman, P. W. Callow and J. J. Luby. 2004, Taxonomic
variation among North and South American subspecies of Fragaria virginiana Miller and
Fragaria chiloensis (L.) Miller. Can. J. Bot., 82: 1632-1644.
Hartmann, H.T., D.E. Kester, J.F.T. Davies and R.L. Geneve. 2007. Plant Hormones. In:
Plant Propagation: Principle and Practices. 7th edition, Prentice-Hall, New Delhi. pp.
292-320.
Hartmann, H.T., Kester, D.E. and Davies, F.T. 1990. Plant Propagation: Principles and
Practices. pp. 246.-247. Prentice Hall, Englwood Cliffs, NJ . ISBN 0-13-681007-1.
Hasegawa P.M. 1980. Factors affecting shoot and root initiation from cultured rose shoot
tips. J. Am. Soc. Hortic. Sci. 105: 216-220.
Hasegawa, P.M. 1979. In vitro propagation of rose. HortScience, 14: 610-612.
Hegde, V., B. Kag, B.N. Sathyanarayana and R. Sharath. 2011. Rapid in vitro multiplication
and shoots regeneration from nodal explants of rose. J. Cell and Tiss Res. 11(3) 2961-
2965.
Heinrichs, F. 2008. International statistics flowers and plants, International Association of
Horticultural Producers/Union Fleurs, Voorhout, The Netherlands, 56: 16-90
Heun, M. 1997. Site of einkorn wheat domestication identified by DNA fingerprinting,
Science 278: 1312-1314.

94
Hibrand-Saint Oyant, L., L. Crespel, S. Rajapakse, L. Zhang, and F. Foucher. 2008. Genetic
linkage maps of rose constructed with new microsatellite markers and locating QTL
controlling flowering traits. Tree Genet. Genomes. 4: 11-23.
Hokanson, S.C., A.K. Szewc-McFadden, W.F. Lamboy and J.R. McFerson. 1998.
Microsatellite (SSR) markers reveal genetic identities, genetic diversity and relationships
in a Malus · domestica Borkh. core subset collection. Theor Appl Genet 97:671-683.
Horst., R.K. 1983. Black spot. In: Compendium of Rose Diseases. APS Press. American
Phytopathological Society, St. Paul, MN, pp 7-11.
Hussey, G. 1976. In vitro release of axillary shoots from apical dominance in
monocotyledonous plantlets. Ann Bot 40: 1323-1325.
Huys, G. 1996. High-resolution genotypic analysis of the genus Aeromonas by AFLP
fingerprinting, Int. J. Syst. Bacteriol. 46, 572-580
Hvarleva, T., K. Rusanov, F. Lefort, I. Tsvetkov, A. Atanassov and I. Atanassov. 2004.
Genotyping of Bulgarian Vitis vinifera L. cultivars by microsatellite analysis. Vitis 1:27-
34.
Hyndman, S.E., P.M. Hasegaw, and R.A. Bressan. 1982. The role of sucrose and nitrogen in
adventitious root formation on cultured rose shoots. Plant Cell Tissue Organ Culture 1:
229-238
Iqbal, M.J., M.M. Khan, B. Fatima, M. Asif and M. Abbas. 2003. In vitro propagation of
"Hybrid Tea" roses. Pak. J. Agri. Sci., 40: 3-4.
Ishioka, N. and S. Tanimoto. 1990. Plant regeneration from Bulgarian rose callus. Plant Cell,
Tissue and Organ Culture 22: 197-199.
Iwata, H., T. Kato and S. Ohno. 2002. Triparental origin of Damask roses. Gene (Elsevier)
259: 53-59.
Jacobs, G., P. Allan and C.H. Bomman. 1969. Tissue culture studies on rose: use of shoot tip
explants. I. Auxin: cytokinin effects. Agroplautae, 1: 179-188.
Jalali-Heravi, M., H. Parastar and H. Sereshti. 2008. Development of a method for analysis
of Iranian damask rose oil: combination of gas chromatography-mass spectrometry with
Chemometric techniques. Anal Chim Acta 2008: 623: 11-21.
Jan, C.H., 1996. Texas A&M University. Rose germplasm identification and analysis with
RAPD markers. Master.s Thesis, Texas A&M University.

95
Jan, C.H., D.H. Byrne, J. Manhart and H. Wilson. 1999. Rose germplasm analysis with
RAPD markers. ASHS Northeast Region Annual Meeting, Cambridge, Mass., ETATS-
UNIS, 34: 206-209.
Jaskani, M.J., M. Qasim, J. Sherani, Z. Hussain and H. Abbas. 2005. Effect of growth
hormones on shoot proliferation of rose cultivars. Pak. J. Bot., 37(4): 875-881.
Jitendra, J., T. Vineeta, K. Ashok, K. Brijesh and P. Singh. 2012. Rosa centifolia: Plant
Review. International Journal of Research in Pharmacy and Chemistry (IJRPC). 2(3)
Jones, C.J., K.J. Edwards, S. Castiglione, M.O. Winfield, F. Sala and C. Van deWiel. 1997.
Reproducibility testing of RAPD, AFLP and SSR markers in plants by a network of
European laboratories. Mol Breeding 3: 381-390.
Joret, C. 1892. La Rose dans l antiquite et au moyen age. Emile Bouillon, Paris.
Kanchanapoom, K., P. Sakpetha and K. Kanchanapoom. 2010. In vitro lowering of shoots
regenerated from cultured nodal explants of Rosa hybrida cv. ‘Heirloom’. Sci. Asia. 36:
161-164.
Kanchanapoon, K., N. Posayapisit and K. Kanchanapoom. 2009. In Vitro Flowering from
Cultured Nodal Explants of Rose (Rosa hybrida L.). Not. Bot. Hort. Agrobot. Cluj. 37:
261-263
Kapchina A.V., H.J. Vanntelgen and E. Yakimova. 2000. Role of phenylurea cytokinin
CPPU in apical dominance release in in vitro cultured Rosa hybrida L. J. Plant Growth
Regul. 19: 232-237.
Kapchina-Toteva, V., M. Somleva and H.J. Van Telgen. 2002. Anticytokinin effect on apical
dominance release in in vitro cultured Rosa hybrida L. Biol. Plant. 45: 183-188.
Karlik, J.F., J.O. Becker, H.B. Pemberton and U.K. Schuch. 2003. Production and
Marketing: Field Rose production. In: Roberts AV, Debener T, Gudin S (eds)
Encyclopedia of Rose Science. Elsevier Ltd.
Kaul, K., S. Karthigeyan, D. Dhyani, N. Kaur, R.K. Sharma and P.S. Ahuja. 2009.
Morphological and molecular analyses of Rosa damascena × Rosa bourboniana
interspecific hybrids. Sci. Hortic.122:258–263.
Kaur, N., R.K. Sharma, M. Sharma, V. Singh and P.S. Ahuja. 2007. Molecular evaluation
and micropropagation of field selected elites of R. damascena. General and Applied Plant
Physiology, 33: 171-186.

96
Keim, P. 1997. Molecular evolution and diversity in Bacillus anthracis as detected by
amplified fragment length polymorphism markers, J. Bacteriol. 179: 818-824
Kermani, M.J., P. Khosravi and S. Kavand. 2010. Optimizing in vitro propagation of Rosa
persica. Irani J. Gene and Plant Breeding, 1: 44-51.
Kesari, V., A. Das and L. Rangan. 2010. Effect of genotype and auxin treatments on rooting
response in stem cuttings of CPTs of Pongamia pinnata a potential biodiesel legume crop.
Current Science, 98(9): 1234-1237.
Khalifah, N.S.A., S. Hadi and F. Khan. 2005. Influence of Sucrose Concentration on in vitro
Growth of Five Rose (Rosa hybrida L.) Cultivars. Plant Tissue Cult. 15: 43-49.
Khan, I.A. and J.J. Shaw. 1988. Biotechnology in Agriculture. Punjab. Agric. Res.
Coordination Board Faisalabad, Pakistan. pp. 2.
Khan, M.A. and S.U. Rehman. 2005. Extraction and analysis of essential oil of Rosa species.
Inter. J. Agri & Bio. 7(6): 973-974
Khan, M.A., S.S. Shaukat, A. Shahzad and W. Ahmed. 2011. Application of waste
stabilization pond’s effluent on cultivation of roses (Rosa damascena mill). Pak. J. Bot.,
43(4): 1919-1923
Khosh-Khui, M. and K.C. Sink, 1982. Micropropagation of new and old world rose species.
J. hort. Sci. 57: 315-319.
Khosravi, P., J.M. Kermani, G.A. Nematzadeh and M.R. Bihamta. 2007. A protocol for mass
production of Rosa hybrida cv. Iceberg through in vitro propagation. Iranian Journal of
Biotechnology, 5: 100-104.
Kiani, M., Z. Zamani, A. Khalighi, M.R.F. Moghadam, and D.H. Byrne. 2010. Microsatellite
analysis of Iranian Damask rose (Rosa damascena Mill.) germplasm. Plant Breeding
129:551-559.
Kiani, M., Z. Zamani, A. Khalighi, R. Fatahi and D.H. Byrne. 2008. Wide genetic diversity
of Rosa damascena Mill. Germplasm in Iran as revealed by RAPD analysis. Scientia
Horticulturae 115: 386-392
Kiani, M., Zamani, Z., Khalighi, A., Fatahi, R., Byrne, D.H., 2007. Wide genetic diversity of
Rosa damascene Mill. germplasm in Iran as revealed by RAPD analysis. Scient Hort,
115: 386-392.

97
Kim, C.K., J.D. Chung, S.K. Jee and J.Y. Oh. 2003-c. Somatic embryogenesis from in vitro
grown leaf explants of Rosa hybrida L. J Plant Biotechnol., 5:161-4.
Kim, C.K., J.Y. Oh, J.D. Chung, A.M. Burrell, D.H. Byrne. 2004-a. Somatic embryogenesis
and plant regeneration from in vitro grown leaf explants of rose. HortScience,
39(6):1378-1380.
Kim, C.K., J.Y. Oh, S.O. Jee and J.D. Chung. 2003. In vitro micropropagtion of Rosa
hybrida L. J. Plant Biotechnol. 5: 115-119.
Kim, S.W., S.C. Oh, D.S. In and J.R. Liu. 2003b. High frequency somatic embryogenesis
and plant regeneration in zygotic embryo cultures of Japanese honeysuckle. Plant Cell
Tissue Organ Cult. 72: 277-280.
Kintzios, S., C. Manos and O. Makri. 1999. Somatic embryogenesis from mature leaves of
rose (Rosa sp.). Plant Cell Rep. 18: 467-472.
Knudsen, J.T., R. Eriksson, J. Gershenzon and B. Stahl. 2006. Diversity and distribution of
floral scent. Bot Rev. 72: 1-120.
Koreth, J., J.J. O’Leary and J.O. McGee. 1996. Microsatellites and PCR genome analysis. J.
Path 178: 239-248.
Kornova, K.M. and J. Michailova. 1994. Study of the in vitro rooting of Kazanlak oil-bearing
rose (Rosa damascena Mill.). - J. Essen. Oil Res. 6: 485-492.
Kovats, E. 1987. Composition of essential oils. Part 7. Bulgarian oil of rose (Rosa
damascena Mill.). J. Chromatogr. 406, 185-222.
Krussmann, G. 1981. The complete book of roses. Portland, Oregon: Timber Press
Kumar, A., A. Sood, U.T. Palni, A.K. Gupta and L.M.S. Palni. 2001. Micropropagation of
Rosa damascena Mill. from mature bushes using thidiazuron. - J. hort. Sci. Biotechnol.
76: 30-34.
Kumar, A., D. Aggarwal, P. Gupta and M.S. Reddy. 2010. Factors affecting in vitro
propagation and field establishment of Chlorophytum borivilianum. Biologia Plantarum.
54 (4): 601-606.
Kunitake, H., H. Imamizo and M. Mii. 1993. Somatic embryogenesis and plant regeneration
from immature seed-derived calli of rugosa rose (Rosa rugosa Thunb.). Plant Sci 90:187-
194.

98
Kurt A. and Yamankaradeniz R. 1983. The composition of rose hip is grown naturally in
Erzurum province and their processing possibilities to different products. Turkish J.
Agric. Forest. 7: 243-248
Lambardi, M. and E. Rugini. 2003. Micropropagation of olive (Olea europaea L.). In:
Micropropagation of woody trees and fruits. (Eds.): S.M. Jain and K. Ishii Kluwer Acad.
Publ., pp. 1-5.
Landrein, S., R. Borosova, J. Osborne, M. Shah, M.T.M. Rajput, S.S. Tahir and J. Zielinski.
2009. Rosaceae-Potentilleae and Roseae. In: Flora of Pakistan. Karachi University Press,
University of Karachi, Karachi, Pakistan. 216-312.
Lawrence, B.M. 1991. Progress in essential oil: rose oil and extracts. Perfum. Flavor. 16: 43-
77.
Lehner, E., and J. Lehner. 1960. Folklore and Symbolism of Flowers, Plants and Trees.
Tudor, New York.
Leus, L., F. Jeanneteau, J.V. Huylenbroeck, E.V. Bockstaele and J.D. Riek. 2004. Molecular
evaluation of a collection of rose species and cultivars by AFLP, ITS, rbc L, and mat K.
Acta Horticul. 651:141-147.
Li, X., S.F. Krasnyanski and S.S. Korban. 2002: Somatic embryogenesis, secondary somatic
embryogenesis and, shoot organogenesis in Rosa. J. Plant Physiol. 159, 313-319.
Lis-Balchin, M. and S.G. Deans. 1997. Bioactivity of selected plant essential oils against
Listeria monocytogenes. Journal of Applied Bacteriology, 82, 759-762.
Lu, L., C. Gu, C. Li, C. Alexander, B. Bartholomew, A.R. Brach, D.E. Boufford, H. Ikeda,
H. Ohba, K.R. Robertson. 2003. Spongberg. Rosaceae. Flora of China, 9: 46-434.
Mahmood, N., S. Piacente, C. Pizza, A. Burke, A.I. Khan and A.J. Hay. 1996. The Anti-HIV
activity and mechanisms of action of pure compounds isolated from Rosa damascene.
Biochemical and Biophysical Research Communications. 229, 73-79.
Mahmoud, I.M.A., A.M.A. Hassanein and N.E. El-keltawi. 2011. Factors Affecting in vitro
Propagation Efficiency of Hybrid Tea Rose. American-Eurasian J. Agric. & Environ.
Sci., 11 (6): 847-853
Maia, N and P. Venard. 1976. Cytotaxonomie du genre Rosa et origine des rosiers cultivés.
In: FNPH (ed) Travaux sur le rosier de serre. INRA Editions, Paris, pp 7-20

99
Mamaghani, B.A., M. Ghorbanli, M.H Assareh and A.G Zare. 2010. In vitro propagation of
three Damask Roses accessions. Iranian Journal of Plant Physiology. 1(2): 85-94.
Marchant. R., M.R. Davey, J.A. Lucas and J.B. Power. 1996. Somatic embryogenesis and
plant regeneration in Floribunda rose (Rosa hybrida L.) cvs. trumpeter and glad tidings.
Plant Sci 120: 95-105.
Martin, C. 1985. Plant breeding in vitro. Endeavour; 9:81-6.
Martin, M., F. Piola, J.M. Chessel and P. Heizmann. 2001. The domestification process of
the Modern Rose: genetic structure and allelic composition of the rose complex. Theor
Appl Genet 102: 398-404.
Mclellan, T. 2000. Geographic variation and plasticity of leaf shape and size in Begonia
dregei and B. homonyma (Begoniaceae). Bot. J. Linn. SOC. 132: 79-95.
Mcnaughton, S.J., T.C. Folsom, T.P. Lee, F. Ark, C. Price, D. Roeder, J. Schmitz and D.
Stockwell. 1974. Heavy metal tolerance in Typha latifolia without the evolution of
tolerant races. Ecology, 47:1163-1165.
Misra, P. and D. Chakrabarty, 2009. “Clonal Propagation of Rosa clinophylla Thory.
Through Axillary Bud Culture,” Scientia Horticulturae. 2:212- 216
Mohan, M.M., S.M. Ibrahim and S.L. Narayanan. 2000. Rooting of rose microshoots.
Research on Crops. 1(1): 67-70.
Mohapatra, A. and G.R. Rout. 2005. Identification and analysis of genetic variation among
rose cultivars using random amplified polymorphic DNA. Z. Naturforsch. C. 60: 611-
617.
Mukhambetzhanov, S., S. Nam, V. Mursalieva and I. Rakhimbaev. 2011. Micropropagation
of rose cultivar ‘Pareo’ Afri. J. Biotechn. 10(46): 9344-9346
Murali, S., D. Sreedhar and T.S. Lokeswari. 1996. Regeneration through somatic
embryogenesis from petal-derived calluses of Rosa hybrida L. ‘Arizona’ (hybrid tea).
Euphytica 91; 271-275.
Murashige, T and F.A. Skoog. 1962. A revised medium for rapid growth and bioassays with
tobacco tissue cultures. Physiol Plant 15:473-497.
Nak-Udom, N., K. Kanchanapoom and K. Kanchanapoom. 2009. Micropropagation from
cultured nodal explants of rose (Rosa hybrida L. cv. ‘Perfume Delight’). Songklanakarin
J. Sci. Technol. 31(6): 583-586.

100
Narayan, D.P. and U. Kumar. 2003. Agro's Dictionary of Medicinal Plants. Publishers
Agrobios. India. p. 292.
Nasir, M.H., R. Nadeem, K. Akhtar, M.A. Hanif and A.M. Khalid. 2007. Efficacy of
modified distillation sludge of rose (Rosa centifolia) petals for lead (II) and zinc (II)
removal from aqueous solutions. J Hazard Mater. 147: 1006-1014.
Nawaz, T., M. Hameed, M. Ashraf, F. AL-Qurainy, M.S.A. Ahmad, A. Younis and M.
Hayat. 2011. Ecological significance of diversity in leaf tissue architecture of some
species/cultivars of the genus Rosa l. Pak. J. Bot., 43(2): 873-883.
Nei, M. and H.W. Li. 1979. Mathematical model for studying genetic variation in terms of
restriction endonucleases. Proc. Natl. Acad. Sci. U.S.A. 76:5269-5273.
Nikbakht, A., M. Kafi, M. Mirmasoumi and M. Babalar. 2005. Micropropagation of Damask
Rose (Rosa damascena Mill.) cvs Azaran and Ghamsar. International Journal of
Agriculture & Biology. 4: 535-538.
Nilsson, O. 1972. Flora of Turkey and the East Aegean Islands. dinburgh University Pres,
Edinburgh. 4: 106-128.
Noodezh, H.M., A. Moieni and A. Baghizadeh. 2012. In vitro propagation of the Damask
rose (Rosa damascena Mill.). In Vitro Cell.Dev.Biol.-Plant. 48: 530-538.
Noriega, C. and M.R. Sondahl. 1991. Somatic embryogenesis in hybrid tea roses. Bio/Tech.
9; 991-993.
Nybom, H., A. Olsson and G. Werlemark. 1996. Morpholmetric variation in Nordic dogroses
(Rosa sect. Caninae, Rosaceae). Acta Univ. Ups. Symb. Bot Ups. 31: 59-68.
Nybom, H., G. Carlson-Nilsson, G. Werlemark and M. Uggla. 1997. Different levels of
morphometric variation in three heterogamous dogrose species (Rosa sect. Caninae,
Rosaceae). Plant. Syst. Evol. 204: 207-224.
Nybom, H., G. Esselink and G. Werlemark. 2004. Microsattelite DNA marker inheritance
indicates preferential pairing between two highly homologous genomes in polyploidy and
hemisexual dog roses, Rosa L. sect Caninae DC. Hered, 92: 139-150.
Olsson, A., H. Nybom and H. Prentice. 2000. Relationships between Nordic dogroses (Rosa
L. Sect. Caninae, Rosaceae) assessed by RAPDs and Fourier analysis of leaflet shape.
Syst. Bot. 25: 511-521.

101
Ozel, A.C and A, Orhan. 2006. Efficient Micropropagation of English Shrub Rose
“Heritage” Under in Vitro Conditions. J. Agr. Bio.626-629.
Ozkan G., O. Sagdic, N.G. Baydar and H. Baydar. 2004. Antioxidant and antibacterial
activities of Rosa damascene flower extracts. Food Science and Technology. 10: 277-
281.
Paek, K.Y., J.H. Seom, S.H. Son, B.H. Han and B.W. Yae. 1998. Highquality microplant
production and reduction of cost through plant tissue culture. - Korean J. hort. Sci.
Technol. 16: 272-278.
Parry, E.J. 1925. Cyclopedia of Perfumery. Blakiston's Son, Philadelphia, PA.
Pati, P.K., M. Sharma, P.S. Ahuja. 2005. Micropropagation, protoplast culture and its
implications in the improvement of scented rose. Acta. Hortic. 547: 147-158.
Pati, P.K., O. Prakash, M. Sharma, A. Sood and P.S. Ahuja. 2004. Growth performance of
cuttings raised from in vitro and in vivo propagated stock plants of Rosa damascena Mill.
Biologia Plantarum 48: 609-611.
Pati, P.K., S.P. Rath, M. Sharma, A. Sood and P.S. Ahuja. 2006. In vitro propagation of rose:
A review. Biotechnol. Adv. 24: 94-114
Pati, P.K., Sharma, M., Ahuja, P.S. 2001. Micropropagation, protoplast culture and its
implications in the improvement of scented rose. - Acta Hort. 547: 147-158.
Pawłowska, B. 2011. The effect of BA and GA3
on the shoot multiplication of in vitro
cultures of Polish wild roses. Folia Hort. 23 (2): 145-149.
Persson, H.A. and B.A. Gustavsson 2001. The extent of clonality and genetic diversity in
lingonberry (Vaccinium vitis-idaea L.) revealed by RAPDs and leaf-shape analysis. Mol.
Ecol.10: 1385-1397.
Peter Bealis, T. 1990. Botanical Rose. Grange Books, London, England.
Phillips, R. and M. Rix. 1988. Roses. Pan Books Ltd London.
Pierik, R.L.M. 1991. Horticulture new technologies and applications proceeding of the
international seminar on new frontiers in horticulture. Curr Plant Sci Biotechnol Agric.
12:141-53.
Pirseyedi, S.M., M. Mardi, S. Davazdahemami, J.M. Kermani and S.A. Mohammadi. 2005.
Analysis of the genetic diversity between 12 Iranian Damask rose (Rosa damascena

102
Mill.) using amplified fragment length polymorphism markers. Iran J Biotech, 3: 225-
230.
Podwyszyfiska, M. and M. Hempel. 1987. Effect of growth regulators applied in a medium
during multiplication and rooting stages on acclimatization of Rosa hybrida. In: Ducote
G.,
Prasad, K.V., S. Kumar and M.L. Choudhary. 2006. Molecular characterisation of fragrant
rose cultivars. Ind J Hort, 63: 229-234.
Pratpakumar, P., M. Sharma and P.S. Ahuja. 2001. Micropropagation, protoplast culture and
its implications in the improvement of scented rose. Acta Hort., 547: 147-58
Priyakumari, I. and V.L. Sheela. 2005. Micropropagation of gladiolus cv. ‘Peach Blossom’
through enhanced release of axillary buds. Journal of Tropical Agriculture 43 (1-2): 47-
50
Rajapakse, S. 2003. Genetics molecular markers, In: A. V. Roberts, T. Debener, and S.
Gudin (eds) Encyclopedia of Rose Science, Vol 1. Elsevier Academic Press Oxford UK.
pp 334-341
Rajapakse, S., D.H. Byrne, L. Zhang, and N. Anderson. 2001. Two genetic linkage maps of
tetraploid roses. Theor. Appl. Genet. 103:575-583.
Ramanayake, S.M.S.D., W.A.V.R. Wanniarachchi and T.M.A. Tennakoon. 2001. Axillary
shoot proliferation and in vitro flowering in an adult giant bamboo, Dendrocalamus
giganteus Wall. Ex Munro. In Vitro Cellu and Develop Biolo Plant. 37: 667-671.
Randhawa, G.S. and A. Mukhopadhyay, 1994. Floriculture in India. Allied Publishers Ltd.,
Bombay. India: 6- 9, 406.
Razavizadeh, R. and A.A. Ehsanpour. 2008. Optimization of in vitro propagation of Rosa
hybrida L. Cultivar Black Red. American- Eurasian J. Agric. Environ. Sci. 3(1): 96-99.
Reed, M.S. 2009. Simple-sequence Repeat Marker Analysis of Genetic Relationships within
Hydrangea paniculata. HORTSCIENCE 44(1):27-31.
Reist, A. 1985. Culture in vitro en pipiniere de rosiers: une alternative au bouturage ou au
greffage des varietes - Rev. Suisse Viticult. Arboricult. Hort. 17: 361-364.
Riaz, A., A. Younis, M. Hameed and M.A. Khan. R. Ahmed and A. Raza. 2007. Assessment
of biodiversity based on morphological characteristics among wild rose genotypes. Pak.
J. Agri. Sci., 44(2) 295-299.

103
Riaz, A., M. Hameed, A.I. Khan, A. Younis and F.S Awan. 2011. Assessment of biodiversity
based on morphological characteristics and RAPD markers among genotypes of wild rose
species. Afri. J Biotech. 10(59):12520-12526.
Ritz, C.M., H. Schmuths and V. Wissemann, 2005. Evolution by reticulation: European
dogroses originated by multiple hybridization across the genus Rosa. J. Heredity, 96(1):
4-14.
Roberts, A.V. and A. Schum. 2003. Cell, Tissue and Organ culture (Micropropagation) In:
Roberts, A.V., Debener, T., Gudin, S., Encyclopedia of Rose Science, Elsevier Academic
Press. 1: 57-63
Roder, M.S., V. Korzun, K. Wendehake, J. Plaschke, M.H. Tixier, P. Leory, and M.W.
Ganal. 1998. A microsatellite map of wheat. Genetics 149, 2007-2023.
Rogers, R.B. and M.A.L. Smith. 1992. Consequence of in vitro and ex vitro root initiation for
miniature rose production. J. Hort. Sci., 67: 535-540.
Rosu, A., R.M. Skirvin, A. Bein, M.A. Norton, M. Kushad, and A.G. Otterbacher. 1995. The
development of putative adventitious shoots from a chimeral thornless rose (Rosa
multiflora Thunb. ex J. Murr.) in vitro. J. Hort. Sci. 70:901-907.
Rout, G.R. and S.M. Jain. 2004. Micropropagation of ornamental plants-cut flowers.
Propagation of Ornamental Plants 4: 3-28.
Rout, G.R., B.K. Debata and P. Das. 1991. Somatic embryogenesis in callus culture of Rosa
hybrida L. cv. Landora. Plant Cell Tissue Organ Cult 27: 65-69.
Roy, P.K., A.N.K. Mamun and G. Ahmed. 2004. In vitro plantlets regeneration of Rose.
Plant Tissue Cult. 14(2): 149-154
Rusanov, K., N. Kovacheva, B. Vosman, L. Zhang, S. Rajapakse, A. Atanassov, and I.
Atanassov. 2005: Microsatellite analysis of Rosa damascena Mill. accessions reveals
genetic similarity between genotypes used for rose oil production and old Damask rose
varieties. Theor. Appl. Genet. 111: 804-809.
Salekjalali, M., B. Jafari and A. Tarinejad. 2011. In vitro multiplication of rose (Rosa hybrid
cv. Baccara). American-Eurasian J. Agric. & Environ. Sci, 11(1): 111-116.
Samiei, L., R. Naderi, A. Khalighi, A.A. Shahnejat-Bushehri, V. Mozaffarian, G.D. Esselink,
S. K. Osaloo and M.J.M. Smulders. 2010. Genetic diversity and genetic similarities
between Iranian rose Species. J. Horticu Sci & Biotechn. 85 (3) 231-237.

104
Savvas, D., 2002. Automated replenishment of recycled greenhouse effluents with individual
nutrients in hydroponics by means of two alternative models. Biosystems Engineering,
83: 225-236.
Scariot, V., A. Akkak and R. Botta. 2006. Characterization and genetic relationships of wild
species and old garden roses based on microsatellite analysis. J. Ameri Soc Hortic Sci.
131: 66-73.
Semblat, J.P. 1998. High-resolution DNA fingerprinting of parthenogenetic root-knot
nematodes using AFLP analysis, Mol. Ecol. 7, 119-125
Semeniuk, P. 1971b. Inheritance of recurrent and non-recurrent blooming in ‘Goldilocks’ x
R. wichuraiana progeny. J. Hered. 62:319-320.
Senapati, S.K. and G.R. Rout. 2008. Study of culture conditions for improved
micropropagation of hybrid rose. Hort. Sci. (Prague), 35 (1): 27-34.
Shabbir A, N. Hameed A. Ali and R. Bajwa. 2009. Effect of different cultural conditions on
micropropagation of rose (Rosa indica L.), Pak. J. Bot., 41: 2877-2882.
Shafiq, M., A. Younas, M. A. Khan, A. A. Khan and A. Riaz. 2006. Correlation Studies in
Rosa Species Under Faisalabad (Pakistan) Conditions. J. Agri &Soc. Sci. 2(1):58-59.
Sheppard-Hanger, S. 1995. The aromatherapy Practitioner Reference Manual. Volume (1).
Institute of Aromatherapy. Tampa. Florida, U.S.A.
Shiva, M.P., A. Lehri and A. Shiva. 2007. Aromatic and medicinal plant. Interational book
distributor. 268-276.
Short, K.C. and A.V. Roberts. 1991. Rosa spp. (Roses): In vitro culture, micropropagation
and the production of secondary products. In: Biotechnology in Agriculture and Forestry,
Bajaj YPS (Ed.), Springer-Verlag, Berlin, pp. 376-379.
Simandi, B., A. Deak, E. Ronyai, G. Yanxiang, T. Veress, E. Lemberkovics, M. Then, A.
Sasskiss and Z. Vamos-Falusi. 1999. Supercritical carbon dioxide extraction and
fractionation of fennel oil. J. Agric. Food Chem., 47: 1635-1640.
Singh, K.S. and M.M. Syamal. 2000. Anti Auxin Enhance Rosa hybrida L. Micropropagation
Biologia Plantarum. 43: 279-281.
Singh, S.P. and R.S. Kayiyar. 2001. Corelation and path coefficient analysis of flower yield
of Rosa damascene. Mill. Journal of Herbs, Spices & Medicinal Plants. 8(1): 43-49.

105
Skirvin R.M., M.C. Chu and H.J. Young. 1990. Rose. In: handbook of plant cell culture. Vol.
5. (Anmirato, P.V., Evans D.A., Sharp W.R. and Bajaja, Y.P.S., Eds.). Mcgraw-Hill
Publishing Inc., Newyork, USA, 716.743.
Sosinski, B., M. Gannavarapu, L.D. Hager, L.E. Beck, G.J. King and C.D. Ryder. 2000.
Characterization of microsatellite markers in peach [Prunus persica (L.) Batsch]. Theor
Appl Genet 101: 421-428.
Sulusoglu, M. and A. Cavusoglu. 2010. Vegetative propagation of Cherry laurel (Prunus
laurocerasus L.) using semi-hardwood cuttings. Afri. J. Agricult. Res. 5(23): 3196-3202.
Suttle, J.C. 1988. Effect of ethylene treatment on polar IAA transport, net IAA uptake and
specific binding of N-1-naphthylphthalamicacid in tissues and microsomes isolated from
etiolated pea epicotyls. Plant Physiol. 88: 795-799.
Tabaei-Aghdaei, S.R., A. Babaei, M. Khosh-Khui, K. Jaimand, M.B. Rezaee, M.H. Assareh
and M.R. Naghavi. 2007. Morphological and oil content variations amongst Damask rose
(Rosa damascena Mill.) landraces from different regions of Iran. Sci. Hort. 113: 44-48.
Tabaei-Aghdaei, S.R., H.H. Monfared, H. Fahimi, H. Ebrahimzadeh, M. Jebbely, M.R.
Naghavi and A. Babaei. 2006. Genetic variation analysis of different populations of Rosa
damascena in NW Iran using RAPD markers. Iran J Bot, 12: 121-127.
Taiz, L. and E. Zeiger. 2006. Auxin: The Growth Hormone. In: Plant Physiology. 4th edition,
Sinauer Associates, Inc. Publ. pp. 467-504.
Tautz, D. 1989. Hypervariability of simple sequences as a general source for polymorphic
DNA. Nucl Acids Res 17: 6463-6471.
Topalov, V. 1978. The Kazanlak rose and the rose production in Bulgaria. Christo G. Danov
Press, Plovdiv, Bulgaria. p. 211.
Torkashvanda, A.M. and V. Shadparvar. 2012. Rooting in hibiscus rosa- sinensis (yellow
double hybrid’) by Indole Butyric Acid and rooting substrates. Inter. J Plant, Animal
Environmental Sci. 2(2): 194-197.
Tuan, D.Q., and S.G. Iiangantileke. 1997. Liquid CO2 extraction of essential oil from star
anise fruits (Illicium verum Hook.f.). J. Food Eng., 31: 47-57.
Tucker, A.O. and M. Maciarello. 1988. Nomenclature and chemistry of the Kazanlak
Damask Rose and some potential alternatives from the horticultural trade of North

106
America and Europe. In: Flavors and Fragrances: a World Perspective, Elsevier,
Amsterdam, pp. 99-114.
Uggla, M., X. Gao and G. Werlemark. 2003. Variation among and within dogrose taxa (Rosa
sect. Caninae) in fruit weight, percentages of fruit flesh and dry matter, and vitamin C
content. Acta Agric. Scand. Section B, Soil and Plant (in-press).
Van Der Salm, T.P.M., C.J.G. Van Der Toorn, C.H. Hanischtencate and H J.M. Dons. 1996.
Somatic embryogenesis and shoot regeneration from excised adventitious roots of the
rootstock Rosa hybrida cv. ‘Money Way’. Plant Cell Rep. 15; 522-526.
Vantu., S. 2011. In vitro multiplication of Rosa canina L. 1II a. Biologie vegetala.1 (2): 19-
22.
Vasudevan., A, N. Selvaraj, P. Sureshkumar and A. Ganapathi. 2001 Multiple shoot
induction from the shoot tip explants of Cucumber (Cucumis sativus L.) Cucurbit
Genetics Cooperative Report 24: 8-12.
Vijaya, N., G. Satyanarayana, J. Prakash and R.L.M. Pierik. 1991. Effect of culture media
and growth regulators on in vitro propagation of rose. Curr Plant Sci Biotechnol Agric.
12:209-214.
Vijaya, N. and G. Satyanarayana. 1991. Effect of culture and growth regulators on in vitro
propagation of rose. 209-214 In: Prakash J. Pierik R.L.M. (eds) Horticulture- New
tecnologies and applications. Kluwer Academic Publishers, Nederlands.
Vos, P. and M. Kuiper. 1997. AFLP analysis, in DNA Markers: Protocols, Applications, and
Overviews (Caetano-Anollés, G. and Gresshoff, P.M., eds), pp. 115-131
Vos, P., R. Hogers, M. Bleeker, M. Reijans, T. van der Lee, M. Homes, A. Frijters, J. Pot, J.
Peleman, M. Kuiper and M. Zabeau. 1995. AFLP: a new technique for DNA
fingerprinting. Nucleic Acids Res. 23: 4407-4414.
Vu, N.H., P.H. Anh and D.T. Nhut. 2006. The role of sucrose and different cytokinins in the
in vitro floral morphogenesis of rose (hybrid tea) cv. ‘‘First Prize’’. Plant Cell Tiss Organ
Cult 87: 315-320.
Weaver, R.J. 1972. Plant growth substances in Agriculture. W.H. Freeman and Company.
San Francisco: p. 91-141.

107
Weisman, Z., J. Riov and E. Epstein. 1988. Characterization and rooting ability of indole-3-
butyric acid conjugates formed during rooting of mung bean cuttings. Plant Physiol., 91:
1080-84.
Weiss, E.A. 1997. Rosaceae. In Essential Oil Crops. E.A. Weiss, ed (Oxon, UK: CAB
International Wallingford), pp. 393-416.
Widrlechner, M.P. 1981. History and Utilization of Rosa damascene. Econ Bot; 35:42-58.
Wilson, D. and N.K. Nayar. 1998. In vitro propagation of rose cv. Folklore. J. Tropical
Agriculture. 36: 12-17.
Wulster G and J. Sacalis. 1980. Effects of auxin and cytokinins on ethylene evolution and
growth of rose callus tissue in sealed vessels. Hort. Sci. 15: 736-737.
Wylie, A.P. 1954. The history of garden roses. J. Roy Hort Soc 79: 555-571.
Xing, W., B. Manzhu, Q. Hongdao and N. Guogui. 2010. Micropropagation of Rosa rugosa
through axillary shoot proliferation. Acta Biologica Cracoviensia Series Botanica 52(2):
69-75.
Yakimova, E., V.K. Toteva, I. Groshkoff and G. Ivanova. 2000. Effect of BA and CPPU on
protease and amylase activity of in vitro cultured explants of Rosa hybrida L. Bulg. J.
Plant Physiol. 26: 39-47.
Yan M, D.H. Byrne and C. Jing. 1996. Propagation of rose species in vitro. In vitro cell Dev-
Pl. 32: 103-108
Yan, Z., C. Denneboom, A. Hattendorf, O. Dolstra, T. Debener, P. Stam and P.B. Visser.
2005. Construction of an integrated map of rose with AFLP, SSR, PK, RGA, RFLP,
SCAR and morphological markers. Theor Appl Genet.110: 766-777
Younis, A. 2006. Photochemical analysis and potential for exploitation of heterosis for
essential oil contents in Rosa species. Ph D Thesis, Institute of Horticultural Sciences.
University of Agriculture, Faisalabad. Pakistan. P.127.
Younis, A., A. Riaz, M.A. Khan and A.A. Khan. 2009. Effect of time of growing season and
time of day for flower harvest on flower yield and essential oil quality and quantity of
four Rosa cultivars. Flori. & Ornamental Biotech., 3: 98-103.
Younis, A., M.A. Khan, A. Ali, M.A. Pervaiz. 2006. Performance of four Rosa species under
Faisalabad agroclimatic conditions. Caderno de Pesquisa Sér. Bio., Santa Cruz do Sul, v.
18:17-27.

108
Zakizadeh, H., T. Debener, S. Sriskandarajah, S. Frello and M. Serek. 2008. Regeneration of
Miniature Potted Rose (Rosa hybrida L.) via Somatic Embryogenesis. Europ.J.Hort.Sci.,
73; 1611-4426
Zeinali, H., S.R. Tabaei-Aghdaei and A. Arzani. 2009 A Study of Morphological Variations
and Their Relationship with Flower Yield and Yield Components in Rosa damascene. J.
Agric. Sci. Technol. 11: 439-448
Zhang, L.H. D.H. Byrne, R.E. Ballard and S. Rajapakse. 2006. Microsatellite marker
development in rose and its application in tetraploid mapping. J Am Soc Hortic Sci 131:
380-387.
Zieslin, N., 1996. Influence of climatic and socio-economical factors on mode of cultivation
and research of rose plants. Acta Hortic. 424, 21-22
Zimmerman, P.W. and F. Wilcoxon. 1935. Several chemical growth substances which cause
initiation of roots and other responses in plants. Contrib . Boyce Thompson Inst . 7: 209-
229.
Zoellick, R.B. 2009. A Climate Smart Future. The Nation Newspapers. Vintage Press
Limited, Lagos, Nigeria. Page 18.
Zuker, A., T. Tzfira and A. Vainstein. 1998. Cut-flower improvement using genetic
engineering. Biotechnol. Adv. 16: 33-79.
Zvereva, E.L. and M.V. Kozlov. 2006. Consequences of simultaneous elevation of carbon
dioxide and temperature for plant herbivore interactions and metaanalysis. Global
Change Biology. 12: 27-41.
Zykov, K.I. and Z.K. Klimenko. 1999. Mutational variability of some qualitative
characteristics of garden roses. Biology Bul. 26(3): 223-229.