molecular characterization of in vivo adjuvant...
TRANSCRIPT

1
Molecular Characterization of in vivo Adjuvant Activity in Influenza-1
Vaccinated Ferrets 2
3
Running title: Adjuvant activity in seasonal flu vaccination 4
5
Yuan Fang1,2,3
, Thomas Rowe1,2
, Alberto J Leon1, David Banner
2, Ali Danesh
2,3, Luoling Xu
2, 6
Longsi Ran2, Steven E. Bosinger
6, Yi Guan
5, Honglin Chen
5, Cheryl C. Cameron
2, Mark J. 7
Cameron2, David J. Kelvin
1,2,3 4 * 8
9 1 Division of Immunology, International Institute of Infection and Immunity, Shantou University 10
Medical College, 22 Xinling Road, Shantou, Guangdong, P.R. China, 515041. 11
12 2 Division of Experimental Therapeutics, Toronto General Research Institute, University Health 13
Network, 101 College Street, Toronto, Ontario, Canada, M5G 1L7. 14
15 3
Department of Immunology, University of Toronto, Toronto, Ontario, Canada. 16
17 4 University di Sassari, Dipartimento di Scienze Biomediche, Sassari,, Italy 18
19 5 Division of Virology, International Institute of Infection and Immunity, Shantou University 20
Medical College, 22 Xinling Road, Shantou, Guangdong, P.R. China, 515041. 21
22 6
Department of Pathology and Laboratory Medicine, University of Pennsylvania, Philadelphia, 23
Pennsylvania, USA. 24
25
*Corresponding author Dr. David J. Kelvin, Division of Immunology, International Institute of 26
Infection and Immunity, Shantou University Medical College, 22 Xinling Road, Shantou, 27
Guangdong, P. R. China 515041. 28
Email: [email protected]; Tel and Fax: (+86)-754-88573991 29
30
Word count for the abstract: 181 31
Word count for the text (except reference, figure legends and table footnotes): 7,285 32
Abbreviation used: 33
ODN, oligodeoxynucleotide; PEG-IFN, pegylated interferon-alpha2b; CFA, Complete Freund’s 34
adjuvant; ISG: interferon stimulated gene; TLR, toll-like receptor; IRF, interferon regulatory 35
factor. 36
Copyright © 2010, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.J. Virol. doi:10.1128/JVI.02305-09 JVI Accepts, published online ahead of print on 9 June 2010
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

2
ABSTRACT 37
The 2009 H1N1 influenza pandemic has prompted a significant need for the development 38
of efficient, single-dose adjuvanted vaccines. Here we investigated the adjuvant potential of CpG 39
Oligodeoxynucleotide (ODN) when used with human seasonal flu vaccine in ferrets. We found 40
that CpG ODN-adjuvanted vaccine effectively increased antibody production and activated type 41
I interferon (IFN) responses, compared to vaccine alone. Based on these findings, pegylated IFN-42
α2b (PEG-IFN) was also evaluated as an adjuvant in comparison to CpG ODN and Complete 43
Freund’s Adjuvant (CFA). Our results showed that all three adjuvant-added vaccines prevented 44
seasonal human H1N1 Brisbane59 virus replication more effectively than vaccine alone. Gene 45
expression profiles indicated that, as well as up-regulating IFN-stimulated genes (ISGs), CpG 46
ODN enhanced B cell activation and increased TLR4 and IRF4 expression, whereas PEG-IFN 47
augmented adaptive immunity by inducing MHC transcription and Ras signaling. In contrast, 48
using CFA as an adjuvant induced limited ISG expression but increased the transcription of 49
MHC, cell adhesion molecules, and B cell activation markers. Taken together, our results better 50
characterize the specific molecular pathways leading to adjuvant activity in different adjuvants-51
mediated flu vaccination. 52
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

3
INTRODUCTION 53
Influenza infection is a prominent threat to human health around the world and can cause 54
severe morbidity and mortality in susceptible individuals due to acute respiratory disease. 55
Among the approaches to limit severe illness caused by influenza, vaccination is a critical 56
component in the prevention of the spread of infection. The human seasonal influenza vaccine 57
usually includes antigens from different influenza subtypes, H1N1, H3N2 and influenza type B, 58
which are predicted to circulate in the following flu season. However, this vaccine provides 59
protection to only 75% of the vaccinated population (15), and the protection efficacy in 60
immunized elderly individuals is lower than 50% (28). 61
For over 70 years, adjuvants have been used to enhance antigen-specific immune 62
responses. CpG ODN and type I IFN have been evaluated for their efficacy in commercial 63
influenza vaccines (4,53) with the general conclusion that adjuvant-mediated vaccines induce 64
stronger antibody responses and elevated cytotoxic T lymphocyte (CTL) activity. CpG ODN is a 65
ligand of toll-like receptor (TLR) 9, which is mainly expressed by plasmacytoid dentritic cells 66
(pDC), lymphocytes B and monocyte/macrophages. TLR9 stimulation by CpG can effectively 67
induce type I IFN responses and augment humoral responses (34,57). Type I IFN signaling is 68
thought to be critical to the initiation of innate immune responses to viral infections, and the IFN 69
stimulated genes (ISG), which include a variety of transcription factors, cytokines and 70
chemokines, appear to be involved in stimulating adaptive immunity and eliminating the virus 71
from the host (65). As an adjuvant, type I IFN has been shown to induce higher CTL 72
proliferation and antibody secretion than alum, and was equal to Complete Freund’s Adjuvant 73
(CFA), considered to be the gold standard adjuvant for use animal models (53). However, the 74
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

4
connection between gene regulated immune protection and adjuvant-mediated vaccination are 75
still unknown. 76
In this study, the domestic ferret (Mustela putorius furo), a well-established model to 77
study the pathogenicity of influenza virus (42), was used to better characterize adjuvant activity 78
following influenza vaccination. CpG-adjuvanted influenza vaccination resulted in an increased 79
antibody response compared to vaccine alone in ferrets. In addition, elevated ISG mRNA levels 80
were observed at an early stage post immunization, presumably in part due to IFN-α signaling 81
activation. Based on these results and the known activities of type I IFN (38,65), it was expected 82
that type I IFN could also be an effective adjuvant in influenza vaccination. Pegylated IFN-α2b 83
(PEG-IFN), which has been approved for use in humans since 2001, was therefore tested for its 84
adjuvant potential in flu vaccination in ferrets along with CFA, which activates innate immune 85
responses partly through NOD-like receptor family member NOD2 (20,27), to compare its 86
effects on immune outcome and gene regulation to those of CpG and PEG-IFN. 87
MATERIALS AND METHODS 88
Animals 89
Male ferrets 4-6 months old were purchased from Marshall Bioresources (New York 90
city, NY, USA) and maintained at the Animal Resources Centre (University Health Network, 91
Toronto, Canada). Ferrets were routinely screened for influenza infection. 92
Immunization 93
The 2007-2008 seasonal human flu vaccine, Fluviral (ID Biomedical Corporation of 94
Quebec), which contains 15 µg HA of each inactivated influenza virus strain, A/Solomon 95
Islands/3/2006 (H1N1), A/Wisconsin/67/2005 (H3N2) and B/Malaysia/2506/2004 in 0.5ml, was 96
used. Fluviral vaccine (100 µl) was mixed with 100 µl of total 500 µg class B CpG ODN (10104, 97
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

5
Coleypharma, USA) four hours before injection. Each group of ferrets was injected with either 98
200 µl PBS, 100 µl PBS plus 100 µl Fluviral or 200 µl CpG added Vaccine. At Day 35 following 99
primary immunization, all ferret groups were boosted with Fluviral vaccine alone. All the 100
animals were vaccinated through intramuscular (i.m.) injection. Fluviral vaccine was abbreviated 101
as V2007
and the Fluviral vaccine administered with PBS and CpG was abbreviated as V2007
-102
Alone and V2007
+ CpG in the text. 103
The 2008-2009 seasonal human flu vaccine Vaxigrip (Sanofi Pasteur Limited), which 104
contains A/Brisbane/59/2007 (H1N1), A/Brisbane/10/2007 (H3N2) and B/Florida/4/2006, was 105
mixed with 100 µl PBS containing 1 µg PEG-IFN-α2b and 100 µl CFA before the injection. 106
Ferrets were also vaccinated intramuscularly with CpG added vaccine, vaccine alone, and PBS. 107
The abbreviations of the vaccinations were recorded as V2008
-Alone, V2008
+ CpG, V2008
+ IFN 108
and V2008
+ CFA. The amount of CpG and CFA were chosen based on the previous studies 109
which showed optimal adjuvanticity with various antigens (50,68). The dose of PEG-IFN 110
(1µµµµg/kg) was chosen according to the manufacture’s recommendations for use of PEG-IFN 111
in humans (UNITRON PEG, Schering-Plough). 112
ELISA for Anti-Influenza antibodies 113
Serum from PBS, V2007
-Alone and V2007
+ CpG injected ferrets was collected at Days 0, 114
14, 21, 28, and 35 after primary immunization and at Day 7 post second vaccine injection. 115
Antibody responses were assessed by ELISA assay. Briefly, ELISA plates were directly coated 116
with 2007-2008 FLUVIRAL vaccine at 5 µg/ml overnight at room temperature. Plates were 117
washed with PBS containing 0.05% Tween 20 (T-PBS) and blocked with 1% bovine serum 118
albumin (BSA) for one hour at 37°C. Antigen-coated plates were washed with T-PBS and 119
incubated with 1:1,000 diluted serum samples overnight at 4°C. After washing with T-PBS, 120
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

6
plates were incubated with goat anti-ferret immunoglobulin (IgM and IgG) HRP conjugates 121
(Rockland Immunochemicals, Gilbertsville, USA) in a 1:10,000 dilution for 2 hours at 37°C. The 122
reaction was developed by o-phenylenediamine for 30 minutes and the optical density was read 123
at 450nm. 124
Haemagglutination Inhibition (HI) Test 125
Ferret serum samples were treated with receptor destroying enzyme (RDE) at 37°C 126
overnight. Fresh turkey red blood cells (TRBC) were washed and diluted in PBS to a 127
concentration of 0.5% (v/v). The non-immunized and immunized ferret sera were serially diluted 128
in PBS in 96-well V bottom cell culture plates. The serial diluted sera from PBS, V2007 + PBS 129
and V2007 + CpG groups were incubated with 25 µl (8HA units/50 µl) of A/Solomon Islands/3, 130
A/Wisconsin/67/2005 and B/Malaysia/2507/2004 strains (CDC at Atlanta, Georgia, USA) 131
separately for 15 minutes. Then, 50 µl of 0.5% TRBC was added and the plates were incubated 132
at room temperature for 30 minutes. The HI titer was the reciprocal of the highest serum dilution 133
to completely prevent agglutination. The same assay was applied on the 2008-2009 vaccine 134
immunized ferret serum by using 8 HA units/50 µl of A/Brisbane/59/2007 (H1N1) and 135
A/Brisbane/10/2007 viruses. 136
Microneutralization Assay 137
The serum neutralizing antibodies of the Fluviral and Vaxigrip immunized ferrets were 138
determined using the viruses A/Solomon Islands/3/2006 (H1N1) and A/Brisbane/59/2007 139
(H1N1) respectively for analysis by microneutralization (MN) assay described in Rowe et al., 140
1999 (58). Briefly, the 50% tissue culture infectious dose (TCID50) of each virus was determined 141
by titration in MDCK cells under biosafety level 2 conditions. The serially 2-fold diluted RDE 142
treated serum at a starting dilution of 1:10 was tested for neutralizing the 100 TCID50/50 µl of 143
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

7
each virus in MDCK cell monolayer. The cytopathic effect was read after incubation for 20 144
hours. 145
Infection and Monitoring of Ferrets 146
All the immunized ferrets and PBS control group animals were moved at least 4 days 147
prior to infection to the BSL-2 animal holding area, where they were housed in cages contained 148
in bioclean portable laminar flow clean room enclosures (Lab Products, Seaford, Del.). Prior to 149
infection, baseline temperatures were measured twice daily for at least 3 days. Ferrets were 150
anesthetized with ketamine (25mg/kg), xylazine (2mg/kg), and atropine (0.05mg/kg) by the 151
intramuscular route and infected intranasally (i.n.) with a total of 1ml of 106 EID50 of virus/ml in 152
phosphate buffered saline (PBS) delivered to the nostrils. Temperatures were measured every 153
day by using a subcutaneous implantable temperature transponder (BioMedic Data Systems, Inc., 154
Seaford, Del.). Pre-infection values were averaged to obtain a baseline temperature for each 155
ferret. The change in temperature (°C) was calculated at each time point for each animal. Clinical 156
signs of sneezing (before anesthesia), inappetence, dyspnea, and level of activity were assessed 157
once daily. A scoring system based on Reuman et al.’s work (56) was used to assess the activity 158
level as follows: 0, alert and playful; 1, alert but playful only when stimulated; 2, alert but not 159
playful when stimulated; and 3, neither alert nor playful when stimulated. A relative inactivity 160
index was calculated as follows: Σ(day 1 to day 5) [score + 1]n/Σ(day 1 to day 5) n, where n equals the total 161
number of observations. A value of 1 was added to each base score so that a score of 0 could be 162
divided by a denominator, resulting in an index value of 1.0. 163
All the infected animals were euthanatized by intracardiac injection of Euthanasia V 164
solution (1ml/kg body weight) at Day 5 p.i. Tissues from nasal turbinates and major organs 165
including lung and spleen were collected either in Trizol or in formalin for later analyses. 166
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

8
Collection of Nasal Wash and Virus Titration 167
Nasal washes were collected on Day 1, 2, 3 and 5 post infection (p.i.). Ferrets were 168
anesthetized as described above, and 0.5 ml of sterile PBS containing 1% bovine serum albumin 169
and penicillin (100U/ml), streptomycin (100µg/ml), and gentamicin (50µg/ml) was injected into 170
each nostril and collected in a petri dish when expelled by the ferret. The volume was brought up 171
to 1 ml with cold sterile PBS plus antibiotics. Sedated ferrets were weighed on Day 0 and Days 172
1, 2, 3, 4, 5 p.i. To determine the viral load in the nasal washes, 20 µl supernatant of nasal wash 173
was added to the 180 µl vDMEM (DMEM, 1% BSA, 50µg/ml gentamicin, 100 U/ml penicillin, 174
100 µg/ml streptomycin, 2.5µg/ml amphotericin B) cultured MDCK cells (2×105/ml) and 175
followed by a 10-fold dilution on a 96-well cell culture plate. After incubation for 2 hours, the 176
media was replaced with the 200 µl/well fresh vDMEM and the cells were cultured for 6 days. 177
On Day Six, 50 µl of the cultured media from each well was transferred to the V bottom 96-well 178
plate and 50 µl 0.5% TRBC was added to run a HI test. The virus titers expressed as TCID50/ml 179
from each nasal wash sample was calculated by Reed-Muench method. 180
Primer Design and Synthesis 181
Each gene primer was designed based on the conserved gene sequences obtained through 182
the alignments of the coding sequences from the various species of dog (Canis lupus), domestic 183
cat (Felis catus), human (Homo sapiens), cattle (Bos taurus) and/or pig (Sus scrofa). Primers 184
were designed specifically for detecting the ferret target genes including the paralog genes 185
of each family to produce a PCR product in the range of 60~250 base pairs (bp) using Primer3 186
software (http://frodo.wi.mit.edu/cgi-bin/primer3/primer3_www.cgi). The primers’ sequences 187
(Table 1) have GC content between 40-60% and penalties were set against self and pair 188
complements to deter primer-dimer formation. The primers were synthesized desalted under 189
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

9
HPLC purity by Operon (Huntsville, AL) and Invitrogen (Shanghai, China). The ferret specific 190
gene primers available at ATCC BEI Resources (http://www.beiresources.org) are listed in Table 191
1. 192
Cloning and Sequencing of Partial Coding Sequences of Ferret Immune Genes 193
Polymerase chain reaction (PCR)-amplified products of target genes derived from 194
mitogen-activated ferret (Mustela putorius furo) PBMCs, splenocytes and lung tissue. Amplified 195
PCR products were cloned using the TOPO-TA cloning kit (Invitrogen, Burlington, ON) per the 196
manufacturer’s instructions. Sequencing was done by the laboratory at International Institute of 197
Infection and Immunity (Shantou, China). Sequences were analyzed using the BLASTN program 198
of the National Center for Biotechnology Information portal and deposited to the public database 199
GenBank (NCBI accession numbers are listed in Table 1). 200
Whole Blood Collection, RNA Extraction and cDNA Synthesis 201
Whole blood (1.5 ml) from each ferret was collected into a Paxgene tube (Qiagen, 202
Mississauga) at Day 1, 3 and 5 after first vaccination and Day 7 post second immunization. RNA 203
was extracted and purified by Qiagen Paxgene blood RNA kit according to the manufacturer’s 204
instructions. RNA quality and concentration were determined by a spectrophotometer 205
(Eppendorf, Mississauga, Canada). 500 ng of total RNA was reverse transcribed using 206
SuperScript II reverse transcriptase (Invitrogen, Burlington, ON) in a 20 µl reaction under the 207
following conditions: 6.25 µM random hexamers primer (Applied Biosystems), 50 mM Tris-HCl 208
pH 8.3, 3 mM MgCl2, 75 mM KCl, 0.5 mM of dATP, dGTP, dTTP, and dCTP, 10 mM DTT, 40 209
U RNase Inhibitor (Applied Biosystems) and 200 U SuperScript II RNAse H- reverse 210
transcriptase at 42ºC for 1 hr. 211
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

10
Real-time PCR 212
Quantitative real time PCR (QRT-PCR) was performed using an ABI-PRISM 7900HT 213
Sequence Detection System and SYBR green PCR Master Mix (Applied Biosystems, Foster 214
City, California). Each primer pair was tested with serially diluted concentrations of a control 215
cDNA to generate a standard curve. Samples and standards were analyzed in triplicate. Each 216
QRT-PCR reaction was performed a 10 µl reaction volume with 0.25 µl of cDNA, 1 µl primers 217
(500 nM each primer) and 5 µl of SYBR green PCR MasterMix in ABI-PRISM optical 384-well 218
plates. β-actin was used as the house-keeping gene for sample normalization. 219
Microarray analysis 220
Peripheral blood RNA isolated from each animal at day 1 post vaccination was analyzed 221
by microarrays in the 2008-2009 vaccine study (n=3/group). Briefly, cRNA was prepared from 222
500 ng total whole blood RNA by two-cycle cRNA synthesis according to the manufacturer’s 223
protocol (Affymetrix, Santa Clara, CA). cRNA samples (20 µg) were labeled and hybridized to 224
Affymetrix Canine 2.0 oligonucleotide arrays to monitor gene expression of over 18,000 Canis 225
familiaris mRNA/EST-based transcripts and over 20,000 non-redundant predicted genes. To 226
minimize batch effects, RNA extraction and cRNA preparation of all samples were 227
performed at the same time. Also, cRNA samples were hybridized on the same batch of 228
microarray chips. As we have established in previous studies (2,59), canine arrays were used 229
due to the high levels of homology between canine and ferret nucleotide sequences (see the 230
homology analysis for the current study in supplemental Table S1). 231
The arrays were scanned during the same session using an Affymetrix GCS3000 7G 232
system according to standard Affymetrix protocols. Probe-level analysis was performed using 233
Probe Logarithmic Error Intensity Estimate (PLIER). The raw intensity values for each 234
individual target on the Affymetrix chips were pre-processed with variance stabilization, log2 -235
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

11
transformation and were normalized against the PBS control group datasets with ArrayAssist V 236
5.5.1 (Stratagene, USA). T-test was performed with Benjamini-Hochberg false discovery rate 237
(FDR) correction. Genes with a significant difference (p ≤ 0.05; fold change ≥ 1.5 or ≤ -1.5) 238
were selected for agglomerative hierarchical clustering with Pearson distance metrics and 239
average linkage distance measurements between clusters using MultiExperimental Viewer 4.1 240
(60). 241
Ingenuity Pathway Analysis (IPA) 5.0 software (Ingenuity Systems Inc., Redwood City, 242
CA) was used to annotate and organize the gene expression data into networks and pathways. 243
Statistics 244
One-way ANOVA was used for statistical analysis of the results represented in Fig. 4A 245
and Fig. 6. Other analyses used the Student’s t-test for comparing two independent populations. 246
Quantitative PCR results from 50 ferret immune-related genes were represented as heat map 247
charts generated by the software MultiExperimental Viewer 4.1. Bar graphs were generated by 248
SigmaPlot 8.0. 249
RESULTS 250
Humoral responses of CpG ODN-mediated immunization 251
We first investigated the effects of CpG ODN as an adjuvant for the 2007-2008 seasonal 252
human flu vaccine, Fluviral (V2007
) in ferrets. Fluviral contains 15µg of HA protein from the 253
following influenza virus strains: A/Solomon Islands/3/2006 (H1N1), A/Wisconsin/67/2005 254
(H3N2) and B/Malaysia/2506/2004. Ferret serum samples were collected at time points 255
following the primary immunization and vaccine alone boost. Immune activation was determined 256
by anti-vaccine IgM and IgG antibody serum levels based on ELISA assays. Serum samples 257
from V2007
+ CpG treated animals had significantly higher IgM levels at Day 14 after primary 258
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

12
immunization (Fig. 1A, left panel), when compared to the IgM level of V2007
-Alone group. 259
Interestingly, there were also statistically higher levels of antigen specific IgG in V2007
+ CpG 260
sera than V2007
-Alone treated animals at all the time points after primary vaccination and also at 261
Day 7 post boost (Fig. 1A, right panel). These results suggest that CpG ODN mediated influenza 262
vaccination increases humoral immunity by inducing antibody production and promoting faster 263
antibody class switching. 264
To confirm the results of ELISA assays, the antibody titers were also assessed by 265
haemagglutination inhibition (HI) and microneutralization (MN) assays. HI assays indicated that 266
the sera from V2007
+ CpG treated animals contained significantly higher levels of antibody titers 267
than the V2007
-Alone treated animals (Fig. 1B) after primary vaccination to Day 7 post boost. The 268
MN assay also showed that the serum antibody from V2007
+ CpG treated animals had 269
significantly higher titers against the A/Solomon Islands/3/2006 (H1N1) virus than the animals 270
treated with vaccine alone at all the time points (Fig. 1C). These data further indicate that using 271
CpG as adjuvant is able to augment the humoral immune response to influenza vaccinations. 272
Regulation of ISGs by CpG adjuvanted vaccination 273
Having shown that the addition of CpG ODN to V2007
upregulated humoral responses 274
after vaccination, we assessed gene expression activity associated with increased 275
immunogenicity at mRNA level. Quantitative PCR (Q-PCR) was used to generate a 50-gene 276
expression profile from the whole blood RNA samples collected at Days 1, 3, and 5 after primary 277
immunization and Day 7 post boost in the vaccination groups V2007
+ CpG, V2007
-Alone and PBS 278
V2007
Boost. The PBS treated group was used as control for normalization at Days 1, 3 and 279
5 after primary vaccination and at Day 7 post boost. The identical gene expression pattern 280
was observed in CpG adjuvanted and non-adjuvanted vaccination groups following 281
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

13
normalization to PBS group at day 0 (data not shown). Since most of the immune related 282
genes were significantly regulated at Day 1 in the V2007
+ CpG group compared to the control 283
group, the Day 1 Q-PCR data of 50 genes was illustrated by heat map (Fig. 2A). The expression 284
levels of the 12 immunologically relevant genes, from Day 1 after primary vaccination to Day 7 285
post boost, were statistically analyzed and represented by bar graphs (Fig. 2B, 3). 286
ISG molecules, such as the anti-viral genes OAS1, RIG-I, Mx-1, ISG15, ISG20; 287
transcription factors IRF1, IRF7, STAT1, STAT2; and the Th1 cell chemokine CXCL10 (54), 288
which are induced by type I IFN during the virus infection, were significantly elevated at Day 1 289
after primary vaccination (Fig. 2A). However, at later time points the expression level of most 290
ISGs decreased to basal level or became downregulated, such as OAS1, IRF7 and RIG-I. Only 291
CXCL10 and STAT1 showed sustained increased expression levels at Day 7 after boost (Fig. 292
2B). Although the ISGs were modulated similarly in the group without adjuvant, at Day 3 and 293
Day 5 post primary vaccination the induced level of genes such as OAS1, RIG-I, ISG15 and 294
STAT2 were markedly lower than with CpG (Fig. 2B). This observation suggested that CpG 295
stimulated a faster and stronger type I IFN response than that in the animals treated with vaccine 296
alone. 297
Given that CpG mediated vaccination strongly induced ISGs, we then determined 298
whether IFN regulated ISG expression in vaccinated animals. We examined the mRNA level of 299
IFN-α and IFN-γ by Q-PCR. IFN-α was elevated in the V2007
+ CpG group at Day 1 after 300
primary vaccination and Day 7 post boost, whereas, IFN-γ levels were not highly increased by 301
vaccination plus CpG (Fig. 3A). Even though V2007
-Alone immunization increased the IFN-α 302
level at Day 3 and Day 5, it was not statically significant when compared to the V2007
+ CpG 303
group (Fig. 3A). Several studies have reported that IFN-stimulated response repressors, such as 304
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

14
IRF-2, ICSBP and IRF4/PIP, likely terminate ISG transcription following IFN induction 305
(18,48,69). In this study, IRF4 was highly upregulated in the V2007
+ CpG group at Day 5 after 306
primary vaccination and Day 7 post boost (Fig. 3B). Since ISG expression was low during these 307
time points, these results implied a negative role for IRF4 in the regulation of IFN-stimulated 308
genes during adjuvant-mediated vaccination. Since IRF4 is also involved in the B cell class 309
switch recombination and plasma cell differentiation (32,40,41), the elevated IRF4 expression in 310
V2007
+ CpG immunized animals may indicate a role in humoral immunogenicity regulation. 311
IFN-α and CFA-mediated vaccination 312
We next evaluated IFN-α as an adjuvant to human influenza vaccination to determine if 313
the CpG-induced IFN response was responsible for the induced humoral response. Pegylated 314
IFN-α (PEG-IFN) was tested as an adjuvant for the 2008-2009 human flu vaccine (V2008
), 315
Vaxigrip, in ferrets. The activity of pegylated IFN-α was evaluated by antibody titer 316
quantification, and compared to the response of CpG ODN (V2008
+ CpG), CFA (V2008
+ CFA), 317
vaccine alone (V2008
-Alone) and PBS control groups. To determine antibody titers, HI and 318
microneutralization assays were run using serum samples collected on Days 14, 21, 28 and 35 319
post V2008
immunizations. Quantification of HI titers for the live influenza A viruses 320
Brisbane/59/2007 H1N1 and Brisbane/10/2007 H3N2 determined that CpG, PEG-IFN and CFA 321
mediated vaccinations induced significantly higher titers than vaccine alone (Fig. 4A). 322
Furthermore, the microneutralization assay showed that V2008
+ CpG, V2008
+ IFN and V2008
+ 323
CFA immunized ferrets had higher titers of Brisbane/59/2007 H1N1 neutralizing antibody than 324
V2008
-Alone at Day 14 after vaccination (Fig. 4B). Later time points showed only CpG and CFA 325
mediated vaccinations stimulated statistically stronger antibody responses. In a further 326
comparison of antibody induction among the adjuvanted immunizations, both CpG and CFA 327
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

15
adjuvanted vaccines stimulated significantly higher antibody titers than the PEG-IFN adjuvanted 328
vaccine. However, significant differences were not observed between the CpG and CFA groups. 329
These results indicate that adjuvant addition increases humoral responses and that V2008
+ CFA 330
produces the strongest responses of the three adjuvant-mediated vaccinations. 331
Virus infectivity in adjuvant-treated animals 332
We next determined whether an increase in humoral immunity led to increased protection 333
from viral infection. Ferrets were immunized with the following combinations: PBS, V2008
-334
Alone, V2008
+ CpG, V2008
+ IFN and V2008
+ CFA. At day 42 following immunization, the 335
ferrets were infected with 106 EID50 Brisbane/59/2007 H1N1 virus to evaluate the protection 336
efficacy of adjuvant-mediated vaccinations. Brisbane/59/2007 H1N1 virus has been found to 337
replicate only in the nasal cavity of ferrets (47,59). From Day 2 p.i., 50-75% of the animals in 338
each group exhibited clinical signs of respiratory disease, including nasal discharge and 339
sneezing. The symptoms lasted for 2-3 days and decreased at Day 5 p.i. All animals exhibited 340
increased temperature at Day 2 p.i., which diminished by Day 3, except the groups of V2008
-341
Alone, V2008
+ IFN and PBS, which showed a second increase in temperature on Day 4 p.i.. In 342
addition, the animals displayed a decrease in activity by Day 3 p.i., except the animals 343
immunized with V2008
+ CFA. The relative inactivity index (Table 2) showed that V2008
+ CFA 344
was 1.1, V2008
+ CpG and V2008
+ IFN were both 1.3, and V2008
-Alone and PBS control groups 345
were 2.0 and 1.6, respectively. These results suggest that adjuvant-mediated vaccination in 346
ferrets dramatically reduces influenza severity compared to receiving the vaccine alone. 347
To evaluate infectivity, nasal washes were collected from animals on each day post 348
infection and viral loads (TCID50/ml) were calculated. Although viral loads in each group were 349
elevated at Day 1 and Day 2 p.i., the virus replication in the nasal turbinates of the V2008
+ CFA 350
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

16
group was significantly lower than in the PBS control group. Furthermore, the Day 3 p.i. virus 351
titer measured in nasal washes of adjuvant groups were markedly diminished compared to those 352
of the PBS control (Fig. 5), whereas no significant decrease was observed in the ferrets that 353
received vaccine alone. By Day 5 p.i., the virus titer was not detectable in the majority of 354
animals. Interestingly, the assessment of antibody titer in serum at Day 5 p.i. showed that 355
adjuvant-immunized animals had significantly higher antibody levels than animals treated with 356
vaccine alone, as determined by HI and neutralization assays (Fig. 6A, B). Additionally, in nasal 357
washes collected at day 5 p.i., the anti-Brisbane/59 IgG levels in the CpG and CFA adjuvanted 358
vaccination groups were significantly higher than the vaccine alone (data not shown). Given the 359
clinical results and the decreased viral loads and increased antibody production observed in 360
animals vaccinated with adjuvant compared to animals receiving the vaccine alone, we suggest 361
that adjuvant induces a faster and stronger memory antibody response upon infection. 362
Microarray analysis of CpG adjuvant immunization 363
Above we showed that the expression of our selected immune related genes changed 364
most significantly at Day 1 after vaccination with CpG compared to control (Fig. 2A). We next 365
investigated the large scale gene expression profile of Day 1 adjuvant vaccinations by microarray 366
analysis. The heat map (Fig. 7) represents the expression of immune-related genes significantly 367
altered at Day1 post vaccination from adjuvant-mediated V2008
vaccinations (one ferret per 368
column). A similar expression pattern was observed by Q-PCR for three genes selected from the 369
antigen presentation, adaptive immunity and complement clusters shown in Fig. 7 (see 370
supplemental Fig. S1). Fold gene expression changes for each adjuvant-treated group compared 371
to PBS controls are listed in Table 3. Also, we used vaccine alone as a comparator and 372
observed a similar expression pattern to that of the PBS group (data not shown). 373
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

17
Consistent with our Q-PCR results of V2007
+ CpG vaccination, which showed 374
upregulation of 10 ISGs, the microarray data determined that 15 ISG members were increased by 375
at least 1.5 fold in V2008
+ CpG immunization (Fig. 8A, Table 3). The Ingenuity Pathway 376
Analysis (IPA), combining the Q-PCR and microarray data, suggests that CpG-adjuvanted 377
vaccination stimulates type I IFN signaling and activates ISG members through STAT1 and 378
STAT2 (Fig. 8B upper panel). Also, toll-like receptor 4 (TLR4), which specifically recognizes 379
bacterial lipopolysaccharides (LPS) and mediates innate immunity (31), was induced following 380
CpG-adjuvanted vaccination (Table 3). Transcription analyses showed that co-stimulatory 381
molecules and MHC genes were not elevated by vaccination with CpG, except for the 382
upregulation of cathepsin family antigen processing genes, CTSB and CTSS. Similarly, neither 383
microarray nor Q-PCR showed increased IFN-γ levels after CpG-mediated vaccination when 384
compared to the vaccine alone. Interestingly, the immune regulator SOCS1 (Table 3), which 385
negatively regulates IFN-γ signaling (10), was found to be upregulated in V2008
+ CpG 386
immunized animals by microarray analysis. 387
Microarray analysis of IFN-α adjuvant immunization 388
Given that CpG-mediated vaccination significantly increased ISGs, we tested IFN-α for 389
its direct adjuvant potential during immunization against influenza. We have found that 390
treatment with PEG-IFN alone resulted in significantly increased mRNA levels of ISGs in ferrets 391
at early time points (unpublished data). Here, microarray analysis revealed that 345 genes were 392
highly upregulated following V2008
+ IFN vaccination. Similar to the gene profile of CpG 393
mediated immunization, V2008
+ IFN stimulated the expression of ISGs, including STAT1, IRF2, 394
OAS1/2, ISG15, USP18, IFIT2, RSAD2, IFI44 and IFI44L (Fig. 8A), although to a lesser extent 395
than the V2008
+ CpG group, shown by the IPA analysis (Fig. 8B lower panel). 396
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

18
V2008
+ IFN immunization also upregulated antigen processing and presentation genes, 397
such as CTSD and MHC class Ib, as well as the T cell adhesion molecule, CD84 (66) (Table 3). 398
We also found the MHC enhanceosome member RFX5, which regulates MHC during antigen 399
presentation (55), was significantly increased following PEG-IFN mediated vaccination. 400
Interestingly, PEG-IFN mediated vaccination also induced the gene expression of intracellular 401
signaling molecules Ras and the MEK member MAP2K2 (Fig. 8C), which are known mediators 402
of B cell proliferation and B cell memory after CD40 stimulation (7,8). JUNB, which is 403
downstream of Ras following BCR stimulation (70), was also up-regulated in V2008
+ IFN 404
vaccination. Additionally, the expression of the guanine nucleotide activating protein (GAP), 405
RGS1, involved in B cell activation (26), which was elevated in the V2008
+ CpG group, was also 406
increased in V2008
+ IFN immunized ferrets (Table 3). 407
Microarray analysis of CFA adjuvant immunization 408
The precise molecular mechanism of CFA-mediated influenza immunization is not 409
completely understood. We therefore investigated the gene profile of CFA-adjuvanted influenza 410
vaccination in ferrets. Microarray analysis revealed that a total of 1,255 genes were regulated at 411
Day 1 post V2008
+ CFA immunization. Since CFA is thought to contain the NOD2 agonist 412
MDP, which induces NF-κB and MAPKs to initiate proinflammatory cytokine expression in 413
innate immune cells (36), we examined the expression of NOD2 signaling genes. The expression 414
of the MAP Kinase family member ERK was induced at Day 1 after CFA mediated vaccination 415
(Table 3), but proinflammatory cytokines were not induced. 416
CFA may also contain a TLR ligand, which could synergistically activate DCs with MDP 417
(19,27). Similar to CpG- and IFN-mediated vaccination, the expression of antigen processing 418
genes, namely CTSD, MHC class I DLA-64, MHC class II HLA-DMA/CLIP (CD74) and MHC 419
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

19
transcription enhancesome RFX5 (Fig. 7, Table 3) were significantly increased in CFA-mediated 420
immunization. However, the MHC class II transactivator (CIITA), which regulates MHC class II 421
gene transcription, was significantly down-regulated (Table 3). We also found that CFA 422
activated the expression of several adhesion molecules, including Integrin-β, VCAM1, BCAM, 423
CD36, CD84 and CD44 (Fig. 9A), and genes such as Zyxin, Talin 1, CRKL, RAP2A and Rho 424
family members involved in integrin signaling and mediating cell adhesion and motility (Fig. 425
9B). Moreover, expression of CXCL14, a chemokine involved in B cell migration and activation 426
(63), was also found elevated (Table 3). 427
B cell intracellular signaling molecules, such as Lyn, Syk, and PI3K, were significantly 428
up-regulated by CFA at Day 1 (Fig. 9A, C). In addition, increased mRNA level of B cell 429
proliferation regulator Bam32 (22,43) was observed in CFA-mediated vacination. In contrast to 430
the PEG-IFN-induced transcription of MEK member MAP2K2, involved in Ras mediated BCR 431
signaling, CFA activated the expression of Ras downstream effectors, c-Raf and MAP Kinase 432
ERK (Fig. 9C). Following BCR engagement, Ras activation is one result of Ca2+
signaling (9) 433
which leads to gene induction through the calcineurin-NFAT pathway to stimulate B cell 434
proliferation (23). Indeed, the calcineurin family members PPP3R1, NFAT molecule NFATC3, 435
and the calcineurin-NFAT activation regulators G protein complex (GNA15, GNB3, GNG2 and 436
GNG11) and CK1 (CSNK1D and CSNK1G2), were all highly induced following CFA-437
adjuvanted vaccination (Fig. 9D). Moreover, we found that the small G protein Rac, another 438
factor which activates NF-κB and JNK signaling (6,25,45), was induced after vaccination with 439
CFA (Fig. 9C) and AICDA (AID), essential for initiating the immunoglobulin gene 440
hypermutation (SHM) and class switch recombination (CSR) in B cells (3), was increased by 2.6 441
fold in CFA mediated vaccination (Table 3). Although AID was induced, only one of its 442
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

20
activator molecules, Oct2 (51), was significantly increased whereas the other, HoxC4, was 443
downregulated (Fig. 9C). 444
Lastly, SOCS family member, SOCS3, was induced by V2008
+ CFA treatment (Table 3). 445
High SOCS3 expression may favour Th2 cell differentiation and inhibit IL12-mediated Th1 446
development (62), which is consistent with our finding that IL12A (Table 3) was significantly 447
down-regulated in CFA-adjuvanted vaccination. Also, more complement components, such as 448
C1QL2, C1R, C5AR1 and CFI (Fig. 7, Table 3), were activated in V2008
+ CFA immunization 449
compared with the other adjuvanted immunizations which upregulated only FCN1 and C3 450
precursor. However, V2008
+ CFA stimulated few ISGs, such as transcription regulator IRF1, 451
IRF2, and antiviral gene RSAD2 (Table 3), when compared to CpG and PEG-IFN adjuvanted 452
vaccinations. 453
DISCUSSION 454
Here we investigated the adjuvant potential of CpG ODN when added to the seasonal 455
human influenza vaccine, and subsequently characterized the molecular gene signatures of the 456
induced immune responses. Since CpG ODN initiated robust IFN responses following 457
vaccination, we evaluated type I IFN as an adjuvant by using PEG-IFN in combination with the 458
human influenza vaccine, Vaxigrip. We also compared the effect of PEG-IFN to the standard 459
adjuvant, CFA. To demonstrate the activity of each adjuvant during immunization, gene 460
expression profiling was performed by microarray. 461
Gene expression profile generated by Affymetrix canine arrays was used to expand 462
on the gene signatures identified with Q-PCR analysis (Fig. 2). At this time, it is not known 463
whether microvariation in the canine probes affects detection of ferret orthologs. Our 464
group has established the utility of canine platform to assess ferret gene expression using a 465
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

21
homology analysis of the limited publicly available ferret and canine cDNA sequences and 466
a ferret-specific Q-PCR validation strategy (2,59). Cross-species microarray analyses are 467
supported by previous studies (11,30,49), however, a lack of publicly available canine whole 468
blood microarray datasets has not allowed us to make the same comparisons. Nonetheless, 469
high homology was identified between numerous ferret and canine genes derived from the 470
current study (Table S1) and extensive ferret-specific Q-PCR validation on surrogate genes 471
was performed. 472
The CpG experiment showed that our genes of interest changed most significantly at day 473
1 following vaccination (Fig. 2), therefore this time point was chosen for more extensive 474
microarray analyses. We demonstrated that although CpG ODN led to the activation of ISGs and 475
subsequent stimulation of humoral responses, when PEG-IFN was used it was unable to 476
reproduce the humoral activation to the same extent as that of CpG ODN. Furthermore, CpG 477
ODN as well as IFN and CFA adjuvant-mediated immunizations stimulated stronger antibody 478
responses than the vaccine alone and gave better protection for animals following seasonal H1N1 479
infection. The microarray analysis for each adjuvant showed differing molecular signatures 480
indicating that distinct molecular pathways were activated depending on the adjuvant used. 481
Based on microarray profiling and Q-PCR analysis, the CpG-adjuvanted immunization 482
activated canonical type I IFN signaling responses. The upregulation of ISGs included the virus 483
sensing RIG-I and PKR, the virus replication inhibitors OAS1, 2 and L, the transcription factors 484
STAT1, STAT2, the IFN-regulating molecules IRF1, IRF7, and the Th1 cell chemoattractant 485
CXCL10. In plasmacytoid DC (pDC), CpG binds to the intracellular receptor TLR9 to activate 486
the IFN-α expression, which initiates the ISG stimulation (21). In our study, the ISG induction 487
was transient after vaccination and the relatively increased IFN-α expression was only observed 488
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

22
at 24 hours post vaccination. Typically, pDCs are thought to be the primary producers of type I 489
IFN and that production of IFN following CpG stimulation leads to maturation of the 490
conventional DCs (cDCs), which in turn augments the B cell response toward a Th1-like 491
phenotype by inducing IFN-γ and IL12 (33). Our gene profiling did not support this mechanism 492
for CpG-induced pDC and cDC-mediated enhancement of humoral response through Th1 493
regulation, since molecular signatures associated with DC maturation and/or Th1 signaling were 494
not observed. Moreover, we found that SOCS1, the suppressor of IFN-γ signaling (10), was 495
significantly up-regulated. Therefore, CpG may function as a non-Th1 biasing adjuvant during 496
immunization, a possibility we are studying further. 497
Several recent studies indicate that CpG can directly activate B cells through TLR9 and 498
MyD88 to promote class switching toward a Th1 phenotype (29,39). In addition, TLR9 signaling 499
has been implicated in plasma cell proliferation and differentiation after immunization with 500
antigen and non-soluble CpG, which leads to IL-6 and IgM secretion (14). Here we found that 501
IRF4, which is involved in antibody class-switching and plasma cell differentiation (17,32,41), 502
was upregulated in CpG-mediated vaccination. Since both ligation of TLR9 and intracellular 503
activation of IRF4 regulate antibody class switching, we postulated that the IRF4 could be 504
induced downstream of TLR9 engagement in B cells. In this scenario, it is possible that 505
downstream activation of IRF4 may occur via MyD88-mediated NF-κB and/or AP-1 activation, 506
which has been a proposed route for B cell activation, proliferation and Ig production after TLR9 507
anchoring (52). High levels of IRF4 occurring after the antigen boost also suggests that IRF4 508
could play a role in the memory plasma cell differentiation, which is consistent with the findings 509
in the study by Klein et al. (32). Taken together, our data suggests that CpG-adjuvanted 510
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

23
vaccination activates B cells via TLR9-mediated expression of genes such as IRF4, involved in 511
plasma cell differentiation and antibody class switching. 512
Interestingly, the microarray profiling of CpG mediated vaccination indicated that TLR9 513
activation correlated with TLR4 upregulation. TLR4 cooperates in BCR signaling to enhance the 514
antibody response through LPS ligation (13,46). Although LPS is the classic antigen for TLR4, 515
an updated list asserts that other pathogens, such as viral protein and parasitic heat shock 516
proteins, also bind to this innate receptor (1). Thus, TLR4 may be also involved in immune 517
activation by recognizing the vaccine antigen in conjunction with TLR9-mediated vaccine 518
responses. 519
As discussed above, type I IFN induces adaptive immunity by stimulating DC cell 520
surface co-stimulatory molecules and MHC antigens which enable DCs to activate B cells (37). 521
Since type I IFN is an inducer of adaptive immunity, we used pegylated IFN-α2b as an adjuvant 522
to compare with CpG-adjuvanted immunization. PEG-IFN induced similar ISG expression to 523
CpG-adjuvanted vaccination as determined by microarray analysis (Fig. 8 b). The MHC class I 524
gene was also induced possibly as a result of increased levels of the MHC transcription 525
enhancesome RFX5. An increase of the MHC class I molecule on the surface of antigen 526
presenting cells (APCs) can then activate T cells to express the adhesion molecule CD84. Since 527
RFX5 was also up-regulated in CFA adjuvanted vaccination, it is possible that RFX5 was 528
activated by the same signaling pathway and may represent as a common mediator of adjuvant 529
activity in both PEG-IFN and CFA mediated vaccination. In the future, it will be important to 530
elucidate the mechanism of RFX5 induction since MHC upregulation in our adjuvant-mediated 531
studies are important components of adaptive immunity. Interestingly, PEG-IFN stimulated Ras 532
and MEK genes involved in Ras-MEK-ERK signaling. Ras-MEK-ERK signaling in B cell plays 533
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

24
an important role in generating the high affinity antibody (61). It is therefore likely that type I 534
IFNs can directly activate B cells through type I IFN receptor to initiate the Ras-MEK-ERK 535
pathway, as type I IFNs can stimulate B cells directly to produce antibody and express IFIT2/3 536
early after influenza infection (5,61). 537
Our microarray data suggest that IFN-α2b activates MHC class I expression which may 538
play a role in activating CD8+ T cells post-vaccination. Furthermore, the data implies that IFN-α 539
activates antibody generation by B cells through Ras signaling. However, our HI and 540
microneutralization assays showed antibody levels induced by PEG-IFN were lower than those 541
stimulated by CpG (Fig. 2). This decreased antibody production may be dose-dependent since 542
previous reports have shown that the Ig subclass was highly increased at a high dose of IFN-α 543
adjuvant administration (67); or possibly the IFN-α2b subtype could not fully activate the DC or 544
B cell, since IFN-α1 is the main subtype produced by pDC after virus infection. 545
CFA has been used for decades in animal models to generate high levels of antibody 546
against antigens. It is known that MDP, the NOD2 ligand, is the minimal essential component in 547
CFA (16). Unlike previous findings (36), the results of our microarray profiling in ferrets showed 548
that CFA-mediated vaccination did not stimulate a proinflammatory milieu but instead only 549
showed the elevation of the MAPK pathway gene ERK. Weak proinflammatory cytokine 550
responses, e.g. absence of IFN-γ gene expression, may be the result of a low dose of CFA 551
treatment (100 µµµµl, one injection) given to the ferrets. Even though a robust inflammatory 552
response was not observed, the transcription enhancesome RFX5 was upregulated upon CFA-553
adjuvanted vaccination along with MHC gene induction, which is associated with DC maturation 554
(44). Previous reports have shown strong synergism between TLR ligands and MDP when 555
administered together (64). Therefore, it is possible that there may be another ligand in CFA that 556
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

25
contributes to the adjuvant activity. In particular, the TLR4 ligand can increase the maturation 557
potential of MDP on human DC by inducing co-stimulatory molecules and the MHC class II 558
gene (19). Furthermore, NOD2/TLR mediated MHC upregulation may be regulated by the 559
induced SOCS3, which is involved in the negative regulation of STAT3 and associated with 560
Th2-type signaling in DC (35). This is supported by our finding that the Th1-directing 561
cytokine IL12A (Table 3) was significantly down-regulated in CFA-adjuvanted 562
vaccination. 563
Mature DCs efficiently present antigens on the cell surface and directly promote 564
expression of B cell high-affinity receptors. This results in strong BCR signaling and subsequent 565
interaction with T helper cells to initiate antibody secretion cells and memory B cells (12). In this 566
study, we found that several important genes involved in B cell activation were induced in CFA 567
mediated vaccination, in particular, the molecules involved in Lyn-Syk-PI3K signaling pathway, 568
calcineurin-NFAT pathway and Ras-MEK-ERK pathway. Furthermore, our pathway analysis 569
suggested that B cell activation may have induced Calcineurin-NFAT, Ras-MEK-ERK and NF-570
κB signaling (24). These pathways in turn initiate the transcription of genes involved in B cell 571
expansion, plasma cell differentiation and antibody production, such as Bam32 (22), and Oct2 572
(POU2F2), which binds to the AICDA promoter and activates AID transcription (51) for 573
determining the antibody SHM and CSR (3). In addition, the molecules engaged in cell adhesion 574
and motility, complement components, and chemoattractant were highly stimulated in CFA 575
adjuvanted vaccination. 576
In contrast to type I IFN activation by CpG ODN or PEG-IFN, CFA stimulated a 577
relatively low level of ISG expression, which is unlike previously published findings that robust 578
type I IFN activation is a hallmark of CFA activity in mouse models (37). Given that CFA was 579
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

26
administered with vaccine intramuscularly, the slow release of this emulsion from the injection 580
site to the peripheral circulation, or the more localized activity of this adjuvant, may result in the 581
limited ISG stimulation in peripheral blood at 24 hours post vaccination. It will be important to 582
evaluate gene regulation in the peripheral blood at later time points following CFA adjuvanted 583
vaccination to demonstrate the role of IFN responses in the adjuvanticity of CFA. 584
Altogether, our data suggests that the low dose of CFA activates the expression of MHC 585
molecules which are associated with DC activation through NOD2 and/or TLR signaling rather 586
than type I IFN receptor ligation. We contend that the activated DC may strongly activate BCR 587
signaling to initiate the B cell proliferation and plasma cell differentiation through high-Ca2+
-588
induced NFAT, ERK and NF-κB-regulated transcription. Given the paucity of ferret-specific 589
reagents, we could not isolate DC in this study to investigate this potential mechanism further. 590
Also, we were limited to one time point of gene expression profiling and cannot rule out 591
that some of the expression differences may be affected by different gene expression 592
kinetics or different dose of adjuvants. 593
In conclusion, we have identified both common and disparate signaling pathways 594
activated downstream of in vivo adjuvant activity during vaccination. Of note, RFX5 was a 595
common transcript induced by both IFN-α and CFA-adjuvanted vaccinations, representing a 596
focal point of adjuvant activity. Additionally, the identified signature molecules in our study 597
could be specifically targeted in future vaccines, thereby facilitating the efficacy of vaccination 598
and the development of host immunogenicity. 599
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

27
ACKNOWLEDGEMENTS 600
We thank Dr. Alexander Klimov (CDC at Atlanta, USA) for the kind gift of the 2007-601
2008 human seasonal influenza viruses, and Lixia Guo and Zujiang Li at Shantou University 602
Medical College (Shantou, China) for assisting with ferret gene cloning. We would also like to 603
thank Jean Flanagan and Roman Skybin, from Animal Resources Centre at Toronto General 604
Hospital (Toronto, Canada), for the collection of ferret samples. We would like to give special 605
thanks to Dr. Alyson Kelvin for reviewing the manuscript. 606
This work is supported by grants from the Li Ka Shing Foundation, the Canadian 607
Institute of Health Research, Sardegna Ricerche and the National Institutes of Health. 608
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

28
REFERENCES 609
610
1. Aosai, F., M. Chen, H. K. Kang, H. S. Mun, K. Norose, L. X. Piao, M. Kobayashi, O. 611
Takeuchi, S. Akira, and A. Yano. 2002. Toxoplasma gondii-derived heat shock protein 612
HSP70 functions as a B cell mitogen. Cell Stress. Chaperones. 7:357-364. 613
2. Cameron, C. M., M. J. Cameron, J. F. Bermejo-Martin, L. Ran, L. Xu, P. V. Turner, 614
R. Ran, A. Danesh, Y. Fang, P. K. Chan, N. Mytle, T. J. Sullivan, T. L. Collins, M. G. 615 Johnson, J. C. Medina, T. Rowe, and D. J. Kelvin. 2008. Gene expression analysis of 616
host innate immune responses during Lethal H5N1 infection in ferrets. J. Virol. 82:11308-617
11317. 618
3. Chaudhuri, J., M. Tian, C. Khuong, K. Chua, E. Pinaud, and F. W. Alt. 2003. 619
Transcription-targeted DNA deamination by the AID antibody diversification enzyme. 620
Nature 422:726-730. 621
4. Cooper, C. L., H. L. Davis, M. L. Morris, S. M. Efler, A. M. Krieg, Y. Li, C. 622
Laframboise, M. J. Al Adhami, Y. Khaliq, I. Seguin, and D. W. Cameron. 2004. Safety 623
and immunogenicity of CPG 7909 injection as an adjuvant to Fluarix influenza vaccine. 624
Vaccine 22:3136-3143. 625
5. Coro, E. S., W. L. Chang, and N. Baumgarth. 2006. Type I IFN receptor signals directly 626
stimulate local B cells early following influenza virus infection. J. Immunol. 176:4343-627
4351. 628
6. Coso, O. A., M. Chiariello, J. C. Yu, H. Teramoto, P. Crespo, N. Xu, T. Miki, and J. S. 629
Gutkind. 1995. The small GTP-binding proteins Rac1 and Cdc42 regulate the activity of 630
the JNK/SAPK signaling pathway. Cell 81:1137-1146. 631
7. Coughlin, J. J., S. L. Stang, N. A. Dower, and J. C. Stone. 2005. RasGRP1 and 632
RasGRP3 regulate B cell proliferation by facilitating B cell receptor-Ras signaling. J. 633
Immunol. 175:7179-7184. 634
8. Coughlin, J. J., S. L. Stang, N. A. Dower, and J. C. Stone. 2006. The role of RasGRPs in 635
regulation of lymphocyte proliferation. Immunol. Lett. 105:77-82. 636
9. Cullen, P. J. and P. J. Lockyer. 2002. Integration of calcium and Ras signalling. Nat. 637
Rev. Mol. Cell Biol. 3:339-348. 638
10. Dalpke, A. H., S. Opper, S. Zimmermann, and K. Heeg. 2001. Suppressors of cytokine 639
signaling (SOCS)-1 and SOCS-3 are induced by CpG-DNA and modulate cytokine 640
responses in APCs. Journal of immunology (Baltimore, Md. : 1950. ) 166:7082-7089. 641
11. Dillman, J. F., III and C. S. Phillips. 2005. Comparison of non-human primate and 642
human whole blood tissue gene expression profiles. Toxicol. Sci. 87:306-314. 643
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

29
12. Dustin, M. L. and L. B. Dustin. 2001. The immunological relay race: B cells take antigen 644
by synapse. Nat. Immunol. 2:480-482. 645
13. Dye, J. R., A. Palvanov, B. Guo, and T. L. Rothstein. 2007. B cell receptor cross-talk: 646
exposure to lipopolysaccharide induces an alternate pathway for B cell receptor-induced 647
ERK phosphorylation and NF-kappa B activation. J. Immunol. 179:229-235. 648
14. Eckl-Dorna, J. and F. D. Batista. 2009. BCR-mediated uptake of antigen linked to TLR9 649
ligand stimulates B-cell proliferation and antigen-specific plasma cell formation. Blood 650
113:3969-3977. 651
15. Edwards, K. M., W. D. Dupont, M. K. Westrich, W. D. Plummer, Jr., P. S. Palmer, 652
and P. F. Wright. 1994. A randomized controlled trial of cold-adapted and inactivated 653
vaccines for the prevention of influenza A disease. The. Journal of infectious. diseases. 654
169:68-76. 655
16. Ellouz, F., A. Adam, R. Ciorbaru, and E. Lederer. 1974. Minimal structural 656
requirements for adjuvant activity of bacterial peptidoglycan derivatives. Biochem. 657
Biophys. Res. Commun. 59:1317-1325. 658
17. Fillatreau, S. and A. Radbruch. 2006. IRF4 - a factor for class switching and antibody 659
secretion. Nat. Immunol. 7:704-706. 660
18. Friedman, R. L., S. P. Manly, M. McMahon, I. M. Kerr, and G. R. Stark. 1984. 661
Transcriptional and posttranscriptional regulation of interferon-induced gene expression in 662
human cells. Cell 38:SP - 745-EP - 755. 663
19. Fritz, J. H., S. E. Girardin, C. Fitting, C. Werts, D. Mengin-Lecreulx, M. Caroff, J. M. 664
Cavaillon, D. J. Philpott, and M. Adib-Conquy. 2005. Synergistic stimulation of human 665
monocytes and dendritic cells by Toll-like receptor 4 and. Eur. J. Immunol. 35:2459-2470. 666
20. Girardin, S. E., I. G. Boneca, J. Viala, M. Chamaillard, A. Labigne, G. Thomas, D. J. 667
Philpott, and P. J. Sansonetti. 2003. Nod2 is a general sensor of peptidoglycan through 668
muramyl dipeptide (MDP) detection. J. Biol. Chem. 278:8869-8872. 669
21. Haller, O., G. Kochs, and F. Weber. 2006. The interferon response circuit: induction and 670
suppression by pathogenic viruses. Virology 344:119-130. 671
22. Han, A., K. Saijo, I. Mecklenbrauker, A. Tarakhovsky, and M. C. Nussenzweig. 2003. 672
Bam32 links the B cell receptor to ERK and JNK and mediates B cell proliferation but not 673
survival. Immunity. 19:621-632. 674
23. Healy, J. I., R. E. Dolmetsch, L. A. Timmerman, J. G. Cyster, M. L. Thomas, G. R. 675
Crabtree, R. S. Lewis, and C. C. Goodnow. 1997. Different nuclear signals are activated 676
by the B cell receptor during positive versus negative signaling. Immunity. 6:419-428. 677
24. Healy, J. I. and C. C. Goodnow. 1998. Positive versus negative signaling by lymphocyte 678
antigen receptors. Annu. Rev. Immunol. 16:645-670. 679
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

30
25. Hill, C. S., J. Wynne, and R. Treisman. 1995. The Rho family GTPases RhoA, Rac1, and 680
CDC42Hs regulate transcriptional activation by SRF. Cell 81:1159-1170. 681
26. Hong, J. X., G. L. Wilson, C. H. Fox, and J. H. Kehrl. 1993. Isolation and 682
characterization of a novel B cell activation gene. Journal of immunology (Baltimore, Md. : 683
1950. ) 150:3895-3904. 684
27. Ishii, K. J. and S. Akira. 2007. Toll or toll-free adjuvant path toward the optimal vaccine 685
development. J. Clin. Immunol. 27:363-371. 686
28. Jefferson, T., D. Rivetti, A. Rivetti, M. Rudin, C. Di Pietrantonj, and V. Demicheli. 687
2005. Efficacy and effectiveness of influenza vaccines in elderly people: a systematic 688
review. Lancet 366:1165-1174. 689
29. Jegerlehner, A., P. Maurer, J. Bessa, H. J. Hinton, M. Kopf, and M. F. Bachmann. 690
2007. TLR9 signaling in B cells determines class switch recombination to IgG2a. Journal 691
of immunology (Baltimore, Md. : 1950. ) 178:2415-2420. 692
30. Ji, W., W. Zhou, K. Gregg, N. Yu, S. Davis, and S. Davis. 2004. A method for cross-693
species gene expression analysis with high-density oligonucleotide arrays. Nucleic Acids 694
Res. 32:e93. 695
31. Jin, M. S. and J. O. Lee. 2008. Structures of the toll-like receptor family and its ligand 696
complexes. Immunity. 29:182-191. 697
32. Klein, U., S. Casola, G. Cattoretti, Q. Shen, M. Lia, T. Mo, T. Ludwig, K. Rajewsky, 698
and R. Dalla-Favera. 2006. Transcription factor IRF4 controls plasma cell differentiation 699
and class-switch recombination. Nature immunology 7:773-782. 700
33. Krieg, A. M. 2002. CpG motifs in bacterial DNA and their immune effects. Annual. 701
Review of Immunology 20:709-760. 702
34. Krieg, A. M., A. K. Yi, S. Matson, T. J. Waldschmidt, G. A. Bishop, R. Teasdale, G. A. 703
Koretzky, and D. M. Klinman. 1995. CpG motifs in bacterial DNA trigger direct B-cell 704
activation. Nature 374:546-549. 705
35. Kubo, M., T. Hanada, and A. Yoshimura. 2003. Suppressors of cytokine signaling and 706
immunity. Nature immunology 4:1169-1176. 707
36. Kufer, T. A. 2008. Signal transduction pathways used by NLR-type innate immune 708
receptors. Mol. Biosyst. 4:380-386. 709
37. Le Bon, A., G. Schiavoni, G. D'Agostino, I. Gresser, F. Belardelli, and D. F. Tough. 710
2001. Type i interferons potently enhance humoral immunity and can promote isotype 711
switching by stimulating dendritic cells in vivo. Immunity. 14:461-470. 712
38. Le Bon, A. and D. F. Tough. 2002. Links between innate and adaptive immunity via type 713
I interferon. Current. opinion. in immunology 14:432-436. 714
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

31
39. Lin, L., A. J. Gerth, and S. L. Peng. 2004. CpG DNA redirects class-switching towards 715
"Th1-like" Ig isotype production via TLR9 and MyD88. Eur. J. Immunol. 34:1483-1487. 716
40. Lohoff, M., H. W. Mittrucker, S. Prechtl, S. Bischof, F. Sommer, S. Kock, D. A. 717
Ferrick, G. S. Duncan, A. Gessner, and T. W. Mak. 2002. Dysregulated T helper cell 718
differentiation in the absence of interferon regulatory factor 4. Proceedings. of the. 719
National. Academy. of Sciences. of the. United. States. of America. 99:11808-11812. 720
41. Lu, R. 2008. Interferon regulatory factor 4 and 8 in B-cell development. Trends in 721
immunology 29:487-492. 722
42. Maher, J. A. and J. DeStefano. 2004. The ferret: an animal model to study influenza 723
virus. Lab animal. 33:50-53. 724
43. Marshall, A. J., H. Niiro, C. G. Lerner, T. J. Yun, S. Thomas, C. M. Disteche, and E. 725
A. Clark. 2000. A novel B lymphocyte-associated adaptor protein, Bam32, regulates 726
antigen receptor signaling downstream of phosphatidylinositol 3-kinase. J. Exp. Med. 727
191:1319-1332. 728
44. Mellman, I. and R. M. Steinman. 2001. Dendritic cells: specialized and regulated antigen 729
processing machines. Cell 106:255-258. 730
45. Minden, A., A. Lin, F. X. Claret, A. Abo, and M. Karin. 1995. Selective activation of 731
the JNK signaling cascade and c-Jun transcriptional activity by the small GTPases Rac and 732
Cdc42Hs. Cell 81:1147-1157. 733
46. Minguet, S., E. P. Dopfer, C. Pollmer, M. A. Freudenberg, C. Galanos, M. Reth, M. 734
Huber, and W. W. Schamel. 2008. Enhanced B-cell activation mediated by TLR4 and 735
BCR crosstalk. Eur. J. Immunol. 38:2475-2487. 736
47. Munster, V. J., E. de Wit, J. M. van den Brand, S. Herfst, E. J. Schrauwen, T. M. 737
Bestebroer, d. van, V, C. A. Boucher, M. Koopmans, G. F. Rimmelzwaan, T. Kuiken, 738 A. D. Osterhaus, and R. A. Fouchier. 2009. Pathogenesis and transmission of swine-739
origin 2009 A(H1N1) influenza virus in ferrets. Science 325:481-483. 740
48. Nelson, N., M. S. Marks, P. H. Driggers, and K. Ozato. 1993. Interferon consensus 741
sequence-binding protein, a member of the interferon regulatory factor family, suppresses 742
interferon-induced gene transcription. Molecular. and cellular. biology. 13:SP - 588-EP - 743
599. 744
49. Nieto-Diaz, M., W. Pita-Thomas, and M. Nieto-Sampedro. 2007. Cross-species analysis 745
of gene expression in non-model mammals: reproducibility of hybridization on high 746
density oligonucleotide microarrays. BMC. Genomics 8:89. 747
50. O'Hagan, D. T., M. Singh, J. Kazzaz, M. Ugozzoli, M. Briones, J. Donnelly, and G. 748
Ott. 2002. Synergistic adjuvant activity of immunostimulatory DNA and oil/water 749
emulsions for immunization with HIV p55 gag antigen. Vaccine 20:3389-3398. 750
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

32
51. Park, S. R., H. Zan, Z. Pal, J. Zhang, A. Al Qahtani, E. J. Pone, Z. Xu, T. Mai, and P. 751
Casali. 2009. HoxC4 binds to the promoter of the cytidine deaminase AID gene to induce 752
AID expression, class-switch DNA recombination and somatic hypermutation. Nat. 753
Immunol. 10:540-550. 754
52. Peng, S. L. 2005. Signaling in B cells via Toll-like receptors. Curr. Opin. Immunol. 755
17:230-236. 756
53. Proietti, E., L. Bracci, S. Puzelli, T. Di Pucchio, P. Sestili, E. De Vincenzi, M. Venditti, 757
I. Capone, I. Seif, E. De Maeyer, D. Tough, I. Donatelli, and F. Belardelli. 2002. Type I 758
IFN as a natural adjuvant for a protective immune response: lessons from the influenza 759
vaccine model. Journal of immunology (Baltimore, Md. : 1950. ) 169:375-383. 760
54. Qian, C., H. An, Y. Yu, S. Liu, and X. Cao. 2007. TLR agonists induce regulatory 761
dendritic cells to recruit Th1 cells via preferential IP-10 secretion and inhibit Th1 762
proliferation. Blood 109:3308-3315. 763
55. Reith, W., S. LeibundGut-Landmann, and J. M. Waldburger. 2005. Regulation of 764
MHC class II gene expression by the class II transactivator. Nat. Rev. Immunol. 5:793-806. 765
56. Reuman, P. D., S. Keely, and G. M. Schiff. 1989. Assessment of signs of influenza illness 766
in the ferret model. J. Virol. Methods 24:27-34. 767
57. Roman, M., E. Martin-Orozco, J. S. Goodman, M. D. Nguyen, Y. Sato, A. Ronaghy, 768
R. S. Kornbluth, D. D. Richman, D. A. Carson, and E. Raz. 1997. Immunostimulatory 769
DNA sequences function as T helper-1-promoting adjuvants. Nature medicine 3:849-854. 770
58. Rowe, T., R. A. Abernathy, J. Hu-Primmer, W. W. Thompson, X. Lu, W. Lim, K. 771
Fukuda, N. J. Cox, and J. M. Katz. 1999. Detection of antibody to avian influenza A 772
(H5N1) virus in human serum by using a combination of serologic assays. Journal of 773
clinical. microbiology 37:937-943. 774
59. Rowe, T., A. J. Leon, C. J. Crevar, D. M. Carter, L. Xu, L. Ran, Y. Fang, C. M. 775
Cameron, M. J. Cameron, D. Banner, D. C. Ng, R. Ran, H. K. Weirback, C. A. Wiley, 776 D. J. Kelvin, and T. M. Ross. 2010. Modeling host responses in ferrets during 777
A/California/07/2009 influenza infection. Virology. 778
60. Saeed, A. I., V. Sharov, J. White, J. Li, W. Liang, N. Bhagabati, J. Braisted, M. Klapa, 779
T. Currier, M. Thiagarajan, A. Sturn, M. Snuffin, A. Rezantsev, D. Popov, A. Ryltsov, 780 E. Kostukovich, I. Borisovsky, Z. Liu, A. Vinsavich, V. Trush, and J. Quackenbush. 781
2003. TM4: a free, open-source system for microarray data management and analysis. 782
BioTechniques 34:374-378. 783
61. Sanjo, H., M. Hikida, Y. Aiba, Y. Mori, N. Hatano, M. Ogata, and T. Kurosaki. 2007. 784
Extracellular signal-regulated protein kinase 2 is required for efficient generation of B cells 785
bearing antigen-specific immunoglobulin G. Mol. Cell Biol. 27:1236-1246. 786
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

33
62. Seki, Y., H. Inoue, N. Nagata, K. Hayashi, S. Fukuyama, K. Matsumoto, O. Komine, 787
S. Hamano, K. Himeno, K. Inagaki-Ohara, N. Cacalano, A. O'Garra, T. Oshida, H. 788 Saito, J. A. Johnston, A. Yoshimura, and M. Kubo. 2003. SOCS-3 regulates onset and 789
maintenance of T(H)2-mediated allergic responses. Nature medicine 9:1047-1054. 790
63. Sleeman, M. A., J. K. Fraser, J. G. Murison, S. L. Kelly, R. L. Prestidge, D. J. Palmer, 791
J. D. Watson, and K. D. Kumble. 2000. B cell- and monocyte-activating chemokine 792
(BMAC), a novel non-ELR alpha-chemokine. International. immunology 12:677-689. 793
64. Tada, H., S. Aiba, K. Shibata, T. Ohteki, and H. Takada. 2005. Synergistic effect of 794
Nod1 and Nod2 agonists with toll-like receptor agonists on human dendritic cells to 795
generate interleukin-12 and T helper type 1 cells. Infect. Immun. 73:7967-7976. 796
65. Takaoka, A. and H. Yanai. 2006. Interferon signalling network in innate defence. Cell 797
Microbiol. 8:907-922. 798
66. Tangye, S. G., K. E. Nichols, N. J. Hare, and B. C. van de Weerdt. 2003. Functional 799
requirements for interactions between CD84 and Src homology 2 domain-containing 800
proteins and their contribution to human T cell activation. J. Immunol. 171:2485-2495. 801
67. Tovey, M. G., C. Lallemand, J. F. Meritet, and C. Maury. 2006. Adjuvant activity of 802
interferon alpha: mechanism(s) of action. Vaccine 24 Suppl 2:S2-46-7. 803
68. Wang, X., T. Uto, T. Akagi, M. Akashi, and M. Baba. 2008. Poly(gamma-glutamic acid) 804
nanoparticles as an efficient antigen delivery and adjuvant system: potential for an AIDS 805
vaccine. J. Med. Virol. 80:11-19. 806
69. Yamagata, T., J. Nishida, S. Tanaka, R. Sakai, K. Mitani, M. Yoshida, T. Taniguchi, 807
Y. Yazaki, and H. Hirai. 1996. A novel interferon regulatory factor family transcription 808
factor, ICSAT/Pip/LSIRF, that negatively regulates the activity of interferon-regulated 809
genes. Molecular. and cellular. biology. 16:1283-1294. 810
70. Yin, Q., X. Wang, J. McBride, C. Fewell, and E. Flemington. 2008. B-cell receptor 811
activation induces BIC/miR-155 expression through a conserved AP-1 element. The. 812
Journal of biological. chemistry. 283:2654-2662. 813
814
815
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

34
Figure 1. CpG ODN-assisted vaccination increased influenza virus-specific antibody levels 816
in serum from immunized ferrets. 817
Influenza virus-specific antibody levels in serum from immunized ferrets were assessed by (A) 818
ELISA, (B) HA inhibition and (C) microneutralization assays. (A) Serum IgM (left panel) and 819
IgG (right panel) antibody levels against the commercial vaccine FLUVIRAL were measured at 820
Days 0, 14, 21, 28, 35 and Day 7 post boost. The average relative absorbance density read at 450 821
nm from three individual samples were plotted graphically. (B) HI titers were measured from 822
ferret sera against inactivated 2007-2008 seasonal Solomon Island A/Solomon Islands/3 H1N1, 823
A/Wisconsin/67/2005 H3N2 and B/Malaysia/2507/2004 viruses. (C) Neutralizing antibody titers 824
for blocking the live A/Solomon Islands/3 H1N1 virus were measure by microneutralization 825
assays. Three independent experiments were performed. Error bars indicate standard deviation. 826
Statistical analysis between adjuvant added vaccine and vaccine alone treated group animals 827
were performed. *: p ≤ 0.05, **: p ≤ 0.01, ***: p ≤ 0.001. 828
Figure 2. CpG ODN vaccine activated ISGs expression early post immunization. 829
mRNA expression profiles from immunized ferrets with and without CpG ODN were determined 830
by Q-PCR. (A) The heat map represents the mRNA expression profile of 50 ferret immune-831
related genes in V2007
+ CpG (V + CpG) and V2007
-Alone (V-Alone) group animals (n = 3/group) 832
at Day 1 post vaccination. The heat map was generated by Multiexperiment Viewer software 833
version 4.1 from the β-actin normalized real-time PCR data relative to PBS control group. Genes 834
were listed by descending mRNA-level (red, upregulation; green, downregulation). V represents 835
the V2007
. (B) Transcription of nine ISG genes (OAS1, RIG-I, Mx-1, CXCL10/IP10, ISG15, 836
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

35
IRF1, IRF7, STAT1 and STAT2) in ferret whole blood was quantified by Q-PCR and displayed 837
graphically. The PBS V2007
boost group animals were treated with PBS at Day 0 and boosted 838
with commercial vaccine at Day 35 after injection. The RNA samples used for expression 839
analysis were extracted from the peripheral blood of three ferrets (n = 3) in each group at every 840
time point. mRNA levels were normalized to β-actin and then to PBS control groups. The data 841
was averaged from three independent experiments and the error bars represent standard 842
deviation. Horizontal bars indicate the statistical analysis performed between the selected two 843
groups. *: p ≤ 0.05, **: p ≤ 0.01. 844
Figure 3. The levels of ISG regulatory genes in CpG ODN adjuvant-mediated 845
immunization. 846
The average transcription levels of (A) IFN-α and IFN-γ and (B) IRF4 in CpG ODN mediated 847
immunization were determined by Q-PCR and plotted graphically from various time points 848
following immunization. Increases in mRNA levels were relative to β-actin and then normalized 849
to PBS control groups. Average was obtained from three independent experiments and the error 850
bars indicate standard deviation. Horizontal bars show the statistical analysis performed between 851
the selected two groups. *: p ≤ 0.05. 852
Figure 4. IFN-α and CFA-mediated vaccination induced influenza virus-specific antibody 853
production. 854
(A) Antibody titers in vaccinated ferret serum against 2008-2009 seasonal live 855
A/Brisbane/59/2007 H1N1 virus and A/Brisbane/10/2007 H3N2 virus were measured by HI 856
inhibition assays. (B) Neutralizing antibody titers for blocking the live A/Brisbane/59/2007 857
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

36
H1N1 virus were measured from immunized ferrets by microneutralization assays. Data 858
represents triplicate measurements of sera collected from three animals. Error bars indicate 859
standard deviation. Statistical analysis between adjuvant added vaccine and vaccine alone treated 860
group animals were performed by either one-way ANOVA in figure (A) or student t test in figure 861
(B). *: p ≤ 0.05, **: p ≤ 0.01, ***: p ≤ 0.001. 862
Figure 5. Adjuvant-mediated vaccination decreased viral load following influenza 863
infection. 864
Viral load in nasal wash at Day 3 post H1N1 influenza challenge was determined. Nasal wash 865
from each group animals (n = 3) was collected post infection, cultured and titrated. The virus 866
titer was calculated by Reed-Muench method and expressed as TCID50/ml. Error bars 867
demonstrate standard error of the mean. Statistical analysis was performed between each 868
adjuvant added vaccination group and PBS control group by student t test. V represents the 869
V2008
. *: p ≤ 0.05, **: p ≤ 0.01. 870
Figure 6. Memory antibody response at Day 5 post H1N1 infection was increased in 871
adjuvant-mediated vaccinated animals. 872
(A) Antibody titers from ferret sera against live seasonal A/Brisbane/59/2007 H1N1 virus was 873
determined by HA inhibition assays at Day 5 post infection for adjuvant and non-adjuvant 874
assisted vaccinations. (B) Neutralizing antibody titers for blocking the A/Brisbane/59/2007 875
H1N1 virus were assessed by microneutralization assays. The statistical analysis determined 876
between Day 35 after vaccination and Day 5 post infection is shown by horizontal bar. The 877
differences between adjuvant groups and vaccine alone group at Day 5 p.i. were analyzed by 878
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

37
one-way ANOVA. Data represents the average of triplicate measurements of sera collected from 879
three animals. Error bars indicate standard deviation. V represents the V2008
. *: p ≤ 0.05, **: p ≤ 880
0.01, ***: p ≤ 0.001. 881
Figure 7. Adjuvant-mediated vaccination activated differing arms of innate and adaptive 882
immunity. 883
The heat maps represent RNA expression determined by microarray analysis Day 1 post 884
vaccination. The heat map generated from the gene expression profile of the four different 885
vaccination groups (n = 3/group) shows the innate, adaptive, Ag processing/presentation and 886
complement immune related genes, which were significantly regulated by at least 1.5 fold 887
change (p ≤ 0.05) in one of the adjuvant additive groups. The fold change of the gene expression 888
is shown in table 3 (red, upregulation; blue, downregulation). V represents the V2008
. 889
Figure 8. CpG ODN and PEG-IFN-adjuvanted vaccine-regulated IFN signaling pathways. 890
(A) IFN-responsive gene expression was determined by microarray analysis from the RNA of 891
vaccinated ferrets at Day 1 post vaccination (red, upregulation; blue, downregulation) and plotted 892
by heat map. (B) A schematic was created from the gene expression profiling data obtained Day 893
1 post immunization by IPA: the upper panel shows the IPA canonical pathway of IFN signaling 894
using the Q-PCR of 2007 vaccine study and microarray data of the 2008 vaccine study from 895
CpG-mediated immunized ferrets; the lower panel exhibits the IFN signaling pathway activated 896
by PEG-IFN-adjuvanted vaccination by using the microarray data of the 2008 vaccine study (red, 897
upregulation). (C) Gene expression heat map of molecules involved in Ras signaling at Day 1 898
post-vaccination (upper panel; red, upregulation), and IPA modeling of JAK/STAT pathway-899
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

38
mediated Ras signaling using the microarray data from the whole blood RNA of PEG-IFN-900
mediated immunized ferrets at Day 1 post vaccination (lower panel; red, upregulation). 901
Figure 9. Cell adhesion and B cell signaling pathways were activated in ferrets immunized 902
with CFA-mediated vaccines. 903
(A) Heat map of cell adhesion molecule gene expression and genes involved in B cell 904
activation/calcineurin-NFAT signaling at Day 1 post vaccination (red, upregulation; blue, 905
downregulation). (B), (C) and (D) IPA canonical pathways of integrin signaling, B cell 906
activation, and calcineurin-NFAT created by using the microarray data of CFA immunized ferret 907
peripheral blood RNA Day 1 post vaccination, (red, upregulation; green, downregulation). V 908
represents the V2008
. 909
Figure S1. The level of ferret_DLA-64-like, ferret_FCN1-like and ferret_SOCS3-like 910
expression as quantified by Q-PCR in adjuvant-mediated vaccination. 911
To confirm the gene changes from the microarray analysis, the average transcription levels of 912
three selected genes (A) Ferret_DLA-64-like, (B) Ferret_FCN1-like and (C) Ferret_SOCS3-like 913
in multiple vaccination groups (n=3/group) were determined by Q-PCR at day 1 following 914
immunization. Increases in mRNA levels were relative to β-actin and then normalized to PBS 915
control groups. Average values were obtained from three independent experiments and the error 916
bars indicate standard deviation. The asterisk showed the statistical difference found between the 917
adjuvanted vaccine group and vaccine alone group. *: p ≤ 0.05. 918
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

Table 1. Ferret Specific Gene Primer list
Primer NameATCC Item
Number
NCBI Gene
Accesion NumberPrimer 5'-3' Sequence
Caspase 4 F NR-8075 EU836041 TCATTGCTTTCTGCTCCTCA
Caspase 4 R NR-8076 EU836041 ATCTGGGCTTTCACATCTGG
C1 Inhibitor F NR-8079 EU835994 GCCTCTCAGAGCCTGTATGG
C1 Inhibitor R NR-8080 EU835994 CTTCCACTTGGCACTCAGGT
Hsp40 F NR-8081 EU836010 CCACCGTGTCGAGCTTTATT
Hsp40 R NR-8082 EU836010 CATGAACGTTGGACTGGTTG
NFAT5 F NR-8083 EU836021 ACCTCTTCCAGCCCTACCAT
NFAT5 R NR-8084 EU836021 CCTCTTCGGTGTTGATGGAT
SOCS5 F NR-8087 EU835993 GTTAGCTCCCGGAATGACTG
SOCS5 R NR-8088 EU835993 CTTTCCAAGCTCCCTGTCTG
IRF2 F NR-8091 EU835486 AGGTGACCACCGAGAGTGAC
IRF2 R NR-8092 EU835486 CCCCATGTTGCTGAGGTACT
CD3 F NR-8097 EF492054 GGCGGTGGCTGCAATC
CD3 R NR-8098 EF492054 TCCAGTAATAGACCAGCAGAAGCA
CD14 F NR-8103 TBA AGTGCCATCGAGGTGGAGAT
CD14 R NR-8104 TBA GCGTACTGCTTCGGGTCTGT
CRLF2 F NR-8109 EU836018 GGCTTGGATGCTGAGAAATGTT
CRLF2 R NR-8110 EU836018 GGGCCATAGCTGGACTCCAT
HLA-DRA_like F NR-8115 EU835995 GAGAGCCCAACATCCTCATCTG
HLA-DRA_like R NR-8116 EU835995 TCGAAGCCACGTGACATTGA
STAT3 F NR-8121 EU835482 CAACCCCAAGAACGTGAACT
STAT3 R NR-8122 EU835482 AGCCCACGTAATCTGACACC
TLR2 F NR-8123 EU836009 TCTTCTGGAGCCCATTGAGAA
TLR2 R NR-8124 EU836009 GTGTTCATTATCTTCCGCAGCTT
NFKBIA F NR-8125 EU835992 CCAGCACCTCTACTCCATCC
NFKBIA R NR-8126 EU835992 CATCAGCACCCAAAGACACC
Sprouty 4 F NR-8127 EU836020 CAGCGGCTCTTGGACCAC
Sprouty 4 R NR-8128 EU836020 ACTTACACTTCCCACAGG
IFN-alpha F NR-8133 EF492061 TCTCCATGTGACAAACCAGAAGA
IFN-alpha R NR-8134 EF492061 CAGAAAGTCCTGAGCACAATTCC
IFN-gamma F NR-8137 EF492064 TCAAAGTGATGAATGATCTCTCACC
IFN-gamma R NR-8138 EF492064 GCCGGGAAACACACTGTGAC
IFI35 F NR-8141 EU835487 GGGCTCCGGCTGAGTGA
IFI35 R NR-8142 EU835487 CCACCCCCATTTCTGGTCTT
P52rIPK_like F NR-8143 EU835488 TACGATGCCGAACTTCTG
P52rIPK_like R NR-8144 EU835488 TGCTCTCCTACAATTCTCC
Mx1 F NR-8147 EU835483 ACATCCTCAGGCAGGAGACA
Mx1 R NR-8148 EU835483 CAGGTCAGGCTTTGTCAAGA
IL-1beta F NR-8149 EU835490 GGACTGCAAATTCCAGGACATAA
IL-1beta R NR-8150 EU835490 TTGGTTCACACTAGTTCCGTTGA
IL-4 F NR-8151 EF492062 TCACCGGCACTTTCATCCA
IL-4 R NR-8152 EF492062 TTCTCGCTGTGAGGATGTTCA
IL-6 F NR-8153 EF492063 AGTGGCTGAAACACGTAACAATTC
IL-6 R NR-8154 EF492063 ATGGCCCTCAGGCTGAACT
IL-8 F NR-8155 EU477256 AAGCAGGAAAACTGCCAAGAGA
IL-8 R NR-8156 EU477256 GCCAGAAGAAACCTGACCAAAG
IL-16 F NR-8157 EU835491 CCCACTCGGACCTTCT
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

IL-16 R NR-8158 EU835491 GAGAGGAGCCAAAGGTCTCAA
NKG7 F NR-8159 EU836023 GCGAGCGGTGGAACCA
NKG7 R NR-8160 EU836023 CAGGTAGAACGACCAGGAGAAGA
CXCL10/IP10 F NR-8161 EF492058 CTTTGAACCAAAGTGCTGTTCTTATC
CXCL10/IP10 R NR-8162 EF492058 AGCGTGTAGTTCTAGAGAGAGGTACTC
TNF-alpha F NR-8167 EF492065 CCAGATGGCCTCCAACTAATCA
TNF-alpha R NR-8168 EF492065 GGCTTGTCACTTGGAGTTCGA
CXCR3 F NR-8169 EF492060 TTTGACCGCTACCTGAGCAT
CXCR3 R NR-8170 EF492060 GCCGACAGGAAGATGAAGTC
Beta-actin F NR-8171 EU836011 TGACCGGATGCAGAAGGA
Beta-actin R NR-8172 EU836011 CCGATCCACACCGAGTACTT
CCL5 F NR-8173 EU835495 GCTGCTTTGCCTACATTTCC
CCL5 R NR-8174 EU835495 CCCATTTCTTCTGTGGGTTG
NF-kB F NR-8177 EU835494 AGGATCGATCAAAGCCTGAA
NF-kB R NR-8178 EU835494 CCCTCACCAGGTAACAGAGC
TLR3 F NR-8179 EU835984 GATGACCTCCCAGCAAACAT
TLR3 R NR-8180 EU835984 GCACAATTCTGGCTCCAGTT
MyD88 F NR-8181 EU836043 GCGTTTTGATGCCTTCATCT
MyD88 R NR-8182 EU836043 GGCAAGACATCACGATCAGA
TRAF6 F NR-8183 EU836040 AGATTGGCAACTTTGGGATG
TRAF6 R NR-8184 EU836040 ACAGTTTGTAGCCGGGTTTG
IRAK4 F NR-8185 EU836022 CCCCTGCAAGTCTTTTGCTA
IRAK4 R NR-8186 EU836022 GATGTCCTGTCTTTGCCACA
OAS1 F NR-8187 EU835484 TGAAGAGCCTCCTACGGCTA
OAS1 R NR-8188 EU835484 CCATCTCCCAGGCGTAGATA
TLR4 F NR-8191 EU835996 AGTGGGTCAAAGACCACAGG
TLR4 R NR-8192 EU835996 GACAGGGTGGCATTCCTAAA
IRF7 F NR-8195 EU835985 ACACTCTACCCCCGTGTCTG
IRF7 R NR-8196 EU835985 GTCAAGTCCAGAGCCTCCAA
ISG20 F NR-8197 TBA GGTGGCCATTGACTGTGAG
ISG20 R NR-8198 TBA GCCGGATGAACTTGTCGTAG
TLR9 F NR-8201 TBA ACTCCGACTTTGTCCACCTG
TLR9 R NR-8202 TBA GGTCATGTGACAGGGGAAGT
CASP8 F NR-9500 EU836044 ATTGCCAATGTCGGACTCTC
CASP8 R NR-9501 EU836044 TACTCAACGCATCTGCATCC
ICAM1 F NR-9508 EU836045 TGGACTACGGTGACTGTGGA
ICAM1 R NR-9509 EU836045 CGGACAATCCCTCTGGTCTA
IL8RB F NR-9512 EU836026 CGCTCCTGAAGGAAGTCAAC
IL8RB R NR-9513 EU836026 AGCAGACTGGGCTGGAGTAG
IRF1 F NR-9514 EU835485 CGATACAAAGCAGGGGAAAA
IRF1 R NR-9515 EU835485 GGCCTTGCACTTAGCATCTC
IRF4 F NR-9516 EU836039 AATCCTCGTGAAGGAGCTGA
IRF4 R NR-9517 EU836039 AGATCCTGCTCTGGCACAGT
ISG15 F NR-9518 EU835986 AGCAGCAGATAGCCCTGAAA
ISG15 R NR-9519 EU835986 CAGTTCTTCACCACCAGCAG
PKR F NR-9522 EU835989 ACGAATACGGCATGAAGACC
PKR R NR-9523 EU835989 TGGAAGGGTCAGGCATTAAG
RIG-I F NR-9524 EU836024 AGAGCACTTGTGGACGCTTT
RIG-I R NR-9525 EU836024 TGCAATGTCAATGCCTTCAT
SOCS3 F NR-9528 EU835987 GCTGGTGCATCACTACATGC
SOCS3 R NR-9529 EU835987 GACCGTCTTCCGACAGAGAT
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

STAT2 F NR-9530 EU835988 AGCTGCTGAAGGAGCTGAAG
STAT2 R NR-9531 EU835988 TGCCTTCCTGGAGTCTCACT
STAT1 F not available EU835493 AGCCTTGCATGCCAACTCA
STAT1 R not available EU835493 ACAGTCCAGCTTCACCGTGAA
FCN1 F NR-8117 TBA CACCAAGGACCAGGACAATGA
FCN1 R NR-8118 TBA CACCAGGCCCCCTGGTA
DLA-64_like F not available TBA CAGGACACAGAGGTTGTGGA
DLA-64_like R not available TBA TGGCACGTGTATCTCTGCTC
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

Table 2. Inactivity of Infected Ferrets
GROUP
Relative
Inactivity Index Day 1 Day 2 Day 3 Day 4 . Day 5
PBS 1.6 0.0 1.5 0.6 0.5 0.5
V + CFA 1.1 0.0 0.0 0.5 0.0 0.0
V + CpG 1.3 0.0 0.6 0.5 0.3 0.3
V + IFN 1.3 0.0 0.5 0.3 0.4 0.4
V + PBS 2.0 0.5 1.4 1.4 0.9 0.6
Relative inactivity index was calculated from daily activity scores as described in Materials and Methods
Average of daily activity score was obtained from 3 animals in each group
2008-2009 human flu vaccine
Daily Mean Inactivity Score Post Infection
a
b
a
b
C
C
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

Table 3. In vivo differential microarray gene expression analysis on whole blood of ferrets immunized with adjuvanted 2008-2009 vaccines (n=3) versus PBS controls (n=3) on day 1 post vaccination.
Gene Symbol
V + CpG V + IFN V + CFA V + PBS
IFN Responding Genes
STAT1 2.7 2.3 1.1 1.0
IRF1 2.3 1.2 1.5 1.1
IRF2 1.5 2.5 2.3 -1.1
OAS1 3.2 2.3 -1.1 -1.0
OAS2 2.4 1.8 -1.1 -1.0
OASL 1.9 1.1 1.0 1.1
ISG15 7.6 6.9 1.4 -1.0
USP18 4.8 3.2 1.1 -1.0
IFI44 3.4 3.3 1.7 -1.1
IFI44L 3.4 3.3 1.7 -1.0
IFIT2 1.7 1.5 1.4 1.1
RSAD2 5.6 3.5 2.2 1.2
EIF2AK2 (PKR) 1.7 1.3 1.1 -1.1
CXCL10 1.6 1.1 1.0 1.3
SOCS1 2.7 1.2 1.0 1.3
Toll-like Receptor
TLR4 1.5 1.1 1.1 1.0
Antigen Processing and Presentation Gene
CTSB 1.6 1.4 1.1 -1.0
CTSS 1.7 1.4 1.2 -1.0
CTSD 1.1 1.9 1.7 -1.0
MHC class Ib 1.1 1.5 1.3 1.0
MHC class I DLA-64 1.1 1.4 1.9 1.1
HLA-DMA 1.1 1.4 2.0 -1.1
CD74 1.0 2.1 1.9 1.0
RFX5 1.3 1.5 1.6 1.0
CIITA 1.1 1.1 -1.8 -1.0
Adaptive Immunity Regulator
LYN 1.2 1.2 1.5 1.0
SYK 1.0 1.4 1.6 -1.1
PIK3CD 1.2 1.6 1.8 -1.1
RAC1 1.2 1.2 2.3 1.1
BAM32 (DAPP1) 1.3 1.1 1.5 -1.1
HRAS 1.3 2.0 2.5 1.0
RAF1 -1.1 1.5 1.8 1.0
MAP2K2 1.0 1.6 1.5 1.1
MAPK1 1.0 1.0 1.6 1.0
NFATC3 1.1 1.2 2.0 -1.2
AKT3 -1.1 -1.1 -1.7 -1.4
Mean of Fold Changea b
C
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

POU2F2 1.1 1.3 1.6 -1.2
HoxC4 1.1 -1.1 -1.7 -1.1
AICDA (AID) 1.0 1.1 2.6 1.0
JUNB 1.3 2.3 2.8 1.2
RGS1 2.3 1.6 1.0 1.2
GNA15 1.3 2.0 2.1 1.2
GNG2 1.1 1.4 1.6 1.1
GNG11 1.0 1.2 1.6 1.1
GNB3 1.1 1.4 1.8 -1.1
Calcineurin (PPP3R1) 1.0 1.2 1.7 -1.0
RCAN1 1.1 -1.1 -2.3 1.0
CSNK1D 1.0 1.1 1.9 1.1
CSNK1G2 1.1 1.1 1.5 -1.1
MEF2C -1.1 -1.2 1.9 -1.2
LAT -1.1 1.1 -1.5 -1.0
IL12A -1.1 -1.4 -1.8 -1.1
SOCS3 1.2 1.2 1.6 -1.0
CXCL14 1.3 1.4 1.6 1.1
Complement
C3 precursor 2.4 2.4 1.1 -1.0
FCN1 2.6 1.9 1.1 -1.0
C1QL2 1.2 1.1 1.5 -1.0
C1R 1.1 1.5 3.9 1.1
C5AR1 1.3 1.3 1.6 -1.0
CFI 1.0 1.0 1.7 -1.0
Cell Adhesion Molecule and Regulator
VCAM1 -1.1 1.3 1.9 -1.1
BCAM 1.1 1.3 2.0 -1.0
CD36 1.1 1.2 1.6 1.0
CD44 1.1 1.1 1.5 1.0
CD84 1.4 1.6 1.7 1.2
Integrin-beta6 1.1 1.1 1.6 1.0
ZYXIN 1.2 1.7 2.6 1.2
TALIN 1 1.1 1.0 1.5 -1.1
RHOG 1.2 1.4 1.9 -1.1
RHOT1 1.0 -1.1 1.8 -1.1
CRKL 1.0 1.0 1.5 -1.1
RAP2A 1.0 1.2 1.9 1.1
The gene symbol is compatible to the human ortholog
The mean fold change is normalized to the corresponding PBS control group. Boldface indicates
the gene expression is significanltly induced by at least 1.5 fold change (p г 0.05) versus controls.
2008-2009 human flu vaccine
b
C
a
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

Ferret Antigen Sepcific IgM
Days
Day0 Day14 Day21 Day28 Day35Day7 Post Boost
OD
45
0
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
**
PBS
V2007
-Alone
V2007
+CpG
H1N1 HA Inhibition Test
Days
Day0 Day14 Day21 Day28 Day35 Day7 Post Boost
HI
Titer
(Lo
g2)
4
6
8
10
12
** ** *****
**PBS
PBS V2007
Boost
V2007
-Alone
V2007
+CpG
Influenza B Strain HA Inhibition Test
Days
Day0 Day14 Day21 Day28 Day35 Day7 Post Boost
HI T
ite
r (L
og
2)
4
6
8
10
12
** * ***
**
Microneutralization Assay on Solomon Islands H1N1 Virus
Days
Day0 Day14 Day21 Day28 Day35 Day7 Post Boost
Ne
utr
aliz
ing
Antibo
dy T
ite
r (L
og
2)
8
10
12
14
16 PBS
PBS V2007
Boost
V2007
-Alone
V2007
+CpG
** **
***
**
Ferret Antigen Specific IgG
Days
Day0 Day14 Day21 Day28 Day35Day7 Post Boost
OD
45
00.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
1.8
2.0
**
****
**
Fig.1
A
H3N2 HA Inhibition Test
Days
Day0 Day14 Day21 Day28 Day35 Day7 Post Boost
HI
Titer
(Lo
g2)
4
6
8
10
12
*** ******
***
***
B
C
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

Fig.2
RIG-I Q-PCR Assay
Days
Day1 Day3 Day5 Day7 Post Boost
Norm
aliz
ed to B
-actin
0
2
4
6
8
* **
* *
Mx-1 Q-PCR Assay
Days
Day1 Day3 Day5 Day7 Post Boost
Norm
aliz
ed to B
-actin
0
1
2
3
4
*
* *
CXCL10 Q-PCR Assay
Days
Day1 Day3 Day5 Day7 Post Boost
Norm
aliz
ed to B
-actin
0
1
2
3
4
5
6
7
*
**
ISG15 Q-PCR Assay
Days
Day1 Day3 Day5 Day7 Post Boost
Norm
aliz
ed t
o B
-actin
0
1
2
3
4
5
6
** **
IRF1 Q-PCR Assay
Days
Day1 Day3 Day5 Day7 Post Boost
No
rmaliz
ed
to B
-actin
0.0
0.5
1.0
1.5
2.0
2.5
*
IRF7 Q-PCR Assay
Days
Day1 Day3 Day5 Day7 Post Boost
Norm
aliz
ed to B
-actin
0.0
0.5
1.0
1.5
2.0
2.5
*** *
STAT1 Q-PCR Assay
Days
Day1 Day3 Day5 Day7 Post Boost
Norm
aliz
ed
to
B-a
ctin
0.0
0.5
1.0
1.5
2.0
2.5
** *
STAT2 Q-PCR Assay
Days
Day1 Day3 Day5 Day7 Post Boost
Norm
aliz
ed
to
B-a
ctin
0.0
0.5
1.0
1.5
2.0
2.5
3.0
* ***
A BOAS1 Q-PCR Assay
Days
Day1 Day3 Day5 Day7 Post Boost
No
rma
lize
d to
B-a
ctin
0
1
2
3
4
5
**
***
*PBS V
2007 Boost
V2007
-Alone
V2007
+CpG
V-Alone V+CpG
-3 30
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
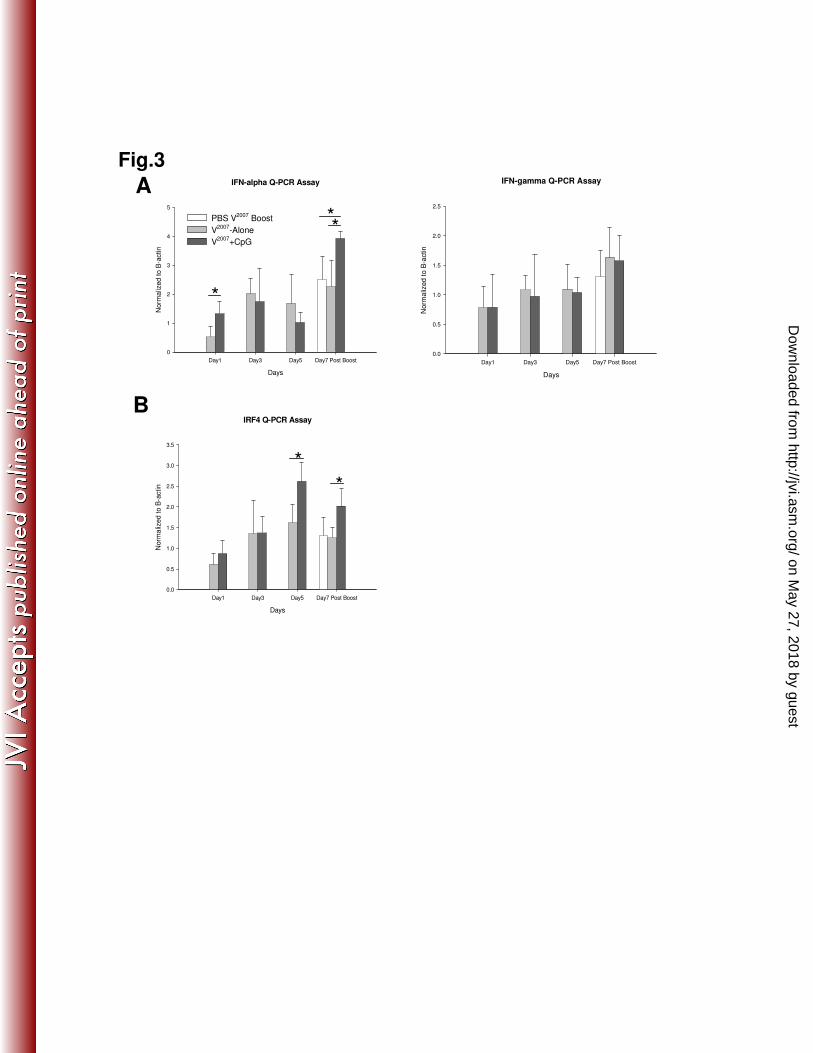
IFN-gamma Q-PCR Assay
Days
Day1 Day3 Day5 Day7 Post Boost
Norm
aliz
ed
to B
-actin
0.0
0.5
1.0
1.5
2.0
2.5
Fig.3IFN-alpha Q-PCR Assay
Days
Day1 Day3 Day5 Day7 Post Boost
Norm
aliz
ed
to
B-a
ctin
0
1
2
3
4
5
*
**PBS V
2007 Boost
V2007
-Alone
V2007
+CpG
IRF4 Q-PCR Assay
Days
Day1 Day3 Day5 Day7 Post Boost
Norm
aliz
ed
to
B-a
ctin
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
*
*
A
B
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

Microneutraliztion Assay on Brisbane59 H1N1 Virus
Days
Day0 Day14 Day21 Day28 Day35
Neu
tralizin
g A
nti
bo
dy T
iter
(Lo
g2)
8
10
12
14
16
PBS
V2008
-Alone
V2008
+CpG
V2008
+IFN
V2008
+CFA
*
*
* **
***
Fig.4
A
B
H1N1 HA Inhibition Test
Days
Day0 Day14 Day21 Day28 Day35
HI
Tite
r (L
og
2)
4
6
8
10
12
PBS
V2008
-Alone
V2008
+CpG
V2008
+IFN
V2008
+CFA
***
***
******
H3N2 HA Inhibition Test
Days
Day0 Day14 Day21 Day28 Day35
HI T
iter
(Log
2)
4
6
8
10
12
** *** *** ***
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

Day 5 Post Infection
Vaccination
PBS V-Alone V+CpG V+IFN V+CFA
HI T
iter
(Log2)
Blo
ckin
g H
1N
1 V
irus
3
4
5
6
7
8
9
***
Fig.5
Viral Load in Nasal Wash at Day 3 Post Infection
Vaccination
PBS V-Alone V+CpG V+IFN V+CFA
TC
ID5
0/m
l (L
og
10)
1.2
1.4
1.6
1.8
2.0
2.2
2.4
2.6
2.8
**
*
**
Fig.6
A BMemory Antibody Response to H1N1
Days
Day14 Day21 Day28 Day35 Day5 Post Infection
Ne
utr
aliz
ing
An
tib
od
y T
ite
r (L
og
2)
8
10
12
14
16PBS
V2008
-Alone
V2008
+CpG
V2008
+IFN
V2008
+CFA
**
*
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

Fig.7
-3 30
Innate Immune Defense Gene
V-AloneV+CFAV+IFNV+CpG
ComplementV-AloneV+CFAV+IFNV+CpG
V-AloneV+CFAV+IFNV+CpG
Ag Processing and Presentation
V-AloneV+CFAV+IFNV+CpG
Adaptive Immune Defense Gene
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

Fig.8
B
-3 30
Interferon Stimulated GeneA
V+CpG
V+IFN
V-AloneV+CFAV+IFNV+CpG
-3 30
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

Fig.8
Ras Signaling MoleculeC
-3 30
V-AloneV+CFAV+IFNV+CpG
-3 30
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

Fig.9
Cell Adhesion and Motility B cell ActivationA
B
-3 30
V-AloneV+CFAV+IFNV+CpGV-AloneV+CFAV+IFNV+CpG
-3 30
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

Fig.9
C
D
-3 30
-3 30
on May 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from