modulationof d-serinelevelsviaubiquitin-dependent ... · fax: 972-4-829-5384 , ......
TRANSCRIPT

Modulation of D-Serine Levels via Ubiquitin-dependentProteasomal Degradation of Serine Racemase*
Received for publication, March 1, 2006, and in revised form, May 18, 2006 Published, JBC Papers in Press, May 19, 2006, DOI 10.1074/jbc.M601971200
Elena Dumin‡, Inna Bendikov‡, Veronika N. Foltyn‡, Yoshio Misumi§, Yukio Ikehara¶, Elena Kartvelishvily‡,and Herman Wolosker‡1
From the ‡Department of Biochemistry, The Bruce Rappaport Faculty of Medicine, Technion-Israel Institute of Technology,Haifa 31096, Israel, §Department of Cell Biology, Fukuoka University School of Medicine, Fukuoka 814-0180, Japan,and ¶Institute for Pharmaceutical Sciences, Daiichi University, Fukuoka 815-8511, Japan
Mammalian serine racemase is a brain-enriched enzyme thatconverts L- into D-serine in the nervous system. D-Serine is anendogenous co-agonist at the “glycine site” of N-methyl D-as-partate (NMDA) receptors that is required for the receptor/channel opening. Factors regulating the synthesis of D-serinehave implications for the NMDA receptor transmission, but lit-tle is known on the signals and events affecting serine racemaselevels. We found that serine racemase interacts with the Golginsubfamily A member 3 (Golga3) protein in yeast two-hybridscreening. The interaction was confirmed in vitro with therecombinant proteins in co-transfected HEK293 cells andin vivo by co-immunoprecipitation studies from brain homoge-nates. Golga3 and serine racemase co-localized at the cytosol,perinuclear Golgi region, and neuronal and glial cell processesin primary cultures. Golga3 significantly increased serine race-mase steady-state levels in co-transfectedHEK293 cells and pri-mary astrocyte cultures. This observation led us to investigatemechanisms regulating serine racemase levels. We found thatserine racemase is degraded through the ubiquitin-proteasomalsystem in a Golga3-modulated manner. Golga3 decreased theubiquitylation of serine racemase both in vitro and in vivo andsignificantly increased the protein half-life in pulse-chaseexperiments. Our results suggest that the ubiquitin system is amain regulator of serine racemase and D-serine levels. Modula-tion of serine racemase degradation, such as that promoted byGolga3, provides a newmechanism for regulating brain D-serinelevels and NMDA receptor activity.
N-Methyl D-aspartate (NMDA)2 type of glutamate receptorsplay key roles in excitatory synaptic transmission and areinvolved in many physiological processes including learningandmemory (1).NMDAreceptor activity is tightly regulated, asits overactivation contributes to pathologic conditions such as
stroke and neurodegenerative diseases (2). An interesting fea-ture of NMDA receptors is the requirement of simultaneousbinding of two agonists for channel opening, i.e. the NMDAchannel only operates when both a glutamate site and a co-agonist site are occupied (1). It has been shown that binding ofglycine to the co-agonist site is an obligatory requirement forNMDA receptor/channel operation (3, 4). Subsequent studieshave shown that brain D-serine is an endogenous ligand of theglycine site of NMDA receptors (5–9).Regulation of NMDA receptor activity by the co-agonist
D-serine plays critical roles. Removal of endogenous D-serinedecreases NMDA receptor responses (8) and blocks NMDA-dependent migration of immature granule cells in the cerebel-lum (9). D-Serine is the dominant endogenous co-agonist forNMDA neurotoxicity, as removal of D-serine abolishes NMDAreceptor-elicited cell death in hippocampal slices (6). In thevertebrate retina, endogenous D-serine mediates the light-de-pendent increase in neuronal activity by activating NMDAreceptors (10). Furthermore, endogenous D-serine is requiredfor the long term potentiation of the synaptic transmission inthe hippocampus, which is thought to be involved in the mem-ory formation (11).Brain D-serine is synthesized by the serine racemase enzyme,
which converts L- into D-serine (12–14). The enzyme occurs inglia (13) and more recently was demonstrated in neurons (15).For its activity, serine racemase requires pyridoxal 5�-phos-phate, ATP, and magnesium as cofactors (16). In addition toconverting L- to D-serine, serine racemase catalyzes the �,�-elimination ofwater from L-serine to formpyruvate and ammo-nia (16). This activity was also observed toward D-serine, whichseems to limit the achievable D-serine concentration and mayplay a role in D-serine metabolism as well (17).
Despite the important roles proposed for the endogenousD-serine, little is known about the regulation of its synthesis byserine racemase. Factors altering serine racemase levels willhave an impact on NMDA receptor activity, since the D-serinebinding site of the receptor is not saturated under normal con-ditions (1) and serine racemase is key for D-serine homeostasis(5, 13). In the present study we sought to look for proteins thatbind and regulate serine racemase using the yeast two-hybridtechnique. We discovered that Golga3, also known as GCP170(18) or golgin-160 (19, 20), strongly binds to serine racemaseboth in vitro and in vivo. We found that serine racemase isdegraded by the ubiquitin-proteasome system and that Golga3greatly increases serine racemase levels by decreasing its ubiq-
* This work was supported by grants from the Israel Science Foundation,Rappaport Institute for Research in the Medical Sciences, Atkins MedicalResearch Fund for Gerontology, and the public committee for the desig-nation of estate funds from the Ministry of Justice (to H. W.). The costs ofpublication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked “advertisement” inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1 To whom correspondence should be addressed. Fax: 972-4-829-5384,E-mail: [email protected].
2 The abbreviations used are: NMDA, N-methyl-D-aspartate; GFP, green fluo-rescent protein; SR, serine racemase; HA, hemagglutinin; HEK293, humanembryonic kidney 293 cells; GST, glutathione S-transferase; ATP�S, adeno-sine 5�-O-(thiotriphosphate); aa, amino acids.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 281, NO. 29, pp. 20291–20302, July 21, 2006© 2006 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
JULY 21, 2006 • VOLUME 281 • NUMBER 29 JOURNAL OF BIOLOGICAL CHEMISTRY 20291
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

uitylation and degradation rate. Our data disclose new mecha-nisms modulating D-serine production, with implications forthe regulation of NMDA receptors.
EXPERIMENTAL PROCEDURES
Materials—L-Serine and D-serine were purchased fromBachem. Acetonitrile, 5-bromo-4-chloro-3-indolyl-�-D-galac-topyranoside (X-gal), deoxyribonuclease I, anti-glial fibrillaryacidic protein, anti-microtubule-associated protein 2 antibod-ies, and imidazole, were obtained from Sigma. Monoclonalanti-hemagglutinin (HA), anti-HA matrix, and anti-Mycmatrixwere obtained fromCovance.Monoclonal anti-Mycwasobtained from Calbiochem. Polyclonal anti-Myc was fromSanta Cruz Biotechnology. Monoclonal anti-GM130 was fromBD Biosciences. Basal Medium Eagle, minimum essentialmedium, fetal bovine serum, glutamine, penicillin-streptomy-cin, trypsin, and soybean trypsin inhibitor were obtained fromBiological Industries (Kibbutz Beit Haemek, Israel). B27 sup-plement was obtained from Invitrogen. Other reagents were ofanalytical grade.Yeast Two-hybrid Screening—The yeast two-hybrid experi-
ments were carried out as previously described (21) using full-length mouse serine racemase cloned into ppC97 as bait andscreening both a rat hippocampal and a human fetal brainlibrary in ppC86 (provided by Prof. P. Worley, Johns HopkinsUniversity). Positive interactors were identified by their abilityto grow on selective plates (Trp�, Leu�, His�) and activate thetranscription of the reporter gene, �-galactosidase.Cell Culture and Transfection—HEK293 cells were cultured
in Dulbecco’s modified Eagle’s medium containing 10% fetalbovine serum and antibiotics. For transfection, cells wereplated into 6-well tissue culture plates (Nunc) at 70–90% con-fluence. On the next day cells were transfected with mousefull-length serine racemase in pRK5 containing HA in the Nterminus (13), Golga3-Myc (golgin-160) in pcDNA 3.1 (pro-vided by Prof. C. E. Machamer, Johns Hopkins University),Myc-LacZ in pRK5, or green fluorescent protein (GFP) in pRK5using Polyfect reagent (Qiagen). Cells were used 24–48 h aftertransfection.In Vitro Binding Assays—Extracts of transfected HEK293
cellswere incubatedwith 5�g/mlGSTorGST-serine racemase(22) bound to glutathione-Sepharose beads for 1 h at 4 °C inbuffer containing 20mMTris-HCl (pH 7.4), 100mMNaCl, 0.1%Triton X-100, and protease inhibitors (Complete, Roche Diag-nostics). The beads were washed 5 times with the same buffersupplemented with 0.3 M NaCl, and bound material wasresolved by SDS-PAGE and detected by Western blot analysis.Co-immunoprecipitation Assays—Transfected HEK293 cells
were lysed by sonication in buffer containing 20 mM Tris-HCl(pH 7.4), 100 mM NaCl, and protease inhibitors (Complete,Roche Diagnostics). Then 0.1% Triton X-100 was added, andthe cell extract was cleared by centrifugation at 18,000 � g for10min. The supernatant was incubated for 4 h at 4 °CwithMycaffinity matrix (Covance), and the immunoprecipitates werewashed 6 times with lysis buffer supplementedwith 0.3 MNaCl.The blots were probed with monoclonal anti-HA (1:2000) ofpolyclonal anti-Myc (1:2000). For co-immunoprecipitation ofendogenous proteins, rat brains were homogenized using a
glass homogenizer with 5 volumes of 20 mM Tris-HCl (pH 7.4),0.3 M NaCl, 0.5% Triton X-100, and protease inhibitors (Com-plete, RocheDiagnostics). The homogenatewas cleared by cen-trifugation (40,000� g) for 40min). Tenmicrogram of purifiedantibodies to serine racemase and glucosamine-6-phosphatedeaminase (23) were coupled to protein G beads using dimeth-ylpimelimidate as previously described (24) and incubated for12 h with rat brain homogenate (1.5 mg/ml). After washing 7times with lysis buffer containing 0.5% Triton X-100 and0.5 M NaCl, the immunoprecipitates were revealed with anti-GCP170 (Golga3) polyclonal antibody generated by Misumiet al. (18).Brain Subcellular Fractionation—Subcellular fractionation
was carried out as described by Huttner et al. (25). Brains fromadult Sprague-Dawley rats were homogenized with in 5 vol-umes of buffer containing 5 mM Hepes-NaOH (pH 7.3), 0.32 Msucrose, 0.4 mM phenylmethylsulfonyl fluoride, and 1 �g/ml ofaprotinin, leupeptin, chymostatin, and pepstatin. The homoge-nate was centrifuged at 800 � g for 15 min to give P1 (crudenuclear pellet) and S1 (supernatant). S1 was centrifuged at9200� g for 15min to give P2 (crude synaptosomal/mitochon-drial pellet) and S2. P3 (microsomal pellet) and S3 (cytosolicfraction) were obtained by centrifuging S2 at 120,000 � g for1 h. P3 washed (washed microsomal pellet) was obtained bywashing the P3membranes in 50ml of buffer containing 20mMTris-HCl (pH 7.4) and 0.3 M NaCl to remove loosely boundproteins. Samples were analyzed by SDS-PAGE and Westernblot, and probed with antibodies anti-Golga3 (1:1000–1:2000),anti-SR serum (1:5000), and anti GM130 (1:2500).EnzymeMarkers—Lactate dehydrogenase activity was deter-
mined bymonitoring the conversion of pyruvate to lactate cou-pled to oxidation of NADH (26). Fumarase activity was deter-mined bymonitoring the conversion ofmalate into fumarate bya photometric assay (26).Primary Cell Cultures—Animalswere killed by quick decapita-
tion with the approval of the Committee for the Supervision ofAnimal Experiments (Technion-Israel Institute of Technology).Neuronal cultures from cerebral cortex were prepared from E16-E18 Sprague-Dawley embryos as previously described (27).Nearlypure glial cultures were prepared from post-natal day 3 rat pupsusing Basal Medium Eagle containing 10% fetal bovine serum.The cells were vigorously washed with Hanks’-buffered salinesolution 4–5 times every 2 days to dislodge neuronal cells andmicroglia. Glial cultures were used 10–14 days after plating.Assay of D-Serine Synthesis—HEK293-transfected cells were
incubated in 6-well plates with 1 ml of Dulbecco’s modifiedEagle’s medium. D-Serine was measured in the culture mediumby high performance liquid chromatography as previouslydescribed (16).Immunocytochemistry Assays—Primary neural culture cells
were fixedwith 4%paraformaldehyde for 20min.The cellswereblocked and permeabilized with 8% normal goat serum, 0.1%Triton X-100 in Tris-buffered saline (50 mM Tris-HCl (pH 7.4)and 150 mM NaCl). Primary antibody incubation was carriedout for 12–16 h at 4 °C with the polyclonal antibody to serineracemase (Ab2, 0.3 �g/ml) previously characterized (15),monoclonal anti-GM130 (1:200, BD Biosciences), and serumanti-GCP170 (Golga3) (1:150). Immunolabeling was detected
Serine Racemase Is Degraded through the Proteasome
20292 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 29 • JULY 21, 2006
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

by using Cy2- and Cy3-labeled secondary antibodies (JacksonImmunoresearch Laboratories). For double-labeling with poly-clonal antibodies (serine racemase and Golga3), serine race-mase was first labeled using Cy3-conjugated anti-Fab fragmentanti-rabbit IgG that blocks all the primary antibody sites. Thenrabbit anti-GCP170 (Golga3) was added followed by Cy-2-con-jugated anti-rabbit IgG. Co-localization was analyzed by confo-cal laser microscopy.Lentivirus Production—N-terminal HA-tagged Golga3
(455–888 fragment) was subcloned into pTK208 lentiviral vec-tor (a gift from Dr. T. Kafri, Univ. of North Carolina, ChapelHill) containing cytomegalovirus promoter. The virus was pro-duced by calcium-mediated co-transfection of the lentiviralvector (5 �g), packing vector pCMV-dR8.74 (3 �g), and vesic-ular stomatitis virus glycoprotein envelope pMD2G (2 �g) (agift from Prof. D. Trono, University of Geneva) inHEK293 cellsgrown in 10-mm culture dishes (29). Control virus consisted ofGFP under the control of cytomegalovirus promoter. Viralstocks were produced by concentrating the viral particles pres-ent in the culture medium by centrifugation at 120,000 � g for2 h. The pellet was washed once by centrifugation with freshculture medium and suspended in a small volume of basalmedium Eagle culture medium. The viruses were stored at�70 °C until use. Virus titer was determined by infectingHEK293 cells and monitoring the immunofluorescence forGFP or immunocytochemistry for HA-Golga3 (455–880) withanti-HA antibody. To infect primary astrocyte cultures, viralstocks were added to cultured cells grown in 24-well plates, andthe medium was replaced after 48 h. The experiments werecarried out 5 days after infection under conditions that virtuallyall cells had been infected.In Vitro Ubiquitylation—Mouse full-length serine racemase
was translated in vitro by the use ofWheat germ extract in vitrotranslation system (Promega) and [H3]methionine (AmershamBiosciences). For in vitro ubiquitylation, 1 �l of the translatedprotein was incubated with 40 mM Tris-HCl (pH 7.6), 2 mM
dithiothreitol, 5mMMgCl2, 1�Mubiquitin aldehyde, 0.6mg/mlubiquitin (Sigma), 2 mM ATP�S (Sigma), and 4 mg/ml trans-fected HEK293 extract for 60 min at 37 °C. Controls were car-ried out by omitting ATP�S and removing endogenous ATPwith 20 mM deoxyglucose and 240 units/ml of hexoquinase(Sigma). The ubiquitin-protein conjugates were resolved bySDS-PAGE followed by phosphorimaging analysis. For produc-ing HEK293 cell extracts, the cells were transfected withGolga3-Myc in pCDNA 3.1 using Polyfect reagent (Qiagen).Forty-eight hours after transfection, the cells were harvestedand lysed by 2 freeze/thaw cycles in liquid N2 in medium con-taining 20mMTris-HCl (pH 7.6), 5mMKCl, 2mMMgCl2, 1mM
dithiothreitol, deoxyribonuclease I (10 units/ml), 0.4 mM phen-ylmethylsulfonyl fluoride, 30 �M MG132, and 1 �g/ml leupep-tin, pepstatin, aprotinin, and chymostatin. The suspension wascleared by centrifugation at 16,000� g for 40min and frozen inaliquots at �70 °C.In Vivo Ubiquitylation—HEK293 cells were transfected with
FLAG-ubiquitin, HA-serine racemase (HA-SR) and eitherGFP-pRK5 or Golga3-Myc in pcDNA3.1. Forty eight hoursafter transfection, the cells were incubated with 30 �M MG132
for 5 h. Stringent immunoprecipitation conditionswere carriedout as previously described (30), with some modifications. Themediumwas aspirated, and the cells were lysed inmedium con-taining 20 mM Tris-HCl (pH 7.4), 1% SDS, 1 mM EDTA, andprotease inhibitor mixture (Complete, Roche Diagnostics), andthe samples were immediately boiled for 5 min. Then the sam-ples were cleared by centrifugation at 16,000� g for 10min anddiluted 10-fold into buffer containing 2% Triton X-100, 0.5%deoxycholate, 1 mM EDTA, and protease inhibitor mixture.After additional centrifugation to remove any insoluble mate-rial, immunoprecipitation was carried out with anti-HA affini-ty-matrix and washed 6 times with radioimmune precipitationassay buffer (150 mM NaCl, 1% Nonidet P-40, 0.5% deoxy-cholate, 0.1% SDS, 50 mM Tris-HCl (pH 7.5)). Then the ubiq-uitin-serine racemase conjugates were revealed with polyclonalanti-FLAG (Sigma).Pulse-chase Experiments—HEK293 cells were transfected
with HA-SR and either Golga3-Myc or GFP at a SR/Golga3cDNA ratios of 1:5 to 1:9. Forty-eight hours after transfection,the medium was replaced by Dulbecco’s modified Eagle’smedium lacking methionine/cysteine (Sigma) for 1 h. Then thecells were pulsed with methionine/cysteine-free medium con-taining 100 �Ci of [35S]methionine/cysteine (PerkinElmer LifeSciences) for 70 min and subsequently chased in completeDulbecco’s modified Eagle’s medium. At the specified times,the cells were harvested and lysed in buffer containing 20 mMTris-HCl (pH 7.4), 0.5 MNaCl, 1%TritonX-100, 30�MMG132,and protease inhibitors (Complete, Roche Diagnostics). Immu-noprecipitation of HA-serine racemase was carried out withanti-HA affinity matrix (Covance), and the immunoprecipi-tates were washed six times with radioimmune precipitationassay buffer. The samples were resolved on SDS, 10% PAGEgels, transferred to nitrocellulose membranes, and quantifiedby phosphorimaging analysis. The amount of immunoprecipi-tated HA-serine racemase was checked by Western blot of themembrane followed by densitometry of the chemiluminescentsignal.
RESULTS
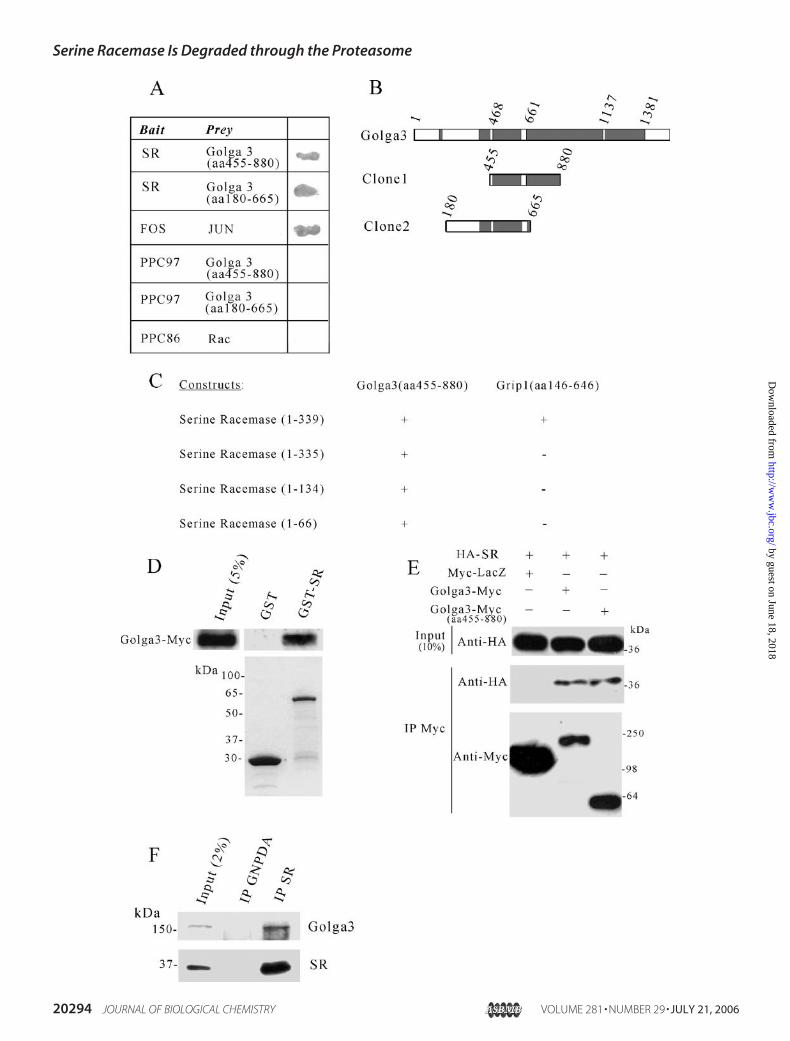
SR Interacts with Golga3—We identified Golga3 protein as anovel interactor of serine racemase in yeast two-hybrid screenings(Fig. 1A). Two independent clones of Golga3 were identified inscreenings of different libraries and encompass the amino acidregions 455–880 and 180–665 (Fig. 1B). Golga3 associates withthe cytosolic surface of the Golgi apparatus, where it may beinvolved in vesicular trafficking (31, 32). Analysis of the primaryamino acid sequence indicates that the interacting region ofGolga3 (amino acids 455–665) is predicted to adopt a coiled-coilconformation (Fig. 1B, shaded boxes). The interaction of Golga3fragments with serine racemase in yeast was specific, as Golga3-transformed cells did not grow on selective plates when co-trans-formed with empty vector pPC97 (Fig. 1A). Also, mouse serineracemase did not activate the reporter genewhen co-transformedwith empty pPC86 vector (Fig. 1A). As a positive control, weobserved that co-transformation of fos and jun strongly activated�-galactosidase expression (Fig. 1A). We further mapped theinteracting region of serine racemase using yeast two-hybrid (Fig.1C). As control, we used glutamate receptor-interacting protein 1
Serine Racemase Is Degraded through the Proteasome
JULY 21, 2006 • VOLUME 281 • NUMBER 29 JOURNAL OF BIOLOGICAL CHEMISTRY 20293
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from
Serine Racemase Is Degraded through the Proteasome
20294 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 29 • JULY 21, 2006
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

(Grip) fragment, previously shown to interactwith theC terminusof serine racemase (9). Contrasting to glutamate receptor-inter-acting protein 1, we found that Golga3 binds to the first 66 aminoacids of the N-terminal region of serine racemase (Fig. 1C).To confirm the interaction, we carried out GST binding
assays. Full-length Golga3 construct containing a C-terminalMyc tag (20) was transfected into HEK293 cells, and the cellextract was incubated with beads containing GST-serine race-
mase (GST-SR) or GST alone.Golga3-Myc bound to GST-SR butnot to GST alone (Fig. 1D).The interaction of Golga3 with
serine racemase was also confirmedin intact cells by co-transfectingHA-serine racemase (HA-SR) withGolga3-Myc (full-length and 455–880 fragment) or Myc-LacZ controlcDNA in HEK293 cells. The Myc-tagged proteins were immunopre-cipitated with anti-Myc, and theblot was probed with anti-HA. Ser-ine racemase robustly co-immuno-precipitated with Golga3 constructsbut not with LacZ (Fig. 1E).To verify that the proteins inter-
act in vivo, we carried out co-im-munoprecipitation experimentsfrom rat brain extracts. Beads cou-pled with purified antibody to ser-ine racemase specifically co-im-munoprecipitated Golga3 (Fig. 1F,upper panel). To ensure specificbinding, the immunoprecipitateswere washed with high salt buffercontaining 0.5% Triton X-100 and0.5 M NaCl. Furthermore, beadscoupled with an antibody againstthe unrelated protein glucosa-mine-6-phosphate deaminase didnot co-immunoprecipitate Golga3(Fig. 1F, upper panel). This sug-gests that Golga3 and serine race-mase specifically interact in vivo.Subcellular Distribution of SR
andGolga3—In case the interactionis physiologically relevant, serineracemase andGolga3 should display
overlapping localizations. We first examined the subcellularlocalizations of endogenous Golga3 and serine racemase inbrain extracts. In the brain we detected enrichment of Golga3in high speed membrane fraction (P3) (Fig. 2A). A significantfraction of Golga3 remained bound to themembrane even afterwashing with buffer containing high salt (P3 washed), indicat-ing a strong interaction with membranes (Fig. 2A). It is note-worthy that significant levels of Golga3 were also found in the
FIGURE 2. Subcellular fractionation of endogenous Golga3, serine racemase, and GM130. A, Western blotanalysis of Golga3 distribution in rat brain subcellular fractions. Golga3 is enriched in microsomal fraction (P3), anda large fraction remains bound to the membrane fraction after washing with buffer containing 20 mM Tris-HCl (pH7.4) and 0.3 M NaCl (P3 washed ). Substantial levels of Golga3 were found in the cytosol. Each lane contained 40 �g ofprotein. B, Western blot analysis of SR distribution in rat brain subcellular fractions. SR is enriched in the cytosolicfraction, but substantial levels of the enzyme are also found in the microsomal fraction (P3) and remains bound afterwashing with high salt buffer (P3 washed ). C, GM130 subcellular distribution (upper panel ) and relative activity ofenzyme markers (lower panel ). The cis-Golgi matrix protein GM130 is enriched in the P3 fraction and remains boundafter washing (P3 washed ). The cytosolic marker lactate dehydrogenase (LDH ) is enriched in the cytosolic fraction,and only trace levels are found in the P3 fraction. BD, below detection. The mitochondrial matrix enzyme fumarase(Fum.) is enriched in the P2 fraction, and only low levels are found in the P3 fraction. The enzyme activities areexpressed as units/mg of protein. H, homogenate; P1, crude nuclear pellet; P2, crude synaptosomal/mitochondrialfraction; P3, microsomal fraction; P3 washed, washed microsomal fraction.
FIGURE 1. Golga3 interacts with serine racemase both in vitro and in vivo. A, interaction of SR with Golga3 in yeast. Positive interaction was revealed by theappearance of �-galactosidase-positive colonies. B, schematic representation of Golga3 protein and clones obtained in the yeast-two hybrid screening. Shadedareas correspond to coiled-coil domains. C, mapping of the interacting region of serine racemase with Golga3 fragment (aa 455– 880) and glutamate receptor-interacting protein 1 (Grip; aa 146 – 646) by yeast-two hybrid analysis. Positive interaction was revealed by �-galactosidase assay. D, interaction of Golga3 withGST-SR and not with GST alone. Cell extracts were prepared from HEK293 cells transfected with full-length Myc-Golga3 and incubated with glutathione-agarose beads containing GST or GST-SR. After washing, the bound material was detected by Western blot using monoclonal anti-Myc antibody. The lowerpanel depicts SDS-PAGE analysis of the recombinant proteins used for the GST binding assay. E, serine racemase was specifically co-immunoprecipitated withGolga3-Myc and Golga3-Myc (aa 455– 880). HEK293 cells were co-transfected with 0.8 �g of HA-SR and 0.2 �g of Golga3-Myc, Golga3-Myc (aa 455– 880), orMyc-LacZ plasmid constructs. Myc-tagged proteins were immunoprecipitated with anti-Myc matrix, and co-immunoprecipitation (IP) with racemase wasrevealed with anti-HA antibody (middle panel). The lower panel demonstrates the immunoprecipitation of the Myc-tagged proteins. F, in vivo interaction ofGolga3 with serine racemase. Rat brain protein homogenate was immunoprecipitated with polyclonal antibody to glucosamine 6-phosphate deaminase(GNPDA) or serine racemase. The immunoprecipitates were washed with buffer containing 0.5% Triton X-100and 0.5 M NaCl. Co-immunoprecipitation ofGolga3 with serine racemase was revealed with anti-Golga3 (GCP170) serum (1:1000) (18) and is depicted in the upper panel. The lower panel demonstrates theimmunoprecipitation of serine racemase protein.
Serine Racemase Is Degraded through the Proteasome
JULY 21, 2006 • VOLUME 281 • NUMBER 29 JOURNAL OF BIOLOGICAL CHEMISTRY 20295
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

Serine Racemase Is Degraded through the Proteasome
20296 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 29 • JULY 21, 2006
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

cytosolic fraction of brain (Fig. 2A). Some bands of smallermolecular weight are also seen in the cytosolic fraction. Thismay be related to the previously described partial cleavage ofGolga3 that releases it from the Golgi membrane (20, 33). Ser-ine racemase was also observed in both soluble and membranefractions of the brain (Fig. 2B). Although most serine racemaseis cytosolic, a portion of serine racemase protein is present inthe microsomal fraction (P3) of brain (Fig. 2B). Association ofserine racemase to the membrane fraction is strong, sincewashing the membranes with high salt solution did not removethe membrane-bound enzyme (Fig. 2B, compare P3 withwashed P3).We found that the cis-Golgimatrix proteinGM130is highly enriched in P3 and washed P3 fraction, whereas onlytrace levels were found in the cytosolic fraction (Fig. 2C). Thisindicates that Golgi membrane fragments are enriched in P3fraction and that our cytosolic fraction is mostly devoid ofmembranes. In addition, we monitored the activity of theenzymes lactate dehydrogenase and fumarase, which are spe-cificmarkers for cytosol andmitochondriamatrix, respectively.We found that lactate dehydrogenase activity is clearlyenriched in the cytosol and virtually absent from the P3 frac-tions, indicating that our microsomal preparation is devoid ofcytosolic contaminants (Fig. 2C). The fumarase activity wasenriched in P2, which corresponds to a fraction enriched inmitochondria and synaptosomes, whereas only traces weredetected in P3 (Fig. 2C). These findings are consistent with thenotion that a part of the racemase molecules in cells may bespecifically associated with microsomal membrane containingGolga3.Co-localization of SR and Golga3—In co-transfected
HEK293 cells, serine racemase and Golga3 exhibited co-local-ization both at the cytosol and perinuclear region (Fig. 3,A–C).Serine racemase was distributed throughout the cytosol (Fig.3A). In some cells, a cytosolic distribution of Golga3 was alsoapparent (Fig. 3B, arrow), whereas in others Golga3 was con-centrated in the perinuclear region with lighter staining of thecytosol (Fig. 3B). This pattern of Golga3 distribution in trans-fected cells was also observed in the absence of serine racemaseco-transfection (data not shown).To confirm that parts of the endogenous proteins co-localize,
we analyzed the localizations of Golga3 and serine racemase inprimary neural cultures. Consistentwith our recent descriptionof serine racemase in primary neuronal cultures using a newantibody to serine racemase (15), we found the enzyme to bepresent in the cytosol as well as in neuronal processes (Fig. 3D).Likewise, Golga3 was detected throughout the cytosol and neu-ronal processes and co-localized to a large extent with serineracemase (Fig. 3, E and F). The identity of the cells as neuronswas routinely checked with an antibody to microtubule-associ-ated protein 2 (data not shown). To further evaluate possibleco-localization of serine racemase and Golga3 at the Golgi
membrane, we carried out double-labeling with the specificGolgi marker GM130. This revealed that a significant fractionof endogenous serine racemase and Golga3 co-localized at theGolgi apparatus in primary neuronal cultures (Fig. 3, G–L). Asimilar pattern was observed in primary astrocyte cultures (Fig.3, M–R). Astrocyte cultures were positive for the astrocyticmarker glial fibrillary acidic protein (data not shown). Togetherwith the immunoprecipitation data (Fig. 1), the results indicatethat serine racemase and Golga3 interact in vivo and displaypartially overlapping localizations at the cytosol and at theGolgi apparatus.Golga3 Increases Steady-state SR Levels—To investigate the
role of serine racemase-Golga3 interaction, we first carried outco-transfection studies in HEK293 cells. Interestingly, weobserved that Golga3-Myc promoted a dose-dependentincrease in serine racemase steady-state protein levels in com-parison to cells co-transfected with GFP (Fig. 4, A and B). Asadditional control, we investigated the effect of the Golga3N-terminal fragment (aa 1–380) that does not interact withserine racemase in yeast two-hybrid assays (data not shown). Asshown in Fig. 4C, this non-interacting construct did notincrease the levels of serine racemase (lanes 3 and 4) as com-pared with the full-length Golga3-Myc (lanes 1 and 2). Thestriking increase in steady-state serine racemase levels pro-moted by Golga3-Myc was associated to a robust increase inD-serine synthesis, which accumulated in the culture media(Fig. 4D). This indicates that Golga3 promotes the accumula-tion of a functional serine racemase enzyme. Co-transfection ofthe Golga3 fragment (aa 455–880) isolated in the yeast two-hybrid screening also promoted an increase in both serine race-mase levels and D-serine synthesis in HEK293 cells (data notshown).To assure that Golga3 modifies endogenous levels of ser-
ine racemase, we produced recombinant lentivirus carryingthe cDNA of the Golga3 fragment (aa 455–880) that inter-acts with serine racemase and investigated its effects on ser-ine racemase expression in primary astrocyte cultures. Infec-tion of the cultures with Golga3 fragment doubled thesteady-state levels of endogenous serine racemase levelswhen compared with lentivirus containing GFP (Fig. 4 E andF). Actin levels were unchanged, indicating the specificity ofthe effect (Fig. 4E).Proteasomal Degradation of SR—To investigate the mecha-
nisms regulating serine racemase levels and the role of Golga3,we first testedwhether serine racemase is degraded by the ubiq-uitin-proteasome system, which mediates selective degrada-tion of many short-lived proteins (34). The addition of the pro-teasome inhibitor MG132 more than doubled the steady-statelevels of endogenous serine racemase in primary astrocyte cul-
FIGURE 3. Co-localization of serine racemase, Golga3, and GM130 in transfected cells and primary cultures. A–C, co-localization of serine racemase (A)and Golga3 (B) in transfected HEK293 cells analyzed by confocal laser microscopy. Significant co-localization was observed in the perinuclear region. In afraction of the cells Golga3 was observed in the cytosol (panel B, arrow), where it also co-localized with serine racemase. D–F, co-localization of endogenousserine racemase (D) and Golga3 (E ) at the neuronal somata and processes of cortical primary neuronal cells (Neu). G–I, co-localization of endogenous serineracemase (G) and GM130 (H ) in the perinuclear region of neurons. J–L, co-localization of endogenous Golga3 (J ) and GM130 (K) in the perinuclear region ofneurons. M–O, co-localization of endogenous serine racemase (M) and Golga3 (N ) at the cytosol and glial processes of cortical primary astrocyte cells (Ast). P–R,co-localization of endogenous Golga3 (P) and GM130 (Q) in the perinuclear region of astrocytes. The bar represents 10 �m. Ast, primary astrocyte culture; Neu,primary neuronal culture; Transf., transfected HEK293 cells.
Serine Racemase Is Degraded through the Proteasome
JULY 21, 2006 • VOLUME 281 • NUMBER 29 JOURNAL OF BIOLOGICAL CHEMISTRY 20297
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

Serine Racemase Is Degraded through the Proteasome
20298 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 29 • JULY 21, 2006
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

tures, whereas actin levels were unchanged (Fig. 5, A and B).Another proteasomal inhibitor, lactacystin (10 �M), also signif-icantly increased serine racemase levels (data not shown). Thus,serine racemase seems to be degraded through the proteasome.In transfected HEK293 cells, treatment with MG132 alsoincreased serine racemase levels when co-transfected with thecontrol protein LacZ (Fig. 5C). However, MG132 did not pro-mote additional serine racemase accumulation beyond thatobserved by co-transfection with Golga3 (Fig. 5, C and D).Thus, wewonderedwhetherGolga3 could increase serine race-
mase levels by slowing down serineracemase degradation through theubiquitin proteasome system.MG132 is known to cause robustaccumulation of polyubiquitin-pro-tein conjugates due to proteasomalinhibition (34). These conjugates,however, were not observed in theWestern blot of Fig. 5C, possiblybecause the polyubiquitin chainsmight conceal the epitopes recog-nized by the antibody to serine race-mase. To directly monitor theeffects of Golga3 in the ubiquityla-tion and degradation rate of serineracemase, we carried out ubiquity-lation studies (Fig. 6) and measure-ment of serine racemase half-life(Fig. 7).Golga3 Decreases SR Ubiquit-
ylation—We first investigated ifserine racemase is a substrate forthe ubiquitin system by employingan in vitro ubiquitylation assay.Serine racemase was translated invitro and incubated with ubiquitinand HEK293 cell extracts to pro-vide the other components of theubiquitin system. We observedformation of high molecularweight species compatible withpolyubiquitylation of serine race-mase (Fig. 6A). As expected, theprocess was ATP-dependent, asremoval of ATP decreased theubiquitylation to a large extent. Toevaluate the role of Golga3, we
investigated the effect of HEK293 cell extracts from cellsoverexpressing Golga3 and monitored serine racemase invitro ubiquitylation (Fig. 6A). Inspection of the ubiquitylatedconjugates revealed that Golga3 decreased the formation ofthe higher molecular weight conjugates of serine racemase,located at the top of the gel (Fig. 6A, region a). The highermolecular weight conjugates are formed by large serine race-mase-polyubiquitin conjugates, which are more efficientlydegraded by the proteasome when compared with lowermolecular weight conjugates (35). Golga3 did not affect the
FIGURE 4. Golga3 increases serine racemase steady-state levels. A, Western blot analysis of co-transfected HEK293 cells extracts showing a dose-dependentincrease in serine racemase steady-state protein levels (upper panel) as a function of Golga3 cDNA employed in the transfection. The cells were transfected with0.1 �g of HA-SR and various amounts of Golga3-Myc and GFP to keep the total DNA concentration at 1 �g. The lower panel depicts Golga3 levels detected withanti-GCP170 antibody. B, densitometric quantification of Western blot of panel A showing the increase in serine racemase levels as a function of the amount ofGolga3-Myc. C, Western blot analysis of HEK293 cells extracts co-transfected with 0.1 �g of HA-SR and either 0.9 �g of Golga3-Myc (lanes 1 and 2) or thenon-interacting Golga3-Myc fragment (1–380) (lanes 3 and 4). The lower panel depicts the expression of Golga3 constructs using anti-Myc antibody. D, analysisof D-serine synthesized and released into the culture media upon the addition of 4 mM L-serine 40 h after co-transfection of HA-SR with either Golga3-Myc(1–380) or Golga3-Myc as described in panel C. The values represent the average � S.E. of four experiments with different cultures. E, infection of primaryastrocyte culture with HA-Golga3 (aa 455– 880) recombinant lentivirus increased the steady-state levels of endogenous serine racemase in comparison withGFP virus control. The upper panel depicts endogenous serine racemase expression monitored with a polyclonal antibody to serine racemase. The middle paneldepicts HA-Golga3 (aa 455– 880) monitored with monoclonal anti-HA. The lower panel depicts actin expression in the infected astrocyte cultures. F, densito-metric quantification of serine racemase per actin expression of the Western blot of panel E. O.D., optical density; Au, arbitrary units.
FIGURE 5. Effect of proteasomal inhibition on the steady-state levels of serine racemase. A, treatmentof primary astrocyte cultures for 10 h with 30 �M MG132 promoted an increase in the endogenous serineracemase levels (upper panel ). Actin levels were not increased (lower panel ). Each lane contained 40 �g ofprotein. B, densitometric quantification of serine racemase per actin expression of the Western blot ofpanel A, which shows that MG132 more than doubled the expression of endogenous serine racemase. C,serine racemase levels in HEK293 cells co-transfected with 0.1 �g of HA-SR and either 0.3 �g of Myc-LacZor Golga3-Myc. Thirty-six hours after transfection, the cells were treated for 10 h with either vehicle Me2SO(�) or 30 �M MG132 (�). MG132 treatment did not increase serine racemase levels beyond the increasepromoted by co-transfection with Golga3. The blot was developed with antibody to serine racemase(upper panel ) or actin (lower panel ). D, densitometric quantification of serine racemase per actin expres-sion of the Western blot of panel C. O.D., optical density; Au, arbitrary units.
Serine Racemase Is Degraded through the Proteasome
JULY 21, 2006 • VOLUME 281 • NUMBER 29 JOURNAL OF BIOLOGICAL CHEMISTRY 20299
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from
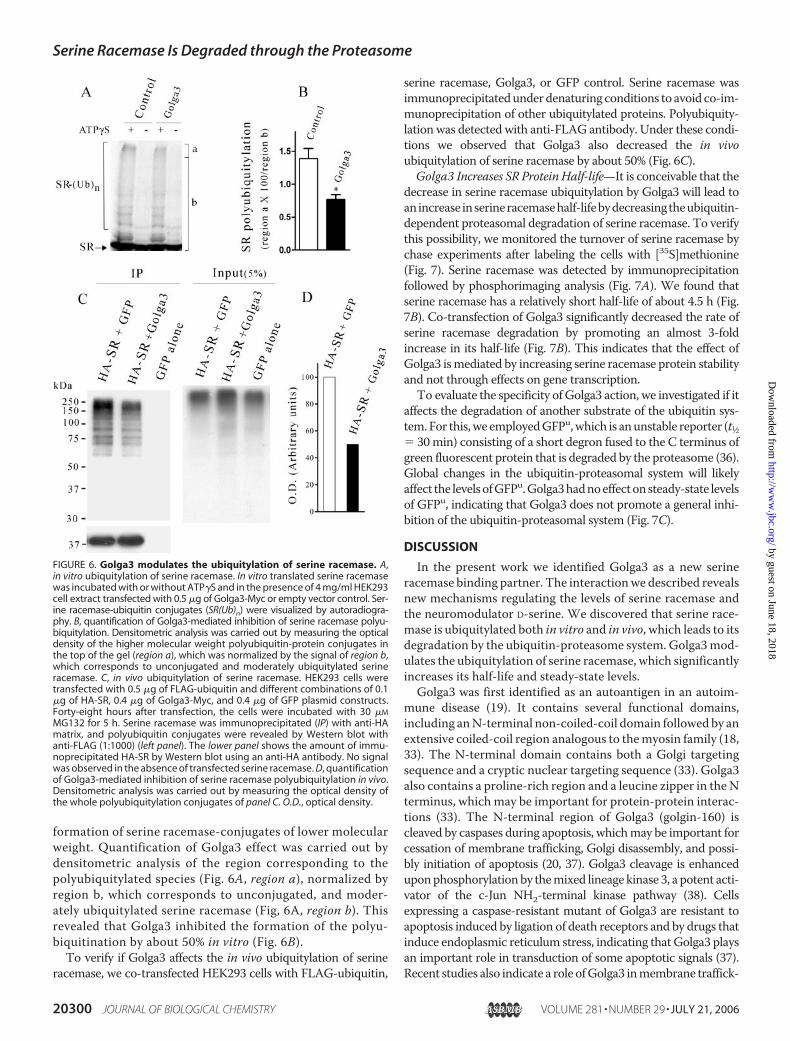
formation of serine racemase-conjugates of lower molecularweight. Quantification of Golga3 effect was carried out bydensitometric analysis of the region corresponding to thepolyubiquitylated species (Fig. 6A, region a), normalized byregion b, which corresponds to unconjugated, and moder-ately ubiquitylated serine racemase (Fig, 6A, region b). Thisrevealed that Golga3 inhibited the formation of the polyu-biquitination by about 50% in vitro (Fig. 6B).To verify if Golga3 affects the in vivo ubiquitylation of serine
racemase, we co-transfected HEK293 cells with FLAG-ubiquitin,
serine racemase, Golga3, or GFP control. Serine racemase wasimmunoprecipitatedunderdenaturing conditions to avoid co-im-munoprecipitation of other ubiquitylated proteins. Polyubiquity-lationwas detected with anti-FLAG antibody. Under these condi-tions we observed that Golga3 also decreased the in vivoubiquitylation of serine racemase by about 50% (Fig. 6C).Golga3 Increases SR ProteinHalf-life—It is conceivable that the
decrease in serine racemase ubiquitylation by Golga3 will lead toanincrease inserineracemasehalf-lifebydecreasingtheubiquitin-dependent proteasomal degradation of serine racemase. To verifythis possibility, we monitored the turnover of serine racemase bychase experiments after labeling the cells with [35S]methionine(Fig. 7). Serine racemase was detected by immunoprecipitationfollowed by phosphorimaging analysis (Fig. 7A). We found thatserine racemase has a relatively short half-life of about 4.5 h (Fig.7B). Co-transfection of Golga3 significantly decreased the rate ofserine racemase degradation by promoting an almost 3-foldincrease in its half-life (Fig. 7B). This indicates that the effect ofGolga3 ismediated by increasing serine racemase protein stabilityand not through effects on gene transcription.To evaluate the specificity ofGolga3 action, we investigated if it
affects the degradation of another substrate of the ubiquitin sys-tem.For this,we employedGFPu,which is anunstable reporter (t1⁄2� 30min) consisting of a short degron fused to the C terminus ofgreen fluorescent protein that is degraded by the proteasome (36).Global changes in the ubiquitin-proteasomal system will likelyaffect the levelsofGFPu.Golga3hadnoeffectonsteady-state levelsof GFPu, indicating that Golga3 does not promote a general inhi-bition of the ubiquitin-proteasomal system (Fig. 7C).
DISCUSSION
In the present work we identified Golga3 as a new serineracemase binding partner. The interactionwe described revealsnew mechanisms regulating the levels of serine racemase andthe neuromodulator D-serine. We discovered that serine race-mase is ubiquitylated both in vitro and in vivo, which leads to itsdegradation by the ubiquitin-proteasome system. Golga3mod-ulates the ubiquitylation of serine racemase, which significantlyincreases its half-life and steady-state levels.Golga3 was first identified as an autoantigen in an autoim-
mune disease (19). It contains several functional domains,including anN-terminal non-coiled-coil domain followedby anextensive coiled-coil region analogous to themyosin family (18,33). The N-terminal domain contains both a Golgi targetingsequence and a cryptic nuclear targeting sequence (33). Golga3also contains a proline-rich region and a leucine zipper in theNterminus, which may be important for protein-protein interac-tions (33). The N-terminal region of Golga3 (golgin-160) iscleaved by caspases during apoptosis, whichmay be important forcessation of membrane trafficking, Golgi disassembly, and possi-bly initiation of apoptosis (20, 37). Golga3 cleavage is enhanceduponphosphorylationby themixed lineage kinase 3, a potent acti-vator of the c-Jun NH2-terminal kinase pathway (38). Cellsexpressing a caspase-resistant mutant of Golga3 are resistant toapoptosis induced by ligation of death receptors and by drugs thatinduce endoplasmic reticulum stress, indicating that Golga3 playsan important role in transduction of some apoptotic signals (37).Recent studies also indicate a role ofGolga3 inmembrane traffick-
FIGURE 6. Golga3 modulates the ubiquitylation of serine racemase. A,in vitro ubiquitylation of serine racemase. In vitro translated serine racemasewas incubated with or without ATP�S and in the presence of 4 mg/ml HEK293cell extract transfected with 0.5 �g of Golga3-Myc or empty vector control. Ser-ine racemase-ubiquitin conjugates (SR(Ub)n) were visualized by autoradiogra-phy. B, quantification of Golga3-mediated inhibition of serine racemase polyu-biquitylation. Densitometric analysis was carried out by measuring the opticaldensity of the higher molecular weight polyubiquitin-protein conjugates inthe top of the gel (region a), which was normalized by the signal of region b,which corresponds to unconjugated and moderately ubiquitylated serineracemase. C, in vivo ubiquitylation of serine racemase. HEK293 cells weretransfected with 0.5 �g of FLAG-ubiquitin and different combinations of 0.1�g of HA-SR, 0.4 �g of Golga3-Myc, and 0.4 �g of GFP plasmid constructs.Forty-eight hours after transfection, the cells were incubated with 30 �M
MG132 for 5 h. Serine racemase was immunoprecipitated (IP) with anti-HAmatrix, and polyubiquitin conjugates were revealed by Western blot withanti-FLAG (1:1000) (left panel). The lower panel shows the amount of immu-noprecipitated HA-SR by Western blot using an anti-HA antibody. No signalwas observed in the absence of transfected serine racemase. D, quantificationof Golga3-mediated inhibition of serine racemase polyubiquitylation in vivo.Densitometric analysis was carried out by measuring the optical density ofthe whole polyubiquitylation conjugates of panel C. O.D., optical density.
Serine Racemase Is Degraded through the Proteasome
20300 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 29 • JULY 21, 2006
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

ing. When expressed in Xenopus oocytes, Golga3 facilitates thetransportof therenal inwardrectifierpotassiumchannel to thecellsurface (39). Golga3 was shown to interact with PIST, a proteininvolved in vesicular trafficking (32). GCP170, a 32-amino acidlonger version ofGolga3, binds toGCP16, an acylatedGolgi-asso-ciated protein thought to be involved in trafficking of cargo to theplasma membrane (31). Despite the above studies, the role ofGolga3 is still unknown, as no specific function has been ascribedto this widely expressed protein.A role of Golga3 in processes unrelated to classic Golgi func-
tions has not been previously investigated, although part ofGolga3was observed in the cytosolic fraction ofHeLa cells (18).We now describe an interaction between Golga3 and serineracemase both in vitro as well as in vivo by co-immunoprecipi-tation studies (Fig. 1). The interaction ismediated by the coiled-coil domain of Golga3, as both isolated clones lack the N-ter-minal non-coiled region (Fig. 1). We observed Golga3 in thecytosolic fraction of brain homogenates (Fig. 2) and also in thecytosol and processes of neuronal and glial cells (Fig. 3). Thissuggests an additional function forGolga3 unrelated to its pres-ence on the cytosolic surface of the Golgi apparatus.An intriguing observation is the presence of serine racemase
strongly bound to the membrane fraction, which was resistantto washing with high salt concentration (Fig. 2). The presenceof serine racemase in the perinuclear region corresponding tothe Golgi apparatus was confirmed by co-localization studieswith a Golgi marker (Fig. 3). Thus, it is possible that Golga3mediates serine racemase binding to the Golgi membrane. Therole of Golgi-associated serine racemase is unclear, since nointracellular target for D-serine has been identified so far. Nev-ertheless, membrane-bound serine racemase displays racemi-zation activity,3 indicating that it may constitute a functionalreservoir of the enzyme to be exported to other places in thecell. In view of the fact that D-serine has been observed in vesi-cle-like structures in some types of cells (40, 41), it is tempting
to speculate that membrane-bound serine racemase could playa role in the cellular compartmentation or release of D-serine.Our presentwork reveals a role forGolga3 inmodulating serine
racemase levels (Fig. 4, A–D). Golga3 also increased the steady-state levels of serine racemase in primary astrocyte cultures, indi-cating that it affects the endogenous serine racemase levels (Fig.4E).We found that the proteasomal activity regulates serine race-mase levels (Fig. 5), and Golga3 promoted a decrease in serineracemase ubiquitylation both in vitro and in vivo (Fig. 6). The datasuggest that a decrease in polyubiquitylation of serine racemaseaccounts for the increase in the steady-state levels (Fig. 4) andhalf-life of serine racemase (Fig. 7) promoted by Golga3.It has been shown that the ubiquitin system regulates traf-
ficking of cargo from Golgi, and some E3 ubiquitin ligases arelocated in this organelle (42–44). However, Golga3 has notbeen previously implicated in the modulation of protein ubiq-uitylation. One possible explanation for Golga3 action is itsinterference with the binding of a still unidentified E3 ubiquitinligase to serine racemase, a class of proteins that promote thetransfer of ubiquitin to proteins and ubiquitin chains (34).Despite the fact that Golga3 did not affect GFPu levels (Fig. 7C),it is possible that Golga3 affects the half-life of additional pro-teins by a mechanism similar to that observed for serine race-mase. The metabolism of D-serine in the forebrain areas is slowdue to the scarcity of the metabolic enzyme D-amino acid oxi-dase enzyme in this region (7). The limitedmetabolismof D-ser-ine may account for the long half-life of D-serine in the brain,which was calculated to be about 12 h (45). On the other hand,we found that serine racemase displays a relatively short half-life of about 4.5 h (Fig. 7), which is compatible with the exist-ence of an efficient degradation/regulatory system. In thisframework, regulation of serine racemase levels by the ubiq-uitin system and through its interactionwithGolga3might playa key role in regulating D-serine synthesis.The co-agonist site of the NMDA receptors that is occupied
by D-serine is an important regulator of the receptor/channelfunction (1). As an endogenous co-agonist, D-serine plays3 E. Dumin and H. Wolosker, unpublished result.
FIGURE 7. Golga3 decreases serine racemase degradation. HEK293 cells were co-transfected with HA-SR and either Golga3-Myc or GFP construct. Cells wereincubated in medium containing [35S]methionine/cysteine, chased for the indicated times, and HA-SR-immunoprecipitated using an anti-HA matrix. A,autoradiography of HA-SR immunoprecipitates. B, quantification of remaining HA-SR in the presence of GFP (E) or Golga3-Myc (F) at different chase times. C,Golga3 does not affect the steady-state levels of GFPu in transfected cells. GFPu was revealed by Western blot using an anti-HA antibody. Each lane contained25 �g of protein. The error bars represent the S.E. of four independent experiments. *, different from HA-SR � GFP at p � 0.05.
Serine Racemase Is Degraded through the Proteasome
JULY 21, 2006 • VOLUME 281 • NUMBER 29 JOURNAL OF BIOLOGICAL CHEMISTRY 20301
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

important roles in regulating NMDA receptor responses andneurotoxicity (6, 8–11, 46). On the other hand, the ubiquitinsystemhas been implicated in protein turnover at synaptic sites,but the mechanisms regulating synaptic proteins by degradationonly started to be unveiled (47). Synaptic scaffold proteins such asShank, AKAP79/150, and guanylate kinase-associated protein(GKAP) are ubiquitylated in an activity-dependent manner andmay regulate the targetingofNMDAreceptors at the synapse (48).Although the NMDA receptor by itself can be ubiquitylated (28),its half-life is quite long (about 20 h (37)). Our data raise the pos-sibility that the ubiquitylation of serine racemase and its modula-tion by Golga3 are likely to play a role in NMDA receptor activityand neurotoxicity as well.In summary, our study discloses new mechanisms regulating
the synthesis of the neuromodulator D-serine, with implicationsfor the regulation ofNMDAactivity and neurotoxicity. Identifica-tion of the E3 ligase responsible for serine racemase ubiquitylationin future studies will allow amore complete understanding of themechanism regulating brain D-serine levels.
Acknowledgments—We thank to Prof. Avram Hershko (Technion-Israel Institute of Technology) for helpful discussions and for provid-ing ATP�S and ubiquitin aldehyde and Drs. Ofer Shenker and EdithSuss-Toby for expert technical assistance with confocal imaging. Weare also indebted to Dr.Michael J. Schell (Uniformed Services Univer-sity of the Health Sciences) for helpful discussions and SR-GFP con-structs and to Dr. Carolyn Machamer (Johns Hopkins University) forgolgin-160/Golga3 cDNA.
REFERENCES1. Danysz, W., and Parsons, A. C. (1998) Pharmacol. Rev. 50, 597–6642. Choi, D.W., and Rothman, S.M. (1990)Annu. Rev. Neurosci. 13, 171–1823. Johnson, J. W., and Ascher, P. (1987) Nature 325, 529–5314. Kleckner, N. W., and Dingledine, R. (1988) Science 241, 835–8375. Wolosker, H., Panizzutti, R., and De Miranda, J. (2002) Neurochem. Int.
41, 327–3326. Shleper, M., Kartvelishvily, E., and Wolosker, H. (2005) J. Neurosci. 25,
9413–94177. Schell, M. J., Molliver,M. E., and Snyder, S. H. (1995) Proc. Natl. Acad. Sci.
U. S. A. 92, 3948–39528. Mothet, J. P., Parent, A. T., Wolosker, H., Brady, R. O., Jr., Linden, D. J.,
Ferris, C. D., Rogawski, M. A., and Snyder, S. H. (2000) Proc. Natl. Acad.Sci. U. S. A. 97, 4926–4931
9. Kim, P. M., Aizawa, H., Kim, P. S., Huang, A. S., Wickramasinghe, S. R.,Kashani, A. H., Barrow, R. K., Huganir, R. L., Ghosh, A., and Snyder, S. H.(2005) Proc. Natl. Acad. Sci. U. S. A. 102, 2105–2110
10. Stevens, E. R., Esguerra, M., Kim, P. M., Newman, E. A., Snyder, S. H.,Zahs, K. R., and Miller, R. F. (2003) Proc. Natl. Acad. Sci. U. S. A. 100,6789–6794
11. Yang, Y., Ge, W., Chen, Y., Zhang, Z., Shen, W., Wu, C., Poo, M., andDuan, S. (2003) Proc. Natl. Acad. Sci. U. S. A. 100, 15194–15199
12. Wolosker, H., Sheth, K. N., Takahashi, M., Mothet, J. P., Brady, R. O., Jr.,Ferris, C. D., and Snyder, S. H. (1999a) Proc. Natl. Acad. Sci. U. S. A. 96,721–725
13. Wolosker, H., Blackshaw, S., and Snyder, S. H. (1999b) Proc. Natl. Acad.Sci. U. S. A. 96, 13409–13414
14. DeMiranda, J., Santoro, A., Engelender, S., andWolosker, H. (2000)Gene(Amst.) 256, 183–188
15. Kartvelishvily, E., Shleper, M., Balan, L., Dumin, E., and Wolosker, H.(2006) J. Biol. Chem. 281, 14151–14162
16. DeMiranda, J., Panizzutti, R., Foltyn, V. N., andWolosker, H. (2002) Proc.Natl. Acad. Sci. U. S. A. 99, 14542–14547
17. Foltyn, V. N., Bendikov, I., De Miranda, J., Panizzutti, R., Dumin, E.,Shleper, M., Li, P., Toney, M. D., Kartvelishvily, E., and Wolosker, H.(2005) J. Biol. Chem. 280, 1754–1763
18. Misumi, Y., Sohda,M., Yano, A., Fujiwara, T., and Ikehara, Y. (1997) J. Biol.Chem. 272, 23851–23858
19. Fritzler, M. J., Hamel, J. C., Ochs, R. L., and Chan, E. K. (1993) J. Exp. Med.178, 49–62
20. Mancini, M., Machamer, C. E., Roy, S., Nicholson, D. W., Thornberry,N. A., Casciola-Rosen, L. A., and Rosen, A. (2000) J. Cell Biol. 149,603–612
21. Liani, E., Eyal, A., Avraham, E., Shemer, R., Szargel, R., Berg, D., Borne-mann, A., Riess, O., Ross, C. A., Rott, R., and Engelender, S. (2004) Proc.Natl. Acad. Sci. U. S. A. 101, 5500–5505
22. Panizzutti, R., DeMiranda, J., Ribeiro, C. S., Engelender, S., andWolosker,H. (2001) Proc. Natl. Acad. Sci. U. S. A. 98, 5294–5299
23. Wolosker, H., Kline, D., Bian, Y., Blackshaw, S., Cameron, A. M., Fralich,T. J., Schnaar, R. L., and Snyder, S. H. (1998) FASEB J. 12, 91–99
24. Harlow, E., and Lane, D. (1988)Antibodies. A LaboratoryManual, 1st Ed.,pp. 343–345, Cold Spring Harbor Laboratory Press, Cold Spring Harbor,NY
25. Huttner, W. B., Schiebler, W., Greengard, P., and De Camilli, P. (1983)J. Cell Biol. 96, 1374–1388
26. Eisenthal, R., and Danson, M. J. (1993) in Enzyme Assays: A PracticalApproach, pp. 75–78, Oxford University Press, Oxford
27. Brewer, G. J., Torricelli, J. R., Evege, E. K., and Price, P. J. (1993) J. Neurosci.Res. 35, 567–576
28. Kato, A., Rouach, N., Nicoll, R. A., and Bredt, D. S. (2005) Proc. Natl. Acad.Sci. U. S. A. 102, 5600–5605
29. Zufferey, R., Nagy, D., Mandel, R. J., Naldini, L., and Trono, D. (1997)Nat.Biotechnol. 15, 871–875
30. Hershko, A., Eytan, E., Ciechanover, A., and Haas, A. L. (1982) J. Biol.Chem. 257, 13964–13970
31. Ohta, E., Misumi, Y., Sohda, M., Fujiwara, T., Yano, A., and Ikehara, Y.(2003) J. Biol. Chem. 278, 51957–51967
32. Hicks, S.W., andMachamer, C. E. (2005) J. Biol. Chem. 280, 28944–2895133. Hicks, S.W., andMachamer, C. E. (2002) J. Biol. Chem. 277, 35833–3583934. Hershko, A., and Ciechanover, A. (1998) Annu. Rev. Biochem. 67,
425–47935. Hershko, A., and Heller, H. (1985) Biochem. Biophys. Res. Commun. 128,
1079–108636. Bence, N. F., Sampat, R. M., and Kopito, R. R. (2001) Science 292,
1552–155537. Maag, R. S.,Mancini,M., Rosen, A., andMachamer, C. E. (2005)Mol. Biol.
Cell 16, 3019–302738. Cha, H., Smith, B. L., Gallo, K., Machamer, C. E., and Shapiro, P. (2004)
J. Cell Sci. 117, 751–76039. Bundis, F., Neagoe, I., Schwappach, B., and Steinmeyer, K. (2006) Cell.
Physiol. Biochem. 17, 1–1240. Mothet, J. P., Pollegioni, L., Ouanounou, G., Martineau, M., Fossier, P.,
and Baux, G. (2005) Proc. Natl. Acad. Sci. U. S. A. 102, 5606–561141. Williams, S. M., Diaz, C. M., Macnab, L. T., Sullivan, R. K., and Pow, D. V.
(2006) Glia 53, 401–41142. Bartke, T., Pohl, C., Pyrowolakis, G., and Jentsch, S. (2004) Mol. Cell 14,
801–81143. Nakamura, N., Fukuda, H., Kato, A., and Hirose, S. (2005)Mol. Biol. Cell
16, 1696–171044. Alroy, I., Tuvia, S., Greener, T., Gordon, D., Barr, H. M., Taglicht, D.,
Mandil-Levin, R., Ben-Avraham, D., Konforty, D., Nir, A., Levius, O., Bi-coviski, V., Dori, M., Cohen, S., Yaar, L., Erez, O., Propheta-Meiran, O.,Koskas,M., Caspi-Bachar, E., Alchanati, I., Sela-Brown,A.,Moskowitz,H.,Tessmer, U., Schubert, U., and Reiss, Y. (2005) Proc. Natl. Acad. Sci.U. S. A. 102, 1478–1483
45. Dunlop, D. S., andNeidle, A. (1997)Biochem. Biophys. Res. Commun. 235,26–30
46. Yang, S., Qiao, H.,Wen, L., Zhou,W., and Zhang, Y. (2005)Neurosci. Lett.379, 7–12
47. Yi, J. J., and Ehlers, M. D. (2005) Neuron 47, 629–63248. Ehlers, M. D. (2003) Nat. Neurosci. 6, 231–242
Serine Racemase Is Degraded through the Proteasome
20302 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 29 • JULY 21, 2006
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

Elena Kartvelishvily and Herman WoloskerElena Dumin, Inna Bendikov, Veronika N. Foltyn, Yoshio Misumi, Yukio Ikehara,
of Serine RacemaseModulation of D-Serine Levels via Ubiquitin-dependent Proteasomal Degradation
doi: 10.1074/jbc.M601971200 originally published online May 19, 20062006, 281:20291-20302.J. Biol. Chem.
10.1074/jbc.M601971200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/281/29/20291.full.html#ref-list-1
This article cites 46 references, 31 of which can be accessed free at
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from