metal-induced inhibition of anaerobic metabolism of volatile fatty acids and hydrogen
TRANSCRIPT

Appl. Microbiol Biotechnol (1994) 42:396-402 © Springer-Verlag 1994
I.-C. Kong • J. S. Hubbard • W. J. Jones
Metal-induced inhibition of anaerobic metabolism of volatile fatty acids and hydrogen
Received: 11 January 1994/Received revision: 31 March 1994/Accepted: 18 April 1994
Abstract The effects of copper (Cu), chromium (Cr), cadmium (Cd), lead (Pb) and zinc (Zn) on the biotrans- formation of organic acids (acetate, propionate and butyrate) and H2 were assessed in serum-bottle micro- cosms. Experiments were performed over a range of metal concentrations (20-200mg/1) using biomass from an anaerobic bioreactor fed continuously with ethanol distillery waste as inoculum. In general, the added metals inhibited the biotransformation of organic acids with increasing metal concentration. However, the extent of inhibition varied for the differ- ent biotransformations and for the different metals tested. For example, the concentration of CuC12 effect- ing a 50% reduction in the rate constant for biotrans- formation of acetate, propionate and butyrate was 60, 75 and 30 mg/1, respectively. Cu and Cr (VI) were the most inhibitory metals in organic acid transformation, whereas Pb was the least toxic. The rate of biotrans- formation of acetate was reduced by half at Cu and Cr concentrations of 60 and 40 rag/1 respectively, whereas Cd, Pb, and Zn concentrations of 160 to 200 rag/1 had little effect. The activities of hydrogenotrophic meth- anogens were much less affected by the same metals and metal concentrations.
I.-C. Kong 1 • J. S. Hubbard School of Biology, Georgia Institute of Technology, Atlanta, GA 30332, USA
W. J. Jones ( ~ ) US Environmental Protection Agency, Environmental Research Laboratory, Athens, GA 30605, USA
Present address: 1 210 Black Hall, Department of Environmental Engineering Science, University of Florida, Gainesville, FL 32611, USA
Introduction
The fixed-film/upflow anaerobic sludge blanket (UASB) bioreactor, containing a biofilm support me- dium in the upper portion and a blanket of settling biomass in the lower section, was designed to increase solids retention, improve gas/solids/liquid separation, and provide a surface for the attachment of biomass (Olthof and Oleszkiewicz 1982; Switzenbaum 1983; Switzenbaum and Grady 1986; Oleszkiewicz and Thadani 1988; Chang 1989). In the anaerobic digestion process, volatile fatty acids (VFAs) are important inter- mediates and are useful indicators of digestor perfor- mance (Mawson et al. 1991). The C2-C7 organic acids are predominant intermediates in the anaerobic diges- tion of organic matter. The anaerobic oxidation of the C3-C7 substrates is coupled to a reduction of protons (H2 formation), and the oxidation of C3 and C4 organic acids is thermodynamically unfavorable (endergonic process) under standard conditions. Maintenance of low H2 partial pressures by H2-oxidizing bacteria such as methanogens provides for a thermodynamically feasible reaction. In this regard, methanogenesis via H2-mediated reduction of CO2 is important in the anaerobic digestion process (Jones et al. 1987; Lin et al. 1989).
The presence of significant amounts of heavy metals may result in the inhibition and failure of sewage diges- tors (McCarty 1964; Stronach et al. 1986). Many stud- ies have demonstrated the toxic effects of heavy metals on anaerobic digestion (Parkin and Owen 1986; Kou- zeli-Katsiri et al. 1988). An important feature that dis- tinguishes heavy metals from other toxic pollutants is that they persist, i.e., are not biodegradable, but in some cases may be biotransformed (Sterrit and Lester 1980; Hickey et al. 1989).
Metals such as mercury (Hg), lead (Pb), cadmium (Cd), copper (Cu), chromium (Cr), and zinc (Zn) are classified as very toxic and readily accessible and are

397
found at elevated levels in many environments (Wood 1974; Keith and Telliard 1979; Nriagu and Pacyna 1988). Hickey et al. (1989) investigated the effects of heavy metals on methane production as well as H2 and carbon monoxide levels in a conventional batch diges- tor. Results indicated that H2 monitoring holds prom- ise as an indicator of digestor performance upset brought on as a consequence of heavy-metal-induced inhibition. More recently, Lin (1992) investigated the effects of heavy metals on VFA degradation in VFA- acclimated sludge. Results from the study of Lin (1992) indicated that the biotransformation of acetate, propionate, and butyrate were rate-limiting reactions in the degradation of synthetic organic waste. Thus, understanding the inhibitory effects of contaminants such as metals on the biotransformation of important intermediates (VFAs, H2) in digestor systems may con- tribute to the knowledge of factors affecting reactor kinetics and overall digestor performance.
In this investigation, the biotransformation of speci- fic organic acids (acetate, propionate, and butyrate) and H2 by anaerobic digestor biomass was assessed in the absence and presence of CuC12, KzCrzOT, CdC12, ZnC12 and PbC12.
Materials and methods
Source of inoculum
Microbial biomass used for serum-bottle microcosm studies was obtained from a methanogenic hybrid bioreactor that had been operated continuously for over 2 years. The laboratory-scale, hybrid bioreactor was of similar design to that described by Chang (1989) and Kitsos et al. (1993). The lower third of the reactor consisted of a granular bed of microbial biomass, and the upper consisted of matrix (aluminum silicate)-attached biomass. The single-stage bioreactor was operated at 36 ± 2°C with a continuous feed of synthetic distillery waste-water. The essential ingredients of the feedstock, containing 30 g Chemical oxygen demand (COD)/1, were 12 g sucrose, 8 g yeast autolysate, 0.3 g glutamate, 0.04 g citric acid, VFAs (1.05 mg acetate, 0.1 mg propionate, and 0.19 mg butyrate), alcohols (1.6 mg ethanol, 0.55 nag methanol, 0.04 mg isobutanol, 0.04 mg amylalcohol, 0.04 mg n-butanol, and 0.04 mg n-propanol), and mineral salts (Kitsos et al. 1993; Balch et al, 1979). A detailed analysis of the pseudo-steady-state operational conditions and per- formance data for the anaerobic bioreactor is described in Table 1.
Inocula for metal-inhibition studies were periodically collected from the middle portion of the bioreactor and consisted predomi- nantly of microbial floes. The biomass was collected in a 500-ml Bellco bottle, purged with O2-free N z gas, capped, and incubated at 36°C for 2 days in order to deplete the background concentration of organic acids. After confirmation of such depletion by gas chromatography (GC), the pH of the inoculum was adjusted to 7.2 by addition of O2-free 1 M HC1. The inoculum was used without any additional modifications.
Biotransformation experiments
Duplicate assays were conducted in serum vials to assess the effects of heavy metals on the anaerobic transformation of acetate, propionate, butyrate, and H 2. Aliquots (15 ml) of the reactor
TabLe 1 Optimal operational conditions and performance data for the methanogenic, hybrid bioreactor (HRT, hydraulic retention time; COD, chemical O 2 demand; CODso ~, soluble COD of effluent; COD~n , introduced COD; CH4, methane)
Parameters Values
Maximum feedstock concentration HRT COD loading rate COD removal Effluent COD~o ~ Total gas production Total CH 4 production CH 4 in effluent gas (%)
30 g COD/1 45 h 23.8 g COD/1 per day 97.8% Approx. 0.6 g COD/1 0.48-0.50 1/g CODin 0.29-0.33 1/g CODi, 62 65%
biomass were dispensed into 60-ml serum vials, and amendments of volatile organic acids and metals were made inside an anaerobic chamber (Coy Laboratory Products, Ann Arbor, Mich., USA). Or- ganic acids were added at an initial concentration of 25 mM; this value was chosen since previous experiments indicated near maxi- mal biotransformation activities at this concentration. Various metals, Cu (II), Cr (VI), Cd (II), Pb (II) or Zn (II), were added to achieve final concentrations of 20 to 200 rag/1. Metal solutions were prepared as concentrated stocks of CuC12, CdCI2, KzCr2OT, PbC12 and ZnC12. All serum vials were sealed with butyl rubber stoppers, flushed with O2-free N 2 gas, and incubated at 36°C with gentle shaking (150 rpm).
To investigate the inhibitory/stimulatory effects of metals on the activity of hydrogenotrophic methanogens, 20-ml aliquots of diges- tor inoculum were distributed into 160-ml serum vials. Methane (CH4) production was determined with time following exchange of the headspace atmosphere with Hz/CO 2 (4:1; 1 atm). The large serum vial headspace-to-liquid ratio ensured that H 2 was not limit- ing; the headspace atmosphere was replaced with the H2/CO 2 mix- ture every 5-10 h. The CH 4 was quantified by GC. All additions and manipulations were performed using strict anaerobic techniques (Balch et al. 1979).
Monitoring
To determine the effects of the added heavy metals on the biotrans- formation reactions, the concentrations of VFAs in serum-vial microcosms were determined every 5-10 h and biotransformation rate constants were determined. Pseudo-first-order biotransforma- tion rate constants . (k) were calculated from plots of the natural logarithm (ln) of substrate concentration versus time according to the integrated, first-order rate equation of Levenspiel (1972):
-- Ln(Ct/Co) = k. t
where C o and C t are the concentrations of VFAs at time zero and time t, respectively.
Analyses
Acetate, propionate, and butyrate were quantified using a Hewlett Packard gas chromatograph (model 5890) equipped with a flame ionization detector, Organic acids were separated using a 2 m (I.D. 3 ram) glass column packed with 80/120 mesh Carbopack B-DA/4% Carbowax 20 M (Supelco, Bellefonte, Pa., USA). In some experi- ments, organic acids were separated and quantified using a 2 m x 3 mm glass column packed with 80-100 mesh Porapak Q (Supelco). In both cases, helium was used as the carrier gas at a flow rate of 20 ml/min.

398
Methane was quantified using a Hewlett Packard gas chromato- graph equipped with a thermal conductivity detector (TCD). Gases were separated on a 2 m x 3 mm glass column packed with 80-100 mesh Porapak Q.
The concentration of biomass collected from the anaerobic diges- tor was estimated by determining total volatile solids (TVS) accord- ing to Standard Methods (APHA 1981).
Results
Effects of metal addition on organic acids biotransformation
The rate of disappearance of added organic acids (25 m~a concentration) in serum-vial microcosms was measured over time at different initial concentrations of added metals. Representative results of the disappear- ance of the added organic acids during a short incuba- tion period in the absence and presence of amended metals are presented as the natural logarithm of the disappearance of organic acid versus time (Fig. la-d). Data from duplicate experiments were comparable and did not differ by more than 10%. Biotransformation rate constants were calculated from the slopes of the pseudo-first-order plots (Fig. 1). The calculated rate Constants (k) for biotransformation of acetate, propion- ate, and butyrate in the absence and presence of various metal solutions (CuClz, CdClz, K2 CrzOT, PbCI2 or ZnClz) over a range of metal concentrations (20-200 rag/l) are presented in Table 2. As expected, biotransformation rate constants for the specific or- ganic acids tested were lower at the higher amended metal concentrations. The biotransformation rate con- stant for butyrate (0.15 h -1) in the absence of added metals was higher than the biotransformation rate con- stant for either propionate (0.08h -1) or acetate (0.11 h-l) .
Generally, the inhibitory effects of added metals on the biotransformation of organic acids increased with higher initial concentrations of metal. At all concentra- tions of CuC1/, the transformation of butyrate was inhibited to a greater extent than that of acetate and propionate (Table 2). For example, at 20 mg Cu/1, the rate constants for butyrate, propionate, and acetate transformation were 60, 88, and 100%, respectively, of the control values. In experiments with 80 mg Cu added per liter, the rate constant for butyrate, propion- ate, and acetate transformation were 13, 50, and 27%, respectively, of the control values. In this case, the transformation of acetate was inhibited to a greater extent than that of propionate. The concentrations of CuCI:, K2Cr/O7, CdC12, ZnC12 and PbClz effecting a 50% reduction in the biotransformation rate con- stants for the various VFAs tested are presented in Table 3. The concentration of CuC12 effecting a 50% inhibition in the biotransformation of propionate was 2.5 and 1.25 times higher than for inhibition of butyrate and acetate transformation, respectively.
The biotransformation of acetate was appreciably inhibited (45% reduction in rate constant) by a low concentration (40rag/l) of added K2Cr/O7 while butyrate and propionate transformation were only moderately (12-20%) inhibited. At 40 mg Cr added per liter, the biotransformation rate constant for acetate was 55% of the level of the control (Table 2). In con- trast, the biotransformation rate constants for propion- ate and butyrate were 88 and 80%, respectively, of the control. However, a different degree of inhibition was noted in experiments amended with higher chro- mium(VI) levels. At 160 mg Cr/1, the biotransformation rate constant for acetate was 36% of the control while the values for propionate and butyrate transformation were 12 and 20%, respectively, of the control. The Cr(VI) concentration effecting a 50% reduction in the biotransformation rate constants for acetate, propionate, and butyrate were 40, 65, and 75 rag/l, respectively.
In microcosms amended with CdC12, the anaerobic transformation of acetate was inhibited to a lesser ex- tent than for either propionate or butyrate transforma- tion (Table 2). At 50 and 200 mg Cd/1, the rate con- stants for acetate biotransformation were 91 and 82%, respectively, of the control value. In contrast, at 50 mg/1 amended Cd (II), the rate constants for biotransforma- tion of butyrate and propionate were 67 and 63%, respectively, of the control values. At a high Cd concen- tration (200 mg/1), the rate constants for butyrate and propionate transformation were 33 and 12%, respec- tively, of the control values. The Cd concentrations effecting a 50% reduction in the rate constants were appreciably different for the different organic acids (Table 3). Cd concentrations of approximately 100, 125, and greater than 200 mg/1 were required to effect a 50% inhibition of the biotransformation of propionate, butyrate, and acetate, respectively.
Among the five metals tested, Pb was the least inhib- itory for the biotransformation of the organic acids (Table 2). Even in microcosms amended with 160 mgPb/l, the rate constants for biotransformation of the three organic acids ranged from 80 to 91% of the control values. At 80 mg Pb/1, essentially no inhibition was observed with any of the organic acids tested. The biotransformation rate constants ranged from 100 to 113% of the control values. The Pb concentrations effecting a 50% reduction in the biotransformation rate constants were greater than 160 mg/1 for all organic acids tested (Table 3).
In microcosms amended with ZnC12, the trans- 'formation of acetate was inhibited to a lesser extent than the biotransformation of either propionate or butyrate (Table 2). At 50 and 200 mg Zn/1, the rate constants for transformation of butyrate were 47 and 20%, respectively, of the control value. In contrast, at 200 mg/1 amended Zn, the rate constant for trans- formation of acetate was 91% of the control. The ZnC12 concentrations effecting a 50% inhibition of
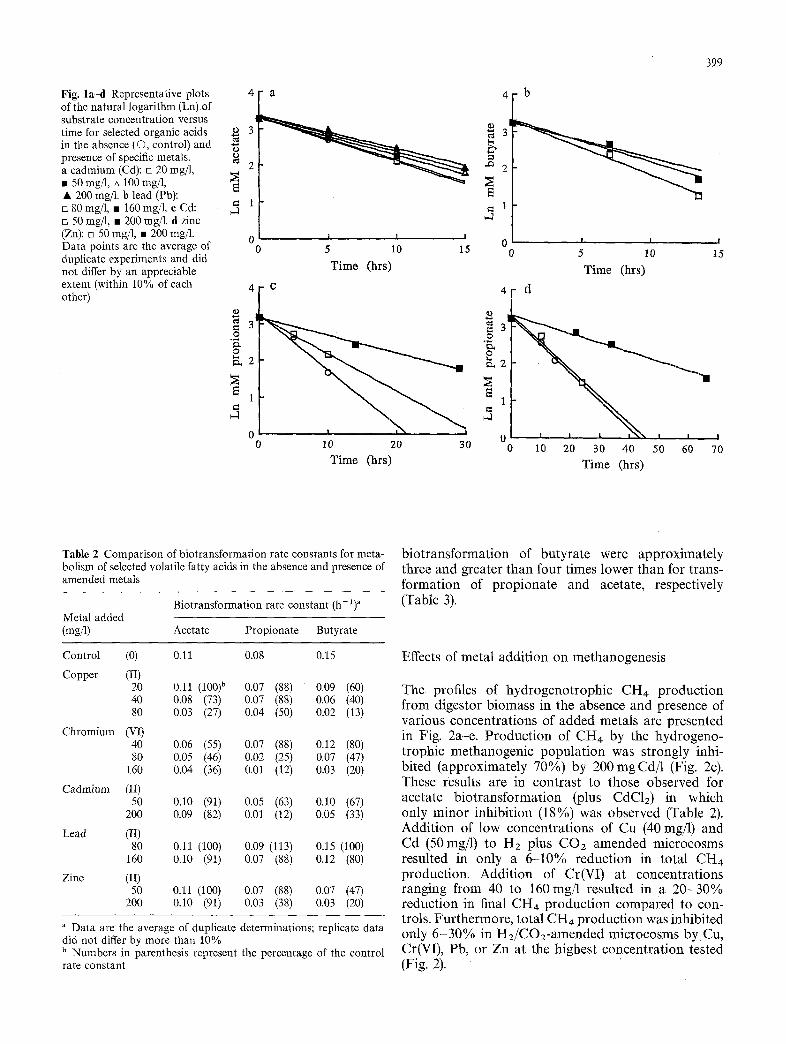
Fig. l a d Representative plots 4 of the natural logarithm (Ln) of substrate concentration versus 3 I time for selected organic acids in the absence (O, control) and ,g presence of specific metals. ,~ a cadmium (Cd): [] 20 mg/1, ~ 2 • 50 mg/1, A 100 rag/l, '~ • 200 mg/1. b lead (Pb):
= 1 [] 80 rag/l, • 160 mg/1. c Cd: [] 50 rag/l, • 200 rag/1, d zinc (Zn): rn 50 rag/l, • 200 mg/1. 0 Data points are the average of 0 duplicate experiments and did not differ by an appreciable extent (within 10% of each 4 other)
3 I 0
~D
~ 2
1
0 0
a 4
31
3 2
1
1 I I 0 5 10 15 0
T ime (hrs)
c 4
~ 2
0 10 20 30 0 T ime (hrs)
399
b
I I I
5 10 15
Time (hrs)
I !
60 70 10 20 30 40 50
T ime (hrs)
Table 2 Comparison of biotransformation rate constants for meta- bolism of selected volatile fatty acids in the absence and presence of amended metals
Biotransformation rate constant (h-~)a Metal added (mg/1) Acetate Propionate Butyrate
Control (0) 0.11 0.08 0.15
Copper (II) 20 0.11 (100) b 0.07 (88) 0.09 (60) 40 0.08 (73) 0.07 (88) 0.06 (40) 80 0.03 (27) 0.04 (50) 0.02 (13)
Chromium (VI) 40 0.06 (55) 0.07 (88) 0.12 (80) 80 0.05 (46) 0.02 (25) 0.07 (47)
160 0.04 (36) 0.01 (12) 0.03 (20)
Cadmium (II) 50 0.10 (91) 0,05 (63) 0.10 (67)
200 0.09 (82) 0.01 (12) 0.05 (33)
Lead (II) 80 0.11 (100) 0.09 (113) 0.15 (100)
160 0.10 (91) 0.07 (88) 0.12 (80)
Zinc (II) 50 0.11 (100) 0.07 (88) 0.07 (47)
200 0.10 (91) 0.03 (38) 0.03 (20)
a Data are the average of duplicate determinations; replicate data did not differ by more than 10% b Numbers in parenthesis represent the percentage of the control rate constant
biotransformation of butyrate were approximately three and greater than four times lower than for trans- formation of propionate and acetate, respectively (Table 3).
Effects of metal addition on methanogenesis
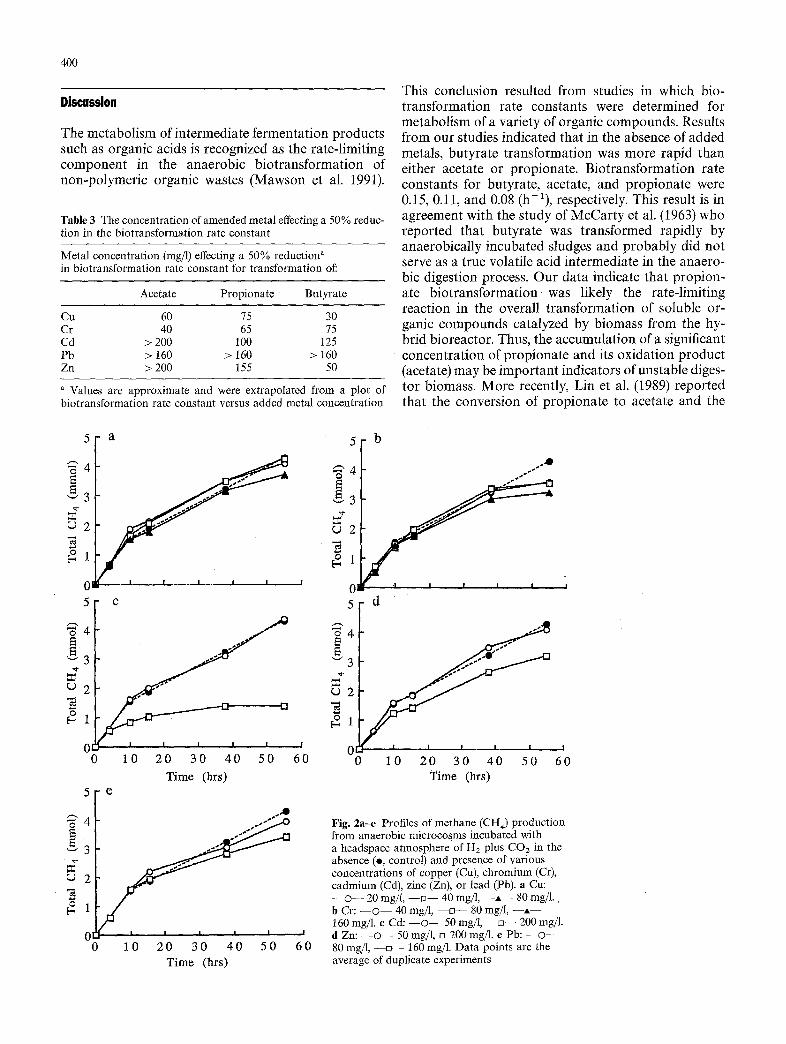
The profiles of hydrogenotrophic CH4 production from digestor biomass in the absence and presence of various concentrations of added metals are presented in Fig. 2a-e. Production of CH4 by the hydrogeno- trophic methanogenic population was strongly inhi- bited (approximately 70%) by 200 mgCd/1 (Fig. 2c). These results are in contrast to those observed for acetate biotransformation (plus CdC12) in which only minor inhibition (18%) was observed (Table 2). Addition of low concentrations of Cu (40 mg/1) and Cd (50mg/1) to H2 plus CO2 amended microcosms resulted in only a 6--10% reduction in total CH4 production. Addition of Cr(VI) at concentrations ranging from 40 to 160 rag/1 resulted in a 20-30% reduction in final CH4 production compared to con- trols. Furthermore, total CH4 production was inhibited only 6-30% in H2/CO2-amended microcosms byCu, Cr(VI), Pb, or Zn at the highest concentration tested (Fig. 2).
400
Discussion
The metabolism of intermediate fermentation products such as organic acids is recognized as the rate-limiting component in the anaerobic biotransformation of non-polymeric organic wastes (Mawson et al. 1991).
Table 3 The concentration of amended metal effecting a 50% reduc- tion in the biotransformation rate constant
Metal concentration (rag/l) effecting a 50% reduction" in biotransformation rate constant for transformation of:
Acetate Propionate Butyrate
Cu 60 75 30 Cr 40 65 75 Cd > 200 100 125 Pb > 160 > 160 > 160 Zn > 200 155 50
a Values are approximate and were extrapolated from a plot of biotransformation rate constant versus added metal concentration
This conclusion resulted from studies in which bio- transformation rate constants were determined for metabolism of a variety of organic compounds. Results from our studies indicated that in the absence of added metals, butyrate transformation was more rapid than either acetate or propionate. Biotransformation rate constants for butyrate, acetate, and propionate were 0.15, 0.11, and 0.08 (h-a), respectively. This result is in agreement with the study of McCarty et al. (1963) who reported that butyrate was transformed rapidly by anaerobically incubated sludges and probably did not serve as a true volatile acid intermediate in the anaero- bic digestion process. Our data indicate that propion- ate biotransformation, was likely the rate-limiting reaction in the overall transformation of soluble or- ganic compounds catalyzed by biomass from the hy- brid bioreactor. Thus, the accumulation of a significant concentration of propionate and its oxidation product (acetate) may be important indicators of unstable diges- tor biomass. More recently, L ine t al. (1989) reported that the conversion of propionate to acetate and the
5 a 5
4
" - ' 3 ~ 3
~ 2 ~ ) 2
2 1 " ~ 1
i I i i i 1
0
"~4
g3 ~ 9 2
O 1
0 0
~ 4 O
~ 3
2
1
c
I I I l I
10 2 0 3 0 4 0 5 0
Time (hrs) e
i l I ! I
10 2 0 3 0 4 0 5 0 Time (hrs)
5
3
¢ 9 2
21
0 6 0 0
!
60
b
,, '1 I I ! I
d
I I I, f l I
10 2 0 3 0 4 0 5 0 6 0 Time (hrs)
Fig. 2a--e Profiles of methane (CH4) production from anaerobic microcosms incubated with a headspace atmosphere of Hz plus CO2 in the absence (e, control) and presence of various concentrations of copper (Cu), chromium (Cr), cadmium (Cd), zinc (Zn), or lead (Pb). a Cu: - - © - - 20 mg/1, - - t~- - 40 rag/l, - - , , - - 80 rag/1. b Cr: - - © - - 40 mg/1, - - o - - 80 rag/l, - - A - - 160 mg/1. e Cd: - - o - - 50 mg/1, - - m - - 200 mg/l. d Zn: - - © - - 50 mg/l, t~ 200 mg/1. e Pb: - - O - - 80 mg/1, - - = - - 160 mg/1. Data points are the average of duplicate experiments

401
conversion of acetate to CH4 are likely the rate-limiting biotransformation processes in methanogenic diges- tors. However, the true rate-limiting step in natural and man-made anaerobic systems is difficult to predict be- cause of the presence of undefined, complex organic compounds as components of the digestion process.
In order to assess the potential impact of inorganic contaminants, we examined the toxicity of various heavy metals on the biotransformation of various organic acids using steady-state digestor biomass that is representative of naturally occurring microbial con- sortia (stable populations). In general, the inhibitory effects of metals on the biotransformation of organic acids increased with increased metal concentration. However, differences in the degree of inhibition were noted for different organic acid substrates and different metals (Tables 2 and 3). Our results indicated that the rate of biotransformation of butyrate was affected to a greater extent by the added metals than that of acetate. These results may be explained as follows: (a) the activity of butyrate-utilizing acetogens is more sus- ceptible to inhibition than that of acetate-utilizing methanogens; (b) the addition of metals inhibited the H2- scavenging population on which the butyrate-oxidizing population depends. Results indicated that the relative toxicity of the tested metals were Cr(VI) > Cu>>Cd Zn ~ Pb for acetate biotransformation, Cr(VI) Cu > Cd > Zn > Pb for propionate biotransforma- tion, and Cu > Cr(VI) ~ Zn > Cd>>Pb for butyrate biotransformation. In general, it was concluded that Cu and Cr(VI) were the most toxic metals tested and Pb was the least toxic for organic acid biotransformation. These results are in full or partial agreement with the results from previous studies (Hickey et al. 1989; Hayes and Theis 1978; Capone et al. 1983; Lin et al. 1989).
The mechanism of metal inhibition in biological sys- tems is not completely understood, but heavy metals are known to bind to thiols and other functional groups of enzymes and may also replace metals that are constituents of the active centers of enzymes, cofactors or other biomolecules (Vallee and Ulmer 1972). For methanogens, coenzymes such as the nickel-containing F43o, and other electron transport intermediates may be affected by the presence of metals. Pankhania and Robinson (1984) reported that the activities of both hydrogenase and formate dehydrogenase in whole cells of Methanospirillum hungatei strain GP1 were lower in experiments amended with 50 mM Hg, Zn, Cu, Cd, or Co.
In the present study, the activity of hydrogeno- trophic methanogens (Fig. 2) was not as adversely affected by the added metals as were the microbial groups responsible for organic acid biotransformation (Table 2). However, the lower inhibitory effects of metals on the bioconversion of H2/CO2 to CH4 may have been due, in part, to the slightly higher initial biomass (7.6 g TVS/1) used in those studies compared to metal-amended studies assessing organic acid biotrans-
formation (4.3 g TVS/1). An important consideration of heavy metal toxicity may be the quantity and type of particulate solids or biomass present in the environ- mental system. Although it is difficult to directly ex- trapolate laboratory results to field situations, laborat- ory investigations are important for identifying factors that directly affect biotransformation reactions. Lamb and Tollefson (1973) observed that the toxic effects of metals on overall digestor performance were reduced by increasing the concentration of sludge solids. Fur- thermore, Hickey et al. (1989) observed that higher total solid or volatile solid concentrations afforded better protection from metal inhibition, presumably through binding of the added metals by ligands. Gener- ally, the toxicity of metals in biological systems is controlled by metal speciation. For instance, heavy metals in the precipitated form are likely of little conse- quence with regard to inhibition of bacterial systems. Furthermore, low concentrations of metals in anaer- obic systems will likely be complexed as insoluble metal sulfides and a metal such as Cr may precipitate as the Cr(III)-hydroxide form (Babich et al. 1982; Richard and Bourg 1991). Precipitated forms are more likely to occur with Zn or Pb than with Cu or Cd. Inhibition due to heavy metals, thus, may be more accurately assessed if expressed as a function of biomass, or in some cases, as total or volatile solids (Hickey et al. 1989; Mosey and Hughes 1975). For these reasons, the concentrations of metals resulting in toxic or inhibitory effects should be defined with regard to the concentra- tion and characteristics of particulate solids and other chemical species (organic and inorganic).
References
APHA (1980) Standard methods for the examination of water and waste water, 15th edn. American Public Health Association, Washington, D.C.
Babich H, Schiffenbauer M, Stotzky G (1982) Comparative toxicity of trivalent and hexavalent chromium to fungi. Bull Environ Contain Toxicol 28:452-459
Balch WE, Fox GE, Magrum LJ, Woese CR, Wolfe RS (1979) Methanogens: reevaluation of a unique biological group. Micro- bial Rev 43:260-296
Capone DG, Reese DD, Kiene RP (1983) Effects of metals on methanogenesis, sulfate reduction, carbon dioxide evolution, and microbial biomass in anoxic salt marsh sediment. Appl Environ Microbiol 45:1586-1591
Chang JE (1989) Treatment of landfill leachate with an upflow anaerobic reactor combining a sludge bed and a filter. Water Sci Technol 21:133-143
Hayes TD, Theis TL (1978) The distribution of heavy metals in anaerobic digestion. J Water Pollut Control Fed 50:61-72
Hickey RF, Vanderwielen J, Switzenbaum MS (1989) The effect of heavy metals on methane production and hydrogen and carbon monoxide levels during batch anaerobic sludge digestion. Water Res 23:207-218
Jones WJ, Nagle DP, Whitman WB (1987) Methanogens and the diversity of archaebacteria. Microbiol Rev 51 : 135-177
Keith LH, Telliard WA (1979) Priority pollutants; a perspective view. Environ Sci Technol 13:416-423

402
Kitsos HM, Jones WJ, Roberts RS, Tornabene, TG (1993) Anaer- obic digestion of simulated distillery waste using floculated and fixed cell reactors, J Environ Sci Health 28:1099-1121
Kouzeli-Katsiri A, Kartsonas N, Priftis A (1988) Assessment of the toxicity of heavy metals to the anaerobic digestion of sewage sludge. Environ Technol Lett 9:261-270
Lamb A, Tollefson EL (1973) Toxic effects of cupric chromate and chromic ions on biological oxidation. Water Res 7:599-605
Levenspiel O (1972) Chemical reaction engineering. Wiley, New York Lin CY (1992) Effect of heavy metals on volatile fatty acids degrada-
tion in anaerobic digestion. Water Res 26:177-183 Lin CY, Sato K, Noike T, Matsumoto J (1989) Methanogenic
digestion using mixed substrate of acetic, propionic and butyric acids. Water Res 20:385-394
Mawson A J, Earle RL, Larsen VF (1991) Degradation of acetic acid and propionic acids in the methanogenic fermentation. Water Res 25:1549-1554
McCarty PL (1964) Anaerobic waste treatment fundamentals part I. Chemistry and microbiology. Public Works 95:107-112
McCarty PL, Jeris JS, Murdoch W (1963) Individual volatile acids in anaerobic treatment. J Water Pollut Control Fed 35:1501-1513
Mosey FE, Hughes DA (1975) The toxicity of heavy metal ions to anaerobic digestion. J Water Pollut Control Fed 74:18-38
Nriagu JO, Pacyna JM (1988) Quantitative assessment of worldwide contamination of air, water and soils by trace metals. Nature 333:134-139
Oleszkiewicz JA, Thadani VJ (1988) Effects of biofilter media on the performance of anaerobic hybrid reactors. Environ Technol Lett 9:89-100
Olthof M, Oleszkiewicz JA (1982) Anaerobic treatment of industrial wastewaters, Chem Eng 15:121-126
Pankhania IP, Robinson JP (1984) Heavy metal inhibition of meth- anogenesis by Methanospirillurn hungatei GP1. FEMS Microbiol Lett 22:277-281
Parkin GF, Owen WF (1986) Fundamentals of anaerobic digestion of wastewater sludges. J Environ Eng 112:867-901
Richard FC, Bourg ACM (1991) Aqueous geochemistry of chro- mium: a review. Water Res 25:807-816
Sterrit RM, Lester JN (1980) Interaction of heavy metals with bacteria. Sci Total Environ 14:6-17
Stronach SM, Rudd T, Lester JN (1986) Anaerobic digestion pro- cessed in industrial wastewater treatment. Springer, Berlin Heidelberg New York
Switzenbaum MS (1983) Anaerobic treatment of wastewater: recent developments. ASM News 49:532-536
Switzenbaum MS, Grady CPL (1986) Anaerobic treatment of domestic wastewater. J Water Pollut Control Fed 58:102-106
Vallee BL, Ulmer DD (1972) Biochemical effects of mercury, cad- mium, and lead. Annu Rev Biochem 41:91-128
Wood JM (1974) Biological systems for toxic elements in the envi- ronment. Science 183:1049-1052