mesenchymal derivative ofs the neural crest: analysi of...
TRANSCRIPT

/ . Embryol. exp. Morph. Vol. 34, 1, pp. 125-154, 1975 125
Printed in Great Britain
Mesenchymal derivatives ofthe neural crest: analysis of chimaeric quail and
chick embryos
By C. S. LE LIEVRE1 AND N. M. LE DOUARIN1
From the Laboratoire d'Embryologie, Universite de Nantes
SUMMARYInterspecific grafts of neural tube and associated neural crest (NC) have been made
between quail and chick embryos. Structural differences of the interphase nucleus in the twospecies make it possible to identify quail from chick cells in the chimaeras after Feulgen-Rossenbeck's staining and at the electron microscope level. Owing to the stability of thenatural quail nuclear marker labelling, migration pattern and developmental fate of thegrafted NC cells could be followed in the host embryo. In previous work it has been demon-strated that the visceral skeleton derives entirely from NC mesenchyme and the variouslevels of the neural axis from which visceral cartilages and bones originate have been estab-lished. In the present work, the contribution to the lower jaw and pharynx of NC mesen-chymal derivatives other than bones and cartilages has been studied. It is shown that thedermis in the face and ventrolateral side of the neck has a neural origin. The wall of thelarge arteries deriving from the branchial arches (systemic aorta, pulmonary arteries, brachio-cephalic trunks and common carotid arteries) are entirely made up of mesectodermal cellsexcept for the endothelial epithelium which is mesodermal in origin. The presence in thewall of the common carotid arteries of fiuorogenic monoamines-containing cells is demon-strated using the formol-induced-fiuorescence technique. Like the secretory cells of thecarotid body, the fluorescent cells of the carotid artery wall originate from the rhomb-encephalic NC.
Connective tissue of the lower jaw, tongue and ventrolateral part of the neck originatefrom the neural crest. Mesectoderm participate in the formation of the glands associatedwith the tongue and pharynx (lingual gland, thymus, thyroid, parathyroids) giving theirmesenchymal component. On the other hand, as demonstrated previously by our group,NC cells are the main cellular component of the UB since they give rise to the calcitonin-producing cells. The wall of the oesophagus and trachea is of mesodermal origin, but adiposetissue around the trachea and parasympathetic enteric ganglia of the digestive tube derivesfrom NC. NC cells participate in the formation of striated muscles of the branchial archesand differentiate there into connective and muscle cells.
It appears from this study that the differentiating capabilities are similar in mesenchymaland mesectodermal cells with the exception of blood vessel endothelia which in our experi-ments are always of host origin in mesectoderm-derived tissues.
The capacity of the NC to give rise to mesenchymal derivatives is restricted to thecephalic neural axis down to the level of the 5th somite in both chick and quail embryos.
1 Authors' address: Laboratoire d'Embryologie, Universite de Nantes, B.P. 1 044, 44037Nantes Cedex, France.

126 C. S. LE LIEVRE AND N. M. LE DOUARIN
INTRODUCTION
Katschenko (1888) was the first to suggest that some of the mesenchyme of thehead originates from the neural crest. He drew this assumption from the ob-servation of selacian development. Later, Goronowitsch (1892,1893) stated thatin teleosts and birds also the neural crest contributes to the formation ofmesenchyme. At the same time Platt (1893) found that in Necturus embryos thecartilage of the visceral arches as well as the dentine of the teeth derive fromectoderm, but she considered the main source of the mesenchyme that gives riseto these structures to be the lateral ectoderm of the head. She proposed the termof mesectoderm for the mesenchyme originating from the ectoderm and mesen-doderm for the mesodermal mesenchyme. The conclusions of Platt were basedon the fact that the yolk platelets of ectodermal cells were conspicuously smallerthan those of either mesoderm or endoderm, making possible the histologicaldistinction of migrating cells of the neural crest and placodal origin from othermesenchymal cells.
Experimental studies on the fate of neural crest cells were later undertaken byseveral investigators, mainly utilizing amphibian embryos. By extirpations andtransplantations Stone (1922, 1926, 1929) was able to demonstrate that theskeletal connective tissues of the upper face and visceral arches were largely ofneural crest origin.
Similar conclusions have been drawn by several authors using interspecificand intergeneric transplantation techniques (Raven, 1931; Holtfreter, 1933,1935a, b; Harrison, 1935, 1938; Andres, 1946, 1949; Wagner, 1949, 1955, 1959;and others: see reviews of Horstadius, 1950 and Weston 1970 for bibliography).Artificial cell marking techniques such as vital dyes (Stone, 1932; Horstadius &Sellman, 1946) and tritiated thymidine (Chibon, 1962,1964) were also applied tothis problem in amphibians.
Less numerous are the studies dealing with the contribution of the neuralcrest to mesenchymal structures in higher vertebrates. After extirpation experi-ments in chick embryo, Hammond & Yntema (1953, 1964) observed severedeficiencies of the nasal septum and visceral skeleton. Johnston (1966) using theexperimental technique of isotopic labelling of nuclei previously developed byWeston (1962, 1963) to study the migration of spinal neural crest cells in thechick embryo, carried out an autoradiographic study of the evolution of thecrest at the cranial level. He clearly showed that crest cells contribute extensivelyto the mesenchyme of upper facial regions and visceral arches and consequentlythat their behaviour is remarkably similar to that of their amphibian counter-parts. He was able to demonstrate that labelled crest cells participate in theformation of cartilage, but due to the instability of the labelling which becomesdiluted through the rapid proliferation of embryonic cells, the isotopic tech-nique cannot provide comprehensive data on the ectodermal mesenchymecapabilities.

Mesenchymal derivatives of avion neural crest 121
The present report deals with the application of a biological cell markingtechnique, which has the advantage of being stable (Le Douarin 1969, 19716,1973a), to the problem of neural crest cell migration and differentiation. Thusit is possible to follow the migrating crest cells until they have reached a fullydifferentiated state. Cell identification is based on structural differences of theinterphase nucleus in two closely related species of birds, the Japanese quail(Coturnix cotwnix japonicd) and the chick {Gallus gallus). In the quail, thenucleus contains one or several large heterochromatic masses associated with thenucleolar RNA, making the nucleolus considerably enlarged in all embryonicand adult cell types. In the chick the chromatin is evenly distributed in thenucleoplasm forming a fine network with some small dispersed chromocentersand the amount of nucleolar-associated chromatin is small. As a result of thisdifferent disposition of the chromatin material quail and chick cells can easilybe distinguished after the application of the Feulgen-Rossenbeck's specificstaining procedure for DNA. They can also be recognized at the electronmicroscope level after routine uranyl acetate-lead citrate staining or by meansof the EDTA preferential staining procedure for RNA according to Bernhard(1968), in which RNA is stained while DNA and most of the proteins areunstained (Le Douarin 1971 a, 1913a, 1973b).
The 'quail-chick marking system' has already been applied to the problem ofmigration and differentiation of neural crest cells by our group (Le Douarin &Le Lievre, 1970; Le Douarin & Teillet, 1970; Teillet & Le Douarin, 1970;Le Douarin, 1971 a, b; Le Douarin & Le Lievre, 1971; Le Douarin & Teillet,19716; Le Lievre, 1971a, b; Teillet, 1911a, b; Le Douarin & Le Lievre, 1972;Le Douarin, Le Lievre & Fontaine, 1972; Le Douarin, 1973a; Le Douarin &Teillet, 1973 a, b; Le Lievre & Le Douarin, 1973; Pearse et al. 1973; Le Douarin,Fontaine & Le Lievre, 1974; Le Lievre, 1974; Le Lievre & Le Douarin, 1974;Polak et al. 1974; Teillet & Le Douarin, 1974) and others (Johnston,Bhakdinaronk & Reid, 1973; Saxod, 1973). In previous papers it was not onlyshown that the entire visceral skeleton originates from mesectodermal cells, butit was also established from what levels of the neural axis the different bonesand cartilages are derived (Le Lievre, 19716, 1974).
The observations reported here concern the capability of neural crest togive rise, in addition to bone and cartilage, to other mesenchymal derivatives.
Due to the stability of the labelling provided by quail cells, it was possible toshow that the neural crest mesenchyme contributes to various tissues of face andneck.
MATERIAL AND METHODS
The eggs employed throughout this study were from the White Leghornstrain of the fowl Gallus gallus and the Japanese quail Coturnix coturnix japonica.They were incubated in a humidified atmosphere at 38 ± 1 °C. The developmentalstage was determined by the number of somites for the early stages until 2\ days
o EMB 34

128 C. S. LE LIEVRE AND N. M. LE DOUARIN
of incubation. After this, staging was done by the days of incubation. For thechick it was useful in some cases to use the staging series of Hamburger &Hamilton (1951).
Isotopic and isochronic grafts of sections of quail neural primordium into thechick have been carried out according to the following technique.
In a first step, a piece of neural tube and associated neural crest is surgicallyremoved at a prescribed transverse level of a chick embryo, between the anteriorlimit of mesencephalon to the level of the 9th somite inclusive. The length andlocation of the excision varies according to the experiment. The more cranialthe intervention the younger the embryo had to be, since the neural crestbecomes progressively established soon after the closure of the neural tube anddisperses in a cranio-caudal sequence.
In a second step the corresponding transverse part is taken from a quailembryo at the same developmental stage and immersed in a 0-1 % solution oftrypsin in Ca2+, Mg2+ free Tyrode solution (10 min at 2 °C) (Moscona &Moscona, 1952). All tissues adhering to the neural tube are manually removedwith fine dissecting needles. Thus the isolated quail neural rudiment is com-pletely devoid of contamination by non-neurectodermal cells (Fig. 1). Thetissue is then rinsed in Tyrode containing horse serum for 10 min and then inTyrode without horse serum for an additional 10 min. After the final rinse theneural tube is orthotopically grafted into the chick in the space resulting fromthe previously excised neural tube.
Four experimental series were made:(1) In 5- to 7-somite embryos the graft consisted of the mesencephalic pri-
mordium (Fig. 2, Expt. 1 a) or the level of the presumptive mesencephalon +anterior rhombencephalon back to the level of the 1st somite (Fig. 2, Expt. 1 b).
(2) In 6- to 10-somite embryos the intervention consisted of either the anteriorrhombencephalon (from the mesencephalo-rhombencephalic constriction to the1st somite (Fig. 2, Expt. 2 a), the posterior rhombencephalon from the 1st to the5th somite (Fig. 2, Expt. 2b), or the whole rhombencephalon (Fig. 2, Expt. 2c).
FIGURE 1
Experimental procedure for isotopic and isochronic transplantations of quail neuralprimordium into chick embryo. Ch, Notochord; NC, neural crest.(A) Transverse section of a 10-somite control embryo at the level of anteriorrhombencephalon. x 180.(B) Ten-somite chick embryo after removal of the rhombencephalic primordium.x 250.(C) Isolated rhombencephalic neural tube and neural folds of a 10-somite quailembryo, x 380.(D) Transverse section at the rhombencephalic level in a chick embryo which hasreceived, 6 h before fixation, the graft of a quail neural primordium. x 250.The same procedure is used for the reverse graft of chick neural primordium intoquail.

Mesenchymal derivatives of avian neural crest 129
. . . .
50 //m 1 B
9-2

130 C. S. LE LIEVRE AND N. M. LE DOUARIN
Exp. 1 Exp.2 Exp. 3
Fig. 2. Levels of heterospecific transplantations of neural rudiment.Expt. 1. The operations are carried out at 5- to 7-somite stage of host and donor
embryos at the level of mesencephalon (a) or mesencephalon + anterior rhomben-cephalon down to the 1st somite (b).
Expt. 2. The transplantations are carried out at 6- to 10-somite stage at the levelof anterior rhombencephalon (between mesencephalo-rhombencephalic constrictionand 1st somite) (a), posterior rhombencephalon (from the 1st to the 5th somites)(b) or concern the whole rhombencephalon (c).
Expt. 3. Isotopic and isochronic transplantations of short fragments of quailneural tube into chick in order to determine the posterior limit of NC capabilityto give rise to mesenchymal derivatives. Stage of intervention: 7—11 somite; graftof a fragment of neural tube corresponding to the length of 3 somites from the 1stto the 9th somites.
(3) In a third series of experiments short fragments of the chick neuralprimordium were removed and replaced by their quail counterparts at the levelof the first somite pairs in order to find out the location of the posterior limit tothe capabilities of the neural crest cells to give rise to mesenchymal derivatives.The embryos employed in these latter experiments were at 7- to 11-somitestages and the length of the neural axis involved in the experiment correspondedto the extent of three somites (Fig. 2, Expt. 3).
(4) In order to control the validity of the results obtained by grafting quailneural tube into chick, the reverse operation was carried out in an experimentalseries, i.e. heterospecific transplantation of rhombencephalic primordium.
Histological procedures
The operated embryos are sacrificed between 3 and 19 days of incubation.Up to the 11th day the anterior half of the body (head, neck and thorax down toand including the anterior limbs) is fixed in Zenker's fluid and cut in serial 5 /.imthick transverse sections.
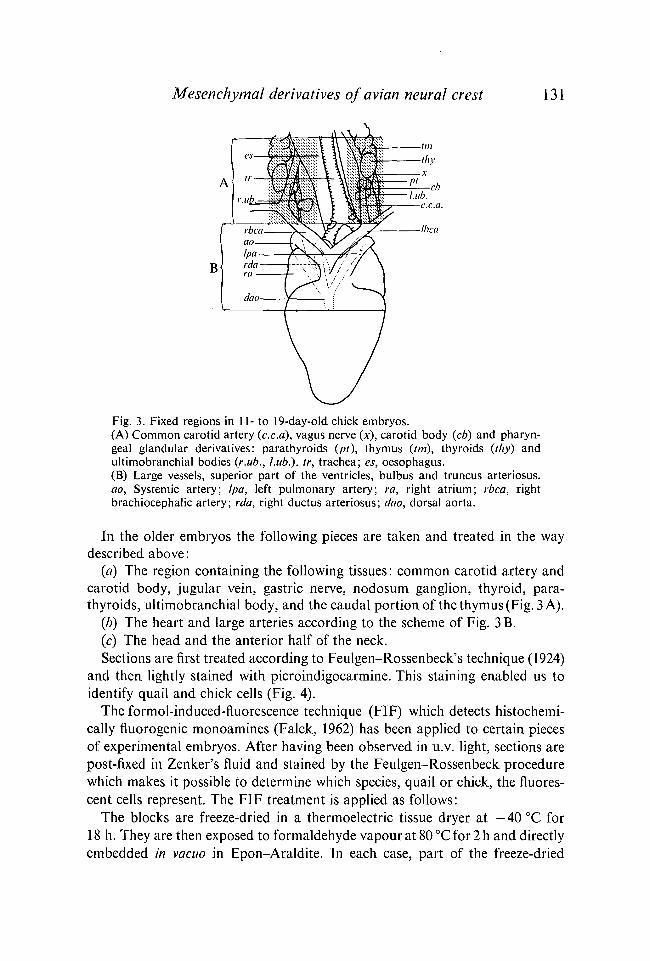
Mesenchymal derivatives of avian neural crest 131
Fig. 3. Fixed regions in 11- to 19-day-old chick embryos.(A) Common carotid artery (c.c.a), vagus nerve (x), carotid body (cb) and pharyn-geal glandular derivatives: parathyroids (pt), thymus (tm), thyroids (thy) andultimobranchial bodies (r.ub., hub.), tr, trachea; es, oesophagus.(B) Large vessels, superior part of the ventricles, bulbus and truncus arteriosus.ao, Systemic artery; /pa, left pulmonary artery; ra, right atrium; rbca, rightbrachiocephalic artery; rda, right ductus arteriosus; dao, dorsal aorta.
In the older embryos the following pieces are taken and treated in the waydescribed above:
(a) The region containing the following tissues: common carotid artery andcarotid body, jugular vein, gastric nerve, nodosum ganglion, thyroid, para-thyroids, ultimobranchial body, and the caudal portion of the thymus (Fig. 3 A).
(b) The heart and large arteries according to the scheme of Fig. 3B.(c) The head and the anterior half of the neck.Sections are first treated according to Feulgen-Rossenbeck's technique (1924)
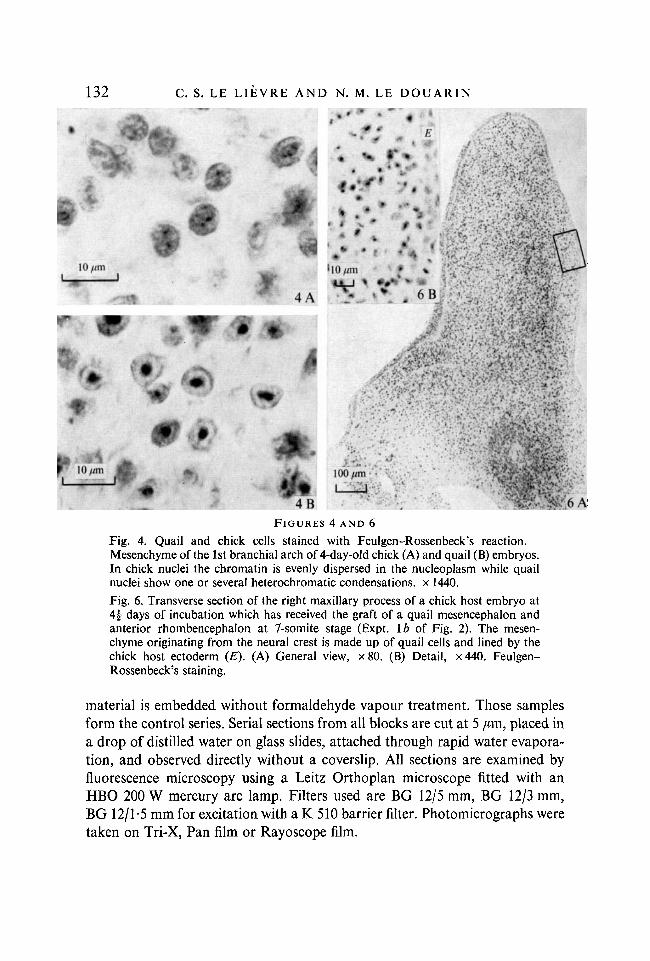
and then lightly stained with picroindigocarmine. This staining enabled us toidentify quail and chick cells (Fig. 4).
The formol-induced-fluorescence technique (FIF) which detects histochemi-cally fluorogenic monoamines (Falck, 1962) has been applied to certain piecesof experimental embryos. After having been observed in u.v. light, sections arepost-fixed in Zenker's fluid and stained by the Feulgen-Rossenbeck procedurewhich makes it possible to determine which species, quail or chick, the fluores-cent cells represent. The FIF treatment is applied as follows:
The blocks are freeze-dried in a thermoelectric tissue dryer at - 4 0 °C for18 h. They are then exposed to formaldehyde vapour at 80 °C for 2 h and directlyembedded in vacuo in Epon-Araldite. In each case, part of the freeze-dried
132 C. S. LE LIEVRE AND N. M. LE DOUARIN
6A
FIGURES 4 AND 6Fig. 4. Quail and chick cells stained with Feulgen-Rossenbeck's reaction.Mesenchyme of the 1st branchial arch of 4-day-old chick (A) and quail (B) embryos.In chick nuclei the chromatin is evenly dispersed in the nucleoplasm while quailnuclei show one or several heterochromatic condensations, x 1440.
Fig. 6. Transverse section of the right maxillary process of a chick host embryo at4 | days of incubation which has received the graft of a quail mesencephalon andanterior rhombencephalon at 7-somite stage (Expt. \b of Fig. 2). The mesen-chyme originating from the neural crest is made up of quail cells and lined by thechick host ectoderm (E). (A) General view, x 80. (B) Detail, x 440. Feulgen-Rossenbeck's staining.
material is embedded without formaldehyde vapour treatment. Those samplesform the control series. Serial sections from all blocks are cut at 5 ju>m, placed ina drop of distilled water on glass slides, attached through rapid water evapora-tion, and observed directly without a coverslip. All sections are examined byfluorescence microscopy using a Leitz Orthoplan microscope fitted with anHBO 200 W mercury arc lamp. Filters used are BG 12/5 mm, BG 12/3 mm,BG 12/1-5 mm for excitation with a K 510 barrier filter. Photomicrographs weretaken on Tri-X, Pan film or Rayoscope film.

Mesenchymal derivatives of avian neural crest 133
B
Fig. 5. Diagrammatic representation of the extension of mesencephalic andrhombencephalic NC cells in facial and pharyngeal structures of 4-day-old embryos.(A) Lateral view. 1-4: 1st, 2nd, 3rd and 4th branchial arches. 6, 7: Level of thecross-sections illustrated in Figs. 6 and 7. mp, Maxillary process; h, heart.(B) Cross-section through the 2nd branchial arch, ao, Dorsal aorta and 2nd aorticarch; ch, notochord; j , anterior cardinal vein; m, muscle plate; ph, pharynx. NCcells are located ventrally (stippled area) and extend dorsally to the level of anteriorcardinal vein. The muscle plate is essentially of host origin.
Electron microscopy
For ultrastructural observation tissues are fixed in 6% glutaraldehyde in01 M phosphate buffer, at pH 7-4 for 20 min at 4 °C, and postfixed in 1 %osmium tetroxide in phosphate buffer for 1 h. The blocks are embedded inEpon, sectioned, stained by lead citrate and uranyl acetate, and observed in anHitachi HS 8 electron microscope.
RESULTS
I. Observation of the host embryos at 3rd and 4th days of incubation
Transverse sections (5 jim) of the anterior part of the embryos were stainedwith the Feulgen-Rossenbeck's technique. The neural crest [NC] cells, whichcontribute to visceral arch morphogenesis, could be recognized as a result of thequail nuclear marker.
The results obtained after the graft of mesencephalic and rhombencephalicprimordia (experimental series represented Fig. 2, Expts. 1 and 2) are summarizedin Table 1.
It appears that the mesenchyme of maxillary buds and branchial arches iscomposed primarily of mesectodermal cells (Figs. 5-7). Chick cells were found

OJ
Tab
le 1
. Par
tici
pati
on
of m
esec
tode
rmal
ce
lls
in t
he g
enes
is o
f fac
ial
and
phar
ynge
al s
truc
ture
s in
a4-
day-
old
chic
k em
bryo
^"""
^--^
App
eara
nce
of h
eter
ospe
cific
Leve
l of
^""
""^-
^^
mes
ecto
derm
alhe
tero
spec
ific
^
^^
ce
lls
in th
egr
aft
^""
"^^
ho
st
Exp
. 1
a.M
esen
ceph
alon
Exp
. 1
h.M
esen
ceph
alon
+ a
nter
ior
part
of
rhom
benc
epha
lon
Exp
. 2a
ante
rior
par
t of
rhom
benc
epha
lon
Exp
. 2b
.Po
ster
ior
part
of
rhom
benc
epha
lon
Exp
. 2 c
.T
otal
rhom
benc
epha
lon
Max
illar
ypr
oces
s1s
tbr
anch
ial
arch
2nd
bran
chia
lar
ch
3rd
bran
chia
lar
ch
4th
bran
chia
lan
d po
ster
ior
to4t
h po
uch
p t-1 w tfl d % r tn d o c!

Mesenchymal derivatives ofavian neural crest 135
forming the endothelium of the aortic arch and the muscle plate, in which,however, some quail cells were dispersed.
Similar observations were done whatever the sense of heterospecific combina-tions was: quail neural tube grafted into chick or inversely. Figs. 7 and 8 showthe distribution of host and donor cells in the 2nd visceral arch in the two kindsof grafts at rhombencephalic level.
II. Further localization of differentiated mesectodermal cells in facial andpharyngeal structures
Due to the stability of the nuclear marker provided by quail cells, the ultimatelocalization of neurectodermal cells in the various structures of the head and neckcould be determined. From the 6th day onwards, the contribution of quail cellsto the visceral arch derivatives has been studied in chick embryos which hadreceived various grafts as indicated in Fig. 2.
Mesectodermal cells which colonize the branchial arches give rise to theentire visceral skeleton as previously reported by Le Lievre (1974). In additionquail NC cells have been found to differentiate into many other kinds of cells.
A. Contribution of NC cells to the dermis
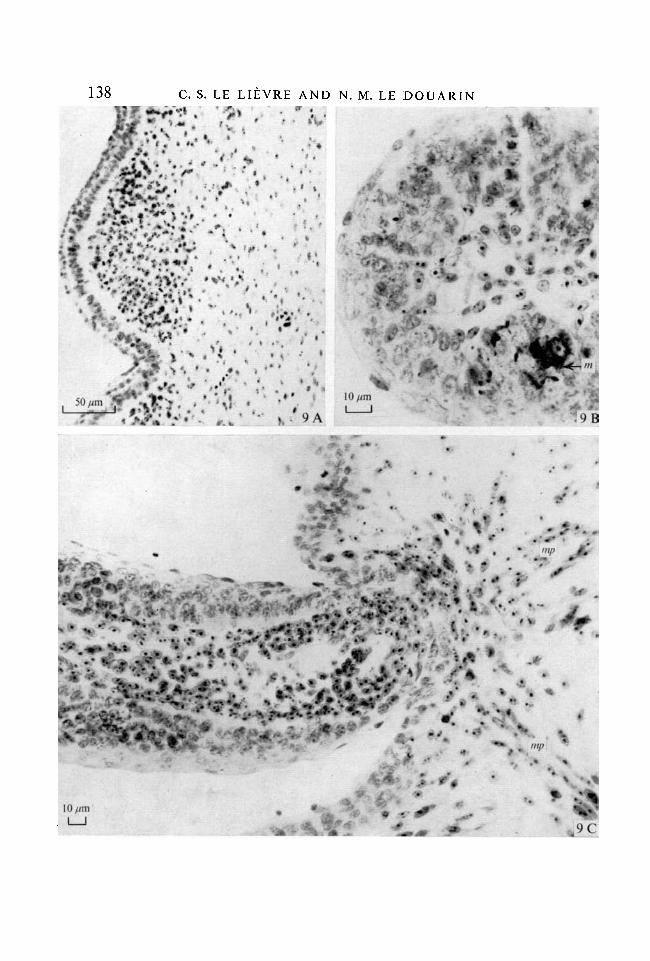
In the face and neck, mesectodermal cells differentiate into dermis. Thisresults in the formation of chimaeric feather buds in which the papilla is madeup of quail cells while the epidermis belongs to the host chick (Fig. 9 A). Quailmelanocytes spread dorso-ventrally and cranio-caudally from the grafted neuraltube in the host skin (Teillet & Le Douarin 1970; Teillet 1971a). They arenumerous inside the chick epidermis of the chimaeric feathers (Fig. 9B). Quailmesectodermal cells form the smooth arrector muscles associated with feathers(Fig. 9C) and the subcutaneous adipose tissue.
B. Connective tissues deriving from the NC
The connective tissue of the lower jaw, the tongue and the ventral part of theneck derives from NC mesenchyme. The dorsal limits of the mesectodermalarea reach the level of the auditory pit and the internal carotid arteries. Mixedwith chick host cells, mesectoderm participates in the histogenesis of the wallof these vessels. No quail cells are encountered dorsally to the notochord.
The loose connective tissue in the tongue and the floor of the mouth is mostlyderived from the NC. Such is the mesenchymal component of salivary glands(Fig. 10) and the connective stroma in the tongue and lower jaw muscles.
The mesenchymal components of the glandular pharyngeal derivatives are ofNC origin. Quail cells are found in the interlobular spaces and in the medulla ofthymic lobes, forming the parafollicular cells of thyroid gland and the connectivetissue located between the cords of parathyroid glandular cells (Fig. 11). Theyare the main cellular component of the ultimobranchial and carotid bodies aspreviously described by our group (Le Douarin & Le Lievre, 1970, 1971, 1972;

136 C. S. LE LIEVRE AND N. M. LE DOUARIN
8B

Mesenchymal derivatives of avion neural crest 137
Le Douarin et al. 1972; Pearse et al. 1973; Le Douarin et al. 1974; Polak et al.1974).
The walls of the oesophagus and trachea are primarily derived from chick hostcells with the exception of the enteric parasympathetic ganglia which arise fromthe grafted NC when the operation involves the hind brain level, as reportedpreviously (Le Douarin & Teillet, 19716, 1973a). During the second half of theincubation period, mesectodermal cells differentiate into adipose tissue in thevicinity of trachea (Fig. 12).
C. Participation of NC cells in striated muscle differentiation
Mesectodermal cells participate to a certain extent in the histogenesis of thestriated muscles of the tongue, the face and the neck though they mainly derivefrom anterior somite mesoderm (Johnston, personal communication). How-ever, as mentioned above, some quail cells are found in the muscle plates of thebranchial arches and are incorporated afterwards in the muscles (Fig. 13).
Striated muscles, which differentiate in the second branchial arch, have beenobserved at the electron microscope level and were seen to be a mixture of quailand chick striated muscle cells (Fig. 14) with a prevalence of host cells. On theother hand, as mentioned above, the muscle connective tissue derives from NCmesenchyme.
D. Mesectodermal origin of the arteries deriving from aortic arches
Due to the elongation of the neck and the anteroposterior movement of theheart during the first half of embryogenesis, the blood vessels which derive fromaortic arches are drawn caudad and incorporated into the thoracic cavity. Ex-cept for the endothelium, the wall of the large arteries deriving from the aorticarches originates entirely from mesectoderm.
When the graft of a quail neural primordium into a chick embryo involves
FIGURES 7 AND 8
Fig. 7. Graft of anterior rhombencephalon of a quail into a chick embryo at7-somite stage (Expt. 2a). Cross-section through the 2nd branchial arch at 4£ daysof incubation. Feulgen-Rossenbeck's staining.(A) General view, x 90.(B) Detail showing that the mesenchyme originates from the grafted quail neuraltissue (Q). The muscle plate (M) is essentially of host origin but contains somedispersed quail cells. Ectoderm (£) and pharyngeal endoderm (En) are made up ofchick cells. x280.Fig. 8. Similar experiment as in Fig. 7: chick rhombencephalon is grafted into a7-somite quail embryo. As in Fig. 7, most of branchial arch mesenchyme is derivedfrom the graft while the muscle plate and the lining ectodermal and endodermalepithelia are of host origin. Feulgen-Rossenbeck's staining.(A) General view, x 220.(B) Detail, x 580.
138 C. S. LE LIEYRE AND N. M. LE DOUARIN
10 ftm
I I

Mesenchymal derivatives of avian neural crest 139
the whole rhombencephalic primordium (Fig. 2, Expt. 2 c) the wall of the arterieswhich arise from the 3rd (brachiocephalic trunks and common carotid arteries),the 4th (systemic aorta) and the 6th (pulmonary arteries) aortic arches, areentirely made up of quail cells (Figs. 15, 16). In a transition zone between thebulbus arteriosus and the aortic trunks arising from the heart, the vessel wall isformed by a mixture of quail and host cells. The same observation is made in thedistal part of the arch of systemic aorta and in ductus arteriosus correspondingto the distal portion of the 6th arches. After the 6th day of incubation, quailmesenchymal cells become distinctly arranged as smooth muscle around thetube formed by chick endothelial cells lining the aortic arch primordium. Thusthe NC cells undergo smooth muscle cell differentiation and elaborate elastinfibrils. The chromatin pattern of quail nuclei is modified during this differentia-tive process. The initial single heterochromatic mass of the nucleus becomesfragmented in two or three smaller Feulgen-positive patches attached to thenuclear membrane (Fig. 16 A). The same disposition is observed in vessel walls ofcontrol quail embryos.
The 3rd visceral arch is the site of differentiation of the carotid body in theclose vicinity of the common carotid artery wall as described by Fontaine (1973)in both quail and chick embryos. The common carotid artery of experimentalembryos (i.e. chick embryos which had received the graft of quail neuralprimordium) was sectioned and treated by FIF technique during the secondhalf of the incubation time. Numerous single or grouped fluorescent cells wereobserved randomly distributed throughout its wall (Fig. 16 B). They show thesame greenish fluorescence as do quail carotid body cells, which has beendemonstrated to be due to dopamine (Pearse et al. 1973). If the sections firstobserved in u.v. light are then post-stained by Feulgen-Rossenbeck's technique,fluorescent cells are identified as belonging to quail species, as are the connectivewall of the artery and the carotid body (Le Douarin et al. 1972).
E. Do NC cells participate in endothelial wall of blood vessels?
Various pharyngeal structures in which the connective tissue derive from themesectoderm were observed at the electron microscope level in order to findout whether the endothelial cells of the blood capillaries belonged to mesodermalor neurectodermal mesenchyme. In all the cases observed (dermis, thymus,
Fig. 9. Experiment \b. Graft of a quail mesencephalon + anterior rhombencephaloninto 6-somite chick embryos.(A) Transverse section in the neck of the host at 9 days. Feather-bud showing hostectoderm and quail mesenchyme. x 280.(B) Feather germ of a 12-day-old host. Quail melanocytes (ra) have invaded thehost ectoderm. Germ pulp is made up of quail cells, x 650.(C) Longitudinal section of a feather germ in the neck of a 12-day-old host chickembryo. Arrector feather muscles in the dermis, made up of quail mesectodermalcells (mp). x 450. Feulgen-Rossenbeck's staining.

140 C. S. LE LIEVRE AND N. M. LE DOUARIN
13

Mesenchymal derivatives of avian neural crest 141
thyroid) none of the endothelial cells have been found to exhibit the quailnuclear marker. The pericytes, however, which line the external side of thecapillaries, are evidently of quail origin (Fig. 17). Therefore, it must be assumedthat the organs deriving from mesectodermal rudiments are invaded by meso-dermal capillary buds during their histogenesis.
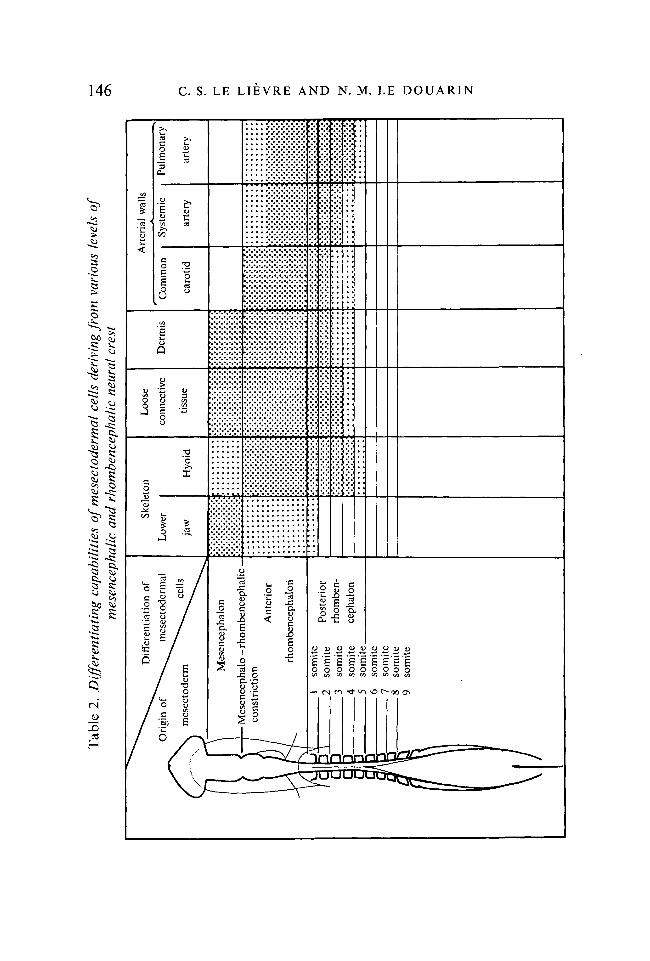
Table 2 and Fig. 18 summarize the observations described above, i.e. thedistribution of NC cells in the various structures of the face and neck accordingto the level at which the graft of the quail primordium has been made.
III. Posterior limit of mesenchymal potentialities of the NC
Grafts of short fragments of quail neural primordium have been madein chick according to the schema of Fig. 2, Expt. 3. Ninety embryoshave been operated and 39 survived and could be observed at 6-12 days ofincubation.
When the graft is made behind the level of the 5th somite, no mesenchymalcells are found in the host embryo. Some dispersed quail cells are encountered inthe 6th aortic arches following a graft involving the levels of 3rd and 4thsomites. In the latter case no quail cells are found in connective tissue of the host,neither in the dermis. Cells originating from the level of somite 1-3 give rise tothe various kinds of connective cells described above. Thus one can considerthat the posterior limit of the mesenchymal capabilities of the crest correspondsroughly to the level of the 5th somite.
In the kind of experiments involving the graft of short fragments of quailneural tube into chick, quail cells are always found mixed to host cells. Thatshows that the same transverse level of the pharynx receives NC cells from alarge portion of the neural axis. Thus it appears necessary to perform extensive
FIGURES 10-13
Fig. 10. Experiment 1 b. Graft of a quail mesencephalon + anterior rhombencephaloninto a 7-somite chick embryo. Section through the body of the tongue of the 14-day-old chick embryo. All mesenchymal derivatives (Q) originate from the graftedneural quail primordium: loose connective tissue (I.e.); pulp of horny papilla (h.p);lingual gland mesenchyme (l.g.); keratinized epithelium (E) and lingual glandularcells are of host origin, x 300. Feulgen-Rossenbeck's staining.Fig. 11. Experiment 2c. Graft of a quail rhombencephalon into a 10-somite chickembryo. Parathyroid of the 8-day-old chick host embryo. Numerous quail mesen-chymal cells (Q) are present between the chick glandular cords (pt). x 510. Feulgen-Rossenbeck's staining.Fig. 12. Experiment 2 c. Graft of a quail rhombencephalon into a 7-somite chickembryo. In the neck of the 12-day-old chick host, quail mesectodermal cells havedifferentiated into adipose cells (A), x 920. Feulgen-Rossenbeck's staining.Fig. 13. Experiment 2c. Graft of a quail rhombencephalon into a 10-somite chickembryo. Branchiomandibularis muscle (3rd branchial arch) of the 10-day-old hostchick embryo. The muscle is made up of a mixture of host and quail cells, x 920.Feulgen-Rossenbeck's staining.

142 C. S. LE LIEVRE AND N. M. LE DOUARIN
Fig. 14. Same experiment as in Fig. 13. Striated muscle cell showing acharacteristic DNA-rich nucleolus of quail, x 27500.
heterospecific transplantation of the neural tube to get a clear view of the normaldistribution and developmental capabilities of crest cells.
DISCUSSION
Interspecific combinations between quail and chick embryos have made itpossible to investigate the normal process of NC cell migration and to follow theNC derivatives until their completely differentiated state. Unknown derivativesof this transitory embryonic structure have thus been identified and the extentof its contribution to cephalic and cervical morphogenesis has been recognized.
1. Reliability of the experimental technique
Experimental techniques devised to investigate embryogenic mapping mustdisturb as little as possible the normal developmental conditions of the rudi-ments. Transplantations of early primordia on the chorioallantois or into thecoelomic cavity have often been used (Willier & Rawles, 1931, Rudnick 1932,1938; Andrew, 1964, 1969, 1970) but they disturb the micro-environment of theembryonic tissues and therefore cannot provide really reliable information ontheir normal developmental fate during embryogenesis. That is especially truewhen the cells concerned undergo extensive migrations in the embryo beforedifferentiating. As demonstrated for NC cells (Weston, 1963; Weston & Butler,

Mesenchymal derivatives ofavian neural crest 143
CCA
SCA
Fig. .15. Aortic-arch-derived arteries. /////, the vessel wall is entirely made
up of mesectodermal cells originating from mesencephalic and rhombencephalic
NC, except for the endothelium which is of mesodermal origin. ,
mesectodermal and mesodermal cells are mixed. | |, the vessel wall is madeup of mesodermal cells in entirety. 3-6, Arteries deriving from the 3rd, 4thand 6th aortic arches; CCA, common carotid artery; DA, dorsal aorta; PA, pul-monary artery; SCA, subclavian artery; TA, aortic trunk; TP, pulmonary trunk.
1966; Le Douarin & Teillet, 1974), migration pathways are indeed highlydependent on the structures and substrates encountered.
Isotopic and isochronic grafts of neural primordium between two species ofbirds closely related in taxonomy provide almost normal developmental condi-tions for both host and grafted structures and result in normally developedembryos, when the operation has been properly done. Compared to extirpationor electrocauterization of the neural tube (see Horstadius, 1950, and Weston,1970, for review), this grafting technique has the advantage of precluding regu-latory mechanisms due to crest cells from anterior and posterior levels.
The developmental rate being a little faster in the quail than in the chickembryo, it was important to perform neural tube exchanges between the twospecies in both directions, i.e. quail into chick and chick into quail and to com-pare the distribution of NC cells in the different combinations. When the quailneural tube is grafted into the chick embryo, the rapidly growing quail cellsmight indeed have an advantage on the chick cells and expand more in the hoststructures than in normal conditions. However, comparison of the embryos showa similar pattern of NC cell migration in both kinds of grafts.
In fact, the difference of growth rate is rather slight in the early developmental10 EMB 34

144 C. S. LE LIEVRE AND N. M. LE DOUARIN

Mesenchymal derivatives of avian neural crest 145
stages when the most significant morphogenetic events occur and becomes signi-ficant only during the second half of the incubation period (Hamburger &Hamilton 1951; Zacchei 1961). Since chick embryos are more resistant thanquail to surgical interventions, most of the results reported in this study havebeen obtained on chick hosts grafted with quail NC.
It can be assumed that, in experimental embryos, the structures composed ofcells with the nuclear characteristics of the grafted primordium are of neuralorigin. Numerous histological controls of the neural tube before the graft andafter trypsination have shown that the neural rudiment is easily isolated fromneighbouring tissues at the early developmental stages and is, at grafting,completely devoid of mesenchymal cell contamination.
2. Differentiating capabilities of mesectodermal cells, and role of inductiveinfluences
Studies on chimaeric quail-chick embryos lead to the conclusion that theneural primordium contributes extensively to structures of the facial andpharyngeal regions. Though the interspecific grafts included the whole neuraltube rather than the neural folds alone, it is reasonable to assume that most ifnot all mesectodermal derivatives observed in the experimental embryos origi-nate from the NC. Some results obtained previously by Johnston (1966), usingtritiated thymidine-labelled transplants of the NC alone, are in complete agree-ment with our findings.
Thus it appears that most of the lateroventral part of head and neck is edifiedby material originating from the mediodorsat embryonic region, which under-goes extensive migration before reaching its definitive site in the face andbranchial arches.
The results obtained here demonstrate that ectodermal mesenchyme has alarge range of developmental capabilities and appears able to give rise to the
FIGURES 16 AND 17
Fig. 16. Experiment 2c. Graft of a quail rhombencephalic primordium on a10-somite chick embryo.(A) Arterial trunk. The endothelium (E) belongs to the host while the musculo-connective tissue of the vessel wall is derived from the quail grafted tissue, x 780.Feulgen-Rossenbeck's staining.(B) FIF technique applied to the area represented in Fig. 3 A of a 11-day-oldembryo. Fluorogenic monoamine-containing cells in the wall of the commoncarotid artery, as carotid body cells, are derived from rhombencephalic NC. x 140.Fig. 17. Capillaries in the dermis, connective cells are of NC origin, endothelial(E) cells belong to the host.(A) Experiment \b. Graft of quail mesencephalon + anterior rhombencephaloninto a 6-somite chick embryo. Chick host was fixed after 12 days of incubation,x 1280. Feulgen-Rossenbeck's staining.(B) Experiment 2c. Graft of a quail rhombencephalic primordium into a 7-somitechick embryo. Chick host was fixed after 16 days of incubation. P, Pericyte.x 11600. Uranyl acetate-lead citrate staining.
Tab
le 2
. D
iffe
rent
iati
ng
capa
bili
ties
of
mes
ecto
derm
al c
ells
der
ivin
g fr
om
vari
ous
leve
ls o
fm
esen
ceph
alic
and
rho
mbe
ncep
hali
c ne
ural
cre
st
( —**
Q
—^^
^ D
iffe
rent
iati
on
of
Ori
gin
of
-\
^ m
esec
tode
rmal
Low
>v
mes
ecto
derm
^
*»
»^
ce
lls
^
I \
Mes
ence
phal
on
>:: :-
:':: :
1 1
co
nstr
icti
on
;;;;
;;
J I /
Ant
erio
r ::
::::
I^L
rh
ombe
ncep
halo
ri"K
:::
:::L
JV
I
som
ite
r^ \
-\
Pos
teri
orU
J 1
2
som
ite
ruai
cuu
i•
1
som
ite
rhom
ben-
U
4
cnm
itp
ceph
alon
U
S
snm
itp
y 6
so
mit
eyt
J 7
so
mit
e\W
8
so
mit
e
Ske
leto
n
/er
v H
yoid
*"v
.*I*
I
Loo
se
conn
ecti
ve
tiss
ue MI! &
£:
Der
mis
Art
eria
l w
alls
Com
mon
Sy
ste
caro
tid
art
'**"
•" *
****
•***
*"! *
!*•*
• *•
*•*•
*•*
I*
, .•
.%•.
•.•.
mic
P
ulm
onar
y
ery
arte
ry
.: : :
: :.:
:.: :
.:.: :
.:.:.:
.:.:.:
.
*.*
f.'i*t*
. I'I
'I'I
'IV
I'.
'I'I
V*

Mesenchymal derivatives of avian neural crest 147same differentiated cell types as mesodermal mesenchyme: general and dif-ferentiated (elastic and fibrous) connective tissues, adipose cells, dermis,smooth and striated muscles, bone and cartilage. In addition mesectodermal cellsare able to differentiate into odontoblasts (Horstadius & Sellman, 1946;Sellman, 1946; De Beer, 1947; Horstadius, 1950; Wagner, 1955; Signoret, 1960;Koch, 1965; Chibon, 1966). However, the only developmental competencywhich neurectodermal mesenchyme does not seem to possess is the formationof endothelium of the blood vessels. In the branchial and facial regions ofexperimental embryos, the vascular endothelia are always host in type, and arethus of mesodermal origin, NC cells participating only in the formation of thevessel wall: i.e. pericytes of capillaries and musculo-connective sheath of largevessels.
During their migration and in their final localization close relationships areestablished between NC cells and various mesodermal, endodermal and ecto-dermal rudiments. The extent to which intertissular inductive reactions betweenneurectodermal cells and those tissues control their subsequent differentiation isnot clear. Until now, the normal fate of the NC in the cephalocervical regionbeing only partially known, this question could not be fully investigated.
Owing to the stability of the quail nuclear marker labelling, the exclusivelyneurectodermal origin of the visceral arch cartilages and membrane bones of thelower jaw is demonstrated. A number of investigators (Horstadius, 1950;Horstadius & Sellman, 1946; Newth, 1954; Holtfreter, 1968) have shown thatchondrogenesis is induced in NC mesenchyme by pharyngeal endoderm. Theheterotopic transplantations of cephalic neural tube into the trunk, recentlycarried out in our laboratory (Le Douarin & Teillet, 1974) show that, in avianembryos, inductive influences from other sources can play the same role:cephalic NC cells grafted into the trunk and thus submitted to the rachismorphogenetic field contribute to vertebral chondrogenesis.
Intimate relationships are established between mesectodermal and mesodermalcells in striated muscle differentiation in branchial arches. The mesodermal coresof each visceral arch derive from somitic mesenchyme (Johnston & Listgarten,1973). From the earliest stages following completion of their migration, crestcells invade this mesodermal core and become intermingled with the somitederiving cells essentially concerned with the formation of skeletal muscle. Theultrastructural study shows that crest cells give rise not only to the connectivetissue elements of muscles but also to striated muscle cells. Mauro (1961), Moss& Leblond (1971) and others have suggested that the growth process of musclefibres involves the addition to the primary myotube of satellite cells. WhetherNC cells play the role of satellite cells in muscles is a hypothesis which would beworth studying.
NC cell differentiating capabilities include secretory elements such as adreno-medullary (Le Douarin et Teillet, 1971 a), calcitonin producing cells (Le Douarin& Le Lievre, 1970) and carotid body glandular cells.

148 C. S. LE LIEVRE AND N. M. LE DOUARIN
hv
Fig. 18. For legend see opposite.

Mesenchymal derivatives of avian neural crest 149
In previous work indeed we have demonstrated the neural crest origin of thecarotid body (CB) the cells of which are characterized by a high content offluorogenic monoamine (Le Douarin et al. 1972; Pearse et al. 1973). Thisglandular structure differentiates in the 3rd branchial arch, in close associationwith the common carotid artery. Among the connective cells which form thevessel wall, some fluorogenic cells of neural origin are present. The nature of themonoamine they contain seems to be the same as that of CB cells: in the chickit is serotonin, with a yellow-greenish fluorescence; in the quail, dopamine, witha greenish fluorescence (Pearse et al. 1973). In chick embryos grafted with aquail rhombencephalon the monoamine containing cells of CB and commoncarotid artery which exhibit the quail nuclear marker after Feulgen-Rossen-beck's staining have a greenish fluorescence, like the homologous structures ofcontrol quail embryos.
3. Extension of NC cell migration in head and neck
In higher vertebrates the competency of neurectoderm to give rise to mesen-chyme is restricted to the cephalic region of the neural axis, while in lower formsmesectoderm also derives from trunk NC (cf. Horstadius, 1950). From ourobservations and those of Johnston et al. (1973) based on the use of the quailchick marker system, it becomes evident that the mesectoderm is quantitativelya more extensive component of cephalocervical morphogenesis than has pre-
F I G U R E 18
Distribution of NC mesenchymal derivatives in the lower jaw and neck of 9-day-oldchick embryo following the transplantation of quail mesencephalon and rhomben-cephalon into the chick host.
(I) Distribution of quail cells in the dermis and inner organs of the chick shown
in lateral and ventral views. *+*+*, Mesencephalic NC derivatives. • • • • ,** w • • • •
Rhombencephalic NC derivatives. A, B, C, Levels of the sections which are repre-sented in figures IIA, B and C. cub, ultimobranchial body; hy, hyoid; thym, thymus;thy, thyroid.(II) Transverse sections at three different levels of the host showing the distributionof NC cells.
(A) Lower jaw and tongue. Cells of mesencephalic origin * ^ * ^ * form {me)
Meckel's cartilage and membrane bones (mb) (angular, supra-angular dentary and
operculary); cells of rhombencephalic origin form the hyoid apparatus(bb, basibranchial; bh, basihyal; cb, ceratobranchials; eb, epibranchials; andentoglossum) and both differentiate into loose connective tissue, dermis and musclecells; ch, notochord; es, oesophagus; ic, internal carotid. (B) Neck. (C) Levelof carotid bodies.
^ ^ ^ H , Tissues entirely made up of quail cells: d, dermis; c, wall of carotid
arteries; ge, enteric ganglia; cb, carotid body. • • • * » Tissues made up of a
mixture of NC and mesodermal cells: cj, loose connective tissues; x, Schwann cellsof the vagus nerve; gn, glial cells of the nodosum ganglion; connective tissue of thy-roid ; p, parathyroid; tm, thymus.
j , Jugular vein; m, tracheohyoideus and tracheolateralis muscles; tr, trachea.

150 C. S. LE LIEVRE AND N. M. LE DOUARIN
viously been estimated. Due to the elongation of the neck, cells originating fromencephalic NC are carried down into the thorax with the heart and the glandularpharyngeal derivatives. During these morphogenetic movements, mesecto-dermal rudiments receive mesoderm-deriving cells, making the neck a verycomposite structure. Most of its ventral part originates from initially anteriormediodorsal ectodermal material which migrates ventrad during the early stagesof embryogenesis and caudad during later morphogenesis.
Although these data result from studies on the avian embryo, it is possibleto infer the general picture of NC migration pattern in mammalian embryos.Thus, a number of spontaneous malformations of the face and brain in thehuman can be explained by defective crest cell migration and differentiation.
RESUME
Des greffes interspecifiques d'ebauches neurales rhombencephaliques et mesencephaliquesont ete realisees entre embryons de Caille et de Poulet. Les caracteres particuliers du noyauchez la Caille permettent de reconnaitre les cellules de l'hote de celles du greffon dans lesembryons chimeres, et d'etudier les voies de migration et les capacites de differenciation descellules issues de la crete neurale (CN) greffee. Le 'marquage' fourni par les cellules de Cailleest en effet stable et permet de suivre les cellules migrantes jusqu'a ce qu'elles aient atteintun stade de complete differenciation. Dans un travail precedent base sur une experimentationsimilaire on avait montre que le squelette visceral derive dans sa totalite des cretes neuralesrhombencephaliques et mesencephaliques. Les recherches rapportees ici concernent lesderives mesenchymateux de la CN autres que les tissus osseux et cartilagineux.
Apres la greffe isotopique et isochronique du rhombencephale et du mesencephale, Jederme de la face et de la partie ventrolaterale du cou de l'hote est forme de cellules issues dugreffon et par consequent d'origine mesectodermique. II en est de meme de la paroi musculo-conjonctive des gros troncs arteriels issus du coeur et derivant des arcs aortiques (troncsbrachiocephaliques, carotides communes, crosse aortique, arteres pulmonaires). Le con-jonctif lache du plancher buccal, de la langue, de la face lateroventrale du cou, ainsi que latrame conjonctive des glandes derivees du pharynx et de la bouche (glandes linguales,thyroide, parathyroides, thymus) sont formes par du mesectoderme neural.
Les cellules mesenchymateuses originates des CN ont la potentialite de se differencier encellules musculaires lisses et participent aussi a l'histogenese des muscles striees des arcsbranchiaux. Elles fournissent du tissu adipeux dans le derme profond et au voisinage de latrachee. Comme cela a ete precedemment demontre par notre groupe, les CN rhomben-cephaliques sont a 1'origine des cellules a calcitonine du corps ultimobranchial et des cellulesglandulaires du corps carotidien. Ces dernieres sont caracterisees par une forte teneur enmonoamines fluorigenes. Des cellules similaires ont ete detectees dans la paroi de l'arterecarotide commune par la methode de fluorescence induite par les vapeurs de formol et nousavons pu montrer qu'elles derivent egalement de la CN.
II apparait done que les potentialites de differenciation des cellules du mesectoderme sontvariees. Cependant il ne semble pas qu'elles soient a 1'origine de l'endothelium des vaisseauxsanguins qui, dans nos experiences, derivent toujours de cellules mesodermiques appartenanta l'embryon hote. La capacite de la crete neurale a fournir des derives mesenchymateux estlimitee a la region du nevraxe anterieure au niveau du 5eme somite.

Mesenchymal derivatives of avian neural crest 151
REFERENCES
ANDRES, G. (1946). Uber Tnduktion und Entwicklung von Kopforganen aus Unkenektodermin Molch (Epidermis, Plakoden und Derivate der Neuralleiste). Revue suisse Zool. 53,502-510.
ANDRES, G. (1949). Untersuchungen an Chimaren von Triton und Bombinator. T. Ent-wicklung xenoplastischer Labyrinthe und Kopfganglien. Genetica 24, 397-534.
ANDREW, A. (1964). The origin of intramural ganglia. I. The early arrival of precursor cellsin the presumptive gut of chick embryo. J. Anat. 98, 421-428.
ANDREW, A. (1969«). The origin of intramural ganglia. IT. The trunk neural crest as a sourceof enteric ganglion cells. / . Anat. 105, 89-10.1.
ANDREW, A. (19696). The origin of intramural ganglia. III. The vagal source of entericganglion cells. / . Anat. 107, 327-336.
ANDREW, A. (1970). The origin of intramural ganglia. IV. The origin of enteric ganglia. Acritical review and discussion of the present state of problem. /. Anat. 108, 169-184.
BERNHARD, W. (1968). Une methode de coloration regressive a l'usage de la microscopieelectronique. C. r. hebd Seanc. Acad. Sci., Paris 267, 2170-2173.
CHIBON, P. (1962). Marquage nucleaire de blastulas et d'explants embryonnaires d'Am-phibiens par la thymidine tritiee. Evolution des noyaux marques au cours du developpe-ment. C. r. hebd Seanc. Acad. Sci., Paris 254, 4508-4510.
CHIBON, P. (1964). Analyse par la methode de marquage nucleaire a la thymidine tritieedes derives de la crete neurale cephalique chez l'Urodele. C. r. hebd. Seanc. Acad. Sci.,Paris 259, 3624-3627.
CHIBON, P. (1966). Analyse experimentale de la regionalisation et des capacites morpho-genetiques de la crete neurale chez 1'Amphibien Urodele {Pleurodeles Waltlii Michah).Mem. Soc. zoo. Fr. 36, 1-107.
DE BEER, G. R. (1947). The differentiation of neural crest cells into visceral cartilages andodontoblasts in Amblystoma and a re-examination of the germ-layer theory. Proc. R.Soc. Loud. B 134, 377-398.
FALCK, B. (1962). Observations on the possibility of the cellular localization of mono-amines by a fluorescence method. Acta physiol. scand. 56, suppl. 197.
FEULGEN, R. & ROSSENBECK, H. (1924). Mikroskopisch-chemischer Nachweis einer Nuclein-saure vom Typus der Thymonucleinsaure und die darauf beruhende elektive Farbungvon Zellkernen der mikroskopischen Praparaten. Hoppe-Seyler's Z. physiol. Chem. 135,203-248.
FONTAINE, J. (1973). Contribution a 1'etude du developpement du corps carotidien et ducorps ultimobranchial des Oiseaux: Le contenu en monoamines et la capacite de capterla L-DOPA des cellules glomiques et des cellules a calcitonine au cours de la vie embryon-naire. Archs Anat. microsc. Morph. exp. 62, 89-100.
GORONOWITSCH, N. (1892). Die axiale und die laterale Kopfmetamerie der Vogelembryonen.Die Rolle der sog. 'Ganglienleisten' im Auf bau der Nervenstamme. Anat. Anz. 7, 454-464.
GORONOWITSCH, N. (1893). Untersuchungen iiber die Entwicklung der sog. 'Ganglienleisten'im Kopfe der Vogelembryonen. Morph. Jb. 20, 187-259.
HAMBURGER, V. & HAMILTON, H. (1951). A series of normal stages in the development ofthe chick embryo. / . Morph. 88, 49-92.
HAMMOND, W. S. & YNTEMA, C. L. (1953). Deficiencies in visceral skeleton of the chickafter removal of cranial neural crest. Anat. Rec. 115, 393-394.
HAMMOND, W. S. & YNTEMA, C. L. (1964). Depletions of pharyngeal arch cartilage followingextirpation of cranial neural crest in chick embryos. Acta anat. 56, 21—34.
HARRISON, R. G. (1935). Heteroplastic grafting in embryology. Harvey Lect. no. 29, pp.1933-1934.
HARRISON, R. G. (1938). Die Neuralleiste. Anat. Anz. 85, 3.HoLTFRETER, J. (1933). Der Einfluss von Wirtsalter und verschiedenen Organbezirken auf
die Differenzierung von angelagertem Gastrulaektoderm. Wilhelm Roux Arch. EntwMech.Org. 127, 619-775.

152 C. S. LE LIEVRE AND N. M. LE DOUARIN
HOLTFRETER, J. (1935a). Morphologische Beinflussung von Urodelenektoderm beixenoplastischer Transplantation. Wilhelm Roux Arch. EntwMech. Org. 133, 367-426.
HOLTFRETER, J. (19356). Uber das Verhalten von Anurenektoderm in Urodelenkeimen.Wilhelm Roux Arch. EntwMech. 133, 423-494.
HOLTFRETER, J. (1968). Mesenchyme and epithelia in inductive and morphogenetic processes.In Epithelial Mesenchyme Interactions. Baltimore: Williams and Wilkins.
HORSTADIUS, S. (1950). The Neural Crest: Its Properties and Derivatives in the Light ofExperimental Research. New York, London: Oxford University Press.
HORSTADIUS, S. & SELLMAN, S. (1946). Experimentelle Untersuchungen uber die Deter-mination des knorpeligen Kopfskelettes bei Urodelen. Nova Acta R. Soc. Scient. upsal.ser. IV, 13, 1-170.
JOHNSTON, M. C. (1966). A radioautographic study of the migration and fate of cranialneural crest cells in the chick embryo. Anat. Rec. 156, 130-143.
JOHNSTON, M. C. & LISTGARTEN, M. A. (1973). Observations on the migration, interactionand early differentiation of orofacial tissues. In Developmental Aspects of Oral Biology.New York: Academic Press.
JOHNSTON, M. C, BHAKDINARONK, A. & REID, Y. C. (1973). An expanded role of theneural crest in oral and pharyngeal development. In Oral Sensation and PerceptionDevelopment in the Fetus and Infant (ed. J. F. Bosma).
KATSCHENKO, N. (1888). Zur Entwicklungsgeschichte der Selachierembryos. Anat. Anz. 3,445-467.
KOCH, W. E. (1965). In vitro development of tooth rudiments of embryonic mice. Anat. Rec.152,513-524.
LE DOUARIN, N. (1969). Particularity du noyau interphasique chez la Caille japonaise(Coturnix coturnix japonicd). Utilisation de ces particularity comme 'marquage biolo-gique' dans les recherches sur les interactions tissulaires et les migrations cellulaires aucours de l'ontogenese. Bull. biol. Fr. Belg. 103, 435-452.
LE DOUARIN, N. (1971a). Etude ultrastructurale comparative du noyau interphasique chezla Caille (Coturnix coturnix japonicd) et le Poulet (Gallus gallus) par la methode de colora-tion regressive a l'EDTA. C. r. hebd. Seanc. Acad. Sci., Paris 272, 2334-2337.
LE DOUARIN, N. (19716). Caracteristiques ultrastructurales du noyau interphasique chezla Caille et chez le Poulet et utilisation de cellules de Caille comme 'marqueurs biolo-giques' en Embryologie experimentale. Annls Embryol. Morph. 4, 125-135.
LE DOUARIN, N. (1973 a). A biological cell labelling technique and its use in experimentalEmbryology. Devi Biol. 30, 217-222.
LE DOUARIN, N. M. (19736). A Feulgen-positive nucleolus. Expl Cell Res. 11, 459-468.
LE DOUARIN, N., FONTAINE, J. & LE LIEVRE, C. (1974). New studies on the neural crestorigin of the avian ultimobranchial glandular cells. Interspecific combinations and cyto-chemical characterization of C cells based on the uptake of biogenic amine precursors.Histochemie 38, 297-305.
LE DOUARIN, N. & LE LIEVRE, C. (1970). Demonstration de l'origine neurale des cellules acalcitonine du corps ultimobranchial chez l'embryon de Poulet. C. r. hebd. Seanc. Acad.Sci., Paris 270, 2857-2860.
LE DOUARIN, N. & LE LIEVRE, C. (1971). Sur l'origine des cellules a calcitonine du corpsultimobranchial de l'embryon d'Oiseau. C. r. Ass. Anat. 152, 558-568.
LE DOUARIN, N. & LE LIEVRE, C. (1972). Demonstration of the neural origin of the ultimo-branchial body glandular cells in the avian embryo. Hlrd Int. Symp. of Endocr., July1971, London.
LE DOUARIN, N., LE LIEVRE, C. & FONTAINE, J. (1972). Recherches experimentales surl'origine embryologique du corps carotidien chez les Oiseaux. C. r. hebd. Seanc. Acad.Sci., Paris 275, 583-586.
LE DOUARIN, N. & TEILLET, M. A. (1970). Sur quelques aspects de la migration des cellulesneurales chez l'embryon de Poulet etudies par la methode des greffes heterospecifiquesde tube nerveux. C. r. Seanc. Soc. Biol. 164, 390-397.

Mesenchymal derivatives of avion neural crest 153LE DOUARIN, N. & TEILLET, M. A. (1971 a). Localisation, par la methode des greffes inter-
specifiques, du territoire neural dont derivent les cellules adrenales surrenaliennes chezl'embryon d'Oiseau. C. r. hebd. Seanc. Acad. Sci., Paris 272, 481-484.
LE DOUARIN, N. & TEILLET, M. A. (19716). L'origine des cellules du systeme ganglionnaireintramural du tractus digestif chez l'embryon d'Oiseau. C. r. hebd. Seanc. Acad. Sci.,Paris 213, 1411-1414.
LE DOUARIN, N. & TEILLET, M. A. (1973a). The migration of neural crest cells to the wallof the digestive tract in avian embryo. / . Embryol. exp. Morph. 30, 31-48.
LE DOUARIN, N. & TEILLET, M. A. (19736). Recherches sur le determinisme de la migrationdes cellules issues de la crete neurale. C. r. hebd. Seanc. Acad. Sci., Paris 277, 1929-1932.
LE DOUARIN, N. M. & TEILLET, M. A. M. (1974). Experimental analysis of the migrationand differentiation of neuroblasts of the autonomic nervous system and of neurectodermalmesenchymal derivatives, using a biological cell marking technique. Devi Biol. 41, 162—184.
LE LIEVRE, C. (1971a). Recherches sur l'origine embryologique des arcs visceraux chezl'embryon d'Oiseau par la methode des greffes interspecifiques entre Caille et Poulet.C. r. Seanc. Soc. Biol. 165, 395-400.
LE LIEVRE, C. (19716). Recherche sur l'origine embryologique du squelette visceral chezl'embryon d'Oiseau. C. r. Ass. Anat. 152, 575-583.
LE LIEVRE, C. (1974). Role des cellules mesectodermiques issues des cretes neuralescephaliques dans la formation des arcs branchiaux et du squelette visceral. / . Embryol.exp. Morph. 31, 453-477.
LE LIEVRE, C. & LE DOUARIN, N. (1973). Contribution du mesectoderme a la genese desarcs aortiques chez l'embryon d'Oiseau. C. r. hebd. Seanc. Acad. Sci., Paris 276, 383-386.
LE LIEVRE, C. & LE DOUARIN, N. (1974). Origine ectodermique du derme de la face et ducou, montree par des combinaisons interspecifiques chez l'embryon d'Oiseau. C. r. hebd.Seanc. Acad. Sci., Paris 278, 517-520.
MAURO, A. (1961). Satellite cell of skeletal muscle fibers. / . biophys. biochem. Cytol. 9,493-494.
MOSCONA, A. & MOSCONA, H. (1952). The dissociation and aggregation of cells from organrudiments of the early chick embryo. / . Anat. 86, 287-301.
Moss, F. P. & LEBLOND, P. (1971). Satellite cells as the source of nuclei in muscles ofgrowing rats. Anat. Rec. 170, 421-436.
NEWTH, D. R. (1954). Determination of the cranial neural crest of the Axolotl. /. Embryol.exp. Morph. 2, 101-105.
PEARSE, A. G. E., POLAK, J. M., ROST, F. W. D., FONTAINE, J., LE LIEVRE, C. & LE DOUARIN,N. (1973). Demonstration of the neural crest origin of type I (APUD) cells in the aviancarotid body using a cytochemical marker system. Histochemie 34, 191-203.
PLATT, J. B. (1893). Ectodermic origin of the cartilages of the head. Anat. Anz. 8, 506-509.POLAK, J. M., PEARSE, A. G. E., LE LIEVRE, C, FONTAINE, J. & LE DOUARIN, N. M. (1974).
Immunocytochemical confirmation of the neural crest origin of avian calcitonin-producingcells. Histochemie 40, 209-214.
RAVEN, C. P. (1931). Zur Entwicklung der Ganglienleiste. I. Die Kinematik der Ganglien-leistenentwicklung bei den Urodelen. Wilhelm Roux Arch. EntwMech. Org. 125, 210-292.
RUDNICK, D. (1932). Thyroid forming potencies of the early chick blastoderm. /. exp.Zool. 62, 287-313.
RUDNICK, D. (1938). Differentiation in culture of pieces of the early chick blastoderm. II.Short primitive streak stages. / . exp. Zool. 79, 399-425.
SAXOD, R. (1973). Developmental origin of the Herbst cutaneous sensory corpuscule.Experimental analysis using cellular markers. Devi Biol. 32, 167-178.
SELLMAN, S. (1946). Some experiments on the determination of the larval teeth in Ambystomamexicanum. Odontologisk Tidskrift. 54, 1—128.
SIGNORET, J. (1960). Cephalogenese chez le triton Pleurodeles Waltlii Michah apres traitementde la gastrula par le chlorure de lithium. Mem. Soc. zool. Fr. 32, 1-117.
STONE, L. S. (1922). Some notes on the migration of neural crest in Ranapalustris. Anat. Rec.23, 39-40.

154 C. S. LE LIEVRE AND N. M. LE DOUARIN
STONE, L. S. (1926). Further experiments on the extirpation and transplantation of mesecto-derm in Ambystoma punctatum. J. exp. Zool. 44, 95-131.
STONE, L. S. (1929). Experiments showing the role of migrating neural crest (mesectoderm)in the formation of the head skeleton and lose connective tissue in Rana palustris. WilhelmRoux Arch. EntwMech. Org. 118, 40-77.
STONE, L. S. (1932). Transplantation of hyobranchial mesentoderm including the right lateralanlage of the second basibranchium in Ambystoma punctatum. J. exp. Zool. 62, 109-123.
TEILLET, M. A. (1971a). Recherches sur le mode de migration et la differenciation desmelanocytes cutanes chez l'embryon d'Oiseau: etude experimental par la methode desgreffes heterospecifiques entre embryons de Caille et de Poulet. Annls Embryol. Morph. 4,95-109.
TEILLET, M. A. (19716). Niveau d'origine de plusieurs derives des cretes neurales cervicalesposterieures, thoraciques et Jombaires chez l'embryon d'Oiseau. C. r. Ass. Anat. 152,734-743.
TEILLET, M. A. & LE DOUARIN, N. (1970). La migration des cellules pigmentaires etudieepar la methode des greffes heterospecifiques de tube nerveux chez l'embryon d'Oiseau.C. r. /rebel. Seanc. Acad. Sci., Paris 270, 3095-3098.
TEILLET, M. A. & LE DOUARIN, N. (1974). Determination par la methode des greffes hetero-specifiques d'ebauches neurales de Caille sur l'embryon de Poulet, du niveau du nevraxedont derivent les cellules medullo-surrenaliennes. Archs Anat. microsc. Morph. exp. 63,51-62.
WAGNER, G. (1949). Die Bedeutung der Neuralleiste fur die Kopfgestaltung der Amphibien-larven. Untersuchungen an Chimaeren von Triton. Revue suisse Zool. 56, 519-620.
WAGNER, G. (1955). Chimaerische Zahnanlagen aus Triton-Schmelzorgan und Bombinator-Papille mit Beobachtungen iiber die Entwicklung von Kiemenzahnchen und Mund-sinnesknospen in den Tritonlarven. /. Embryol. exp. Morph. 3, 160-188.
WAGNER, G. (1959). Untersuchungen an Bombinator-Triton Chimaeren. Das Skelettlarvalen Tritonkopfe mit Bombinator Mesektoderm. Wilhelm Roux Arch. EntwMech. Org.151, 136-158.
WESTON, J. A. (1962). The migration and localization of neural crest cells labelled withtriated thymidine. Am. Zool. 2, 458.
WESTON, J. A. (1963). A radioautographic analysis of the migration and localization of trunkneural crest cells in the chick. Devi Biol. 6, 279-310.
WESTON, J. A. (1970). The migration and differentiation of neural crest cells. Advances inMorphogenesis 8, 41-114.
WESTON, J. A. & BULTER, S. L. (1966). Temporal factors affecting localization of neuralcrest cells in the chicken embryo. Devi Biol. 14, 246-266.
WILLIER, B. H. & RAWLES, M. E. (1931). Developmental relations of the heart and liver inchorioallantoic grafts of whole chick blastoderm. Anat. Rec. 48, 277-301.
ZACCHEI, A. M. (1961). Lo Sviluppo embrionale della quaglia giapponese (Coturnix coturnixjaponica T.S.). Archo ital. Anat. Embriol. 66, 36-62.
(Received 13 December 1974)