mercury accumulation in marine bivalves: influences of biodynamics and feeding niche
TRANSCRIPT

lable at ScienceDirect
Environmental Pollution 159 (2011) 2500e2506
Contents lists avai
Environmental Pollution
journal homepage: www.elsevier .com/locate/envpol
Mercury accumulation in marine bivalves: Influences of biodynamicsand feeding niche
Ke Pan, Wen-Xiong Wang*
Division of Life Science, The Hong Kong University of Science and Technology (HKUST), Clear Water Bay, Kowloon, Hong Kong
a r t i c l e i n f o
Article history:Received 9 March 2011Received in revised form15 June 2011Accepted 17 June 2011
Keywords:MercuryMethylmercuryBivalvesBiodynamicsSubcellular distributionFeeding niche
* Corresponding author.E-mail address: [email protected] (W.-X. Wang).
0269-7491/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.envpol.2011.06.029
a b s t r a c t
Differences in the accumulation of mercury (Hg) in five species of marine bivalves, including scallopsChlamys nobilis, clams Ruditapes philippinarum, oysters Saccostrea cucullata, green mussels Perna viridis,and black mussels Septifer virgatus, were investigated. The bivalves displayed different patterns of Hgaccumulation in terms of the body concentrations of methylmercury (MeHg) and total Hg (THg), as wellas the ratio of MeHg to THg. Parameters of the biodynamics of the accumulation of Hg(II) and MeHgcould reflect the species-dependent Hg concentrations in the bivalves. With the exception of blackmussels, we found a significant relationship between the efflux rates of Hg(II) and the THg concentra-tions in the bivalves. The interspecific variations in the MeHg to THg ratio were largely controlled by therelative difference between the elimination rates of Hg(II) and MeHg. Stable isotope (d13C) analysisindicated that the five bivalve species had contrasting feeding niches, which may also affect the Hgaccumulation.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Bioaccumulation of mercury (Hg) has been a long-standing topicsince themercury poisoning of thousands of peoplewho consumedHg-contaminated fish and shellfish in Minamata Bay, Japan (Ditri,1991). Hg is ubiquitous in natural waters, sediments, soils and air,making it a global health concern. The organometallic form of Hg,methylmercury (MeHg), is a potent neurotoxin and has a strongtendency to be biomagnified in aquatic food chains, despite its lowenvironmental concentrations (Watras and Bloom, 1992; Ullrichet al., 2001). One basic question surrounding Hg bioaccumulationis how Hg concentrations of parts per trillion in water can yield Hgconcentrations of parts per million in fish (Morel et al., 1998; Chenet al., 2008). Although considerable progress has been made inunderstanding the Hg accumulation in aquatic organisms over thepast decades (Mason et al., 1995, 1996; Gagnon and Fisher, 1997;Wang et al., 1998; Pan and Wang, 2004), there are still gaps inour understanding of the biodynamics involved.
The accumulation of Hg in bivalve molluscs is of great interestbecause they are widely consumed by humans worldwide. Bivalvesare strong metal accumulators and may act as an important vectorfor ‘pumping’ Hg to higher trophic levels. Previous studies have
All rights reserved.
indicated that bivalves can efficiently absorb Hg(II) and MeHg fromwater and sediments (Gagnon and Fisher, 1997; Pan and Wang,2004), and environmental variables such as pH, temperature, dis-solved organic carbon, and salinity have substantial effects on theuptake of Hg (Inza et al., 1998; Tsui and Wang, 2004; Wang andWang, 2010). These studies are indeed valuable for interpretingthe crucial steps controlling the entry of Hg(II) and MeHg intothe food web. The interspecific divergence of Hg accumulation inaquatic organisms is however less well known. Bivalve molluscsreduce their metal toxicity through a series of mechanismsincluding sequestration in metallothionein (MT), forming inertgranules by lysosomal vesicles for storage or exocytosis (Dallinger,1993; Marigömez et al., 2002). The choice of detoxification strategyaffects the biodynamics of metals, leading to great variations inmetal concentrations among species.
Differences in the body concentrations of Hg in aquatic organ-isms may reflect the interspecific biodynamics of Hg, and may alsobe caused by the choice of feeding habits by the animals. Foodpartitioning is a common mechanism by which substantial differ-ences in resource use occur between cohabiting species (Ross,1986).Bivalves constitute a diverse taxonomical group occupying a widehabitat in coastal environments. These filter-feeders generally feedopportunistically on any phytoplankton, detritus, or benthic algaeavailable in their habitats. Closely related species may feed on subtlydifferent food sources resulting in different accumulations of metals(Rainbow, 1995). Our previous study has shown that changes in the

K. Pan, W.-X. Wang / Environmental Pollution 159 (2011) 2500e2506 2501
growth environment can have significant impact on food avail-ability, quality and growth for scallops Chlamys nobilis, thus affectingthe related accumulation of Cd and Zn (Pan and Wang, 2008). Chenet al. (2009) also demonstrated that concentrations of MeHg werehigher for pelagic-feeding fish than for benthic-feeding fish. Thesestudies imply that the feeding niche may have a great impact onmetal bioaccumulation, yet the link between feeding niche and thebioaccumulation of Hg remains underexplored.
In this study, we therefore conducted a comprehensive investi-gation on the inter-species difference in Hg(II) and MeHg bio-accumulation in five species of marine bivalves. We determined (1)the difference in Hg bioaccumulation from water and food; (2) thedetoxification strategies and their links with the interspecific Hgbody concentrations; and (3) how different feeding niches affect Hgaccumulation. Our overall objective was to understand the inter-species difference of Hg bioaccumulation in marine bivalves basedon the biodynamics of Hg and the feeding niche of the bivalves.
2. Materials and methods
2.1. Marine bivalves
Five species of marine bivalves commonly found along the east coasts of HongKong were collected. Scallops Chlamys nobilis and green mussels Perna viridis werecollected from Dapeng Bay and Tolo Harbor, respectively. Clams Ruditapes philip-pinarum, black mussels Septifer virgatus and rock oysters Saccostrea cucullata werecollected from Clear Water Bay. Individuals of a wide range of body sizes werecollected, which were acclimatized in circulating seawater (20 �C and 33 psu) ina 200 L water tank for one week. During the acclimation periods, the bivalves werefed with diatoms Thalassiosira weissflogii twice a day.
2.2. Total Hg and MeHg in seawater, sediments and bivalves
To measure the total Hg (THg), surface seawaters were sampled from DapengBay, Tolo Harbor and Clear Water Bay. The seawater samples were filtered withMillipore 0.45 mm polycarbonate filters in a clean bench. Bromine monochloride(BrCl) was added to filtered (for dissolved Hg) and unfiltered (for THg) samples toproduce a final concentration of 0.5% (v/v). The samples were digested at roomtemperature for 12 h prior to analysis by cold vapor atomic fluorescence spec-trometer (CVAFS) according to EPA method 1631E. Total suspended particulatematter (SPM) was also determined by filtering an aliquot of seawater sample onto anAdvantec� GC-50 glass filter, which was rinsed with isotonic ammonium formate toremove the salts. Surface sediments (1e2 cm, <63 mm, n ¼ 3) were collected fromClear Water Bay.
For measurements of background THg and MeHg concentrations in the bivalves,individuals of different sizes for each bivalve species were randomly selected fromthe collection. Selected bivalves were depurated without feeding for two days.Afterwards the bivalves were dissected, freeze-dried, and ground into a fine powder.For small individuals (shell length <1 cm), 10e20 individuals of similar size werepooled to make up a composite sample. For each site, totally 50 samples were made,among which the number of composite samples was between 10 and 15. Driedtissues or sediments were weighed and approximately 0.2 g of which were digestedat 190 �C with aqua regia (2 mL HNO3:6 mL HCl) in a microwave digestion system.MeHg in bivalves were extracted by digesting approximately 40 mg of homogenizedsoft tissues with 25% KOH in methanol at 60 �C for 3 h.
All containers for sampling and analysis of Hg were vigorously cleaned with hot4 N HCl acid. THg was analyzed using the single gold trap amalgamation techniqueby CVAFS (QuickTrace� 8000, USA). Method detection limit (MDL) of THg was0.18 ng L�1. All bottle and filter blanks had THg below the MDL. Matrix recoveries forseawater samples were 85e95%. MeHgwasmeasured for biological samples with anautomated analytical system (MERX, Brooks Rand). Briefly, the extract of bivalvetissues was buffered with sodium acetate at pH 4.9, and ethylated by sodium tet-raethylborate in a 40 mL Teflon line borate glass bottle. The quantification of MeHgwas automatically carried out by the MeHg analyzer. Analytical accuracy of bio-logical and sediment samples was checked by concurrent digestion and analysis ofcertified referencematerial IAEA-142 (mussel homogenate) and NIST 1646a (estuarysediments), with a recovery of 91e94% for THg and 89e96% for MeHg.
2.3. Biodynamics of Hg(II) and MeHg in bivalves
Bivalves of 20e30 mm shell height were used in the biodynamics experiments.The radioisotope 203Hg(II) (t1/2 ¼ 46.6 d, specific activity:162e200 GBq g�1) used inthe study was purchased from Isotope Products Laboratories, Eckert & Ziegler,Germany. Me203Hg was synthesized from the 203Hg(II) following a well-establishedmethod (Rouleau and Block, 1997). Parameters of the biodynamics of Hg(II) and
MeHg, the dissolved uptake constant, assimilation efficiency, and excretion rateconstant, were then quantified.
To measure the influx rates of waterborne Hg(II) and MeHg, the radioisotope203Hg(II) or Me203Hg were added into the 0.22 mm filtered seawater to give differentHg concentrations (25, 50, 100, 200, 400 ng L�1 for Hg(II), and 10, 20, 50, 100,200 ng L�1 for MeHg). Each concentration treatment contained four replicateindividuals, and the bivalves were placed individually in 500 mL medium held ina Teflon beaker. The exposure period was 30 min for Hg (II) and 5e10 min for MeHg,during which the decline of metal concentration was within 20%. Afterwards thebivalves were carefully rinsed with non-radioactive seawater and dissected, andtheir radioactivities were determined. The whole animals were completely dried at60 �C to determine the dry weights. Hg(II) or MeHg influx rate (Iw, mg g�1 day�1) wascalculated as follows:
Iw ¼ AtissueSA�W � t
;
where Atissue is the radioactivity in the whole soft tissue after exposure (ccpm), SA isthe specific activity of Hg(II) or MeHg in seawater (ccpm m g�1), W is the dry weightof the soft tissue (g), and t is the duration of exposure (d). The dissolved uptake rateconstant (ku) can be calculated by the influx rate divided by the metal concentrationin the dissolved phase (Cw).
The assimilation efficiencies (AEs) of Hg in marine diatoms T. weissflogii andsediments were determined using the pulse feeding and gamma radioactive tracermethod (Wang and Fisher, 1996; Pan and Wang, 2009a). Gamma detection isnondestructive, thus the radioactivity in the same individual bivalve can be assessedover time. The diatoms were grown in an f/2mediumwith N, P, Si, vitamins and tracemetals minus Zn, Cu, and EDTA. A stock solution (500mg L�1) of sediments (<63 mm)was prepared with 0.22 mm filtered seawater. The radioisotope 203Hg(II) or Me203Hgwas added to exponentially growing diatoms at a concentration of 370 kBq L�1 andthe diatoms were grown for two days. The sediment was radiolabeled at 740 kBq L�1
for two days as well. The radiolabeled diatoms or sediments were collected bycentrifugation, and were washed in non-radioactive filtered seawater. There were 10replicate individuals in each treatment. The bivalves were fed diatoms at a concen-tration of 5 � 104 cells mL�1 or sediments at a concentration of 2 mg L�1 for 30 min.After the pulse feeding, the bivalves were rinsed thoroughly with filtered seawaterand their initial radioactivities determined, and then placed separately into poly-propylene beakers containing 200mL of seawater held in a 10 L enclosed recirculatingflow-through aerated seawater aquarium. non-radioactive algae T. weissflogii werefed to the bivalves twice a day to depurate their guts of the ingested radiolabeled food.Feces were removed frequently and the seawater was changed twice a day during theexperiment. Radioactivity in each bivalve was assayed at 12 h intervals over 48 h.Assimilation efficiency (AE) was determined as the percentage of the initial radio-activity retained in bivalves after 48 h of depuration.
To measure the Hg(II) and MeHg efflux rates of bivalves, fifteen individuals of eachbivalve species were exposed to dissolved 203Hg(II) or Me203Hg for 1 h per day (at37 kBq L�1). During the radioactive exposure, diatoms T. weissflogii radiolabeled with203Hg(II) or Me203Hgwere also fed to the bivalves. The dual-exposure lasted for six daysand all the bivalves were assayed for their initial activities on the seventh day. Twobivalveswere dissected, and the radioactivity in the tissues and shells was also counted.The bivalves were depurated in non-radioactive seawater for 24 days. The radioactivityin each bivalve as a whole animal was assayed periodically throughout the experiment.Meanwhile, three individuals were sampled on the sixth day for subcellular metaldistribution analysis. The efflux rate constant (ke, day�1) was calculated from the slopeof the regression between the natural log of the percentage of 203Hg(II) or Me203Hgretained in the slow exchange compartment and the period of depuration.
2.4. Subcellular fractionation of Hg(II) and MeHg
The subcellular fractionation was carried out according to Wallace et al. (2003)and five fractions were separated (cellular debris, metal-rich granule fraction-MRG,organelle, heat-sensitive protein [HSP], and metallothionein-like protein [MTLP]).Briefly, the soft tissueswere homogenized in 5mL of 30mMTriseNaCl buffer (pH 8.0;0.15 M sodium chloride; 5 mM freshly prepared antiprotease, 2-mercaptoethanol;0.1 mM phenylmethylsulfonyl fluoride (PMSF)). The homogenate was centrifuged at1450g at 4 �C for 15 min. The separated pellet was the cellular debris fraction, whichwas digested in 1 N NaOH at 80 �C for 10 min, and centrifuged at 5000 g again for10 min. The resulting pellet was theMRG and the supernatant was the cellular debrisfraction. The supernatant from the first centrifugation was further subjected tocentrifugation at 100,000g at 4 �C for 1 h. The pellet from this step was the organellefraction (organelles). After 80 �C treatment of the supernatant from this step, the HSPwas denatured and separated by centrifugation at 50,000g and 4 �C. The finalsupernatant was the MTLP fraction. Each fraction was assayed for radioactivity. Themetal subcellular distribution was defined as the percentage of 203Hg(II) or Me203Hgin each fraction.
2.5. Stable isotopic signature (d13C and d15N) for bivalves
Selected bivalves (shell length: w20 mm) were depurated in 0.22 mm filteredseawater for two days to allow the evacuation of gut contents. The soft tissues were

Table 1Summary of THg andMeHg concentrations in seawater and bivalves collected from the studied sites. Data presented asmean� SD. n¼ 3 for THg, total dissolved Hg, and SPM inseawater. n ¼ 50 for the body concentrations of THg and MeHg in bivalves. NA: not applicable.
Site Total Hg(ng L�1)
Total dissolved Hg(ng L�1)
SPM load(mg L�1)
Species Body burden of Hg(ng g�1)
MeHg/THg(%)
Allometric function(Y ¼ aWb)
THg MeHg THg MeHg
Dapeng Bay 0.39 � 0.05 0.21 � 0.04 2.0 � 0.3 Scallop C. nobilis 60.3 � 14.2 18.8 � 5.7 31.4 � 6.9 53.5W�0.13 17.0W�0.11
Tolo Harbor 0.65 � 0.10 0.35 � 0.01 3.1 � 0.2 Green mussel P. viridis 30.2 � 4.1 8.9 � 2.0 29.9 � 6.4 26.3W�0.10 NAClam R. philippinarum 47.4 � 15.3 16.6 � 3.5 37.6 � 11.8 NA NA
Clear Water Bay 0.38 � 0.02 0.20 � 0.01 1.7 � 0.1 Oyster S. cucullata 70.3 � 15.3 15.2 � 3.3 22.4 � 6. 8 NA NABlack mussel S. virgatus 91.9 � 18.8 10.4 � 1.5 11.6 � 2.2 NA NA
Scallop
0 1 2 3 4
-1)
0
50
100
150
200THg
MeHg
Green mussel
0.0 0.4 0.8 1.2 1.60
50
100
150
200
Black mussel
Tissue dry weight (g)
0.0 0.2 0.4 0.6 0.8 1.00
50
100
150
200
Clam
0.0 0.3 0.6 0.9 1.20
50
100
150
200
Oyster
Tissue dry weight (g)
0.0 0.2 0.4 0.6 0.8
Con
cent
ratio
n (n
g g
-1)
Con
cent
ratio
n (n
g g
0
50
100
150
200
Fig. 1. Concentrations of THg and MeHg (ng g�1) relative to the body dry weights infive species of bivalves.
K. Pan, W.-X. Wang / Environmental Pollution 159 (2011) 2500e25062502
dissected and freeze-dried, which were then ground into a fine powder. Five indi-viduals were combined into one composite sample in order to obtain a representa-tive sample (Herman et al., 2000). Stable isotopic measurements were performedwith Finnigan MAT Delta V advantage isotope ratio mass spectrometer. Results arereported as comparisons with atmospheric nitrogen (for N) and Vienna Pee DeeBelemnite (for C) as reference standards, and were calculated as: d13C ord15N ¼ (Rsample/Rstandard � 1) � 1000, where R is the 13C/12C or 15N/14N ratio for d13Cor d15N, respectively. The analysis errors of reproducibility were within �0.2& forboth d13C and d15N.
2.6. Radioactivity measurements, data treatments and statistical analysis
The radioactivity of 203Hg(II) and Me203Hg were determined using a wallac1480 NaI(T1) gamma counter (Turku, Finland). All counts were related tostandards and spillover. The gamma emission of 203Hg(II) and Me203Hg wasdetermined at 232 keV. Counting times were adjusted to yield a propagatedcounting error of less than 5%. All counts data were corrected for decay beforeany calculations. The statistical significance of the regressions was tested byanalysis of variance, and the regressions were quantified by the coefficient ofdetermination, r2. Differences in AE and subcellular metal distribution amongthe treatments were detected by one-way analysis of variance with a least-significant-difference (LSD) post hoc test in which all the percentage data werearcsine transformed before analysis. Analysis of covariance (ANCOVA) was per-formed to compare the THg and MeHg between different species of bivalves(Fialkowski et al., 2003).
3. Results and discussion
3.1. Total Hg concentrations in seawater and sediments
Total dissolved Hg concentrations in the seawater were 0.20,0.35, and 0.21 ng L�1 for Dapeng Bay, Tolo Harbor and Clear WaterBay, respectively (Table 1). Total dissolved Hg accounted for about50% of THg, which was 0.39 ng L�1 for Dapeng Bay, 0.64 ng L�1 forTolo Harbor, and 0.38 ng L�1 for Clear Water Bay. The calculatedTHg concentrations in SPMwere 90, 97 and 106 ng g�1 for the threesites. THg concentration in the sediments from ClearWater Bay wasonly 65.8 � 3.0 ng g�1 (n ¼ 3), lower than that of sediments foundin SPM (p < 0.01). Generally, THg concentrations in the uncon-taminated marine environments were in the range of0.1e0.8 ng L�1 in seawater and in the range of 20e400 ng g�1 insediments (Ullrich et al., 2001). These data indicate that the threestudied sites were not affected by Hg contamination and thebivalves were exposed to a similarly low level of Hg.
3.2. Interspecific accumulation of THg and MeHgin bivalves from the filed
THg andMeHg concentrations in each bivalve species are shownin Table 1 and Fig. 1. Overall, the bivalves showed various accu-mulation patterns for Hg in terms of concentration and theMeHg toTHg ratio. THg concentration was the highest in the black musselsS. virgatus (67.7e165.1 ng g�1, mean of 91.9 ng g�1), followed byoysters S. cucullata (34.3e106.4 ng g�1, mean of 70.3 ng g�1) andscallops C. nobilis (39.8e99.5 ng g�1, mean of 60.3 ng g�1). ClamsR. philippinarum and green mussels P. viridis accumulated the least
THg, with an average concentration of 47.4 ng g�1
(32.4e108.5 ng g�1) and 30.1 ng g�1 (24.7e42.9 ng g�1), respec-tively. In contrast, the total dissolved Hg concentration from theirsites of collection was the highest (0.35 ng L�1, Tolo Harbor)(Table 1). With regard to MeHg, the scallops had the highestaverage MeHg concentration of 18.8 ng g�1. Comparable MeHgconcentrations were observed for oysters and clams (15.2 ng g�1
and 16.6 ng g�1, respectively), which were higher than in greenmussels and black mussels (8.5 ng g�1 and 10.4 ng g�1 respec-tively). The effect of growth dilution on Hg accumulation was onlyobserved in scallops and green mussels (Fig. 1, Table 2). Interest-ingly, the proportion of MeHg also varied among bivalve species.For example, the black mussels typically had a lower MeHg to THgratio of 11% (Table 1), three times lower than that of clams (37%).
3.3. Biodynamics of Hg(II) and MeHg in the bivalves and itsrelationship with subcellular distribution of Hg(II) and MeHg
The dissolved uptake rate of Hg(II) and MeHg increased linearlywith exposure concentration (Fig. 2). The ku (calculated by Iu/Cw) ofHg(II) for the scallops was 32.8 L g�1 d�1, approximately three timesthat of the greenmussels (11.4 L g�1 d�1) and clams R. philippinarum

Table 2Summary of biokinetics parameters in five species of bivalves (Mean� SD). ku: dissolved uptake constant; AE: assimilation efficiency (n¼ 10); ke: efflux rate constant (n ¼ 10);aw: absorption efficiency; FR: filtration rate (from Pan and Wang, 2009a, b).
Species ku(L g�1 day�1)
FR(L g�1 h�1)
aw (%) AE (%) ke (day�1) Biological half-life(t1/2 ¼ ln2/ke, days)
Hg(II) MeHg Hg(II) MeHg Hg(II) MeHg Hg(II) MeHg Hg(II) MeHg
Scallop C. nobilis 32.8 595.8 36.4 � 9.4 3.8 68.2 15.5e28.4 50.1e86.3 0.042 � 0.007 0.005 � 0.003 16.5 138.6Clam R. philippinarum 8.9 82.4 4.79 � 1.87 7.7 71.7 9.2e46.3 67.5e86.0 0.039 � 0.007 0.005 � 0.002 17.8 138.6Oyster S. cucullata 4.4 62.5 3.29 � 1.76 5.5 79.2 18.4e48.2 70.5e91.9 0.021 � 0.005 0.003 � 0.001 33.0 231.0Green mussel P. viridis 11.4 132.7 12.5 � 3.33 3.8 44.2 5.5e31.6 52.9e76.5 0.060 � 0.018 0.007 � 0.006 11.6 99.0Black mussel S. virgatus 3.5 71.4 6.25 � 1.07 2.8 56.7 10.6e34.1 72.5e81.5 0.035 � 0.006 0.006 � 0.002 19.8 115.5
100 100
ined
K. Pan, W.-X. Wang / Environmental Pollution 159 (2011) 2500e2506 2503
(8.9 L g�1 d�1). Oysters S. cucullata and the black mussels hadcomparatively lower kus (4.4 and 3.5 L g�1 d�1 respectively).Remarkably high kus of MeHg were observed in the bivalves, whichwere 10e20 times higher than those of Hg(II). The ku of MeHg wasthe highest for the scallops (596 L g�1 d�1), and the lowest for theoysters (62.5 L g�1 d�1). The absorption efficiency (aw) was 3e8% forHg(II) and 44e79% for MeHg (Table 2).
The depuration patterns of radiotracers 203Hg(II) and Me203Hgafter the pulse feeding in the five bivalve species are shown inFig. 3. The radioactivity of 203Hg adsorbed on the shells was lessthan 5% of the overall accumulation. With diatoms as food, theoysters and clams had relatively higher AEs of 46% and 47%respectively, while the other bivalve species had an AE of around30% for Hg(II). MeHg was more efficiently absorbed from food thanHg(II) regardless of the type of food. Nearly 80e90% of MeHg indiatoms and 50e72% MeHg in sediments was assimilated by thebivalves. The AEs of Hg(II) from sediments were only 5e20%. Thereported AEs of Hg(II) and MeHg for zooplanktonwere only around15% and 60%, respectively; for a deposit-feeding polychaete Nereissuccinea the values were 7e28% and 66e84%, respectively (Masonet al., 1995, 1996; Wang et al., 1998). The high kus and AEs indi-cate that the bivalves have a strong ability to absorb Hg from bothdissolved and dietary phases.
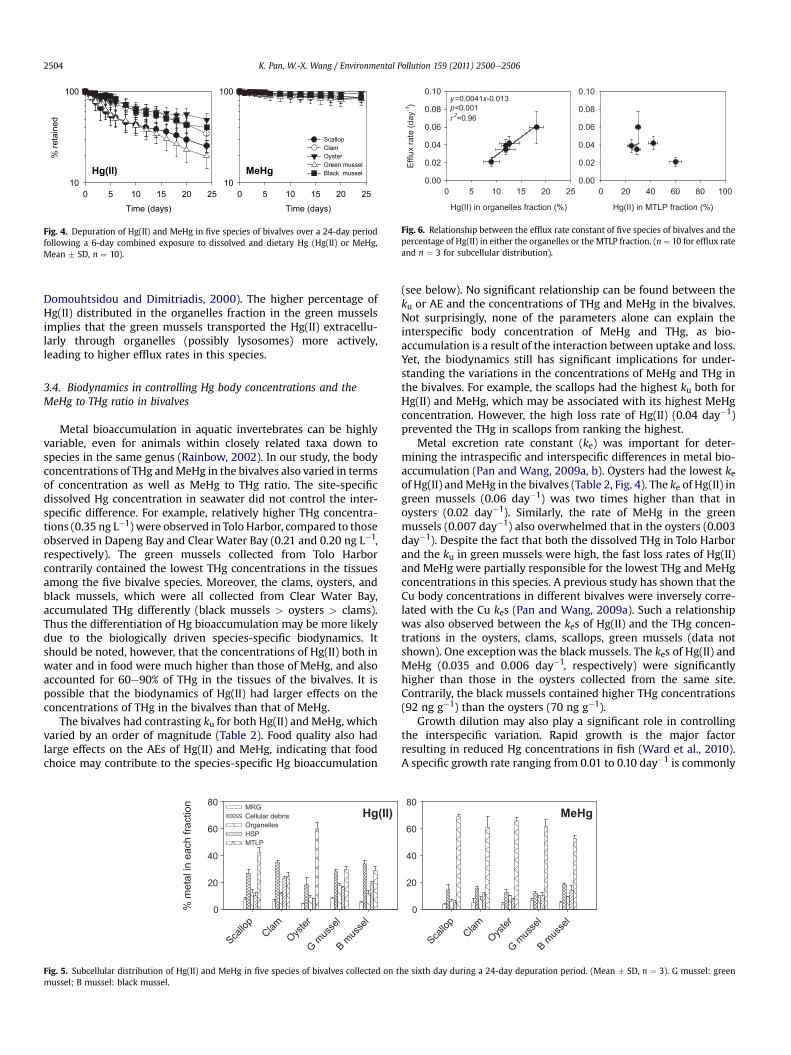
There were considerable differences in the loss rates of Hg(II) inthe bivalves (Fig. 4, Table 2). The green mussels had the highestefflux rate (0.060 day�1), followed by the scallops (0.042 day�1), theclams (0.039 day�1) and the black mussels (0.035 day�1) (Table 2).The oysters had the slowest efflux rate for Hg(II) of 0.021 day�1.Depuration of MeHg was clearly slower than that of Hg(II) inbivalves. Again, the oysters had the lowest efflux rate for MeHg(0.003 day�1), while the green mussels and the black mussels hadrelatively higher rates (0.006e0.007 day�1). The biological half-life(t1/2 ¼ ln2/ke) for Hg(II) in bivalves was only 12e33 days, but was99e231 days for MeHg.
Hg(II)
Dissolved concentrations (µg L-1)
0.01 0.1 1
Influ
x ra
te (µ
g g
d )-1
-1
0.01
0.1
1
10
100
1000
MeHg
0.01 0.1 10.01
0.1
1
10
100
1000Scallop ClamOysterGreen mussel Black mussel
Fig. 2. Influx rates of Hg(II) and MeHg in five species of bivalves at different ambientconcentrations. Data presented as mean � SD (n ¼ 4). The relationships between metalinflux rate (Iw, mg g�1 d�1) and metal concentration in the dissolved phase (Cw, mg L�1)in the bivalves are as follows (r2 ¼ 0.99, p < 0.001). For Hg(II), Iw ¼ 32.8Cw1.02 forscallops; Iw ¼ 8.9Cw0.87 for clams; Iw ¼ 4.4Cw1.05 for oyster; Iw ¼ 11.4Cw1.06 for greenmussels; Iw ¼ 3.5Cw1.29 for black mussels; For MeHg, Iw ¼ 595.8Cw1.16 for scallops;Iw ¼ 82.4Cw1.13 for clams; Iw ¼ 62.5Cw1.14 for oysters; Iw ¼ 132.7Cw1.24 for green mussels;Iw ¼ 71.4Cw0.96 for black mussels.
The subcellular distributions of Hg(II) and MeHg in the fivebivalve species are shown in Fig. 5. Both Hg(II) and MeHg weremainly distributed in MTLP (30e60% for Hg(II) and 60% for MeHg).MT can be a rather important protein for Hg, as both Hg(II) andMeHg are characterized by an extremely high affinity for SH-residues (Viarengo et al., 1994; Canesi et al., 1999). There was alsoa major difference in the distributions of Hg(II) among the fivebivalve species. The oysters typically stored over 60% of the accu-mulated Hg(II) in the MTLP. This was obviously higher than in theother four species. The scallops stored 40% of Hg(II) in this fractionand the percentages for the other three species were similar (30%).The green mussels had about 20% of Hg(II) distributed in theorganelles fraction, whereas the oysters had 9%. For MeHg, thebivalves shared rather similar partitioning patterns, in which theMeHg in MTLP fraction contributed to around 60% of the overallMeHg and other fractions only made up a minor portion.
A significant positive relationship was found between the effluxrate of Hg(II) and the percentage of Hg(II) distribution in theorganelles fraction, indicating that this subcellular fraction hasa close relationship with the Hg(II) elimination process (Fig. 6).Besides being stored in MT, metals could be diverted from MT andother cytosolic ligands to various forms of insoluble storage, wherethe organelles lysosomes may play a significant role (Fowler, 1978;Marigömez et al., 2002). In bivalves, mussels in particular, theabundant lysosomes for intracellular and extracellular digestion areparticularly important organelles for forming vesicle-bound gran-ules which are further eliminated from the body by exocytosisin digestive gland (Langston et al., 1998; Dimitriadis et al., 2003;
MeHg-Sediment
Time (h)
0 10 20 30 40 501
10
100
Hg(II)-Algae
0 10 20 30 40 50
% re
tain
ed
1
10
MeHg-Algae
0 10 20 30 40 501
10Scallop Clam Oyster Green mussel Black mussel
Hg(II)-Sediment
Time (h)
0 10 20 30 40 50
% re
ta
1
10
100
Fig. 3. Retention of Hg(II) and MeHg in five species of bivalves following a pulsefeeding of radiolabeled diatoms or sediments. Data presented as mean � SD (n ¼ 10).
MeHg
0 5 10 15 20 2510
100
Hg(II)
Time (days)0 5 10 15 20 25
% re
tain
ed
10
100
Scallop Clam Oyster Green mussel Black mussel
Time (days)
Fig. 4. Depuration of Hg(II) and MeHg in five species of bivalves over a 24-day periodfollowing a 6-day combined exposure to dissolved and dietary Hg (Hg(II) or MeHg,Mean � SD, n ¼ 10).
Hg(II) in organelles fraction (%)
0 5 10 15 20 25
Efflu
x ra
te (d
ay )
-1
0.00
0.02
0.04
0.06
0.08
0.10
Hg(II) in MTLP fraction (%)
0 20 40 60 80 1000.00
0.02
0.04
0.06
0.08
0.10 y =0.0041 x -0.013 p<0.001 r
2=0.96
Fig. 6. Relationship between the efflux rate constant of five species of bivalves and thepercentage of Hg(II) in either the organelles or the MTLP fraction. (n ¼ 10 for efflux rateand n ¼ 3 for subcellular distribution).
K. Pan, W.-X. Wang / Environmental Pollution 159 (2011) 2500e25062504
Domouhtsidou and Dimitriadis, 2000). The higher percentage ofHg(II) distributed in the organelles fraction in the green musselsimplies that the green mussels transported the Hg(II) extracellu-larly through organelles (possibly lysosomes) more actively,leading to higher efflux rates in this species.
3.4. Biodynamics in controlling Hg body concentrations and theMeHg to THg ratio in bivalves
Metal bioaccumulation in aquatic invertebrates can be highlyvariable, even for animals within closely related taxa down tospecies in the same genus (Rainbow, 2002). In our study, the bodyconcentrations of THg andMeHg in the bivalves also varied in termsof concentration as well as MeHg to THg ratio. The site-specificdissolved Hg concentration in seawater did not control the inter-specific difference. For example, relatively higher THg concentra-tions (0.35 ng L�1) were observed in Tolo Harbor, compared to thoseobserved in Dapeng Bay and Clear Water Bay (0.21 and 0.20 ng L�1,respectively). The green mussels collected from Tolo Harborcontrarily contained the lowest THg concentrations in the tissuesamong the five bivalve species. Moreover, the clams, oysters, andblack mussels, which were all collected from Clear Water Bay,accumulated THg differently (black mussels > oysters > clams).Thus the differentiation of Hg bioaccumulation may be more likelydue to the biologically driven species-specific biodynamics. Itshould be noted, however, that the concentrations of Hg(II) both inwater and in food were much higher than those of MeHg, and alsoaccounted for 60e90% of THg in the tissues of the bivalves. It ispossible that the biodynamics of Hg(II) had larger effects on theconcentrations of THg in the bivalves than that of MeHg.
The bivalves had contrasting ku for both Hg(II) andMeHg, whichvaried by an order of magnitude (Table 2). Food quality also hadlarge effects on the AEs of Hg(II) and MeHg, indicating that foodchoice may contribute to the species-specific Hg bioaccumulation
Scallo
p Clam
Oyster
B mus
sel
G mus
sel
0
20
40
60
80 MRG Cellular debris Organelles HSP MTLP
Hg(II)
% m
etal
in e
ach
fract
ion
Fig. 5. Subcellular distribution of Hg(II) and MeHg in five species of bivalves collected on tmussel; B mussel: black mussel.
(see below). No significant relationship can be found between theku or AE and the concentrations of THg and MeHg in the bivalves.Not surprisingly, none of the parameters alone can explain theinterspecific body concentration of MeHg and THg, as bio-accumulation is a result of the interaction between uptake and loss.Yet, the biodynamics still has significant implications for under-standing the variations in the concentrations of MeHg and THg inthe bivalves. For example, the scallops had the highest ku both forHg(II) and MeHg, which may be associated with its highest MeHgconcentration. However, the high loss rate of Hg(II) (0.04 day�1)prevented the THg in scallops from ranking the highest.
Metal excretion rate constant (ke) was important for deter-mining the intraspecific and interspecific differences in metal bio-accumulation (Pan and Wang, 2009a, b). Oysters had the lowest keof Hg(II) andMeHg in the bivalves (Table 2, Fig. 4). The ke of Hg(II) ingreen mussels (0.06 day�1) was two times higher than that inoysters (0.02 day�1). Similarly, the rate of MeHg in the greenmussels (0.007 day�1) also overwhelmed that in the oysters (0.003day�1). Despite the fact that both the dissolved THg in Tolo Harborand the ku in green mussels were high, the fast loss rates of Hg(II)and MeHg were partially responsible for the lowest THg and MeHgconcentrations in this species. A previous study has shown that theCu body concentrations in different bivalves were inversely corre-lated with the Cu kes (Pan and Wang, 2009a). Such a relationshipwas also observed between the kes of Hg(II) and the THg concen-trations in the oysters, clams, scallops, green mussels (data notshown). One exceptionwas the black mussels. The kes of Hg(II) andMeHg (0.035 and 0.006 day�1, respectively) were significantlyhigher than those in the oysters collected from the same site.Contrarily, the black mussels contained higher THg concentrations(92 ng g�1) than the oysters (70 ng g�1).
Growth dilution may also play a significant role in controllingthe interspecific variation. Rapid growth is the major factorresulting in reduced Hg concentrations in fish (Ward et al., 2010).A specific growth rate ranging from 0.01 to 0.10 day�1 is commonly
Scallo
p Clam
Oyster
B mus
sel
G mus
sel
0
20
40
60
80 MeHg
he sixth day during a 24-day depuration period. (Mean � SD, n ¼ 3). G mussel: green

Table 3The values of d13C and d15N in five species of bivalves (mean � SD, n ¼ 3).
Species Collection site d13C (&) d15N (&)
Scallop C. nobilis Dapeng Bay �18.7 � 0.1 9.0 � 0.1Green mussel P. viridis Tolo Harbor �20.7 � 0.0 8.6 � 0.2Clam R. philippinarum Clear Water Bay �17.1 � 0.0 8.3 � 0.2Oyster S. cucullata Clear Water Bay �18.8 � 0.0 8.5 � 0.1Black mussel S. virgatus Clear Water Bay �19.5 � 0.0 8.0 � 0.2
K. Pan, W.-X. Wang / Environmental Pollution 159 (2011) 2500e2506 2505
observed for marine bivalves (Clausen and Riisgärd, 1996;Jørgensen, 1996). Thus, growth dilution can contribute to 10e30%of the overall efflux for Hg(II) when the specific growth rateconstant (g) is 0.01 per day, but can be up to 60e80% when the g isas high as 0.10 per day. It is possible that the five bivalve specieshave different g and thus have different degrees of dilution for THgand MeHg. However, the effects of growth may be complicated bythe rate of weight change compared to the rate of uptake and lossfor Hg, which may be the reason that the effect of growth dilutionon Hg accumulation was only observed in scallops and greenmussels (Fig. 1).
Biodynamics not only controls the body concentrations of Hg(II)and MeHg, but also has confounding effects on the MeHg to THgratios. Bivalves effectively absorbed Hg(II) and MeHg from bothdissolved and dietary phases, with MeHg being accumulated moreefficiently. Consequently, the MeHg to THg ratios in bivalves weresignificantly elevated compared to those in water and sediments(less than 0.5%, Ullrich et al., 2001). Although MeHg concentrationsin bivalves did not vary widely (a difference ofw2 times), therewasa difference of nearly four times (11e37%) in the MeHg to THg ratio(Table 1). On the one hand, the relatively low percentage of Hg inthe form of MeHg in bivalves was largely determined by the muchhigher concentrations of Hg(II) in water and food as compared toMeHg. On the other hand, the difference in the MeHg to THg ratiowas affected by the differences between the biodynamics of Hg(II)and MeHg in each species. However, it appeared that the ku and AEcould not explain the species-dependent ratio. For example,difference in ku between Hg(II) and MeHg was most pronounced inthe black mussel (a difference ofw20 times), whereas the ratio wasthe lowest in this species. The proportion of MeHg was positivelycorrelated with the ratio of the efflux rate of Hg(II) to that of MeHg(Fig. 7, p ¼ 0.059), indicating that the species-specific MeHg to THgratio was possibly controlled by the relative difference between theefflux rate of Hg(II) andMeHg in each species. The ratio of the effluxrate of Hg(II) to that of MeHg in bivalves was found to be 6e9.A similar difference (3e7 times) has also been observed in fish(Wang and Wong, 2003; Wang et al., 2010). However, thepredominance of dietary exposure and ingestion of food of a higherMeHg to THg ratio made the proportion of MeHg in high-trophic-level fishes overwhelm that in bivalves.
3.5. Feeding niche of bivalves and its effects on Hg accumulation
While the species-specific accumulation of Hgwas driven by thedetoxification strategies of bivalves, the impacts of ecologicalfactors cannot be ruled out. Field studies have shown an average15N enrichment of 3.4& per trophic level, and a lower value of 2.5&was observed for herbivores (Vander Zanden and Rasmussen,
ke(Hg(II)) /ke(MeHg)
5 6 7 8 9 10
MeH
g/TH
g ra
tio (%
)
0
10
20
30
40
50
y=7.84x-32.4p=0.059r
2
=0.74
Fig. 7. Relationship between the ratio of MeHg to THg and the ratio of the eliminationrates of Hg(II) and MeHg.
2001). The d15N only varied within a small range of 1& betweenthe five bivalve species (Table 3), implying a similar trophic levelbetween the bivalves. There were however significant differencesin d13C signals among the five species of bivalves (Table 3), indi-cating that they may rely on different carbon sources. The d13Csignals varied largely between clams (�17.1&) and black mussels(�19.5&) collected from Clear Water Bay, with an intermediatevalue of �18.8& for oysters. Numerous studies have shown thatphytoplankton or SPM are more depleted in d13C than sedimentaryorganic matter (SOM) or benthic microalgae (France, 1995;Middelburg and Nieuwenhuize, 1998; Ishihi, 2003), resulting inpelagic feeders that are more depleted in d13C than benthicconsumers (Hobson et al., 1995). For example, Currin et al. (1995)suggested that an average d13C of �14.9& was found for benthicmicroalgae and an average of �21.1& for marine phytoplankton. Inour study, the clams appeared to feed on resuspended sediments,whereas the blackmussels relied on phytoplankton or SPM inwatercolumn. The rock oysters possibly relied on both sediments andSPM. The different food sources and habitats may lead to differentbioavailability of Hg for clams, oysters and black mussels. It wasfound that the THg concentration in the SPM (w100 ng g�1) washigher than that in the sediments (w65 ng g�1), while the AEs ofHg(II) and MeHg were both lower for sediments than phyto-plankton. The black mussels may have a higher potential foraccumulating Hg from food than the clams and oysters. As a result,the concentration of THg in black mussels was much higher thanthat in oysters even though the oysters had the lowest eliminationrates of both Hg(II) and MeHg.
4. Conclusions
Our study has clearly demonstrated that bivalves havesubstantial capacity to accumulate Hg(II) andMeHg fromwater andfood. All five bivalve species, scallops, clams, oysters, green musselsand black mussels, significantly accumulated Hg from the low Hgenvironments. The bivalves showed distinct detoxification strate-gies for Hg, which was reflected in their subcellular partitioningand biodynamics. Hg(II) distributing in the organelles fraction wasclosely related to the species-dependent Hg(II) elimination rate,suggesting an interaction between the subcellular partitioning andbiokinetics of Hg(II) in bivalves. The biodynamics parameters wereonly able to partially explain the different body concentrations ofTHg and MeHg in bivalves, and the variations in the MeHg to THgratios were largely controlled by the relative difference betweenthe elimination rates of Hg(II) and MeHg. The interspecific bodyconcentrations of MeHg and THg were complicated by the differ-ences in the biodynamics of Hg(II) and MeHg, and the differentfeeding niches of bivalves.
Acknowledgements
We thank Prof. Ling-Feng Huang and his student Mr. Xin-QingZheng of Xiamen University for their help in analyzing stableisotopes in this study. This study was supported by the GeneralResearch Funds from the Hong Kong Research Grants Council(663009 and 662610).

K. Pan, W.-X. Wang / Environmental Pollution 159 (2011) 2500e25062506
References
Canesi, L., Viarengo, A., Leonzio, C., Filippelli, M., Gallo, G., 1999. Heavy metals andglutathione metabolism in mussel tissues. Aquatic Toxicology 46, 67e76.
Chen, C.Y., Serrell, N., Evers, D.C., Fleishman, B.J., Lambert, K.F., Weiss, J., Mason, R.P.,Bank, M.S., 2008. Meeting report: methylmercury in marine ecosystems-fromsources to seafood consumers. Environmental Health Perspectives 116,1706e1712.
Chen, C.Y., Dionne, M., Mayes, B.M., Ward, D.M., Sturup, S., Jackson, B.P., 2009.Mercury bioavailability and bioaccumulation in estuarine food webs in the gulfof Maine. Environmental Science and Technology 43, 1804e1810.
Clausen, I., Riisgärd, H.U., 1996. Growth, filtration and respiration in the musselMytilus edulis: no evidence for physiological regulation of the filter-pump tonutritional needs. Marine Ecology Progress Series 141, 37e45.
Currin, C.A., Newell, S.Y., Paerl, H.W., 1995. The role of standing dead spartina-alterniflora and benthic microalgae in salt-marsh food webs e considerationsbased on multiple stable-isotope analysis. Marine Ecology Progress Series 121,99e116.
Dallinger, R., 1993. Strategies of metal detoxification in terrestrial invertebrates. In:Dallinger, R., Rainbow, P.S. (Eds.), Ecotoxicology of Metals in Invertebrates.Lewis Publisher, Boca Raton, USA, pp. 245e289.
Dimitriadis, V.K., Domouhtsidou, G.P., Raftopoulou, E., 2003. Localization of Hg andPb in the palps, the digestive gland and the gills in Mytilus galloprovincialis (L.)using autometallography and X-ray microanalysis. Environmental Pollution125, 345e353.
Ditri, F.M., 1991. Mercury contamination e what we have learned since Minamata.Environmental Monitoring and Assessment 19, 165e182.
Domouhtsidou, G.P., Dimitriadis, V.K., 2000. Ultrastructural localization of heavymetals (Hg, Ag, Pb, and Cu) in gills and digestive gland of mussels, Mytilusgalloprovincialis (L.). Archives of Environmental Contamination and Toxicology38, 472e478.
Fialkowski, W., Rainbow, P.S., Smith, B.D., Zmudzinski, L., 2003. Seasonal variation intrace metal concentrations in three talitrid amphipods from the gulf of Gdansk,Poland. Journal of Experimental Marine Biology and Ecology 288, 81e93.
Fowler, B.A., 1978. General subcellular effects of lead, mercury, cadmium, andarsenic. Environmental Health Perspectives 22, 37e41.
France, R.L., 1995. C-13 enrichment in benthic compared to planktonic algae e
foodweb implications. Marine Ecology Progress Series 124, 307e312.Gagnon, C., Fisher, N.S., 1997. Bioavailability of sediment-bound methyl and inor-
ganic mercury to a marine bivalve. Environmental Science and Technology 31,993e998.
Herman, P.M.J., Middelburg, J.J., Widdows, J., Lucas, C.H., Heip, C.H.R., 2000. Stableisotopes’ as trophic tracers: combining field sampling and manipulative label-ling of food resources for macrobenthos. Marine Ecology Progress Series 204,79e92.
Hobson, K.A., Ambrose, W.G., Renaud, P.E., 1995. Sources of primary production,benthic-pelagic coupling, and trophic relationships within the Northeast waterPolynya: insights from delta C-13 and delta N-15 analysis. Marine EcologyProgress Series 128, 1e10.
Inza, B., Ribeyre, F., Boudou, A., 1998. Dynamics of cadmium and mercurycompounds (inorganic mercury or methylmercury): uptake and depuration inCorbicula fluminea. Effects of temperature and pH. Aquatic Toxicology 43,273e285.
Ishihi, Y., 2003. Feeding of the bivalve Theora lubrica on benthic microalgae: isotopicevidence. Marine Ecology Progress Series 255, 303e309.
Jørgensen, C.B., 1996. Bivalve filter feeding revisited. Marine Ecology Progress Series142, 287e302.
Langston, W.J., Bebianno, M.J., Burt, G.R., 1998. Metal handling strategies inmolluscs. In: Langston, W.J., Bebianno, M.J. (Eds.), Metal Metabolism in AquaticEnvironments. Chapman & Hall, London, pp. 219e283.
Marigömez, I., Soto, M., Cajaraville, M.P., Angulo, E., Giamberini, L., 2002. Cellularand subcellular distribution of metals in molluscs. Microscopy Research andTechnique 56, 358e392.
Mason, R.P., Reinfelder, J.R., Morel, F.M.M., 1995. Bioaccumulation of mercury andmethylmercury. Water Air and Soil Pollution 80, 915e921.
Mason, R.P., Reinfelder, J.R., Morel, F.M.M., 1996. Uptake, toxicity, and trophictransfer of mercury in a coastal diatom. Environmental Science and Technology30, 1835e1845.
Middelburg, J.J., Nieuwenhuize, J., 1998. Carbon and nitrogen stable isotopes insuspended matter and sediments from the Schelde estuary. Marine Chemistry60, 217e225.
Morel, F.M.M., Kraepiel, A.M.L., Amyot, M., 1998. The chemical cycle and bio-accumulation of mercury. Annual Review of Ecology and Systematics 29,543e566.
Pan, J.F., Wang, W.-X., 2004. Uptake of Hg(II) and methylmercury by the greenmussel Perna viridis under different organic carbon conditions. Marine EcologyProgress Series 276, 125e136.
Pan, K., Wang, W.-X., 2008. Validation of biokinetic model of metals in the scallopChlamys nobilis in complex field environments. Environmental Science andTechnology 42, 6285e6290.
Pan, K., Wang, W.-X., 2009a. Biodynamics to explain the difference of copper bodyconcentrations in five marine bivalve species. Environmental Science andTechnology 43, 2137e2143.
Pan, K., Wang, W.-X., 2009b. Inter-individual variations in cadmium and zincbiodynamics in the scallop Chlamys nobilis. Marine Ecology Progress Series 383,151e160.
Rainbow, P.S., 1995. Biomonitoring of heavy metal availability in the marine envi-ronment. Marine Pollution Bulletin 31, 183e192.
Rainbow, P.S., 2002. Trace metal concentrations in aquatic invertebrates: why andso what? Environmental Pollution 120, 497e507.
Ross, S.T., 1986. Resource partitioning in fish assemblages e a review of fieldstudies. Copeia, 352e388.
Rouleau, C., Block, M., 1997. Working methods paper e fast and high-yield synthesis ofradioactive (CH3Hg)-Hg-203(II). Applied Organometallic Chemistry 11, 751e753.
Tsui, M.T.K., Wang, W.-X., 2004. Temperature influences on the accumulation andelimination of mercury in a freshwater cladoceran, Daphnia magna. AquaticToxicology 70, 245e256.
Ullrich, S.M., Tanton, T.W., Abdrashitova, S.A., 2001. Mercury in the aquatic envi-ronment: a review of factors affecting methylation. Critical Reviews in Envi-ronmental Science and Technology 31, 241e293.
Vander Zanden, M.J., Rasmussen, J.B., 2001. Variation in delta N-15 and delta C-13trophic fractionation: implications for aquatic food web studies. Limnology andOceanography 46, 2061e2066.
Viarengo, A., Canesi, L., Moore, M.N., Orunesu, M., 1994. Effects of Hg2þ and Cu2þ onthe cytosolic Ca2þ level in molluscan blood-cells evaluated by confocalmicroscopy and spectrofluorometry. Marine Biology 119, 557e564.
Wallace, W.G., Lee, B.G., Luoma, S.N., 2003. Subcellular compartmentalization of Cdand Zn in two bivalves. I. Significance of metal-sensitive fractions (MSF) andbiologically detoxified metal (BDM). Marine Ecology Progress Series 249,183e197.
Wang, W.-X., Fisher, N.S., 1996. Assimilation of trace elements and carbon by themussel Mytilus edulis: effects of food composition. Limnology and Oceanog-raphy 41, 197e207.
Wang, R., Wang, W.-X., 2010. Importance of speciation in understanding mercurybioaccumulation in tilapia controlled by salinity and dissolved organic matter.Environmental Science and Technology 44, 7964e7969.
Wang, W.-X., Wong, R.S.K., 2003. Bioaccumulation kinetics and exposure pathwaysof inorganic mercury and methylmercury in a marine fish, the sweetlips Plec-torhinchus gibbosus. Marine Ecology Progress Series 261, 257e268.
Wang, W.-X., Stupakoff, I., Gagnon, C., Fisher, N.S., 1998. Bioavailability of inorganicand methylmercury to a marine deposit feeding polychaete. EnvironmentalScience and Technology 32, 2564e2571.
Wang, R., Wong, M.H., Wang, W.-X., 2010. Mercury exposure in the freshwatertilapia Oreochromis niloticus. Environmental Pollution 158, 2694e2701.
Ward, D.M., Nislow, K.H., Chen, C.Y., Folt, C.L., 2010. Rapid, efficient growth reducesmercury concentrations in stream-dwelling Atlantic salmon. Transactions of theAmerican Fisheries Society 139, 1e10.
Watras, C.J., Bloom, N.S., 1992. Mercury and methylmercury in individualzooplankton e implications for bioaccumulation. Limnology and Oceanography37, 1313e1318.