mechanism of the ketogenic effect of ammonium chloride · 264 ketogenic effect of ammonium chloride...
TRANSCRIPT

MECHANISM OF THE KETOGENIC EFFECT OF AMMONIUM CHLORIDE
BY RICHARD 0. RECKNAGEL* AND VAN R. POTTER
(FTo~ the McAT& ikfemorial Laboratory, Medical School, University of Wisconsin, Madison, Wisconsin)
(Received for publication, January 9, 1951)
When ammonium chloride is added to liver mince (1) or to rat liver slices (2), there is an increase in ketone body production. In the ab- sence of other possibilities, it was earlier suggested (3, 4) that the ketone bodies arising on addition of ammonium chloride were coming from ace- tate formed during the “acetylation amination” of cr-keto acids. Ac- cording to this view, amination of a-keto acids is an oxidative process involving condensation with ammonia and pyruvate to yield carbon di- oxide and the corresponding N-acetylamino acid, which, on hydrolysis, would then yield the corresponding a-amino acid and acetate, the latter condensing to form acetoacetate. However, it has not been possible to bring experimental evidence to bear on the formation of ketone bodies through such a mechanism.
More recently, a knowledge of the reductive amination of cr-ketoglutaric acid to glutamic acid (5-7) has led to the concept of ammonia fixation by reductive amination, followed by transamination (8-10). In addi- tion, experiments with liver homogenate by Krebs et al. (ll), and un- published experiments’ from this laboratory, indicated that the primary locus of action of added ammonia, even under aerobic conditions, was in the reductive amination of cr-ketoglutaric acid. Meanwhile, the ex- periments of Lehninger (12) had clearly shown that ketone body pre- cursors (pyruvate, octanoate) would form acetoacetate in liver in the absence of oxalacetate, and that degradation via the Krebs citric acid cycle was the preferred pathway provided oxalacetate was available. These considerations suggested that the ketosis induced by ammonium chloride might be due fundamentally to the absence of oxalacetate, a con- dition brought about by the preferential removal of cr-ketoglutarate, in the presence of added ammonia, to glutamic acid. The experimental data presented in this paper support such a mechanism for the ketosis induced by ammonium chloride.
* National Research Council Fellow in Zoology,1949-50. 1 Ackermann, W. W., and Potter, V. R., unpublished.
263
by guest on February 14, 2020http://w
ww
.jbc.org/D
ownloaded from

264 KETOGENIC EFFECT OF AMMONIUM CHLORIDE
EXPERIMENTAL
General-The conventional Warburg apparatus was used. Well fed male rats,2 3 to 6 months old, weighing between 200 and 230 gm., were de- capitated, bled, and the livers excised and chilled to 0” before further treatment. Liver slices were cut in the cold to a thickness of 0.5 mm. in an apparatus essentially that of Stadie and Riggs (13), and suspended in Medium Al of Robinson (14), to which the appropriate substrates were added. Glucose was omitted. Oxygen was used in the gas phase; the temperature was 38”. One slice was used per flask. Final dry weights varied between 10 and 20 mg. Data from the liver slice experiments are expressed as & values; i.e., microliters per mg. of dry weight per hour. 1 PM of any substance is taken as equivalent to 22.4 ~1.
Homogenates were prepared in the Potter and Elvehjem homogenizer (15) at concentrations of 10 to 20 per cent, in an ice-cold medium of the following composition: 5.75 gm. of KCl, 16 mg. of KHCO,, 494 ml. of H20, pH adjusted to 7.6. The ice-cold homogenates were pipetted directly into the iced Warburg flasks containing all the necessary additions, and equilibrated for 10 minutes at 38” before oxygen uptake measurements were made.
The homogenate system employed was essentially the “oxalacetic acid oxidase” system of Potter et al. (16). The main constituents were not varied and had the following final concentrations: 0.067 M KC1 (not in- cluding the KC1 of the homogenate), 3.3 X 10m3 M MgC12, 6.7 X 10e3 M
potassium phosphate buffer of pH 7.4, 1 X 1O-3 M adenosinetriphosphate (K salt). The final pH was 7.2; the final volume was 3.0 ml. Other additions, such as cytochrome c, diphosphopyridine nucleotide (DPN), and nicotinamide, are indicated in connection with the pertinent data.
Pure adenosinetriphosphate and 80 per cent pure DPN were obtained from Dr. G. A. LePage. Cytochrome c was prepared according to the method of Keilin and Hartree (17), as described by Potter ((18) p. 211). The oxalacetic and cr-ketoglutaric acids used were supplied through the courtesy of C. Heidelberger and W. W. Ackermann, respectively.
Analytical Methods
Pyruvic acid disappearance was followed by the direct method for total keto acids (19). Although cr-ketoglutaric and oxalacetic acids react under these conditions, it was shown by chromatography (20) that the level of c+ketoglutaric acid did not exceed 0.2 pM per flask, while the level of oxal- acetic acid was not detectable. Acetoacetic acid interferes to the extent of 10 per cent in the direct method, but the maximum amount present
2 Holtzman-Rolfsmeyer Company, Madison, Wisconsin.
by guest on February 14, 2020http://w
ww
.jbc.org/D
ownloaded from

R. 0. RECKNAGEL AND V. R. POTTER 265
was always small enough to make corrections of little significance. p- Hydroxybutyric acid was determined by the method of Greenberg and Lester (21) after boiling to remove acetoacetic acid and after treating with copper lime (22) and then with Ca(OH)2 alone to remove citric acid. Acetoacetic acid was determined by Lehninger’s modification3 of the method of Greenberg and Lester (21). Protein-free filtrates containing aceto- acetic acid equivalent to 5 to 30 y of acetone were brought to pH 5.0 in a volume of 1.5 ml. Molar acetate buffer (pH 5.0, 0.5 ml.) and 4 M aniline hydrochloride (2.0 ml.) were added, and the mixture was incubated in glass-stoppered test-tubes for 90 minutes at 30”. The acetone 2,4-di- nitrophenylhydrazone was then formed (21) and extracted into 5.0 ml. of CC14 with prolonged (50 minutes) shaking. The CC14 layer was washed twice with H20 and once with 0.5 N NaOH to remove any excess reagent and pyruvic acid 2,4-dinitrophenylhydrazone, and read at 420 mp. p- Hydroxybutyric acid is completely inert in the procedure, and pyruvic acid does not interfere. When this method was tested with acetoacetic acid prepared according to Ljunggren (23)) the results agreed with those obtained by the use of the aniline citrate method ((2)) (18) p. 175) within +3 per cent.
Glutamic acid was measured according to the manometric method of Gale (24). The glutamic decarboxylase of Clostridium welchii SR12 was a gift from Dr. P. P. Cohen. This enzyme has some activity against aspartic acid (10). In some experiments the specific glutamic decarboxy- lase of Escherichia coli (American Type Culture Collection strain 4157) was used. The enzyme was prepared according to Umbreit and Gunsalus (25) with slight modifications. At pH 5.0 and 38”) the enzyme had a QcoZ (glutamic acid) of 60 and was completely inactive against n-aspartic acid and nn-alanine. Lactic acid was measured according to the method of Barker and Summerson (22)) as described by LePage ((18) p. 192)) citric acid by the method of Natelson et al. (26)) as employed by Potter et al. (16). Flask contents were deproteinized by addition of 0.3 ml. of 50 per cent perchloric acid. If glutamic acid was to be measured, 0.5 ml. of 2.00 N HCl was added; the pH was then adjusted to 5.0, and the pre- cipitated protein was removed by centrifugation.
Results
Liver Slices-Four experiments with liver slices from fed rats were in close agreement and completely confirmed the ketogenic effect of am- monium chloride as described by Edson (2). In the presence of 20 PM of pyruvate, the QAeAc was 0.73 without added ammonium chloride. When
8 We are indebted to Dr. A. L. Lehninger for giving us unpublished details, made available here through his permission.
by guest on February 14, 2020http://w
ww
.jbc.org/D
ownloaded from
266 KETOGENIC EFFECT OF AMMONIUM CHLORIDE
ammonium chloride was added to a final concentration of 0.04 M, ace- toacetate production was increased almost C-fold (QAcAc = 2.67). The endogenous production of acetoacetate was low (QAcac = 0.51) and rose to Qua = 0.78 on addition of ammonium chloride. With pyruvate as substrate, /3-hydroxybutyric acid production was increased from &a+-,= = 0.18 without added NH, to 0.26 when ammonium chloride was added. No endogenous /3-hydroxybutyric acid was found. Addition of ammonium
100 K I
QUANTITATIVE CONVERSION ( MALONATE)
01 1 1 I 0 5 IO 20
JJM F”i:RATE 60 80
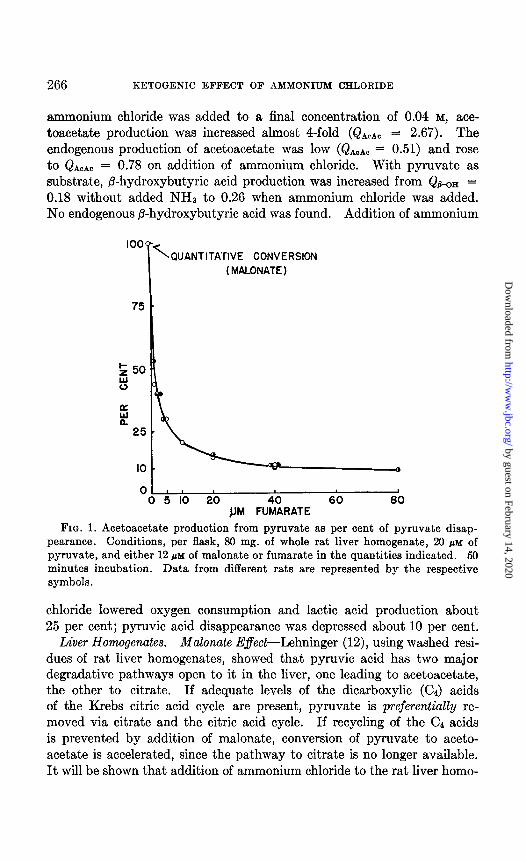
FIG. 1. Acetoacetate production from pyruvate as per cent of pyruvate disap- pearance. Conditions, per flask, 80 mg. of whole rat liver homogenate, 20 PM of pyruvate, and either 12 PM of malonate or fumarate in the quantities indicated. 50 minutes incubation. Data from different rats are represented by the respective symbols.
chloride lowered oxygen consumption and lactic acid production about 25 per cent; pyruvic acid disappearance was depressed about 10 per cent.
Liver Homogenates. Malonate E$ect-Lehninger (12)) using washed resi- dues of rat liver homogenates, showed that pyruvic acid has two major degradative pathways open to it in the liver, one leading to acetoacetate, the other to citrate. If adequate levels of the dicarboxylic (Cd) acids of the Krebs citric acid cycle are present, pyruvate is preferentially re- moved via citrate and the citric acid cycle. If recycling of the Ca acids is prevented by addition of malonate, conversion of pyruvate to aceto- acetate is accelerated, since the pathway to citrate is no longer available. It will be shown that addition of ammonium chloride to the rat liver homo-
by guest on February 14, 2020http://w
ww
.jbc.org/D
ownloaded from

R. 0. RECKNAGEL AND V. R. POTTER 267
genate results in a similar increase in the production of acetoacetate from pyruvate.
We first established the properties of the whole homogenate with respect to the oxidation of pyruvic acid, both in the presence and in the ab- sence of a functioning Krebs citric acid cycle. The data of Fig. 1, com- piled from a number of experiments, confirm Lehninger’s basic observa- tion concerning the alternative pathways open to pyruvate. Without added intermediates of the Krebs citric acid cycle, 75 per cent of added pyruvate is converted to acetoacetate. That this is due simply to the di- lution of the intermediates of the Krebs cycle is evident from the marked depression in acetoacetic acid production resulting from addition of fu- maric acid. Similar results were obtained on addition of oxalacetate, malate, succinate, a-ketoglutarate, and citrate. Even with the highest levels of added fumaric acid, only 1 PM of P-hydroxybutyric acid was found, indicating that the decrease in the yield of acetoacetic acid in the presence of fumarate was not due to extensive formation of /?-hydroxy- butyrate. When malonate was added, pyruvate was converted to acetoacetate quantitatively. These experiments show that in the whole homogenate, in the presence of adequate amounts of the Cd acids, the path- way of pyruvate to citrate has a high priority over the pathway to aceto- acetate.
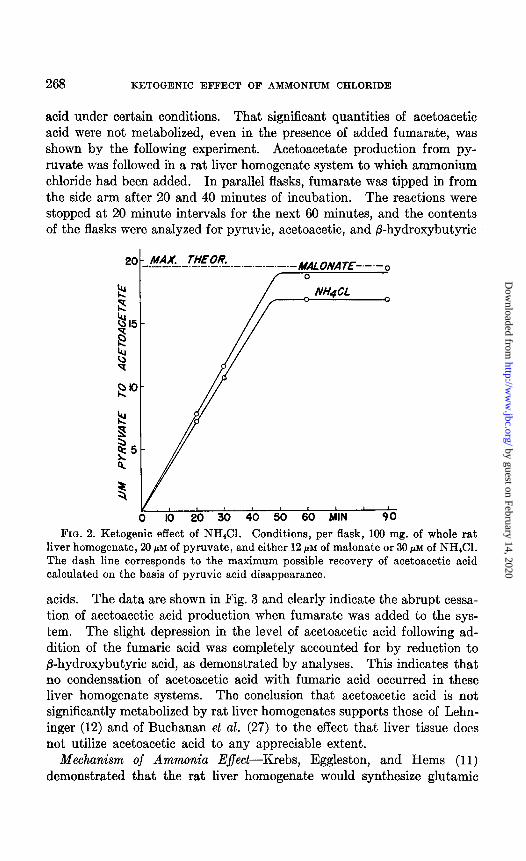
Ammonia Effect-The ketosis resulting from addition of ammonium chloride to the whole homogenate is shown in Fig. 2. In experiments in which the reaction was followed at shorter time intervals, it was estab- lished that the linear rate of conversion of pyruvate to acetoacetate was maintained down to levels of pyruvate approaching 0.1 PM per flask or less. A maximum ketogenic effect can be obtained with as little as 9 PM
of added ammonium chloride. The ketosis induced by ammonia is easily prevented by addition of an equivalent quantity of fumarate. Glutamate acts like a-ketoglutarate, and not like ammonia; i.e., addition of glutamate depresses acetoacetic acid production from pyruvate. Occasionally, in experiments with added ammonium chloride, and rarely with added malo- nate, conversion of pyruvate to acetoacetate was less than quantitative (cf. Fig. 3). The reason for this finding is not clear, but is believed to be due to correspondingly high endogenous levels of the Krebs cycle inter- mediates convertible to oxalacetate under the respective experimental conditions.
Non-Utilization of Acetoacetate-The quantitative estimation of the ex- tent of the conversion of pyruvic acid to acetoacetic acid in these homo- genate systems and the effects of various additions on this conversion depend on the fact that acetoacetate, once formed, is not further metab- olized, except for the very small amounts reduced to fl-hydroxybutyric
by guest on February 14, 2020http://w
ww
.jbc.org/D
ownloaded from
268 KETOGENIC EFFECT OF AMMONIUM CHLORIDE
acid under certain conditions. That significant quantities of acetoacetic acid were not metabolized, even in the presence of added fumarate, was shown by the following experiment. Acetoacetate production from py- ruvate was followed in a rat liver homogenate system to which ammonium chloride had been added. In parallel flasks, fumarate was tipped in from the side arm after 20 and 40 minutes of incubation. The reactions were stopped at 20 minute intervals for the next 60 minutes, and the contents of the flasks were analyzed for pyruvic, acetoacetic, and &hydroxybutyric
20 MAX THEOR. t -_____ ‘--.-.-.-.-.-.-.---.-.-M~O~ATE-.-.-o
/ 0
FIG. 2. Ketogenic effect of NH&l. Conditions, per flask, 100 mg. of whole rat liver homogenate, 20 PM of pyruvate, and either 12 pM of malonate or 30 PM of NH&l. The dash line corresponds to the maximum possible recovery of acetoacetic acid calculated on the basis of pyruvic acid disappearance.
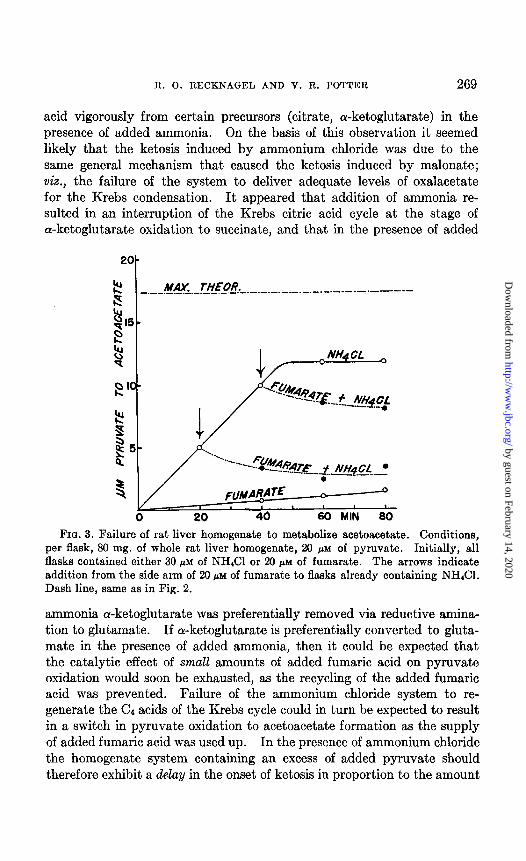
acids. The data are shown in Fig. 3 and clearly indicate the abrupt cessa- tion of acetoacetic acid production when fumarate was added to the sys- tem. The slight depression in the level of acetoacetic acid following ad- dition of the fumaric acid was completely accounted for by reduction to fl-hydroxybutyric acid, as demonstrated by analyses. This indicates that no condensation of acetoacetic acid with fumaric acid occurred in these liver homogenate systems. The conclusion that acetoacetic acid is not significantly metabolized by rat liver homogenates supports those of Lehn- inger (12) and of Buchanan et al. (27) to the effect that liver tissue does not utilize acetoacetic acid to any appreciable extent.
Mechanism of Ammonia E$ect--Krebs, Eggleston, and Hems (11) demonstrated that the rat liver homogenate would synthesize glutamic
by guest on February 14, 2020http://w
ww
.jbc.org/D
ownloaded from
R. 0. RECKNAGEL AND V. R. POTTER 269
acid vigorously from certain precursors (citrate, cu-ketoglutarate) in the presence of added ammonia. On the basis of this observation it seemed likely that the ketosis induced by ammonium chloride was due to the same general mechanism that caused the ketosis induced by malonate; viz., the failure of the system to deliver adequate levels of oxalacetate for the Krebs condensation. It appeared that addition of ammonia re- sulted in an interruption of the Krebs citric acid cycle at the stage of a-ketoglutarate oxidation to succinate, and that in the presence of added
20 t
k MAX THEOR ________. z __._____._. .-.-.-.-----.-----------.- --me E iu $15. 2
0 20 46 60 MIN 50
FIQ. 3. Failure of rat liver homogenate to metabolize acetoacetate. Conditions, per flask, 80 mg. of whole rat liver homogenate, 20 PM of pyruvate. Initially, all flasks contained either 30 pM of NH&l or 20 PM of fumarate. The arrows indicate addition from the side arm of 20 PM of fumarate to flasks already containing NH&I. Dash line, same as in Fig. 2.
ammonia a-ketoglutarate was preferentially removed via reductive amina- tion to glutamate. If cr-ketoglutarate is preferentially converted to gluta- mate in the presence of added ammonia, then it could be expected that the catalytic effect of small amounts of added fumaric acid on pyruvate oxidation would soon be exhausted, as the recycling of the added fumaric acid was prevented. Failure of the ammonium chloride system to re- generate the Cd acids of the Krebs cycle could in turn be expected to result in a switch in pyruvate oxidation to acetoacetate formation as the supply of added fumaric acid was used up. In the presence of ammonium chloride the homogenate system containing an excess of added pyruvate should therefore exhibit a delay in the onset of ketosis in proportion to the amount
by guest on February 14, 2020http://w
ww
.jbc.org/D
ownloaded from

270 KETOGENIC EFFECT OF hMMONIUM CHLORIDE
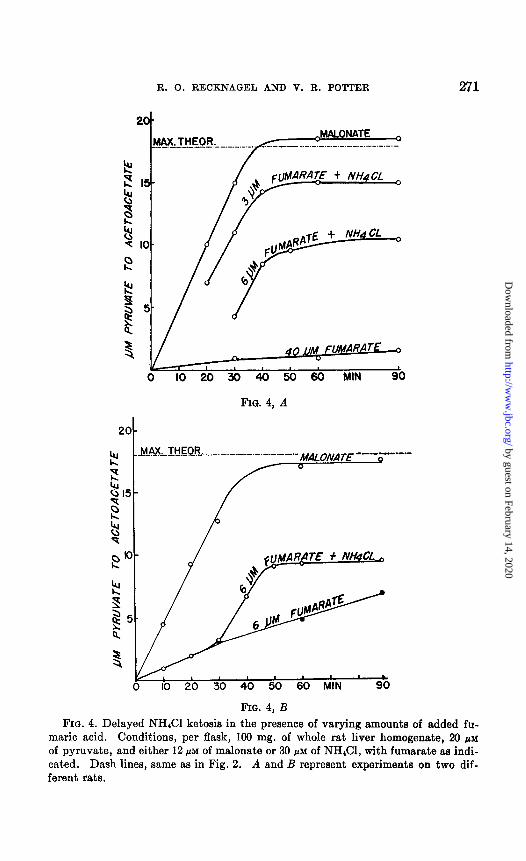
of added fumaric acid. Furthermore, once initiated, the rate of the con- version of pyruvate to acetoacetate should be independent of the time of onset of the ketosis, and, finally, since recycling of the added fumarate is prevented, there should, be a deficit in the final quantities of acetoacetic acid produced corresponding stoichiometrically to the amount of fumarate added. All of these expectations were repeatedly borne out by experiment. Typical data are shown in Fig. 4, A and 4, B, which present data for two different rats.
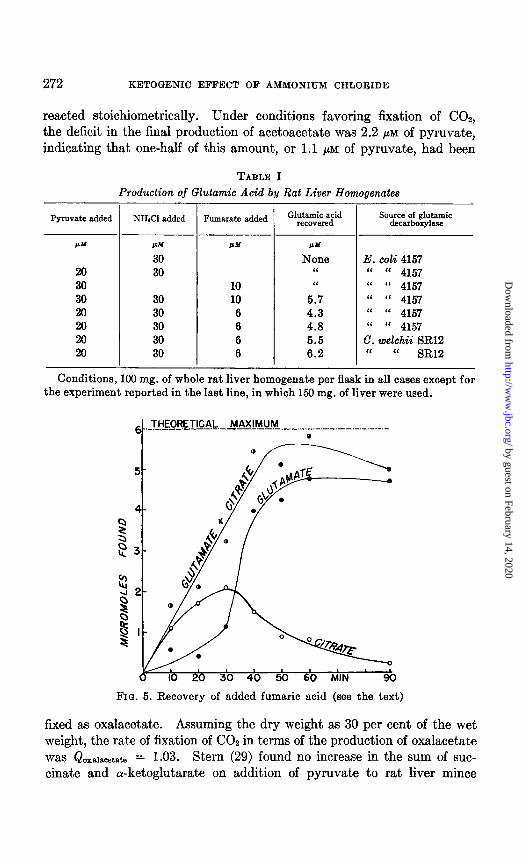
Recovery of Added Fumaric Acid-Net recoveries of glutamic acid from experiments on five different rats are shown in Table I. Addition of fumaric acid to the whole rat liver homogenate, in the presence of pyruvate and ammonium chloride, leads to a marked production of glutamic acid. No net increase in glutamic acid was observed if either ammonium chloride or fumaric acid was omitted from the system. If the theory advanced in this paper to explain the ketogenic effect of ammonium chloride is correct, the added fumaric acid should all be metabolized beyond the stage of oxalacetate at the time of onset of the ketosis, and should therefore be accountable as glutamic and citric acids at this time. In Fig. 5 are shown the results of analyses for these two acids for the experiment re- corded in Fig. 4, B, in which the ketosis began after 30 minutes of incuba- tion. The plot of the sum of citrate plus glutamate reveals that about 85 per cent of the added fumaric acid had already been metabolized past the level of oxalacetate after 40 minutes of incubation. The glutamic decarboxylase of C. welchii was used to measure production of glutamic acid. In a similar experiment in which the glutamic decarboxylase of E. coli was used for measurement of glutamic acid, the sum of citrate plus glutamate accounted for 60 per cent of added fumaric acid at 30 minutes of incubation, but a maximum recovery of only 75 per cent was reached on further incubation. The same early rapid production of citric acid was observed, followed by a fall in the citric acid and a rise in the glutamic acid concentration of the medium. These data support the hypothesis that, in the presence of added ammonium chloride, the onset of the ketosis is coincident with the reduction of the concentration of the Cd acids to a point at which the condensation reaction can no longer occur.
E$ect of CO-Formation of oxalacetate via fixation of CO2 in the Wood- Werkman reaction (28) could be expected to result in a decrease in the conversion of pyruvate to acetoacetate. Data from an experiment de- signed to test this possibility are shown in Table II. Addition of ammonium chloride alone exerted a pronounced ketogenic effect. If 4 PM
of fumarate were added in addition to ammonia, the deficit in the final recovery of acetoacetate corresponded exactly to 4 PM of pyruvate, indi- cating that, in the presence of added ammonia, fumarate and pyruvate
by guest on February 14, 2020http://w
ww
.jbc.org/D
ownloaded from
R. 0. RECKNAGEL AND V. Ft. POTTER 271
4o UM FUMARATE 4o UM FUMARATE a a
0 0 IO IO 20 20 30 30 40 40 50 50 60 60 MN MN 90 90
FIG. 4, A
0 IO 20 30 40 50 60 MN 90
FIG. 4, B FIG. 4. Delayed NH&l ketosis in the presence of varying amounts of added fu-
maric acid. Conditions, per flask, 100 mg. of whole rat liver homogenate, 20 PM of pyruvate, and either 12 pi of malonate or 30 PM of NH&l, with fumarate aa indi- cated. Dash lines, same as in Fig. 2. A and B represent experiments on two dif- ferent rats.
by guest on February 14, 2020http://w
ww
.jbc.org/D
ownloaded from
272 KETOGENIC EFFECT OF AMMONIUM CHLORIDE
reacted stoichiometrically. Under conditions favoring fixation of COz, the deficit in the final production of acetoacetate was 2.2 PM of pyruvate, indicating that one-half of this amount, or 1.1 PM of pyruvate, had been
TABLE I Production of Glutamic Acid by Rat Liver Homogenates
Pymvate added
PM
20
30 30 20 20 20 20
T
-
-
NH&I added
PM
30 30
30 30 30 30 30
- 1
.-
Fumarate added
10 10 6 6 6 6
Glutamic acid recovered
Source of glutamic decarboxyhe
PM
None ‘I “
5.7 4.3 4.3 5.5 6.2
E. coli 4157 “ I‘ 4157 “ ‘I 4157 “ “ 4157 “ “ 4157 “ “ 4157
C. weld& SR12 ‘I “ SR12
Conditions, 100 mg. of whole rat liver homogenate per flask in all cases except for the experiment reported in the last line, in which 150 mg. of liver were used.
THEORETICAL MAXIMUM ._________._._. -.-.-~-_--._-.- ._._._._._. -_.--.-._.- .-.- 0
IO 20 30 40 50 60 MIN 90
FIG. 5. Recovery of added fumaric acid (see the text)
fixed as oxalacetate. Assuming the dry weight as 30 per cent of the wet weight, the rate of fixation of CO2 in terms of the production of oxalacetate was Qoxa~scetate = 1.03. Stern (29) found no increase in the sum of suc- cinate and a-ketoglutarate on addition of pyruvate to rat liver mince
by guest on February 14, 2020http://w
ww
.jbc.org/D
ownloaded from
R. 0. RECKNAGEL AND V. R. POTTER 273
treated with malonate. He emphasized that the failure of his method to detect fixation of CO2 indicated only that the rate of the Wood-Werkman reaction in the mammalian liver was below Qoxalacetste = 1.0. Our ex- periments, in which conversion of pyruvate to acetoacetate was used as a more sensitive measure of the availability of oxalacetate, indicate that the rate of this reaction in rat liver is near the limit of detection in the experi- ments of Stern.
E#ect of Cytochrome c and DPN-Addition of cytochrome c to a final concentration of 2.67 X 10m5 M had a slight stimulatory effect on the rate of 02 consumption. It had no effect on the rate of pyruvic acid removal or acetoacetate production, either with added malonate or with added fumarate. However, as in the case of kidney (30), cytochrome c increased the maintenance of activity by 30 per cent, as indicated by the time to
TABLE II Effect of CO, on NH&l Ketosis
Additions Gas
30 JAM NH&I.. . Air 30 “ “ 45 /kM KHCOa. 5% CO2 30 “ “ 4 ” fumarate. . Air 4 “ fumarate....................... I‘
Pynwate to acetoacetate
PM
16.6 14.4 12.6 3.8
Deficit
PM
2.2 4.0
12.8
Conditions, 80 mg. of whole rat liver and 20 PM of pyruvate per flask. Incu- bation time, 60 minutes. No alkali absorbers in flasks containing added bicarbon- ate-CO,.
half the maximum rate of 02 consumption. Addition of cytochrome c 2.67 X 10e5 M, DPN, 1200 y, and nicotinamide, 30 pM, had little effect on the rate of conversion of pyruvate to acetoacetate in the malonate- treated system, although the rate of removal of pyruvate from the system was increased about 20 per cent. Analysis for lactic acid in the DPN- fortified system revealed that all the pyruvate disappearing from the sys- tem which could not be accounted for as acetoacetate was present as lactic acid. In the absence of added DPN, less than 0.5 PM of lactate was found.
Octanoate Oxidation-With 112 mg. of whole rat liver homogenate and 3 PM of added octanoic acid, and in the presence of 12 I.LM of malonate, we observed a vigorous 02 uptake, and recovered acetoacetate in yields of 91 and 93 per cent of the theoretical 2 moles of acetoacetate per mole of octanoate utilized. With 48 and 80 mg. of tissue, the yields were 80 per cent. In the absence of malonate, and on addition of fumarate, we were able to depress the yield of acetoacetate down to, but not below, 1 mole of acetoacetate per mole of octanoate. Lehninger and Kennedy (31),
by guest on February 14, 2020http://w
ww
.jbc.org/D
ownloaded from

274 KETOGENIC EFFECT OF AMMONIUM CHLORIDE
using washed residues from rat liver, obtained a much greater depression of octanoate conversion to acetoacetate in the malate-supplemented sys- tem (acetoacetate yield, only 13 per cent of the theoretical maximum). However, our experiments with octanoate oxidation confirm in general those of Lehninger (12) and of Lehninger and Kennedy (31), which indi- cate that availability of oxalacetate is the controlling mechanism in the disposition of ketone body precursors arising from oxidation of octanoic acid.
DISCUSSION
These data have certain implications regarding the control of metabolic pathways in the whole organism. In two separate instances, a substrate having two alternative metabolic pathways open to it was shown to pro- ceed preferentially by one pathway when a second substrate was available and to proceed via another pathway when the second substrate was not available. In these experiments it was possible to arrange the conditions so that other potential reactions were minimized and the maximum shifts were obtained. It is recognized that the controlling mechanisms in the utilization of pyruvate in the liver of the intact animal will involve many controlling factors not operating in the reaction systems employed here. However, by limiting the reactions to two main alternatives, it became feasible to demonstrate the decisive r81e played by the delivery of the second substrate. The extent, to which these mechanisms operate in the intact animal can only be shown by experiments designed to this end. Unpublished studies on rats injected with fluoroacetate have shown that the accumulation of citrate in liver in vivo can be greatly modified by al- tering the conditions of the experiment along lines suggested by the data reported above.
SUMMARY
The ketogenic effect of added ammonium chloride in the rat liver slice has been confirmed and the phenomenon has been extended to the rat liver homogenate, supplemented with adenosinetriphosphate, magnesium, potassium, and phosphate. Data are presented which indicate that the ketosis induced by ammonium chloride involves the following steps: (1) a primary vigorous reductive amination of cr-ketoglutaric acid to form glutamic acid; (2) because of the interruption of the Krebs cycle at the stage of a-ketoglutarate oxidation to succinate, the system fails to supply levels of oxalacetate sufficient for the Krebs citric acid condensation; (3) with the preferential pathway to citric acid formation no longer avail- able, the metabolism of ketone body precursors is shunted in the direction of acetoacetate formation.
by guest on February 14, 2020http://w
ww
.jbc.org/D
ownloaded from

R. 0. RECKNAGEL AND V. R. PO’ITER 27.5
BIBLIOGRAPHY
1. Annau, E., 2. physiol. Chem., 224, 141 (1934). 2. Edson, N. L., Biochem. J., 29, 2082 (1935). 3. Krebs, H. A., Ann. Rev. B&hem., 6, 247 (1936). 4. Bloch, K., Physiol. Rev., 27. 574 (1947). 5. Dewan, J. G., Biochem. J., 32, 1378 (1938). 6. Adler, E., von Euler, H., Gunther, E., and Plass, M., Biochem. J., 33,1028 (1939). 7. Krebs, H. A., and Cohen, P. P., Biochem. J., 33, 1895 (1939). 8. Braunstein, A. E., and Kritzmann, M. G., Biokhimiya, 2, 859 (1937); Chem.
Abstr., 32, 2548 (1938). 9. Kritamann, M. G., J. Biol. Chem., 167, 77 (1947).
10. Cammarata, P. S., and Cohen, P. P., J. Biol. Chem., 187, 439 (1950). 11. Krebs, H. A., Eggleston, L. V., and Hems, R., Biochem. J., 43, 406 (1948). 12. Lehninger, A. L., .I. Biol. Chem., 164, 291 (1946). 13. Stadie, W. C., and Riggs, B. C., J. BioZ. Chem., 164, 687 (1944). 14. Robinson, J. R., Biochem. J., 46, 68 (1949). 15. Potter, V. R., and Elvehjem, C. A., J. BioZ. Chem., 114, 495 (1936). 16. Potter, V. R., Pardee, A. B., and Lyle, G. G., J. BioZ. Chem., 176, 1075 (1948). 17. Keilin, D., and Hartree, E. R., Proc. Roy. Sot. London, Series B, 122, 298 (1937). 18. Umbreit, W. W., BuTriB, R. H., and Stauffer, J. F., Manometrio techniques and
tissue metabolism, Minneapolis, 2nd edition (1949). 19. Friedemann, T. E., and Haugen, G. E., J. BioZ. Chem., 147, 415 (1943). 20. LePage, G. A., Cancer Res., 10, 393 (1950). 21. Greenberg, L. A., and Lester, D., J. BioZ. Chem., 164, 177 (1944). 22. Barker, S. B., and Summerson, W. H., J. BioZ. Chem., 138, 535 (1941). 23. Ljunggren, G., Ber. them. Ges., 66, 2469 (1923). 24. Gale, E. F., Biochem. J., 39, 46 (1946). 25. Umbreit, W. W., and Gunsalus, I. C., J. BioZ. Chem., 169, 333 (1945). 26. Natelson, S., Luguvoy, J. K., and Pincus, J. B., J. BioZ. Chem., 170, 597 (1947). 27. Buchanan, J. M., Sakami, W., and Gurin, S., J. BioZ. Chem., 169, 411 (1947). 28. Wood, H. G., and Werkman, C. H., Biochem. J., 32, 1262 (1938). 29. Stern, J. R., Biochem. J., 43, 616 (1948). 30. Pardee, A. B., and Potter, V. R., J. BioZ. Chem., 181,739 (1949). 31. Lehninger, A. L., and Kennedy, E. P., J. BioZ. Chem., 173, 753 (1948).
by guest on February 14, 2020http://w
ww
.jbc.org/D
ownloaded from

Richard O. Recknagel and Van R. PotterEFFECT OF AMMONIUM CHLORIDE
MECHANISM OF THE KETOGENIC
1951, 191:263-276.J. Biol. Chem.
http://www.jbc.org/content/191/1/263.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/191/1/263.citation.full.haccessed free atThis article cites 0 references, 0 of which can be
by guest on February 14, 2020http://w
ww
.jbc.org/D
ownloaded from