life history patterns in tropical south american …colli2010.pdf · 50 life history patterns in...
TRANSCRIPT

CAPÍTULO II
LIFE HISTORY PATTERNS IN TROPICAL SOUTH AMERICAN LIZARDS
Daniel O. Mesquita y Guarino R. Colli


ABSTRACT
We use comparative methods to investigate the relative contributions of environmental conditions and histori-cal effects upon life history aspects of tropical South American lizards. We assembled a dataset based on our sampling in 25 localities in tropical South America and additional data from 31 localities based on the litera-ture. To investigate the roles of phylogenetic history and environmental parameters in life history variables, we used Canonical Phylogenetic Ordination – CPO. Most populations from Cerrado, Amazonian Savannas, and Restinga are single-brooded, while most populations from Amazon Forest are multiple-brooded, corroborat-ing the hypothesis that species from seasonal regions
tend to reproduce ciclically, whereas species from less seasonal regions reproduce continuously. We found no significant differences in clutch size among regions. The CPO detected a significant historical effect in life his-tory aspects mainly in the node separating Iguania and Scleroglossa and for Cnemidophorus lizards, similar to other results reported elsewhere. Probably, the advanced age of the separation between Iguania and Scleroglossa (late Triassic), when probably most of the environmen-tal influence occurred, are responsible for the historical significant effect encountered. Much of the life history variation exhibits today simply reflect phylogenetic conservatism, thus having an historical basis.


Life history studies are essential for understanding the diversity and complexity of the vital cycles of living organisms (Roff, 1992; Stearns, 1992). Lizards are excellent subjects for ecological studies, because they are often abundant and easy to observe and capture. As a consequence, lizard studies have contributed enormously to the development of several areas of research, including foraging and life-history theory, population and community ecology, and the growing field of comparative biology (Huey et al., 1983; Vitt and Pianka, 1994). The number of studies on life-history variation has increased steadily since the classic comparative studies of Tinkle (Tinkle, 1969; Tinkle et al., 1970), and has developed enormously in the last decades (see Ballinger, 1983; Dunham et al., 1988; Shine and Schwarzkopf, 1992; Shine and Charnov, 1992; Stearns, 1984; Vitt, 1992). However, the majority of these studies was conducted either on species from temperate areas or on tropical anoles. The relevance and application of such theoretical developments to a great number of poorly known tropical species remains to be determined. For
instance, the gathering and analysis of massive amounts of data indicates that both phylogenetic inertia and adaptive responses to environmental conditions seem to influence lizard life-history patterns (Dunham and Miles, 1985; Dunham et al., 1988; Vitt, 1992). Therefore, several lineages restricted to tropical regions and under-represented in life-history studies, such as Gymnophthalmidae, Hoplocercidae, and Leiosauridae, might possess unique attributes that can substantially affect both the generality and the predictions of life-history models. Conversely, tropical regions contain unique ecogeographic features and conditions that can influence lizard ecologies in ways that disagree with current life-history theory. Nowadays, we know that there is a great variation on life-history patterns of South American tropical lizards (e.g., Colli et al., 1997; Colli et al., 2003; Mesquita and Colli, 2003b; Van Sluys, 1993; Van Sluys, 2000; Vitt, 1992; Wiederhecker et al., 2002).
Life-history variation among populations or species can have genetic and non genetic causes (Ballinger, 1983; Dunham et al., 1988). Several environmental
INTRODUCTION

LIFE HISTORY PATTERNS IN TROPICAL SOUTH AMERICAN LIZARDS50
factors can influence the life history of organisms, such as temperature, precipitation and photoperiod (Censky, 1995; Wiederhecker et al., 2002), the availability of favorable sites for egg development (Andrews, 1988), environmental predictability (Colli, 1991; Mesquita and Colli, 2003b; Vitt and Colli, 1994) and food availability (Vrcibradic and Rocha, 1998b). In addition, foraging mode can affect life history traits. Vitt and Congdon (1978) proposed that foraging mode, body shape, and relative clutch mass have coevolved in lizards. Actively foraging lizards rely on speed to avoid predation, have typically streamlined bodies, and clutches that comprise a relatively low proportion of total body mass. Conversely, sit-and-wait lizards rely on crypsis against predators, have a stocky body shape, and high relative clutch mass (Vitt and Congdon, 1978). This dichotomy was corroborated by several studies (e.g., Anderson and Karasov, 1981; Huey and Pianka, 1981; Vitt and Price, 1982; Dunham and Miles, 1985). Further, habitat specialization can also constrain life history parameters. For instance, many species of anoles have fixed clutch size of a single egg, which is compensated by multiple clutches spread throughout
the reproductive season, presumably related to the microhabitat used by these species (arboreal), which can limit the production of larger clutches because of the weight excess (Dunham et al., 1988; Roff, 1992; Stearns, 1992). Likewise, the utilization of rock crevices as shelter to avoid predators has strong influences in the morphology of Tropidurus semitaeniatus, resulting in a reduced clutch size (Vitt, 1981).
Although environmental conditions, like precipitation, temperature, and environmental predictability no doubt influence life history traits (e.g., Vitt and Colli, 1994; Censky, 1995; Wiederhecker et al., 2002; Mesquita and Colli, 2003b), it is becoming increasingly clear that life history traits could also have their origins deep in evolutionary history (e.g., Dunham and Miles, 1985; Dunham et al., 1988; Vitt, 1992). The aim of this work is to compare life history attributes of tropical South American lizards, identifying the relative contributions of environmental conditions and evolutionary history, using comparative methods combining life-history data with current phylogenetic hypotheses.

REPRODUCCIÓN EN REPTILES: MORFOLOGÍA, ECOLOGÍA Y EVOLUCIÓN 51
Study localities. 1- Cuyabeno, Colombia, 2- Santander, Colombia, 3- Boa Vista, RR,
4- Porto Walter, AC, 5- Guajará-Mirim, RO, 6- Santa Cruz da Serra, RO, 7- Ariquemes,
RO, 8- Santa Barbara, RO, 9- Humaitá, AM, 10- Amapá, AP, 11- Monte Alegre, PA, 12-
Alter do Chão, PA, 13- Curuá-Una, PA, 14- Altamira, PA, 15- Carajás, PA, 16- São Luís,
MA, 17- Serra do Cachimbo, PA, 18- Chapada dos Guimarães, MT, 19- Jaru, PA, 20- Ilha
do Bananal, TO, 21- Alto Araguaia, MT, 22- Barra do Garças, MT, 23- Pirenópolis,
GO, 24- Minaçu, GO, 25- Jalapão, TO, 26- São Domingos, GO, 27- Dianópolis, TO,
28- Alto Paraíso, GO, 29- Alvorada do Norte, GO, 30- Paranã, TO, 31- Correntina, BA,
32- Coribe, BA, 33- Brasília, DF, 34- Cristalina, GO, 35- Paracatu, MG, 36- Mirorós,
MATERIAL AND METHODS
Study sites
Used data collected by the authors during ca. two decades, from 25 localities distributed in tropical South America (Fig. 1). In addition, we searched the literature for the same kind of data, adding 31 more localities (Fig. 1). All lizards collected by authors were deposited in Coleção Herpetológica da Universidade de Brasília (CHUNB).
Reproduction
We sexed lizards by dissection and direct examination of gonads. Females were considered reproductive if vitellogenic follicles or oviductal eggs were present. We regarded the simultaneous presence of enlarged vitellogenic follicles and either oviductal eggs or corpora lutea as evidence for the sequential production of more than one clutch of eggs during the year. We considered clutch size as the number of vitellogenic follicles or oviductal eggs in mature females. For each lizard, we recorded the snout-vent length (SVL) with Mitutoyo® electronic calipers to the nearest 0.01 mm.
The data
We recorded for each population the following variables: mean SVL, mean clutch size (number of offspring per
Figure 1

LIFE HISTORY PATTERNS IN TROPICAL SOUTH AMERICAN LIZARDS52
clutch for all reproductive females in the population), clutch frequency (single or multiple-brooded) and preferred habitat type. We collected life history data of tropical South American lizards from 151 populations. Data from 68 populations (45%) were collected by the
authors and data the remaining 83 populations (55%) were obtained from the literature. All data collected are described in Appendix 1. From this data, we extracted common patterns and mean clutch size and SVL for species that were represented by more than one population (Table 1). We obtained climatic data, like mean annual temperature, total annual precipitation, and annual variation in precipitation for each study locality (available in Instituto Nacional de Meteorologia, 1998). To estimate annual variation in precipitation we used the coefficient of variation of total monthly precipitation.
Individual groups used in canonical phylogenetic ordination for life history data. Phylogeny based in Estes et al. (1988), Reeder et al. (2002), Frost et al. (2001) and
Giugliano (2003).
Figure 2
BA, 37- Irecê, BA, 38- Petrolina, PE, 39- Exu, PE, 40- Prado, BA, 41- Buzios, RJ,
42- Barra de Maricá, RJ, 43- Ilha Grande, RJ, 44- Ubatuba, SP, 45- Caraguatatuba, SP,
46- São Sebastião, SP, 47- Bertioga, SP, 48- Alcatrazes, SP, 49- Enseada, SP, 50- Queimada
Grande, SP, 51- Peruíbe, SP, 52- Valinhos, SP, 53- Campinas, SP, 54- Itirapina, SP,
55- Serra do Cipó, MG, 56- Vitória, ES, 57- Vilhena, RO and 58- Itatiaia, RJ. Closed
symbols: data collected by authors; open symbols: data from literature.

REPRODUCCIÓN EN REPTILES: MORFOLOGÍA, ECOLOGÍA Y EVOLUCIÓN 53
Statistical analyses
To assess the role of evolutionary history and environmental parameters (mean annual temperature, total annual precipitation, annual variation in precipitation, preferred habitat type and biome) in life history traits, we used Canonical Phylogenetic Ordination-CPO (Giannini, 2003). CPO is a modification of Canonical Correspondence Analysis-CCA (Ter Braak, 1986), a constrained ordination method that promotes the ordination of a set of variables in such a way that its association with a second set of variables is maximized. The significance of the association is tested via randomizations of one or both of the data sets. In our CPO, one of the matrices (Y) contained life history data (clutch size and clutch condition) measured over
the lizard populations, whereas the second matrix (X) consisted of a tree matrix that contained all monophyletic groups (Fig. 2). Each coded separately as a binary variable, and environmental parameters (mean annual temperature, total annual precipitation, annual variation in precipitation, preferred habitat type and biome). The analysis thus consisted of finding the subset of X that best explained the variation in Y, using CCA coupled with Monte Carlo permutations. Because SVL does influence life-history parameters, like clutch size (e.g., Colli, 1991; Vitt and Zani, 1996a; Vitt and Zani, 1996b; Colli et al., 2003; Mesquita and Colli, 2003a), we used mean SVL as a covariate. We performed CPO in CANOCO 4.5 for Windows, using the following parameters: symmetric scaling, biplot scaling, manual selection of environmental variables (monophyletic groups and environmental
Lizard Species
Gekkonidae
Coleodactylus meridionalis
Gonatodes humeralis
Gymnodactylus geckoides
Phyllopezus pollicaris
Thecadactylus rapicauda
Gymnophthalmidae
Colobosaura modesta
Micrablepharus maximiliani
SVL
24.78 (2)
36.40 (1)
40.71 (8)
76.10 (2)
109.10 (2)
45.35 (3)
39.04 (5)
Clutch size
1.00 (2)
1.00 (2)
1.72 (8)
2.00 (2)
1.00 (2)
2.00 (3)
2.20 (5)
Clutch condition
Fixed
Fixed
Not fixed
Fixed
Fixed
Fixed
Not fixed
Table 1

LIFE HISTORY PATTERNS IN TROPICAL SOUTH AMERICAN LIZARDS54
Polychrotidae
Anolis meridionalis
Polychrus acutirostris
Scincidae
Mabuya caissara
Mabuya frenata
Mabuya guaporicola
Mabuya heathi
Mabuya macrorhyncha
Mabuya nigropunctata
Teiidae
Ameiva ameiva
Cnemidophorus cryptus
Cnemidophorus lemniscatus
Cnemidophorus mumbuca
Cnemidophorus cf ocellifer
Kentropyx striata
Polychrotidae
Anolis meridionalis
Polychrus acutirostris
Tropiduridae
Tropidurus hispidus
Tropidurus itambere
Tropidurus cf oreadicus
54.92 (2)
109.34 (3)
71.58 (1)
65.96 (4)
65.27 (3)
63.30 (2)
66.80 (1)
84.15 (5)
114.77 (14)
59.19 (3)
58.51 (4)
50.07 (2)
59.07 (19)
93.45 (2)
54.92 (2)
109.34 (3)
81.19 (3)
66.65 (4)
71.72 (11)
1.54 (2)
15.77 (3)
4.85 (4)
4.05 (4)
4.08 (3)
4.00 (2)
2.78 (7)
4.26 (5)
1.46 (14)
1.49 (3)
1.67 (4)
1.00 (2)
2.11 (19)
5.30 (2)
1.54 (2)
15.77 (3)
5.82 (3)
3.51 (4)
4.32 (11)
Not fixed
Not fixed
Not fixed
Not fixed
Not fixed
Not fixed
Not fixed
Not fixed
Not fixed
Not fixed
Not fixed
Fixed
Not fixed
Not fixed
Not fixed
Not fixed
Not fixed
Not fixed
Not fixed
Summary of life history data of Tropical South American lizard species. Note: Clutch sizes (in parenthesis) are relative to populations.
Continuation...

REPRODUCCIÓN EN REPTILES: MORFOLOGÍA, ECOLOGÍA Y EVOLUCIÓN 55
Biome
All biomes
Amazon Forest
Amazonian Savannas
Caatinga
Cerrado
Restinga
Single-brooded
52.38% (55)
10.53% (2)
61.54% (8)
53.33% (8)
64.58% (31)
62.50% (5)
Multiple-brooded
47.62% (50)
89.47% (17)
38.46% (5)
46.67% (7)
35.42% (17)
37.50% (3)
Comparison
x2 = 0.238; df = 1; P = 0.626
x2 = 11.842; df = 1; P = 0.001
x2 = 0.692; df = 1; P = 0.405
x2 = 0.067; df = 1; P = 0.796
x2 = 4.083; df = 1; P = 0.043
x2 = 0.500; df = 1; P = 0.480
Percentages of populations of South American Tropical lizards per biome according with clutch frequency. Sample sizes are in parenthesis.
Table 2
parameters), 9,999 permutations, and unrestricted permutations. We carried out other statistical analyses using SYSTAT 11.0 for Windows, with a significance level of 5% to reject null hypotheses. Throughout the text, means appear ± 1 SD.
RESULTS
In 31% of studied populations clutch size was fixed (Table 1). Apparently, similar numbers of populations of tropical South American lizards are single brooded or multiple-brooded (Table 2). Considering populations per biome, most populations from Cerrado, Amazonian Savannas, and Restinga were single-brooded, while most populations from Amazon Forest were multiple-brooded (Table 2; Appendix 1). We found no significant difference in clutch size among biomes, independently of SVL (ANCOVA F4,1,60 = 1.450, P = 0.538; Adjusted Means Amazon
Forest = 2.74 ± 2.03, Caatinga = 2.99 ± 1.77, Cerrado = 3.35 ± 1.33, Restinga = 3.41 ± 1.10, and Amazonian Savannas = 3.71 ± 1.87).
The CPO revealed a significant phylogenetic effect on life history aspects at the node separating Iguania and Scleroglossa, which accounted for 22% of total life history variation (Table 3). In addition, historical significant effects were detected in the clade containing the Cnemidophorus species from Amazonian Savannas (E’), the clade with Cnemidophorus from central and eastern Brazil (I’), in Autarchoglossa (M’), Tropidurinae and Liolaeminae (E), in the node separating Teioidea and Anguidae (L’), in Tropidurinae (F), and in Tropidurinae without Uracentron (G), accounting respectively 18, 14, 14, 12, 12, 12, and 10% of the life history variation (Table 3). No significant phylogenetic effects were detected in any other clade or in any environmental parameter (Table 3), indicating that life history parameters are shaped primarily by historical factors.

LIFE HISTORY PATTERNS IN TROPICAL SOUTH AMERICAN LIZARDS56
J
C
K
J’
L
M
A
Preferred habitat type
X
C’
O
S
Mean annual temperature
T
0.004
0.003
0.003
0.002
0.002
0.002
0.001
0.001
0.001
0.001
0.001
0.001
0.001
0.001
8.00
6.00
6.00
4.00
4.00
4.00
2.00
2.00
2.00
2.00
2.00
2.00
2.00
2.00
3.056
2.395
2.390
1.864
1.809
1.452
1.120
1.118
1.040
0.972
0.945
0.942
0.881
0.836
0.0856
0.1163
0.1301
0.1803
0.1862
0.2275
0.3590
0.2954
0.4023
0.3232
0.3541
0.3389
0.3574
0.3675
Group(s)
D/R
E’
I’
M’
E
L’
F
G
H
Z
I
B
K’
Variation
0.011
0.009
0.007
0.007
0.006
0.006
0.006
0.005
0.005
0.004
0.004
0.004
0.004
Variation %
22.00
18.00
14.00
14.00
12.00
12.00
12.00
10.00
10.00
8.00
8.00
8.00
8.00
F
9.142
6.811
5.804
5.545
4.774
4.659
4.385
4.150
3.803
3.396
3.311
3.209
3.098
P
0.0023
0.0056
0.0165
0.0204
0.0287
0.0341
0.0394
0.0444
0.0520
0.0667
0.0724
0.0616
0.0770
Table 3

REPRODUCCIÓN EN REPTILES: MORFOLOGÍA, ECOLOGÍA Y EVOLUCIÓN 57
Continuation....
Historical effects on the life history parameters South American Tropical lizards. Results of Monte Carlo permutation tests of individual groups (defined in Figs. 2) and environmental
variables, for the Y matrix of life history data, with mean SVL as a covariate. Percentage of the variation explained (relative to total unconstrained variation), and F and P values for
each variable are given (9999 permutations were used) for each main matrix.
V
W
Biome
D’
P
N
B’
G’
H’
Annual variation in precipitation
Y
Q
U
A’
F’
Total annual precipitation
0.001
0.001
0.001
0.001
0.001
0.001
0.001
0.000
0.000
0.000
0.000
0.000
0.000
0.000
0.000
0.000
2.00
2.00
2.00
2.00
2.00
2.00
2.00
0.00
0.00
0.00
0.00
0.00
0.00
0.00
0.00
0.00
0.818
0.771
0.749
0.670
0.638
0.546
0.446
0.295
0.242
0.205
0.167
0.139
0.089
0.058
0.054
0.016
0.4179
0.4571
0.3849
0.4205
0.4295
0.4726
0.5227
0.5899
0.6279
0.6489
0.6782
0.7128
0.7756
0.8695
0.8277
0.9001
DISCUSSION
Reproductive parameters are often related to environmental factors that limit reproduction (Tinkle et al., 1970; Dunham et al., 1988). In temperate regions, the main limiting factor is the rigorous winter, being the reproductive timing and the clutch size affected (McCoy and Hoddenbach, 1966; Pianka, 1970). In tropical regions, the reproduction is also affected by climatic variables. Some species
reproduce continuously with usually several clutches per reproductive season, where the precipitation is better distributed throughout the year (e.g., Amazon Forest) or is unpredictable (e.g., Caatinga), and reproduce cyclically with only one or a few clutches per reproductive season, in regions with seasonal climate (e.g., Cerrado and Amazonian Savannas) (see Vitt, 1982a; Vitt and Blackburn, 1983; Vitt, 1983; Vitt and Goldberg, 1983; Colli, 1991; Vitt and Colli, 1994; Wiederhecker et al., 2002; Colli et

LIFE HISTORY PATTERNS IN TROPICAL SOUTH AMERICAN LIZARDS58
al., 2003; Mesquita and Colli, 2003b). Apparently, in temperate regions reproduction is affected by a cold-warm seasonality; whereas in tropical regions by a wet-dry seasonality.
The seasonality in reproduction influences directly the clutch sizes. The production of bigger clutches, distributed in only one or a few hatches, in populations that live in regions with seasonal climate appear to be an adaptation to concentrate the reproductive effort in a short period, differently of what occur in unpredictable environments and in places where the precipitation is well distributed throughout the year, where generally the species reproduce continuously with smaller clutches distributed in several hatches (Colli, 1991; Vitt and Colli, 1994; Colli et al., 2003; Mesquita and Colli, 2003b). This was reported for several species. For example, Ameiva ameiva reproduce continuously with smaller clutches in the Caatinga and Amazon Forest, and seasonally with bigger clutches in Cerrado (Vitt, 1982a; Colli, 1991; Vitt and Colli, 1994), and Cnemidophorus ocellifer and Gymnodactylus geckoides reproduce seasonally with bigger clutches in Cerrado and continuously with smaller clutches in the Caatinga (Vitt and Goldberg, 1983; Vitt, 1983; Colli et al., 2003; Mesquita and Colli, 2003b). Our results partially support this tendency. We showed that most lizard populations from Cerrado, Restinga and Amazonian Savannas (seasonal biomes) are single-brooded and most populations from Amazon Forest
are multiple-brooded, but our results for Caatinga contradict this hypothesis. However, data from a very well studied Caatinga site in Northeast Brazil, Pernambuco State, clearly reveal this tendency. From 13 species from this area, nine shows prolonged reproductive period and are multiple-brooded (Vitt, 1992). Regarding the clutch size, even which our results showed the tendency of clutch sizes of lizards from Cerrado, Restinga and Amazonian Savannas being bigger than from Amazon Forest and Caatinga, the comparisons were not significant. However, studies comparing reproduction aspects from the same species among biomes, like Ameiva ameiva, Cnemidophorus ocellifer and Gymnodactylus geckoides from Caatinga and Cerrado, confirm this trend (Colli, 1991; Vitt and Colli, 1994; Colli et al., 2003; Mesquita and Colli, 2003b).
Several Anoline lizards are characterized by a clutch size of a single egg as a consequence of an extremely low relation between clutch and body weight, which is partly compensated by multiple broods (Andrews and Rand, 1974). A characteristic of the genus Anolis is the possession of expanded subdigital lamellae or adhesive toe pads. These are adaptively associated with arboreal habitats in most species (Andrews and Rand, 1974; Roff, 1992). The number of subdigital lamellas is positively correlated with the degree of arboreality in Anolis species; the loading capacity of a female anole may limit the amount of additional weight she can

REPRODUCCIÓN EN REPTILES: MORFOLOGÍA, ECOLOGÍA Y EVOLUCIÓN 59
carry and still climb effectively (Collette, 1961). In contrast, arboreal lizard species without toe pads (e.g., Polychrus, Iguana and Chamaleo) have a large clutch size (see Vitt and Lacher, 1981; Campos, 2004). Thus, the low clutch number in some anoles is explained by their climbing habitats (Andrews and Rand, 1974; Roff, 1992). Also, the use of crevices to avoid predators could also play an important role in the evolution of body and egg morphology and clutch size in the lizard Tropidurus semitaeniatus (Vitt, 1981). Unlike most Tropidurus species, which lay hatches of 3-8 eggs (This work, Vitt, 1991c; Van Sluys, 1993; Vitt, 1993; Van Sluys et al., 2002; Wiederhecker et al., 2002), T. semitaeniatus lay only two elongated eggs as an adaptation for its habit (Vitt, 1993). In addition, an unrelated genus with crevice-dwelling habits, the African Cordylidae Platysaurus showed similar modifications to those in T. semitaeniatus (Roff, 1992). However, our results do not corroborate this hypothesis. The CPO analysis indicated none significant effect of preferred habitat type on life history traits, accounting only 2% of the total variation. We have no doubt about the constraining of life history traits by mechanical factors imposed by habitat type; nevertheless, this constrains appear to be uncommon, in whole South America, only T. semitaeniatus appear to have this trait. Also, the anoles species used in the analysis (Anolis meridionalis and A. nitens), are not arboreal and probably do not suffer this constrains.
The CPO detected a significant historical effect in life history aspects mainly in the node separating Iguania – Scleroglossa and for the Cnemidophorus lizards. Similar results, based on dietary shifts, were reported elsewhere. In a study with diet data of 184 lizard species of four families from four continents reveal significant historic effects on dietary shifts, being the most striking divergence in the node separating Iguania – Scleroglossa (Vitt et al., 2003; Vitt and Pianka, 2005). The authors suggested the hypothesis that ancient events in squamate cladogenesis, rather than present–day interactions, caused dietary shifts in these major clades, which promoted that some lizards gained access to new resources, influencing much of the biodiversity observed today (Vitt et al., 2003; Vitt and Pianka, 2005). Even that the environmental variables influenced and still influence in life history traits of lizards, the separation between Iguania and Scleroglossa, when probably most of this influence occurred, was long ago in the evolutionary history of these clades (late Triassic). In addition, much of the life history variation exhibit today simply reflect phylogenetic conservatism, thus having an historical basis (see Harvey and Pagel, 1991; Brooks and McLennan, 1993). Regarding the historical effects encountered in Cnemidophorus lizards, the genus occurs from Argentina to north of Central America (Zug et al., 2001; Reeder et al., 2002). In spite of their wide distribution, they exhibit similar

LIFE HISTORY PATTERNS IN TROPICAL SOUTH AMERICAN LIZARDS60
ecological traits along them (see Pianka, 1970; Vitt et al., 1997b; Mesquita and Colli, 2003b). Studies with South American Cnemidophorus, show that their ecological traits are very conservative, varying a little, even among drastically different biomes, which is an indication that these parameters have an historical basis (Vitt et al., 1997b; Mesquita and Colli, 2003b), corroborating with the results encountered in the present work.
Although there are some analyses to examine the historical influences on ecological aspects, the use of phylogenetically based analyses is in its infancy, and the nature of forces acting on species remains unclear. Previous studies have suggested that ecological aspects have both historical and non historical basis (Ballinger, 1983; Dunham et al., 1988; Vitt and Colli, 1994; Mesquita and Colli, 2003b). In addition, observations on ecological characteristics of species and the use of great amount of data, especially involving a significant portion of clades (global ecology) are essential to elucidate the origins of life history variations.
LITERATURE CITED
Anderson, R. A. and W. H. Karasov. (1981). “Contrasts in energy intake and expenditure in sit-and-wait and widely foraging lizards”. Oecologia. 49: 67-72.
Andrews, R. and A. S. Rand. (1974). “Reproductive effort in anoline lizards”. Ecology. 55: 1317-1327.
Andrews, R. M. (1988). “Demographic correlates of variable egg survival for a tropical lizard”. Oecologia (Berlin). 76: 376-382.
Anjos, I. A., M. C. Kieferiefer and R. J. Sawaya. (2002). “Kentropyx paulensis (NCN). Reproduction”. Herp. Rev. 33: 52.
Araújo, A. F. B. (1991). “Structure of a white sand-dune lizard community of coastal Brazil”. Rev. Bras. Biol. 51: 857-865.
Ballinger, R. E. (1983). “Life-history variations”. In: Huey R. B., E. R. Pianka, and T. W. Schoener (eds.). Lizard Eco-logy: Studies of a Model Organism. Harvard University Press, Cambridge, Massachusetts. Pages 241-260.
Brooks, D. R. and D. A. McLennan. (1993). “Historical ecol-ogy: examining phylogenetic components of commu-nity evolution”. In: Ricklefs R. E. and D. Schluter (eds.). Species Diversity in Ecological Communities, Historical and Geographical Perspectives. The University of Chi-cago Press, Chicago, Illinois. Pages 267-280.
Campos, Z. (2004). “Iguana iguana (sinimbu, green iguana). Reproduction”. Herp. Rev. 35: 169-169.
Censky, E. J. (1995). “Reproduction in two Lesser Antil-lean populations of Ameiva plei (Teiidae)”. J. Herpetol. 29: 553-560.
Collette, B. B. (1961). “Correlations between ecology and morphology in anoline lizards from Havana, Cuba and southern Florida”. Bull. Mus. Comp. Zool. 125: 137-162.
Colli, G. R. (1989). “O ciclo reprodutivo e o dimorfismo sexual em Ameiva ameiva (Sauria, Teiidae) nos cerrados do Distrito Federal”. In: Departamento de Ecologia. Univer-

REPRODUCCIÓN EN REPTILES: MORFOLOGÍA, ECOLOGÍA Y EVOLUCIÓN 61
sidade de Brasília, Brasília, Distrito Federal. Page 75.Colli, G. R. (1991). “Reproductive ecology of Ameiva amei-
va (Sauria: Teiidae) in the cerrado of central Brazil”. Copeia. 1991: 1002-1012.
Colli, G. R., D. O. Mesquita, P. V. V. Rodrigues and K. Kitayama. (2003). “The ecology of the gecko Gymno-dactylus geckoides amarali in a neotropical savanna”. J. Herpetol. 37: 694-706.
Colli, G. R., A. K. Péres, Jr. and M. G. Zatz. (1997). “For-aging mode and reproductive seasonality in tropical lizards”. J. Herpetol. 31: 490-499.
Dunham, A. E. and D. B. Miles. (1985). “Patterns of covariation in life history traits of squamate reptiles: the effects of size and phylogeny reconsidered”. Am. Nat. 126: 231-257.
Dunham, A. E., D. B. Miles and D. N. Reznick. (1988). “Life history patterns in squamate reptiles”. In: Gans C. and R. B. Huey (eds.). Biology of the Reptilia. Vol. 16, Ecology B. Defense and Life History. Alan R. Liss, Inc., New York. Pages 441-522.
Estes, R., K. de Queiroz and J. Gauthier. (1988). “Phylogenetic relationships within Squamata”. In: Estes R. and G. Pregill (eds.). Phylogenetic Relationships of the Lizard Families. Essays Commemorating Charles L. Camp. Stanford Uni-Stanford Uni-versity Press, Stanford, California. Pages 119-281.
Faria, R. G. (2001). “Ecologia de duas espécies simpátricas de Tropiduridae (Tropidurus itambere e Tropidurus oreadicus) no Cerrado do Brasil Central”. In: Departamento de Eco-logia. Universidade de Brasília, Brasília, Distrito Federal.
Frost, D. R., M. T. Rodrigues, T. Grant and T. A. Titus.
(2001). “Phylogenetics of the lizard genus Tropidurus (Squamata: Tropiduridae: Tropidurinae): direct optimization, descriptive, efficiency, and sensitivity analysis of congruence between molecular data and morphology”. Mol. Phylogenet. Evol. 21: 352-371.
Galdino, C. A. B., V. B. Assis, M. C. Kiefer and M. Van Sluys. (2003). “Reproduction and fat body cycle of Eurolophosaurus nanuzae (Sauria; Tropiduridae) from a seasonal montane habitat of southeastern Brazil”. J. Herpetol. 37: 687-694.
Giannini, N. P. (2003). “Canonical phylogenetic ordina-tion”. Syst. Biol. 52: 684-695.
Giugliano, L. G. (2003). “Variação aloenzimática e sistemá-tica de uma nova espécie de Cnemidophorus (Reptilia, Teiidae) de Rondônia”. In: Faculdade de Medicina de Ribeirão Preto. Universidade de São Paulo, Ribeirão Preto, São Paulo. Page 57.
Harvey, P. H. and M. D. Pagel. (1991). “The Compara-tive Method in Evolutionary Biology”. Oxford Univ. Press, New York.
Huey, R. B. and E. R. Pianka (1981). “Ecological conse-quences of foraging mode”. Ecology. 62: 991-999.
Huey, R. B., E. R. Pianka and A. Schoener. (1983). “Liz-ard Ecology: Studies of a Model Organism”. Harvard University Press, Cambridge, Mass.
Instituto Nacional de Meteorologia. (1998). “Normais Climatológicas”. Ministério da Agricultura, Instituto Nacional de Meteorologia, Rio de Janeiro.
Luedemann, G., G. R. Colli and R. A. Brandão. (1997). “Polychrus acutirostris (Bicho-preguiça)”. Reproduction.

LIFE HISTORY PATTERNS IN TROPICAL SOUTH AMERICAN LIZARDS62
Herp. Rev. 28: 43.McCoy, C. J. and G. A. Hoddenbach. (1966). “Geographic
variation ovarian cycles and clutch size in Cnemido-phorus tigris (Teiidae)”. Science. 1671-1672.
Menezes, V. A., C. F. D. Rocha and G. F. Dutra. (2004). “Reproductive ecology of the partenogenetic whiptail lizard Cnemidophorus nativo in a Brazilian restinga habitat”. J. Herpetol. 38: 280-282.
Mesquita, D. O. (2001). “Uma análise comparativa da ecolo-gia de populações do lagarto Cnemidophorus (Squamata: Teiidae) do Brasil”. In: Departamento de Ecologia. Uni-versidade de Brasília, Brasília, Distrito Federal. Page 73.
Mesquita, D. O. and G. R. Colli. (2003a). “The ecology of Cnemidophorus ocellifer (Squamata, Teiidae) in a neotropical savanna”. J. Herpetol. 37: 498-509.
Mesquita, D. O. and G. R. Colli. (2003b). “Geographical variation in the ecology of populations of some Brazil-ian species of Cnemidophorus (Squamata, Teiidae)”. Copeia. 2003: 285-298.
Mesquita, D. O., A. K. P. Jr, G. H. C. Vieira and G. R. Colli. (2000). “Natural History: Mabuya guaporicola”. Herp. Rev. 31: 240-241.
Miranda, J. P. and G. V. Andrade. (2003). “Seasonality in diet, perch use, and reproduction of the gecko Gonatodes humeralis from eastern Brazilian Amazon”. J. Herpetol. 37: 433-438.
Mojica, B. H., B. H. Rey, V. H. Serrano and M. P. Ramírez-Pinilla. (2003). “Annual reproductive activity of a population of Cnemidophorus lemniscatus (Squamata: Teiidae)”. J. Herpetol. 37: 35-42.
Pianka, E. R. (1970). “Comparative autecology of the lizard Cnemidophorus tigris in different parts of its geographic range”. Ecology. 51: 703-720.
Pinto, A. C. S. (1999a). “Dimorfismo sexual e comporta-mento social do lagarto Tropidurus torquatus (Squama-ta, Tropiduridae) em uma área de Cerrado no Distrito Federal”. In: Departamento de Ecologia. Universidade de Brasília, Brasília, Distrito Federal.
Pinto, M. G. M. (1999b). “Ecologia das espécies de lagartos simpátricos Mabuya nigropunctata e Mabuya frenata (Scincidae), no Cerrado de Brasília e Serra da Mesa (GO)”. In: Departamento de Ecologia. Universidade de Brasília, Brasília, Distrito Federal. Page 101.
Reeder, T. W., C. J. Cole and H. C. Dessauer. (2002). “Phy-logenetic relationships of Whiptail lizards of the genus Cnemidophorus (Squamata: Teiidae): a test of mono-phyly, reevaluation of karyotypic evolution, and review of hybrid origins”. Am. Mus. Novit. 3365: 1-61.
Rocha, C. F. D. (1992). “Reproductive and fat body cycles of the tropical sand lizard (Liolaemus lutzae) of South-eastern Brazil”. J. Herpetol. 26: 17-23.
Rocha, C. F. D. and D. Vrcibradic. (1996). “Thermal ecol-ogy of two sympatric skinks (Mabuya macrorhyncha and Mabuya agilis) in a brazilian restinga habitat”. Aust. J. Ecol. 21: 110-113.
Rocha, C. F. D. and D. Vrcibradic. (1999). “Reproductive traits of two sympatric viviparous skinks (Mabuya macrorhyncha and Mabuya agilis) in a brazilian restinga habitat”. Herp. J. 9: 43-53.
Roff, D. A. (1992). “The Evolution of Life Histories:

REPRODUCCIÓN EN REPTILES: MORFOLOGÍA, ECOLOGÍA Y EVOLUCIÓN 63
Theory and Analysis”. Chapman and Hall, London.Shine, R. and E. L. Charnov. (1992). “Patterns of survival,
growth, and maturation in snakes and lizards”. Am. Nat. 139: 1257-1269.
Shine, R. and L. Schwarzkopf. (1992). “The evolution of reproductive effort in lizards and snakes”. Evolution. 46: 62-75.
Stearns, S. C. (1984). “The effects of size and phylogeny on patterns of covariation in the life history traits of lizards and snakes”. Am. Nat. 123: 56-72.
Stearns, S. C. (1992). “The Evolution of Life Histories”. Oxford University Press, Oxford.
Ter Braak, C. J. F. (1986). “Canonical correspondence analysis: a new eigenvector technique for multivariate direct gradient analysis”. Ecology. 76: 1167-1179.
Tinkle, D. W. (1969). “The concept of reproductive effort and its relation to the evolution of life histories of lizards”. Am. Nat. 103: 501-516.
Tinkle, D. W., H. M. Wilbur and S. G. Tilley. (1970). “Evolutionary strategies in lizard reproduction”. Evo-lution. 24: 55-74.
Van Sluys, M. (1993). “The reproductive cycle of Tropi-durus itambere (Sauria: Tropiduridae) in Southeastern Brazil”. J. Herpetol. 27: 28-32.
Van Sluys, M. (2000). “Population dynamics of the saxicolous lizard Tropidurus itambere (Tropiduridae) in a seasonal habitat of Southeastern Brazil”. Herpetologica. 56: 55-62.
Van Sluys, M., V. M. Ferreira and C. F. D. Rocha. (2004). “Natural history of the lizard Enyalius brasiliensis (Les-son, 1828) (Leiosauridae) from an Atlantic Forest of
southeastern Brazil”. Braz. J. Biol. 64: 353-356.Van Sluys, M., H. M. A. Mendes, V. B. Assis and M. C.
Kiefer. (2002). “Reproduction of Tropidurus montanus Rodrigues, 1987 (Tropiduridae), a lizard from a seaso-nal habitat of south-eastern Brazil, and a comparison with other Tropidurus species”. Herp. J. 12: 89-97.
Vanzolini, P. E. and R. Rebouças-Spieker. (1976). “Distri-bution and differentiation of animals along the coast and on continental islands of the state of São Paulo, Brazil. 3. Reproductive differences between and within Mabuya caissara and M. macrorhyncha (Sauria: Scinci-dae)”. Pap. Avulsos Zool. 29: 95-109.
Vieira, G. H. C., D. O. Mesquita, A. K. P. Jr, K. Kitayama and G. R. Colli. (2000). “Natural History: Micra-blepharus atticolus”. Herp. Rev. 31: 241-242.
Vitt, L. J. (1981). “Lizard reproduction: habitat specific-ity and constraints on relative clutch mass”. Am. Nat. 117: 506-514.
Vitt, L. J. (1982a). “Reproductive tactics of Ameiva ameiva (Lacertilia: Teiidae) in a seasonally fluctuating tropical habitat”. Can. J. Zool. 60: 3113-3120.
Vitt, L. J. (1982b). “Sexual dimorphism and reproduction in the microteiid lizard, Gymnophthalmus multiscuta-tus”. J. Herpetol. 16: 325-329.
Vitt, L. J. (1983). “Reproduction and sexual dimorphism in the tropical teiid lizard Cnemidophorus ocellifer”. Copeia. 1983: 359-366.
Vitt, L. J. (1985). “On the biology of the little known lizard, Diploglossus lessonae in Northeast Brazil”. Pap. Avulsos Zool. 36: 69-76.

LIFE HISTORY PATTERNS IN TROPICAL SOUTH AMERICAN LIZARDS64
Vitt, L. J. (1986). “Reproductive tactics of sympatric gek-konid lizards with a comment on the evolutionary and ecological consequences of invariant clutch size”. Copeia. 1986: 773-786.
Vitt, L. J. (1991a). “Ecology and life history of the scanso-rial arboreal lizard Plica plica (Iguanidae) in Amazo-nian Brazil”. Can. J. Zool. 69: 504-511.
Vitt, L. J. (1991b). “Ecology and life history of the wide-foraging lizard Kentropyx calcarata (Teiidae) in Ama-zonian Brazil”. Can. J. Zool. 69: 2791-2799.
Vitt, L. J. (1991c). “An introduction to the ecology of Cerrado lizards”. J. Herpetol. 25: 79-90.
Vitt, L. J. (1992). “Diversity of reproductive strategies among brazilian lizards and snakes: the significance of lineage and adaptation”. In: Hamlett W. C. (ed.). Reproductive Biology of South American Vertebrates. Springer-Verlag, New York. Pages 135-149.
Vitt, L. J. (1993). “Ecology of isolated open-formation Tropidurus (Reptilia: Tropiduridae) in Amazonian lowland rain forest”. Can. J. Zool. 71: 2370-2390.
Vitt, L. J. (1995). “The ecology of tropical lizards in the Caatinga of northeast Brazil”. Occas. Pap. Oklahoma Mus. Nat. Hist. 1: 1-29.
Vitt, L. J. and T. C. S. Ávila-Pires. (1998). “Ecology of two sympatric species of Neusticurus (Sauria: Gym-nophthalmidae) in the Western Amazon of Brazil”. Copeia. 1998: 570-582.
Vitt, L. J., T. C. S. Ávila-Pires and P. A. Zani. (1996). “Observations on the ecology of the rare amazonian
lizard, Enyalius leechii (Polychrotidae)”. Herpetol. Nat. Hist. 4: 77-82.
Vitt, L. J. and D. G. Blackburn. (1983). “Reproduction in the lizard Mabuya heathi (Scincidae): a commentary on viviparity in new world Mabuya”. Can. J. Zool. 61: 2798-2806.
Vitt, L. J. and D. G. Blackburn. (1991). “Ecology and life history of the viviparous lizard Mabuya bistriata (Scincidae) in the Brazilian Amazon”. Copeia. 1991: 916-927.
Vitt, L. J. and J. P. Caldwell. (1993). “Ecological obser-vations on Cerrado lizards in Rondônia, Brazil”. J. Herpetol. 27: 46-52.
Vitt, L. J. and C. M. Carvalho. (1992). “Life in the trees: the ecology and life-history of Kentropyx striatus (Teiidae) in the Lavrado area of Roraima, Brazil, with comments on tropical teiid life histories”. Can. J. Zool. 70: 1995-2006.
Vitt, L. J. and G. R. Colli. (1994). “Geographical ecology of a neotropical lizard: Ameiva ameiva (Teiidae) in Brazil”. Can. J. Zool. 72: 1986-2008.
Vitt, L. J. and J. D. Congdon. (1978). “Body shape, re-productive effort, and relative clutch mass in lizards: resolution of a paradox”. Am. Nat. 112: 595-608.
Vitt, L. J. and S. R. Goldberg. (1983). “Reproductive ecology of two tropical iguanid lizards: Tropidurus torquatus and Platynotus semitaeniatus”. Copeia. 1983: 131-141.
Vitt, L. J. and T. E. Lacher, Jr. (1981). “Behaviour, habitat, diet and reproduction of the iguanid lizard Polychrus

REPRODUCCIÓN EN REPTILES: MORFOLOGÍA, ECOLOGÍA Y EVOLUCIÓN 65
acutirostris in the Caatinga of Northeastern Brazil”. Herpetologica. 37: 53-63.
Vitt, L. J. and E. R. Pianka. (1994). “Lizard Ecology”. Princeton University Press, Princeton.
Vitt, L. J. and E. R. Pianka. (2005). “Deep history impacts present-day ecology and biodiversity”. Proc. Natl. Acad. Sci. 102: 7877-7881.
Vitt, L. J., E. R. Pianka, W. E. Cooper, Jr. and K. Schwenk. (2003). “History and the global ecology of squamate reptiles”. Am. Nat. 162: 44-60.
Vitt, L. J., and H. J. Price. (1982). “Ecological and evolu-tionary determinants of relative clutch mass in lizards”. Herpetologica. 38: 237-255.
Vitt, L. J., R. A. Souza, S. S. Sartorius, T. C. d. Avilla-Pires and M. C. Espósito. (2000). “Comparative ecology of sympatric Gonatodes (Squamata: Gekkonidae) in western Amazon of Brazil”. Copeia. 2000: 83-95.
Vitt, L. J. and P. A. Zani. (1996a). “Ecology of the elu-sive tropical lizard Tropidurus [=Uracentron] flaviceps (Tropiduridae) in lowland rain forest of Ecuador”. Herpetologica. 52: 121-132.
Vitt, L. J. and P. A. Zani. (1996b). “Ecology of the lizard Ameiva festiva (Teiidae) in Southeastern Nicaragua”. J. Herpetol. 30: 110-117.
Vitt, L. J. and P. A. Zani. (1997). “Ecology of the nocturnal lizard Thecadactylus rapicauda (Sauria: Gekkonidae) in the Amazon region”. Herpetologica. 53: 165-179.
Vitt, L. J., P. A. Zani and T. C. S. Ávila-Pires. (1997a).
“Ecology of the arboreal tropidurid lizard Tropidurus (= Plica) umbra in the Amazon region”. Can. J. Zool. 75: 1876-1882.
Vitt, L. J., P. A. Zani, J. P. Caldwell, M. C. D. Araujo and W. E. Magnusson. (1997b). “Ecology of whiptail lizards (Cnemidophorus) in the Amazon region of Brazil”. Copeia. 1997: 745-757.
Vitt, L. J., P. A. Zani, J. P. Caldwell and E. O. Carrillo. (1995). “Ecology of the lizard Kentropyx pelviceps (Sauria: Teiidae) in lowland rain forest of Ecuador”. Can. J. Zool. 73: 691-703.
Vrcibradic, D., M. Cunha-Barros and C. F. D. Rocha. (2004). “Ecological observations on Mabuya dorsivitta-ta (Squamata; Scincidae) from a high latitude habitat in South-Eastern Brazil”. Herp. J. 14: 109-112.
Vrcibradic, D. and C. F. D. Rocha. (1998a). “The ecology of the skink Mabuya frenata in an area of rock outcrops in Southeastern Brazil”. J. Herpetol. 32: 229-237.
Vrcibradic, D. and C. F. D. Rocha. (1998b). “Reproductive cycle and life-history traits of the viviparous skink Mabuya frenata in Southeastern Brazil”. Copeia. 1998: 612-619.
Wiederhecker, H. C., A. C. S. Pinto and G. R. Colli. (2002). “Reproductive ecology of Tropidurus torquatus (Squamata: Tropiduridae) in the highly seasonal Cerra-do biome of central Brazil”. J. Herpetol. 36: 82-91.
Zug, G. R., L. J. Vitt and J. P. Caldwell. (2001). “Herpe-tology: an Introductory Biology of Amphibians and Reptiles”. Academic Press, San Diego, California.

LIFE HISTORY PATTERNS IN TROPICAL SOUTH AMERICAN LIZARDS66
LS
Scleroglossa
Anguidae
Diploglossus lessonae
Gekkonidae
Coleodactylus meridionalis
Coleodactylus meridionalis
Gonatodes humeralis
Gonatodes humeralis
Gonatodes hasemani
Gymnodactylus geckoides
Gymnodactylus geckoides
Gymnodactylus geckoides
Gymnodactylus geckoides
Gymnodactylus geckoides
Gymnodactylus geckoides
Gymnodactylus geckoides
Gymnodactylus geckoides
Lygodactylus klugei
Hemidactylus mabouia
Phyllopezus pollicaris
Phyllopezus pollicaris
Thecadactylus rapicauda
Thecadactylus rapicauda
Gymnophthalmidae
Bachia bresslaui
Colobosaura modesta
Colobosaura modesta
B
CA
CE
CE
AF
AF
AF
CE
CE
CE
CE
CA
CE
CE
CA
CA
CA
CA
CE
AF
AF
CE
CE
CE
POP
39
30
46
5
16
5
30
46
25
38
23
24
39
39
39
39
46
13
1
35
35
46
SVL
141.4 (28)
23.16 (7)
26.40 (25)
36.40 (7)
40.10 (11)
41.98 (85)
39.67 (75)
39.55 (370)
47.17 (107)
37.49 (5)
38.29 (61)
39.74 (287)
41.8 (191)
28.85 (355)
57.90 (29)
75.25 (70)
76.95 (19)
105.90 (29)
112.30 (24)
90.67 (12)
47.43 (14)
51.33 (6)
CS
3.33 (9)
1.00 (1)
1.00 (4)
1.00 (7)
1.00 (14)
1.00 (11)
1.67 (6)
1.75 (4)
1.65 (32)
2.00 (1)
1.00 (2)
2.00 (1)
1.71 (28)
2.00 (47)
2.00 (94)
2.00 (2)
2.00 (12)
2.00 (3)
1.00 (7)
1.00 (7)
2.00 (1)
2.00 (5)
2.00 (1)
CF
1
-
1
+ 1
+ 1
+ 1
1
1
1
-
-
-
1
+ 1
+ 1
1
+ 1
-
+ 1
+ 1
-
1
-
PHT
SF
LL
LL
TR
TR
LG
TN
UL
UR
TN
UR
UR
UR
UR
TR
B
RO
RO
B
TR
LL
LL
LL
REF
(Vitt, 1985)
This work
This work
(Vitt et al., 2000)
(Miranda and Andrade, 2003)
(Vitt et al., 2000)
This work
This work
(Colli et al., 2003)
This work
This work
This work
This work
(Vitt, 1986; Vitt, 1995)
(Vitt, 1986; Vitt, 1995)
(Vitt, 1986; Vitt, 1995)
(Vitt, 1986; Vitt, 1995)
This work
(Vitt and Zani, 1997)
(Vitt and Zani, 1997)
This work
This work
This work
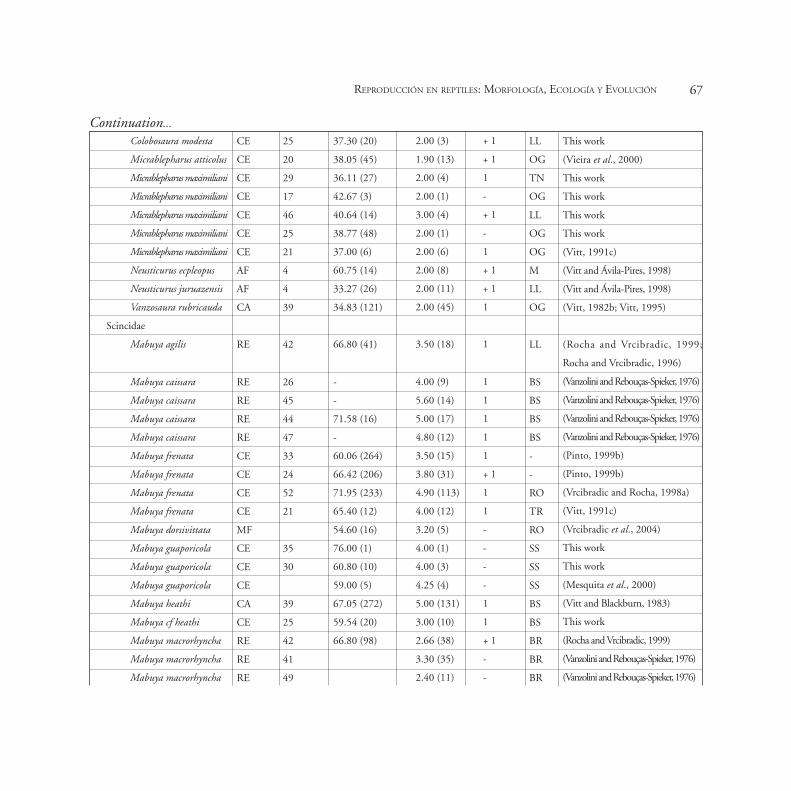
Appendix 1
REPRODUCCIÓN EN REPTILES: MORFOLOGÍA, ECOLOGÍA Y EVOLUCIÓN 67
Colobosaura modesta
Micrablepharus atticolus
Micrablepharus maximiliani
Micrablepharus maximiliani
Micrablepharus maximiliani
Micrablepharus maximiliani
Micrablepharus maximiliani
Neusticurus ecpleopus
Neusticurus juruazensis
Vanzosaura rubricauda
Scincidae
Mabuya agilis
Mabuya caissara
Mabuya caissara
Mabuya caissara
Mabuya caissara
Mabuya frenata
Mabuya frenata
Mabuya frenata
Mabuya frenata
Mabuya dorsivittata
Mabuya guaporicola
Mabuya guaporicola
Mabuya guaporicola
Mabuya heathi
Mabuya cf heathi
Mabuya macrorhyncha
Mabuya macrorhyncha
Mabuya macrorhyncha
CE
CE
CE
CE
CE
CE
CE
AF
AF
CA
RE
RE
RE
RE
RE
CE
CE
CE
CE
MF
CE
CE
CE
CA
CE
RE
RE
RE
25
20
29
17
46
25
21
4
4
39
42
26
45
44
47
33
24
52
21
35
30
39
25
42
41
49
37.30 (20)
38.05 (45)
36.11 (27)
42.67 (3)
40.64 (14)
38.77 (48)
37.00 (6)
60.75 (14)
33.27 (26)
34.83 (121)
66.80 (41)
-
-
71.58 (16)
-
60.06 (264)
66.42 (206)
71.95 (233)
65.40 (12)
54.60 (16)
76.00 (1)
60.80 (10)
59.00 (5)
67.05 (272)
59.54 (20)
66.80 (98)
2.00 (3)
1.90 (13)
2.00 (4)
2.00 (1)
3.00 (4)
2.00 (1)
2.00 (6)
2.00 (8)
2.00 (11)
2.00 (45)
3.50 (18)
4.00 (9)
5.60 (14)
5.00 (17)
4.80 (12)
3.50 (15)
3.80 (31)
4.90 (113)
4.00 (12)
3.20 (5)
4.00 (1)
4.00 (3)
4.25 (4)
5.00 (131)
3.00 (10)
2.66 (38)
3.30 (35)
2.40 (11)
+ 1
+ 1
1
-
+ 1
-
1
+ 1
+ 1
1
1
1
1
1
1
1
+ 1
1
1
-
-
-
-
1
1
+ 1
-
-
LL
OG
TN
OG
LL
OG
OG
M
LL
OG
LL
BS
BS
BS
BS
-
-
RO
TR
RO
SS
SS
SS
BS
BS
BR
BR
BR
This work
(Vieira et al., 2000)
This work
This work
This work
This work
(Vitt, 1991c)
(Vitt and Ávila-Pires, 1998)
(Vitt and Ávila-Pires, 1998)
(Vitt, 1982b; Vitt, 1995)
(Rocha and Vrcibradic, 1999;
Rocha and Vrcibradic, 1996)
(Vanzolini and Rebouças-Spieker, 1976)
(Vanzolini and Rebouças-Spieker, 1976)
(Vanzolini and Rebouças-Spieker, 1976)
(Vanzolini and Rebouças-Spieker, 1976)
(Pinto, 1999b)
(Pinto, 1999b)
(Vrcibradic and Rocha, 1998a)
(Vitt, 1991c)
(Vrcibradic et al., 2004)
This work
This work
(Mesquita et al., 2000)
(Vitt and Blackburn, 1983)
This work
(Rocha and Vrcibradic, 1999)
(Vanzolini and Rebouças-Spieker, 1976)
(Vanzolini and Rebouças-Spieker, 1976)
Continuation...

LIFE HISTORY PATTERNS IN TROPICAL SOUTH AMERICAN LIZARDS68
Mabuya macrorhyncha
Mabuya macrorhyncha
Mabuya macrorhyncha
Mabuya macrorhyncha
Mabuya nigropunctata
Mabuya nigropunctata
Mabuya nigropunctata
Mabuya nigropunctata
Mabuya nigropunctata
Mabuya sp.
Teiidae
Ameiva ameiva
Ameiva ameiva
Ameiva ameiva
Ameiva ameiva
Ameiva ameiva
Ameiva ameiva
Ameiva ameiva
Ameiva ameiva
Ameiva ameiva
Ameiva ameiva
Ameiva ameiva
Ameiva ameiva
Ameiva ameiva
Ameiva ameiva
Cnemidophorus cryptus
Cnemidophorus cryptus
Cnemidophorus cryptus
Cnemidophorus gramivagus
Cnemidophorus lemniscatus
RE
RE
RE
RE
AS
AS
CE
CE
AF
CE
CE
CA
AS
CE
CE
CE
AS
CE
AS
CE
AF
AF
AS
AS
AS
AS
AS
AS
AS
50
51
56
48
11
25
33
24
46
17
39
11
35
46
33
12
24
3
21
14
19
3
15
11
10
14
9
12
82.30 (10)
80.17 (6)
80.40 (338)
88.71 (307)
89.15 (143)
55.79 (35)
85.11 (115)
135.10 (316)
84.86 (80)
126.71 (61)
128.00 (6)
133.69 (358)
78.07 (43)
113.42 (31)
118.26 (116)
122.80 (11)
125.30 (21)
114.00 (137)
126.50 (33)
115.00 (34)
58.89 (85)
53.77 (81)
64.90 (21)
55.84 (93)
51.13 (54)
2.20 (51)
3.20 (12)
3.40 (5)
2.33 (3)
3.00 (2)
4.00 (4)
5.30 (9)
4.30 (97)
4.70 (94)
3.54 (13)
2.60 (5)
5.65 (106)
3.33 (3)
5.93 (14)
5.50 (2)
6.40 (83)
8.00 (1)
3.67 (3)
4.20 (5)
4.30 (10)
4.40 (12)
3.20 (110)
3.90 (28)
4.80 (5)
1.09 (23)
1.48 (21)
1.90 (15)
1.68 (3)
1.13 (8)
-
-
-
-
-
-
1
1
-
1
1
+ 1
-
+ 1
-
+ 1
-
-
1
+ 1
+ 1
+ 1
+ 1
1
1
+ 1
-
1
1
BR
BR
BR
BR
BS
BS
LG
LL
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
(Vanzolini and Rebouças-Spieker, 1976)
(Vanzolini and Rebouças-Spieker, 1976)
(Vanzolini and Rebouças-Spieker, 1976)
(Vanzolini and Rebouças-Spieker, 1976)
This work
This work
(Pinto, 1999b)
(Pinto, 1999b)
(Vitt and Blackburn, 1991)
This work
This work
(Vitt, 1982a)
This work
This work
This work
(Colli, 1989; Colli, 1991)
This work
This work
This work
(Vitt and Colli, 1994)
(Vitt and Colli, 1994)
(Vitt and Colli, 1994)
(Vitt and Colli, 1994)
This work
This work
(Mesquita, 2001; Mesquita and Colli, 2003b)
(Vitt et al., 1997b)
(Mesquita, 2001; Mesquita and Colli, 2003b)
This work
Continuation...

REPRODUCCIÓN EN REPTILES: MORFOLOGÍA, ECOLOGÍA Y EVOLUCIÓN 69
Cnemidophorus lemniscatus
Cnemidophorus lemniscatus
Cnemidophorus lemniscatus
Cnemidophorus nativo
Cnemidophorus mumbuca
Cnemidophorus mumbuca
Cnemidophorus cf ocellifer
Cnemidophorus cf ocellifer
Cnemidophorus cf ocellifer
Cnemidophorus cf ocellifer
Cnemidophorus cf ocellifer
Cnemidophorus cf ocellifer
Cnemidophorus cf ocellifer
Cnemidophorus cf ocellifer
Cnemidophorus cf ocellifer
Cnemidophorus cf ocellifer
Cnemidophorus cf ocellifer
Cnemidophorus cf ocellifer
Cnemidophorus cf ocellifer
Cnemidophorus cf ocellifer
Cnemidophorus cf ocellifer
Cnemidophorus cf ocellifer
Cnemidophorus cf ocellifer
Cnemidophorus cf ocellifer
Cnemidophorus cf ocellifer
Cnemidophorus parecis
Dracaena guianensis
Kentropyx calcarata
AS
AS
DF
RE
CE
CE
CE
CE
CE
CE
CE
CE
CE
CE
CA
CA
CE
CE
CA
CA
CA
CE
CE
CA
CA
CE
AF
AF
3
2
40
27
25
21
29
35
30
24
28
22
33
32
31
34
18
37
36
38
23
39
57
10
62.44 (125)
57.45 (74)
63.03 (250)
56.10 (48)
50.63 (27)
49.51 (193)
57.40 (12)
50.46 (77)
57.46 (22)
52.33 (162)
53.69 (142)
54.62 (29)
53.29 (59)
58.33 (28)
59.75 (20)
56.50 (14)
56.22 (37)
59.49 (36)
69.64 (45)
59.91 (22)
65.14 (63)
60.33 (81)
59.35 (322)
72.52 (464)
65.85 (99)
65.78 (99)
303.38 (6)
80.28 (122)
1.88 (8)
1.50 (16)
2.17 (46)
2.20 (37)
1.00 (3)
1.00 (43)
2.30 (12)
1.50 (2)
3.00 (2)
1.80 (5)
2.25 (16)
2.00 (2)
1.75 (4)
2.60 (5)
2.00 (5)
2.25 (4)
3.25 (4)
1.83 (6)
1.50 (2)
1.60 (5)
1.75 (4)
2.20 (15)
2.07 (41)
2.67 (41)
1.83 (23)
1.58 (12)
6.00 (1)
6.00 (11)
1
1
+ 1
+ 1
-
1
1
-
-
1
1
-
-
1
1
-
-
1
-
1
-
1
1
+ 1
1
1
+ 1
-
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
OG
TE
BS
This work
(Mesquita, 2001; Mesquita and Colli, 2003b)
(Mojica et al., 2003)
(Menezes et al., 2004)
This work
This work
(Vitt, 1991c)
This work
This work
This work
This work
This work
This work
This work
This work
This work
This work
This work
This work
This work
This work
This work
(Mesquita and Colli, 2003a; Mesquita
and Colli, 2003b)
(Vitt, 1983)
(Mesquita, 2001; Mesquita and Colli, 2003b)
(Mesquita, 2001; Mesquita and Colli, 2003b)
This work
(Vitt, 1991b)
(Anjos et al., 2002)
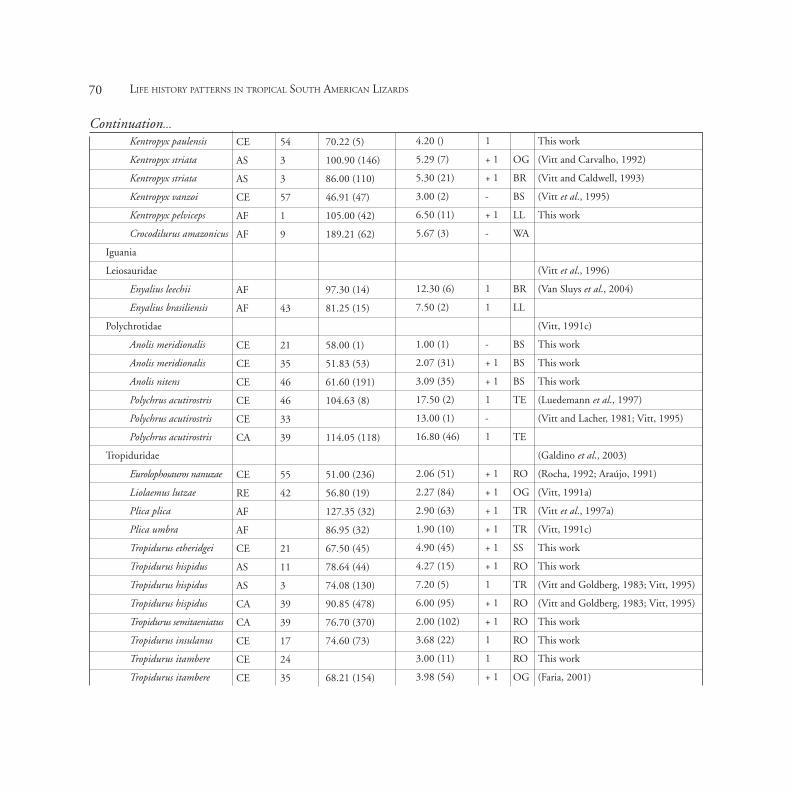
Continuation...
LIFE HISTORY PATTERNS IN TROPICAL SOUTH AMERICAN LIZARDS70
Kentropyx paulensis
Kentropyx striata
Kentropyx striata
Kentropyx vanzoi
Kentropyx pelviceps
Crocodilurus amazonicus
Iguania
Leiosauridae
Enyalius leechii
Enyalius brasiliensis
Polychrotidae
Anolis meridionalis
Anolis meridionalis
Anolis nitens
Polychrus acutirostris
Polychrus acutirostris
Polychrus acutirostris
Tropiduridae
Eurolophosauros nanuzae
Liolaemus lutzae
Plica plica
Plica umbra
Tropidurus etheridgei
Tropidurus hispidus
Tropidurus hispidus
Tropidurus hispidus
Tropidurus semitaeniatus
Tropidurus insulanus
Tropidurus itambere
Tropidurus itambere
CE
AS
AS
CE
AF
AF
AF
AF
CE
CE
CE
CE
CE
CA
CE
RE
AF
AF
CE
AS
AS
CA
CA
CE
CE
CE
54
3
3
57
1
9
43
21
35
46
46
33
39
55
42
21
11
3
39
39
17
24
35
70.22 (5)
100.90 (146)
86.00 (110)
46.91 (47)
105.00 (42)
189.21 (62)
97.30 (14)
81.25 (15)
58.00 (1)
51.83 (53)
61.60 (191)
104.63 (8)
114.05 (118)
51.00 (236)
56.80 (19)
127.35 (32)
86.95 (32)
67.50 (45)
78.64 (44)
74.08 (130)
90.85 (478)
76.70 (370)
74.60 (73)
68.21 (154)
4.20 ()
5.29 (7)
5.30 (21)
3.00 (2)
6.50 (11)
5.67 (3)
12.30 (6)
7.50 (2)
1.00 (1)
2.07 (31)
3.09 (35)
17.50 (2)
13.00 (1)
16.80 (46)
2.06 (51)
2.27 (84)
2.90 (63)
1.90 (10)
4.90 (45)
4.27 (15)
7.20 (5)
6.00 (95)
2.00 (102)
3.68 (22)
3.00 (11)
3.98 (54)
1
+ 1
+ 1
-
+ 1
-
1
1
-
+ 1
+ 1
1
-
1
+ 1
+ 1
+ 1
+ 1
+ 1
+ 1
1
+ 1
+ 1
1
1
+ 1
OG
BR
BS
LL
WA
BR
LL
BS
BS
BS
TE
TE
RO
OG
TR
TR
SS
RO
TR
RO
RO
RO
RO
OG
This work
(Vitt and Carvalho, 1992)
(Vitt and Caldwell, 1993)
(Vitt et al., 1995)
This work
(Vitt et al., 1996)
(Van Sluys et al., 2004)
(Vitt, 1991c)
This work
This work
This work
(Luedemann et al., 1997)
(Vitt and Lacher, 1981; Vitt, 1995)
(Galdino et al., 2003)
(Rocha, 1992; Araújo, 1991)
(Vitt, 1991a)
(Vitt et al., 1997a)
(Vitt, 1991c)
This work
This work
(Vitt and Goldberg, 1983; Vitt, 1995)
(Vitt and Goldberg, 1983; Vitt, 1995)
This work
This work
This work
(Faria, 2001)
Continuation...

REPRODUCCIÓN EN REPTILES: MORFOLOGÍA, ECOLOGÍA Y EVOLUCIÓN 71
Tropidurus itambere
Tropidurus itambere
Tropidurus montanus
Tropidurus spinulosus
Tropidurus torquatus
Tropidurus cf oreadicus
Tropidurus cf oreadicus
Tropidurus cf oreadicus
Tropidurus cf oreadicus
Tropidurus cf oreadicus
Tropidurus cf oreadicus
Tropidurus cf oreadicus
Tropidurus cf oreadicus
Tropidurus cf oreadicus
Tropidurus cf oreadicus
Tropidurus cf oreadicus
Uracentron flaviceps
CE
CE
CE
CE
CE
CE
CE
CE
CE
AS
CE
CE
AF
AF
AF
AF
AF
23
17
55
21
33
29
27
30
46
15
25
23
8
7
6
14
1
60.13 (194)
71.60 (176)
73.95 (243)
86.20 (21)
94.72 (299)
57.17 (41)
69.31 (48)
55.18 (51)
79.73 (26)
65.47 (72)
42.62 (165)
71.94 (136)
80.00 (56)
89.65 (53)
85.25 (82)
92.60 (38)
98.69 (20)
3.57 (35)
3.50 (40)
3.48 (52)
4.00 (21)
6.10 (56)
6.00 (1)
4.12 (17)
4.50 (2)
5.00 (3)
5.33 (3)
4.00 (1)
3.65 (26)
3.40 (17)
3.50 (18)
3.80 (45)
4.20 (24)
2.00 (5)
+ 1
+ 1
+ 1
+ 1
+ 1
-
1
-
-
-
-
1
1
+ 1
+ 1
+ 1
+ 1
RO
RO
RO
TR
RO
OG
RO
RO
RO
RO/OG
RO
RO
RO
RO
RO
TR
(Van Sluys, 1993)
(Van Sluys et al., 2002)
(Vitt, 1991c)
(Wiederhecker et al., 2002; Pinto, 1999a)
This work
This work
This work
This work
This work
This work
(Faria, 2001)
(Vitt, 1993)
(Vitt, 1993)
(Vitt, 1993)
(Vitt, 1993)
(Vitt and Zani, 1996a)
Life history data per population of Tropical South American lizards. Note: LS- lizard species, B- biome, POP- population, SVL- mean Snout vent-length, CS- clutch size,
CF- clutch frequency, FM- foraging mode, PHT- preferred habitat type, REF- Reference, AC- active foragers, SW- sit and wait foragers, B- building, BR- branches, BR-
bromeliads, BS- bushes, LG- log, LL- leaf litter, OG- open ground, RO- rock outcrops, SF- semi-fossorial, SS- sandy soils, TE- trees, TN- termite nests, TR- trunks, UL-
under log, M- Mud, UR- under rock, WA- water, AF- Amazon Forest, CA- Caatinga, CE- Cerrado, DF- Dry Forest, RE- Restinga, MF- Montane Fields and AS- Amazonian
Savanna. Population codes are defined in Figure 1. Sample sizes (in parenthesis) refer to individuals in each population.
Continuation...