ldentification and characterization of mpgi, gene … plant cell, vol. 5, 1575-1590, november 1993 0...
TRANSCRIPT

The Plant Cell, Vol. 5, 1575-1590, November 1993 0 1993 American Society of Plant Physiologists
ldentification and Characterization of MPGI, a Gene lnvolved in Pathogenicity from the Rice Blast Fungus Magnaporthe grisea
Nicholas J. Talbot,'>' Daniel J. Ebbole,b and John E. liamera,* a Department of Biological Sciences, Hansen Life Science Research Building, Purdue University, West Lafayette, Indiana 47907-1392
Department of Microbiology and Plant Pathology, Texas A & M University, College Station, Texas 77843-2132
Differential cDNA cloning was used to identify genes expressed during infectious growth of the fungal pathogen Mag- naporthe grisea in its host, the rice plant. We characterized one of these genes, MPG1, in detail. Using a nove1 assay to determine the proportion of fungal biomass present in the plant, we determined that the MPGl transcript was 60-fold more abundant during growth in the plant than in culture. Mpgl mutants have a reduced ability to cause disease symp- toms that appears to result from an impaired ability to undergo appressorium formation. MPGl mRNA was highly abundant very early in plant infection concomitant with appressorium formation and was also abundant at the time of symptom development. The MPGl mRNA was also expressed during conidiation and in mycelial cultures starved for nitrogen or carbon. MPGl potentially encodes a small, secreted, cysteine-rich, moderately hydrophobic protein with the characteris- tics of a fungal hydrophobin. Consistent with the role of the MPGl gene product as a hydrophobin, Mpgl mutants show an basily wettable" phenotype. Our results suggest that hydrophobins may have a role in the elaboration of infective structures by fungi and may fulfill other functions in fungal phytopathogenesis.
INTRODUCTION
The ability of a plant pathogenic fungus to infect and cause disease on its host plant is an intricate process encompass- ing the entire life cycle of the pathogen. Pathogenicity is therefore a complex phenotype involving such disparate abili- ties of the fungus as its infection efficiency, the extent to which it colonizes its host, its derangement of plant metabolism, and its capacity for disseminating infective propagules to new host plants.
The complexity of the pathogenic lifestyle is exemplified by the ascomycete fungus Magnaporthe grisea. The fungus is a pathogen of a large number of grass species but is best known as the causal agent of rice blast disease, the most se- rious disease of cultivated rice (Ou, 1985). The infective cycle of the rice blast fungus starts when an asexual spore (a conidium) lands on the leaf surface. The events that follow de- pend on the prevailing phyllosphere environment and the ability of the spore to enter a complex morphogenetic sequence. Hydration of the conidium results in the release of an adhe- sive from a periplasmic space at the spore apex which binds it tightly to the hydrophobic leaf surface (Hamer et al., 1988). The spore germinates within 2 hr of binding to the leaf surface
' Current address: Department of Biological Sciences, University of Exeter, Exeter EX4 4QG, U.K.
To whom correspondence should be addressed.
and forms a short germ tube which differentiates at the tip to produce the infective cellular structure, the appressorium (Howard et al., 1991a). The formation of this cell involves the cessation of apical growth and extensive melanization of the cell wall (Howard and Ferrari, 1989).
lnfection proceeds by tight adhesion of the appressorium to the leaf surface, followed by the build-up of a high interna1 turgor pressure (Howard et al., 1991b) which allows a penetra- tion peg to break through the cuticle and into the underlying epidermal cell layer. This process may involve actin-associated events as actin has been immunolocalized to penetration pegs of M. grisea (Bourett and Howard, 1992). lnfectious hyphae ramify throughout the underlying epidermal cells differentiat- ing into bulbous, branched secondary hyphae which spread both intercellularly and intracellularly (Heath et al.; 199Oa, 1990b). At 96 hr after the initial infection, the first visible symp- toms become apparent. Small ellipsoid lesions appear on leaves resulting from rapid chlorosis and necrosis of host tis- sues (Peng and Shishiyama, 1988; Valent et al., 1991). In heavy infection the lesions ultimately coalesce, covering much of the leaf surface. The fungus then sends out aerial conidiophores which sporulate extensively to further spread disease.
The successful colonization of the rice plant by M. grisea thus requires a large variety of morphogenetic and metabolic processes. Genetic characterization of pathogenesis has

1576 The Plant Cell
proceeded by the identification of single gene mutations thatbring about a loss of pathogenicity primarily by altering thefunction or formation of appressoria (for review, see Valent andChumley, 1991). Conversely, molecular genetic approacheshave shown that some putative pathogenicity genes such ascutinase may be dispensable for full pathogenicity (Sweigardet al., 1992a, 1992b). Physiological studies of rice blast dis-ease have identified a number of phytotoxic compoundsproduced by M. grisea including tenuazonic acid (3-acetyl5-sec-butyl pyrrolidine-2,4-dione) and pyricularin (for review,see Ou, 1985), although their precise role in pathogenesisawaits further characterization (Lebrun et al., 1990). Thus, manydiverse physiological and morphogenetic processes may beclassifiable as being mediated by "pathogenicity" genes. Thegenetic evidence for the complexity of pathogenesis is appar-ent from the study of Valent et al. (1991) in which polygenicfactors governing lesion density and size were observed inprogeny segregating for major host or cultivar specificity (aviru-lence) genes.
Given the complexity of infectious growth, we reasoned thatone strategy to identify genes playing important roles inpathogenicity would be to study genes expressed preferen-tially during growth of M. grisea in the plant. This paperdescribes the results of a differential cDNA screening designedto identify such genes. One gene, designated WPG7, encodesan mRNA that is ~60-fold more abundant during infectiousgrowth than in conditions used to grow M. grisea in culture.MPG1 is a single copy gene, maps to a new locus on the endof linkage group H (Romao and Hamer, 1992), and is conservedin most if not all isolates of M. grisea. Null mutations at theMPG1 locus produce a reduced pathogenicity phenotype oncompatible rice cultivars. The reduced pathogenicity pheno-type is associated with the reduced frequency of appressoriumdevelopment by Mpg1 mutants. DNA sequencing showed thatthe putative MPG1 gene product is related to the fungalhydrophobins, a recently described group of secreted proteins(Stringer etal., 1991; Wesselsetal., 1991a; Lauteretal., 1992;St. Leger et al., 1992). Hydrophobins have been implicatedin various aspects of fungal morphogenesis, including theproduction of aerial hyphae and reproductive structures. Ourresults suggest these proteins may also play important rolesin pathogenesis.
RESULTS
Identification of Abundant mRNAs duringM. grisea Infections
Rice plants of the cultivar CO-39 were infected with the com-patible M. grisea isolate Guy-11. The infection was allowed toproceed for 72 hr. At this point few visible disease symptomswere apparent, but considerable host colonization had alreadyoccurred. Total RNA was extracted from the infected rice leaves,enriched for poly(A)+ RNA, and used to construct a directional
cDNA library (see Methods). A low-density differential screenwas then carried out. Replicate filters were hybridized withcDNA probes derived from uninfected rice leaf poly(A)+ RNAand M. grisea-infected rice leaf poly(A)+ RNA. We selected42 cDNAs that hybridized preferentially to the infection cDNAprobe.
These cDNAs were converted to plasmid clones and clas-sified as being of rice or M. grisea origin by DNA gel blotanalysis (data not shown). The cDNAs were then classifiedby their hybridization patterns to RNA gel blots, and represen-tative clones are shown in Figure 1. Blots were prepared with
1 2 3—————————— kb
A - -0.9
8 <f • -0.9
-1.5
-1.1
-0.7
-0.7
G
Figure 1. Classification of cDNA Clones by RNA Gel Blot Hybridiza-tion Analysis.Poly(A)+ RNA (5 ng per lane) was fractionated on agarose gels un-der denaturing conditions and blotted to Hybond N membranes. Lane1 contains poly(A)+ RNA from M. grisea grown in CM; lane 2 containspoly(A)+ RNA from uninfected rice leaves; lane 3 contains polyfA)*RNA from M. gr/sea-mfected rice leaves. Blots were hybridized withthe following DNA probes.(A) pNJT-15 (18-hr autoradiograph exposure).(B) pNJT-15 (48-hr autoradiograph exposure).(C) pNJT-23.(D) pNJT-70.(E) pNJT-49.(F) pNJT-14.(G) Ethidium bromide-stained RNA gel of the same RNA preparations.

A Pathogenicity Gene from Magnaporthe 1577
equal amounts of poly(A)+ RNA from M. grisea grown in ax-enic culture, uninfected rice leaves, and M. grisea-infectedrice leaves. NJT-15 (Figure 1A) represents a M. grisea mRNAspecies expressed abundantly during plant infection. A longerexposure of the autoradiogram is given in Figure 1B, showingthat low level expression of the NJT-15 gene can be detectedin axenic culture. NJT-23 (Figure 1C) represents an abundantM. grisea mRNA species produced constitutively in culture andduring growth in the plant. NJT-70 (Figure 1D) represents aless abundant M. grisea mRNA species expressed at a slightlyhigher level in the plant than in culture based on estimationof fungal mRNA levels in the plant (see below). NJT-49 (Fig-ure 1E) represents a rice mRNA species produced abundantlyduring M. grisea infection. NJT-14 (Figure 1F) represents a ricemRNA species found specifically during M. grisea infection.
Fungal Genomic ONA (ug) Infected Rice Plant GenomicDNA (ug)
o o.- cv
Fungal DNA Probe
Rice DNA Probe
BHours Post-Inoculation
96 72 48 H
Fungal DNA Probe
Estimation of mRNA Levels in the Plant
Quantitative measurements of fungal gene expression duringgrowth in the plant is difficult because it is dependent on therelative proportions of fungal and plant biomass. This propor-tion changes with time and is likely to vary from experimentto experiment. Correlating RNA gel blot signals of fungal genesto known single copy genes was unsuccessful because thelevel of expression of several of these genes (e.g., ILV1 andp-tubulin) was almost undetectable in mRNA preparation frominfected plants and too little is known about infectious fungalgrowth to assume that any gene will be constitutivelyexpressed.
Assuming that the proportion of fungal and plant biomasspresent at any given time during an infection is equivalent tothe proportion of plant and fungal DNA, DNA gel blot analysiscan be used to estimate the extent of fungal growth in the plant.We found that plant and fungal DNA could be quantitativelyextracted by a single protocol and that DNA extraction by thismethod was mass dependent (see Methods). Figure 2 showshybridization of the ILV1 probe, a single copy M. grisea geneencoding isoleucine valine synthase (Valent and Chumley,1991), to total genomic DNA extracted from rice plants 72 hrafter inoculation with M. grisea. This is the time at which theinfection mRNA was routinely extracted and from which thecDNA library was constructed. The hybridization signals fromthis analysis were compared to a dilution series of DNA sam-ples extracted from M. grisea nuclei (Figure 2). Laserdensitometry of the hybridization signals showed that 500 ngof M. grisea genomic DNA gave the same hybridization signalas 5 ug of blast-infected rice genomic DNA. Therefore, 10%of the infected rice DNA is composed of M. grisea genomicDNA. Because DNA extraction is mass dependent, 10% ofthe biomass of infected leaves can be accounted for asrepresenting M. grisea mycelium. When this value was usedto normalize the densitometry data from the RNA gel blot hy-bridization experiments, the mRNA detected with the NJT-15clone (Figures 1A and 1B) was 62 times more abundant in the
Rice DNA Probe
Figure 2. Estimation of Fungal Biomass Present in M. gy/sea-lnfectedRice Leaves.
(A) Total genomic DNA was extracted from M. gnsea-infected rice plants(72 hr after inoculation) and from M. grisea liquid grown hyphae. DNAwas quantified by fluorometry and a dilution series prepared. The DNAsamples were digested with EcoRI, fractionated in a 0.8% agarosegel, blotted, and hybridized with a 6.4-kb Sail fragment of the singlecopy gene ILV1 excised from pCB573 (Valent and Chumley, 1991). Thisprobe failed to hybridize to rice genomic DNA (see [B]). Here 5.0 ugof infected rice DNA hybridized to the ILV1 probe with equal intensityas did 0.5 ng of M. grisea DNA, suggesting that 10% of the DNA ex-tracted from infected rice leaves is fungal DNA. This was confirmedby laser densitometry of the autoradiographs (data not shown). Theamount of fungal biomass present was calculated from this figure (seetext for details).(B) The progress of fungal infection was assessed by performing thisanalysis on DNA extracted at various time points following inoculationof rice plants (lane H contains DNA extracted from healthy uninfectedrice plants). No hybridization was observed prior to 48 hr after inocu-lation. Between 72 and 96 hr, the proportion of fungal DNA in the infectedrice plant samples doubled.Control hybridizations for all experiments were performed with a single-copy rice gene (pNJT-14) as shown in the lower panels.
plant than in culture, and the mRNA detected with the NJT-70clone (Figure 1D) was 3.5 times more abundant.
The biomass estimates were also used to follow the rate offungal ingress, as shown in Figure 2B. M. grisea biomass ap-pears to have more than doubled between 72 and 96 hr afterinoculation. This dramatic increase is consistent with micro-scopic observations that show very rapid and extensivecolonization of the plant leaf area prior to symptom develop-ment, which occurs approximately 96 hr after initial infection(Peng and Shishiyama, 1988; Heath et al., 1990a).
We also compared the signals of infection-induced M. griseacDNAs with NJT-23, a putative constitutively expressed M.grisea cDNA (Figure 1C). Later we show that the NJT-23 tran-script is probably not constitutively expressed at all times during
1578 The Plant Cell
the infection but is abundant at the 72-hr time point examinedin this experiment. Normalization of the hybridization signalssuggested that the NJT-15 cDNA represents an mRNA spe-cies 40-fold more abundant in the plant than in culture andthe NJT-70 cDNA, an mRNA species three times more abun-dant in the plant than in culture. Thus, the estimates of fungalmRNA levels in the plant normalized relative to another geneare similar to those derived from estimations of fungal biomass.
Characterization and Genetic Mapping of MPG1
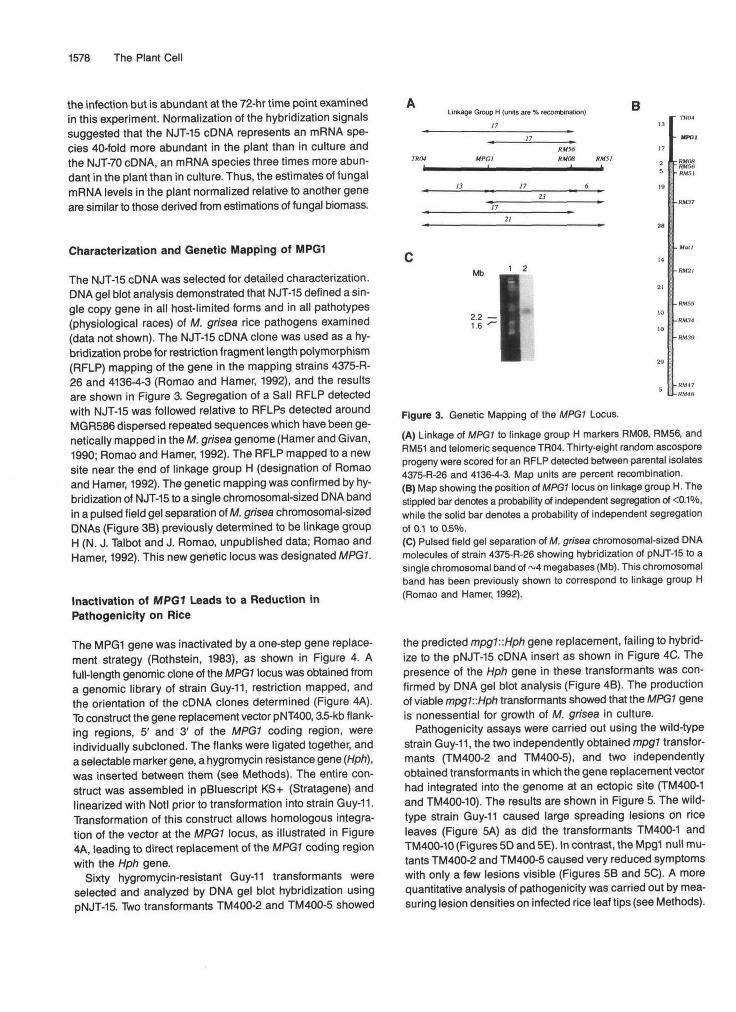
The NJT-15 cDNA was selected for detailed characterization.DNA gel blot analysis demonstrated that NJT-15 defined a sin-gle copy gene in all host-limited forms and in all pathotypes(physiological races) of M. grisea rice pathogens examined(data not shown). The NJT-15 cDNA clone was used as a hy-bridization probe for restriction fragment length polymorphism(RFLP) mapping of the gene in the mapping strains 4375-R-26 and 4136-4-3 (Romao and Hamer, 1992), and the resultsare shown in Figure 3. Segregation of a Sail RFLP detectedwith NJT-15 was followed relative to RFLPs detected aroundMGR586 dispersed repeated sequences which have been ge-netically mapped in the M. grisea genome (Hamer and Givan,1990; Romao and Hamer, 1992). The RFLP mapped to a newsite near the end of linkage group H (designation of Romaoand Hamer, 1992). The genetic mapping was confirmed by hy-bridization of NJT-15 to a single chromosomal-sized DNA bandin a pulsed field gel separation of M. grisea chromosomal-sizedDNAs (Figure 3B) previously determined to be linkage groupH (N. J. Talbot and J. Romao, unpublished data; Romao andHamer, 1992). This new genetic locus was designated MPG1.
Inactivation of MPG1 Leads to a Reduction inPathogenicity on Rice
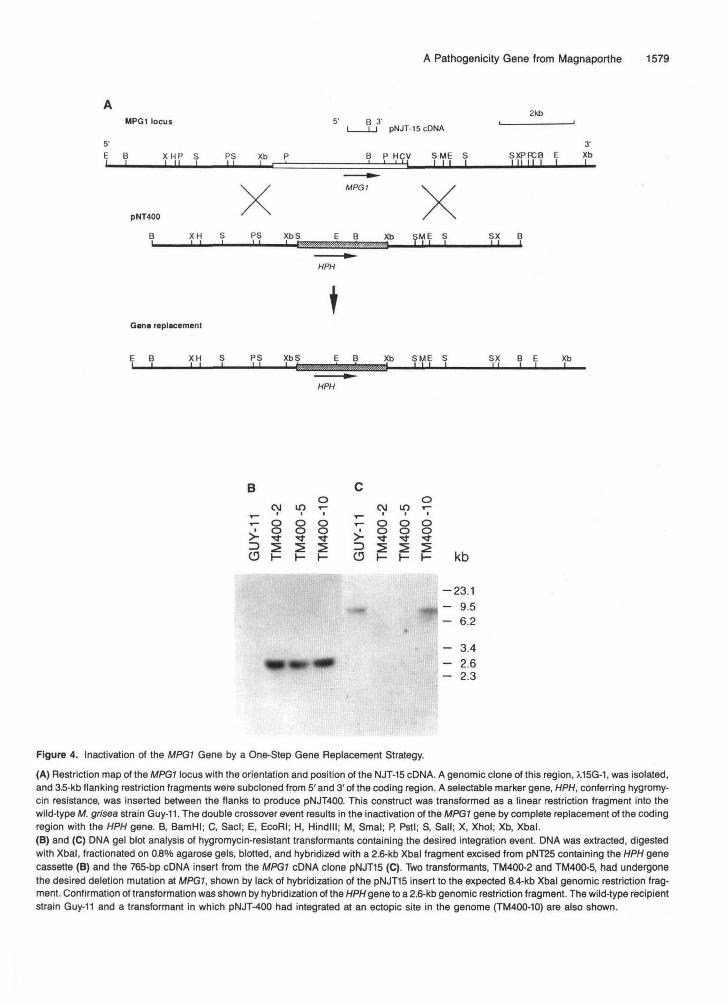
The MPG1 gene was inactivated by a one-step gene replace-ment strategy (Rothstein, 1983), as shown in Figure 4. Afull-length genomic clone of the MPG1 locus was obtained froma genomic library of strain Guy-11, restriction mapped, andthe orientation of the cDNA clones determined (Figure 4A).To construct the gene replacement vector pNT400,3.5-kb flank-ing regions, 5' and 3' of the MPG1 coding region, wereindividually subcloned. The flanks were ligated together, anda selectable marker gene, a hygromycin resistance gene (Hph),was inserted between them (see Methods). The entire con-struct was assembled in pBluescript KS+ (Stralagene) andlinearized with Notl prior to transformation into strain Guy-11.Transformation of this construct allows homologous integra-tion of the vector at the MPG1 locus, as illustrated in Figure4A, leading to direct replacement of the MPG1 coding regionwith the Hph gene.
Sixty hygromycin-resistant Guy-11 transformants wereselected and analyzed by DNA gel blot hybridization usingpNJT-15. Two transformants TM400-2 and TM400-5 showed
Linkage Group H (units are % recombination)
17
B
HMDS RM5I
1 2
~ TOM
MPGl
rRMOSRM56RM51
-RM55
-RM34
-RM39
-KM47-RM4S
Figure 3. Genetic Mapping of the MPGl Locus.(A) Linkage of MPG1 to linkage group H markers RM08, RM56, andRM51 and telomeric sequence TR04. Thirty-eight random ascosporeprogeny were scored for an RFLP detected between parental isolates4375-R-26 and 4136-4-3. Map units are percent recombination.(B) Map showing the position of MPG1 locus on linkage group H. Thestippled bar denotes a probability of independent segregation of <0.1°/o,while the solid bar denotes a probability of independent segregationof 0.1 to 0.5%.(C) Pulsed field gel separation of M. grisea chromosomal-sized DNAmolecules of strain 4375-R-26 showing hybridization of pNJT-15 to asingle chromosomal band of ~4 megabases (Mb). This chromosomalband has been previously shown to correspond to linkage group H(Romao and Hamer, 1992).
the predicted mpgT.:Hph gene replacement, failing to hybrid-ize to the pNJT-15 cDNA insert as shown in Figure 4C. Thepresence of the Hph gene in these transformants was con-firmed by DNA gel blot analysis (Figure 4B). The productionof viable mpg1::Hph transformants showed that the MPG1 geneis nonessential for growth of M. grisea in culture.
Pathogenicity assays were carried out using the wild-typestrain Guy-11, the two independently obtained mpgl transfor-mants (TM400-2 and TM400-5), and two independentlyobtained transformants in which the gene replacement vectorhad integrated into the genome at an ectopic site (TM400-1and TM400-10). The results are shown in Figure 5. The wild-type strain Guy-11 caused large spreading lesions on riceleaves (Figure 5A) as did the transformants TM400-1 andTM400-10 (Figures 5D and 5E). In contrast, the Mpg1 null mu-tants TM400-2 and TM400-5 caused very reduced symptomswith only a few lesions visible (Figures 5B and 5C). A morequantitative analysis of pathogenicity was carried out by mea-suring lesion densities on infected rice leaf tips (see Methods).
A Pathogenicity Gene from Magnaporthe 1579
2KbMPG1 locus 5' B 31 ,_______
I___u pNJT15cDNA
E B XHP S PS Xb P B P HCV S ME S SXPPCB E XbI I________I II I______II I ,——I————————————————I——1——L-LJ III I I II I I I I_____I
B P HCV SME S- I I l-M Ml |
————»MPG1
PNT400
B_____XH S PS XbS_____JL..JL,,,,,,,Jfb ?ME s SX B
HPH
Gene replacement
E B XH S PS XbS E B Xb SME S SX B E XbI——I————LJ——I———U———L-immmmiii>;ml>>#m>#»i———UJ—I______" ' '———L_
HPH
B Co oCM m r- CM in i-,_ , , ^ • . •
T - O O O i - O O O• ooo • o o o> - ^ r t t > - • * • < * • « *
O K K H O H H H k b
-23.1
~ 9'5_ 62
- 3.4- 2.6- 2.3
Figure 4. Inactivation of the MPG1 Gene by a One-Step Gene Replacement Strategy.
(A) Restriction map of the MPG1 locus with the orientation and position of the NJT-15 cDNA. A genomic clone of this region, A.15G-1, was isolated,and 3.5-kb flanking restriction fragments were subcloned from 5' and 3' of the coding region. A selectable marker gene, HPH, conferring hygromy-cin resistance, was inserted between the flanks to produce pNJT400. This construct was transformed as a linear restriction fragment into thewild-type M. grisea strain Guy-11. The double crossover event results in the inactivation of the MPG1 gene by complete replacement of the codingregion with the HPH gene. B, BamHI; C, Sacl; E, EcoRI; H, Hindlll; M, Smal; P, Pstl; S, Sail; X, Xhol; Xb, Xbal.(B) and (C) DMA gel blot analysis of hygromycin-resistant transformants containing the desired integration event. DNA was extracted, digestedwith Xbal, fractionated on 0.8°/o agarose gels, blotted, and hybridized with a 2.6-kb Xbal fragment excised from pNT25 containing the HPH genecassette (B) and the 765-bp cDNA insert from the MPG1 cDNA clone pNJT15 (C). Two transformants, TM400-2 and TM400-5, had undergonethe desired deletion mutation at MPG1, shown by lack of hybridization of the pNJT15 insert to the expected 8.4-kb Xbal genomic restriction frag-ment. Confirmation of transformation was shown by hybridization of the HPH gene to a 2.6-kb genomic restriction fragment. The wild-type recipientstrain Guy-11 and a transformant in which pNJT-400 had integrated at an ectopic site in the genome (TM400-10) are also shown.

1580 The Plant Cell
Figure 5. Inactivation of MPG1 Leads to Reduced Pathogenicity onRice.Conidial suspensions were prepared from Mpg1 deletion mutants andcontrol strains and sprayed onto 14-day-old seedlings of the compati-ble rice cultivar CO-39. The plants were incubated to allow diseasedevelopment for 96 hr. Leaves shown were from plants inoculated withthe following strains.(A) Wild-type Guy-11.(B) Mpg1 strain TM400-2.(C) Mpg1 strain TM400-5.(D) Ectopic transformant TM400-1.(E) Ectopic transformant TM400-10.Mpg1 mutants produced significantly fewer lesions. The experimentwas repeated four times with identical results using 160 seedlings perexperiment.
Mean lesion density was reduced from 37.4 per 5-cm leaf tipin the wild-type isolate Guy-11 to 7.5 per 5-cm leaf tip in TM400-2(f = 12.65, P < 0.001, df = 78). Mean lesion density was simi-larly reduced in TM400-5 to 7.3 per 5-cm leaf tip, but wasunchanged in both TM400-1 and TM400-10.
mpg1::Hph Strains Show a Reduced Ability ToForm Appressoria
The reduced lesion density suggested that the Mpg1 null mu-tants were defective in early steps in the infection process. Theability of the mpg1::Hph strains to form appressoria was as-sayed on Teflon (DuPont) membranes (Hamer et al., 1988).Figure 6A shows differentiated appressoria from the wild-typestrain Guy-11 in which 70.8% of the conidia examined (n =1200) produced appressoria. The pyriformed three-celledconidia germinate and form a round melanized appressoria.The transformants TM400-1 and TM400-10 were unchangedin their ability to form appressoria relative to strain Guy-11 (datanot shown). Figure 6B shows the reduced frequency of ap-pressorial formation in strain TM400-2. Conidia from this straingerminate normally (data not shown) but failed to form appres-soria at a high frequency. We found that only 18.3% of conidia
examined on Teflon (n = 1200) from TM400-2 formed appres-soria (x2 = 39.09, P < 0.001). Similarly, 20.6% of conidia (n =1200) from TM400-5 produced appressoria (x2 = 35.96, P <0.001). The inset (Figure 6C) shows a close-up of a typicalconidium that failed to produce an appressorium. A long germtube has formed that appears to have undergone multiple hook-ing and swelling events. Previous studies (Bourett and Howard,1990) have shown that hooking and swelling of the germ tubeoccur prior to appressorium formation. Thus, Mpg1 null mu-tants appear to undergo primary differentiation events that
A» **̂ "2*,: "" ^V 0 » » -r'%*
* * • ̂ ^$&*^>J '- *«P
?si*f?A,L,/ -y^v.-g''a. • -•'3fc<<i • *v .c4*t-/*-̂ f-r;ri.s«S|TrFigure 6. Reduced Ability of Mpg1 Mutants To Undergo Infection-Related Morphogenesis.
Conidial suspensions were prepared from Mpg1 mutants and the wild-type strain Guy-11 and tested for their ability to make appressoria onTeflon membranes. Photomicrographs of typical fields of view fromthis analysis are shown.(A) Three-celled pyriform conidia of Guy-11 producing highly melanizedround appressoria. Bar = 40 urn.(B) The Mpg1 mutant TM400-2 that has a significantly reduced abilityto undergo appressorial development. Bar = 40 urn.(C) Detailed view of the phenotype of conidia from the Mpg1 mutant.Long germ tubes appear to undergo swelling at multiple points alongthe hyp ha but do not differentiate fully into an appressorium. Bar =15 urn.Assays for appressorial development were repeated four times count-ing a total of 1200 conidia from each strain.
A Pathogenicity Gene from Magnaporthe 1581
Time after inoculation (h)
12 18 24 48 72 96
B
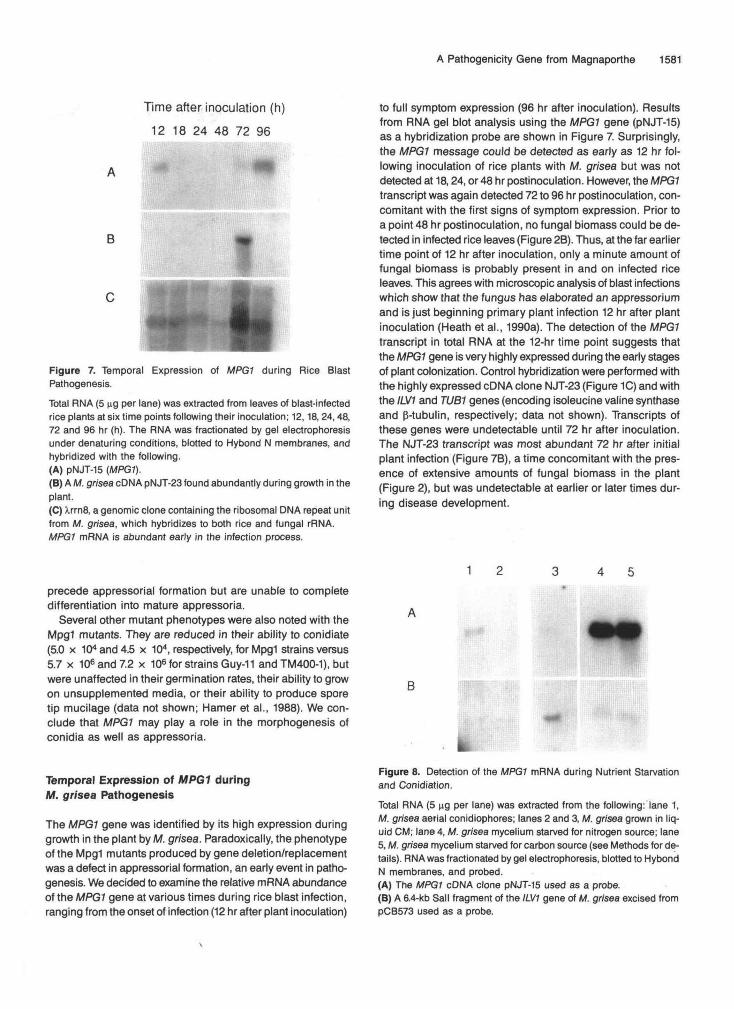
Figure 7. Temporal Expression of MPGf during Rice BlastPathogenesis.Total RNA (5 ng per lane) was extracted from leaves of blast-infectedrice plants at six time points following their inoculation; 12,18, 24,48,72 and 96 hr (h). The RNA was fractionated by gel electrophoresisunder denaturing conditions, blotted to Hybond N membranes, andhybridized with the following.(A) pNJT-15 (MPG1).(B) A M. grisea cDNA pNJT-23 found abundantly during growth in theplant.(C) XrrnS, a genomic clone containing the ribosomal DMA repeat unitfrom M. grisea, which hybridizes to both rice and fungal rRNA.MPG1 mRNA is abundant early in the infection process.
to full symptom expression (96 hr after inoculation). Resultsfrom RNA gel blot analysis using the MPG1 gene (pNJT-15)as a hybridization probe are shown in Figure 7. Surprisingly,the MPG1 message could be detected as early as 12 hr fol-lowing inoculation of rice plants with M. grisea but was notdetected at 18,24, or 48 hr postinoculation. However, the MPG1transcript was again detected 72 to 96 hr postinoculation, con-comitant with the first signs of symptom expression. Prior toa point 48 hr postinoculation, no fungal biomass could be de-tected in infected rice leaves (Figure 2B). Thus, at the far earliertime point of 12 hr after inoculation, only a minute amount offungal biomass is probably present in and on infected riceleaves. This agrees with microscopic analysis of blast infectionswhich show that the fungus has elaborated an appressoriumand is just beginning primary plant infection 12 hr after plantinoculation (Heath et al., 1990a). The detection of the MPG1transcript in total RNA at the 12-hr time point suggests thatthe MPG1 gene is very highly expressed during the early stagesof plant colonization. Control hybridization were performed withthe highly expressed cDNA clone NJT-23 (Figure 1C) and withthe ILV1 and TUB1 genes (encoding isoleucine valine synthaseand p-tubulin, respectively; data not shown). Transcripts ofthese genes were undetectable until 72 hr after inoculation.The NJT-23 transcript was most abundant 72 hr after initialplant infection (Figure 7B), a time concomitant with the pres-ence of extensive amounts of fungal biomass in the plant(Figure 2), but was undetectable at earlier or later times dur-ing disease development.
1 2 3 4 5precede appressorial formation but are unable to completedifferentiation into mature appressoria.
Several other mutant phenotypes were also noted with theMpg1 mutants. They are reduced in their ability to conidiate(5.0 x 10* and 4.5 x 104, respectively, for Mpg1 strains versus5.7 x 106 and 7.2 x 106 for strains Guy-11 and TM400-1), butwere unaffected in their germination rates, their ability to growon unsupplemented media, or their ability to produce sporetip mucilage (data not shown; Hamer et al., 1988). We con-clude that MPG1 may play a role in the morphogenesis ofconidia as well as appressoria.
Temporal Expression of MPG1 duringM. grisea Pathogenesis
The MPG1 gene was identified by its high expression duringgrowth in the plant by M. grisea. Paradoxically, the phenotypeof the Mpg1 mutants produced by gene deletion/replacementwas a defect in appressorial formation, an early event in patho-genesis. We decided to examine the relative mRNA abundanceof the MPG1 gene at various times during rice blast infection,ranging from the onset of infection (12 hr after plant inoculation)
Figure 8. Detection of the MPG1 mRNA during Nutrient Starvationand Conidiation.
Total RNA (5 ng per lane) was extracted from the following: lane 1,M. grisea aerial conidiophores; lanes 2 and 3, M. grisea grown in liq-uid CM; lane 4, M. grisea mycelium starved for nitrogen source; lane5, M. grisea mycelium starved for carbon source (see Methods for de-tails). RNA was fractionated by gel electrophoresis, blotted to HybondN membranes, and probed.(A) The MPG1 cDNA clone pNJT-15 used as a probe.(B) A 6.4-kb Sail fragment of the ILV1 gene of M. grisea excised frompCB573 used as a probe.

1582 The Plant Cel!
A
B
-148
-69
11
90
156 14
216 34
276 54
336 74
396 94
4 68 101
533
612
691
770
849
s S PS Xb P B P HCV SME S SXPFCB 3'
MPGl E"-
1 k b d J pNJT-15 (cDNA) - pNJTG15XB
GGCCCTGCGTTGAGGCCCGAGCTGCCTGCCTGCCTGACCCTGACCCTGACTAACTGACTGAGTGACTGAGGGACT~GA
I+- CGGTCCCGGATCGTGGATAAATAGAAGGC'ITCCTCCCTCCCGATCAACAAACTTTTCCTGTCTCTTCTCATCA~CCAA
ATGCTCACCATAGCAATAGCCACTCTTTAGACTCCCACTTCCAAACTACTTAATATCTAATTCCTTGTC'IT~CCCGAG
ACCCCTCTCAACCCAACCTAA'M'CAAG ATG TTC TCC CTC AAG ACC GTT GTC CTG GCC C T T GCC GCC met phe ser leu lys thr val val leu ala leu ala ala
GCC GCC TTC GTC CAG GCC ATC CCC GCC CCT GGT GAG GGT CCT TCC GTC T C C ATG GCT CAG ala ala phe val gln ala ile pro ala pro gly glu gly pro ser val ser met ala gln
CAG AAG TGC GGT GCC GAG AAG GTC GTC TCT TGC TGC AAC AGC AAG GAG C T T AAG AAC TCC gln lys cys gly ala glu lys val val ser cys cys asn ser lys glu leu lys asn ser
AAG T C T GGT GCC GAG ATC CCC ATC GAC GTT CTC AGC GGC GAG TGC AAG AAC ATC CCG ATC lys ser gly ala glu ile pro ile asp val leu ser gly glu cys l y s asn ile pro ile
AAC ATC CTC ACC ATC AAC CAG CTC ATC CCC ATC AAC AAC T T C TGC TCG GAC ACC G T T TCG asn ile leu thr ile asn gln leu ile pro ile asn asn phe cys ser asp thr val ser
TGC TGC TCC GGC GAG CAG ATT gtgagttcctcttgtgacgctcctttctcattcaacacacagtctactgac cys cys ser gly glu gln ile
cacagct tc t tc t t t cacaaatacag GGT CTG GTC AAC ATC CAG TGC ACT C C T ATC CTG AGC TAA gly leu val asn ile gln cys thr pro ile leu ser -
ATGGCTTAATGTCTTTACCGACATCTAGCTA?TTTCTCACATCAGGGATGCATGCGATCACGAGGGGATGATGGTGGTG
GAGGACGTTTGGAGTl~CTGTGAGGGAGCCGGAGTCCGGCGATCAGCGTTGAGTGAAAGCCGGTCCTGGCTGCTGGGGG
CCCTGGTTTCGTCCTAAGCACCTGGCGGCCGTGGAATGGCTAGGCACACAATCCGCC~TGTC~GC~GAGGGA
T T G A T G T C A A G T T T T G G A A G C A T C T G G A A G G A T G C C C G C T A G A G A A G T C ~ ~ C T C G ~ C C A C ~ ~ A C C C C T T C
CAGACCTATTTA
BamHI
Figure 9. Transcriptional Organization and Sequence of the MPG7 Locus.
(A) Restriction map of the MPGl locus showing orientation of the cDNA clone pNJT-15 and the genomic subclone pNJT15GXB. 6, BamHI; E, ECORI; H, Hindlll; M, Smal; P, Pstl; S, Sall; V, EcoRV; X, Xhol; Xb, Xbal. (B) DNA sequence of the transcriptional unit of MPG7. The transcription initiation site was determined by primer extension mapping. The two 'major 5' ends are indicated by the arrow at positions +1 and +4. A putative TATA box (-52) in the 5' region of the gene is overlined. A 77-bp intron was identified by comparison of the cDNA and genomic sequence and is shown in lowercase letters. The hexanucleotide polyadenylation signal sequence in the 3'untranslated region of the gene is underlined at position 835. The poly(A) addition sites are underlined and were deduced from the cDNA sequences of two independent clones.

A Pathogenicity Gene from Magnaporthe 1583
1 50 80
sc 4 ...... mrFs LAlLALpAlA AAaPvpgggk ....... ..g Agqa CnS..Gp.Vq CCNe...... sc 3 mfarlpwF1 YAfvAFgAlv AAlPgghpgt typpstttia ............. :..Aggt Ctt..Gs.ls CCNq......
....... mFk alIvALAAvA AAiPtqqps. ......... 9 . . . . . . . . . . . . . . . . n e m CdS..G..Vy CC Nk...... ssg Roda .... mkfsia aAVvAFAAsV AAlPpahdsq fagngvgnkg nsnvkfpvpenvtvkqAsdk Cgd..qaqls CCNkatyagd ..... d a L k tvVLALM.. MfvQaipap .................. gegpavadqqk Cga..akvVa CCNS ...... lanl Cerato-Ulmin . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . sdsydp Ctgllqkspq CCNt . . . . . . Eas .... mqftsv FtILAiAmtA AAaPaewpr ...................... attigpnt C.Siddykpy CCqs . . . . . .
sc 1 ...... mpFs LAILALpvlA AAta vp... r ......... g ................ gask CnS..Gp.Vq CCNt... ... ................
Consensus --------F- LA1LAM-A =-e------ _ _ _ _ _ _ _ _ _ - ----------------A--- C-S--G--V- CCN-------
81 100 lvdtkdkhqt nivgaLlGldlGsltgLaGv ... nCS ..... PvsV1gV.g. GnsCstQlV. tttvanaqkq glLggLlGvwGpitgLvG1 .. nCS ..... PisWgVlt. GnsCtAQlV. vqsassspvt alLg.LlGivlsdlnvLvGi .. sCS..... PLtV1gV.g. GsgCsAQlV. v . . . . . aqnt giwpidal ......... ss .. tCg .... d tLklVtVda1 ndkCtsQlV. ............. keLknaksQaeipidv1 ageCknipin LLtInqlipi nnfcsdtva. ........................ diLgva nlahgppsv Ptspsq ... f qasCvAdggr . . . msgpags pgLlnLipvdlsa ....... sl gc...... ..... vVgvi GsqCgAsv..
tttvdeglls gaLsgLiGagsGaegl..Gl fdqCSkldva vLigIqd1.v nqkckqnia.
_ _ _ _ _ _ _ _ _ _ --L--L-G---G----L-G- ---CS----- pL-VI-V--- G--C-AQW-
150 172 ... CCeg ..... tqENGLVN Vg.CT.PInV gl ... CC dh..... vtqNGLVN Vg.CT.PIs1 ... CCen . . . . . tq€NGLIN Ig.CT.PIn1 1. . . . CQm ..... vqqNGLVN Va.CT.PIdV ... CCqnspssadgnligVg 1p.Cv.algs i1 ... CC ......OgeQiQLVN Iq.CT.PI1S ..
..
..
sarCC ....... tlsllgla 1v.CTdPVgI . ..kCCkddvtntgnsfliIN aanCva . . . . . . ---CC----------NGLlJN V--Cp-pI-- _ _
Figure 10. Comparison of the Polypeptide Sequence Predicted by MPG7 with the Funga1 Hydrophobins.
The predicted MPGl polypeptide sequence was aligned with the known hydrophobins. Scl, Sc3, and Sc4 are proteinsfrom S. commune (Wessels, 1992); ssgA, rodA, and Eas are translated from genes of M. anisopliae (St. Leger et al., 1992). A. nidulans, and N. crassa (Bell-Pederson et al., 1992; Lauter et al., 1992); cerato-ulmin is a peptide from O. ulmi (Bolyard and Sticklen, 1992; Stringer and Timberlake, 1993). The sequences were aligned based on the conserved cysteine residues (shown in bold) using the GCG PILEUP program (Devereaux et al., 1984). Highly con- served amino acid residues are shown in uppercase letters and as a consensus sequence.
MPG7 1s Expressed during Nutrient Starvation and Conidiation
We reasoned that the elevated expression of M E 7 during rice blast infection could be due to regulation by either an exoge- nous plant signal and/or by nutrient deprivation. To dis!inguish between these possibilities, strain Guy-11 was grown in com- plete media(CM) for 4 days. The mycelium was then removed to a medium lacking either a nitrogen or a carbon source for a further 12 hr (for details, see Methods). RNA was extracted from the mycelium and gel blots were prepared and hybrid- ized with the MPGl gene. Figure 8A shows that the steady state MPGl mRNA leve1 was significantly elevated by starva- tion for either nitrogen or carbon.
The late expression of MPGl in rice blast pathogenesis is concomitant with lesion formation and the production of aer- ia1 conidiophores. Mpgl mutants also produce fewer conidia. We thus examined the expression of MPGl during conidia- tion in culture. Total RNA was extracted from aerial conidiating hyphae of M. grisea, and RNA gel blots were prepared and hybridized with the MPG7 gene. Figure 8 also shows that the MPGl message can be detected in conidiating cultures but not in mycelium growing in CM.
MPG7 Encodes a Hydrophobin-like Protein
A detailed map of the MPGl locus is illustrated in Figure 9A, showing the positions of the NJT-15 cDNA and the pNJT15GXB genomic subclone. DNA sequence information was obtained from both of these clones, encompassing the entire 2.7-kb re- gion of the MPGl locus which was deleted in the mpg7::Hph
transformants. RNA gel blot analysis with pNJT15GXB con- firmed that only a single 850-bp mRNA was transcribed from this region. The longest open reading frame identified within this 2.7-kb Xbal-BamHI fragment was 336 bp long and predicted a protein of 112 amino acids with a molecular mass of 11.5 kD. The DNA and predicted amino acid sequences together with various landmark features of the transcription unit of MPGl are shown and described in Figure 96. We noted that the N terminus of the predicted MPGl protein contained a tract of amino acids with features consistent with those of a signal se- quence (Heijne, 1983), suggesting that MPGl encodes a secreted peptide. A single 77-bp intron was located in the MPG7 gene by comparison of the genomic and cDNA sequences con- taining consensus funga1 splice sequences (Ballance, 1986).
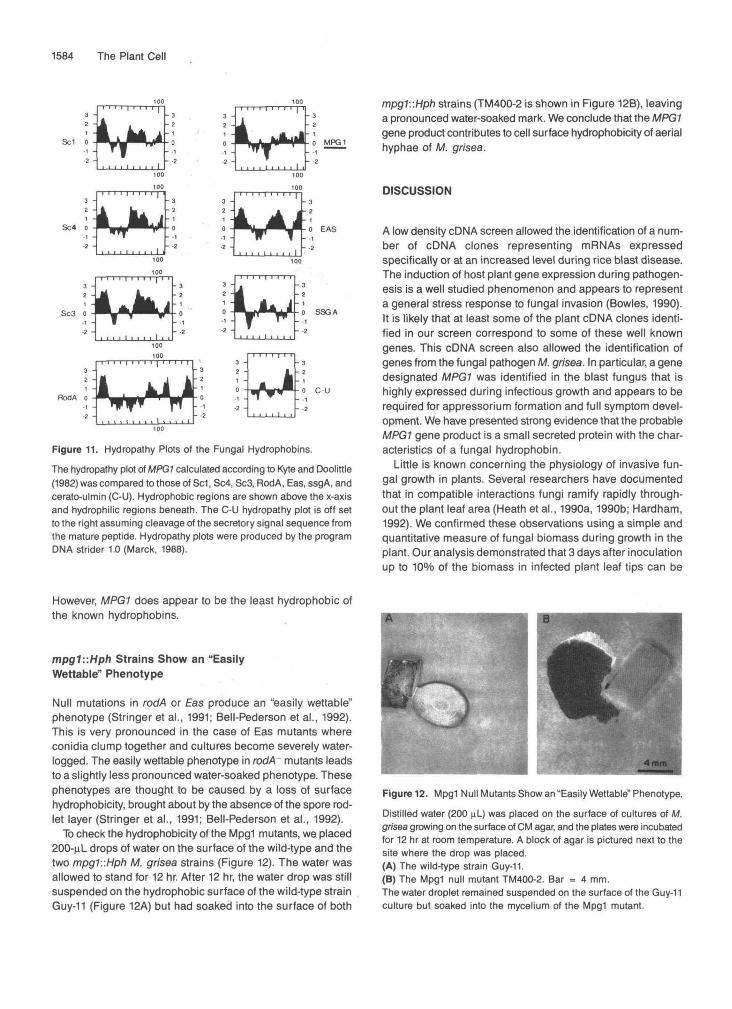
The MPGl sequence predicted a small secreted, moderately hydrophobic protein containing eight cysteine residues. These features are characteristic of a class of proteins known as the hydrophobins which have been recently identified in Schizophyllum commune (for review, see Wessels, 1992), Aspergillus nidulans (Stringer et al., 1991), and Neurospora crassa (Bell-Pederson et al., 1992; Lauter et al., 1992). The amino acid sequences of the hydrophobins have two main distinguishing characteristics: the conservation of the spac- ing of eight cysteine residues and the conservation of hydrophobic domains within the polypeptide sequence. Computer-generated amino acids sequence alignments of MPGl with the known hydrophobin amino acid sequences, given in Figure 10, showed a similar spacing of the cysteine residues in MPG7. Figure 11 shows an alignment of the hydropathy plot of the MPGl amino acid sequence with those of the known hydrophobins. This comparison shows that MPGl has the characteristic hydropathy profile of a hydrophobin.
1584 The Plant Cell
Sc1
Sc4
Sc3
MPG1
EAS
SSGA
C U
Figure 11. Hydropathy Plots of the Fungal Hydrophobins.The hydropathy plot of MPG1 calculated according to Kyte and Doolittle(1982) was compared to those of Sc1, Sc4, Sc3, RodA, Eas, ssgA, andcerato-ulmin (C-U). Hydrophobic regions are shown above the x-axisand hydrophilic regions beneath. The C-U hydropathy plot is off setto the right assuming cleavage of the secretory signal sequence fromthe mature peptide. Hydropathy plots were produced by the programDMA strider 1.0 (Marck, 1988).
mpg1::Hph strains (TM400-2 is shown in Figure 12B), leavinga pronounced water-soaked mark. We conclude that the MPG1gene product contributes to cell surface hydrophobicity of aerialhyphae of M. grisea.
DISCUSSION
A low density cDNA screen allowed the identification of a num-ber of cDNA clones representing mRNAs expressedspecifically or at an increased level during rice blast disease.The induction of host plant gene expression during pathogen-esis is a well studied phenomenon and appears to representa general stress response to fungal invasion (Bowles, 1990).It is likely that at least some of the plant cDNA clones identi-fied in our screen correspond to some of these well knowngenes. This cDNA screen also allowed the identification ofgenes from the fungal pathogen M. grisea. In particular, a genedesignated MPG1 was identified in the blast fungus that ishighly expressed during infectious growth and appears to berequired for appressorium formation and full symptom devel-opment. We have presented strong evidence that the probableMPG1 gene product is a small secreted protein with the char-acteristics of a fungal hydrophobin.
Little is known concerning the physiology of invasive fun-gal growth in plants. Several researchers have documentedthat in compatible interactions fungi ramify rapidly through-out the plant leaf area (Heath et al., 1990a, 1990b; Hardham,1992). We confirmed these observations using a simple andquantitative measure of fungal biomass during growth in theplant. Our analysis demonstrated that 3 days after inoculationup to 10% of the biomass in infected plant leaf tips can be
However, MPG1 does appear to be the least hydrophobic ofthe known hydrophobins.
mpg1::Hph Strains Show an "EasilyWettable" Phenotype
Null mutations in rodA or Eas produce an "easily wettable"phenotype (Stringer et al., 1991; Bell-Pederson et al., 1992).This is very pronounced in the case of Eas mutants whereconidia clump together and cultures become severely water-logged. The easily wettable phenotype in rodA~ mutants leadsto a slightly less pronounced water-soaked phenotype. Thesephenotypes are thought to be caused by a loss of surfacehydrophobicity, brought about by the absence of the spore rod-let layer (Stringer et al., 1991; Bell-Pederson et al., 1992).
To check the hydrophobicity of the Mpg1 mutants, we placed200-nL drops of water on the surface of the wild-type and thetwo mpgT.'.Hph M. grisea strains (Figure 12). The water wasallowed to stand for 12 hr. After 12 hr, the water drop was stillsuspended on the hydrophobic surface of the wild-type strainGuy-11 (Figure 12A) but had soaked into the surface of both
Figure 12. Mpg1 Null Mutants Show an "Easily Wettable" Phenotype.
Distilled water (200 nL) was placed on the surface of cultures of M.grisea growing on the surface of CM agar, and the plates were incubatedfor 12 hr at room temperature. A block of agar is pictured next to thesite where the drop was placed.(A) The wild-type strain Guy-11.(B) The Mpg1 null mutant TM400-2. Bar = 4 mm.The water droplet remained suspended on the surface of the Guy-11culture but soaked into the mycelium of the Mpg1 mutant.

A Pathogenicity Gene from Magnaporthe 1585
attributed to M. grisea. Furthermore, this proportion appears to increase dramatically as infection continues. DNA gel blot biomass measurements can be used to quantify race-culti- var interactions and is broadly applicable to analyzing various properties of infectious growth, including the effects of vari- ous antifungal chemicals. The technical simplicity of this approach contrasts with other methods that utilize fungal spe- cific polymers, such as chitin or ergosterol, or rely on the use of transformed fungal pathogens carrying reporter gene constructs.
The Role of MPG7 in Pathogenesis
MPG7 was selected as a gene likely to be important in the es- tablishment of plant infection because of its high leve1 of expression in the plant and its conservation in a broad collec- tion of different races and host-specific forms of M. grisea. Gene deletionlreplacement mutants of MPGl are severely reduced in their ability to cause disease symptoms on susceptible rice cultivars due to an inability to form appressoria. In this report, we have shown that the probable MPG7 gene product has char- acteristic features of a fungal hydrophobin. These include conservation of the spacing of eight cysteine residues, including a conserved tripeptide, CCN, present in seven of the eight known hydrophobins, and conservation of hydrophobic do- mains throughout the polypeptide. MPGl also shows a high degree of amino acid similarity, including a large block of iden- tical amino acids, to the S. commune hydrophobins and ssgA at its C terminus. Furthermore, the Mpgl null mutants have an “easily wettable” phenotype, suggesting a reduction in cell surface hydrophobicity, similar to that observed in rodA- and €as- mutants of A. nidulans and N. crassa.
The highest levelsof expression of MPG7 during pathogen- esis were found to be very early in the infection process during appressorial development (12 hr after plant inoculation) and again very late during the disease cycle at a point coincident with symptom expression. The finding that MPG7 transcript levels are very abundant during appressorial differentiation and early plant infection is consistent with the observation that Mpgl mutants are impaired in their ability to undergo ap- pressorial development. Although a generalized role for hydrophobins in the emergence of aerial structures in fungi has been proposed (Chasan, 1991; Wessels, 1992; Wosten et al., 1993), this model would not readily explain the role of these molecules in plant pathogenesis.
Appressorial development occurs in an aqueous environ- ment and is triggered by interactions with a hydrophobic surface (Hamer et al., 1988). Indeed, the mechanical action of appressoria appears to require the influx of water to create the high interna1 cell turgor pressure needed for penetration (Howard et al., 1991b). According to recent models for hydro- phobin function, the MPGl hydrophobin would not be expected to accumulate in appressorial cell walls in an aqueous envi- ronment. Rather, it would be secreted onto the plant surface and most probably bind to the leaf cuticle through hydrophobic
interactions. Recently, it has been shown that purified hydrophobins can self-assemble at any hydrophilic/hydropho- bic interphase (Wosten et al., 1993). Such an interphase may be present between the infecting fungal cell and the plant sur- face and thus MPG7 could play a role in attachment, infection court preparation, or topological signaling (for review, see Mendgen and Deising, 1993). Experiments to localize and study the accumulation of the MPGl gene product during the infection process are currently underway.
The late expression of MPGl during growth in the plant and the recent report that the wilt toxin cerato-ulmin is a fungal hydrophobin (Stringer and Timberlake, 1993) point to another potential role for the MPG7 protein. Late in pathogenesis the fungus is causing disease symptoms that, in a heavy infec- tion such as the one studied here, include the production of large spreading lesions across the leaf surface and a general dessication and wilting of the leaf tissue. This phenotype is coincident with high levels of MPGl transcript accumulation and could potentially be caused by secretion of large amounts of a small hydrophobic protein that could block xylem func- tion. However, it is difficult to test for the desiccation of leaves by the Mpgl null mutants due to their infection deficiency phenotype. We are currently investigating whether the MPGl hydrophobin can exert phytopathic effects.
Regulated Expression of MPG7
We also showed that MPGl transcript levels were elevated dur- ing conidiation and that Mpgl mutants are impaired in their ability to conidiate, suggesting a role for the gene product in conidiation. 60th the rodA and €as genes encode hy- drophobins that form the rodlet layer of conidial cell walls in A. nidulans and N. crassa, respectively (Stringer et al., 1991; Lauter et al., 1992), but neither mutant is impaired in conidia- tion. The MPGl gene product may be fulfilling a different role from rodA and €as during conidial development. Thus, while the presence of the MPGl gene contributes to cell surface hydrophobicity, it may not encode the major rodlet protein. Studies are in progress to localize and characterize the rodlet proteins of M. grisea.
Hydrophobin genes are regulated by a diverse array of regulatory genes and environmental cues. In the case of MPG7, transcript levels were elevated in RNA from cultures that had been starved for either a carbon or a nitrogen source. This suggests that one of the main environmental cues for appres- sorial morphogenesis may be nutrient starvation. Consistent with this idea, the ssgA gene from the entomopathogen M. anisopliae is similar to MPG7 in that it is regulated by nutrient deprivation and is also highly expressed during appressorium formation (St. Leger et al., 1992). The S. commune dikaryon- specific hydrophobins Sc7 and Sc4 are regulated by the mating-type genes and by wide domain regulators essential for basidiocarp elaboration (Wessels et al., 1991b; Wessels, 1992). It has been proposed that the rodA gene from A. nidu- lans is controlled by the early regulator of conidiation, BrlA

1586 The Plant Cell
(Stringer et al., 1991). The €as gene is regulated by circadian control (Bell-Pederson et al., 1992), blue light (Pandit and Russo, 1991), and may also be regulated by genes necessary for early conidiophore development in N. crassa (Lauter et al., 1992). This diversity of regulation of hydrophobin gene expres- sion highlights the complexity of environmental contingencies by which fungal cell differentiation is controlled.
Hydrophobins appear to be primarily related by structural features, and many fungi have now been shown to contain mul- tiple secreted hydrophobin-like proteins (de Vries et al., 1993). The predicted structure of hydrophobins, based on their amino acid sequences, is reminiscent of a variety of toxins including snake venoms and neurotoxins. The presence of eight cys- teine residues within these small(-lOO-amino acid) proteins predicts the formation of a small looped protein built around a core of four disulfide bridges (Drenth et al., 1980; St. Leger et al., 1992). An analysis of the three-dimensional structure of a hydrophobin may prove instrumental in understanding how they are able to play such diverse roles in development and pathogenesis.
METHODS
Funga1 lsolates and Culture Conditions
Strains of Magnaporthe grisea described in this study are stored in the laboratory of J.E. Hamer (Purdue University, West Lafayette, IN). Standard procedures for the culture and storage of M. grisea were used (Crawford et al., 1986) except complete medium (CM) was 10 gL-’ glu- cose, 2 gL-’ peptone, 1 gL-l yeast extract, 1 gL-’ casamino acids, O.lo/o (vlv) trace elements, O.lO/o (v/v) vitamin supplement, 6 gL-’ NaN03, 0.5 gL-’ KCI, 0.5 gL-l MgSO,, 1.5 gL-l KH2P04, pH 6.5.
M. grisea strain Guy-11 was grown in CM for 4 days, at 23OC, in the dark with vigorous aeration. At this time mycelium was removed by filtration, washed three times with sterile distilled water, and transferred to either CM-C containing all the constituents of CM except glucose, yeast extract, peptone, and casamino acids or CM-N containing all the constituents of CM except sodium nitrate, yeast extract, and casamino acids. Mycelium was incubated in this media for 12 hr with vigorous aeration before removal for RNA extraction.
To produce synchronous M. grisea conidiophores, a 2-cm2 agar block of strain Guy-11, grown on oatmeal agar, was fragmented in a blender and used to inoculate 50 mL of CM. After 2 days of growth at room temperature with shaking at 200 rpm, the culture was again blended and used to inoculate three 50-mL CM cultures, and these were grown for 2 days. At this stage some green pigmentation was observed but the hyphae in the culture were not darkly pigmented. Cultures were harvested by vacuum filtration onto 7-cm diameter filter paper circles (Whatman No. 541) to produce a thin mycelial pad. RNA was prepared from some pads immediately. For the remaining pads, the filter paper was placed mycelium-side down onto complete medium agar plates and placed in a 25°C-incubator with constant illumination. After 1 day, hyphae began to protrude through the filter paper. After 3 days, a uniform layer of aerial hyphae was present. After 5 days, a dense layer of gray aerial hyphae was observed with many mature and developing conidiophores present. The filter paper was removed
from the agar plate to a hard surface with the aerial hyphae-side up. A razor blade was used to shave the aerial hyphae from the filter pa- per, and the aerial material was used immediately to prepare RNA.
Rice lnfections
Blast infections were carried out using 14-day-old rice seedlings of cultivar CO-39. Seedlings were sprayed with conidial suspensions of the compatible M. grisea isolate Guy-11 or the engineered isolates de- scribed in this study derived from Guy-11. Conidial suspensions were routinely 104 conidia mL-’. lnoculations and plant incubation condi- tions were as described by Valent et al. (1991). Lesions were counted and mean lesion densities determined from randomly chosen 5-cm leaf tips. Mean lesion densities were compared using a t test (Sokal and Rohlf, 1981). Significant differences were confirmed using a non- parametric statistical analysis (Sokal and Rohlf, 1981), the Mann-Whitney two-sample test (data not shown).
Nucleic Acid lsolations and Analyses
Rice leaves were excised and quickly frozen in liquid NP for RNA ex- traction. Leaves were routinely stored in this form at -8OOC. RNA was extracted from plant tissues by a guanidine thiocyanate method (Sambrook et al., 1989). RNA was isolated from M. grisea by the method of Timberlake (1980). RNA was poly(A)+ enriched by oligo(dT) cellu- lose chromatography (Pharmacia) following manufacturer‘s sug- gestions. Small scale enrichment of poly(A)+ RNA was performed using the Polyattract kit (Promega).
Total or poly(A)+ RNA samples were denatured in 50% formamide, 2.2 M formaldehyde, 1 x MopslEDTA buffer (1 x MopdEDTA is 20 mM 3-(Kmorpholino)-propanesulfonic acid, 5 mM sodium acetate, 1 mM EDTA) at 65OC for 15 min. Gel electrophoresis was performed on 1.2% agarose gels containing 2.2 M formaldehyde in 1 x MopslEDTA. Gels were blotted onto Hybond-N (Amersham) and hybridizations were conducted according to manufacturer’s suggestions.
Genomic DNA extractions were carried out from rice leaves by the method of Dellaporta et al. (1983) and from M. grisea as described by Talbot et al. (1993).
Mass dependence of DNA extraction from rice and fungal tissues was shown in the following way. Two grams of rice leaf tissue or 2 g of fungal mycelium was taken, ground in liquid NP, and DNA extracted by the method of Dellaporta et al. (1983). A total of 10 replicates was performed. The extracted DNA was quantified in a TKO-100-min fluorimeter (Hoefer Scientific Instruments, San Francisco, CA) using the DNA-specific dye Hoechst 33258. Quantification was confirmed by spectrophotometry and direct visual inspection of ethidium bro- mide-stained DNA separated on agarose gels. Two grams of rice tissue and 2 g of fungal mycelium both typically yielded 400 pg (plus or mi- nus 50 pg) (data not shown). Total genomic DNA was then extracted from rice plants of cultivar CO-39 that had been inoculated with M. grisea strain Guy-11 at various time points during pathogenesis. The DNA was quantified, digested with restriction enzymes, fractionated by electrophoresis, blotted to nylon membranes, and probed with a single-copy M. grisea gene and then a single-copy rice gene, as de- scribed in Figure 2. DNA hybridization probes were labeled by the random primer method (Feinberg and Vogelstein, 1983) using the Stratagene Prime-lt kit. DNA gel blot hybridizations were conducted by standard methods (Sambrook et al., 1989) and washed as described previously.

A Pathogenicity Gene from Magnaporthe 1587
lntact chromosomal-sized DNA was prepared from M. grisea pro- toplasts and separated by contour clamped homogeneous electric field electrophoresis (CHEF) as described by Hamer et al. (1989). Contour clamped homogeneous electric field electrophoresis was carried out for 120 hr at 45 V with a pulse time of 90 min at 4OC. DNA was subse- quently transferred to nylon membranes as described above.
Autoradiographs were analyzed by laser densitometry using an Ultrascan II (LKB Instruments, Uppsala, Sweden) at 630 nm. Absor- bance values were integrated using the interna1 integrator function. The data were subsequently analyzed using Gelscan software (LKB- Pharmacia) on an IBM PC-AT.
cDNA and Genomic Llbrary Constructions
The blast-infected rice cDNA library was constructed in X GEM-4 (Promega) by an adaptation of the methods of Brown and Kafatos (1988) and Polites and Marrotti (1986). Poly(A)+ RNA (10 pg) was annealed to an Xbal-oligo(dT) primer adaptor. First strand cDNA synthesis was performed in modified RT buffer (0.8 mM dGTP, dATP, dTTP, 4 mM N&P207, 8 mM MgC12, 40 mM KCI, 10 mM DTT, 100 mM Tris-HCI, pH 8.7, 50 wg mL-I actinomycin D) in the presence of 0.8 mM 5-methyl dCTP (Pharmacia) and 1 pL RNasin (Promega), using 400 units MMLV- reverse transcriptase (Gibco-BRL), for 1 hr at 40OC. DNA:RNA hybrids were collected by ethanol precipitation, and second strand synthesis carried out in a reaction mixture containing the following: 100 mM Hepes, pH 7.6,6 mM MgC12, 10 mM DTT, 60 mM KCI, 0.5 mM dATP, dGTP, dTTP, 0.15 mM P-NAD, 100 pCi a-=P dCTP, 150 units DNA poly- merase I, 6 units RNaseH, 10 units E. coli DNA ligase, for 1 hr at 14OC. The DNA was phenol extracted, ethanol precipitated, and EcoRl linkers ligated to the double stranded DNA. The cDNA inserts were digested with EcoRl and Xbal and size fractionated prior to ligation to X GEM 4 and packaging, which were carried out according to manufacturer's suggestions (Promega).
Genomic DNA was isolated from purified nuclei prepared from strain Guy-11 as described by Hamer et al. (1989). The DNA was partially digested with Sau3Al and cloned into X GEMll Xhol half-site arms according to manufacturer's instructions (Promega).
cDNA Library Screenlng
Low-density screening of a total of 5000 plaque-forming units of the cDNA library was carried out. Duplicate nitrocellulose filters were pre- pared according to standard methods (Sambrook et al., 1989). Radiolabeled cDNA probes were constructed by an adaptation of the first strand cDNA synthesis protocol described above. One microgram of poly(A)+ RNA was annealed to oligo(dT) primer and reverse tran- scribed in RT buffer in the presence of 50 pCi a-32P-dCTP for 1 hr at 40OC. The probes were purified through Biogel P50 columns (Bio- Rad) constructed in 5-mL disposable pipettes. Plaque lift filters were prehybridized for 4 hr at 65OC in 6 x SSPE (1 x SSPE is 0.15 M NaCI, 10 mM sodium phosphate, 1 mM EDTA, pH 7.4). 2% SDS, 5 x Den- hardt's buffer (1 x Denhardt's solution is 0.02% Ficoll, 0.02% PVP, 0.02% BSA). Hybridizations were conducted under the same condi- tions for 20 hr. The filters were washed under high-stringency conditions (65OC in 0.1% SDS, 0.1% sodium pyrophosphate [PPi], 10 mM NaH2P04.H20, 1 mM EDTA, pH 7.4) and exposed to x-ray film (Amer- sham). Escherichia coli KW251 (Promega) was routinely used for the propagation of bacteriophage X.
Genetic Mapplng
Restriction fragment length polymorphism (RFLP) mapping was car- ried out using progeny from the 7-R cross between M. grisea strains 4136-4 and 43754-26 (Romao and Hamer, 1992). DNA was isolated from the parents and progeny, digested with Sall, and probed with pNJT-15. The segregation of an RFLP detected between the parenta1 isolates was compared with the segregation of RFLPs around MGR586 dispersed repeated elements (Hamer and Givan, 1990; Romao and Hamer, 1992).
Construction of the Gene Replacement Vector pNT400
The gene replacement vector was constructed as outlined in Figure 3. All plasmid clones were constructed in pBluescript SK+ (Stratagene). A full-length genomic clone of MPG7, X 15G-1, was selected by screening the Guy-11 genomic librarywith pNT-15. A5'and 3'proximal6.25- and 45kb BamHl fragments were subcloned tocreate pNT108 and pNT211, respectively. Xbal linkers were added to the unique EcoRV site in pNT211 and a 3.4-kb BamHI-Xbal fragment was subcloned and desig- nated pNT220. A 3' 3.4-kb Xbal fragment derived from pNT108 was then cloned into the unique Xbal site in pNT220. The resulting plas- mid contained 3.4-kb flanking regions from both 5'and 3'of the MPG7 coding region as a single 6.8-kb BamHl fragment. This was subcloned into a modified version of pBluescript SK+ in which the Xbal site had been removed and designated pNT320. The hygromycin resistance (HPH) gene was excised as a 2.6-kb Sacl-Xbal fragment from pAN7-I (Punt et al., 1987), Xbal linkers were added, and the fragment was inserted into the unique Xbal site of pNT320 to give the gene replace- ment vector pNT400. The insert of this construct is shown in Figure 3B.
E. coli strains DH5a (Gibco-BRL) and XLI-Blue (Stratagene) were routinely used for the propagation of plasmids. Recombinant DNA tech- niques were carried out according to standard protocols (Sambrook et al., 1989).
Funga1 Transformations
Transformation of M. grisea was carried out by an adaptation of the previously described methods (Parsons et al., 1987; Leung et al., 1990). Briefly, a 25 cm2 square of M. grisea strain Guy-11 mycelium was cut from the surface of an oatmeal agar plate, macerated in 50 mL of CM liquid media, and incubated overnight at 23OC on a rotary shaker at 120 rpm. The culture was blended again and incubated for a second 24-hr period. The culture was harvested by filtration and protoplasts produced by Novozym 234 digestion (Novo Nordisk, Danbury, CT) in OS buffer (1.2 M MgS04, 10 mM Na2HPOJNaH2P04, pH 5.8). Pro- toplasts were collected at the interface of OS overlayed with ST buffe,r (0.6 M sorbitol, 0.1 M Tris-HCI, pH 7.0) after centrifugation at 3750s. The protoplasts were washed in STC buffer (1.2 M sorbitol, I0 mM Tris- HCI, pH 7.5, 10 mM CaCI2). Transformation was carried out using 5 x 106 protoplasts, with 2 pg DNA, in the presence of 1 mL PTC buffer (60% polyethylene glycol 4000, 1 M sorbitol, 10 mM Tris-HCI, pH 7.5, 10 mM CaCId. The gene replacement vector pNT400 was transformed as a linear Notl fragment. Protoplasts were incubated with PTC for 5 min and then added to 150 mL of molten CM agar, osmotically'stabi- lized by addition of 1 M of sucrose, and poured to 150-mm plates. Protoplasts were allowed to regenerate for 18 hr at 23OC. At this time a selective overlay of 1% CM agar containing 200 pg mL-1 Hygromy- cin B (Boehringer Mannheim) was added, and the plates were incubated

1588 The Plant Cell
for 7 days at 3OoC in the dark. Monoconidial isolations were conducted on all hygromycin-resistant transformants.
Assays for Infection-Related Morphogenesis
Appressorial formation by M. grisea was observed on Teflon membranes (Du Pont) as previously described (Hamer et al., 1988; Howard et al., 1991a). Briefly, 200 pL of a conidial suspension at a concentration of 103 mL-I was placed on the surface of a Teflon coverslip and left in a humid chamber for 12 to 14 hr. The coverslip was then gently in- verted and viewed by phase contrast or Nomarski interference microscopy. The frequency of appressorium formation was determined from a sample size of 300 conidia per membrane. Frequencies of ap- pressorial formation were determined and tested for significant departure from the expected wild-type frequency by Chi-square tests (Sokal and Rohlf, 1981). The production of spore tip mucilage was as- sayed by fluorescein isothiocyanate-concanavalin A staining (Hamer et al., 1988). Conidial germinatíon assays were carried out on polysty- rene coverslips using conidial suspensions at a concentration of 103 mL-1. Estimations of conidial production were carried out by flood- ing the surface of plates, containing single 4-cm-diameter mycelial colonies, with 2 mL of sterile distilled water and determining conidial concentrations by haemocytometer counting.
DNA Sequencing
A 2.7-kb BamHI-Xbal fragment containing the MPGl gene was sub- cloned and designated pl5GXB. Nested deletions were generated by exonuclease 111 digestion (Henikoff, 1984) throughout the genomíc sub- clone pl5GXB and the cDNA clone pNJT-15, and DNA sequence information was obtained from both strands using the dideoxy chain termination method (Sanger et al., 1977) using Sequenase (United States Biochemical Corp.). DNA sequence analysis was performed using the Sequence analysis software, version 7.2, of the Genetics Computer Group (GCG) of the University of Wisconsin (Devereux et al., 1984) made available by the AlDS Center Laboratory for Computa- tional Biochemistry at Purdue University. The screening of DNA sequence data bases was performed using the BLAST algorithm (Altschul et al., 1990) at the NCBl or the GCG program FASTA. Amino acid alignments were made by eye and by using BESTFIT, ALIGN, and PILEl.JP programs of the GCG package. Kyte-Doolittle hydropa- thy plots (Kyte and Doolíttle, 1982) were constructed with a window size of seven, using the program DNAStrider Version 1.0 (Marck, 1988).
Primer Extensions
The 5' end of the MPGl transcript was determined by primer exten- sion using total RNA from nitrogen-starved cultures of M. grisea (see below). The oligonucleotide 5'-GCCAGGATCCACGGTCTTGAGGGA- GAACATCTTG-3' is complementary to the MPGl mRNA for 24 nucleotides starting four nucleotides upstream of the ATG initiation codon. The primer was end labeled with Y-~~P-ATP using T4 poly- nucleotide kinase according to standard protocols (Sambrook et al., 1989). Labeled primer (105 cpm) was annealed to 10 pg of RNA and coprecipitated with 0.1 volume of 3 M sodium acetate and 2.5 volume ethanol. The RNA-oligonucleotide mixture was resuspended in 20 pL of RNase-free distílled water and incubated at 65% for 5 min. Five microliters of avian myeloblastosis virus reverse transcriptase reac- tion buffer was added (final concentrations: 50 mM Tris-HCI, pH 8.3, 50 mM KCI, 10 mM MgClp, 0.5 mM spermidine, 10 mM DTT, 4 mM
PPi, and 1 mM each of dTTP, dGTP, and dCTP). The reaction mixture was moved from 65 to 42OC for 5 min and then to room temperature for 5 min to complete annealing of the primer to the MPGl mRNA. Finally, 25 units of RNasin (Promega) and 10 units of AMV reverse transcriptase (Gibco-Bethesda Research Laboratories) were added, and the reaction was incubated for 60 min at 42OC. The primer exten- sion products were analyzed by electrophoresis on an 8% (wh) urea polyacrylamide sequencing gel and visualized by autoradiography (data not shown). DNA sequencing ladders were used as size standards to estimate the size of the reverse transcribed products.
ACKNOWLEDGMENTS
We are grateful to Larry Dunkle, Mark Young, and Sally MacKenzie for critical reading of the manuscript and members of the Hamer lab for useful discussions and suggestions. We are also grateful to Jos Wessels (University of Groningen), Mary Stringer and Bill Timberlake (University of Georgia), and Raymond St. Leger (Boyce Thompson) who communicated results prior to publication and gave us invalua- ble insights on the function of hydrophobins. We are also indebted to Mary Stringer who helped us with the sequence alignments, and Jeni Keenan and David McGee for technical assistance. D.J.E. is sup- ported by the Texas Agricultura1 Experiment Station and by a grant from the Office of University Research, Texas A&M University. This work was supported by grants to J.E.H. from the Biotechnology Re- search and Development Consortium, the National Science Foundation, and the David and Lucille Packard Foundation.
Received June 15, 1993; accepted September 15, 1993.
REFERENCES
Altschul, S.F., Gish, W., Miller, W., Myen, C.W., and Lipman, D.L. (1990). Basic local alignment search tool. J. MOI. Biol. 215,403-410.
Ballance, D.J. (1986). Sequences important for gene expression in filamentous fungi. Yeast 2, 229-236.
Bell-Pedenon, D., Dunlap, J.C., and Loros, J.J. (1992). The Neu- rospora circadian clock-controlled gene, ccg-2, is allelic to eas and encodes a fungal hydrophobin requíred for formation of the conidial rodlet layer. Genes Dev. 6, 2382-2394.
Bolyard, M.J., and Sticklen, M.B. (1992). Expression of a modified Dutch elm disease toxin in Escherichia coli. MOI. Plant-Microbe In- teract. 5, 520-524.
Bourett, T. M., and Howard, R.J. (1990). In vitro development of penetration structures in the rice blast fungus Magnaporthe grisea. Can. J. Bot. 68, 329-342.
Bourett, T.M., and Howard, R.J. (1992). Actin in penetration pegs of the fungal rice blast pathogen, Magnaporfhe grisea. Protoplasma
Bowles, D.J. (1990). Defense related proteins in higher plants. Annu.
Bmwn, N.H., and Kafatos, F.C. (1988). Functional cDNA libraries from
Chasan, R. (1991). Building hyphae in the air. Plant Cell3, 1048-1050.
168, 20-26.
Rev. Biochem. 59, 873-907.
Drosophila embryos. J. MOI. Biol. 203, 425-437.

A Pathogenicity Gene from Magnaporthe 1589
Crawford, M.S., Chumley, F.G., Weaver, C.G., andValent, B. (1986). Characterization of the heterokaryotic and vegetative diploid phases of Magnaporthe grisea. Genetics 114, 1111-1129.
Dellaporta, S., Wood, J., and Hicks, J.B. (1983). A plant DNA minipreparation: Version II. Plant MOI. Biol. Rep. 1, 19-21.
Devereux, J., Haeberli, P., and Smithies, O. (1984). A comprehen- sive set of sequence analysis programs of the VAX. Nucl. Acids Res. 12, 387-395.
deVries, O.M.H., Fekkes, M.P., Wosten, H.A.B., and Wessels, J.G.H. (1993). lnsoluble hydrophobin complexes in the walls of Schizophyl- /um commune and other filamentous fungi. Arch. Microbiol. 159,
Drenth, J., Low, B. W., Richardson, J.S., and Wright, C.S. (1980). The toxin-agglutinin fold: A new group of small protein structures organized around a four disulfide core. J. Biol. Chem. 255,
Feinberg, A.P., and Vogelsteln, B. (1983). A technique for radiolabeling DNA restriction endonuclease fragments to high specific activity. Anal. Biochem. 132, 6-13.
Hamer, J.E., and Givan, S. (1990). Genetic mapping with dispersed repeated sequences in the rice blast fungus: Mapping the SMO 10- cus. MOI. Gen. Genet. 223, 487-495.
Hamer, J. E., Howard, R.J., Chumley, F.G., and Valent, B. (1988). A mechanism for surface attachment in spores of a plant patho- genic fungus. Science 239, 288-290.
Hamer, J.E., Farrall, L., Orbach, M.J., Valent, B., and Chumley, F.G. (1989). Host species-specific conservation of a family of repeated DNA sequences in the genome of a fungal plant pathogen. Proc. Natl. Acad. Sci. USA 86, 9981-9985.
Hardham, A. R. (1992). Cell biology of pathogenesis. Annu. Rev. Plant Physiol. Plant MOI. Biol. 43, 491-526.
Heath, M.C, Valent, B., Howard, R.J., and Chumley, F. (199Oa). Corre- lations between cytologically detected plant-funga1 interactions and pathogenicity of Magnaporthe grisea toward weeping lovegrass. Phytopathology 80, 1382-1386.
Heath, M.C., Valent, B., Howard, R.J., and Chumley, F.G. (1990b). lnteractions of two strains of Magnaporthe grisea with rice, goosegrass and weeping lovegrass. Can. J. Bot. 68, 1627-1637.
Heijne, G.V. (1983). Patterns of amino acids near signal sequence cleav- age sites. Eur. J. Biochem. 133, 17-21.
Henikoff, S. (1984). Unidirectional digestion with exonuclease 111 cre- ates targeted breakpoints for DNA sequencing. Gene 28,351-359.
Howard, R.J., and Ferrari, M.A. (1989). Role of melanin in appresso- rium function. Exp. Mycol. 13, 403-418.
Howard, R.J., Bourett, T.M., and Ferrari, M.A. (1991a). lnfection by Magnaporthe grisea: An in vitro analysis. In Electron Microscopy of Plant Pathogens, K. Mendgen and D.E. Lesemann, eds (Berlin: Springer-Verlag), pp. 251-264.
Howard, R.J., Ferrari, M.A., Roach, D.H., and Money, N.P. (1991b). Penetration of hard substances by a fungus employing enormous turgor pressures. Proc. Natl. Acad. Sci. USA 88, 11281-11284.
Kyte, J., and Doolittle, R.F. (1982). A simple methcd for displaying the hydropathy character of a protein. J. MOI. Biol. 157, 105-132.
Lauter, F.-R., Russo, V.E.A., and Yanofsky, C. (1992). Developmen- tal and light regulation of €as, the structural gene for the rodlet protein of Neurospora crassa. Genes Dev. 6, 2373-2381.
Lebrun, M.-H., Dutfoy, F., Gaudemer, F., Kunesch, G., and Gaudemer, A. (1990). Detection and quantification of the fungal
330-335.
2e52-2655.
phytotoxin tenuazonic acid produced by Pyricularia bfyzae. Phyiochemistry 29, 3777-3783.
Leung, H., Lehtinen, U., Karjalainen, R., Skinner, D., Tooley, P., Leong, S.A., and Ellingboe, A. (1990). Transformation of the rice blast fungus Magnaporthe grisea to hygromycin B resistance. Curr. Genet. 17, 409-411.
Marck, C. (1988). 'DNA strider': A 'C' program for the fast analysis of" DNA and protein sequences for the Apple Macintosh family of com- puters. Nucl. Acids Res. 16, 1829-1836.
Mendgen, K., and Deising, H. (1993). lnfection structures of fungal' plant pathogens: A cytological and physiological evaluation. New
Ou, S.H. (1985). Rice Diseases (Surrey: Commonwealth Mycological Institute).
Pandit, N.N., and Russo, V.E.A. (1991). The bli regulon: A network of blue light inducible genes of Neurcspora crassa. Funga1 Genet. Newslett. 38, 93-95.
Parsons, K.A., Chumley, F.G., and Valent, 8. (1987). Genetic trans- formation of the fungal pathogen responsible for rice blast disease. Proc. Natl. Acad. Sci. USA 84, 4161-4165.
Peng, Y.-L., and Shishiyama, J. (1988). Temporal sequences of cyto- logical events in rice leaves infected with Pyricularia ofyzae. Can.
Polites, H.G., and Marrotti, K.R. (1986). Astep-wise protocol for cDNA synthesis. BioTechniques 4, 514-520.
Punt, P.J., Oliver, R.P., Dingemanse, M.A., Pouwels, P.H., and van den Hondel, C.A.M.J.J. (1987). Transformation of Aspergillus based on the hygromycin B resistance marker from Escherichia coli. Gene 56, 117-124.
Romao, J., and Hamer, J.E. (1992). Genetic organization of a repeated DNA sequence family in the rice blast fungus. Proc. Natl. Acad. Sci.
Rothstein, R.J. (1983). One-step Gene Disruption in Yeast (New York: Academic Press, Inc.).
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989). Molecular Clon- ing: A Laboratory Manual (Cold Spring Harbor, NY Cold Spring Harbor Laboratory Press).
Sanger, F., Nicklen, S., and Coulson, A.R. (1977). DNAsequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74,
Sokal, R.R., and Rohlf, F.J. (1981). Biometry (San Francisco, CA: W.H. Freeman and Company).
St. Leger, R.J., Staples, R.C., and Roberts, D.W. (1992). Cloning and regulatory analysis of the starvation-stress gene ssgA, encoding a hydrophobin-like protein from the entomopathogenic fungus Metarhizium anisopliae. Gene 120, 119-124.
Stringer, M.A., and Timberlake, W.E. (1993). Cerato-ulmin, a toxin involved in Dutch elm disease, is a fungal hydrophobin. Plant Cell
Stringer, M.A., Dean, R.A., Sewell, T.C., and Timberlak, W.E. (1991). Rodletless, a new Aspergillus developmental mutant induced by directed gene inactivation. Genes Dev. 5, 1161-1171.
Sweigard, J.A., Chumley, F.G., and Valent, B. (1992a). Cloning and analysis of CUT7, a cutinase gene from Magnaporthe grisea. MOI. Gen. Genet. 232, 174-182.
Sweigard, J.A., Chumley, F.G., and Valent, 8. (1992b). Disruption of a Magnaporthe grisea cutinase gene. MOI. Gen. Genet. 232,
Phytol. 124, 193-213.
J. BOt. 66, 730-735.
USA 89, 5316-5320.
5463-5467.
5, 145-146.
183-190.

1590 The Plant Cell
Talbot, N.J., Salch, Y., Ma, M., and Hamer, J.E. (1993). Karyotype variation within clonal lineages of the rice blast fungus Magnaporthe grisea. Appl. Environ. Microbiol. 59, 585-593.
Tlmberlake, W.E. (1980). Developmental gene regulation in Asper- gillus nidulans. Dev. Biol. 78, 497-510.
Valent, B., and Chumley, F.G. (1991). Molecular genetic analysis of the rice blast fungus, Magnaporthe grisea. Annu. Rev. Phytopathol.
Valent, B., Farrall, L., and Chumley, F.G. (1991). Magnaporthe grisea genes for pathogenicity and virulence identified through a series of backcrosses. Genetics 127, 87-101.
29, 443-467.
Wessels, J.G.H. (1992). Gene expression during fruiting in Schizophyl- /um commune. Mycol. Res. 96, 609-620.
Wessels, J.G.H., de Vries, O.M.H., Asgeirsd6ttir, S.A., and Schuren, F.H.J. (1991a). Hydrophobin genes involved in forrnation of aerial hyphae and truit bodies in Schizophyllum. Plant Cell 3, 793-799.
Wessels, J.G.H., de Vries, O.M.H., Asgeirsdbttir, S.A., and Springer, J. (1991b). The fhn mutation of Schizophyllum commune which sup- presses formation of aerial hyphae, affects expression of the Sc3 hydrophobin gene. J. Gen. Microbiol. 137, 2439-2445.
Wbsten, H.A.B., de Vrles, O.M.H., and Wessels, J.G.H. (1993). In- terfacial self-assembly of a funga1 hydrophobin into a hydrophobic rodlet layer. Plant Cell, 5, 1567-1574.

DOI 10.1105/tpc.5.11.1575 1993;5;1575-1590Plant Cell
N J Talbot, D J Ebbole and J E Hamerfungus Magnaporthe grisea.
Identification and characterization of MPG1, a gene involved in pathogenicity from the rice blast
This information is current as of May 18, 2018
Permissions X
https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists