j. gen. appl. microbiol., 56(1): 19-29(2010)
TRANSCRIPT

Introduction
Microbial fuel cells (MFCs) can utilize special micro-organisms to oxidize organics in wastewater, while si-multaneously generating electricity. Thus, the MFC is potentially a very innovative energy-generating device, and is becoming a research focus in both the environ-mental and energy fi elds (Bullen et al., 2006; Holmes et al., 2004a; Kim et al., 2002; Logan and Regan, 2006).
Mediator-less MFCs are considered to have more commercial applications, with potentially lower cost and lower toxicity than biofuel cells with mediators (Bond and Lovley, 2003). While certain bacteria such as Shewanella putrefaciens can transfer electrons to electrodes, electrochemically active redox enzymes always play an important role in mediator-less MFCs (Kim et al., 2002; Leang et al., 2003). The microorgan-isms on the surface of the anode in MFCs are key to the reduction of chemical oxygen demand (COD) as well as electricity generation (Bond and Lovley, 2003; Lee et al., 2003; Liu et al., 2004; Reimers et al., 2001; Sokhee and John, 2007). Presently, this topic is expe-
J. Gen. Appl. Microbiol., 56, 19‒29 (2010)
Single-chamber microbial fuel cells (MFCs), inoculated with anaerobic sludge and continuously run with two kinds of organic wastewater infl uents, were systemically investigated. The diversity of microbes, determined by 16S rDNA analysis, was analyzed on three anodes under different conditions. One anode was in a closed circuit in synthetic wastewater containing glucose. The other two anodes, in open or closed circuits, were fed effl uent from an anaerobic reactor treating starch wastewater. The chemical oxygen demand (COD) removal effi ciency was about 70%, and the exported voltages were about 450 mV. The 16S rDNA molecular clones of microbes on anode surfaces showed signifi cant changes in Eubacterial structure under different conditions. γ-Proteobacteria and the high G+C gram-positive groups were predominant in the synthetic wastewater, while ε-Proteobacteria predominated in the anaerobic reactor effl uent. Known exo-electrogenic bacterial species composition also changed greatly depending on substrate. On the artifi cial substrate, 28% of the bacterial sequences were affi liated with Aeromonas, Pseudomo-nas, Geobacter, and Desulfobulbus. On the anaerobic effl uent, only 6% were affi liated with Geobacter or Clostridium. Because only a few exoelectrogenic bacteria from MFCs have been directly isolated and studied, we compared the community structures of two bacterial anodes, in open and closed circuits, under the same substrate of anaerobic effl uent in order to identify ad-ditional exoelectrogenic bacterial strains. Alcaligenes monasteriensis, Comamonas denitrifi cans, and Dechloromonas sp. were found to be potential exoelectrogenic bacteria worthy of further research.
Key Words—anode; community structure; exoelectrogenic bacteria; microbial fuel cell (MFC)
* Address reprint requests to: Dr. Yujiao Sun, College of Wa-ter Sciences, Beijing Normal University, Beijing 100875, China. E-mail: [email protected]
Full Paper
Diversity of microbes and potential exoelectrogenic bacteria on anode surface in microbial fuel cells
Yujiao Sun,1,2,* Jiane Zuo,2 Longtao Cui,2 Qian Deng,2 and Yan Dang2
1 College of Water Sciences, Beijing Normal University, Beijing 100875, China2 Department of Environmental Science and Engineering, Tsinghua University, Beijing, 100084, China
(Received June 12, 2009; Accepted September 12, 2009)

20 Vol. 56SUN et al.
riencing much research activity (Kim et al., 2004; Nguyet et al., 2004). Bacterial communities and exoelectrogenic bacteria have been studied in MFCs. Different substrates great-ly affect which bacterial group will predominate. In general, δ-Proteobacteria were found enriched on cur-rent-generating anodes of MFCs inoculated with a va-riety of sediments rich in acetate (Holmes et al., 2004b) β-Proteobacteria were found to predominate on the anodes of ethanol-fed MFCs (Kim et al., 2007). γ-Proteobacteria predominated in cysteine-fed MFCs (Logan et al., 2005), while glucose and glutamate yielded diverse bacterial classes that varied with sub-strate concentration (Choo et al., 2006; Kim et al., 2006). Mixed-culture MFCs have generated higher power densities than have their pure-culture counter-parts, perhaps due to synergistic interaction within the complex anode communities and mechanisms. Some bacteria might also be participating in non-relevant metabolisms, such as fermentation. At this point in time, very few exoelectrogenic bacte-ria have been directly isolated from MFCs. The failure to detect known exoelectrogens in some systems sug-gests that there is a greater diversity of this phenotype than is presently understood. Several phylogenetically diverse bacteria are known to generate electricity in MFCs without the provision of an exogenous mediator. These include Acidobacteria (Geothrix) (Bond and Lovley, 2005), Firmicutes (Clostridium) (Park et al., 2001), α-Proteobacteria (Ochrobactrum) (Zuo et al., 2008), β-Proteobacteria (Rhodoferax) (Chaudhuri and Lovley, 2003), γ-Proteobacteria (Pseudomonas and Shewanella) (Kim et al., 1999, 2002; Rabaey et al., 2004) and δ-Proteobacteria (Aeromonas, Geobacter, Geopsychrobacter, Desulfuromonas and Desulfobul-
bus) (Bond et al., 2002; Bond and Lovley, 2003; Hol-mes et al., 2004b, c; Pham et al., 2003). Some researchers have reported that some bacteria in the family of Geobacteraceae are able transfer elec-trons to extracellular electron acceptors such as Fe3+ oxides as organic chemicals are degraded (Kaufmann and Lovley, 2001; Magnuson et al., 2000). Reguera reported that pili of Geobacter served as biological nanowires, transferring electrons from the cell surface to the surface of Fe3+(Reguera et al., 2005). The mechanisms used for exocellular transport of elec-trons have been demonstrated, in which cell-bound outer membrane cytochromes and conductive nano-wires might play key roles in electron transfer for some
Geobacter and Shewanella species (Gorby et al., 2006; Lovley et al., 2004; Myers and Myers, 1992; Reguera et al., 2005). Alternatively, some exoelectrogens, such as Pseudomonas aeruginosa and Geothrix fermentans, excrete mediators in order to shuttle electrons to sur-faces (Bond and Lovley, 2005; Rabaey et al., 2004). In the present research, we have used a molecular cloning technique to document the diversity of anode bacterial communities. The quantities of exoelectro-genic bacteria exposed to different mixing infl uents were also studied by observing the resulting electro-generated effects. The relationship between the elec-trochemically active microbes and the other nonexo-electrogenic microbes were analyzed. Comparison and analysis of microbes in open and closed anodes using the same MFCs can help us to identify other po-tentially exoelectrogenic bacteria.
Materials and Methods
Structure and system components of MFCs. The MFC discussed in this paper was made of poly(methyl methacrylate) (PMMA) and consisted of a cubic box with a total volume about 1.1 L. The anode was made of carbon fabric and divided into 5 pieces. The super-fi cial area dimensions were 50 mm×50 mm. The cath-ode was also made of carbon fabric, but contained platinum (1 mg/cm2) as the catalyst. Its dimensions were also 50 mm×50 mm. The outside of the cathode was exposed to the air, the inner side was pressed and connected tightly with the proton exchange membrane (PEM), and the other side of the PEM directly contact-ed the solution of the reactor. The distance between anode and cathode is 1.5 cm, the same as the dis-tance between anodes. The anode and cathode were connected to the resistance (1,000 Ω) in the circuit through platinum and copper wires, respectively. A multimeter (UT803 digital multimeter; Youlide Science and Technology Co., Ltd., Hong Kong) was used to automatically measure the exported voltage. The elec-trodes used were 218 Ag/AgCl (Luosu Science and Technology Co., Ltd., Shanghai). The structure and system components of the MFC are shown in Fig. 1. Infl uent wastewater was pumped into the MFC reactor at a fl ow rate of 0.3‒1.0 L/d. The Hydraulic Retention Time (HRT) was about 1‒3 days. The MFC was incu-bated at 32°C. The pH (TPX-90i pH meter) in the reac-tor was kept at about 7.0 by adding NaHCO3 solution to the infl uent.

2010 21Diversity of microbes and potential exoelectrogens in MFC
Seed sludge. The seed sludge was taken from an anaerobic reactor that was treating starch-processing wastewater. Before inoculating the MFC, the sludge was pre-treated as follows: (1) Thirty ml raw anaerobic sludge was put into a mortar and ground with a pestle. (2) Twenty ml oxygen-free water was added and the mixture was moved into a conical fl ask and fl ushed with high-purity nitrogen gas for 1 h. (3) The suspen-sion was then used to inoculate the MFC reactor. Wastewater types and characteristics. There were two different kinds of wastewater used as the infl uent for the MFC in this study. One was synthetic, prepared fresh by dissolving glucose in tap water until the COD concentration was about 300 mg/L. To 1 L of this solu-tion were added 50 ml of a phosphate buffer solution (Na2HPO4・12H2O (177.99 g/L), KH2PO4 (67.8 g/L), and 10 ml of a micro-nutrients solution (MgSO4 (10 g/L), CaCl2 (1.5 g/L), FeCl3・6H2O (0.1 g/L), MnSO4・H2O (2 g/L), NH4CO3 (67 g/L)). The other infl uent was the actual effl uent from an anaerobic reactor that was be-ing used to treat starch-processing wastewater. Its COD concentration was about 300‒400 mg/L, and we added the same micro-nutrient solution prior to use in the MFC. The second infl uent was designated ‘anaer-obic effl uent’. The COD level and sludge concentra-tion were monitored following standard methods (Fusheng et al., 2002). Sludge samples. The MFC was operated in syn-thetic wastewater for 50 days, and then in anaerobic effl uent for 20 days, consecutively. Three sludge sam-ples from the anode surface were taken by knife scraped from the MFC after it had been operating for 15 days in each liquid, and the anode installed was not reduced as the cultivation time elapsed. Sample A rep-resented the sludge on the surface of an anode in a closed circuit, with synthetic wastewater. Sample B represented sludge on an anode in a closed circuit in
anaerobic effl uent, and sample C represented sludge on an anode in an open circuit, in anaerobic effl uent. These different conditions are listed in Table 1. DNA extraction and PCR amplifi cation of Eubacteria with 16S rDNA. Sludge samples were scraped off 10 cm2 of each of the different anodes. The bacteria living on the surface of the anodes were scraped off with a sharp, sterilized knife. The bead beating method (Yeates et al., 1998) was applied to break the cell wall of the sludge samples. All chromosomal DNA was ex-tracted using a DP301 DNA extraction kit (Tiangen Bio-technology, China), and then was purifi ed with an A140-2 DNA purifi cation kit (Dingguo Biotechnology, China). Using the purifi ed DNA extracts as templates, the 16S rDNA fragments of the Eubacteria were ampli-fi ed using primers of P8 (5′-AGA GTT TGA TCC TGG CTC AG-3′) (Liu et al., 1997) and P1492 (5′-GGC TAC CTT GTT ACG ACT T-3′), yielding a fragment of about 1,500 bp (Lane, 1991). One percent agarose gel elec-trophoresis was used to separate the PCR products, and the bands were reclaimed and purifi ed with an A140-2 DNA purifi cation kit (Dingguo Biotechnology) before being ligated into vectors. Construction of 16S rDNA clone library and phyloge-
netic analysis. The purifi ed 16S rDNA amplifi cation fragments were ligated into a pGEM-T vector system (Promega Company) before they were transformed into the competent E. coli DH5α cells. The transformed cells were spread on Luria-Bertani plates containing 100 μg/ml of ampicillin, 80 μg/ml of X-GAL, and 0.5 mM of IPTG. The plates were incubated at 37°C for 18 h. The white clones were chosen as the possible positive clones (Sambrook et al., 1989). Positive clones of each sample were randomly selected for further analysis. Plasmid DNA extraction and PCR amplifi cation were conducted with these possible positive clones. Plasmids were analyzed for restriction fragment length polymorphism (RFLP) using enzyme pairs (RsaI and MspI) to select different clones (Weisburg et al., 1991). The 16S rDNA fragments of selected clones were sequenced by an ABI sequencer (Sanbo Bio-
Fig. 1. Schematic chart of MFC structure and its system components.
Table 1. Operational conditions of MFC while sampling the three sludge samples.
Sample Infl uent Anode status
A Synthetic wastewater Connected in closed circuitB Anaerobic effl uent Connected in closed circuitC Anaerobic effl uent Connected in open circuit

22 Vol. 56SUN et al.
technology Company, Beijing, China). All different se-quences were believed to be operational taxonomic units (OTUs). The sequences were compared with all accessible sequences in the databases using the BLAST server at NCBI (National Centre of Biotechnol-ogy Information). The sequences were aligned with those belonging to representative organisms of the domain bacteria. The phylogenetic trees were con-structed by the neighbor-joining method using the Mo-lecular Evolecular Evolutionary Genetics Analysis package (MEGA, version 3) (Kimura, 1980; Kumar et al., 2004). The stability of the phylogenetic trees was analyzed by Bootstrap. The sequences obtained in this study are available in GenBank under accession numbers GQ390367‒ GQ390427.
Results
Startup and performance of MFCs with synthetic waste-
water
After inoculation with the pre-treated anaerobic sludge, the MFC was started using synthetic wastewa-ter as the infl uent, and operated for a total of 50 days. The performance results are shown in Fig. 2 and Fig. 3. At the time the MFC was started, the infl uent COD level was about 300 mg/L and the fl ow rate was about 250 ml/d. The HRT of the infl uent in the MFCs was about 4 days, and the loading rate was about 0.07 kg COD/m3 d. The voltage over the resistance increased gradually, and on day 18, the highest exported volt-age, that of 530 mV, was observed. Calculations indi-cated that, at this moment, the power density was 112.36 mW/m2 (253.06 mW/m3). The COD removal ef-fi ciency was about 60%, and the effl uent COD was about 100 mg/L. By day 20, the operation and perfor-
mance of the MFC were quite stable, and the starting period was over. Subsequently, the MFC operated stably for more than 30 days, and the infl uent loading rate was in-creased stepwise, primarily by increasing the infl uent fl ow rate. In the last period of operation, the infl uent fl ow rate was increased to about 1,000 ml/d, and the loading rate was 0.27 kg COD/m3 d. The exported volt-ages were kept stable at 430‒440 mV, and the average power density was about 77.43 mW/m2 (193.56 mW/m3). The COD removal effi ciency was about 70%, and the effl uent COD was 90 mg/L. In the stable operation phase with synthetic wastewater, the coulombic effi -ciency was about 1.5.
Performance of MFCs with anaerobic effl uent
The performance results of the MFCs are shown in Fig. 4 and Fig. 5. After infl uent changing, the anaero-bic effl uent was diluted with tap water for the fi rst 3 days. The COD level was only about 90 mg/L, the infl u-ent fl ow rate was kept at about 500 ml/d, and the aver-age export voltage was about 393 mV. After the sixth day, the infl uent was not diluted further, so the infl uent COD was about 300 mg/L. Therefore, the loading rate
Fig. 2. Loading rates and exported voltages of MFC fed by synthetic wastewater.
Fig. 3. COD removal in MFC fed by synthetic wastewater.
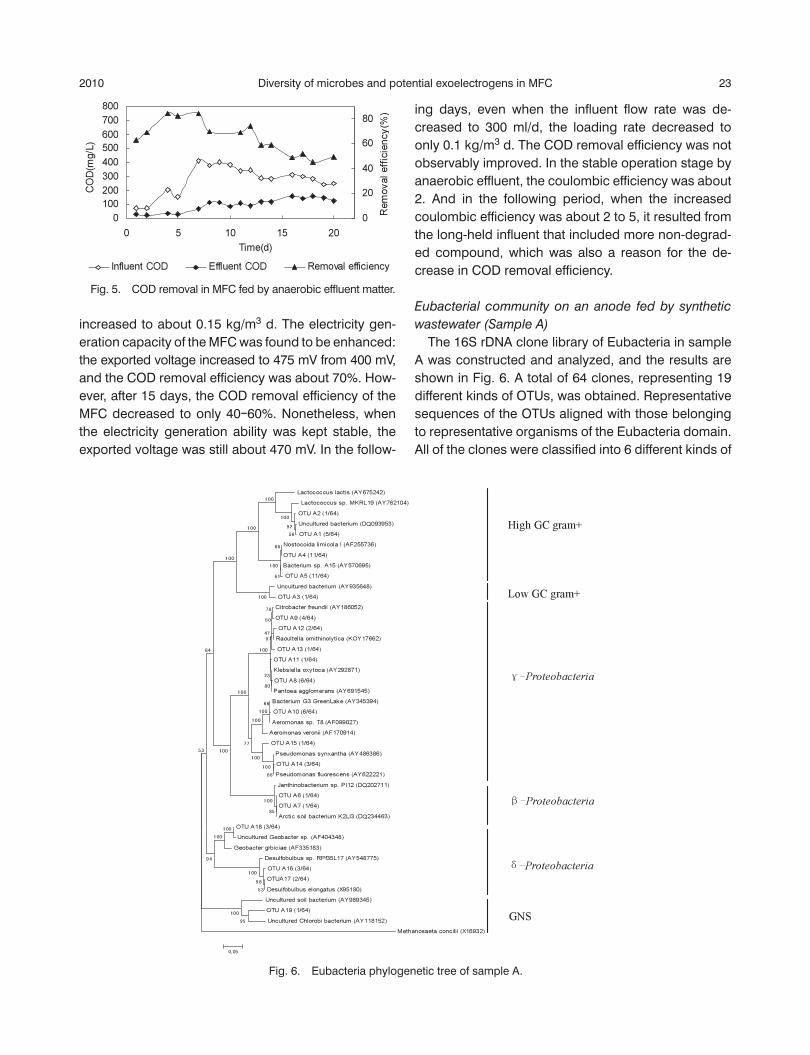
Fig. 4. Loading rates and exported voltages of MFC fed by anaerobic effl uent matter.
2010 23Diversity of microbes and potential exoelectrogens in MFC
increased to about 0.15 kg/m3 d. The electricity gen-eration capacity of the MFC was found to be enhanced: the exported voltage increased to 475 mV from 400 mV, and the COD removal effi ciency was about 70%. How-ever, after 15 days, the COD removal effi ciency of the MFC decreased to only 40‒60%. Nonetheless, when the electricity generation ability was kept stable, the exported voltage was still about 470 mV. In the follow-
ing days, even when the infl uent fl ow rate was de-creased to 300 ml/d, the loading rate decreased to only 0.1 kg/m3 d. The COD removal effi ciency was not observably improved. In the stable operation stage by anaerobic effl uent, the coulombic effi ciency was about 2. And in the following period, when the increased coulombic effi ciency was about 2 to 5, it resulted from the long-held infl uent that included more non-degrad-ed compound, which was also a reason for the de-crease in COD removal effi ciency.
Eubacterial community on an anode fed by synthetic
wastewater (Sample A)
The 16S rDNA clone library of Eubacteria in sample A was constructed and analyzed, and the results are shown in Fig. 6. A total of 64 clones, representing 19 different kinds of OTUs, was obtained. Representative sequences of the OTUs aligned with those belonging to representative organisms of the Eubacteria domain. All of the clones were classifi ed into 6 different kinds of
Fig. 5. COD removal in MFC fed by anaerobic effl uent matter.
Fig. 6. Eubacteria phylogenetic tree of sample A.

24 Vol. 56SUN et al.
Eubacteria: high-GC gram-positive bacteria, low-GC gram-positive bacteria, β-Proteobacteria, γ-Proteobacteria, δ-Proteobacteria, and green nonsulfur bacteria. Among these, the high-GC gram-positive bacteria and the γ-Proteobacteria were the dominant groups, and were present in the respective proportions of 34.38% and 37.5%. Among the high-GC gram-positive bacteria, OTU A4 and OTU A5 were the dominant species. These were related, respectively, to Nostocoida limicola I (AF255736) and Bacterium sp. (AY 570695) (with a >98% se-quence similarity), which belong to Actinobacteria. OTU A8 and OTU A9 were the predominant species among the γ-Proteobacteria, and were related closely to the Enterococcaceae family bacteria Citrobacter fre-
undii and Klebsiella oxytoca, respectively. The phylo-genetic tree for sample A was constructed, and is shown in Fig. 6. Among the exoelectrogenic bacteria,
there were 28% clone sequences (OTU 10, OTU 14, OTU 15, OTU 16, OTU 17, and OTU 18) that were af-fi liated with the known exoelectrogenic bacteria in-cluding Aeromonas (Pham et al., 2003), Pseudomonas (Rabaey et al., 2004), Geobacter (Holmes et al., 2004c), and Desulfobulbus (Holmes et al., 2004b).
Eubacterial community on an anode fed by an anaero-
bic effl uent (Sample B)
For sample B, a total of 64 clones representing 25 different OTUs were obtained. All of these OTUs had been blasted and aligned with those belonging to rep-resentative organisms of the domain Eubacteria, with a sequence similarity of more than 98%. All of the clones were classifi ed into 8 different Eubacterial groups, including low-GC gram-positive bacteria, Spi-
rochaetes, SRI, OPII, CFB-Bacteroidetes, β-Proteobacteria, δ-Proteobacteria, and ε-Proteobacteria, as shown in
Fig. 7. Eubacteria phylogenetic tree of sample B.

2010 25Diversity of microbes and potential exoelectrogens in MFC
Fig. 7. Compared with the results of sample A, after the infl uent was changed, the Eubacterial diversity of sam-ple B changed substantially. The ε-Proteobacteria and low-GC gram-positive bacteria clearly became the dominant groups, as each one accounted for 57.81% and 17.19% of the content, respectively. The predomi-nant species were very closely related to an uncultured ε-Proteobacteria (AF458288), and to Arcobacter but-
zleri (AY621116). The predominant species in the low-GC gram-positive bacterial group were mainly related to some acid-producing bacteria, such as Trichococ-
cus, Streptococcus, and Clostridium. The phylogenet-ic tree for sample B was constructed, and is shown in Fig. 7. According to the reported exoelectrogenic bac-teria, 6% of the clone sequences (OTU B20, OTU B21, and OTU B4) were affi liated with reported exoelectro-genic bacteria of Geobacter (Holmes et al., 2004c) and Clostridium (Park et al., 2001).
Eubacterial community on an anode in an anaerobic
effl uent (Sample C)
For sample C, a total of 66 clones representing 17 different kinds of OTUs were obtained. All the OTUs
had been blasted and aligned with those belonging to representative organisms of the domain Eubacteria, with a sequence similarity of more than 98%. All clones could be classifi ed into 8 different Eubacteria groups: low-GC gram-positive bacteria, Bacteroidetes, SRI, green nonsulfur bacteria (GNS), Chlamydiae, β-Proteobacteria, δ-Proteobacteria, and ε-Proteobacteria. These are shown in Fig. 8. Even though sample C was taken from the anode that did not connect into a circuit, the Eubacterial community structure on it was nearly the same as that from sample B. The ε-Proteobacteria types from sample C were still the predominant groups, but their proportions were even as high as 68.18%. The most dominant species in the ε-Proteobacteria group was very closely related to an uncultured ε-Proteobacteria (AF458288), and to Arcobacter but-
zleri (AY621116), the same result as found with sample B. The phylogenetic tree for sample C was constructed, and is shown in Fig. 8. According to the reported exo-electrogenic bacteria, 1% of the clone sequences (OTU 12) were affi liated with the reported exoelectro-genic bacteria Geobacter (Holmes et al., 2004c) which occasionally appeared on the open anode.
Fig. 8. Eubacteria phylogenetic tree of sample C.

26 Vol. 56SUN et al.
Discussion
In this study, seeded with pre-treated anaerobic sludge, an MFC was operated using either synthetic glucose wastewater or the effl uent from an anaerobic reactor that was treating starch-processing wastewa-ter successfully. Upon consecutive testing using these media, each infl uent’s COD was decreased effi ciently, and electricity was simultaneously generated. The contents of the different main species in the three samples are summarized in Table 2. Comparing sample A and sample B, there were very obvious dif-ferences in Eubacterial diversity between these two anodes. The dominant species were completely differ-ent from each other, due to different organic chemicals contained in the different infl uents. In sample A, in which glucose was the main carbon source, the domi-nant species were a number of high-GC gram-positive bacteria affi liated with Nostocoida limicola, which has a function similar to fl oc-forming bacteria that decrease Biochemical Oxygen Demand (BOD) quite well. γ-Proteobacteria were predominantly affi liated with both Citrobacter freundii and Klebsiella oxytoca, which can degrade organic chemicals, ferment glu-cose to fatty acids and produce CO2 gas. This could have reduced the nitrate levels (Holt et al., 1994). In sample B, whose infl uent contained many complicat-ed organic chemicals, the dominant clone sequences were affi liated with ε-Proteobacteria, such as Arco-
bacter butzleri and uncultured ε-Pro teobacteria, which made it similar to sample C. The infl uent content in sample B suggested that the bacteria in it is less likely to degrade sugar, but rather gets its energy from the metabolism of small molecular organic acids. The
dominant species on the anode surface was probably not the actual electrochemically active bacteria. In fact, most of the above-mentioned dominant species in samples A and B were fermentative and acidogenic bacteria. Their main functions were preliminarily to de-grade organic chemicals in the infl uent, which in turn provided suitable substrates for the actual electro-chemically active bacteria. The latter were possibly not the dominant species on the anode surface. Regarding the exoelectrogenic bacteria, as seen in Fig. 6, there were 28% clone sequences (OTU 10, OTU 14, OTU 15, OTU 16, OTU 17, and OTU 18) affi liated with the reported exoelectrogenic bacteria Aeromo-
nas, Pseudomonas, Geobacter, and Desulfobulbus (Holmes et al., 2004b, c; Pham et al., 2003; Rabaey et al., 2004) in sample A. Because most of the exoelec-trogenic bacteria studied here were fed by artifi cial substrates, more exoelectrogenic bacteria appeared on substrate A than on substrate B. As seen in Fig. 7, only 6% of some clone sequences (OTU 20 and OTU 21) were affi liated with the reported exoelectrogenic bacteria Geobacter and Clostridium (Holmes et al., 2004c; Park et al., 2001) in sample B. There might be other exoelectrogens in this system that are not yet characterized. Figure 2‒Fig. 5 show that the COD re-moval effi ciency and the exported voltage were similar in the two MFC systems in infl uents A and B, while the result was somewhat better in the latter system. Thus, we can presume that there are two possible reasons for the exoelectrogens in sample B. One is that the small numbers of primary exoelectrogenic bacteria that were closely affi liated with Geobacteria grbiciae were especially active, with a high exoelectrogenic ability. However, the Geobacteria grbiciae also ap-peared in MFCs on substrate A. Therefore, the other presumption is more reasonable, which is that there were other exoelectrogenic bacteria in the MFC sys-tem on substrate B that are new. Because very few exoelectrogenic bacteria have been directly isolated from MFCs and studied, we compared the community structures of two anode bacteria (sample B and sample C) in open and closed circuits on the same substrate of anaerobic effl uent, in order to detect more potentially exoelectrogenic bac-teria in the MFCs. The dominant species were similar in both samples B and C, and they were affi liated with both Arcobacter butzleri and with an uncultured ε-Proteobacteria. These were fermentative and aci-dogenic bacteria, which could not use glucose, and
Table 2. Contents of different main species in three samples.
Microbial classifi cationSample A
(%)Sample B
(%)Sample C
(%)
High-GC gram-positive 34.38 ― ―Low-GC gram-positive 1.56 17.19 6.06β-Proteobacteria 3.13 6.25 6.06γ-Proteobacteria 37.50 ― ―δ-Proteobacteria 12.50 6.25 1.52ε-Proteobacteria ― 57.81 68.18Spirochaetes ― 3.13 ―SRI ― 1.56 10.61OPII ― 1.56 ―Green nonsulfur bacterium 1.56 ― 1.52CFB ― 6.25 4.55

2010 27Diversity of microbes and potential exoelectrogens in MFC
their energy was obtained from degrading organic and amino acids (Holt et al., 1994). Their main functions were not to generate electrons, but rather to carry out preliminary degradation of organic matter. This gave them their indirect, secondary function, which was to provide suitable substrates for the real exoelectronic bacteria, thus improving their electrochemical activity. In order to shorten the electron transfer distance, most exoelectrogens will attach to the anode surface that is connected into a closed circuit. However, exo-electrogens might occasionally appear on the open circuit anode because the two anodes are laid in one MFC system at the same time. Therefore, any unique bacterial species appearing in sample B, but not in sample C, should have more attention paid to it in fu-ture. In addition to some uncultured bacteria, seven species were the only species that appeared in sample B with closed circuits. Bacteria not appearing in sam-ple C from open circuits included Trichococcus fl oc-
culiformis, Streptococcus bovis, Spirochaeta sp., Al-
caligenes monasteriensis, Comamonas denitrifi cans, and Dechloromonas sp. In the mechanism used for exocellular transport of electrons, as has been demon-strated elsewhere, cell-bound outer membrane cyto-chromes and conductive fl agella may play a key role in electron transfer for some Geobacter and Shewanel-
la species (Gorby et al., 2006; Lovley et al., 2004; My-ers and Myers, 1992; Reguera et al., 2005). Alterna-tively, some exoelectrogens, such as Pseudomonas aeruginosa and Geothrix fermentans, excrete media-tors that allow shuttling of electrons to surfaces (Bond and Lovley, 2005; Rabaey et al., 2004). Some sequences of OTUs, B7 and B8, belonging to low-GC positive-gram bacteria, are affi liated, respec-tively, with Trichococcus fl occuliformis and Strepto-
coccus bovis. These bacteria are characterized by heterotrophic fermentation, are without fl agella, and are usually found in active sludge (Holt et al., 1994). They may act in MFCs for degrading organic matter. The sequence OTU9, affi liated with Spirochaeta sp., usually appears in wastewater, with the main charac-teristic of fermenting carbohydrates into simple organ-ic acids (Holt et al., 1994). The OTU sequences B17, B18 and B19 are associated with β-Proteobacteria, and are affi liated with Alcaligenes monasteriensis, Co-
mamonas denitrifi cans and Dechloromonas sp., re-spectively. They all have a nanowire formation and carry out anaerobic respiration (Holt et al., 1994). Al-
caligenes monasteriensis and Comamonas denitrifi -
cans easily decompose organic acids, and can trans-fer electrons to dioxide nitrate (Holt et al., 1994; Tianshen et al., 2007). Dechloromonas genus can use chlorate as the terminal electron acceptor, and demon-strates nitrogenase activity (Tan and Reinhold-Hurek, 2003). The current study found that most exoelectrogenic bacteria carry on anaerobic respiration. Some of the exoelectrogens were able to transfer electrons to ex-tracellular electron acceptors, such as Fe3+ oxides, when the organic chemicals were degraded. Included among these are bacteria such as Geobacter spp. and Shewanella putrefaciens (Kim et al., 2002; Leang et al., 2003). Nitrates and sulfates also served as terminal elec-tron acceptors for the exoelectrogens of Pseudomonas aeruginosa (Carlson and Ingraham, 1983), Ochrobac-
trum anthropi (Kesseru et al., 2002), Desulfobulbus propionicu (Holmes et al., 2004b), and Desulfi tobacte-
rium hafniense (Milliken and May, 2007). On the other hand, Geobacter sulfurreducens (Bond and Lovley, 2003) and Desulfobulbus propionicus (Holmes et al., 2004b) have the ability to use both sulphates and Fe3+ as electron acceptors. Thus, Alcaligenes monasteriensis, Comamonas denitrifi cans, and Dechloromonas sp. can be deduced to be potential exoelectrogenic bac-teria in MFCs.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (50708008), Special fund of State Key Joint Laboratory of Environment Simulation and Pollution Con-trol of China (08K07ESPCT).
References
Bond, D. R., Holmes, D. E., Tender, L. M., and Lovley, D. R. (2002) Electrode-reducing microorganisms that harvest energy from marine sediments. Science, 295, 483‒485.
Bond, D. R. and Lovley, D. R. (2003) Electricity production by Geobacter sulfurreducens attached to electrodes. Appl. Environ. Microbiol., 69, 1548‒1555.
Bond, D. R. and Lovley, D. R. (2005) Evidence for involvement of an electron shuttle in electricity generation by Geothrix fermentans. Appl. Environ. Microbiol., 71, 2186‒2189.
Bullen, R. A., Arnot, T. C., Lakemanc, J. B., and Walsh, F. C. (2006) Biofuel cells and their development. Biosens. Bio-
electron., 21, 2015‒2045.Carlson, C. A. and Ingraham, J. L. (1983) Comparison of deni-
trifi cation by Pseudomonas stutzeri, Pseudomonas aerugi-
nosa, and Paracoccus denitrifi cans. Appl. Environ. Micro-

28 Vol. 56SUN et al.
biol., 45, 1247‒1253.Chaudhuri, S. K. and Lovley, D. R. (2003) Electricity generation
by direct oxidation of glucose in mediator-less microbial fuel cells. Nat. Biotechnol., 21, 1229‒1232.
Choo, Y. F., Lee, J., Chang, I. S., and Kim, H. (2006) Bacterial communities in microbial fuel cells enriched with high con-centrations of glucose and glutamate. J. Microbiol. Bio-
technol., 16, 1481‒1484.Fusheng, W. , Wenqi, Q., and Xinfang, W. (2002) China Environ-
mental Protection Bureau, Water and Wastewater Analytic Methods, 4th ed., China Environmental Science Press, Bei-jing.
Gorby, Y. A., Yanina, S., Mclean, J. S., Rosso, K. M., Moyles, D., and Dohnalkova, A. (2006). Electrically conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and other microorganisms. Proc. Natl. Acad. Sci. USA, 103, 11358‒11363.
Holmes, D. E., Bond, D. R., O’Neil, R. A. , Reimers, C. E. , Ten-der, L. R. , and Lovley, D. R. (2004a) Microbial communities associated with electrodes harvesting electricity from a va-riety of aquatic sediments. Microb. Ecol., 48, 178‒190.
Holmes, D. E., Bond, D. R., and Lovley, D. R. (2004b) Electron transfer by Desulfobulbus propionicus to Fe(III) and graph-ite electrodes. Appl. Environ. Microbiol., 70, 1234‒1237.
Holmes, D. E., Nicoll, J. S., Bond, D. R., and Lovley, D. R. (2004c) Potential role of a novel psychrotolerant member of the family Geobacteraceae, Geopsychrobacter electrodiphilus gen. nov., sp. nov., in electricity production by a marine sediment fuel cell. Appl. Environ. Microbiol., 70, 6023‒6030.
Holt, J. G., Krieg, N. R., Sneath, P. H. A., Staley, J. T., and Wil-liams, S. T. (eds.) (1994). Bergey’s Manual of Determinative Bacteriology, 9th ed., Lippincott Williams and Wilkins, Bal-timore, MD.
Kaufmann, F. and Lovley, D. R. (2001) Isolation and character-ization of a soluble NADPH-dependent Fe(III) reductase from Geobacter sulfurreducens. J. Bacteriol., 183, 4468‒4476.
Kesseru, P., Kiss, I., Bihari, Z., and Polyák, B. (2002) The effects of NaCl and some heavy metals on the denitrifi cation activ-ity of Ochrobactrum anthropi. J. Basic Microbiol., 42, 268‒276.
Kim, B. H., Kim, H. J., Hyun, M. S., and Park, D. S. (1999) Direct electrode reaction of Fe(III) reducing bacterium, Shewanel-
la putrefaciens. J. Microbiol. Biotechnol., 9, 127‒131.Kim, B. H., Park, H. S., Kim, H. J., Kim, G. T., Chan, I. S., Lee, J.,
and Phung, N. T. (2004) Enrichment of electrochemically active anaerobic bacteria using a fuel cell type electro-chemical cell. Appl. Microbiol. Biotechnol., 63, 672‒681.
Kim, G. T., Webster, G., Wimpenny, J. W. T. , Kim, B. H. , Kim, H. J., and Weightman, A. J. (2006) Bacterial community struc-ture, compartmentalization and activity in a microbial fuel cell. J. Appl. Microbiol., 101, 698‒710.
Kim, H. J., Park, H. S., Hyun, M. S., Chang, I. S., Kim, M., and
Kim, B. H. (2002) A mediator-less microbial fuel cell using a metal reducing bacterium, Shewanella putrefaciens. En-
zyme Microb. Technol., 30, 145‒152.Kim, J. R., Jung, S. H., Regan, J. M., and Logan, B. E. (2007)
Electricity generation and microbial community analysis of alcohol powered microbial fuel cells. Bioresour. Technol., 98, 2568‒2577.
Kimura, M. (1980) A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol., 16, 111‒120.
Kumar, S., Tamura, K., and Nei, M. (2004) MEGA3, integrated software for molecular evolutionary genetics analysis and sequence alignment. Brief. Bioinform., 2, 150‒163.
Lane, D. J. (1991) 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics, ed. by Stackebrandt, E. and Goodfellow, M., John Wiley & Sons, Inc., New York, pp. 115‒175.
Leang, C., Coppi, M. V., and Lovley, D. R. (2003) A c-type poly-heme cytochrome, involved in Fe(III) reduction in Geobacter sulfurreducens . J. Bacteriol., 185, 2096‒2103.
Lee, J., Phung, N. T., Chang, I. S., Kim, B. H., and Sung, H. C. (2003) Use of acetate for enrichment of electrochemically active microorganisms and their 16S rDNA analyses. FEMS Microbiol. Lett., 223, 185‒191.
Liu, H., Cheng, S., and Logan, B. E. (2004) Production of elec-tricity from acetate or butyrate using a single-chamber mi-crobial fuel cell. Environ. Sci. Technol., 39, 658‒662.
Liu, W. T., Marsh, T. L., Cheng, H., and Forney, L. J.(1997) Char-acterization of microbial diversity by determining terminal restriction fragment length polymorphisms of genes en-coding 16S rRNA. Appl. Environ. Microbiol., 63, 4516‒4522.
Logan, B. E., Murano, C., Scott, K., Gray, N. D., and Head, I. M. (2005) Electricity generation from cysteine in a microbial fuel cell. Wat. Res., 39, 942‒952.
Logan, B. E. and Regan, J. M. (2006) Microbial fuel cells: Chal-lenges and applications. Environ. Sci. Technol., 40, 5172‒5180.
Lovley, D. R., Holmes, D. E., and Nevin, K. P. (2004) Dissimila-tory Fe(III) and Mn(IV) reduction. Adv. Microb. Physiol., 49, 219‒286.
Magnuson, T. S., Hodges-Myerson, A. L., and Lovley, D. R. (2000) Characterization of a membrane-bound NADH-de-pendent Fe3+ reductase from the dissimilatory Fe3+-re-ducing bacterium Geobacter sulfurreducens. FEMS Micro-
biol. Lett., 185, 205‒211.Milliken, C. E. and May, H. D. (2007) Sustained generation of
electricity by the spore-forming, Gram-positive, Desulfi to-
bacterium hafniense strain DCB2. Appl. Microbiol. Biotech-
nol., 73, 1180‒1189.Myers, C. R. and Myers, J. M. (1992) Localization of cytochromes
to the outer membrane of anaerobically grown Shewanella putrefaciens MR-1. J. Bacteriol., 174, 3429‒3438.
Nguyet, T. P., Jiyoung, L., and Kui, H. K. (2004) Analysis of mi-

2010 29Diversity of microbes and potential exoelectrogens in MFC
crobial diversity in oligotrophic microbial fuel cells using 16S rDNA sequences. FEMS Microbiol. Lett., 233, 77‒82.
Park, H. S., Kim, B. H., Kim, H. S., and Kim, H. J. (2001) A novel electrochemically active and Fe(III)-reducing bacterium phylogenetically related to Clostridium butyricum isolated from a microbial fuel cell. Anaerobe, 7, 297‒306.
Pham, C. A., Jung, S. J., Phung, N. T., Lee, J.,Chang, I. S., Kim, B. H., Yi, H., and Chun, J. (2003) A novel electrochemically active and Fe(III)-reducing bacterium phylogenetically re-lated to Aeromonas hydrophila, isolated from a microbial fuel cell. FEMS Microbiol. Lett., 223, 129‒134.
Rabaey, K., Boon, N., Siciliano, S. D., Verhaege, M., and Ver-straete, W. (2004) Biofuel cells select for microbial consor-tia that self-mediate electron transfer. Appl. Environ. Micro-
biol., 70, 5373‒5382.Reguera, G., McCarthy, K. D., Mehta, T., Nicoll, J. S., Tuominen,
M. T., and Lovley, D. R. (2005) Extracellular electron trans-fer via microbial nanowires. Nature, 435, 1098‒1101.
Reimers, C. E., Tender, L. M., Fertig, S., and Wang, W. (2001) Harvesting energy from the marine sediment-water inter-face. Environ. Sci. Technol., 35, 192‒195.
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) Molecular Cloning, a Laboratory Manual, 2nd ed., Cold Spring Har-
bor Laboratory Press, New York, pp. 1.85‒1.86.Sokhee, J. and John, M. R. (2007) Comparison of anode bacte-
rial communities and performance in microbial fuel cells with different electron donors. Appl. Microbiol. Biotechnol., 77, 393‒402.
Tan, Z. and Reinhold-Hurek, B. (2003) Dechlorosoma suillum Achenbach et al. 2001 is a later subjective synonym of Azospira oryzae Reinhold-Hurek and Hurek 2000. Int. J. Syst. Evol. Microbiol., 53, 1139‒1142.
Tianshen, T., Ruifu, Y., and Xiuzhu, D.(2007) Systematics Prokaryota, Chemical Industry Publishing Company, Bei-jing, China.
Weisburg, W. G., Barns, S. M., Pelletier, D. A., and Lane, D. J. (1991) 16S ribosomal DNA amplifi cation for phylogenetic study. J. Bacteriol., 173, 697‒703.
Yeates, C., Gillings, M.R., Davison, A.D., Altavilla, N., and Veal, D.A. (1998) Methods for microbial DNA extraction from soil for PCR amplifi cation. Biol. Proced. Online., 1, 40‒47.
Zuo, Y., Xing, D., John, M. R., and Logan, B. E. (2008) Isolation of the exoelectrogenic bacterium Ochrobactrum anthropi
YZ-1 by using a U-tube microbial fuel cell. Appl. Environ., Microbiol., 74, 3130‒3137.