intertidal gracilaria farming in southern chile: differences of the algal proveniance
TRANSCRIPT

Aquatic Botany, 42 (1992) 327-337 327 Elsevier Science Publishers B.V., Amsterdam
Intertidal Gracilaria farming in southern Chile: differences of the algal proveniance
Alejandro H. Buschmann, Felix A. Kuschel, Pedro A. Vergara and Jacqueline Schulz
Instituto Profesional de Osorno, Departamento de Acuicultura y l~ecursos Acudticos, Casilla 933, Osorno, Chile
(Accepted 25 November 1991 )
ABSTRACT
Buschmann, A.H., Kuschel, EA., Vergara, P.A. and Schulz, J., 1992. Intertidal Gracilaria farming in southern Chile: differences of the algal proveniance. Aquat. Bot., 42: 327-337.
The transplantation of Gracilaria for the establishment of cultivation centres has been a common practice in Chile, but so far differences in the initial transplants and their eventual morphologies have not been noted. Subtidal populations of Gracilaria obtained in the Maullin area ( 41 ° 34' S, 73 ° 39' W) were initially morphologically different from intertidal Gracilaria populations collected in the Quil- laipe area ( 41 ° 33' S, 72 ° 45' w). Both populations were grown under the same experimental intertidal conditions in Metri Bay (41 ° 36'S, 72°42'W). After several months of cultivation, the morphological differences still existed. Significant differences in the yield and gel strength ofagar, and susceptibility to epiphytism, were found. However, no differences were detected in the productivity of each initially different population. This evidence indicates that the success of Gracilaria farming will depend on the origin of the initial Gracilaria inoculum.
INTRODUCTION
Intraspecific var ia t ions in the morphologica l and funct ional characteris t ics o f benth ic algae may be a reflection o f differences in habi ta t ( N o r t o n et al., 1981; L o b b a n et al., 1985; Chapman , 1986) . Han i sak et al. ( 1 9 8 8 ) found that the morphological in t rapopula t ion differences o f Gracilaria t ikvahiae Mc- Lachlan are reflected in different funct ional responses o f the plant. F r o m a produc t ive poin t o f view, these differences could de te rmine the success o f a new cul ture (Santel ices and Doty , 1989; Han i sak et al., 1990) .
The cul t ivat ion o f Gracilaria in Chile has exper ienced rapid deve lopment dur ing the last 5 years (Ponce , 1989) , A t radi t ional pract ice for the establish- ment o f new cult ivation centres involves the t ransplantat ion o f Gracilaria f rom
Correspondence to: A.H. Buscbmann, lnstituto Profesional de Osomo, Departamento de Acui- cultura y Recursos AcmRicos, Casilla 933, Osorno, Chile.
© 1992 Elsevier Science Publishers B.V. All rights reserved 0304-3770/92/$05.00

328 A.H. BUSCHMANN ET AL.
different localities on the Chilean coastline (Santelices, 1989). This has not only created difficulties in the reliable identification of the Chilean Gracilaria species (Bird et al., 1986), but further difficulties arise when ar'dficial com- mercial Gracilaria beds are installed without taking eventual ecological inter- population differences into consideration (Santelices, 1989; Santelices and Dory, 1989). Using tank and laboratory experiments, Santelices and Ugarte (1990) demonstrated that subtidal Gracilaria populations from northern, central and southern Chile show some ecological differences that might affect the success of their cultivation. The different environmental systems used for the Gracilaria mariculture activities, such as wave-sheltered bays, estuaries and intertidal flats (Santelices and Doty, 1989), can increase the risks when initiating aquaculture activities using algae with different ecological charac- teristics with respect to abiotic factors (e.g. irradiance, temperature, sand abrasion and sand burial) and biological factors (e.g. resistance to epiphy- tism and herbivory).
In southern Chile, algae from two environmental systems have been used for the cultivation of Gracilaria, namely subtidal sheltered estuaries and in- tertidal mud flats. The cultivation in intertidal mud flats occupies approxi- mately 1000 ha, representing about 30% of the ste.te area licensed for Graci- laria cultivation in southern Chile (Ponce, 1989). These mud flats possess specific environmental characteristics suitable for testing a contrasting set of conditions shown by a subtidal estuarine habitat on given populations of Gra- cilaria. For these reasons, it is important from a productive point of view to establish whether the different Gracilaria populations used for planting com- mercial intertidal farms respond in a different or the same way. We tested the hypothesis that the response of subtidal Gracilaria estuarine populations will differ from that of the intertidal populations when they are cultivated in in- tertidal flats. Growth responses were measured in terms of changes in mor- phology, biomass production, agar yield and gel strength, epiphyte load and grazing resistance.
MATERIALS A N D M E T H O D S
Study site, algal provenance and experimental 4esign
The Gracilaria populations were collected in two localities in Chile: Maullin (41 °34'S, 73°39'W) and Quillaipe (41 °33'S, 72°45'W) (Fig. 1A). The first locality is a wave-protected subtidal estuarine habitat and the Gra- cilaria beds extend between 1 and 3 m on a sand bottom, with temperatures and salinities ranging between 9 and 16°C and 18 and 32%0, respectively (Westermeier et al., 1984, 1988). The Quillaipe area represents an intertidal flat created as a consequence of the high tidal amplitudes found in this area,
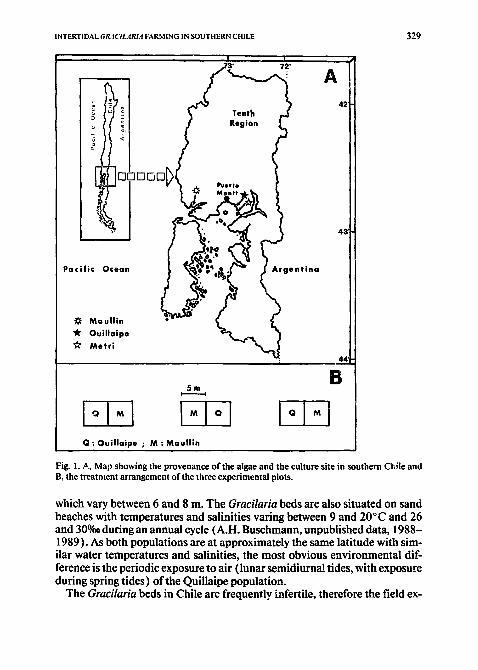
INTERTIDAL GI~4CILARIA FARMING IN SOUTHERN CHILE
I I
A 42'
329
43'
Pac n t i n a
44'
B 5m
I I IMlol 101 l O : O u i l l a i p e ; M : M a u l l i n
Fig. 1. A, Map showing the provenance of the algae and the culture site in southern Chile and B, the treatntent arrangement of the three experimental plots.
which vary between 6 and 8 m. The Gracilaria beds are also situated on sand beaches with temperatures and salinities varing between 9 and 20°C and 26 and 30%o during an annual cycle (A.H. Buschmann, unpublished data, 1988- 1989). As both populations are at approximately the same latitude with sim- ilar water temperatures and salinities, the most obvious environmental dif- ference is the periodic exposure to air (lunar semidiumal tides, with exposure during spring tides) of the Quillaipe population.
The Gracilaria beds in Chile are frequently infertile, therefore the field ex-

330 A.H. BUSCHMANN ET AL.
periments were performed without establishing the specific identity of the plants used. However, it is known that the Maullin algae correspond mainly to Gracilaria chilensis Bird, McLachlan & Oliveira. Observations on Quil- laipe Gracilaria indicate that it corresponds to Gracilaria chilensis, but a more detailed study must be carried out. Both populations differ morphologically: the Quillaipe algae have thinner thalli (Mann Whitney U-test; U=85; P<0.01 ) and are also significantly more ramified (Mann Whitney U-test; U=96; P< 0.01 ).
The algae (about 400 kg from each site) were transported within 6 h to Metri Bay (41 °36'S, 72°42'W), l0 km southeast of Quillaipe Bay (Fig. IA). Metri Bay is a wave-sheltered bay with tidal amplitudes also ranging between 6 and 8 m. Three experimental plots (5 m × l0 m each) were installed in this bay. Each plot was divided into two (5 m × 5 m each) in order to test the effects of two different treatments (Fig. l B). Both populations studied were planted using polyethylene tubes filled with sand (see Pizarro, (1986) and Santelices and Doty( 1989 ) for more details). The three experimental plots were situated between 0.80 and 1.20 m of the tidal high above mean-low- water and 2 kg of algae m -2 were planted in August 1988.
Morphological analysis
Ten 15.0 cm apical portions of Gracilaria were obtained from the Maullin and the Quillaipe algae at the start of the experimental culture, and after 8 months of cultivation. Each of the thaUi was weighed (wet weight) on a Sar- torius balance (0.001 g accuracy) and the number of primary ramifications (those emerging from the central axis) counted. Subsequently, the wet weight of the central axis was determined by removing the laterals and the percent weight of the central axis as a percentage of the total weight (with ramifica- tions) was determined.
Biomass production
During the growing season, spring-summer (between September 1988 and February 1989), all the harvestable biomass was removed manually 15 cm above the bottom. The harvested algae were put in labelled plastic bags and brought to the laboratory where the wet weight was determined on a Sartorius balance ( l g accuracy), after a standardized drainage time.
Agar analysis
Six months after the experimental culture was initiated (March 1988), three samples of Gracilaria were obtained from Maullin and another three from Quillaipe. Agar was extracted from each sample according to the procedure

INTERTIDAL GRACTLARIA FARMING IN SOUTHERN CHILE :3:31
described by Canting and Orellana (1987), and the percent dry weight was calculated. The gel strength was obtained by measuring a 1.5% agar solution prepared in I00 ml distilled water and gelled at room temperature (16°C) for 12 h. The gel strength measurement was carried out utilizing a 'Marine Colloidal Gel Tester'. A total of five measurements was taken for each replicate.
Epiphytism resistance
Before installing the experimental plots, the algae from Manllin and Quil- laipe were cleaned by hand. The Maullln algae showed a very small load of epiphytes (mainly Ceramiales), but the algae collected in Quillaipe had a higher epiphyte load (up to 30% Ceramiales; wet weight). The clean algae were then planted in the experimental plots as described earlier and, after 3 months of culture, between October and November when the epiphyte load owing to the filamentous brown alga Giffordia sp. is highest (Buschmann ct al., 1990; Kuschel and Buschmann, 1991 ), three subsamples (0.5×0.5 m) were taken from each replicate plot and the epiphytes were removed by hand in the laboratory. The Gracilaria and the epiphytes (Giffordia) were then weighed on a Sartorius balance ( __.0.01 g accuracy). The epiphyte load was then expressed as the weight of epiphytes divided by the total algal weight (~0).
Herbivor2 resistance
During May 1990, samples of Maullin and Quillaipe algae were collected from the experimental plots in Metri (22 month acclimatized algae). Maullin algae which had not been acclimatized were also brought to Metri. All the algae were transported to the laboratory and epiphytes removed. Using the algae collected, six bundles of I00 g (wet weight) ofthe acclimatized Maullfn Gracilaria were prepared, six of the non-acclimatized Manllin and six of the QuiUaipe algae. A bundle of each ofthe three different types ofalgae was fixed to short polyethylene tubes (30 em long).
The snail Te~la atra Lesson is the most common ~'azer in the intertidal fiats in southern Chile, reaching over 30 individuals m -= (A.H. Buschmann, unpublished results, 1990). Experimental cages were used to maintain a fixed number of snails for food preference experiments in the field. The cages (50 cm X 50 cm X 50 cm) were anchored in the sand bottom and algal offerings placed inside them (Maullln and Quillaipe algae), as previously described. Thirty individuals of T. atra were added to each ofthe three cages. Consump- tion rates were calculated after 30 days based on weight differences of the algae measured on a balance ( +_ 1 g accuracy), and after making corrections for weight changes which occurred in three other unf~.ed controls. Con- sumption rates were then standardized by dividing the fresh weight of the

332 A.H. BUSCHMANN El" AL.
algae consumed by the fresh weight of the snails, multiplied by the number of experimental days.
Statistical analysis
The morphological data were analysed using the non-parametric U-test (Sokal and Rohlf, 1979) because some of the data did not meet the homoge- neity of variance and normality requirements. All other data were compared using univariate statistical analysis (t-test and ANOVA) throughout (Sokal and Rohlf, 1979).
RESULTS
Morphology
After 8 months of culture, the initial morphological differences observed between the Maullin and Quillaipe Gracilaria populations remained. The to- tal weight of the 15 cm long thalli was not significantly different in the two algal populations (Table 1 ). However, the number of ramifications was sig- nificantly higher (Mann-Whi tney U test; U= 83; P < 0.01 ) in the Quillaipe algae than in the Maullin algae (Table 1 ). The weight of the central axis was significantly higher (Mann-Whi tney U test; U=I00 ; P<0 .001) in the Maullin than in the Quillaipe Gracilaria populations (Table 1 ). As a conse- quence, given that the Quillaipe algae were more branched and had thinner thalli, this population presented a higher surface-volume ratio during the pe- riod studied than the Maullin algae.
TABLE 1
Morphological characterization of 15 cm long thalli of Gracilaria originating from Maullin (M) and Quillaipe (Q). A, total wet weight; E, number of primary ramifications; C, wet weight of the central axis/total weight
Morphological M Q characteristics (mean_+ 1 SE) (mean_+ 1 SE)
A 226.3_+ 37.9 226.7 _+ 15.8* B 20.1 _+ 3.4 32.9_+ 4.4** C 60.2_+ 7.4 29.7_+ 2.3***
SE, standard error. *Not significant (Mann-Whitney test, U=63); **P<0.01 (Mann-Whitney test, U=83); ***P<0.001 (Mann-Whitney test, U= ! 00).
INTERTIDAL GRACILARIA FARMING IN SOUTHERN CHILE 333
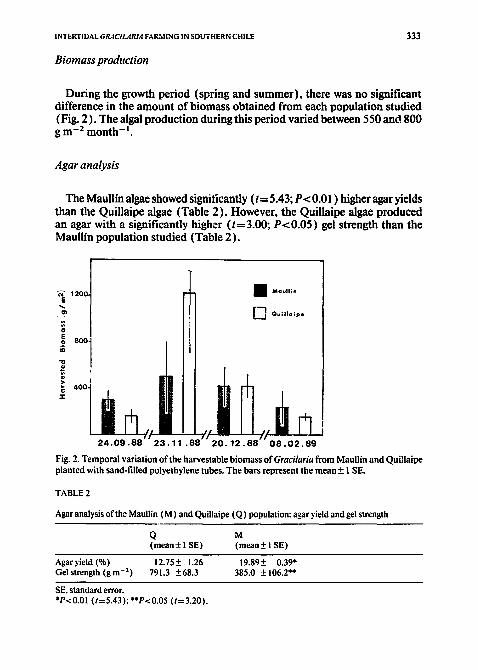
Biomass production
During the growth period (spring and summer), there was no significant difference in the amount of biomass obtained from each population studied (Fig. 2). The algal production during this period varied between 550 and 800 g m -2 month- i.
Agar analysis
The Maullin algae showed significantly (t = 5.43; P< 0.01 ) higher agar yields than the Quillaipe algae (Table 2). However, the Quillaipe algae produced an agar with a significantly higher (t=3.00; P<0.05) gel strength than the Maullin population studied (Table 2).
1200- - I Maullin E
~1~ I ' l l Quil laip e
.o 800. IB
i • • 2 3 . 1 1 2 0 1 0 8 0 2 8 9
Fig. 2. Temporal variation oftbe harvestable biomass of Gracilaria from Maullin and Quillaipe planted with sand-filled polyethylene tubes. The bars represent the mean + 1 SE.
TABLE 2
Agar analysis of the Maullin (M) and Quillaipe (Q) population: agar yield and gel strength
Q M (mean+ 1 SE) (mean+ 1 SE)
Agaryield (%) 12.75+ 1.26 19.89+ 0.39* Gel strength (g m -2) 791.3 +68.3 385.0 +106.2"*
SE, standard error. *P<0.01 (t=5.43),**P<0.05 (t=3.20).

334 A,H. BUSCHMANN ET AL,
TABLE 3
Epiphytism susceptibility ( (g epiphytes/g algae) × i 00) of Gracilaria from Maullln (M) and Quil- laip¢ (Q) populations
Gracilaria population Epiphytism load (%) (mean+- 1 SE)
M 45.6+-6.8
Q 23.2+_3.3
SE, standard error. *P<0.01 (tffi3.48).
TABLE 4
Herbivory susceptibility of Oracilaria (g algae consumed per g Teluga per month) from the acclima- tized Maullfn (Ma), not acclimatized Maullin (Mn) and Quillaipe (Q) populations
Consumption rate (mean+_ 1 SE)
Ma 0.33+0.18 Mn 0.23+-0.16 * Q 0.19+-0.12
SE, standard error. • Not significant (ANOVA: Fffi0.20).
Epiphytism resistance
During the period of a higher degree of epiphytism by Giffordia sp. (Octo- ber-November; Kuschel and Buschmann, 1991 ), the Maullfn algae showed significantly higher (tffi 3.48; P< 0.05) levels of epiphytism than the Quil- laipe algae (Table 3 ).
Herbivory resistance
The food preference experiment carried out during the winter failed to demonstrate any significant difference (ANOVA; F---0.20) between the ac- climatized/non-acclimatized Maullin algae and the Quillaipe Gracilaria (Ta- ble 4).
DISCUSSIOY
Hanisak et al. (1988) found that different morphologies of G. tikvahiae had different productive capacities, depending principally on the surface- volume ratios. According to the predictions made by Littler and Littler

INTERTIDAL ORACILARIA FARMING IN SOUTHERN CHILE ~35
(1980), Gracilaria with a higher surface-volume ratio had higher production capacities (Hanisak ct al., 1988, 1990). Following this argument, it would be expected that the Quillaipe algae with a higher surface-volume ratio would also have higher biomass production rates. However, in the field it was Found that this potential was not realized. This evidence suuests that both algae could be equally productive, but several other aspects must be considered be/ore transplanting Gracilaria to an intertidal cultivation centre.
The agar yield and gel strength of the Maullin algae differed from that of the QuiUaipe algae. The Maullfn algae had common yield values (20%), but the Quillaipe algae had low yields (below 15%) compared with previous re. sults (McLachlan and Bird, 1986). Several Factors could account for these differences. The results regarding the gel strength also varied between the populations studied. It is known that both agar characteristics of yield and gel strength are independent variables (McLachlan and Bird, 1986). As different algal clones can show differences in the chemical constituents of the agar (Craigie and Wen, 1984), it is not surprising that the gel strength varied be- tween the Maullfn and Quillaipe area. Again, the reasons for these differences are still unknown. However, from a production point of view, the quantity and quality or'the final product will vary depending on the source of the algae used for the initial planting.
Epiphytism could be a major problem for the establishment of cultivation areas in southern Chile. The results obtained in this study show the same pattern as that reported previously by Kuschel and Buschmann (I 991 ). The Maullfn algae were more susceptible to epiphytism by Giffordia. The initial epiphyte load could not account for these differences because the Quillaipe (more resistant population) had the highest initial epiphyte load. Further- more, if epiphytism were to depend on the surface area available, it would be expected that the morphology of the Quillaipe algae (higher surface-volume ratio) could enhance epiphytism by Giffordia. However, we Found that the Quillaipe algae had the lower epiphyte level. Santelices and Ugarte (1990) also Found in their experiments that the Maullfn population showed recruit- ment of the brown alga Ectocarp~ sp. ( = Giffordia). As Giffordia can signif- icantly suppress Gracilaria production in intertidal systems (Kuschel and Buschmann, 1991 ), it is a very important variable which must be considered in the establishment of commercial intertidal beds.
No differences were Found between the subtidal algae (Maullfn) which had been acclimatized to intertidal environmental conditions (r~=::: • stressful conditions) and subtidal algae which had not been acclimatiz~ :' ]~ naud et al. (1990) have shown that desiccation can modify the susceptibility of G. tikvahiae to sea urchin grazing. We have not Found such differences between the acclimatized and non-acclimatized Maullfn algae, and our results do not confirm those of Renaud et al. (1990). Our results also indicate that no dif- Ferences exist between the algae from the Maullfn area and those from the Quillaipe area.

336 A.H. BUSCHMANN ET AL.
Considering that in Chile propagation is carried out through fragmentation of the thai!i, it is probable that the mariculture practices have already shown some sort of selection, diminishing the natural variability both within and between populations. However, the results obtained in this study indicate that some differences exist when algae of different provenience, cultivated in the same field conditions, are compared, and they must be considered before es- tablishing a commercial Gracilaria farm in intertidal systems in southern Chile. The evidence also indicates that there is a need to introduce a Graci- laria strain-selection program in Chile in order to obtain better production results in southern Chile.
ACKNOWLEDGEMENTS
We are very grateful for the help of Eugenio P6rez and the students from the 'Seaweed Culture' class 1988. The supply of algae by PACTO S.A. and Pesquera Alerce is specially acknowledged. We recognize the collaboration and help of Juan Cancino and Gonzalo Benavides with the agar analysis. Our special thanks go to Bernab~ Santelices and Daniel L6pez for revising and commenting on early Spanish versions of the manuscript. We are also grateful for the cfiticism ma0e by the referees and a review of the English by Susan Angus. This research was supported by a FONDECYT grant (No. 103-88) by the Direcci6n de Investigaci6n of the Instituto Profesional de Osorno (D.I.- 1.P.O. No. 304.14) and a IFS-Sweden gralit ( No. A/1600- l ).
REFERENCES
Bird, C.J., McLachlan, J. and de Oliveira, E.C., 1986. Gracilaria chilensis sp. nov. (Rhodo- phyta, Gigartinales) from Pacific South America. Can. J. Bot., 64: 2928-2934.
Buschmann, A.H., Kuschel, F.A. and P6rez, E., 1990. Field assessment of intertidal culture of Gracilaria (Rhodophyta: Gigartinales) in southern Chile. In: E.C. de Oliveira and N. Kaut- sky (Editors), Cultivation of Seaweeds in Latin America. Universidad de Sao Paulo, Brazil, pp. 69-74.
Cancino, J. and Orellana, M.C., 1987. Interacci6n entre Gracilaria verrucosa y su fauna epifita: pe;spectivas para la utilizaci6n de mitilidos como fuente de nutrientes en cultivos de Graci- laria en piscinas litorales. In: J.A.J. Verreth, M. Carriilo, S. Zanuy and E.A. Huisman (Edi- tors), Investigaci6n Acuicola en America Latina. Pudoc, Wageningen, pp. 380-392.
Chapman, A.R.O., 1986. Population and community ecology of seaweeds. Adv. Mar. Sci., 16: 1-161.
Craigie, J.S. and Wen, Z.C., 1984. Interspecific, intraspecific and nutritionally-determined vari- ations in the composition of agars from Gracilaria spp. Bot. Mar., 27: 55-61.
Hanisak, M.D., Littler, M.M. and Littler, D.S., 1988. Significance ofmacroalgal polymorphism: intraspecific tests of the functional-fo~m model. Mar. Biol., 99:157-165.
Hanisak, M.D., Littler, M.M. and Littler, D.S., 1990. Application of the functionai-fornJ model to the culture of seaweeds. Hydrobiologia, 204/205: 73-77.
Kuschel, FA. and Buschmann, A.H., 1991. Abundance, effects and management of epiphytism in intertidal cultures of Gracilaria (Rhodophyta) in southern Chile. Aquaculture, 92: 7-19.
Littler, M.M. and Littler, D.S., 1980. The evolution of thallus form and survival strategies in

INTERTIDAL GRACILARIA FARMING IN SOUTHERN CHILE 337
benthic marine macroalgae: field and laboratory test of a functional form model. Am. Nat., 116: 25-44.
Lobban, C.S., Harrison, P.J. and Duncan, M.J., 1985. The Physiological Ecology of Seaweeds. Cambridge University Press, Cambridge, 242 pp.
McLachlan, J. and Bird, C.J., 1986. Gracilaria (Gigartina!es, Rhodophyta) and productivity. Aqual. Bot., 26: 27-49.
Norton, T.A., Mathieson, A.C. and Neushul, M., 1981. Morphology and environment. In: C.S. Lobban and M.J. Wynne (Editors), The Biology of Seaweeds. Blackwell Scientific, Oxford, pp. 421-451.
Pizarro, A., 1986. Conocimiento actual y avances recientes sobre el manejo y cultivo de Grac# laria en Chile. Monogr. Biol., 4: 63-96.
Ponce, F., 1989. Un eslab6n en la acuicultura: E! cultivo de Gracilaria en Chile, Acuanoticias Int., 1: 30-37.
Renaud, P.E., Hay, M.E. and Schmitt, T.M., 1990. Interactions of plant stress and herbivory: intraspecific variation in the susceptibility of a palatable versus an unpalatable seaweed to sea urchin grazing. Oecologia (Berlin), 82:217-226.
Santelices, B., 1989. Algas Marinas de Chile. Distribuci6n, ecologia, utilizaci6n y diversidad. Ediciones Universidad Cat61ica de Chile, Santiago, 399 pp.
Santelices, B. and Dory, M.S., 1989. A review of Gracilaria farming. Aquaculture, 78: 95-133. Santelices, B. and Ugarte, R., 1990. Ecological differences among Chilean populations of com-
mercial Gracilaria. J. Appl. Phycol., 2:17-26. Sokal, R.R. and Rohlf, F.J., 1979. Biometria. Principios y M~todos Estadisticos en la Investi-
gaci6n Biol6gica. H. Blumc, Spain, 832 pp. Westermeier, R., Steubing, L., Rivera, P.J. and Wenzel, H., 1984. Gracilaria verracosa (Hud-
son) Papenfuss en la X Regi6n (Maulliny Quenuir-Provincia Lianquihue), Chile. Mem. Asoc. Latinoam. Acuicult., 5:413-430.
Westermeier, R., Rivera, P. and G6mez, I., 1988. El uso de mangas de polietileno como sustrato en el repoblamiento de Gracilaria sp. (Rhodophyta, Gigartinaceae) en el sur de Chile. Gay- ana Bot., 45: 95-106.