inmunologia - el blog de israel masa · las propias membranas mucosas secretan sustancias ... con...
TRANSCRIPT

INMUNOLOGIA 1. INTRODUCCIÓN 2. DEFENSAS INESPECÍFICAS O MECANISMOS INNATOS EXTERNOS
INFECCIONES DEL EPITELIO MUCOSO Barreras físicas, químicas y flora autóctona
3. MECANISMOS INTERNOS INFECCIÓN DE LOS TEJIDOS SUBEPITELIALES 3.1. DEFENSAS CONSTITUTIVAS DEL HOSPEDADOR Inflamación 3.2. DEFENSAS INDUCIBLES. LA RESPUESTA INMUNE. DEFENSAS ESPECÍFICAS O MECANISMOS ADQUIRIDOS Inmunidad humoral. Anticuerpos
Estructura Producción Genética de la producción Tolerancia del organismo a sus propios antígenos Reacción antígenoanticuerpo Complemento Complejo Mayor de Histocompatibilidad
Inmunidad celular Fases de la inmunidad celular Producción de linfocitos T
4. INMUNIDAD ACTIVA Y PASIVA. INMUNIDAD ARTIFICIAL Vacunas Sueroterapia Serovacunación
5. ALERGIAS Y AUTOINMUNIDAD 5.1. HIPERSENSIBILIDAD Inmediata Retardada 5.2. AUTOINMUNIDAD
6. EL CÁNCER Y LA RESPUESTA INMUNITARIA

1. INTRODUCCIÓN Para que una enfermedad infecciosa se desarrolle es necesario, no sólo una capacidad patógena por parte del microorganismo, sino también una predisposición o receptividad por parte del individuo atacado. Si éste ofrece una resistencia eficaz, la infección no se lleva a cabo y se dice que el organismo es inmune a dicha enfermedad. La inmunidad sería el estado de resistencia que presentan los organismos frente a la infección y frente a la aparición de tumores. En los vertebrados implica que los organismos diferencian lo propio de lo ajeno, es decir, reconocen todos sus tipos celulares. El sistema inmune es el responsable de conferir inmunidad. La ciencia que se ocupa de su estudio es la Inmunología. El sistema inmune Es un sistema complejo que se encuentra distribuido por todos los órganos y fluidos vasculares e intersticiales. Órganos especializados: médula ósea, bazo, timo y nódulos linfáticos. Células: linfocitos, macrófagos y granulocitos. Moléculas solubles: anticuerpos, linfocinas y complemento.

Las células y moléculas llegan a todos los tejidos por el torrente sanguíneo, pueden abandonarlo a través de las paredes de los capilares y pueden regresar a él por el sistema linfático. Funciones de los órganos linfoides
Primarios: origen, desarrollo y maduración de células del sistema inmune.
Médula ósea: origen de las células del sistema inmunológico. Timo: Maduración de linfocitos T.
Secundarios: las células inmunes maduras son activadas por los antígenos.
Adenoides, amígdalas y placas de Peyer: activación de linfocitos por los antígenos.
Ganglios linfáticos y bazo: activación de los linfocitos T y B.

Defensas del organismo frente a la infección Mecanismos innatos: Externos: barreras físicas, químicas y flora autóctona.
Internos: células fagocitarias (neutrófilos y macrófagos), células asesinas, interferón y complemento.
Mecanismos adquiridos: Celulares: linfocitos. Moleculares: anticuerpos.


2. DEFENSAS INESPECÍFICAS O MECANISMOS INNATOS EXTERNOS
2.1. INFECCIONES DEL EPITELIO MUCOSO
Están presentes en el organismo de forma natural. Conjunto de mecanismos que tienden a evitar la invasión de los microorganismos. Unos impiden la entrada del agente invasor y otros lo combaten una vez que ha penetrado.
2.1.1. Barreras físicas, químicas y flora autóctona La piel es infranqueable, la descamación continua, el pH ácido del sudor y la flora bacteriana normal impiden el asentamiento de microbios. Sólo cuando se producen heridas la piel puede ser traspasada. Casi todas las infecciones microbianas empiezan en las superficies de las membranas mucosas. Desde ahí pueden:progresar hasta tejidos subepiteliales
permanecer localizadas en la superficie. Los epitelios del tracto respiratorio, intestinal y vaginal llevan una densa capa de microorganismos o "flora normal" que constituye una fuerte barrera para el establecimiento de patógenos invasores por competencia con ellos. Las propias membranas mucosas secretan sustancias antimicrobianas. En el tracto respiratorio las células epiteliales agitan sus cilios barriendo la película de mucus y los microorganismos poco adheridos hacia las cavidades exteriores. El tabaco, por ejemplo, disminuye los movimientos ciliares.

3. MECANISMOS INTERNOS
3.1.INFECCION DE LOS TEJIDOS SUBEPITELIALES Una vez cruzado el epitelio mucoso, los microorganismos se encuentran otros mecanismos de defensa. Su poder invasor depende de su capacidad para multiplicarse en el ambiente de los tejidos y fluidos del hospedador y de su capacidad para superar las defensas. Los microorganismos invasores se han ido adaptando a desarrollarse en tejidos específicos del hospedador. Esto indica que hay diferencias entre tejidos en lo que se refiere al medio bioquímico que proporcionan.
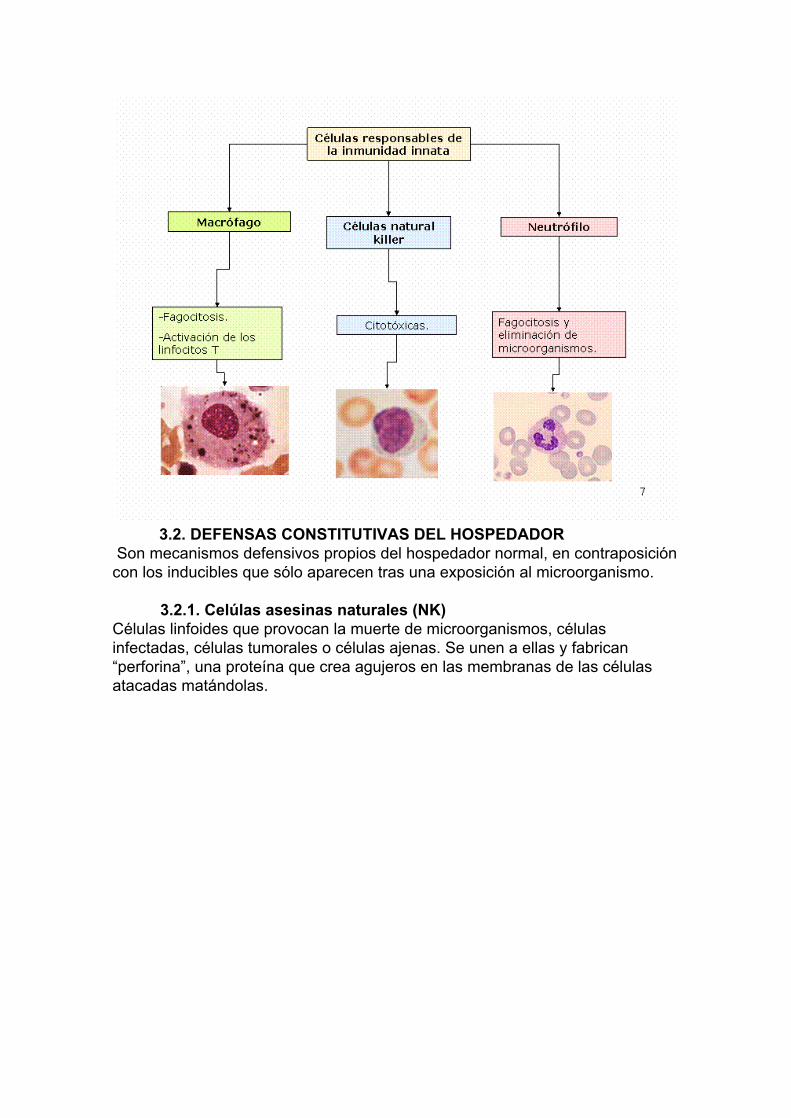
3.2. DEFENSAS CONSTITUTIVAS DEL HOSPEDADOR
Son mecanismos defensivos propios del hospedador normal, en contraposición con los inducibles que sólo aparecen tras una exposición al microorganismo.
3.2.1. Celúlas asesinas naturales (NK) Células linfoides que provocan la muerte de microorganismos, células infectadas, células tumorales o células ajenas. Se unen a ellas y fabrican “perforina”, una proteína que crea agujeros en las membranas de las células atacadas matándolas.

3.2.2. Inteferón Moléculas proteicas segregadas por células atacadas por virus que estimulan a las células adyacentes a sintetizar enzimas antivirales evitando la proliferación viral, inhibiendo la replicación del genoma vírico, inhibiendo la síntesis de proteínas o activando a las células NK para destruir a las células infectadas
3.2.3. Complemento Complejos macromoleculares de proteínas que circulan por la sangre. Activados por anticuerpos dan lugar a una cadena de reacciones que termina con la lisis de las células al adosarse a sus membranas. Pueden unirse a proteínas de la superficie bacteriana y atraer a los fagocitos que digerirán los microorganismos recubiertos por el complemento.
3.2.4. Células fagocitarias Rodean y digieren la materia particulada extraña. De los leucocitos, dos tipos son activamente fagocitarios: Monocitos
Granulocitos (neutrófilos) Se encuentran en la sangre y en los tejidos. Cuando tiene lugar una infección de un tejido se liberan sustancias quimiotácticas que atraen a estas células. Los monocitos circulantes en el torrente sanguíneo se transforman en macrófagos al migrar a los diferentes tejidos y órganos y, en su conjunto, se denomina sistema retículoendotelial. Los neutrófilos circulan por la sangre y salen de los vasos sanguíneos atraídos quimiotácticamente por sustancias que liberan los tejidos infectados.

La fagocitosis se realiza por fijación a la membrana e ingestión por medio de pseudópodos. La destrucción, por enzimas microbicidas de lisosomas.
3.2.5. Sustancias antimicrobianas En los tejidos y fluidos del cuerpo existen sustancias antimicrobianas como la lisozima, betalisina, etc.
3.3. Inflamación Cuando un tejido de un animal superior está sometido a una irritación, se inflama. En dicha zona los capilares están dilatados, provocando enrojecimiento. Las paredes de los capilares se hacen más porosas, de manera que ciertas proteínas solubles salen de los vasos y causan un movimiento osmótico de fluido hacia los tejidos, provocando hinchazón. La inflamación estaría causada por una sustancia liberada de las células dañadas o activadas en los fluidos del cuerpo. Las más estudiadas son la histamina y la serotonina. Si la inflamación ha sido iniciada por una infección bacteriana, los granulocitos se desplazan hacia el foco de la infección y liberan enzimas lisosómicos que dañan y destruyen células vecinas del tejido. Posteriormente son reemplazados por monocitos. También acuden linfocitos productores de anticuerpos (Ac).

La inflamación, como mecanismo defensivo, provoca las siguientes consecuencias: Fuerte aporte de leucocitos. Aporte de plasma e incremento de la concentración de factores antimicrobianos del suero y de Ac. Acumulación de células muertas del hospedador que liberan sustancias tisulares antimicrobianas. Acumulación de ácido láctico, creando condiciones adversas para el crecimiento de muchas bacterias. La elevada temperatura retrasa la multiplicación de algunos virus.
3.4. DEFENSAS INDUCIBLES: LA RESPUESTA INMUNE DEFENSAS ESPECÍFICAS O MECANISMOS ADQUIRIDOS
El contacto prolongado del hospedador vertebrado con el parásito microbiano conduce a la inducción de nuevas y específicas reacciones de defensa a las que se denomina respuesta inmunitaria. Al contrario que los mecanismos inespecíficos, que siempre están presentes, únicamente se desarrollan como respuesta a la invasión por un agente extraño concreto. Este sistema nos defiende específicamente de parásitos, órganos trasplantados, células cancerosas, microorganismos y sustancias tóxicas fabricadas por ellos. Existen dos clases: Inmunidad humoral: producción de Ac.
Inmunidad celular: producción de células sensibilizadas específicamente.

La respuesta inmunitaria reduce la toxigenicidad y el poder invasor del parásito hasta el punto que las defensas constitutivas pueden llegar a eliminarlo. Los individuos nacen con un sistema inmunológico capaz de responder ante lo propio y lo ajeno. Durante las primeras fases del desarrollo este sistema “aprende” a reconocer lo propio y esta capacidad se denomina tolerancia inmunológica; cuando esta tolerancia se pierde aparecen las enfermedades autoinmunes. En algunas enfermedades infecciosas, la respuesta inmunitaria en sí daña seriamente los tejidos del hospedador o es una respuesta frente a sustancias en principio inocuas; dicha reacción es conocida como hipersensibilidad (alergia). Las sustancias que se comportan como extrañas para el organismo y contra las cuales se desarrolla una repuesta inmune específica se denominan antígenos. Casi cualquier macromolécula (proteína o polisacárido más concretamente) con masa molecular de 5000 daltons o más puede desencadenar la respuesta inmunitaria, siempre que sea extraña al receptor. Los nódulos linfáticos actúan como filtro de la circulación a los microbios, partículas extrañas, restos titulares y células muertas. Contienen linfocitos y macrófagos y es en su interior donde ocurren las interacciones responsables de la respuesta inmune.
3.4.1. Inmunidad humoral Linfocitos
Se originan a partir de las células madre hematopoyéticas, células totipotentes que se fabrican en la médula ósea y dan lugar a eritrocitos, leucocitos y plaquetas. Los linfocitos maduran posteriormente en el timo (linfocitos T) o en la propia médula ósea (linfocitos B).

Anticuerpos a. Estructura de los anticuerpos
Los anticuerpos (Ac) son un grupo de proteínas globulínicas relacionadas, capaces de unirse específicamente, de manera no covalente, a las moléculas que inducen su formación. Las moléculas inductoras son los Ag. Prácticamente todas las proteínas son antigénicas, muchos polisacáridos y moléculas que sólo son antigénícas cuando se unen a proteínas. Para que una sustancia sea antigénica ha de cumplir: Tamaño molecular suficientemente grande. Complejidad molecular, con cadenas laterales que puedan ser reconocidas. Carácter extraño para el organismo productor de Ac. Hay varias clases de Ac., pero todos poseen ciertas características estructurales comunes. En el plasma humano hay 5 clases de inmunoglobulinas: IgG, IgA, IgM, IgD, IgE. Las IgG son las más abundantes y mejor conocidas. Contienen 4 cadenas polipeptídicas: 2 cadenas pesadas H, iguales entre sí 2 cadenas ligeras L, iguales entre sí. Están unidas por puentes disulfuro formando una Y flexible. Las cadenas pesadas poseen un oligosacárido unido covalentemente. Cada cadena tiene : 1 región de secuencia aminoacídica constante C

1 región de secuencia aminoacídica variable V Los sitios de unión del Ag son las porciones variables de las cadenas L y H. Allí también se distinguen regiones hipervariables, con elevada frecuencia de sustitución de aminoácidos. La región constante desencadena la respuesta celular. Al unirse el Ac al microorganismo por su región variable, se produce un cambio en la región constante, cambio detectado por los macrófagos que fagocitarán aquello que lleve anticuerpos pegados. Los Ac libres en la sangre no desencadenan la respuesta celuar. Los Ac tienen también una zona bisagra, que le permite adaptarse mejor para su unión óptima al Ag. Los brazos de la Y se escinden con papaína, dando dos fragmentos Fab y un fragmento Fc. Hay dos tipos de cadenas L: kappa
lambda En las cadenas H hay varios tipos antigénicos. Las regiones C y V de cada cadena se cree que están codificadas por genes diferentes. La hipótesis es que dos genes codifican una sola cadena polipeptídica. La reacción AgAc se asemeja la de enzimasustrato. Proyecciones del Ag encajan en cavidades del Ac. Estas proyecciones se llaman determinantes antigénicos y la porción exacta que encaja en el Ac se llama grupo hapténico. Las distintas zonas de las cadenas polipeptídicas del Ac se llaman dominios y tienen distintas funciones: Dominios variables: unión del Ag. 1º dominio constante: une el fragmento C4b del complemento. 2º dominio constante: fija el complemento y controla la velocidad catabólica. 3º dominio constante: se une al receptor Fc de macrófagos y monocitos. 2º y 3º dominio constante: se une al repceptro Fc de neutrófilos y células K.
b. Producción de anticuerpos Los Ac son producidos por linfocitos. Hay dos tipos de linfocitos: B: secretan activamente Ac. T: determinan la inmunidad celular y tienen un papel en el proceso por el cual los Ag inducen la síntesis de Ac en los linfocitos B. Durante la diferenciación de los linfocitos B sólo una dotación de genes para la síntesis de polipéptidos de Ig es activado en cada célula. Las diferentes células B producen diferentes Igs. Las sucesivas divisiones celulares de una célula activada dan lugar a un clon de células formadoras de un Ac específico.

Las células T sufren un proceso paralelo de diversificación. Cada clon contiene un receptor antigénico único en su membrana plasmática. Ambos tipos entran en circulación como linfocitos pequeños. Los linfocitos B pequeños apenas secretan Ac, pero sí están presentes sobre su membrana Ac de superficie. Sólo se dividen cuando el Ag específico se combina con el Ac de su membrana. Entonces la célula crece y se divide. Sus descendientes, grandes, son células plasmáticas con alta tasa de síntesis y secreción de Ac libres. Las células T también son estimuladas por el Ag y entre los productos que liberan algunos potencian la respuesta de las células B relacionadas y de otras células. El proceso de proliferación de clones específicos de linfocitos se llama selección clonal. Cuando la infección termina disminuye la producción de Ac. Muchos linfocitos agrandadados revierten al tipo pequeño. El individuo posee ahora clones de células con memoria capaces de producir, ante un próximo estímulo, Ac específicos mucho más rápidamente.
c. Genética de la producción de anticuerpos Se necesita que un número limitado de genes puedan fabricar alrededor de 100.106 Ac distintos. Los genes para producir Ac no existen como tales en el

genoma de las células germinales. Estas células contienen trozos y piezas de genes que se mezclarán en los linfocitos B cuando maduren. La mezcla de trozos de genes y la mutación amplía enormemente el nº de Ac posibles. En los descendientes maduros de cada línea celular se monta un único gen que se traducirá en un único Ac. La activación de genes durante el desarrollo se acompaña de recombinación somática. Cadenas ligeras: tres tipos de segmentos génicos: V (para la región variable): 150 C (para la región constante): 1 J (de unión entre ambas): 5 150 . 5 = 750 genes distintos. (Más diversidad en una región cercana al sitio de unión VJ donde el sitio de recombinación no está definido. Aumenta la diversidad 10 veces). Cadenas pesadas: en otro cromosoma y con potencial de diversidad mayor, ya que hay otro segmento (D). Hay 2,4 . 106 posibles cadenas pesadas. 2,4 . 106 . 7500 = 18000 . 106 Ac diferentes, a partir de 300 fragmentos génicos distintos. Además se producen mutaciones durante el desarrollo del linfocito.
d. Tolerancia del organismo a sus propios Ag. Teoría de la aborción clonal
Cuando un Ag entra en contacto con las células productoras de Ac mientras éstas están madurando, provoca la aborción de estas células. Así, Ag propios del organismo, causan aborción de células productoras de Ac contra ellos.
e. Reacción antígenoanticuerpo Es una reacción específica, aunque pueden ocurrir algunas reacciones cruzadas. La unión es firme, pero no de tipo covalente. Hay varios mecanismos mediante los cuales el Ac puede destruir o inactivar el Ag: Neutralización: Los viriones o toxinas bacterianas deben unirse a receptores de membrana para provocar daño en la célula del hospedador. La combinación con un Ac bloquea esta fijación a los receptores.

Opsonización: Normalmente el combinado AgAc es fagocitado por macrófagos y destruido. El Ac se fija a células fagocitarias que tienen receptores de Fc en su superficie.

Precipitado: Si las proporciones de AgAc son adecuadas, suelen formar una red o malla que precipita.
Aglutinación:
Lisis por complemento: La fijación de Ac hace a las bacterias G sensibles a la lisis por un conjunto de proteínas normales de la sangre llamadas complemento. Respuesta inflamatoria: La combinación AgAc desencadena una mayor

respuesta inflamatoria.
Complemento La lisis de las células bacterianas requiere un Ac específico y un componente no específico del suero normal, el complemento, que es un grupo de proteínas (en el suero humano son 11). Las bacterias G+ son resistentes a este mecanismo de defensa y aun no se sabe porqué. La reacción de fijación del complemento se ha estudiado en eritrocitos que han fijado Ac en su superficie. El Ac fijado es alterado de manera que fija el complemento C´1. C´1 no tiene actividad enzimática, pero una vez fijado se transforma y reacciona con C´4 rompiéndolo en fragmentos, y el C´4b se une al complejo. Ahora C´1 también rompe a C´2, fijándose el fragmento C´2a. Se van añadiendo componentes hasta que la membrana del eritrocito empieza a cubrirse de lesiones y lisa cuando se ha fijado C´8 y C´9. Tanto la fagocitosis como el aumento de la respuesta inflamatoria se ven favorecidos por la unión del complemento al complejo AgAc.
Complejo mayor de histocompatibilidad (MHC) Conjunto de proteínas responsables del rechazo en los trasplantes de órganos o tejidos. En los humanos se llama HLA. Son moléculas de las superficies celulares, distintas en cada individuo. Tienen dos funciones: Son el blanco del sistema inmunitario en caso de trasplante. En la infección presentan los Ag a los linfocitos B y T. Las moléculas HLA dependen de genes con alto número alélico, variando en cada individuo. Hay dos grupos de moléculas HLA: Clase I: en la superficie de todas las células del organismo. Clase II: sólo en los linfocitos B, macrófagos y algunos linfocitos T (linfocitos T DTA).




3.4.2. Inmunidad celular Los linfocitos T son los responsables de la inmunidad celular. Los linfocitos T no reconocen Ag solubles, sino fijados a una superficie celular. Los mecanismos de la inmunidad son fagocitosis y lisis por contacto.
Fases de la inmunidad celular El primer paso es la fagocitosis por macrófagos. La partícula digerida se fragmenta y el macrófago expone en su membrana algunos fragmentos antigénicos de la partícula unidos a moléculas del MHC. Así, los macrófagos tienen dos funciones: Inespecífica: fagocitar y secretar metabolitos antimicrobianos. Específica: presentar el Ag a otras células. Los linfocitos TH (auxiliares) se activan ante el Ag presentado por los macrófagos junto con el HLA clase II. La interacción con las células presentadoras de antígenos hace que se transformen en linfoblastos TH que liberan interleucinas (moléculas señalizadoras) que provocan activación y proliferación de linfocitos T citotóxicos y linfocitos B, y también aumenta la capacidad fagocítica de los macrófagos.

Los linfocitos T DTA al interaccionar con macrófagos infectados se transforman en linfoblastos TCDA, que producen sustancias químicas capaces

de transformar los macrófagos en células enfadadas, con gran capacidad fagocitaria. Los linfocitos TC (citotóxicos) se activan al interaccionar con macrófagos infectados , transformándose en linfoblastos TC que lisan la célula blanco que lleva el Ag. Mecanismo de lisis: Adhesión Exocitosis de enzimas que forman poros en la membrana de la célula blanco que acaba lisando. Muerte de la célula blanco y reciclaje de la célula TC. Los linfoblastos TC son capaces de destruir virus y células tumorales. También se conservan células citotóxicas de memoria. Los linfocitos TS (supresores) o Treg (células reguladoras) suprimen la respuesta inmune cuando ya no es necesaria. Evitan la autoinmunidad e influyen en la respuesta del sistema inmunitario ante agentes infecciosos, cáncer, trasplante de órganos y embarazo.
Producción de linfocitos T La diversidad de linfocitos T es similar a la de linfocitos B. Para generar esta diversidad los mecanismos son similares en ambos casos: genes partidos y recombinación. Cada clon de linfocitos T contine un receptor antigénico único en sus membranas.



4. INMUNIDAD ACTIVA Y PASIVA INMUNIDAD ARTIFICIAL
Inmunidad activa: adquirida tras superar una enfermedad (espontánea) o por medio de vacunas (artificial). Inmunidad pasiva: Ac que pasan de la madre al feto a través de la placenta o la leche materna (espontánea). Inmunidad que se adquiere al inyectar suero con Ac procedentes de un individuo que ha pasado la enfermedad a otro individuo (artificial).
4.1.INMUNIDAD ARTIFICIAL 4.1.1. Vacunas
Inmunización activa con carácter preventivo y, en algunos casos, tratamiento de ataque de algunas infecciones. Su objetivo es desencadena la producción de células inmunitarias de memoria. Vacunas preventivas: se obtienen por tratamiento físico o químico de gérmenes vivos o de extractos bacterianos. La inmunización se realiza en diversas etapas, necesitándose inoculaciones repetidas. La inmunidad se

mantiene por recuerdos. Vacunas vivas: se inocula el gérmen causal atenuada su virulencia, o mediante gérmenes próximos al patógeno capaces de crear inmunidad cruzada. Una única inoculación es duradera.
4.1.2. Sueroterapia Inmunización pasiva mediante el aporte de Ac. Su actividad es inmediata pero no permanente. Sueros heterólogos: de origen animal. Pueden provocar intolerancia y son de breve duración. Entre ellos están las antitoxinas, sueros para combatir el veneno de serpientes, escorpiones, arañas, etc. Sueros homólogos: de origen humano. Más efectivos pero difíciles de obtener.
4.1.3. Serovacunación Conjunto de medidas preventivas que combinan la vacunación con los tratamientos de sueros adecuados. El suero, con Ac, actúa en los primeros momentos de urgencia y, posteriormente, se desencadena la inmunidad activa producida por la vacuna. Se emplea en el tratamiento del tétanos, botulismo o rabia.
5. ALERGIAS Y AUTOINMUNIDAD

5.1. HIPERSENSIBILIDAD Los resultados patológicos de la sensibilización inmunitaria se llaman hipersensibilidad o alergia. El sistema inmunitario actúa de manera excesiva ante moléculas inocuas o poco peligrosas. La reacción se produce al 2º contacto con el alérgeno. Inmediata Los efectos nocivos se presentan a los pocos minutos del contacto con el alérgeno (Ag que provoca la hipersensibilidad). Está mediada por Ig. 1º contacto con el alérgeno: Captación por macrófagos Exposición de Ag por MHC Reconocimiento por linfocitos T Secreción de interleucinas Maduración de linfocitos B a células plasmáticas Producción de Ig que se unen a leucocitos del tejido conjuntivo (mastocitos) y sanguíneos (basófilos). 2º contacto: El alérgeno se une a las IgE adheridas a los mastocitos Liberación de histamina y otras sustancias responsables de los síntomas alérgicos Consecuencias: Shock anafiláctico con reacción inflamatoria excesiva (típica alergia al polen, heno, etc.)
Tracto respiratorio superior: estornudos y congestión nasal (rinitis alérgica). Tracto respiratorio inferior: constricción y obstrucción bronquial
(asma) Obstrucción de capilares pulmonares. Tracto gastrointestinal: nauseas, espasmos abdominales, diarrea, vómitos y hemorragias intestinales. Insuficiencia cardiaca (anafilaxis).
Retardada
Efectos nocivos al cabo de días u horas. Mediada por células. 1º contacto: Linfocitos T activados 2º contacto: Secreción de interleucinas Activación de macrófagos Los macrófagos se unen unos a otros en la zona afectada formando nódulos y liberando enzimas hidrolíticos que destruyen los tejidos (por ejemplo las lesiones que se formen en torno a los bacilos tuberculosos).

5.2. AUTOINMUNIDAD Las enfermedades de autoinmunidad se dan cuando los linfocitos responden ante elementos del propio cuerpo. Pueden afectar a cualquier órgano, si bien algunos se ven afectados con más frecuencia que otros, por ejemplo:
La sustancia blanca del cerebro y de la médula espinal en la esclerosis múltiple.
Los revestimientos de las articulaciones en la artritis reumatoide. Las células secretotas de insulina en la diabetes mellitas juvenil. Destrucción de las conexiones entre nervios y músculos en la
miastenia gravis. Producción de exceso de hormona tiroidea en la enfermedad de
Graves. Producción de ampollas en la piel en el pénfigo vulgar. Destrucción de los riñones y otros órganos en el lupus eritematoso
sistémico.
REGULACIÓN DE LA AUTOINMUNIDAD (Investigación y Ciencia, diciembre 2006) Enfermedades como la esclerosis múltiple, diabetes dependiente de insulina, artritis reumatoide, derivan de un comportamiento rebelde de los linfocitos T CD4+. Las células T reguladoras (subpoblación de las CD4+) bloquean el comportamiento aberrante del

sistema inmunitario: evitan la autoinmunidad e influyen en la respuesta del SI (sistema inmunitartio) ante agentes infecciosos, cáncer, trasplante de órganos y embarazo. ¿Cómo se consigue la autotolerancia (no atacar órganos del propio individuo)? En el timo las células inmaduras se programan para no reaccionar con intensidad contra tejidos del organismo. Las células rebeldes son destruidas, pero hay células autoagresivas que se salvan. Los vasos sanguíneos y linfáticos constituyen una 2º barrera protectora, mediante patrullas de células Treg. Tejidos como el cerebro y la médula espinal tienen escasos vasos sanguíneos y linfáticos, ocultas al SI las células autorreactivas hallan una vía de entrada si el tejido sufre una lesión. En los años 60 se propuso la existencia de una población de células T capaz de sofocar la respuesta inmunitaria y se las denominó células T supresoras, pero nadie logró localizarlas. En 1995 se identificó un marcador de células TS, la molécula CD25. Al eliminar las células T CD4+ que tenían esta molécula, varios órganos de ratones eran objeto de un ataque autoinmunitario. A estas células T CD4+ y CD25 se las llamó T reguladoras CD25+ o Treg. Las Treg reprimen una amplia gama de células del SI e impiden su multiplicación y la secreción de señales químicas intercelulares (citoquinas). Las Treg se activan por contacto directo célulacélula. Contienen alta concentración de molécula intracelular Foxp3 (es un factor de transcripción, es decir, regula la actividad de determinados genes y controla la síntesis de proteínas codificadas por estos genes). Foxp3 convierte células T en desarrollo en células Treg. En ratones privados de Treg, el SI también atacaba a la población bacteriana intestinal, provocando daños en el intestino. La transferencia de Treg evitaba la enfermedad intestinal. Pero también parece que el fracaso en la erradicación de Helicobacter pylori (causante de úlcera de estómago) podría guardar relación con el bloqueo de la respuesta celular inmunitaria por parte de las Treg. Dejar que sobrevivan algunos organismos invasores no tiene por qué ser malo. Permitiendo la permanencia de un número reducido de parásitos en ratones, la reinfección desencadena una respuesta rápida y eficaz. Sin Treg el parásito es eliminado por completo, pero ante la reinfección la respuesta es ineficaz. Treg debe, por tanto, contribuir al mantenimiento de la memoria inmunitaia. Treg es importante en la protección de los embarazos. El trofoblasto (tejido placentario que adhiere el feto a la pared del útero) produce moléculas inmunosupresoras. El propio SI inmunitario de la madre sufre cambios, con un aumento de la actividad de las Treg. La ausencia de Treg provoca el rechazo del feto. El uso de las Treg sería un gran aliado terapéutico ante una amplia gama de trastornos:
Intensificar la actividad de las Treg para luchar contra: enfermedades autoinmunitarias alergias
rechazo de órganos trasplantados Eliminación selectiva de las Treg para reforzar respuestas inmunitarias beneficiosas
(sólo aquellas Treg que bloquean una respuesta inmunitaria útil) tratamiento de tuberculosis, sida, etc. tratamiento de cáncer (las Treg dificultan la vigilancia de las células inmunitarias que detectan el tránsito de una célula normal a cancerosa)
Desafíos técnicos: Saber qué tipos de Treg hay que eliminar Conseguir grandes poblaciones de Treg fuera del organismo Conseguir células Treg mediante transferencia del gen Foxp3 a otras células T.
Identificar los procesos que activan la producción de la proteína Foxp3 Para órganos trasplantados: Extracción de Treg del receptor y clutivarlas en
presencia de células del donante, para que se multipliquen las Treg con mayor capacidad de suprimir el rechazo. El SI queda intacto para repler la invasión de microorganismos.

6. EL CÁNCER Y LA RESPUESTA INMUNITARIA Las células cancerígenas tienen Ag en su superficie celular que difieren de los Ag de las células normales y pueden ser identificadas como extrañas por lo que el organismo puede organizar una respuesta inmunitaria. Parece que esta respuesta podría suprimir las células cancerígenas mucho antes de que se detecte el cáncer. Los cánceres que se desarrollan representarían fallos ocasionales del sistema inmunitario. Reforzar a respuesta inmunitaria podría ser un avance en la lucha contra el cáncer.