inhibitors: morphactin
TRANSCRIPT

Plant Physiol. (1975) 55, 1082-1086
Auxin and the Response of Pea Roots to Auxin TransportInhibitors: Morphactin
Received for publication September 12, 1974 and in revised form January 22, 1975
DOUGLAS H. GAITHER1Vegetation Control Division, Fort Detrick, Frederick, Maryland 21701
ABSTRACT
The auxin transport inhibitor methyl-2-chloro-9-hydroxy-fluorene-9-carboxylate (CFM), a morphactin, inhibits negativegeotropism, causes cellular swelling, and induces root hairformation in roots of intact Pisum sativum L. seedlings. Inexcised pea root tips, CFM inhibits elongation more than in-crease in fresh weight (swell ratio = 1.3 at 20 /Am CFM). CFMgrowth inhibition was expressed in the presence of ethylene.Indoleacetic acid (IAA) prevented the expression of CFMgrowth inhibition possibly because IAA inhibited the accumula-tion of CFM into the tissue sections. CFM inhibited theaccumulation of IAA and 2,4-dichlorophenoxyacetic acid intoexcised root tips. Applying Leopold's (1963. Brookhaven Symp.Biol. 16: 213-234) model for polar auxin transport, thisresult suggests a possible explanation for CFM inhibition ofgeotropism in pea roots, i.e. disruption of auxin transport byinterfering with auxin binding.
Work with suspensions of intact cells (19) and work withparticulate fractions from homogenates of shoot tissue (14-16,23-25) indicates that some chemicals may inhibit auxin trans-port by interfering with auxin binding at auxin-specific receptorsites. CFM2 effects on geotropism, phototropism, apical dom-inance, and hypocotyl curvature correlate with the ability ofthis morphactin to inhibit auxin transport (6, 7, 18, 20, 21, 27).It occurred to me that a simple explanation for the CFMalteration of root geotropism might involve a mechanismwhereby CFM interferes with auxin binding. The experimentsreported in this paper indicate that this hypothesis may be truefor pea roots.
MATERIALS AND METHODS
Plant Material. Seeds of Pisum sativum L. var. Alaska wereimbibed for 6 hr, sown in moist vermiculite, and grown in thedark at 23 C.
Studies with Intact Seedlings. Two-day-old seedlings (roots2-3 cm long) were soaked in water or 20 iM CFM for 5 minthen placed horizontally in Petri dishes containing vermiculitemoistened with the treatment solution. The experiment was
1 Present address: 4597 Knight Cove, Memphis, Tenn. 38118.2Abbreviations: CFM: methyl-2-chloro-9-hydroxyfluorene-9-
carboxylate; DPX 1840: 3, 3a-dihydro-2-(p-methoxyphenyl)-8H-pyrazolo [5, I-a]isoindol-8-one; NPA: N-1-naphthylphthalamic acid;TIBA: 2,3, 5-triiodobenzoic acid.
then conducted in 10-liter desiccators as described elsewhere(8). Downward curvature of the root tips was measured atdifferent time periods. After 24 hr, photomicrographs weremade of freehand longitudinal sections taken from the elonga-tion zones of the roots.Growth Studies and Ethylene Evolution. Ten terminal 5-mm
sections from 3-day-old seedlings (roots 5-6 cm long) were in-cubated in sealed 125-ml Erlenmeyer flasks as described pre-viously (8) except that 6 ml of an incubation medium (pH 5.2)containing 5 mm dibasic potassium phosphate, 2 mm citricacid, 1 mm Ca(NO,)2- 4H20, 1% sucrose (w/v), and appropriateconcentrations of CFM or IAA were used. Ethylene evolutionwas assayed by gas chromatography using a flame ionizationdetector (1).
Studies with '4C-CFM and "4C Auxins. Tissue sections wereincubated as described above, except that the buffer also con-tained 9-'4C-CFM (10.4 mCi/mmole, Cela Merck, Ingelheim,Germany), 1-14C-IAA (8.9 mCi/mmole, New England Nu-clear), or I-"4C-2,4-D (3.03 mCi/mmole, Tracerlab). Radio-activity in the incubation medium was determined by countingan aliquot by liquid scintillation. After a given incubationperiod, radioactivity in the gas phase was determined by bub-bling a 10-ml gas sample into 3 ml of NCS tissue solubilizer,then counting by liquid scintillation. The solution was thenfiltered, and the tissue was rinsed with ice water. After blottingand weighing, the sections were placed in a scintillation vialcontaining 2 ml of NCS tissue solubilizer. The vial was shakenat 45 C at least 4 hr. The radioactivity which had accumulatedin the sections was then determined by liquid scintillationcounting.
Metabolism of '4C-CFM was studied by homogenizing rinsedroots at 4 C in 5 ml of 50 mm potassium phosphate buffer, pH6.8. The brei was extracted twice with 10 ml of chloroform.After evaporation of the chloroform under reduced pressure,the residue was dissolved in 50 kdI of acetone. This residue and4C-CFM were co-chromatographed separately with unlabeledCFM on an Eastman silica gel sheet. After development inbenzene-glacial acetic acid (4:1, v/v), the chromatogram wascut into 1-cm strips. Each strip was placed directly into a vialfor counting of the radioactivity by liquid scintillation.
RESULTS
CFM rapidly inhibited the normal geotropic response inroots of intact pea seedlings (Table I; 20). By 24 hr, the elonga-tion zone of the CFM-treated root showed swelling of theepidermal and cortical cells and many long root hairs (Fig. 1).
Dose-response data for growth of excised pea root tips(Fig. 2) showed that CFM inhibits elongation to a greater ex-tent than increase in fresh weight. The increase in weight toincrease in length ratio is 1.3 at 20 /M CFM (Fig. 2).
Application of auxin to pea roots increases ethylene pro-1082
www.plantphysiol.orgon April 11, 2018 - Published by Downloaded from Copyright © 1975 American Society of Plant Biologists. All rights reserved.

AUXIN, MORPHACTIN, AND PEA ROOTS
CONTROL CFM
FIG. 1. Freehand longitudinal sections from the elongation zone of roots from pea seedlings exposed for 24 hr to water or 20 ,uM CFM. Seed-lings were 2 days old when initially treated. Magnification is the same for both photomicrographs.
duction (8, 9). Since CFM-induced growth inhibition was ex-pressed (Fig. 2) in the presence of an ethylene concentrationwhich is supraoptimal for growth inhibition (8), synergisticgrowth inhibition was expected in sections treated with CFM +IAA. Synergistic growth inhibition occurred only for lengthgrowth at 1 btM IAA and the two lowest CFM concentrations(Fig. 2). A mutual antagonism between all other CFM andIAA combinations seemed apparent for elongation (Fig. 2).IAA completely prevented CFM-induced fresh weight in-hibition (Fig. 2).The growth curve for CFM-treated roots was sigmoidal
(Fig. 3); complete growth rate inhibition did not occur untilafter 12 hr. The pattern of IAA-induced growth inhibition(Fig. 3) was similar to that reported previously (9). In rootstreated with CFM + IAA (Fig. 3), IAA controlled weightgrowth at all times and length growth for the first 12 hr. Be-tween 16 and 24 hr, the elongation rate of roots treated withCFM + IAA exceeded the control elongation rate (Fig. 3).Thus, absolute elongation of roots subjected to the simulta-neous treatment was greater than elongation of roots treatedwith CFM or IAA alone (Fig. 3). Figure 3 also shows that atno time did CFM increase ethylene production nor did CFMaffect IAA-induced ethylene production.The growth rate reduction caused by CFM after 12 hr (Fig.
3) might be due to tissue transformation of CFM into someother entity or possibly to slow tissue uptake of CFM. CFMremained unchanged during the 24-hr incubation period (Fig.4). Presumably the initial rapid uptake of "4C-CFM (Fig. 5)represents uptake into the free space. '4C-CFM uptake beganto level off after 2 hr (Fig. 5).IAA had no effect on 14C-CFM metabolism (Fig. 4). After
the first hour, accumulation of "C-CFM in pea roots was in-hibited by IAA (Fig. 5) concentrations as low as 0.1 fiM (curveA, Fig. 6). This inhibition occurred in the presence of asaturating dose of unlabeled CFM (experiment B, Table II).In a 3-hr experiment, CFM also inhibited the accumulation of4C-IAA in roots (Table III). Difficulties arise, however, inlong term accumulation experiments using carboxyl-labeledIAA since much of the applied IAA is conjugated with asparticacid or is decarboxylated (3, 9). 2,4-D remains intact in 24-hrexperiments with pea roots (3); therefore, "4C-2,4-D was theradioactive auxin used for further accumulation studies. CFM
Table I. Time Course for CFM Inihibitiont of Geotropism in Rootsof Ijitact Pea Seedlings
Degrees Negative Curvature'Treatment
I hr 4 hr
Control 7.8 d 2.7 26.4 i 1.8CFM, 20,uM -3.5 i 1.6 -8.2 + 3.2
1 Mean ± confidence limit (P < 5%,-c) from 24 seedlings pertreatment.
20 0uM CFM
FIG. 2. Effect of IAA and ethylene on CFM-induced inhibitionof elongation and fresh weight increase in pea root tips. Incubationtime was 24 hr. Controls grew 145% in length and 180% in freshweight.
concentrations as low as 0.01 fiM inhibited accumulation of4C-2,4-D in pea roots (curve B, Fig. 6). This inhibition oc-curred in the presence of a saturating dose of unlabeled 2,4-D(experiment A, Table II). Ethylene (1000 pl/l-0.1 ,ull), ABA(100 jtM-1 /iM), BA (10 itM), GA3 (100 ftM), and the auxintransport inhibitors (5, 10) TIBA (50 ,uM-10 ,uM) and DPX1840(10 [pM) had no effect on 14C-CFM accumulation (data notshown). NPA, another auxin transport inhibitor, did inhibit"C-CFM accumulation (Gaither and Abeles, in preparation).
I
1083Plant Physiol. Vol. 55, 1975
www.plantphysiol.orgon April 11, 2018 - Published by Downloaded from Copyright © 1975 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 55, 1975
TIME, hr
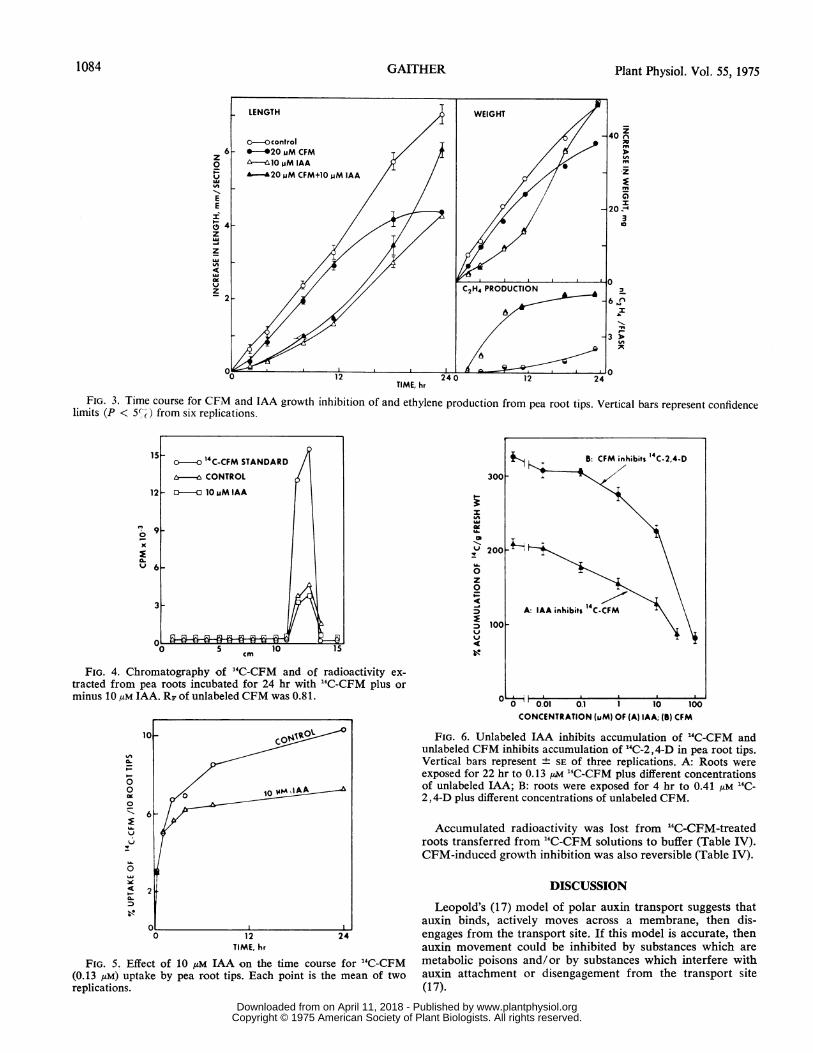
FIG. 3. Time course for CFM and IAA growth inhibition of and ethylene production from pea root tips. Vertical bars represent confidencelimits (P < 5%-) from six replications.
'9
63
0 5 10 15cm
FIG. 4. Chromatography of "C-CFM and of radioactivity ex-tracted from pea roots incubated for 24 hr with "C-CFM plus orminus 10 ,uM IAA. RF of unlabeled CFM was 0.81.
10 _ _
VN0
0,v -
2
0 12 24TIME, hr
FIG. 5. Effect of 10 AmM LAAon the time course for '4C-CFM(0.13 ,M) uptake by pea root tips. Each point is the mean of tworeplications.
3001
z
-
v 200i
0z0I-
E 100vvI.
50
o '' 0.01 0.1 I 10 100CONCENTRATION (uM) OF (A) IAA; (B) CFM
FIG. 6. Unlabeled IAA inhibits accumulation of 14C-CFM andunlabeled CFM inhibits accumulation of 14C-2 ,4-D in pea root tips.Vertical bars represent ± SE of three replications. A: Roots wereexposed for 22 hr to 0.13 pm '4C-CFM plus different concentrationsof unlabeled IAA; B: roots were exposed for 4 hr to 0.41 MM 14C-2, 4-D plus different concentrations of unlabeled CFM.
Accumulated radioactivity was lost from "4C-CFM-treatedroots transferred from "C-CFM solutions to buffer (Table IV).CFM-induced growth inhibition was also reversible (Table IV).
DISCUSSION
Leopold's (17) model of polar auxin transport suggests thatauxin binds, actively moves across a membrane, then dis-engages from the transport site. If this model is accurate, thenauxin movement could be inhibited by substances which aremetabolic poisons and/or by substances which interfere withauxin attachment or disengagement from the transport site(17).
1084 GAITHER
www.plantphysiol.orgon April 11, 2018 - Published by Downloaded from Copyright © 1975 American Society of Plant Biologists. All rights reserved.

AUXIN, MORPHACTIN, AND PEA ROOTS
Table II. CFM Inhibits Accumulationi of 14C-2,4-D (Experimelnt A)antd IAA Inihibits Accumulationt of 14C-CFM (Experiment B)Pea root tips (three replications) were treated with the appro-
priate 14C compound plus saturating doses of the unlabeledchemicals, either singularly or in combination.were (A) 16 hr and (B) 22 hr.
Incubation times
ExperimtFresht '4C AccumulatedExperiment FehAt In Roots2
mg % applied dpmA. 0.41 ,uM 14C-2,4-DControl 36.2 + 0.7 11.79 4 3.57100 MM2,4-D 34.5 4 1.1 1.51 ± 0.22100,/M CFM 41.3 i 0.7 2.42 i 0.34100 ,uM 2,4-D + 100 ,uM 32.9 4 0.9 0.88 4 0.13CFM
B. 0.79 AM 14C-CFMControl 56.7 ± 0.8 10.45 i 1.46100yM CFM 49.1 ± 0.7 1.96 ± 0.09100 AM IAA 29.6 ± 0.9 2.56 ± 0.30100 ,M CFM + 100 ;M 29.6 ± 0.6 0.47± 0.01IAA
1 Mean i SE.2 Mean + confidence limit (P < 5%1-).
I have not demonstrated a direct inhibition of auxin trans-port in pea roots by CFM. However, I observed, as others have(6, 20, 21, 27), that CFM inhibits root geotropism (Table I),a response thought dependent upon the root possessing afunctional auxin transport system (22).CFM does not inhibit respiration in several test systems (20,
21, 27); therefore, Leopold's (17) model predicts that CFMis not interfering with active auxin movement across a mem-brane. Thus, CFM may be interfering with auxin transport atthe level of the auxin transport site. Indeed CFM did displaceauxin from pea root tip sections (Fig. 6, Table III). Auxin alsodisplaced CFM (Figs. 5 and 6).To test the idea that an interference with binding might ex-
plain the CFM inhibition of auxin accumulation, experimentA of Table II was performed. I assumed that the presence of asaturating concentration (100 ,uM) of unlabeled 2,4-D wouldallow "4C-2, 4-D to accumulate in the tissue only where asteady state exists between '4C-2,4-D and unlabeled 2,4-D,i.e. in tissue areas specific for the auxin. Unlabeled 2,4-D didinhibit 14C-2 ,4-D accumulation by 87%. In the presence ofsaturating doses of the auxin and CFM, 14C-2,4-D accumula-tion was reduced further by 6%. This figure represents CFMdisplacement of 2,4-D from tissue areas specific for the auxin.The tissue areas in question could be auxin binding sites.
Using a similar argument, auxin may be interfering withCFM binding. A saturating dose of unlabeled CFM displaced81% of the 14C-CFM applied to excised tissue sections (experi-ment B, Table II). When a saturating dose of IAA was alsopresent, 14C-CFM displacement was increased by 14% (experi-ment B, Table II).
It seems likely that the site of primary auxin action in rootsis located at the cell wall or the plasma membrane (3, 22).Primary CFM action in pea roots may also occur at the cellsurface. Two observations support this conclusion. First, ac-cumulated CFM exists in an exchangeable, diffusable statewhich probably accounts for the reversibility of CFM-inducedgrowth inhibition (Table IV). Second, CFM causes rapid in-hibition of geotropism in intact roots (Table I) and rapid in-hibition of growth in excised root tips (Fig. 3).
In time course studies, CFM showed two different effects on
growth (Fig. 3). The first effect was a rapid inhibition fromwhich the tissue was able to recover (Fig. 3). No growth raterecovery occurred from the second effect which was not ex-pressed until after 12 hr (Fig. 3). This pattern of CFM-inducedgrowth inhibition was not related to altered metabolism (Fig.4) or uptake (Fig. 5) of CFM.A suspected antagonism between CFM and IAA for elonga-
tion (Fig. 2) was not confirmed by time course studies (Fig. 3).Prior to 12 hr, elongation of sections treated with CFM + IAAwas controlled by IAA-regulated processes, in this case prob-ably ethylene production (Fig. 3) (8, 9). Ethylene has no effecton CFM accumulation (data not shown). IAA does not affectCFM metabolism (Fig. 4). Lack of expression of CFM-inducedgrowth inhibition in the presence of IAA (Fig. 3) might berelated to IAA reducing the level of CFM within the tissue(Figs. 5 and 6).
Tissue sections treated with CFM + IAA show an amplifica-tion of recovery from IAA-induced inhibition of elongation(Fig. 3). This response cannot be related to any known physio-logical process, but other workers (26) have reported a similarstimulation of root growth in seedlings which have been treatedwith auxin then transferred to an auxin-free growth medium.Aberg (2) has discussed this phenomenon of auxin-treated rootsdisplaying positive after-effects.
Table III. Effect of CFM oni 14C-IAA (0.14 pixf) Accumulationand Destruction by Pea Root Tips
Means are based on three replications. Incubation time was 3 hr.In each column, means followed by the same letter are not signifi-cantly different (Student's t test).
RadioactivityTreatment Fresh Wt
Tissuel Solution2 Gas Phase
mg % applied dpmtControl 32.6a 13.3b 40.3d 40.4fCFM, 100M 32.3a 9.9c 42.1 e 41.Of
Significant at the 0.1%o level.2 Significant at the 5%c level.
Table IV. Time Course Relating Reversibility of CFM ElongationInihibition to Reversibility of CFM Accumulationi
For the elongation inhibition experiment, root tips were treatedwith 20 Mm unlabeled CFM. For the accumulation experiment,root tips were treated with 0.13 AM '4C-CFM. Mean ± SE basedon three replications.
Experiment
Measured at 4 hrControlCFM
Measured at 16 hrControlCFM
Measured at 24 hrControlCFMCFM 4 hr, then buffer 20 hrCFM 16 hr, then buffer 8 hr
14C-CFMIElongation Accumulated
In Roots
mm %C applied dpm
1.10 + 0.050.70 ± 0.03 8.50 ± 0.30
4.60 ± 0.203.20 ±t 0.09
7.104.706.205.30
± 0.32± 0.18± 0.30± 0.25
12.93 ± 0.04
13.75 ± 0.302.00 ± 0.1016.25 ± 0.102
1 Buffer contained 7.10 ± 0.20% of applied dpm.2 Buffer contained 6.00 ± 0.40% of applied dpm.
Plant Physiol. Vol. 55, 1975 1085
www.plantphysiol.orgon April 11, 2018 - Published by Downloaded from Copyright © 1975 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 55, 1975
No increased ethylene production was detectable from ex-cised root tips treated with CFM (Fig. 3). Thus, in this tissue,CFM or ethylene can separately cause cellular swelling, roothair induction (Fig. 1; refs. 1, 8), inhibition of geotropism(Table I; refs. 1, 8, 9), and inhibition of cell division (4, 27).
Polar auxin transport has been studied mainly in coleoptiles(10). From this work the concept has arisen that auxin trans-port and primary auxin action may have a common initial step(11, 13), namely auxin binding at auxin-specific receptor sites(12, 14). In pea roots, I have shown that CFM could be in-hibiting auxin transport at the level of auxin binding; yet, CFMdid not alter the pattern of fresh weight increase or ethyleneproduction in the presence of auxin (Fig. 3). This result in-dicates that in this tissue, auxin transport may not be directlyrelated to primary auxin action.
Acknouledgmeot-I thank Donald H. Lutz for performing superb technicalassistance, Frederick B. Abeles for proffering many salient ideas, R. C. Frenchiand R. A. Darrow for their interest in this work, J. P. Sterrett for help withliquid scintillation counting techniques, D. H. "Buster" Saucier, Jr. and A. T.McManus for graphic art assistance, S. Gaither for emending the manuscript,Cela Meck, Ingelheim, Germany, for CFM, EMD-IT 3456, and 14C-CFM, Uni-Royal, Inc. for N-PA, and C. W. Bingeman of E. I. du Pont de Nemours andCo., for DPX1840.
LITERATURE CITED
1. ABELES, F. B. 1973. Ethylene in Plant Biology. Academic Press, New York.2. ABERG, B. 1957. Auxin relations in roots. Annu. Rev. Plant Physiol. 8: 153-180.3. A-NDREAE, W. A. 1967. Uptake and metabolism of indole acetic acid,
naphthaleneacetic acid, and 2,4-dichlorophenoxyacetic acid by pea rootsegments in relation to growth inhibition during and after auxin applica-tion. Can. J. Bot. 45: 737-753.
4. APELBAUNI, A. AND S. P. BURG. 1972. Effect of ethylene on cell division anddeoxyribonucleic acid synthesis in Pisum sativum. Plant Physiol. 50: 117-124.
5. BEYER, E. 'M., JR. 1972. Auxin transport: a new synthetic inhibitor PlantPhysiol. 50: 322-327.
6. Bopp, Al. 1972. On the effect of morphactin. In: H. Kaldewey and Y. Vardar,eds., Hormonal Regulation in Plant Growth and Development. VerlagChemie, WVeinheim, Germany. pp. 333-348.
7. BRIDGES, I. G. AND M. B. WILKINS. 1973. Effects of morphactin on indol-3yl-acetic acid transport, growth, and geotropic response in cereal coleoptiles.J. Exp. Bot. 81: 711-723.
8. CHADWICK, A. V'. AN-D S. P. BURG. 1967. An explanation of the inhibition ofroot growth caused by indole-3-acetic acid. Plant Physiol. 42: 415-420.
9 CHADWICK, A. V. AND S. P. BURG. 1970. Regulation of root growth by auxin-ethylene interaction. Plant Physiol. 45: 192-200.
10. GOLDSMITH, MI. H. M1. 1968. The transport of auxin. Annu. Rev. PlantPhysiol. 19: 347-360.
11. HERTEL, R., M. L. EVANS, A. C. LEOPOLD, AND H. M. SELL. 1969. Thespecificity of the auxin transport system. Planta 85: 238-249.
12. HERTEL, R. AND R. FLORY. 1968. Auxin movement in corn coleoptiles. Planta82: 123-144.
13. HERTEL, A. AND A. C. LEOPOLD. 1963. V ersuche zur Analyse des Auxin-transports in der Koleoptile von Zea mays L. Planta 59: 535-562.
14. HERTEL, R., K.-S. THOMSON, AND V. E. A. Russo. 1972. In vitro auxin bind-ing to particulate cell fractions from corn coleoptiles. Planta 107: 325-340.
15. JABLANOVIC, NM. AND L. D. NOODEN. 1974. Changes in competible IAA bindingin relation to bud development in pea seedlings. Plant Cell Physiol. 15:687-692.
16. LE.MBI, C. A., D. J. MIORRt, K.-S. THo.IsoN, AND R. HERTEL. 1971. N-1-N-aphthylphthalamic-acid-binding activity of a plasma ml1embrane-richfraction from maize coleoptiles. Planta 99: 37-45.
17. LEOPOLD, A. C. 1963. The polarity of auxin transport. Brookha-en Symp.Biol. 16: 218-234.
18. -NAQVI, S. MI. 1972. The effect of morphactin on the kinetics of indol-3yl-acetic acid-2-14C transport in Zea mays L. coleoptile segments. J. Exp.Bot. 23: 763-767.
19. RUBERY, P. H. AND A. R. SHELDRAKE. 1974. Carrier-mediated auxin transport.Planta 118: 101-121.
20. SCHNEIDER, G. 1970. 'Morphactins: physiology and peiformance. Annu. Rev.Plant Physiol. 21: 499-536.
21. SCHNEIDER, G. 1972. 'Morphactins and plant growth regulation. In: H.Kaldewey and Y. V'ardar, eds., Hormonal Regulation in Plant Growthand Development. Verlag Chemie, Weinheim, Germany. pp. 317-331.
22. SCOTT, T. K. 1972. Auxin and roots. Annu. Rev. Plant Physiol. 23: 235-258.23. THOMSON, K.-S. 1972. The binding of 1-N-naphthylphthalamic acid (NPA),
an inhibitor of auxin transport, to particulate fractions of corn coleoptiles.In: H. Kaldewey and Y. Vardar, eds., Hormonal Regulation in PlantGrowth and Development. Verlag Chemie, Weinheim, Germany. pp. 83-88.
24. THOMSON, K.-S., R. HERTEL, S. MULLER, AND J. E. TAVARES. 1973. 1-N-Naphthylphthalamic acid and 2,3,5-triiodobenzoic acid: in vitro bindingto particulate cell fractions and action on auxin transport in corn coleoptiles.Planta 109: 337-352.
25. THOMSON, K.-S. AND A. C. LEOPOLD. 1974. In vitro binding of morphactinsand 1-N-naphthylphthalamic acid in corn coleoptiles and their effects on
auxin transport. Planta 115: 259-270.26. THIMANN, K. V. AND R. H. LANE. 1938. After-effects of the treatment of seed
with auxin. Am. J. Bot. 25: 535-543.27. ZIEGLER, H. 1970. Morphactins. Endeavour 19: 112-116.
I1086 GAITHER
www.plantphysiol.orgon April 11, 2018 - Published by Downloaded from Copyright © 1975 American Society of Plant Biologists. All rights reserved.