informe tcnico final del proyecto de...
TRANSCRIPT

INFORME TÉCNICO FINAL DEL PROYECTO DE INVESTIGACIÓN “POSIBLE PARTICIPACIÓN DEL ÓXIDO NÍTRICO EN EL ESTRÉS OXIDATÍVO GENERADO
DURANTE LOS ESTADOS CONVULSIVOS”
PARTICIPANTES: DRA. LOURDES A. VEGA RASGADO, DR. GUILLERMO CEBALLOS REYES Y DR. FERNANDO VEGA DÍAZ
CLAVE CGPI: 20080742
RESUMEN Se ha postulado que el óxido nítrico (NO) como un radical libre que produce estrés oxidativo, podría
tener un importante papel en la etiología de diversas enfermedades neurológicas. Metabólicamente, el NO se forma a partir de la L‐arginina (L‐Arg), reacción catalizada por la óxido nítrico sintasa (NOS). Además, el NO es considerado como un neurotransmisor en el cerebro. En este trabajo se estudia la posible participación del NO en el mecanismo de las convulsiones a través del estrés oxidativo.
Con esta finalidad, se estudia el efecto de drogas convulsionantes (metrazol, tiosemicarbazida, estricnina y bicuculina), anticonvulsionantes (amino oxiacético, valproato, diazepan y gabapentina) y antioxidantes (vitaminas C y E) sobre la generación del estrés oxidativo en cerebro de ratón, determinando la formación de carbonilos libres (método de Dalle y Rossi) y ditirosinas (método de Malencik y Cols), así como sobre los niveles de NO, determinado de forma indirecta como la cantidad de nitratos transformada en nitritos (método basado en la reacción de Griess).
Los resultados obtenidos indican que todos los convulsionantes empleados produjeron estrés oxidativo de diferente magnitud. En el caso del metrazol (PTZ), la formación de carbonilos libres es dosis‐dependiente. Contrariamente, los anticonvulsionantes no producen estrés oxidativo, excepto el amino oxiacético (AAOA). Cuando los ratones se protegen de la mortalidad inducida por los convulsionantes con AAOA, se consigue revertir el aumento en el estrés oxidativo producido por los mismos. También es posible antagonizar el efecto del PTZ cuando se protege a los animales con los anticonvulsionantes. Los antioxidantes también disminuyeron el estrés oxidativo y revertir el efecto del PTZ (aunque algunas veces se requieren altas dosis).
Aparentemente el estrés oxidativo producido por el PTZ tiene relación con el NO, puesto que la inhibición de la NOS con N‐Nitro‐L‐Argininina‐Metil‐Ester (L‐NAME), protegió de las convulsiones inducidas por el PTZ y disminuyó es estrés oxidativo, mientras que la L‐Arg, que promueve la síntesis del NO, incrementó el estrés oxidativo.
Esta suposición se ve apoyada por los efectos de los convulsionantes que incrementan los niveles de NO (incluso de forma dependiente de la dosis en el caso del PTZ), en tanto que los anticonvulsionantes los disminuyen. También es posible revertir los efectos de los convulsionantes cuando se protege a los animales con AAOA, y revertir los efectos del PTZ si se protege a los animales con anticonvulsionantes que actúan sobre el sistema GABAérgico. La vitamina “C” parece proteger de las convulsiones no solo por su capacidad antioxidante, sino por sus efectos por los niveles de NO, en tanto que la vitamina E solo revierte el efecto del PTZ sobre los niveles de NO en altas dosis.
En su conjunto, los resultados anteriores indican que durante los estados convulsivos inducidos por fármacos que actúan sobre el sistema GABAérgico se genera estrés oxidativo que, al menos en parte, aparentemente es causado por el NO, el cual también parece estar relacionado con el efecto anticonvulsionante de fármacos cuyo mecanismo de acción se relaciona con el GABA.

INTRODUCCIÓN El estrés oxidativo puede definirse como el estado en que entra la célula cuando se
presenta un desequilibrio entre los sistemas antioxidantes y prooxidantes en favor de estos últimos, y se ha descrito su participación en procesos tanto fisiológicos (envejecimiento) como en la etiología o progresión de numerosas patologías (Halliwell B, 1999).
Puesto que los organismos están constantemente expuestos a muy diferentes agentes oxidantes (como especies reactivas de oxígeno y nitrógeno), prácticamente todo tipo de células sufren estrés oxidativo. Sin embargo, dado que el cerebro consume una desproporcionada cantidad del oxígeno del organismo por obtener su energía casi exclusivamente del metabolismo oxidativo de la cadena respiratoria mitocondrial, es particularmente vulnerable. De hecho, se considera que el estrés oxidativo juega un importante papel en diversos padecimientos degenerativos del sistema nervioso tales como las enfermedades de Alzheimer, Parkinson y Huntington, ictus, trauma y convulsiones entre otras (Coyle y Puttfarcken, 1993).
Se considera que las convulsiones son el resultado del desbalance entre los sistemas excitatorios e inhibitorios (Engelborghs y col, 2000). Así, la mayoría de los anticonvulsionantes actúan incrementando la neurotransmisión mediada por neurotransmisores inhibitorios, reduciendo la neurotransmisión mediada por neurotransmisores excitatorios o una combinación de ambos (White, 1997). El ácido gama aminobutírico (GABA) es el principal neurotransmisor inhibidor en el cerebro, y el mecanismo de acción predominante de las drogas anticonvulsionantes consiste en incrementar la transmisión GABAergica mediante los receptores GABAa. Sin embargo, estos medicamentos pierden eficacia a medida que se desarrollan las convulsiones (Holtkamp y Meierkord, 2007). Por otra parte, el ácido glutámico (glu) es el neurotransmisor excitador más importante, y sus receptores ionotrópicos serían un blanco molecular ideal para drogas anticonvulsionantes, de modo que el bloque de los mismos resulta una eficiente estrategia contra las convulsiones en las últimas etapas (Holtkamp y Meierkord, 2007).
Aunque las mencionadas estrategias farmacológicas podrían considerarse como las más racionales, y estos anticonvulsionantes conocidos como de primera línea pueden ser muy efectivos, una proporción significativa de pacientes presentan convulsiones que son resistentes a los anticonvulsionantes tradicionales. Este hecho, junto con los efectos secundarios de los anticonvulsionantes convencionales, muestra la importancia de diseñar nuevos tratamientos en los que intervengan nuevos blancos moleculares (Holtkamp y Meierkord, 2007).
Mas allá de las dianas farmacológicas anteriormente mencionadas, recientemente se han identificado otros blancos para el tratamiento de las convulsiones. Por una parte, la disrupción de la barrera hematoencefálica inducida por las convulsiones, la inflamación o una combinación de ambos ha conducido a la utilización de antiinflamatorios (Vezzani y Granata, 2005). Estudios experimentales demostraron que las reacciones inflamatorias en el cerebro pueden incrementar la excitabilidad neuronal (Vezzani y col, 1999), y los tratamientos antinflamatorios podrían reducir las convulsiones clínicas y las inducidas experimentalmente (Vezzani y Granata, 2005). El otro blanco potencial actualmente considerado con posibilidades terapéuticas es el anteriormente conocido como factor de crecimiento vascular endotelial, y

que actualmente se ha identificado como óxido nítrico, y se considera que podría proporcionar junto con los metabolitos relacionados puntos a los cuales dirigir el diseño de nuevos fármacos (Holtkamp y Meierkord, 2007).
El óxido nítrico (NO), es un radical libre gaseoso que se sintetiza a partir del aminoácido L‐Arginina mediante la óxido nítrico sintasa (NOS), la cual presenta tres diferentes isoenzimas: Las isoformas neuronal (nNOS) y endotelial (ambas activadas por calcio) y la inducible (activada por una enzima independiente de calcio). El NO actúa sobre la guanilato ciclasa soluble, incrementando así los niveles intracelulares de GMP cíclico (Snyder y Bredt, 1991), el cual es capaz de modular diversas funciones celulares tales como la excitabilidad normal y patológica, la plasticidad neuronal, etc…(Dawson, 1994). En el sistema nervioso central, el NO actúa como un neurotransmisor no convencional, de hecho, contribuye a la liberación de otros neurotransmisores (por ejemplo glutamato, GABA, Dopamina, etc.), y participar en la plasticidad sináptica, elongación neuronal y otras funciones celulares (Moncada y col, 1991; Kiss y Vizi, 2001). Además, al NO se le ha involucrado en una gran variedad de desordenes neurológicos como isquemia, trauma, diferentes enfermedades neurodegenerativas, mostrando así su particular relevancia en la fisiopatología de procesos neurotóxicos y neuroprotectores.
Dentro de los desordenes del SNC, se considerado que el NO juega un papel fundamental en la generación y propagación de las convulsiones epilépticas (Przegalinski y col, 1996). Particularmente, numerosos investigadores han demostrado la implicación funcional del NO tanto en fenómenos proconvulsivos (De Sarro y col, 1991; Buisson y col, 1993; Osonoe y col, 1994; Van Leeuwen y col, 1995; Nidhi y col, 1999) como anticonvulsivos (Starr y Starr, 1993; Theard y col, 1995; Tsuda y col, 1997), pero aún no se llega a conclusiones definitivas al respecto.
Por lo anterior, en este proyecto de investigación se investigó si durante las convulsiones se produce estrés oxidativo y la posible participación del NO en la generación del mismo.
JUSTIFICACIÓN Más de 50 millones de personas en el mundo padecen epilepsia, muchas de las cuales son
refractarias al tratamiento con drogas anticonvulsionantes estándares. Este hecho pone de manifiesto la relevancia de estudios preclínicos en animales que conduzcan a una mayor comprensión del mecanismo de las convulsiones, ya que el conocimiento de los sitios de acción de las drogas antiepilépticas incrementará el entendimiento de su eficacia clínica, proporcionará importante información acerca de esta neuropatología y facilitará el desarrollo de nuevas terapias con mayor efectividad en el tratamiento de la epilepsia y otros desordenes psiquiátricos.
Recientemente, el estrés oxidativo ha surgido como un mecanismo que podría jugar un importante papel en la etiología de la muerte neuronal inducida por las convulsiones (Patel, 2002). El mencionado estrés oxidativo podría estar relacionado con el NO, tanto por ser un radical libre como por ser un neurotransmisor que contribuye a la regulación de los niveles de

GABA y glutamato. Estudios comparativos muestran que en la corteza cerebral de pacientes epilépticos, particularmente en aquellos con un largo historial de convulsiones, el número de neuronas NOS‐positivas se encuentra elevado (Gonzalez‐Hernandez y col, 2000).
HIPÓTESIS DE TRABAJO Si durante los estados convulsivos se genera estrés oxidativo, y el óxido nítrico participa en
este proceso, entonces drogas convulsionantes, anticonvulsionantes y antioxidantes modificaran los niveles de estrés oxidativo, así como la síntesis de óxido nítrico.
OBJETIVO GENERAL Investigar si el estrés oxidativo generado durante los estados convulsivos es causado, al
menos en parte, por el óxido nítrico, con la finalidad de establecer su posible participación en el mecanismo de las convulsiones.
OBJETIVOS ESPECÍFICOS
• Estudiar el efecto de diferentes convulsionantes sobre el estrés oxidativo en cerebro de ratón.
• Estudiar el efecto de diferentes agentes anticonvulsionantes sobre el estrés oxidativo en cerebro de ratón.
• Estudiar el efecto de diferentes agentes antioxidantes sobre el estrés oxidativo en cerebro de ratón.
• Estudiar el efecto de diferentes convulsionantes sobre los niveles de óxido nítrico en cerebro de ratón.
• Estudiar el efecto de diferentes anticonvulsionantes sobre los niveles de óxido nítrico en cerebro de ratón.
• Estudiar el efecto de diferentes antioxidantes sobre los niveles de óxido nítrico en cerebro de ratón.

MATERIALES Y MÉTODOS
ANIMALES DE EXPERIMENTACIÓN
Se emplearon ratones machos de 20 a 25 g de peso de la cepa CD1, con libre acceso al alimento al agua de bebida y ciclo luz‐obscuridad invertido.
FÁRMACOS UTILIZADOS
CONVULSIONANTES:
Metrazol: De la casa Sigma Chemical Co EUA., empleando a diferentes dosis disuelto en agua destilada.
Tiosemicarbazida: De la casa Sigma Chemical Co EUA., disuelto en agua destilada, a una dosis de 20 mg/Kg de peso.
Estricnina: De la casa Sigma Chemical Co EUA., a una dosis de 2 mg/Kg de peso y disuelto en agua destilada.
Bicuculina: De la casa Sigma Chemical Co EUA, a una dosis de 4 mg/Kg de peso, disuelto en agua destilada y pH ajustado a 5 para su disolución total.
ANTICONVULSIONANTES:
Aminooxiacético: De la casa Sigma Chemical Co EUA, a una dosis de 30 mg/Kg de peso, disuelto en agua destilada y pH ajustado a 5 para su disolución total.
Diazepán: Se utilizó en su presentación farmacéutica de ampolleta para administración intramuscular a una dosis de 10 mg/Kg de peso.

Valproato de Sodio: De la casa Sigma Chemical Co EUA, disuelto en agua destilada a una dosis de 200 mg/Kg de peso.
Gabapentina: De la casa Sigma Chemical Co EUA, disuelto en agua destilada a una dosis de 300 mg/Kg de peso.
ANTIOXIDANTES
Ácido Ascórbico (Vitamina “C”): De la casa Sigma Chemical Co EUA, disuelto en agua destilada a diferentes dosis.
α−Tocoferol (Vitamina “E”): De la casa Sigma Chemical Co EUA, disuelto en agua destilada a diferentes dosis.
EFECTORES SOBRE LA SÍNTESIS DEL ÓXIDO NÍTRICO
L‐Arginina: Precursor del NO, disuelto en agua destilada a diferentes dosis.
Nω‐Nitro‐L‐Arginina‐Metil Ester (L‐NAME): Inhibidor de la Sintasa del óxido nítrico, disuelto en agua destilada y usado a diferentes dosis.
Todos los fármacos fueron administrados por vía intraperitoneal (excepto la estricnina, que se administró vía subcutánea).
Los reactivos estándares de laboratorio utilizados fueron de la casas Merck o Sigma.
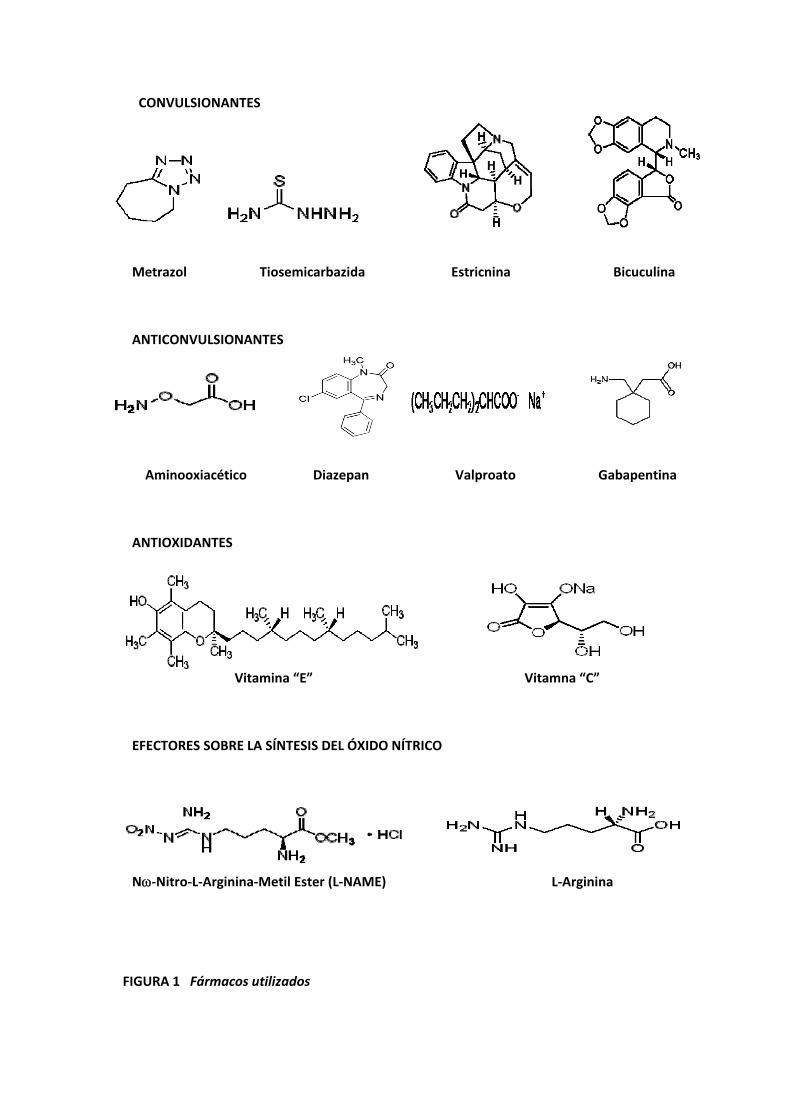
CONVULSIONANTES
Metrazol Tiosemicarbazida Estricnina Bicuculina
ANTICONVULSIONANTES
Aminooxiacético Diazepan Valproato Gabapentina
ANTIOXIDANTES
Vitamina “E” Vitamna “C”
EFECTORES SOBRE LA SÍNTESIS DEL ÓXIDO NÍTRICO
Nω‐Nitro‐L‐Arginina‐Metil Ester (L‐NAME) L‐Arginina
FIGURA 1 Fármacos utilizados

PROCESAMIENTO DE LOS CEREBROS
Los animales se escogen al azar formándose grupos de 5 animales. Los ratones testigos o tratados con los fármacos correspondientes se procesaron como sigue:
• Los animales se sacrifican por decapitación. Se extraen, congelan y pesan los cerebros rápidamente (Mezcla hielo seco‐acetona, ‐40oC)
• Los cerebros se homogenizan en frío con potter Glass‐Col añadiendo 4 volúmenes de Regulador HEPES pH=7.4
A partir del homogeneizado obtenido anteriormente se procede a la determinación de del grado de estrés oxidativo, así como los niveles de óxido nítrico.
DETERMINACIÓN DEL GRADO DE ESTRÉS OXIDATIVO
La evaluación de este parámetro la llevaremos a cabo por medio de dos indicadores, que son la cuantificación de carbonilos libres y ditirosinas, las cuales se describen a continuación:
Cuantificación de Carbonilos Libres:
Esta determinación esta basada en el método espectrofotométrico de Dalle y Rossi (Dalle‐Donne y col, 2003); y se modificó ligeramente para su uso en extracto de cerebro. Consiste en lo siguiente:
• Una vez procesados los cerebros, el homogeneizado de cerebro se centrifuga a 100,000 X g durante 5 minutos

• A partir del sobrenadante se prepara, para cada muestra, una serie de tres tubos como sigue:
TUBO MUESTRA PROBLEMA DPNH 10 mM HCL 2.5 M
BLANCO 50 μl 0 μl 500 μl
PROBLEMA 50 μl 500 μl 0 μl
• Incubar 1 hora en oscuridad agitando cada 15 minutos.
• Agregar 500 ml de TCA al 10%.
• Centrifugar 5 minutos a 3000 rpm.
• Desechar el sobrenadante y agregar 1 ml. De TCA al 5% y disolver la pastilla.
• Centrifugar 5 minutos a 3000 rpm.
• Desechar el sobrenadante y agregar 2 ml. De la solución de extracción etanol‐acetato de etilo 1:1 (v:v) y disolver la pastilla
• Centrifugar 5 minutos a 3000 rpm.
• Eliminar el sobrenadante y secar perfectamente la pastilla
• Disolver la pastilla en 1 ml. De solución de Guanidina 6M y adicionar 2 ml. de regulador HEPES.
• Incubar 10 min. A 37oC
• Leer absorbancia a 370 nm. en espectrofotómetro, utilizando solución de guanidina para calibrar.
Cuantificación de Ditirosinas
Para la cuantificación de ditirosinas se empleó el método fluorométrico basado en el método de Malencik y Cols (Malencik y col, 1996) que se detalla a continuación:
• Una vez procesados los cerebros, se toman 500 μl del homogeneizado de cerebro,
a los cuales se añaden 500 μl de TCA al 10%
• Centrifugar a 300 rpm. Durante 10 min.

• Eliminar el sobrenadante y secar el precipitado lo más posible.
• Adicionar 2 ml. de Urea 6M y disolver el precipitado.
• Incubar 30 min. a temperatura ambiente.
• Calibrar el fluorómetro con una mezcla de 5 μl de sulfato de quinina + 245 μl de urea 6M (en el volumen necesario respetando la proporción indicada)
• Leer la fluorescencia de las muestras problema a una λ de excitación de 400 nm.
CUANTIFICACION DE LOS NIVELES DE ÓXIDO NÍTRICO
Los niveles de óxido nítrico se determinaron en un método basado en la reacción de Griess, (Wu y Brosnan, 1992; Starr y Starr, 1993; Verdon y col, 1995), que consistio en lo siguiente:
• Une vez procesados los cerebros, centrifugar el homogeneizado a 100000 X g durante 5 minutos.
• Separar el sobrenadante, y tomar 250 μl de muestra.
• Adicionar 50 μl de Escherichia coli ATCC 11775, e incubar durante 1 hora a 37oC , agitando a 100 RPM.
• Después de este tiempo, centrifugar a 3000 RPM durante 5 min.
• Pipetear 240 μl del sobrenadante en una placa de 96 pocillos.
• Agregar 40 μl de N‐(1‐naftil)etilendiamina o solució A de Griess (ésta se prepara disolviendo 0.1 g de N‐(1‐naftil)etilendiamina en 5.8 ml de H3PO4 85%, aforar a 100 ml. con agua bidestilada).
• Añadir 40 μl de sulfanilamida o solución B de Griess (ésta se preparada disolviendo 1 g de sulfanilamida en 5.8 ml. de H3PO4 85% y aforar a 100 ml. con agua bidestilada).
• Agitar durante 1 minuto y leer la absorbancia a 554 nm.
RESULTADOS
ESTRÉS OXIDATIVO
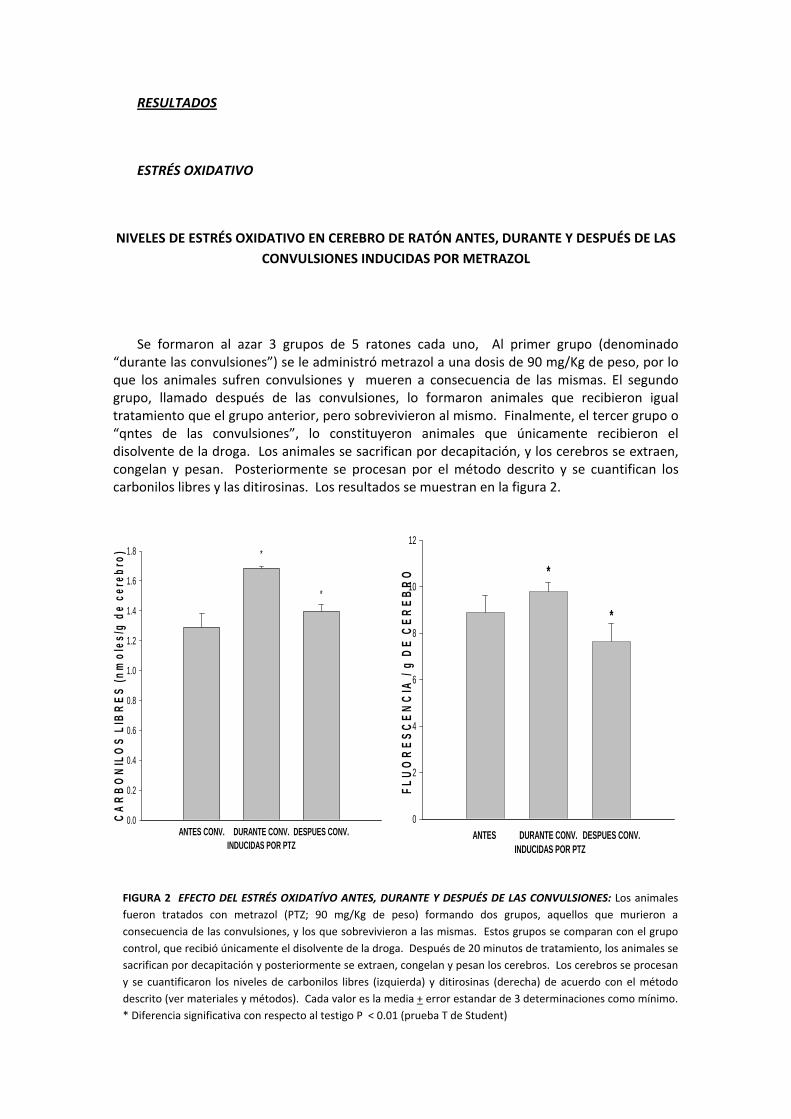
NIVELES DE ESTRÉS OXIDATIVO EN CEREBRO DE RATÓN ANTES, DURANTE Y DESPUÉS DE LAS CONVULSIONES INDUCIDAS POR METRAZOL
Se formaron al azar 3 grupos de 5 ratones cada uno, Al primer grupo (denominado “durante las convulsiones”) se le administró metrazol a una dosis de 90 mg/Kg de peso, por lo que los animales sufren convulsiones y mueren a consecuencia de las mismas. El segundo grupo, llamado después de las convulsiones, lo formaron animales que recibieron igual tratamiento que el grupo anterior, pero sobrevivieron al mismo. Finalmente, el tercer grupo o “qntes de las convulsiones”, lo constituyeron animales que únicamente recibieron el disolvente de la droga. Los animales se sacrifican por decapitación, y los cerebros se extraen, congelan y pesan. Posteriormente se procesan por el método descrito y se cuantifican los carbonilos libres y las ditirosinas. Los resultados se muestran en la figura 2.
ANTES DURANTE CONV. DESPUES CONV.
FLU
OR
ESC
ENC
IA /
g D
E C
EREB
RO
0
2
4
6
8
10
12
INDUCIDAS POR PTZ
*
*
INDUCIDAS POR PTZANTES CONV. DURANTE CONV. DESPUES CONV.
CA
RB
ON
ILO
S LI
BR
ES (n
mol
es/g
de
cere
bro)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
1.8 *
*
FIGURA 2 EFECTO DEL ESTRÉS OXIDATÍVO ANTES, DURANTE Y DESPUÉS DE LAS CONVULSIONES: Los animales fueron tratados con metrazol (PTZ; 90 mg/Kg de peso) formando dos grupos, aquellos que murieron a consecuencia de las convulsiones, y los que sobrevivieron a las mismas. Estos grupos se comparan con el grupo control, que recibió únicamente el disolvente de la droga. Después de 20 minutos de tratamiento, los animales se sacrifican por decapitación y posteriormente se extraen, congelan y pesan los cerebros. Los cerebros se procesan y se cuantificaron los niveles de carbonilos libres (izquierda) y ditirosinas (derecha) de acuerdo con el método descrito (ver materiales y métodos). Cada valor es la media + error estandar de 3 determinaciones como mínimo. * Diferencia significativa con respecto al testigo P < 0.01 (prueba T de Student)

EFECTO DE DIFERENTES DOSIS DE METRAZOL SOBRE EL ESTRÉS OXIDATIVO EN CEREBRO DE RATÓN
Se formaron al azar grupos de 5 ratones cada uno, a los cuales se les aplicó por vía intraperitoneal el metrazol a dosis de 20, 50, 80 y 100 mg/Kg de peso respectivamente., los cuales se compararon con el grupo control, que recibe únicamente el disolvente de la droga. Transcurridos 20 minutos después de la administración del fármaco, los animales se sacrifican por decapitación, y rápidamente los cerebros se extraen, congelan y pesan. A continuación se procesan por el método descrito y se determinan los carbonilos libres y las ditirosinas. Los resultados pueden apreciarse en la figura 3.
DOSIS METRAZOL (mg/Kg)
0 20 40 60 80 1
CA
RB
ON
ILO
S LI
BR
ES (n
mol
es/g
de
cere
bro)
000.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
DOSIS METRAZOL (mg / Kg)0 mg 25 mg 50mg 80mg
FLU
OR
ESC
ENC
IA /
g D
E C
EREB
RO
0
1
2
3
4
5
6
FIGURA 3 EFECTO DE DIFERENTES DOSIS DE METRAZOL SOBRE EL ESTRÉS OXIDATÍVO EN CEREBRO DE RATÓN: Los animales fueron tratados con dosis de 25, 50, 80 y 100 mg/Kg de peso de metrazol. Después de 20 minutos, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros, los cuales se procesan para cuantificar los carbonilos libres (izquierda) y las ditirosinas (derecha) de acuerdo con el método descrito (ver materiales y métodos).
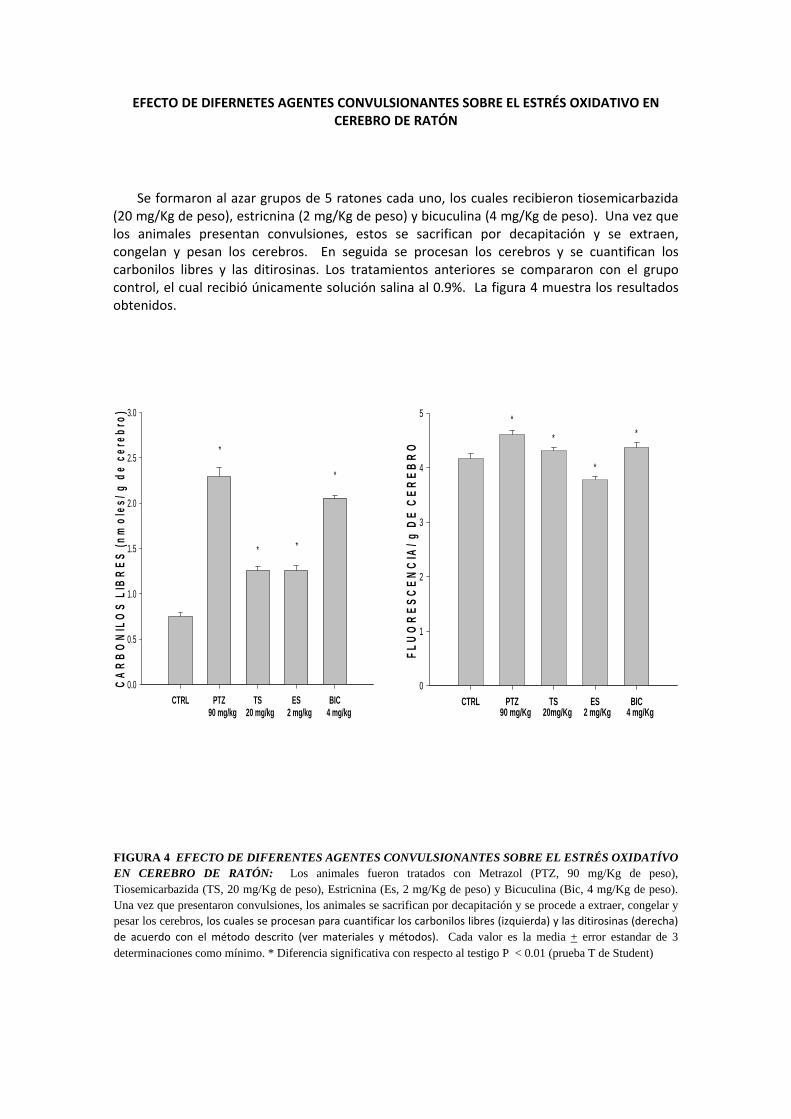
EFECTO DE DIFERNETES AGENTES CONVULSIONANTES SOBRE EL ESTRÉS OXIDATIVO EN CEREBRO DE RATÓN
Se formaron al azar grupos de 5 ratones cada uno, los cuales recibieron tiosemicarbazida (20 mg/Kg de peso), estricnina (2 mg/Kg de peso) y bicuculina (4 mg/Kg de peso). Una vez que los animales presentan convulsiones, estos se sacrifican por decapitación y se extraen, congelan y pesan los cerebros. En seguida se procesan los cerebros y se cuantifican los carbonilos libres y las ditirosinas. Los tratamientos anteriores se compararon con el grupo control, el cual recibió únicamente solución salina al 0.9%. La figura 4 muestra los resultados obtenidos.
*
CTRL PTZ TS ES BIC
CA
RB
ON
ILO
S LI
BR
ES (n
mol
es/ g
de
cere
bro)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
90 mg/kg 20 mg/kg 2 mg/kg 4 mg/kg
*
* *
*
90 mg/Kg 20mg/Kg 2 mg/Kg 4 mg/KgCTRL PTZ TS ES BIC
FLU
OR
ESC
ENC
IA/ g
DE
CER
EBR
O
0
1
2
3
4
5 *
* *
*
FIGURA 4 EFECTO DE DIFERENTES AGENTES CONVULSIONANTES SOBRE EL ESTRÉS OXIDATÍVO EN CEREBRO DE RATÓN: Los animales fueron tratados con Metrazol (PTZ, 90 mg/Kg de peso), Tiosemicarbazida (TS, 20 mg/Kg de peso), Estricnina (Es, 2 mg/Kg de peso) y Bicuculina (Bic, 4 mg/Kg de peso). Una vez que presentaron convulsiones, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros, los cuales se procesan para cuantificar los carbonilos libres (izquierda) y las ditirosinas (derecha) de acuerdo con el método descrito (ver materiales y métodos). Cada valor es la media + error estandar de 3 determinaciones como mínimo. * Diferencia significativa con respecto al testigo P < 0.01 (prueba T de Student)
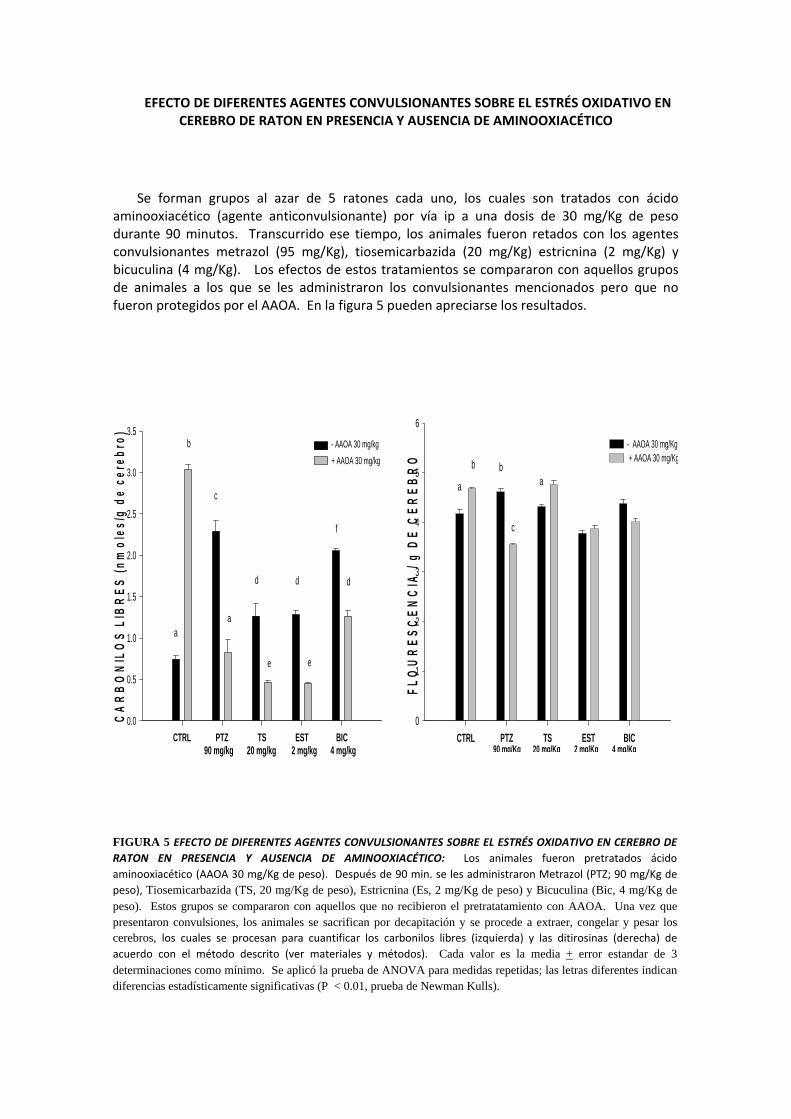
EFECTO DE DIFERENTES AGENTES CONVULSIONANTES SOBRE EL ESTRÉS OXIDATIVO EN CEREBRO DE RATON EN PRESENCIA Y AUSENCIA DE AMINOOXIACÉTICO
Se forman grupos al azar de 5 ratones cada uno, los cuales son tratados con ácido aminooxiacético (agente anticonvulsionante) por vía ip a una dosis de 30 mg/Kg de peso durante 90 minutos. Transcurrido ese tiempo, los animales fueron retados con los agentes convulsionantes metrazol (95 mg/Kg), tiosemicarbazida (20 mg/Kg) estricnina (2 mg/Kg) y bicuculina (4 mg/Kg). Los efectos de estos tratamientos se compararon con aquellos grupos de animales a los que se les administraron los convulsionantes mencionados pero que no fueron protegidos por el AAOA. En la figura 5 pueden apreciarse los resultados.
CTRL PTZ TS EST BIC
FLO
UR
ESC
ENC
IA /
g D
E C
EREB
RO
0
1
2
3
4
5
6
- AAOA 30 mg/Kg + AAOA 30 mg/Kg
a a b b
c
90 mg/Kg 20 mg/Kg 2 mg/Kg 4 mg/Kg
90 mg/kg 20 mg/kg 2 mg/kg 4 mg/kgCTRL PTZ TS EST BIC
CA
RB
ON
ILO
S LI
BR
ES (n
mol
es/g
de
cere
bro)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5- AAOA 30 mg/kg+ g AAOA 30 mg/k
b
c
a
d
e
d
e
f
d
a
FIGURA 5 EFECTO DE DIFERENTES AGENTES CONVULSIONANTES SOBRE EL ESTRÉS OXIDATIVO EN CEREBRO DE RATON EN PRESENCIA Y AUSENCIA DE AMINOOXIACÉTICO: Los animales fueron pretratados ácido aminooxiacético (AAOA 30 mg/Kg de peso). Después de 90 min. se les administraron Metrazol (PTZ; 90 mg/Kg de peso), Tiosemicarbazida (TS, 20 mg/Kg de peso), Estricnina (Es, 2 mg/Kg de peso) y Bicuculina (Bic, 4 mg/Kg de peso). Estos grupos se compararon con aquellos que no recibieron el pretratatamiento con AAOA. Una vez que presentaron convulsiones, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros, los cuales se procesan para cuantificar los carbonilos libres (izquierda) y las ditirosinas (derecha) de acuerdo con el método descrito (ver materiales y métodos). Cada valor es la media + error estandar de 3 determinaciones como mínimo. Se aplicó la prueba de ANOVA para medidas repetidas; las letras diferentes indican diferencias estadísticamente significativas (P < 0.01, prueba de Newman Kulls).

EFECTO DE DIFERNETES AGENTES ANTICONVULSIONANTES SOBRE EL ESTRÉS OXIDATIVO EN CEREBRO DE RATÓN
Se formaron al azar grupos de 5 ratones cada uno, a los cuales se les administraron ácido aminooxiacético (30 mg/Kg), diazepan (10 mg/Kg), valproato (200 mg/Kg) y gabapentina (300 mg/Kg). Una vez que los fármacos han ejercido su efecto anticonvulsionante, los animales se sacrifican por decapitación y los cerebros se extraen, congelan y pesan. A continuación se les procesa y se cuantifican tanto los carbonilos libres como las ditirosinas. La figura 6 muestra los resultados obtenidos.
30 mg/ Kg 10 mg/Kg 200 mg/Kg 300 mg/KgCTRL AAOA DIAZ VALP GBPTNA
FLU
OR
ESC
ENC
IA/ g
DE
CER
EBR
O
0.0
0.5
1.0
1.5
2.0
2.5
3.0* *
**
30 mg/Kg 10 mg/Kg 200 mg/kg 300 mg/Kg
CTRL AAOA DIAZ VALP GBPTNA
CA
RB
ON
ILO
S LI
BR
ES (n
mol
es/g
de
cere
bro)
0.0
0.5
1.0
1.5
2.0
2.5
*
*
FIGURA 6 EFECTO DE DIFERNETES AGENTES ANTICONVULSIONANTES SOBRE EL ESTRÉS OXIDATIVO EN CEREBRO DE RATÓN Los animales fueron tratados con ácido aminooxiacético (AAOA, 30 mg/Kg de peso, 90 min.), diazepan (DIAZ, 10 mg/Kg de peso, 60 min.), valproato (VALP, 200 mg/Kg de peso, 60 min.) y gabapentina (GBPTNA, 300 mg/Kg de peso, 90 min.). Una vez transcurrido el tiempo de tratamiento, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros, los cuales se procesan para cuantificar los carbonilos libres (izquierda) y las ditirosinas (derecha) de acuerdo con el método descrito (ver materiales y métodos). Cada valor es la media + error estandar de 3 determinaciones como mínimo. * Diferencia significativa con respecto al testigo P < 0.01 (prueba T de Student)

EFECTO DE DIFERENTES AGENTES ANTICONVULSIONANTES SOBRE EL ESTRÉS OXIDATIVO EN CEREBRO DE RATÓN EN PRESENCIA Y AUSENCIA DE METRAZOL
Se formaron al azar grupos de 5 ratones cada uno, a los cuales se les administraron ácido aminooxiacético (30 mg/Kg), diazepan (10 mg/Kg), valproato (200 mg/Kg) y gabapentina (300 mg/Kg). Cuando los anticonvulsionantes han actuado, se inyecta a los animales con metrazol (90 mg/Kg), y 20 minutos mas tarde los animales se sacrifican por decapitación. Estos se compararon con los grupos que recibieron solamente los anticonvulsionantes. En todos los casos se extraen los cerebros, los cuales se congelan y pesan rápidamente, y se procesan como se describió anteriormente. En la figura 7, pueden apreciarse los resultados alcanzados.
CTRL VALP GPTNA DIAZ
FLU
OR
ESC
ENC
IA /
g D
E C
EREB
RO
0
1
2
3
4 - PTZ 90 mg/Kg + PTZ 90 mg/Kg
a
b
ac
d e
f
g
CTRL VALP GPTINA DIAZ
CA
RB
ON
ILO
S LI
BR
ES (
nmol
es /
g de
cer
ebro
)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5 - PTZ 90 mg/Kg + PTZ 90 mg/Kg
a
b
c
a
d
e e
f
FIGURA 7 EFECTO DE DIFERENTES AGENTES ANTICONVULSIONANTES SOBRE EL ESTRÉS OXIDATIVO EN CEREBRO DE RATON EN PRESENCIA Y AUSENCIA DE METRAZOL: Los animales fueron tratados con ácido aminooxiacético (AAOA, 30 mg/Kg de peso, 90 min.), diazepan (DIAZ, 10 mg/Kg de peso, 60 min.), valproato (VALP, 200 mg/Kg de peso, 60 min.) y gabapentina (GBPTNA, 300 mg/Kg de peso, 90 min.). Una vez transcurrido el tiempo de tratamiento, los animales fueron retados con metrazol (PTZ, 95 mg/Kg de peso). Después de 20 minutos, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros, los cuales se procesan para cuantificar los carbonilos libres (izquierda) y las ditirosinas (derecha) de acuerdo con el método descrito (ver materiales y métodos). Estos grupos se compararon con aquellos que no recibieron el PTZ. Cada valor es la media + error estandar de 3 determinaciones como mínimo. Se aplicó la prueba de ANOVA para medidas repetidas; las letras diferentes indican diferencias estadísticamente significativas (P < 0.01, prueba de Newman Kulls).
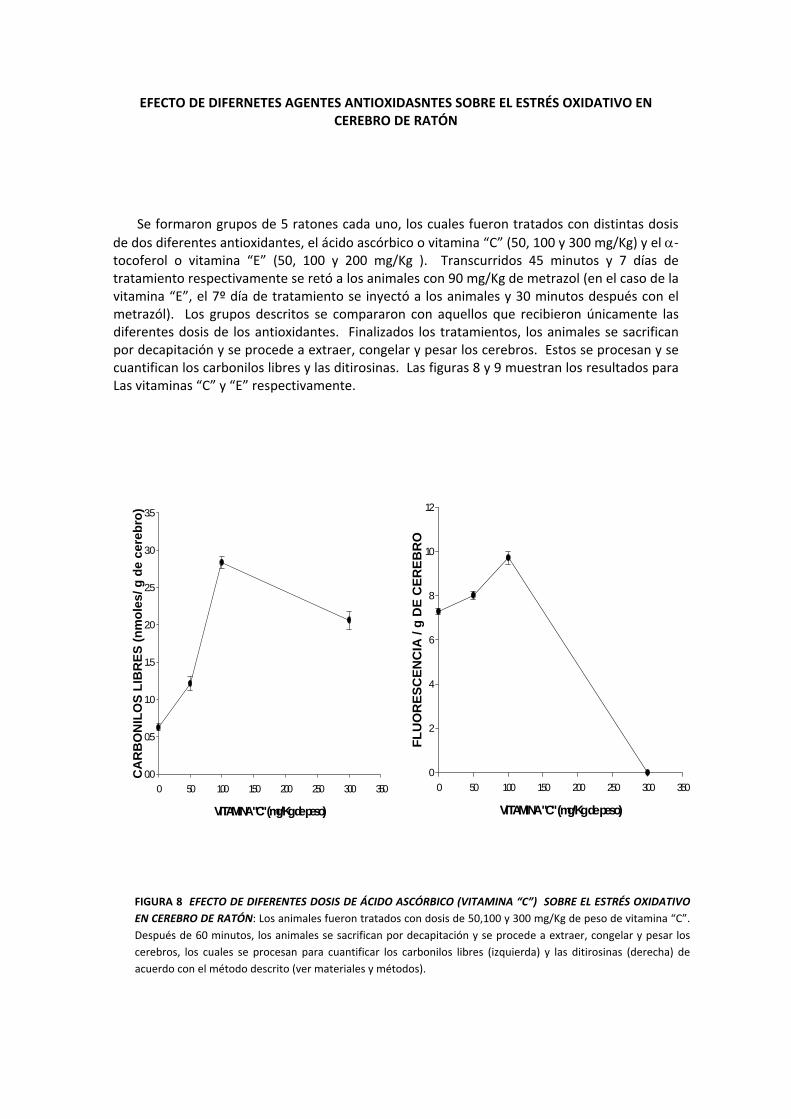
EFECTO DE DIFERNETES AGENTES ANTIOXIDASNTES SOBRE EL ESTRÉS OXIDATIVO EN CEREBRO DE RATÓN
Se formaron grupos de 5 ratones cada uno, los cuales fueron tratados con distintas dosis de dos diferentes antioxidantes, el ácido ascórbico o vitamina “C” (50, 100 y 300 mg/Kg) y el α‐tocoferol o vitamina “E” (50, 100 y 200 mg/Kg ). Transcurridos 45 minutos y 7 días de tratamiento respectivamente se retó a los animales con 90 mg/Kg de metrazol (en el caso de la vitamina “E”, el 7º día de tratamiento se inyectó a los animales y 30 minutos después con el metrazól). Los grupos descritos se compararon con aquellos que recibieron únicamente las diferentes dosis de los antioxidantes. Finalizados los tratamientos, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros. Estos se procesan y se cuantifican los carbonilos libres y las ditirosinas. Las figuras 8 y 9 muestran los resultados para Las vitaminas “C” y “E” respectivamente.
VITAMINA "C" (mg/Kg de peso)
0 50 100 150 200 250 300 350
CA
RB
ON
ILO
S LI
BR
ES (n
mol
es/ g
de
cere
bro)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
VITAMINA "C" (mg/Kg de peso)
0 50 100 150 200 250 300 350
FLU
OR
ESC
ENC
IA /
g D
E C
EREB
RO
0
2
4
6
8
10
12
FIGURA 8 EFECTO DE DIFERENTES DOSIS DE ÁCIDO ASCÓRBICO (VITAMINA “C”) SOBRE EL ESTRÉS OXIDATIVO EN CEREBRO DE RATÓN: Los animales fueron tratados con dosis de 50,100 y 300 mg/Kg de peso de vitamina “C”. Después de 60 minutos, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros, los cuales se procesan para cuantificar los carbonilos libres (izquierda) y las ditirosinas (derecha) de acuerdo con el método descrito (ver materiales y métodos).
VITAMINA "E" (mg/Kg de peso)
0 50 100 150 200 250
CA
RB
ON
ILO
S LI
BR
ES (n
mol
es/ g
de
cere
bro)
0.0
0.5
1.0
1.5
VITAMINA "E" (mg/Kg de peso)
0 50 100 150 200 250
FLU
OR
ESC
ENC
IA /
g D
E C
EREB
RO
0
2
4
6
8
10
12
14
16
4.
0
5
0
5
0
3. 3. 2. 2.
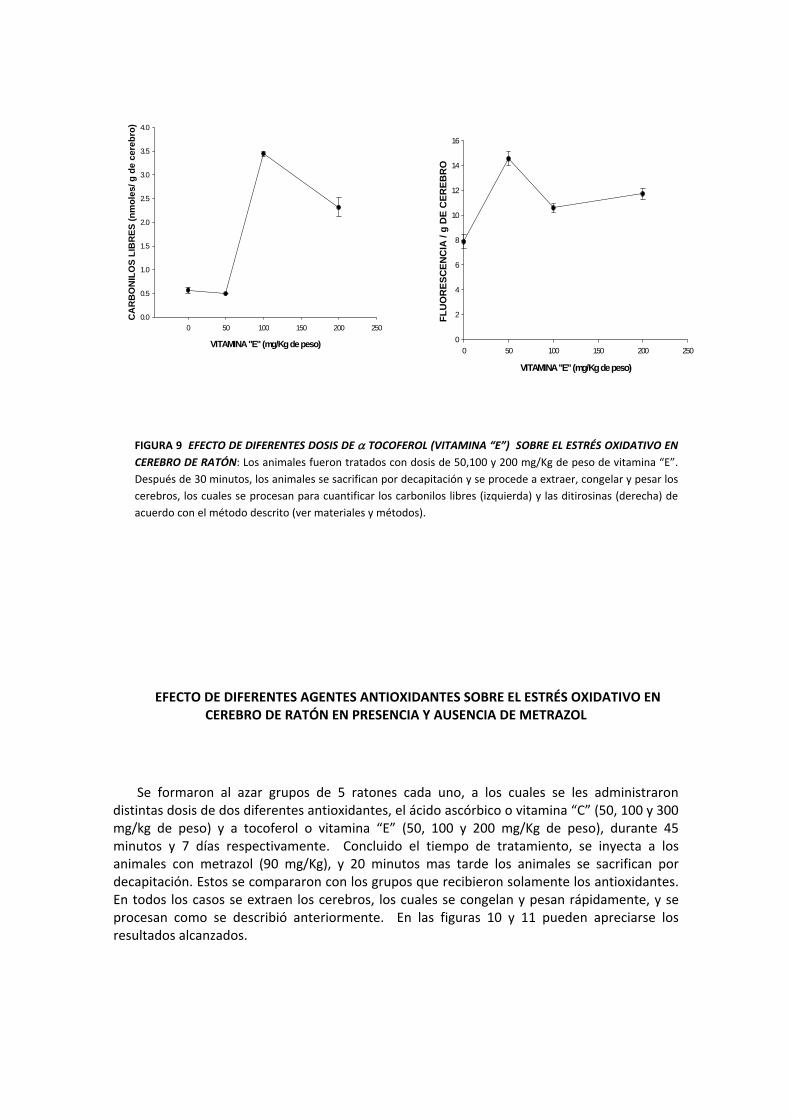
FIGURA 9 EFECTO DE DIFERENTES DOSIS DE α TOCOFEROL (VITAMINA “E”) SOBRE EL ESTRÉS OXIDATIVO EN CEREBRO DE RATÓN: Los animales fueron tratados con dosis de 50,100 y 200 mg/Kg de peso de vitamina “E”. Después de 30 minutos, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros, los cuales se procesan para cuantificar los carbonilos libres (izquierda) y las ditirosinas (derecha) de acuerdo con el método descrito (ver materiales y métodos).
EFECTO DE DIFERENTES AGENTES ANTIOXIDANTES SOBRE EL ESTRÉS OXIDATIVO EN CEREBRO DE RATÓN EN PRESENCIA Y AUSENCIA DE METRAZOL
Se formaron al azar grupos de 5 ratones cada uno, a los cuales se les administraron distintas dosis de dos diferentes antioxidantes, el ácido ascórbico o vitamina “C” (50, 100 y 300 mg/kg de peso) y a tocoferol o vitamina “E” (50, 100 y 200 mg/Kg de peso), durante 45 minutos y 7 días respectivamente. Concluido el tiempo de tratamiento, se inyecta a los animales con metrazol (90 mg/Kg), y 20 minutos mas tarde los animales se sacrifican por decapitación. Estos se compararon con los grupos que recibieron solamente los antioxidantes. En todos los casos se extraen los cerebros, los cuales se congelan y pesan rápidamente, y se procesan como se describió anteriormente. En las figuras 10 y 11 pueden apreciarse los resultados alcanzados.
CONTROL VIT C 50 VIT C 100 VIT C 300
CA
RB
ON
ILO
S LI
BR
ES (n
mol
es/ g
de
cere
bro)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
- PTZ +PTZ
a
b
c
d
e
f
g
f
CONTROL VIT C 50 VIT C 100 VIT C 300
FLU
OR
ESC
ENC
IA/g
DE
CER
EBR
O
0
2
4
6
8
10
12- PTZ+ PTZ
CONTROL VIT E 50 VIT E 100 VIT E 200
FLU
OR
ESC
ENC
IA/g
DE
CER
EBR
O
0
2
4
6
8
10
12
14
16-PTZ + PTZ
a
b
c
d
e
f
g
h
CONTROL VIT E 50 VIT E 100 VIT E 200
CA
RB
ON
ILO
S LI
BR
ES (n
mol
es/g
de
cere
bro)
0
1
2
3
4
- PTZ + PTZ
a
b
c
d
e
f
g
f
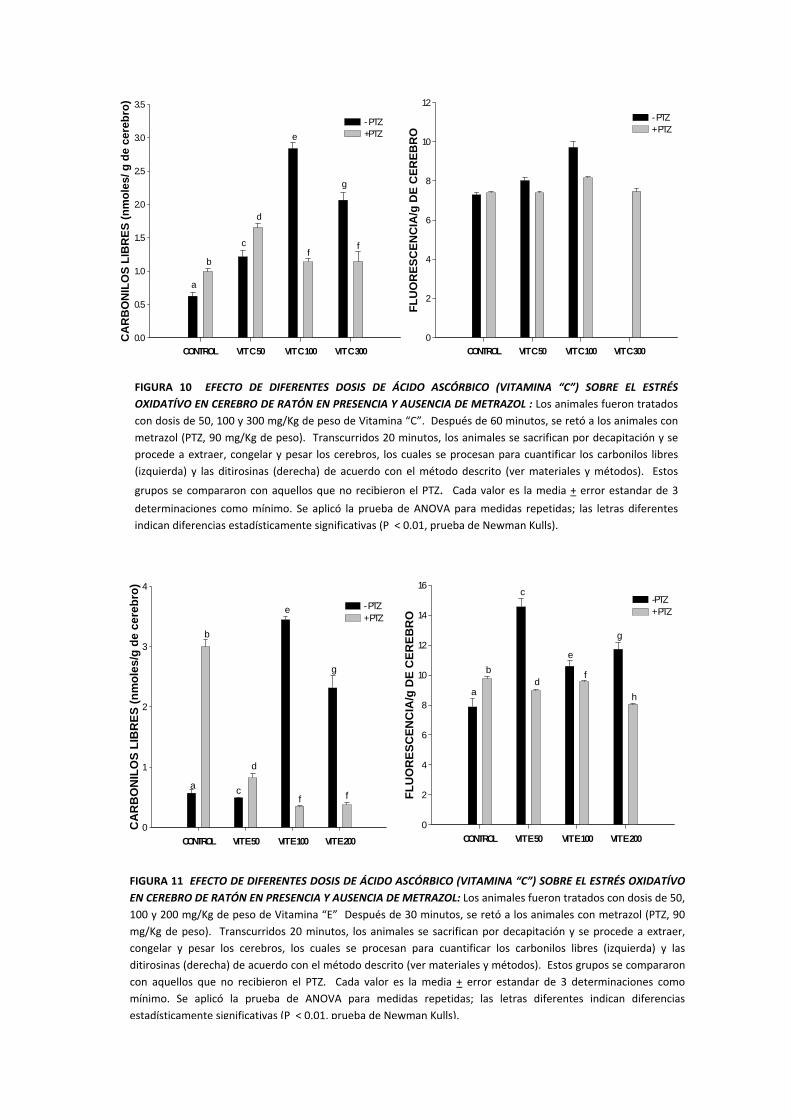
FIGURA 10 EFECTO DE DIFERENTES DOSIS DE ÁCIDO ASCÓRBICO (VITAMINA “C”) SOBRE EL ESTRÉS OXIDATÍVO EN CEREBRO DE RATÓN EN PRESENCIA Y AUSENCIA DE METRAZOL : Los animales fueron tratados con dosis de 50, 100 y 300 mg/Kg de peso de Vitamina “C”. Después de 60 minutos, se retó a los animales con metrazol (PTZ, 90 mg/Kg de peso). Transcurridos 20 minutos, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros, los cuales se procesan para cuantificar los carbonilos libres (izquierda) y las ditirosinas (derecha) de acuerdo con el método descrito (ver materiales y métodos). Estos
grupos se compararon con aquellos que no recibieron el PTZ. Cada valor es la media
+ error estandar de 3
determinaciones como mínimo. Se aplicó la prueba de ANOVA para medidas repetidas; las letras diferentes indican diferencias estadísticamente significativas (P < 0.01, prueba de Newman Kulls).
FIGURA 11 EFECTO DE DIFERENTES DOSIS DE ÁCIDO ASCÓRBICO (VITAMINA “C”) SOBRE EL ESTRÉS OXIDATÍVO EN CEREBRO DE RATÓN EN PRESENCIA Y AUSENCIA DE METRAZOL: Los animales fueron tratados con dosis de 50, 100 y 200 mg/Kg de peso de Vitamina “E” Después de 30 minutos, se retó a los animales con metrazol (PTZ, 90 mg/Kg de peso). Transcurridos 20 minutos, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros, los cuales se procesan para cuantificar los carbonilos libres (izquierda) y las ditirosinas (derecha) de acuerdo con el método descrito (ver materiales y métodos). Estos grupos se compararon con aquellos que no recibieron el PTZ. Cada valor es la media + error estandar de 3 determinaciones como mínimo. Se aplicó la prueba de ANOVA para medidas repetidas; las letras diferentes indican diferencias estadísticamente significativas (P < 0.01, prueba de Newman Kulls).

EFECTO DE LA PROMOCIÓN E INHIBICIÓN DE LA SÍNTESIS DE ÓXIDO NÍTRICO SOBRE EL ESTRÉS OXIDATIVO GENERADO POR EL METRAZOL EN CEREBRO DE RATÓN
A grupos de 5 ratones formados al azar se les administra L‐ Arginina y L‐NAME (precursor e inhibidor de la síntesis de óxido nítrico respectivamente) a una dosis de 125 y 100 mg/Kg de peso respectivamente. A los 45 minutos de tratamiento se les administró metrazol a una dosis de 80 mg/Kg de peso. Transcurridos 20 minutos, los animales se sacrifican por decapitación, los cerebros se extraen, congelan y pesan, para ser procesados como se describió anteriormente. Enseguida se cuantifican los carbonilos libres y las ditirosinas. La figura 12 muestra los resultados obtenidos.
FLU
OR
ES
CE
NC
IA /
g D
E C
ER
EB
RO
0
2
4
6
8
10CTRLL-ARGL-NAME
EN AUSENCIA DE PTZ EN PRESENCIA DE PTZ
a
b
cd
e
f
EN AUSENCIA DE PTZ EN PRESENCIA DE PTZ
CAR
BON
ILO
S L
IBR
ES (n
mol
es/g
de
cere
bro)
0.0
0.5
1.0
1.5
2.0
2.5CTRLL-ARGL-NAME
a
b
c
d
e
f
FIGURA 12 EFECTO DE LA PROMOCIÓN E INHIBICIÓN DE LA SÍNTESIS DE ÓXIDO NÍTRICO SOBRE EL ESTRÉS OXIDATIVO GENERADO POR EL METRAZOL EN CEREBRO DE RATÓN Los animales fueron tratados con L‐Arginina (L‐ARG; 125 mg/kg), y con L‐NAME (L‐NAME; 100 mg/Kg), promotor e inhibidor de la síntesis de NO respectivamente. Después de 45 minutos, se reta a los animales con metrazol (PTZ, 80 mg/Kg). Transcurridos 20 minutos, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros, los cuales se procesan para cuantificar los carbonilos libres (izquierda) y las ditirosinas (derecha) de acuerdo con el método descrito (ver materiales y métodos). Estos grupos se compararon con aquellos que no recibieron el PTZ. Cada valor es la media + error estandar de 3 determinaciones como mínimo. Se aplicó la prueba de ANOVA para medidas repetidas; las letras diferentes indican diferencias estadísticamente significativas (P < 0.01, prueba de Newman Kulls).

EFECTO DE LA PROMOCIÓN E INHIBICIÓN DE LA SÍNTESIS DE ÓXIDO NÍTRICO SOBRE EL ESTRÉS OXIDATÍVO INDUCIDO POR DIFERENTES DOSIS DE METRAZOL EN CEREBRO DE
RATÓN
A grupos de 5 ratones formados al azar se les administra L‐ Arginina y L‐NAME (precursor e inhibidor de la síntesis de óxido nítrico respectivamente) a una dosis de 125 y 100 mg/Kg de peso respectivamente. Posteriormente se administra a los animales diferentes dosis de metrazol (20, 50, 80 y 100 mg/Kg de peso). Estos grupos se comparan con aquellos que recibieron únicamente las distintas dosis de metrazol. Pasados 20 minutos, los animales se sacrifican por decapitación, y rápidamente los cerebros se extraen, congelan y pesan. Estos se procesan por el método descrito y se determinan los carbonilos libres y las ditirosinas. La figura 13 muestra los resultados.
DOSIS DE METRAZOL (mg/Kg)
0 20 40 60 80 100
CA
RB
ON
ILO
S LI
BR
ES (n
mol
es/g
de
cere
bro)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
PTZPTZ + L-NAMEPTZ + L-ARG
DOSIS DE METRAZOL (mg / kg)
0 20 40 60 80 100
FLU
OR
ESC
ENC
IA /g
DE
CER
EBR
O
0
6
7
8
9
10
PTZPTZ + L-NAMEPTZ + L-ARG
FIGURA 13 EFECTO DE LA PROMOCIÓN E INHIBICIÓN DE LA SÍNTESIS DE ÓXIDO NÍTRICO SOBRE EL ESTRÉS OXIDATÍVO INDUCIDO POR DIFERENTES DOSIS DE METRAZOL EN CEREBRO DE RATÓN: Los animales fueron pretratados con L-Arginina (L-ARG; 125 mg/kg) o con L-NAME (L-NAME; 100 mg/Kg), promotor e inhibidor de la síntesis de NO respectivamente. Después de 45 minutos, se reta a los animales con diferentes dosis de metrazol (PTZ, 20, 50, 80 y 100 80 mg/Kg). Estos grupos se comparan con aquellos que recibieron únicamente las distintas dosis de PTZ. Después de 20 minutos de haberles administrado el PTZ, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros, los cuales se procesan para cuantificar los carbonilos libres (izquierda) y las ditirosinas (derecha) de acuerdo con el método descrito (ver materiales y métodos).

NIVELES DE ÓXIDO NITRICO
NIVELES DE ÓXIDO NÍTRICO EN CEREBRO DE RATÓN ANTES, DURANTE Y DESPUÉS DE LAS
CONVULSIONES INDUCIDAS POR METRAZOL
Se formaron al azar 3 grupos de 5 ratones cada uno, Al primer grupo (denominado “durante las convulsiones”) se le administró metrazol a una dosis de 90 mg/Kg de peso, por lo que los animales sufren convulsiones y mueren a consecuencia de las mismas. El segundo grupo, llamado después de las convulsiones, lo formaron animales que recibieron igual tratamiento que el grupo anterior, pero sobrevivieron al mismo. Finalmente, el tercer grupo o “antes de las convulsiones”, lo constituyeron animales que únicamente recibieron el disolvente de la droga. Los animales se sacrifican por decapitación, y los cerebros se extraen, congelan y pesan. Posteriormente se procesan por el método descrito y se cuantifican los niveles de óxido nítrico. Los resultados pueden observarse en la figura 14.
INDUCIDAS POR PTZANTES CONV. DURANTE CONV. POST. CONV.
μGR
AM
OS
DE
NIT
RIT
OS
/ g D
E C
EREB
RO
0.0
0.1
0.2
0.3
0.4
*
*
FIGURA 14 NIVELES DE ÓXIDO NÍTRICO EN CEREBRO DE RATÓN ANTES, DURANTE Y DESPUÉS DE LAS CONVULSIONES INDUCIDAS POR METRAZOL: Los animales fueron tratados con metrazol (PTZ; 90 mg/Kg de peso) formando dos grupos, aquellos que murieron a consecuencia de las convulsiones, y los que sobrevivieron a las mismas. Estos grupos se comparan con el grupo control, que recibió únicamente el disolvente de la droga. Después de 20 minutos de tratamiento, los animales se sacrifican por decapitación y posteriormente se extraen, congelan y pesan los cerebros. Los cerebros se procesan y se cuantificaron los niveles de óxido nítrico de acuerdo con el método descrito (ver materiales y métodos). Cada valor es la media + error estandar de 3 determinaciones como mínimo. * Diferencia significativa con respecto al testigo P < 0.01 (prueba T de Student)
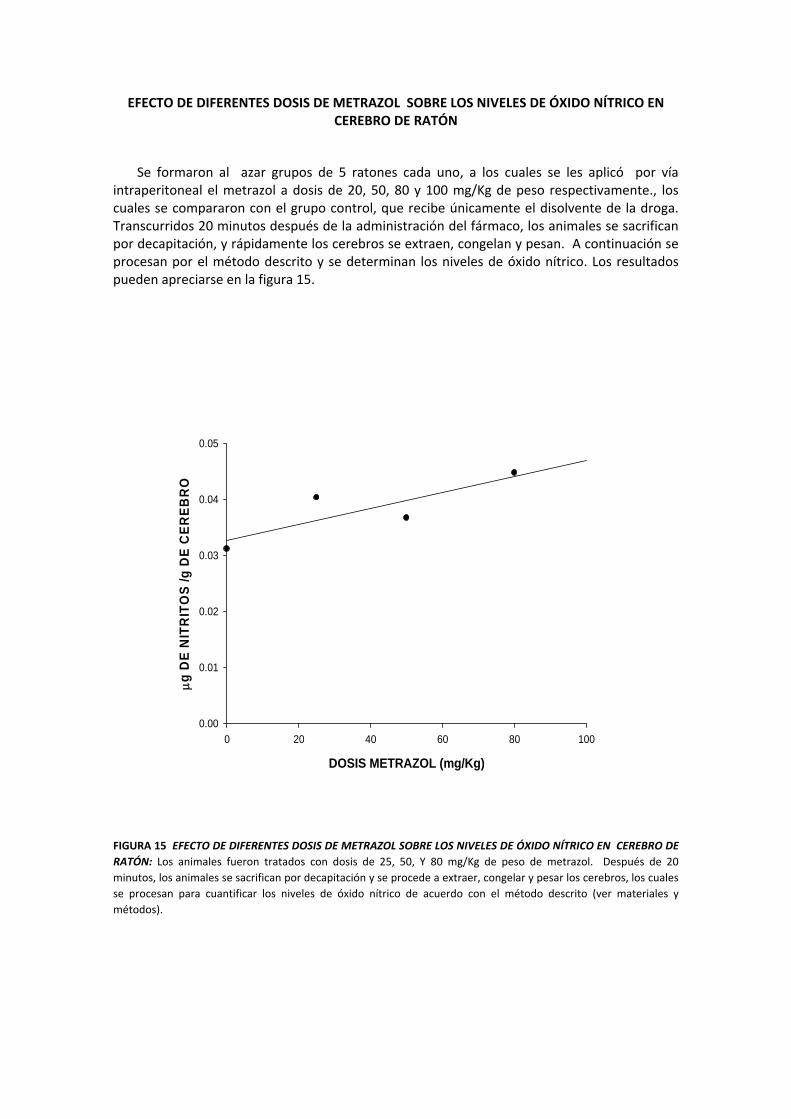
EFECTO DE DIFERENTES DOSIS DE METRAZOL SOBRE LOS NIVELES DE ÓXIDO NÍTRICO EN CEREBRO DE RATÓN
Se formaron al azar grupos de 5 ratones cada uno, a los cuales se les aplicó por vía intraperitoneal el metrazol a dosis de 20, 50, 80 y 100 mg/Kg de peso respectivamente., los cuales se compararon con el grupo control, que recibe únicamente el disolvente de la droga. Transcurridos 20 minutos después de la administración del fármaco, los animales se sacrifican por decapitación, y rápidamente los cerebros se extraen, congelan y pesan. A continuación se procesan por el método descrito y se determinan los niveles de óxido nítrico. Los resultados pueden apreciarse en la figura 15.
DOSIS METRAZOL (mg/Kg)
0 20 40 60 80
μg D
E N
ITR
ITO
S /g
DE
CER
EBR
O
1000.00
0.01
0.02
0.03
0.04
0.05
FIGURA 15 EFECTO DE DIFERENTES DOSIS DE METRAZOL SOBRE LOS NIVELES DE ÓXIDO NÍTRICO EN CEREBRO DE RATÓN: Los animales fueron tratados con dosis de 25, 50, Y 80 mg/Kg de peso de metrazol. Después de 20 minutos, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros, los cuales se procesan para cuantificar los niveles de óxido nítrico de acuerdo con el método descrito (ver materiales y métodos).

EFECTO DE DIFERNETES AGENTES CONVULSIONANTES SOBRE LOS NIVELES DE ÓXIDO NÍTRICO EN CEREBRO DE RATÓN
Se formaron al azar grupos de 5 ratones cada uno, los cuales recibieron metrazol (95 mg/Kg), tiosemicarbazida (20 mg/Kg de peso), estricnina (2 mg/Kg de peso) y bicuculina (4 mg/Kg de peso). Una vez que los animales presentan convulsiones, estos se sacrifican por decapitación y se extraen, congelan y pesan los cerebros. En seguida se procesan los cerebros y se cuantifican los niveles de óxido nítrico. Los tratamientos anteriores se compararon con el grupo control, el cual recibió únicamente solución salina al 0.9%. La figura 16 muestra los resultados obtenidos.
CTRL PTZ TS EST BIC
μGR
AM
OS
DE
NIT
RIT
OS
/ g D
E C
EREB
RO
0.0
0.2
0.4
0.6
0.8
1.0
**
*
*
FIGURA 16 EFECTO DE DIFERENTES AGENTES CONVULSIONANTES SOBRE LOS NIVELES DE ÓXIDO NÍTRICO EN CEREBRO DE RATÓN: Los animales fueron tratados con Metrazol (PTZ, 90 mg/Kg de peso), Tiosemicarbazida (TS, 20 mg/Kg de peso), Estricnina (Es, 2 mg/Kg de peso) y Bicuculina (Bic, 4 mg/Kg de peso). Una vez que presentaron convulsiones, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros, los cuales se procesan para cuantificar los niveles de óxido nítrico de acuerdo con el método descrito (ver materiales y métodos). Cada valor es la media + error estandar de 3 determinaciones como mínimo. * Diferencia significativa con respecto al testigo P < 0.01 (prueba T de Student)

EFECTO DE DIFERENTES AGENTES CONVULSIONANTES SOBRE LOS NIVELES DE ÓXIDO NÍTRICO EN CEREBRO DE RATÓN EN PRESENCIA Y AUSENCIA DE AMINOOXIACÉTICO
Se forman grupos al azar de 5 ratones cada uno, los cuales son tratados con ácido aminooxiacético (agente anticonvulsionante) por vía ip a una dosis de 30 mg/Kg de peso durante 90 minutos. Transcurrido ese tiempo, los animales fueron retados con los agentes convulsionantes metrazol (95 mg/Kg), tiosemicarbazida (20 mg/Kg) estricnina (2 mg/Kg) y bicuculina (4 mg/Kg). Los efectos de la protección con el AAOA podrán notarse al compararse con aquellos grupos de animales a los que se les administraron los convulsionantes mencionados pero que no fueron protegidos por el AAOA. En la figura 17 pueden apreciarse los resultados.
CONTROL PTZ TS EST BIC
μg n
itrito
s / g
de
cere
bro
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7- AAOA+ AAOA
a
b
c
d
e
f
g
hi
j
FIGURA 17 EFECTO DE DIFERENTES AGENTES CONVULSIONANTES SOBRE LOS NIVELES DE ÓXIDO NÍTRICO EN CEREBRO DE RATÓN EN PRESENCIA Y AUSENCIA DE AMINOOXIACÉTICO: Los animales fueron pretratados con ácido aminooxiacético (AAOA 30 mg/Kg de peso). Después de 90 min. se les administró Metrazol (PTZ; 90 mg/Kg de peso), Tiosemicarbazida (TS, 20 mg/Kg de peso), Estricnina (Es, 2 mg/Kg de peso) y Bicuculina (Bic, 4 mg/Kg de peso). Estos grupos se compararon con aquellos que no recibieron el pretratatamiento con AAOA. Una vez que presentaron convulsiones, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros, los cuales se procesan para cuantificar los niveles de óxido nítrico de acuerdo con el método descrito (ver materiales y métodos). Cada valor es la media
+ error estandar de 3 determinaciones como mínimo. Se aplicó la prueba de ANOVA para medidas repetidas; las letras diferentes indican diferencias estadísticamente significativas (P < 0.01, prueba de Newman Kulls).
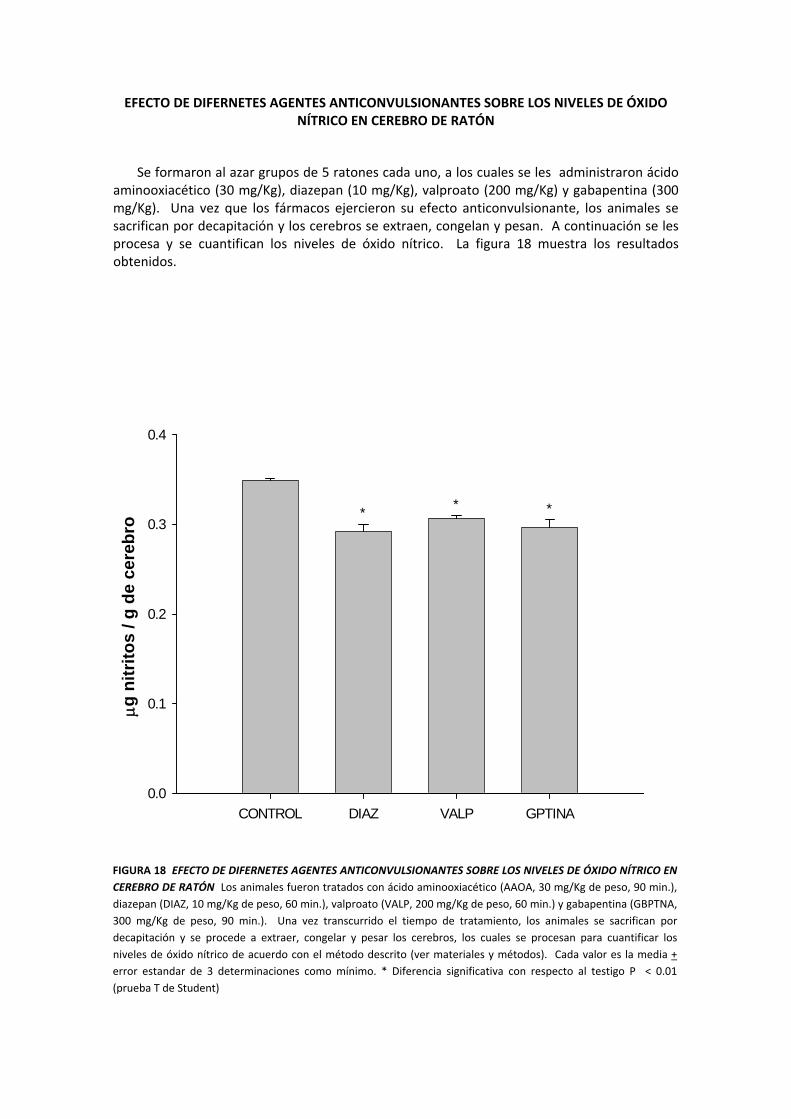
EFECTO DE DIFERNETES AGENTES ANTICONVULSIONANTES SOBRE LOS NIVELES DE ÓXIDO NÍTRICO EN CEREBRO DE RATÓN
Se formaron al azar grupos de 5 ratones cada uno, a los cuales se les administraron ácido aminooxiacético (30 mg/Kg), diazepan (10 mg/Kg), valproato (200 mg/Kg) y gabapentina (300 mg/Kg). Una vez que los fármacos ejercieron su efecto anticonvulsionante, los animales se sacrifican por decapitación y los cerebros se extraen, congelan y pesan. A continuación se les procesa y se cuantifican los niveles de óxido nítrico. La figura 18 muestra los resultados obtenidos.
CONTROL DIAZ VALP GPTINA
μg n
itrito
s / g
de
cere
bro
0.0
0.1
0.2
0.3
0.4
* **
FIGURA 18 EFECTO DE DIFERNETES AGENTES ANTICONVULSIONANTES SOBRE LOS NIVELES DE ÓXIDO NÍTRICO EN CEREBRO DE RATÓN Los animales fueron tratados con ácido aminooxiacético (AAOA, 30 mg/Kg de peso, 90 min.), diazepan (DIAZ, 10 mg/Kg de peso, 60 min.), valproato (VALP, 200 mg/Kg de peso, 60 min.) y gabapentina (GBPTNA, 300 mg/Kg de peso, 90 min.). Una vez transcurrido el tiempo de tratamiento, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros, los cuales se procesan para cuantificar los niveles de óxido nítrico de acuerdo con el método descrito (ver materiales y métodos). Cada valor es la media + error estandar de 3 determinaciones como mínimo. * Diferencia significativa con respecto al testigo P < 0.01 (prueba T de Student)
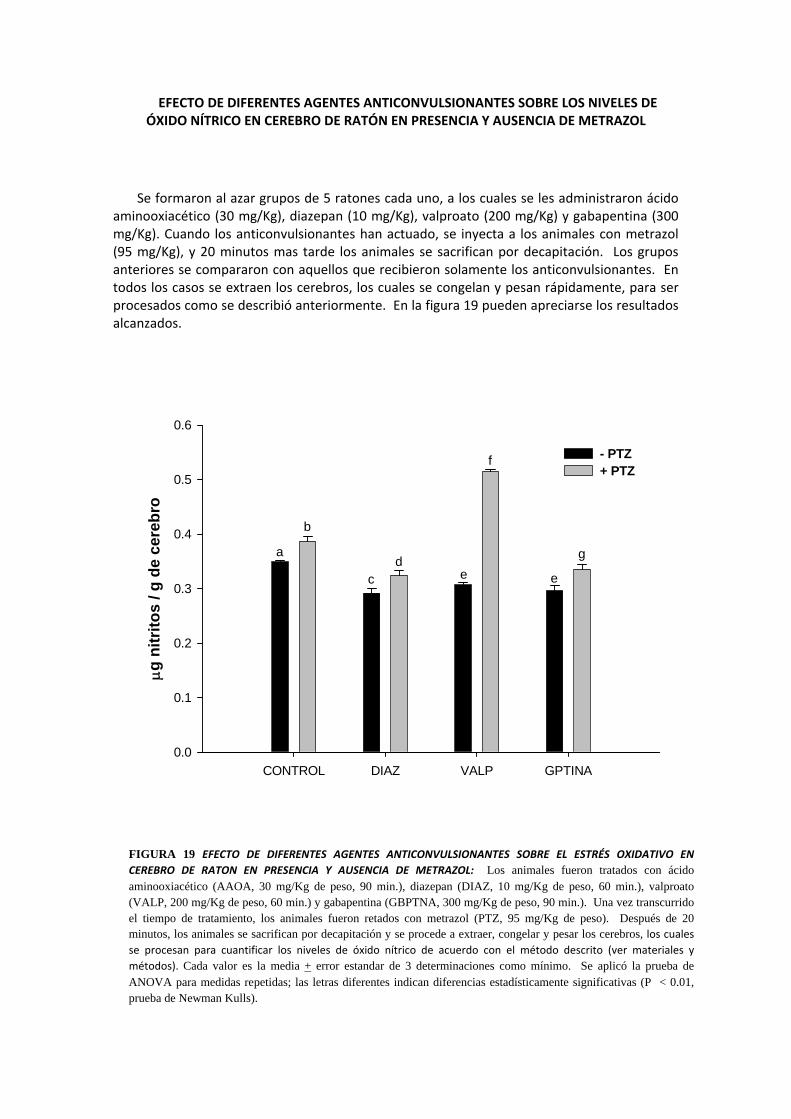
EFECTO DE DIFERENTES AGENTES ANTICONVULSIONANTES SOBRE LOS NIVELES DE ÓXIDO NÍTRICO EN CEREBRO DE RATÓN EN PRESENCIA Y AUSENCIA DE METRAZOL
Se formaron al azar grupos de 5 ratones cada uno, a los cuales se les administraron ácido aminooxiacético (30 mg/Kg), diazepan (10 mg/Kg), valproato (200 mg/Kg) y gabapentina (300 mg/Kg). Cuando los anticonvulsionantes han actuado, se inyecta a los animales con metrazol (95 mg/Kg), y 20 minutos mas tarde los animales se sacrifican por decapitación. Los grupos anteriores se compararon con aquellos que recibieron solamente los anticonvulsionantes. En todos los casos se extraen los cerebros, los cuales se congelan y pesan rápidamente, para ser procesados como se describió anteriormente. En la figura 19 pueden apreciarse los resultados alcanzados.
CONTROL DIAZ VALP GPTINA
μg n
itrito
s / g
de
cere
bro
0.0
0.1
0.2
0.3
0.4
0.5
0.6
- PTZ+ PTZ
a
b
cd
e
f
g
e
FIGURA 19 EFECTO DE DIFERENTES AGENTES ANTICONVULSIONANTES SOBRE EL ESTRÉS OXIDATIVO EN CEREBRO DE RATON EN PRESENCIA Y AUSENCIA DE METRAZOL: Los animales fueron tratados con ácido aminooxiacético (AAOA, 30 mg/Kg de peso, 90 min.), diazepan (DIAZ, 10 mg/Kg de peso, 60 min.), valproato (VALP, 200 mg/Kg de peso, 60 min.) y gabapentina (GBPTNA, 300 mg/Kg de peso, 90 min.). Una vez transcurrido el tiempo de tratamiento, los animales fueron retados con metrazol (PTZ, 95 mg/Kg de peso). Después de 20 minutos, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros, los cuales se procesan para cuantificar los niveles de óxido nítrico de acuerdo con el método descrito (ver materiales y métodos). Cada valor es la media + error estandar de 3 determinaciones como mínimo. Se aplicó la prueba de ANOVA para medidas repetidas; las letras diferentes indican diferencias estadísticamente significativas (P < 0.01, prueba de Newman Kulls).
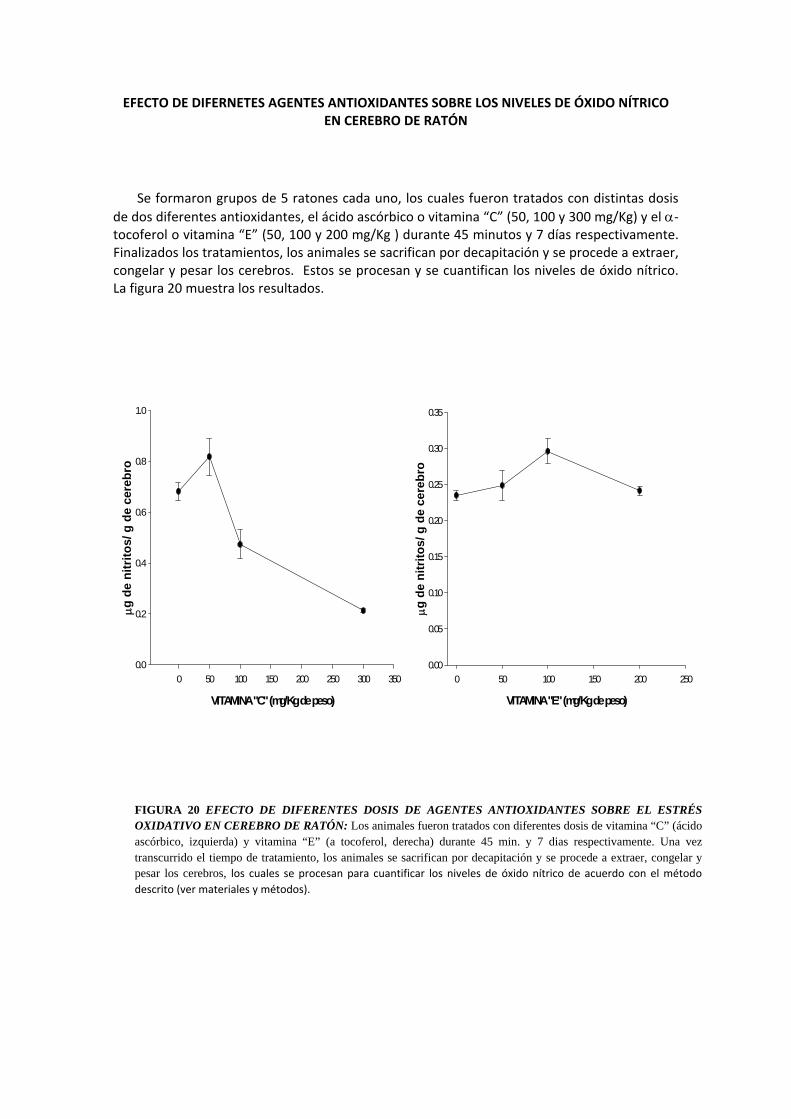
EFECTO DE DIFERNETES AGENTES ANTIOXIDANTES SOBRE LOS NIVELES DE ÓXIDO NÍTRICO EN CEREBRO DE RATÓN
Se formaron grupos de 5 ratones cada uno, los cuales fueron tratados con distintas dosis de dos diferentes antioxidantes, el ácido ascórbico o vitamina “C” (50, 100 y 300 mg/Kg) y el α‐tocoferol o vitamina “E” (50, 100 y 200 mg/Kg ) durante 45 minutos y 7 días respectivamente. Finalizados los tratamientos, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros. Estos se procesan y se cuantifican los niveles de óxido nítrico. La figura 20 muestra los resultados.
VITAMINA "C" (mg/Kg de peso)
0 50 100 150 200 250 300 350
μg d
e ni
trito
s/ g
de
cere
bro
0.0
0.2
0.4
0.6
0.8
1.0
VITAMINA "E" (mg/Kg de peso)
0 50 100 150 200 250
μg d
e ni
trito
s/ g
de
cere
bro
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
FIGURA 20 EFECTO DE DIFERENTES DOSIS DE AGENTES ANTIOXIDANTES SOBRE EL ESTRÉS OXIDATIVO EN CEREBRO DE RATÓN: Los animales fueron tratados con diferentes dosis de vitamina “C” (ácido ascórbico, izquierda) y vitamina “E” (a tocoferol, derecha) durante 45 min. y 7 dias respectivamente. Una vez transcurrido el tiempo de tratamiento, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros, los cuales se procesan para cuantificar los niveles de óxido nítrico de acuerdo con el método descrito (ver materiales y métodos).

EFECTO DE DIFERENTES AGENTES ANTIOXIDANTES SOBRE LOS NIVELES DE ÓXIDO NÍTRICO EN CEREBRO DE RATÓN EN PRESENCIA Y AUSENCIA DE METRAZOL
Se formaron grupos de 5 ratones cada uno, los cuales fueron tratados con distintas dosis de dos diferentes antioxidantes, el ácido ascórbico o vitamina “C” (50, 100 y 300 mg/Kg) y el α‐tocoferol o vitamina “E” (50, 100 y 200 mg/Kg ). Transcurridos 45 minutos y 7 días de tratamiento respectivamente se retó a los animales con 90 mg/Kg de metrazol (en el caso de la vitamina “E”, el 7º día de tratamiento se inyectó a los animales y 30 minutos después con el metrazól). Los grupos descritos se compararon con aquellos que recibieron únicamente las diferentes dosis de los antioxidantes. Finalizados los tratamientos, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros. Estos se procesan y se cuantifican los niveles de óxido nítrico. La figura 21 muestra los resultados.
VITAMINA "C" (mg/Kg de peso)
0 50 100 300
μg n
itrito
s / g
de
cere
bro
0.0
0.2
0.4
0.6
0.8
1.0
- PTZ+ PTZ
ab
c
d
ef
g g
VITAMINA "E" (mg/Kg)
0 50 100 200
μg n
itrito
s / g
de
cere
bro
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35- PTZ+ PTZ
a
b
cd
e
f
c
g
FIGURA 21 EFECTO DE DIFERENTES DOSIS DE AGENTES ANTIOXIDANTES SOBRE EL ESTRÉS OXIDATIVO EN CEREBRO DE RATON EN PRESENCIA Y AUSENCIA DE METRAZOL: Los animales fueron tratados con diferentes dosis de vitamina “C” (ácido ascórbico, izquierda) y vitamina “E” (a tocoferol, derecha) durante 45 minutos y 7 días respectivamente. Una vez transcurrido el tiempo de tratamiento, los animales fueron retados con metrazol (PTZ, 95 mg/Kg de peso). Después de 20 minutos, los animales se sacrifican por decapitación y se procede a extraer, congelar y pesar los cerebros, los cuales se procesan para cuantificar los niveles de óxido nítrico de acuerdo con el método descrito (ver materiales y métodos). Cada valor es la media
+ error estandar de 3 determinaciones como mínimo. Se aplicó la prueba de ANOVA para medidas repetidas; las letras diferentes indican diferencias estadísticamente significativas (P < 0.01, prueba de Newman Kulls).

DISCUSIÓN
Los resultados obtenidos muestran que, al menos en el caso del metrazol, las convulsiones inducidas por esta droga producen estrés oxidativo en el cerebro de ratón, como lo indican las modificaciones producidas en las proteínas que produce este fármaco como son el aumento de carbonilos libres y ditirosinas (figura 2). De hecho, se encontró que este incremento en el estrés oxidativo es dosis dependiente de acuerdo con los resultados mostrados por los carbonilos libres, aunque en el caso de las ditirosinas después de los 50 mg/Kg estas parecen decaer (figura 3).
La idea de que las convulsiones se asocian con un incremento en el estrés oxidativo, parece verse reforzada por el hecho de que el incremento en radicales libres y ditirosinas observado anteriormente no es exclusivo del metrazol, sino que también sucede cuando se emplean distintos convulsionantes que actúan por diferentes mecanismos, como puede apreciarse en la figura 4 (excepto en el caso de la estricnina, donde prácticamente no alteran los niveles de las ditirosinas, aunque si los carbonilos libres).
Otra evidencia de la importancia del estrés oxidativo en el mecanismo de las convulsiones la constituye el hecho de que cuando se protege a los animales de las convulsiones inducidas por los fármacos con aminooxiacético, el aumento en los niveles de estrés oxidativo parece revertirse. Así se tiene que, en el caso de los carbonilos libres, los incrementos producidos por los convulsionantes utilizados se antagonizan cuando se protege a los animales con el AAOA. Las ditiroinas muestran una disminución como resultado del pretratamiento con AAOA en el caso del PTZ y la BIC, pero no así para la TS y la estricnina (figura 5). Probablemente esto pueda explicarse por el hecho de que tanto el PTZ como la BIC actúan sobre receptores gabaérgicos tipo A.
Contrariamente a lo que sucede en el caso de los convulsionantes puede decirse que, en general, los anticonvulsionantes no afectan o bien disminuyen el estrés oxidativo. Así lo indican los resultados mostrados en la figura 6, donde puede verse que los carbonilos libres no modifican sus niveles o bien disminuyen (excepto en el caso del AAOA). En el caso de las ditirosinas, estas prácticamente no se alteran, pero el AAOA y el DIAZ producen un aumento significativo de las mismas. Cuando se reta con PTZ a los animales previamente protegidos con los anticonvulsionantes, se observa que el estrés oxidativo promovido por el convulsionante se revierte (aunque el DIAZ, a pesar de disminuir los carbonilos libres, aumenta las ditirosinas), como puede apreciarse en la figura 7. Cabría esperar que el AAOA disminuyera el estrés oxidativo como lo hace cuando se combina con los convulsionantes, pero tanto los carbonilos libres como las ditirosinas, indican que este fármaco produce el efecto contrario.
Para corroborar la importancia del estrés oxidativo en el mecanismo de las convulsiones se emplearon antioxidantes, con el propósito de evitar la generación del mismo y observar la influencia de estos compuestos. Los resultados indican que la vitamina “C” no pudo antagonizar el estrés oxidativo inducido por el PTZ a ninguna de las dosis empleadas (Figura 10). Por el contrario, la vitamina “E” si logra evitar la generación del estrés oxidativo inducido por el convulsionante con todas las dosis ensayadas (Figura 11).
Debe recordarse que el NO tiene una doble característica, el ser un radical libre además de un neurotransmisor, por lo que podría ser responsable, al menos en parte, del aumento de estrés oxidativo producido por las convulsiones que indican los resultados anteriores. Para averiguarlo, se determinó el efecto de las convulsiones inducidas por PTZ sobre los carbonilos libres y las ditirosinas tanto en presencia como en ausencia de L‐ARG y L‐NAME, promotor e inhibidor de la síntesis de NO respectivamente. Los resultados, consignados en la figura (12),

muestran que el estrés oxidativo producido por el PTZ aumenta en los animales pretratados con L‐ARG, mientras que disminuye cuando se administra L‐NAME. Cuando se hace el ensayo anterior, pero con diferentes dosis de PTZ, se encuentra que tanto los carbonilos libres como las ditirosinas muestran un efecto bifásico tanto de la L‐Arg como del L‐NAME, lo que probablemente se deba a los efectos de los fármacos sobre distintas isoformas de la NOS (figura 13).
Todo lo anterior sugiere que el NO esta relacionado con el estrés oxidativo generado por las convulsiones inducidas por el PTZ y podría explicar, al menos parcialmente, el estrés oxidativo que produce este convulsionante.
Una vez establecida la posible relación entre el estrés oxidativo y el NO, se determinaron los niveles del mismo de forma indirecta, estimados como nitratos convertidos en nitritos, en las mismas condiciones experimentales empleadas para el estrés oxidativo.
Los resultados muestran que, de manera semejante a lo que sucede con el estrés oxidativo, las convulsiones inducidas por el PTZ incrementan los niveles de óxido nítrico, los cuales prácticamente se reestablecen en los animales que sobreviven a las mismas (figura 13). Al igual que sucede en el estrés oxidativo, el incremento producido por el PTZ en los niveles de NO es dependiente de la dosis (fig. 15), y se presenta también con los demás convulsionanantes ensayados (fig. 16). Otra similitud entre el efecto de los convulsionantes sobre el estrés oxidativo y el NO se presenta cuando se protege a los animales con AAOA y se retan con los diferentes convulsionantes (figura 17), pues también en este caso se observa que el pretratamiento con AAOA consigue revertir el aumento en los niveles de NO producido por los convulsionantes (excepto en el caso de la estricnina). Por su parte, los anticonvulsionantes disminuyeron los niveles de NO (fig 18), es decir que tienen un efecto semejante al observado sobre el estrés oxidativo. Además, de acuerdo con lo mostrado en la figura 19, los anticovulsionantes consiguen también revertir el aumento en los niveles del NO inducido por el PTZ (excepto el valproato).
En cuanto a los antioxidantes, los resultados indican que la vitamina “C” disminuye los niveles de óxido nítrico de manera dosis dependiente (fig. 20), y consigue antagonizar el efecto del PTZ sobre los niveles de NO en las dosis de 100 y 300 mg/Kg de peso (fig. 21). En el caso de la vitamina “E” puede observarse que, a pesar de no disminuir a ninguna de las dosis empleadas los niveles de óxido nítrico (fig. 20), logra revertir los efectos del PTZ sobre este parámetro en cualquiera de las dosis empleadas (figura 21).
El aumento en los niveles de NO producido por el PTZ coincide con lo reportado por oros autores, (Osonoey col., 1994; Yokoyama y col, 1995; Han y col, 2000), sin embargo también existen publicaciones en sentido opuesto (Kata y col, 1998). En el curso de estos estudios se encontró que, hasta aproximadamente a una dosis de 80 mg/Kg de peso, la respuesta en cuanto al estrés oxidativo y los niveles de NO es muy reproducible. Sin embargo, a medida que se incrementa la dosis y con ello la mortalidad convulsiva, la respuesta se torna muy variada llegando a observarse incluso importantes disminuciones, lo que aparentemente se asocia con la sensibilidad de los animales a la droga. (datos no mostrados). Es decir, que el aumento en el estrés oxidativo y los niveles de NO parece relacionarse con la aparición de convulsiones y su duración, más que con la mortalidad convulsiva, lo que podría explicar lo contrastante de los reportes en la literatura.
Si se considera que el PTZ actúa impidiendo la transmisión GABAérgica a nivel de receptores tipo “A”, la TS actúa impidiendo la síntesis de GABA, pero a nivel enzimático (inhibiendo la DAG) y la bicuculina actúa también a nivel de receptores GABAérgicos tipo A, de forma colectiva los resultados parecen sugerir que alteraciones en el sistema GABAérgico (lo que finalmente se traduciría en una disminución de los niveles cerebrales de GABA), ocasionan aumentos en el estrés oxidativo que aparentemente se relacionan con un aumento en los

niveles de NO. Esto podría explicar el hecho de que la estricnina, que actúa a nivel de la médula espinal antagonizando competitivamente a la glicina en las células de Renshaw, o lo que es lo mismo, no actúa a través del sistema GABAérgico, muestre un incremento en los niveles de ditirosinas y que tampoco pueda revertir el efecto del PTZ sobre los niveles de NO.
Así, el hecho de que los efectos de los convulsionantes sobre el estrés oxidativo y los niveles de NO se reviertan cuando se protege a los animales con AAOA podría explicarse con base en el mecanismo de acción de este fármaco, cuyo efecto anticonvulsionante se atribuye al incremento que produce en los niveles de GABA producido por la inhibición de la transaminasa encargada del catabolismo de este neurotransmisor inhibidor (T‐GABA). Existen reportes que indican que el NO tiene la capacidad de inhibir la T‐GABA, lo que promueve el incremento en sus niveles de este neurotransmisor inhibidor (Jayakumar y col, 1999), además de promover su liberación en la corteza cerebral (Kuriyama y Ohkuma, 1995), hipocampo (Lonart y col, 1992) y estriatum (Segovia y Mora, 1998). De modo que el incremento en los niveles de NO, podría ser el mecanismo por el cual esta droga inhibe la T‐GABA y con ello promover el incremento en los niveles de GABA. Así mismo, esta acción del NO podría explicar la observación de que, a pesar de alcanzar un máximo los niveles de GABA a las 6 horas de administración del AAOA, este ya no tiene efecto anticonvulsionante, lo que indicaría que además del mero hecho de incrementar el GABA parece intervenir otro mecanismo distinto, que bien podría ser este efecto sobre el NO, poniendo de manifiesto el importante papel que éste podría desempeñar en el mecanismo de las convulsiones.
El DIAZ disminuye tanto el estrés oxidativo como los niveles de NO, lo cual coincide con lo reportado con otros autores, que indican que el NO podría participar en los efectos antiepilepticos de las benzodiacepinas (Talarek y Fidecka, 2003), y podría afirmar esta aparente relación entre el sistema GABAérgico y el NO. Esto explicaría que los resultados obtenidos con la GBPTINA sean similares, pero en cambio no exista esta correlación entre estrés oxidativo y niveles de NO en el caso del VALP, que actúa inactivando los canales de sodio.
El hecho de que la vitamina “C” aparentemente no disminuya el estrés oxidativo, pero si disminuya los niveles de NO y revierta el efecto del PTZ sobre estos, podría indicar, por una parte, que el efecto anticonvulsionantes atribuido a la vitamina “C” (MacGregor y col, 1996; Fighera y col, 1999; Yamamoto y col, 2002) no se debe a su acción antioxidante, sino que podría atribuirse a su efecto sobre los niveles de NO. Lo contrario parece presentarse en el caso de la vitamina “E”, que de acuerdo a nuestros resultados y lo consignado por otros autores (Ribeiro y col, 2005) impide el la formación de carbonilos libres inducida por el PTZ, pero solo a altas dosis revierte los efectos del PTZ sobre los niveles de NO.
En su conjunto, los resultados anteriores indican que durante los estados convulsivos inducidos por fármacos que actúan sobre el sistema GABAergico se genera estrés oxidativo que, al menos en parte, aparentemente es causado por el NO, el cual también parece estar relacionado con el efecto anticonvulsionante de fármacos cuyo mecanismo de acción se relaciona con el GABA.
Finalmente, cabe aclarar que una de las metas propuestas en este proyecto era la evaluación del estrés oxidativo y los niveles de oxidativo en pacientes epilépticos, pero los resultados técnicos llevaron a modificar lo planeado con anterioridad, mostrando la conveniencia de profundizar en los aspectos básicos, antes de continuar con las muestras de pacientes.

IMPACTO
Los resultados obtenidos en este proyecto indican que el NO podría desempeñar un importante papel en el mecanismo de las convulsiones, lo que sugiere que inhibidores de la óxido nítrico sintasa podrían tener actividad anticonvulsionante y un potencial uso terapéutico en los pacientes epilépticos. Si se logra diseñar inhibidores de la NOS lo suficientemente específicos, dependiendo de la isoforma de la NOS que participe, estos anticonvulsionantes podrían tener gran eficacia y menores efectos secundarios.
Por otra parte, el hecho de que durante los estados convulsivos se genere estrés oxidativo, sugiere que como complemento a los fármacos anticonvulsionantes empleados, sería conveniente el uso de antioxidantes como parte del tratamiento.
BIBLIOGRAFÍA
1. Buisson A., Lakhmeche N., Verrecchia C., Plotkine M. y Boulu R. G. (1993) Nitric
oxide: an endogenous anticonvulsant substance Neuroreport, 4, 444‐6 2. Coyle J. T. y Puttfarcken P. (1993) Oxidative stress, glutamate, and neurodegenerative
disorders Science, 262, 689‐95 3. Dalle‐Donne I., Rossi R., Giustarini D., Milzani A. y Colombo R. (2003) Protein
carbonyl groups as biomarkers of oxidative stress Clin Chim Acta, 329, 23‐38 4. Dawson P. S. (1994) Large animal pathology courses Vet Rec, 135, 47 5. De Sarro G. B., Donato Di Paola E., De Sarro A. y Vidal M. J. (1991) Role of nitric oxide
in the genesis of excitatory amino acid‐induced seizures from the deep prepiriform cortex Fundam Clin Pharmacol, 5, 503‐11
6. Engelborghs S., D'Hooge R. y De Deyn P. P. (2000) Pathophysiology of epilepsy Acta Neurol Belg, 100, 201‐13
7. Fighera M. R., Queiroz C. M., Stracke M. P., Brauer M. C., Gonzalez‐Rodriguez L. L., Frussa‐Filho R., Wajner M. y de Mello C. F. (1999) Ascorbic acid and alpha‐tocopherol attenuate methylmalonic acid‐induced convulsions Neuroreport, 10, 2039‐43
8. Gonzalez‐Hernandez T., Garcia‐Marin V., Perez‐Delgado M. M., Gonzalez‐Gonzalez M. L., Rancel‐Torres N. y Gonzalez‐Feria L. (2000) Nitric oxide synthase expression in the cerebral cortex of patients with epilepsy Epilepsia, 41, 1259‐68
9. Halliwell B G. j. (1999) Free radicals in biology and medicine, Oxford Univ. Press, NY. 10. Han D., Yamada K., Senzaki K., Xiong H., Nawa H. y Nabeshima T. (2000) Involvement
of nitric oxide in pentylenetetrazole‐induced kindling in rats J Neurochem, 74, 792‐8 11. Holtkamp M. y Meierkord H. (2007) Anticonvulsant, antiepileptogenic, and
antiictogenic pharmacostrategies Cell Mol Life Sci, 64, 2023‐41

12. Jayakumar A. R., Sujatha R., Paul V., Asokan C., Govindasamy S. y Jayakumar R. (1999) Role of nitric oxide on GABA, glutamic acid, activities of GABA‐T and GAD in rat brain cerebral cortex Brain Res, 837, 229‐35
13. Kata M., Baran L., Siwanowicz J. y Dziedzicka‐Wasylewska M. (1998) Effect of kainic acid and pentetrazole administration on the activity of nitric oxide synthase in the mouse brain Pol J Pharmacol, 50, 259‐64
14. Kiss J. P. y Vizi E. S. (2001) Nitric oxide: a novel link between synaptic and nonsynaptic transmission Trends Neurosci, 24, 211‐5
15. Kuriyama K. y Ohkuma S. (1995) Role of nitric oxide in central synaptic transmission: effects on neurotransmitter release Jpn J Pharmacol, 69, 1‐8
16. Lonart G., Wang J. y Johnson K. M. (1992) Nitric oxide induces neurotransmitter release from hippocampal slices Eur J Pharmacol, 220, 271‐2
17. MacGregor D. G., Higgins M. J., Jones P. A., Maxwell W. L., Watson M. W., Graham D. I. y Stone T. W. (1996) Ascorbate attenuates the systemic kainate‐induced neurotoxicity in the rat hippocampus Brain Res, 727, 133‐44
18. Malencik D. A., Sprouse J. F., Swanson C. A. y Anderson S. R. (1996) Dityrosine: preparation, isolation, and analysis Anal Biochem, 242, 202‐13
19. Moncada S., Palmer R. M. y Higgs E. A. (1991) Nitric oxide: physiology, pathophysiology, and pharmacology Pharmacol Rev, 43, 109‐42
20. Nidhi G., Balakrishnan S. y Pandhi P. (1999) Role of nitric oxide in electroshock and pentylenetetrazole seizure threshold in rats Methods Find Exp Clin Pharmacol, 21, 609‐12
21. Osonoe K., Mori N., Suzuki K. y Osonoe M. (1994) Antiepileptic effects of inhibitors of nitric oxide synthase examined in pentylenetetrazol‐induced seizures in rats Brain Res, 663, 338‐40
22. Patel M. N. (2002) Oxidative stress, mitochondrial dysfunction, and epilepsy Free Radic Res, 36, 1139‐46
23. Przegalinski E., Baran L. y Siwanowicz J. (1996) The role of nitric oxide in chemically‐ and electrically‐induced seizures in mice Neurosci Lett, 217, 145‐8
24. Ribeiro M. C., de Avila D. S., Schneider C. Y., Hermes F. S., Furian A. F., Oliveira M. S., Rubin M. A., Lehmann M., Krieglstein J. y Mello C. F. (2005) alpha‐Tocopherol protects against pentylenetetrazol‐ and methylmalonate‐induced convulsions Epilepsy Res, 66, 185‐94
25. Segovia G. y Mora F. (1998) Role of nitric oxide in modulating the release of dopamine, glutamate, and GABA in striatum of the freely moving rat Brain Res Bull, 45, 275‐9
26. Snyder S. H. y Bredt D. S. (1991) Nitric oxide as a neuronal messenger Trends Pharmacol Sci, 12, 125‐8
27. Starr M. S. y Starr B. S. (1993) Paradoxical facilitation of pilocarpine‐induced seizures in the mouse by MK‐801 and the nitric oxide synthesis inhibitor L‐NAME Pharmacol Biochem Behav, 45, 321‐5
28. Talarek S. y Fidecka S. (2003) Role of nitric oxide in anticonvulsant effects of benzodiazepines in mice Pol J Pharmacol, 55, 181‐91
29. Theard M. A., Baughman V. L., Wang Q., Pelligrino D. A. y Albrecht R. F. (1995) The role of nitric oxide in modulating brain activity and blood flow during seizure Neuroreport, 6, 921‐4
30. Tsuda M., Suzuki T. y Misawa M. (1997) Aggravation of DMCM‐induced seizure by nitric oxide synthase inhibitors in mice Life Sci, 60, PL339‐43
31. Van Leeuwen R., De Vries R. y Dzoljic M. R. (1995) 7‐Nitro indazole, an inhibitor of neuronal nitric oxide synthase, attenuates pilocarpine‐induced seizures Eur J Pharmacol, 287, 211‐3

32. Verdon C. P., Burton B. A. y Prior R. L. (1995) Sample pretreatment with nitrate reductase and glucose‐6‐phosphate dehydrogenase quantitatively reduces nitrate while avoiding interference by NADP+ when the Griess reaction is used to assay for nitrite Anal Biochem, 224, 502‐8
33. Vezzani A., Conti M., De Luigi A., Ravizza T., Moneta D., Marchesi F. y De Simoni M. G. (1999) Interleukin‐1beta immunoreactivity and microglia are enhanced in the rat hippocampus by focal kainate application: functional evidence for enhancement of electrographic seizures J Neurosci, 19, 5054‐65
34. Vezzani A. y Granata T. (2005) Brain inflammation in epilepsy: experimental and clinical evidence Epilepsia, 46, 1724‐43
35. White H. S. (1997) Clinical significance of animal seizure models and mechanism of action studies of potential antiepileptic drugs Epilepsia, 38 Suppl 1, S9‐17
36. Wu G. Y. y Brosnan J. T. (1992) Macrophages can convert citrulline into arginine Biochem J, 281 ( Pt 1), 45‐8
37. Yamamoto N., Kabuto H., Matsumoto S., Ogawa N. y Yokoi I. (2002) alpha‐Tocopheryl‐L‐ascorbate‐2‐O‐phosphate diester, a hydroxyl radical scavenger, prevents the occurrence of epileptic foci in a rat model of post‐traumatic epilepsy Pathophysiology, 8, 205‐214
38. Yokoyama H., Mori N., Osonoe K., Kasai N., Hiramatsu M., Yoshimura T., Matsue T., Uchida I., Kobayashi N., Tsuchihashi N. y et al. (1995) Increase of central nitric oxide during pentylenetetrazol‐induced seizures in rats Psychiatry Clin Neurosci, 49, S277‐9