infectious diseases of wild birds
TRANSCRIPT

InfectiousDiseases of Wild Birds
Edited by Nancy J. ThomasD. Bruce Hunter
Carter T. Atkinson


Infectious Diseasesof Wild Birds


InfectiousDiseases of Wild Birds
Edited by Nancy J. ThomasD. Bruce Hunter
Carter T. Atkinson

Nancy J. Thomas, DVM, MS, Diplomate, AmericanCollege of Veterinary Pathologists, is an endangeredspecies specialist for the U.S. Geological Survey,National Wildlife Health Center. She has more than20 years of experience in diagnostic pathology andresearch on wild bird diseases.
D. Bruce Hunter DVM, MSC. is a professor in theDepartment of Pathobiology, Ontario VeterinaryCollege, University of Guelph. His specialtyencompasses avian, fur-bearing, and wildlifepathology and ecosystem health. His research interestshave included infectious diseases of commercialpoultry, West Nile Virus in owls, and diseases of mink.
Carter T. Atkinson, Ph.D., is a research microbiolo-gist with the U.S. Geological Survey, Paci c IslandEcosystems Research Center. His research focuses onprotozoan parasites of vertebrates, particularly birds,with a recent emphasis on the effects of introducedavian malaria on Hawaiian forest birds.
'2007 Blackwell PublishingChapters 2, 4, 6, 12, 14, 17, 20, and 21 remain withthe U.S. Government
Blackwell Publishing Professional2121 State Avenue, Ames, Iowa 50014, USA
Orders: 1-800-862-6657Office: 1-515-292-0140Fax: 1-515-292-3348Web site: www.blackwellprofessional.com
Blackwell Publishing Ltd9600 Garsington Road, Oxford OX4 2DQ, UKTel.: +44 (0)1865 776868
Blackwell Publishing Asia550 Swanston Street, Carlton, Victoria 3053,AustraliaTel.: +61 (0)3 8359 1011
Authorization to photocopy items for internal orpersonal use, or the internal or personal use ofspeci c clients, is granted by Blackwell Publishing,provided that the base fee is paid directly to theCopyright Clearance Center, 222 Rosewood Drive,Danvers, MA 01923. For those organizations thathave been granted a photocopy license by CCC, aseparate system of payments has been arranged. Thefee code for users of the Transactional ReportingService is ISBN-13: 978-0-8138-2812-1.
Library of Congress Cataloging-in-Publication DataInfectious diseases of wild birds / edited by Nancy J.Thomas, D. Bruce Hunter, Carter T. Atkinson.
p. cm.Includes bibliographical references and index.
ISBN-13: 978-0-8138-2812-1 (alk. paper)ISBN-10: 0-8138-2812-0 (alk. paper)1. Birds Infections. 2. Wildlife diseases.
3. Communicable diseases in animals. I. Thomas, Nancy J. (Nancy Jeanne), 1948—. II.Hunter, D. Bruce III. Atkinson, Carter T.
SF994.I54 2007636.5 08969 dc22
2006021665
The last digit is the print number: 9 8 7 6 5 4 3 2 1

v
Preface viiContributors ix
Section 1: Viral Diseases 1
1 Newcastle Disease and Related Avian Paramyxoviruses 3F.A. Leighton, R.A. Heckert
2 Arboviruses in Birds 17R.G. McLean, S.R. Ubico
3 Avian Herpesviruses 63E.F. Kaleta, D.E. Docherty
4 Duck Plague (Duck Virus Enteritis) 87W.R. Hansen, R.E. Gough
5 Avian In uenza 108D.E. Stallknecht, E. Nagy, D.B. Hunter, R.D. Slemons
6 Avian Pox 131C. van Riper, D.J. Forrester
7 Orthoreoviruses 177T. Hollm n, D.E. Docherty
8 Avian Adenoviruses 182S.D. Fitzgerald
9 Circovirus 194J.A. Par , N. Robert
10 Papillomaviruses and Polyomaviruses 206D.N. Phalen
11 Retroviral Infections 216M.L. Drew
Section 2: Bacterial and Fungal Diseases 237
12 Avian Cholera 239M.D. Samuel, R.G. Botzler, G.A. Wobeser
13 Salmonellosis 270P. Daoust, J.F. Prescott
14 Avian Tuberculosis 289K.A. Converse
15 Avian Chlamydiosis 303A.A. Andersen, J.C. Franson
16 Mycoplasmosis 317P. Luttrell, J.R. Fischer
17 Erysipelas 332M.J. Wolcott
Contents

Contentsvi
18 Borrelia 341B. Olsen
19 Tularemia 352T. M rner
20 Aspergillosis 360K.A. Converse
Section 3: Biotoxins 375
21 Avian Botulism 377T.E. Rocke, T.K. Bollinger
22 Mycotoxicosis 417C.F. Quist, T. Cornish, R.D. Wyatt
23 Algal Biotoxins 431J.H. Landsberg, G.A. Vargo, L.J. Flewelling, F.E. Wiley
Index 457

vii
At its inception, this book was a revision of Infectiousand Parasitic Diseases of Wild Birds, edited by JohnW. Davis, Roy C. Anderson, Lars Karstad, and DanielO. Trainer and published by the Iowa State UniversityPress in 1971. Advances in the eld and the volume ofresulting material led us to expand the original workand necessitated launching this work as a new volumesolely devoted to infectious diseases of wild, free-living birds. Nevertheless, this book is patterned afterthe original volume that has been the mainstay of wildbird disease study, despite its dated condition. Thisbook is planned as a companion to Infectious Diseasesof Wild Mammals, 3rd Edition, edited by Elizabeth S.Williams and Ian K. Barker, and Parasitic Diseases ofWild Mammals, 2nd Edition, edited by William M.Samuel, Margo J. Pybus, and A. Alan Kocan (IowaState University Press). We gratefully acknowledgeour colleagues who established such excellent modelsfor us to follow.
This book focuses on diseases affecting free-livingwild birds and the agents that cause them. Relevantinformation and examples are drawn from captivebirds or poultry in order to ll in gaps in data or toprovide lessons for managers of captive-rearingprograms, as captive reintroduction programs arebecoming increasingly important for supplementingwild populations of threatened and endangeredspecies. Biologists and wildlife managers, wildlifeand veterinary students, professionals in the elds ofanimal health and wildlife disease, and evolutionarybiologists with interests in disease ecology should allnd this book to be a valuable reference. The chapterscover classical waterfowl diseases, such as aviancholera, botulism, and poultry disease agents that havetaken on new dimensions in wild birds (Newcastledisease, mycoplasmosis, and duck plague). New dis-eases (circoviral, papilloma and polyomaviral dis-eases) have risen since the original volume in 1971,and some older diseases, such as avian in uenza,haveacquired global signi cance in new zoonotic forms.Included among the chapters are disease agents that are less signi cant to wild bird health but areimportant to human health, in which wild birds play
an important role in the epizootiologic cycle (certainarboviruses and Borrelias).
The chapter authors were selected for their expertiseand familiarity with the agents, disease processes, andeffects on wild bird populations. This book is thecumulative product of their considerable knowledgeand experience. Each chapter provides a classicaldescription of the history, disease, and causative agent,but the authors were also challenged to provide per-spectives on the signi cance of the disease to wildbirds and to document population impacts, an aspectthat is particularly difficult to quantify in the wild.Chapters concentrate more on the disease processes,recognition, and epizootiologic factors than ontreatment. Authors were encouraged to identify unre-solved questions and to provide balanced reviews ofcontroversies. It is inevitable that rapid advances inknowledge and the fast pace of environmental changesin todays world will quickly render some aspects ofthis book outdated, but the authorstreatments of rap-idly evolving diseases like avian in uenza and WestNile virus are state of the art at this time. The author-ity for avian nomenclature, both scienti c and Englishnames, was the American Ornithologists UnionChecklist of North American Birds, 7th Edition (http://www.aou.org/checklist/), supplemented for anyunlisted species, by James F. Clements Birds of theWorld: A Checklist (Ibis Publishing Company, 2000).Because many unpublished data on wild bird diseaseshave been compiled in laboratory and diagnostic les,citations of unpublished data were allowed for reposi-tories of large, permanent, accessible institutions, suchas the Canadian Cooperative Wildlife Health Centre,USGS National Wildlife Health Center, and Southeast-ern Cooperative Wildlife Disease Study.
Grateful acknowledgement goes to Donald J.Forrester, University of Florida, the Iowa State Uni-versity Press, who guided this project through its ini-tial stages, and to Blackwell Publishing, who took itover and shepherded it through to completion. We alsothank Daina Hunter for her signi cant contribution inthe technical editing of this book. We acknowledgethe support of the U.S. Geological Survey, Wildlife
Preface

and Terrestrial Resources Program and the Universityof Guelph. This book is dedicated to the Wildlife Dis-ease Association, whose members initiated the revi-sion of this book series and who continue to providethe backbone of growing knowledge in the eld of
wildlife disease. Royalties that accrue from sales ofthis book will be provided to the Wildlife DiseaseAssociation.
Nancy J. ThomasD. Bruce Hunter
Carter T. Atkinson
Prefaceviii

ix
Arthur A. AndersenNational Animal Disease CenterAgricultural Research ServiceU.S. Department of AgricultureAmes, Iowa, U.S.A.
Trent K. BollingerCanadian Cooperative Wildlife Health CentreWestern College of Veterinary MedicineDepartment of Veterinary PathologyUniversity of SaskatchewanSaskatoon, Saskatchewan, Canada
Richard G. BotzlerDepartment of WildlifeHumboldt State UniversityArcata, California, U.S.A.
Kathryn A. ConverseU.S. Geological SurveyNational Wildlife Health CenterMadison, Wisconsin U.S.A.
Todd CornishWyoming State Veterinary Diagnostic LaboratoryDepartment of Veterinary ScienceUniversity of WyomingLaramie, Wyoming, U.S.A.
Pierre-Yves DaoustCanadian Cooperative Wildlife Health CentreDepartment of Pathology and MicrobiologyAtlantic Veterinary CollegeUniversity of Prince Edward IslandCharlottetown, Prince Edward Island, Canada
Douglas E. DochertyU.S. Geological SurveyNational Wildlife Health CenterMadison, Wisconsin, U.S.A.
Mark L. DrewIdaho Department of Fish and GameWildlife Health LaboratoryCaldwell, Idaho, U.S.A.
John R. FischerSoutheastern Cooperative Wildlife Disease Study, andDepartment of Population HealthCollege of Veterinary MedicineThe University of GeorgiaAthens, Georgia, U.S.A.
Scott D. FitzgeraldDepartment of Pathobiology and DiagnosticInvestigationCollege of Veterinary MedicineMichigan State UniversityLansing, Michigan, U.S.A.
Leanne J. FlewellingFish and Wildlife Research InstituteFlorida Fish and Wildlife Conservation CommissionSt. Petersburg, Florida, U.S.A.
Donald J. ForresterDepartment of Infectious Diseases and PathologyCollege of Veterinary MedicineUniversity of FloridaGainesville, Florida, U.S.A.
J. Christian FransonU.S. Geological SurveyNational Wildlife Health CenterMadison, Wisconsin U.S.A.
Richard E. GoughAvian VirologyVeterinary Laboratories Agency (Weybridge)New Haw, Addlestone, Surrey, United Kingdom
Wallace R. HansenU.S. Geological SurveyNational Wildlife Health CenterMadison, Wisconsin, U.S.A.
Robert A. HeckertU.S. Department of AgricultureAgricultural Research ServiceAnimal Health National ProgramBeltsville, Maryland, U.S.A.
Contributors

Tuula Hollm nAlaska SeaLife Center, andUniversity of Alaska FairbanksSchool of Fisheries and Ocean SciencesSeward, Alaska, U.S.A.
D. Bruce HunterDepartment of PathobiologyOntario Veterinary CollegeUniversity of GuelphGuelph, Ontario, Canada
Erhard F. KaletaKlinik f r V gel, Reptilien, Amphibien und FischeJustus-Liebig-Universit t GiessenGiessen, Germany
Jan H. LandsbergFish and Wildlife Research InstituteFlorida Fish and Wildlife Conservation CommissionSt. Petersburg, Florida, U.S.A.
Frederick A. LeightonCanadian Cooperative Wildlife Health CentreDepartment of Veterinary PathologyWestern College of Veterinary MedicineUniversity of SaskatchewanSaskatoon, Saskatchewan, Canada
Page LuttrellSoutheastern Cooperative Wildlife Disease Study, andDepartment of Population HealthCollege of Veterinary MedicineThe University of GeorgiaAthens, Georgia, U.S.A.
Robert G. McLeanU.S. Department of AgricultureAnimal and Plant Health Inspection Service-WildlifeServicesNational Wildlife Research CenterFort Collins, Colorado, U.S.A.
Torsten M r nerDepartment of Wildlife, Fish and EnvironmentNational Veterinary InstituteUppsala, Sweden
Eva NagyDepartment of PathobiologyOntario Veterinary CollegeUniversity of GuelphGuelph, Ontario, Canada
Bj r n OlsenDepartment of Infectious DiseasesUme Uni versityUme , Sweden andBiology and Environmental SciencesInstitute for Zoonotic Ecology and EpidemiologyKalmar UniversityKalmar, Sweden
Jean ParAnimal Health CentreToronto ZooScarborough, Ontario, Canada
David N. PhalenWildlife Health and Conservation CentreCamden, New South Wales, Australia
John F. PrescottDepartment of PathobiologyOntario Veterinary CollegeUniversity of GuelphGuelph, Ontario, Canada
Charlotte F. QuistWildlife Health Associates, Inc.Dillon, Montana, U.S.A.
Charles van Riper IIIU.S. Geological SurveySouthwest Biological Science CenterSonoran Desert Field StationThe University of ArizonaTucson, Arizona, U.S.A.
Nadia RobertCenter for Fish and Wildlife HealthInstitute for Animal PathologyUniversity of BerneBerne, Switzerland
Tonie E. RockeU.S. Geological SurveyNational Wildlife Health CenterMadison, Wisconsin, U.S.A.
Michael D. SamuelU.S. Geological Survey (USGS)National Wildlife Health Center, andUSGS-Wisconsin Cooperative Wildlife Research CenterUniversity of WisconsinMadison, Wisconsin, U.S.A.
Contributorsx

Richard D. SlemonsDepartment of Veterinary Preventive MedicineCollege of Veterinary MedicineThe Ohio State UniversityColumbus, Ohio, U.S.A.
David E. StallknechtSoutheastern Cooperative Wildlife Disease Study, and Department of Population HealthCollege of Veterinary MedicineThe University of GeorgiaAthens, Georgia, U.S.A.
Sonya R. UbicoAtitlan Green and WildFort Collins, Colorado, U.S.A.
Gabriel A. VargoCollege of Marine ScienceUniversity of South FloridaSt. Petersburg, Florida, U.S.A.
Faith E. WileyDepartment of Forestry and Natural ResourcesClemson UniversityClemson, South Carolina, U.S.A.
Gary A. WobeserCanadian Cooperative Wildlife Health CentreDepartment of Veterinary PathologyWestern College of Veterinary MedicineUniversity of SaskatchewanSaskatoon, Saskatchewan, Canada
Mark J. WolcottField Operations and Training Branch, andSpecial Pathogens BranchU.S. Army Medical Research Institute of InfectiousDiseasesFort Detrick, Maryland, U.S.A.
Roger D. WyattDepartment of Poultry ScienceUniversity of GeorgiaAthens, Georgia, U.S.A.
Contributors xi


Section 1:Viral Diseases


3
INTRODUCTIONSeveral related paramyxoviruses infect and causedisease in wild and domestic birds. Of these, avianparamyxovirus type 1 (APMV-1), also known asNewcastle Disease virus, is the best studied. Muchless is known about the other eight avian paramyx-oviruses (APMV-2 to APMV-9). This chapter is con-cerned primarily with APMV-1 and the disease itcauses: Newcastle Disease (ND). However, someinformation on the other avian paramyxoviruses alsois provided.
Newcastle Disease virus is widespread among sev-eral different taxonomic groups of wild birds, andappears capable of infecting all species of birds andsome other vertebrates, including humans. ND hascaused substantial mortality in free-ranging popula-tions of Double-crested Cormorants (Phalacrocoraxauritus) and Rock Pigeons (Columba livia), andamong psittacine birds and other tropical species cap-tured and shipped internationally in the pet bird trade.All of these species have been sources of infection fordomestic poultry. ND is one of the most economicallyimportant diseases of domestic poultry world-wide.Newcastle Disease and Avian In uenza are the onlytwo diseases of birds included by the World Organi-zation for Animal Health (formerly OIE) among the15 infectious diseases deemed most economicallyimportant to international trade in animals and animalproducts (World Organization for Animal Health2004). There are many strains of ND virus, and theyvary greatly in their capacity to cause disease in differ-ent bird species. Infection may be entirely unapparentor may result in disease that can range from mild torapidly fatal. Thus, the term Ne wcastle Diseasevirus, in its various contexts, refers to a complex ofmany virus strains, global in distribution, that infect awide range of avian hosts and that manifest them-selves very differently in different settings.
SYNONYMSAPMV-1:Newcastle Disease, pseudo-fowl pest,pseudovogel-pest, atypische gefugelpest, pseudo-poultry plague, avian pest, avian distemper, Ranikhetdisease, Tetelo disease, Korean fowl plague, avianpneumoencephalitisAPMV-2: Yucaipa virusAPMV-5: Kunitachi virus
In general, the terms Ne wcastle Disease virusand Avian Paramyxovirus Type 1 (APMV-1) are syn-onyms. However, this broad application of the nameNe wcastle Disease has proved to be problematic forgovernment regulatory veterinary agencies that seek toregulate only those particular strains of the virus thatcause signi cant disease in commercial poultry. Thus, inthe context of international trade in poultry and poultryproducts, the names Ne wcastle Disease and Ne wcas-tle Disease virus are reserved exclusively for strains ofAPMV-1 that are highly pathogenic for domestic chick-ens. Scientists who do not work within this regulatoryframework do not often adhere to this restricted use ofthe name Ne wcastle Disease, and more often use theterms APMV-1 and ND virus synonymously.
In the past, diseases in birds caused by avian paramyx-oviruses other than APMV-1 also may have been calledNewcastle Disease. As of this writing, nine differentavian paramyxoviruses are now recognized. Virologicaltechniques to distinguish some of these viruses fromAPMV-1 have been available for a relatively short periodof time. Thus, some reports in the literature of ND virusmay have been of diseases and viruses that were not ND(Kaleta and Baldauf 1988; Alexander 2000a).
HISTORYThe earliest record of Newcastle Disease virus inwild birds may be from 1897 in Great Cormorants
1Newcastle Disease and Related
Avian Paramyxoviruses
Frederick A. Leighton and Robert A. Heckert

(Phalacrocorax carbo) and European Shags (Pha-lacrocorax aristotelis) in Scotland (MacPherson1956; Kuiken 1999). This inference is made from apoem in Scottish Gaelic, Call nan cearc ( The Loss ofthe Hens ) that recounts a die-of f of domestic chick-ens that was strikingly similar to an epidemic of NDthat occurred in the same coastal locations in Scotlandand in Ireland in 1949—1951. The source of infectionfor the hens in the 1949—1951 epizootic w as deter-mined to be the two cormorant species, which regu-larly were hunted for food and from which offal wasfed to chickens. In the 1949—1951 outbreak, ND viruswas isolated from European Shags and also from aNorthern Gannet (Morus bassanus), and one GreatCormorant was found with a high serological titre toND virus, indicating recent infection (Wilson 1950;Blaxland 1951; MacPherson 1956). In this mid-century outbreak, there was no evidence that the cor-morants suffered disease because of their infection,but lesions that might have been due to ND wereobserved in the gannet. Whether the ND virus in thisoutbreak originated from infected domestic poultrycarried to wild bird populations by scavenging gulls orwas enzootic in the wild seabird populations them-selves has been discussed but not resolved (Kuiken1999; MacPherson 1956). About 20% of GreatCormorants sampled in eastern France from 1997 to1999 were serologically positive to one or morestrains of ND virus (Artois et al. 2002).
From the 1950s onward, numerous serological sur-veys of free-ranging and captive wild birds wereundertaken, some accompanied by attempts at virusisolation. Results of these surveys showed that expo-sure to ND viruses was widespread, particularly amongfree-living waterfowl. Published reports of infectionof wild birds with ND virus were compiled by Palmerand Trainer (1971) and Kaleta and Baldauf (1988), thelatter recording infection in 241 different species ofbirds encompassing 27 taxonomic Orders. Severalstrains of the virus were isolated from wild waterfowl;these were of very low virulence to poultry and didnot appear to cause disease in the source species(Palmer and Trainer 1971; Vickers and Hanson 1982;Vickers and Hanson 1980; Alexander 1988b; Kaletaand Baldauf 1988; Stallknecht et al. 1991). Nonethe-less, from circumstantial evidence it has been inferredthat wild waterfowl may have been responsible forspread of a strain of ND virus highly pathogenic tochickens across Europe in 1996—1997 (Ale xanderet al. 1998).
An epizootic of ND emerged in Rock Pigeons inEurope in the 1980s (Vindevogel and Duchatel 1988).The disease was recognized and followed primarilyin domestic racing pigeons (domesticated RockPigeons). Its occurrence in wild populations is notwell documented. The epizootic may have begun in
the Middle East in the 1970s and spread westward andthen around the world. The strain of ND virus respon-sible for this epizootic is distinguishable from otherstrains by antigenic and molecular criteria, and gener-ally is at least moderately pathogenic in chickens. Thevirus infected Rock Pigeons inhabiting grain storagefacilities in England and was spread to commercialpoultry when virus from the pigeons was incorporatedinto poultry feeds made from contaminated grains(Alexander et al. 1984; Alexander et al. 1985).
Epizootic ND with high rates of morbidity andmortality was observed in young-of-the-year (YOY)Double-crested Cormorants in Canada in 1990 andsubsequently recurred both in Canada and the UnitedStates throughout that decade (Wobeser et al. 1993;Kuiken et al. 1998b; Kuiken 1999). This is the onlywild bird species in which large-scale mortality fromND has been recognized; reported mortality hasranged from <1% to 92% of YOY per affected colony.ND virus also was isolated from one American WhitePelican (Pelecanus erythrorhynchos), one CaspianTern (Sterna caspia) and one Ring-billed Gull (Larusdelawarensis) during these epizootics. A single strainof ND virus was isolated consistently from these out-breaks and was highly pathogenic for chickens. Thissame strain of ND virus spread from wild cormorantsto one commercial turkey ock in the United States in1992 (Heckert et al. 1996).
In domestic poultry, Newcastle Disease was rstrecognized in the mid-1920s more or less simultane-ously at locations that are currently within India, SriLanka, Indonesia, Korea, Japan and England. The nameof the disease derives from the description of theoutbreak in 1926 at Newcastle-Upon-Tyne, England,by T.M. Doyle (Doyle 1927). Three possible explana-tions for the emergence of these ND virus strainshighly pathogenic to chickens have been advanced:1) that these ND virus strains existed historically indomestic chickens in southeast Asia, and that NDemerged as a major international disease of poultrywhen large-scale commercial poultry farming andrapid international trade developed in the rst half ofthe twentieth century; 2) that these strains of NDviruses were enzootic among wild birds in tropicalrain forests and spread to poultry when human settle-ment intruded into their natural habitat; and 3) thatthese strains of ND viruses arose directly by mutationfrom the many strains of low pathogenicity to chick-ens found in wild birds. Alexander (2000a) consideredthe rst explanation likely, the second unlikely, andthe third at least possible.
The rst world-wide occurrence of ND in poultrylasted from the mid-1920s to the early 1960s. A sec-ond world-wide epizootic in poultry occurred from1969 to 1973, and a regional epizootic occurred inWestern Europe throughout the 1990s.
Infectious Diseases of Wild Birds4

Avian Paramyxovirus serotypes 2 to 9 were describedin domestic or wild birds between 1956 and 1978, asindicated in Table 1.1. None of these viruses has beenrecognized to cause disease in free-living wild birds, butall are assumed to persist in wild bird populations.APMV serotypes 2, 3, 6, and 7 have produced disease indomestic poultry (Alexander 2000).
DISTRIBUTIONND viruses are worldwide in distribution, but detailedknowledge of the distribution of the many differentstrains of the virus in different host species is fragmen-tary and incomplete. Natural migration in many groupsof wild birds and translocation by humans of wild anddomestic birds or bird products occur on so rapid andglobal a scale that any strain of ND virus has the poten-tial to infect wild or domestic birds in all parts of theplanet. Any attempt to de ne the geographic distribu-tion of ND viruses at one point in time would be radi-cally out of date in a very short while. Antibodies to
ND virus have been detected in Antarctic penguinsand arctic-nesting geese (Morgan and Westbury 1981;Bradshaw and Trainer 1966). Small poultry ocksin southeast Asia, Central America, parts of SouthAmerica, and perhaps parts of Africa probably are theprincipal reservoirs of ND virus strains highly patho-genic to chickens (Alexander 2000a). Double-crestedCormorants, which range across the full width ofNorth America and from the Canadian boreal forestsouth to Mexico, and Rock Pigeons world-wide appearto maintain within their populations virus strains path-ogenic both to themselves and to other species, includ-ing domestic poultry. Too little is known about theother avian paramyxoviruses to make an accuratestatement about their geographic distributions.
HOST RANGEAll species of birds probably can be infected with oneor more of the strains of ND virus. Most infections areasymptomatic and do not result in disease. Infection
Newcastle Disease 5
Table 1.1. Avian paramyxovirus serotypes.
Related diseases inPrototype virus Common hosts Other hosts Poultry
PMV-1 (Newcastle disease Many different Spectrum of diseasevirus) avian species
PMV-2/chicken/California/ turkeys, chickens, Respiratory disease,Yucaipa/56 passerines psittacines, egg production losses,
rails serious if complicated
PMV-3/turkey/Wisconsin/68 turkeys only none egg production losses,respiratory disease
PMV-3/parakeet/Netherlands/ psittacines, none No infections known449/75 passerines
PMV-4/duck/Hong Kong/D3/75 ducks geese Inapparent infections incommercial ducks
PMV-5/budgerigar/Japan/ Budgerigars lorikeets No infections knownKunitachi/75
PMV-6/duck/Hong Kong/199/77 ducks geese, turkeys, Inapparent in ducksrails and geese, respiratory
disease and egg losses in turkeys
PMV-7/dove/Tennessee/4/75 pigeons, doves turkeys, ostriches No infections known
PMV-8/goose/Delaware/1053/75 ducks, geese No infections known
PMV-9/duck/New York/22/78 ducks Inapparent infections incommercial ducks
Source: Alexander, D. J. 1997 (with permission).

with ND virus has been reported in more than 241different avian species (Kaleta and Baldauf 1988;Wobeser et al. 1993; Bailey et al. 1996; Alexander2000b).
The other eight avian paramyxoviruses are notnearly as well studied as is ND virus (APMV-1), andthe full range of hosts that each infects remains to bedetermined (Table 1.1).
ETIOLOGYThe causative agent of Newcastle Disease (ND) isavian paramyxovirus serotype 1 (APMV-1), classi edas belonging to the Order Mononegavirales, FamilyParamyxoviridae, Subfamily Paramyxovirinae, GenusRubulavirus (Rima et al. 1995). The virus contains alinear, noninfectious, negative sense, ssRNA genome of15—16 kb in size with a Mr of 3.5—5 × 106 thatcodes for six proteins, including an RNA directed RNApolymerase (L), hemagglutinin-neuraminidase (HN)protein, fusion (F) protein, matrix (M) protein, phos-phoprotein (P), and nucleoprotein (N) (de Leeuwand Peeters 1999). The virions are approximately150 nm or more in diameter, pleomorphic, with a lipidenvelope surrounding a helical nucleocapsid. Embeddedin the lipid envelope are the HN and F proteins, whichform the surface spikes. The virus is sensitive to lipidsolvents, unstable at very high or low pH, and showsheat lability, especially above 40¡C (Beard et al. 1984).
EPIZOOTIOLOGYND virus is readily transmitted among susceptiblebirds. Virus can be shed by infected birds in feces,body uids,and eggs, and potentially is present in alltissues, including meat and viscera. The virus cansurvive for long periods of time outside living hosts,and transmission via contamination of inanimateobjects is likely.
Most infections in wild birds appear to cause littleor no disease but result both in a detectable immuneresponse and a period of virus replication and shed-ding by the infected bird. Cormorants shed virus for21—32 days after e xperimental infection (MacPherson1956; Kuiken et al. 1998a). Such experimental results,and the many isolations of ND viruses from appar-ently healthy birds, indicate that long periods of virusshedding, and thus of potential transmission of thevirus to other birds, is usual in infections with NDvirus. As noted in the historical account (above),among wild bird populations, Double-crested Cor-morants and Rock Pigeons appear to maintain virusstrains pathogenic to themselves and to other species,while wild waterfowl (ducks and geese) appear tomaintain virus strains generally of low virulence tothemselves and to other species.
The virus is relatively stable in nature, remaininginfective for weeks at low temperatures and survivingfor several hours over a wide pH range. Protected byassociated organic matter, it can survive for days inlitter, water, soil, carcasses, eggs, and feathers. Virusremained infectious on feather down for 123 days at atemperature of 20¡C to 30¡C, 255 days when tempera-tures varied from 11¡C to 36¡C, and for 538 days at3¡C to 6¡C. The virus survived pH extremes of pH 3and pH 11 for up to one week. It remained infectiousin meat and bone for six months at 1¡C (Olesiuk 1951;Moses et al. 1947).
Infected Rock Pigeons are reported to shed virusstarting two days after infection and to continue shed-ding for about two weeks. Virus persisted in the intes-tine for up to three weeks after infection and in thebrain for up to ve weeks after infection. RockPigeons were considered no longer potential sourcesof infection for other birds six weeks after theybecame ill with ND. However, infectious virus maypersist in pigeon feces for more than six months undernatural conditions. In domestic ocks of Roc k Pigeons,new cases of clinical ND ceased to appear about veweeks after infection rst reached the ock (Kaletaand Baldauf 1988).
Epizootic ND in wild birds has been documented inonly two settings: Double-crested Cormorants inNorth America and Rock Pigeons, initially in the Mid-dle East, Africa, and Europe, and then North America,Japan, and worldwide. Epizootic ND occurred inRock Pigeons in southern Europe in the early 1980s,preceded by occurrences in the Middle East andAfrica, and spread north and west across Europe. Thevirus was translocated to North America and to Asiaby unknown means in the 1980s (Vindevogel andDuchatel 1988; Johnston and Key 1992; Barton et al.1992). Although documented primarily as a disease ofdomestic Rock Pigeons (racing pigeons), wild andferal birds were affected as well. Among naive domes-tic Rock Pigeons, morbidity rates have been reportedto range from 30% to 70%, with many affected birdsrecovering from the disease and mortality seldomexceeding 10%. Morbidity and mortality rates havenot been estimated in wild Rock Pigeons, but it islikely that morbidity rates equivalent to those reportedin domestic birds would result in much higher mortal-ity rates in wild Rock Pigeons, which must feed them-selves, avoid predators, and maintain their balance onroosts. Newcastle Disease rst affected free-livingurban Rock Pigeons in Saskatoon, Saskatchewan,Canada in the summer of 1990, and this epizootic maybe typical of epizootics elsewhere. Initially, ND pro-duced highly visible mortality. There was a slow butsteady rain of affected birds from their roosting areasunder the citys bridges into the South Saskatchewan
Infectious Diseases of Wild Birds6

River below that could be witnessed any day duringthe summer and early fall of 1990. Dead birds accu-mulated in such quantities around other urban rooststhat special collection and disposal were undertaken.Many birds were submitted to the local veterinary col-lege for diagnosis. Within two years, however, suchhighly visible mortality had ended, although the viruspersisted in the pigeon population. A single strain ofND virus was responsible for this world-wide epizooticamong Rock Pigeons (Alexander et al. 1985; Pearsonet al. 1987). Thus, there appears to be a stable relation-ship between this strain of ND virus and Rock Pigeonpopulations in many parts of the world.
Mortality of young-of-the-year (YOY) cormorantscaused by ND was rst recognized in North Americain Saskatchewan in 1990 and subsequently was recog-nized at various locations in 1992, 1995, 1996, 1997,1999, 2001, and 2003 (F.A. Leighton, unpublisheddata; Wobeser et al. 1993; Kuiken 1999; Meteyer et al.1997). At one breeding colony in Saskatchewan, mon-itored regularly for epidemic disease, ND occurredat two-year intervals in 1995, 1997, 1999, 2001, and2003. A single strain of virus was the cause of all ofthese occurrences of ND. Thus, there may be a stablerelationship between this ND virus strain and Double-crested Cormorants. Antibodies to APMV-1 (virusstrain not determined) were found in 26% to 56% ofmigratory Double-crested Cormorants sampled in win-ter in the southern United States from 1997 to 1999,and in 78% of eggs laid by the nonmigratory Floridasubspecies in 1998—1999 (F arley et al. 2001). It ismost likely that ND is enzootic in Double-crestedCormorant populations, with periodic epizooticoccurrence among YOY. Morbidity rates among cor-morants are not known. Rough estimates of mortalityrates among YOY have ranged from less than 1% to92%. A mortality rate within the range of 32% to 64%was estimated in one intensively studied outbreak(Kuiken 1999). ND virus in cormorants is more patho-genic to young birds than to older birds. This also isthe case in poultry (Kuiken et al. 1998a; Alexander1997). An ND virus isolated from an epizootic thatkilled at least 32% of YOY cormorants on their breed-ing colony produced minimal disease or none at allwhen previously unexposed, hand-reared birds fromthe same colony in the same year were infected at 16weeks of age. The birds on the colony had beenexposed to natural infection at about six weeks of age.The many outbreaks in Double-crested Cormorantshave caused mortality and clinical disease exclusivelyin YOY birds (Meteyer et al. 1997; Kuiken 1999).
Major outbreaks of ND in poultry in the early 1970swere traced to imported infected psittacine birds, andND virus strains highly pathogenic for chickens havefrequently been isolated from dead, sick, or asympto-
matic psittacines and other species imported intovarious countries in the pet bird trade (Clavijo et al.2000; Ashton 1984; Walker et al. 1973). Because of this,it has been assumed that wild populations of thesespecies, particularly psittacines, are reservoirs for theseND virus strains so highly pathogenic to chickens. Allevidence to date indicates that this is a false assump-tion. Pathogenic ND viruses have not been found inwild populations of these species, whereas highlypathogenic ND viruses continue to be found in ruralpoultry ocks in many tropical areas. It is probablethat tropical wild birds captured for the pet bird tradeare placed in contact with small rural ocks of domes-tic chickens during transportation, marketing, and inholding facilities, and that the captured birds becomeinfected with ND viruses only after capture (Johnsonet al. 1986; Goodman and Hanson 1988; Kaleta andBaldauf 1988).
CLINICAL SIGNSIn both Rock Pigeons and Double-crested Cor-morants, the signs most characteristic of ND are man-ifestations of central nervous system dysfunctionassociated with infection and in ammation of thebrain and spinal cord (Kaleta and Baldauf 1988;Barton et al. 1992; Kuiken et al. 1998b). Affectedbirds may have uncoordinated gait and movements,abnormal positioning of the head and neck, poor bal-ance, and unilateral or bilateral partial or completeparalysis of legs and wings (Figure 1.1). Similar clini-cal signs in Double-crested Cormorants are describedin detail by Kuiken et al. 1998b. Among cormorants,paralysis of legs and wings appears to persist as a per-manent debility in many birds that survive acute ND(Figure 1.1). Birds with unilateral wing paralysisthrash across the water with their one functional wingacting like a paddle wheel. They are able to dive butnot to y. Although unilateral wing dysfunction canhave many causes, observation of numbers of birdsexhibiting such single-wing attempts to y can betaken as good evidence of a current or recent epizooticof ND in cormorants. In epizootic years, cormorantswith such paralysed wings remain on colony lakesafter the rest of the birds have departed on southwardmigration, and can be seen on these lakes until freeze-up in late fall. Live birds falling from their roosts isone manifestation of clinical ND in Rock Pigeons.
Clinical signs associated with dysfunction of thecentral nervous system are not always evident in cor-morants and pigeons with ND. Many affected birdsare systemically ill and show only general weaknessand prostration. Diarrhea, sometimes with hemorrhage,is a usual feature of ND in Rock Pigeons and is theclassical clinical sign of highly pathogenic (visceral,velogenic) ND in domestic chickens (Vindevogel and
Newcastle Disease 7

Infectious Diseases of Wild Birds8
Figure 1.1. Clinical signs ofNewcastle Disease in Double-crested Cormorants. A. Bilateral leg paralysis. Thecormorant is trying to moveforward by use of its wingspivoted against the ground.B. Unilateral wing paralysis. Thenormal wing is spread and theaffected wing is held close tothe body. C. Loss of balance.The cormorant has fallen on itsback and has difficulty inrighting itself (From Kuiken1999, used with permission).
A
B
C

Duchatel 1988; Barton et al. 1992). For all species ofbirds, ND must be considered as one potential cause ofany clinical disease that includes bloody diarrhea,signs of central nervous system dysfunction, or generalprostration.
PATHOGENESISThe pathogenicity of ND virus varies greatly with thestrain of virus and the species of host bird. Nearly allresearch on the pathogenesis of ND virus infectionhas been done with domestic chickens, thus most ofthe information presented here is derived from experi-mental infections in chickens.
The period between infection and the appearance ofclinical disease usually is two to six days, but can beup to 15 to 21 days. An outbreak in chickens may beso severe that almost all of an affected ock dieswithin 72 hours without prior noticeable signs, oftencausing a suspicion of poisoning.
The primary determinant of virulence of ND virusesin chickens is the amino acid composition of the fusion(F) protein on the virus surface and its effect on theability of host cell enzymes to cleave this protein. Dur-ing infection with ND virus, it is necessary for the pre-cursor protein, F0, to be cleaved to proteins F1 and F2in order for further steps in the process of virus infec-tion to occur. This cleavage of F0 is mediated by hostcell proteases, which recognize a speci c motif ofamino acids at the F protein cleavage site. For exam-ple, in chickens it was found that if the cleavage motifcontained several basic amino acids, the cleavage andsubsequent virus replication could occur in most cellsthroughout the body, whereas if the cleavage motifcontained few or no basic amino acids, cleavage couldbe mediated only by enzymes found in the respiratoryor intestinal tracts, thus con ning virus infection tothese sites (Nagai et al. 1976; Alexander 2001; Aldouset al. 2001). Mutational changes in this cleavage site,resulting in the addition of basic amino acids, has ledto changes in viruses from low virulence to high, asseen in ND outbreaks in Ireland in 1990 and inAustralia in 1998—2000 (Collins et al. 1998; Westbury2001). The marked variation in pathogenicity of NDvirus strains in different species and ages of birds mayhave a similar basis in the interaction between theamino acid structure of virus surface proteins and hostenzyme locations and con gurations.
PATHOLOGYNewcastle Disease viruses typically cause lesions in oneor more of four organs or body systems: central nervoussystem, kidney, alimentary tract, and respiratory system.
ND viruses pathogenic to wild bird species appearmost often to affect the central nervous system andkidney, or to cause generalized, rapidly fatal diseaseaccompanied by few recognizable gross or histo-logical lesions. In both Double-crested Cormorantsand Rock Pigeons, pathological changes often arerestricted to the central nervous system and kidneyand are evident only microscopically (Kuiken et al.1999; Meteyer et al. 1997; Wobeser et al. 1993; Kaletaand Baldauf 1988; Barton et al. 1992). Histologicallesions in the central nervous system have consisted ofnonsuppurative in ammation in the brain and cord,sometimes including the meninges, with cuffs of lym-phocytes around blood vessels and with associatedgliosis, necrosis of neurons, and swelling of endothe-lial cells. These lesions occurred most regularly in thebrain stem and cerebellum in cormorants but alsowere evident elsewhere. Nonsuppurative nephritis,consisting of multiple small areas of in ltration ofthe renal parenchyma with lymphocytes and plasmacells, also has been a regular feature of infection withND virus in cormorants and pigeons. Small foci ofnecrosis of renal tubule cells have been observed inassociation with the in ammatory cells. Focal non-suppurative pancreatitis has been noted in infectedRock Pigeons. These lesions are particularly wellillustrated in Barton et al.(1992) and Kuiken et al.(1999).
Kaleta and Baldauf (1988) tabulated reported clin-ical signs and lesions from published accounts ofND virus infection in 222 different species of birdsinfected either naturally or experimentally with NDviruses. The birds in their survey either had no lesionsat all or had combinations of lesions in the alimentary,central nervous, and respiratory systems. Becausemany of the reports did not include microscopic evalu-ation of tissues, many of the birds said to have clinicalsigns of central nervous dysfunction but no lesionsprobably also had encephalomyelitis. This tabulationand the abundant literature about ND in domesticpoultry demonstrate that ND viruses can cause a widerange of lesions in any particular species of bird, andfurther, that none of these lesions is uniquely attributa-ble to ND.
The pathology of ND is well known and welldescribed in domestic chickens and turkeys (Alexander1997). Highly pathogenic strains of NDV usually causesevere hemorrhage and necrosis of the alimentary tract.Strains of lesser virulence for chickens often affect boththe respiratory and central nervous systems. In the respi-ratory system, lesions typically include hemorrhage andnecrosis of the mucosa of trachea and bronchi, some-times accompanied by pneumonia due to secondary
Newcastle Disease 9

bacterial infection of the lung. Conjunctivitis and rhini-tis also may result from these infections. Lesions in thecentral nervous system are evident only microscopicallyand consist of a nonsuppurative encephalomyelitis.
DIAGNOSISNeither clinical signs nor post mortem lesions aloneare a reliable basis for diagnosis of ND. Con rmationrequires identi cation and pathotyping of the virus,demonstration of viral genetic material within lesions,or a signi cant rise in antibody titre between acute andconvalescent sera coinciding with a disease outbreak.
When there is suspicion of ND, it is usual to attemptvirus isolation from recently dead or moribund birds.Samples from dead birds should include trachea or tra-cheal swabs and samples of lung, kidney, intestine(including contents), spleen, brain, liver, and heart. Tis-sue samples may be collected separately or as a pool,but the intestinal samples should be packaged sepa-rately. Where possible, a separate set of tissues alsoshould be collected in 10% neutral-buffered formalinfor histopathology. Samples from live birds shouldinclude tracheal swabs and cloacal swabs visibly coatedwith fecal material. Small birds may be harmed byswabbing; therefore the collection of freshly voidedfeces can serve as an adequate alternative. For trans-portation of samples, it is recommended that appropri-ate virus transport media be used (Alexander 2004),and if there is a delay longer than 72 hr in getting sam-ples to a diagnostic laboratory, they should be frozen.
Although routine virus isolation procedures inchicken embryos for NDV are generally adequate(Alexander 2004), some highly pathogenic strains ofND virus have failed to cause hemagglutination afterisolation in embryonated eggs and were detected onlyby use of an indirect immunoperoxidase assay. Forexample, 19 of 21 viruses isolated from epizootic NDin Double-crested Cormorants rapidly killed thechicken embryos used for primary isolation but failedto show hemagglutination with the standard screeningtest (Kuiken et al. 1999). Such failure of hemaggluti-nation may have been due to the highly pathogenicvirus killing the chicken embryos before sufficientvirus replication had occurred to produce a high con-centration of virus in the allantoic uid tested byhemagglutination. Alternatively, this virus strain maylack hemagglutinating properties.
After an ND virus has been isolated, its virulence indomestic chickens is assessed by an established pro-cedure of experimental infections, referred to as path-ogenicity testing. In many countries, it is a legalrequirement that identi cation of an ND virus be com-municated to national veterinary authorities and testedby those authorities for pathogenicity in chickens.Although several potential in vitro tests for establish-
ing virulence are being investigated by various groupsaround the world, at present the assessment of patho-genicity is based on one or more of the following invivo tests (Alexander 2004):
a. Mean death time in eggs: The mean death time(MDT) is the mean time in hours for the minimumlethal dose to kill chicken embryos. The MDT hasbeen used to classify ND virus strains as highlypathogenic (taking less than 60 hours to kill); mod-erately pathogenic (taking between 60 and 90 hoursto kill); and weakly pathogenic (taking more than90 hours to kill).
b. Intracerebral pathogenicity index: The intracerebralpathogenicity index (ICPI) is a weighted score ofclinical signs after intracerebral injection of thevirus into each of 10 ND-free day-old chickens.The most virulent viruses will give indices thatapproach the maximum score of 2.0, whereasstrains with low pathogenicity will give valuesclose to 0.0.
c. Intravenous pathogenicity index: The intravenouspathogenicity index (IVPI) is a weighted score ofclinical signs after intravenous injection of the virusinto 10 six-week-old ND-free chickens. Low patho-genic strains and some moderately pathogenicstrains will have IVPI values of 0.0, whereas theindices for virulent strains will approach 3.0.
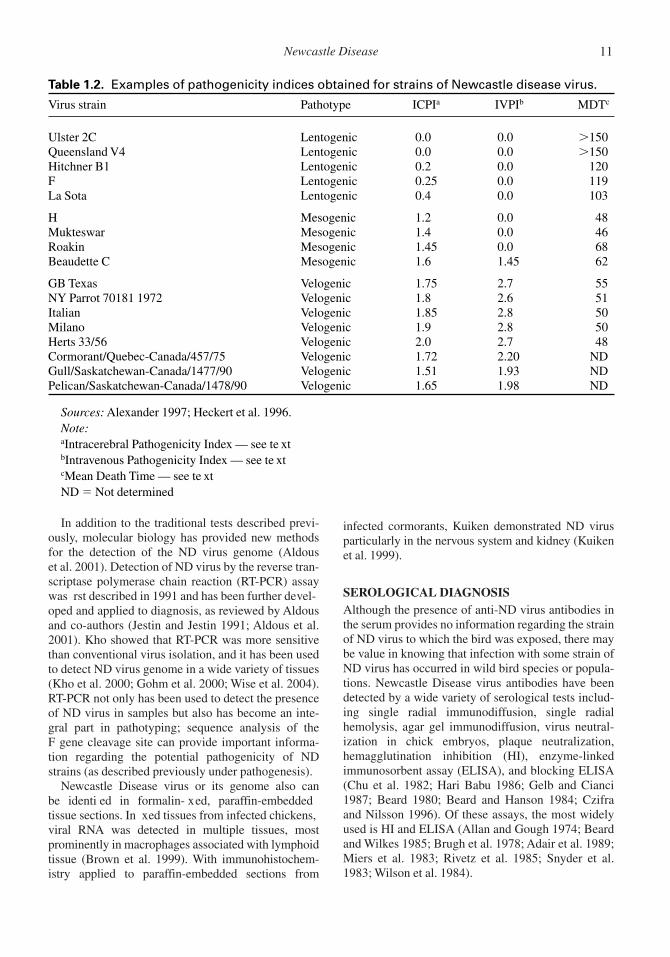
Based upon these tests, strains of ND virus wereclassically grouped into three pathotypes: velogenic(highly virulent), mesogenic (moderately virulent),and lentogenic (low virulence) (Table 1.2). In addi-tion, the velogenic viruses have also been further sub-classi ed based on the organ most severely affectedin the experimentally-infected chickens: viscerotropicfor the intestinal tract, neurotropic for the centralnervous system. In the regulation of internationaltrade in domestic poultry and poultry products, thename Ne wcastle Disease virus now is appliedexclusively to ND strains that are highly pathogenic(velogenic) for chickens.
Mouse monoclonal antibodies (MAbs) directedagainst strains of ND virus have been used in HI teststo allow rapid identi cation of ND viruses without thepossible cross-reactions with other APMV serotypesthat may occur with polyclonal sera. MAbs have beenproduced that give reactions in HI tests that are spe-ci c for particular strains or variant ND virus isolates.Panels of MAbs have been used to establish antigenicpro les of ND virus isolates. This has proven to be avaluable method for grouping and differentiating iso-lates of ND virus and has been particularly valuablein understanding the epizootiology of outbreaks(Alexander et al. 1997).
Infectious Diseases of Wild Birds10
In addition to the traditional tests described previ-ously, molecular biology has provided new methodsfor the detection of the ND virus genome (Aldouset al. 2001). Detection of ND virus by the reverse tran-scriptase polymerase chain reaction (RT-PCR) assaywas rst described in 1991 and has been further devel-oped and applied to diagnosis, as reviewed by Aldousand co-authors (Jestin and Jestin 1991; Aldous et al.2001). Kho showed that RT-PCR was more sensitivethan conventional virus isolation, and it has been usedto detect ND virus genome in a wide variety of tissues(Kho et al. 2000; Gohm et al. 2000; Wise et al. 2004).RT-PCR not only has been used to detect the presenceof ND virus in samples but also has become an inte-gral part in pathotyping; sequence analysis of theF gene cleavage site can provide important informa-tion regarding the potential pathogenicity of NDstrains (as described previously under pathogenesis).
Newcastle Disease virus or its genome also canbe identi ed in formalin- xed, paraffin-embeddedtissue sections. In xed tissues from infected chickens,viral RNA was detected in multiple tissues, mostprominently in macrophages associated with lymphoidtissue (Brown et al. 1999). With immunohistochem-istry applied to paraffin-embedded sections from
infected cormorants, Kuiken demonstrated ND virusparticularly in the nervous system and kidney (Kuikenet al. 1999).
SEROLOGICAL DIAGNOSISAlthough the presence of anti-ND virus antibodies inthe serum provides no information regarding the strainof ND virus to which the bird was exposed, there maybe value in knowing that infection with some strain ofND virus has occurred in wild bird species or popula-tions. Newcastle Disease virus antibodies have beendetected by a wide variety of serological tests includ-ing single radial immunodiffusion, single radialhemolysis, agar gel immunodiffusion, virus neutral-ization in chick embryos, plaque neutralization,hemagglutination inhibition (HI), enzyme-linkedimmunosorbent assay (ELISA), and blocking ELISA(Chu et al. 1982; Hari Babu 1986; Gelb and Cianci1987; Beard 1980; Beard and Hanson 1984; Czifraand Nilsson 1996). Of these assays, the most widelyused is HI and ELISA (Allan and Gough 1974; Beardand Wilkes 1985; Brugh et al. 1978; Adair et al. 1989;Miers et al. 1983; Rivetz et al. 1985; Snyder et al.1983; Wilson et al. 1984).
Newcastle Disease 11
Table 1.2. Examples of pathogenicity indices obtained for strains of Newcastle disease virus.
Virus strain Pathotype ICPIa IVPIb MDTc
Ulster 2C Lentogenic 0.0 0.0 �150Queensland V4 Lentogenic 0.0 0.0 �150Hitchner B1 Lentogenic 0.2 0.0 120F Lentogenic 0.25 0.0 119La Sota Lentogenic 0.4 0.0 103
H Mesogenic 1.2 0.0 48Mukteswar Mesogenic 1.4 0.0 46Roakin Mesogenic 1.45 0.0 68Beaudette C Mesogenic 1.6 1.45 62
GB Texas Velogenic 1.75 2.7 55NY Parrot 70181 1972 Velogenic 1.8 2.6 51Italian Velogenic 1.85 2.8 50Milano Velogenic 1.9 2.8 50Herts 33/56 Velogenic 2.0 2.7 48Cormorant/Quebec-Canada/457/75 Velogenic 1.72 2.20 NDGull/Saskatchewan-Canada/1477/90 Velogenic 1.51 1.93 NDPelican/Saskatchewan-Canada/1478/90 Velogenic 1.65 1.98 ND
Sources: Alexander 1997; Heckert et al. 1996.Note:aIntracerebral Pathogenicity Index — see te xtbIntravenous Pathogenicity Index — see te xtcMean Death Time — see te xtND � Not determined

A variety of commercial ELISA kits are availableand are based on several different strategies for thedetection of ND virus antibodies, including indirect,sandwich, and blocking or competitive tests usingmonoclonal antibodies. Usually such tests have beenevaluated and validated by the manufacturer, and it istherefore important that the instructions speci ed fortheir use be followed carefully. The ELISA lends itselfwell to screening large numbers of sera, and theresults correlate well with those of HI (Adair et al.1989; Brown et al. 1990; Cvelic-Cabrilo et al. 1992).The blocking ELISA may be most useful when testingsera from a wide variety of bird species. Althoughmore sera are probably tested by ELISA than by HI,the HI test is still the most widely used test interna-tionally due to its simplicity and ease in interpretation.Chicken sera rarely give nonspeci c positive reactionsin this test, and pretreatment of the sera is unneces-sary. Sera from species other than chickens maysometimes cause agglutination of chicken red bloodcells (CRBC), so this property should rst be deter-mined and then removed by absorption of the serumwith CRBC.
HI titers may be regarded as being positive if thereis inhibition of hemagglutination at an initial serumdilution of 1:16 or more against 4 HA units of antigen.Some laboratories prefer to use 8 HA units in HI tests.This is permissible, but it affects the interpretation ofresults such that a positive titer becomes 1:8 or more.Hemagglutination inhibition also has been used todetect antibodies to ND virus in egg yolk in epidemio-logical monitoring of exposure to the virus (Kuikenet al. 1998b; Farley et al. 2001).
IMMUNITYMost if not all avian species will produce an immuneresponse upon exposure to ND virus (Kaleta andBaldauf 1988; Alexander 2004; Sousa et al. 1999;Kuiken et al. 1998a). In commercial poultry, thisresponse has been shown to be both humoral (anti-body) and cellular (cell mediated). The initial immuneresponse to infection with ND virus is cell mediatedand may be detectable as early as two to three daysafter infection with live vaccine strains (Ghummanand Bankowski 1976; Timms and Alexander 1977).The importance of cell-mediated immunity is stillunclear. One study showed that it contributed to pro-tection but in itself was insufficient to provide com-plete protection (Reynolds and Maraqa 2000a).
When chickens and some other species have beenexposed to ND virus, antibodies generally weredetectable in the serum within 6 to 10 days. Antibodiesagainst the HN and F proteins are neutralizing antibod-ies (Russell 1988; Reynolds and Maraqa 2000b). Theamount of antibody produced is dependent upon the
infecting strain and generally peaks at approximatelythree to four weeks post infection. As determined by HI,antibodies can persist for as long as one year after infec-tion (Allan and Gough 1974). Secreted antibodies, inparticular from the Harderian gland near the eye, areimportant in providing upper respiratory tract protectionin chickens (Holmes 1979a; Holmes 1979b; Parry andAitken 1977). These antibodies have been shown to beprimarily of the IgM and IgA class (Russell 1993; Rus-sell and Ezeifeka 1995).
Antibody titers in experimentally infected cor-morants reached a maximum 21 days after infectionand were still detectable 70 days after infection whenthe experiment ended. From the steady rate of declinein titer following the peak, it was predicted that titerswould have become undetectable about 126 days afterinfection (Kuiken et al. 1998a). It is not known whensuch birds might become susceptible to re-infection orhow long virus would be shed from re-infected birds.
PUBLIC HEALTH CONCERNSNewcastle Disease virus is a human pathogen of minorimportance. The Advisory Committee on DangerousPathogens of the United Kingdom has assigned NDvirus to Hazard Group 2, each member of which isde ned as a biological agent that can cause humandisease and may be a hazard to employees; it isunlikely to spread to the community (Alexander2000a). Persons most likely to become infected arethose who handle infected birds, such as farmers andpigeon fanciers, veterinary health care workers,including those who vaccinate birds on poultry farms,abattoir workers, and personnel of diagnostic laborato-ries. There are no records of human infections acquiredfrom consumption of infected eggs or meat. Avirulentvirus strains used in live vaccines and eld strainspathogenic for birds appear to be equally pathogenicfor people. Human disease caused by ND virus hasbeen reviewed by M.I. Khan (1994). The commonresult of ND virus infection in humans is conjunctivi-tis, which may be severe but is of only a few daysduration and without residual effect once resolved,unless complicated by secondary pathogens. Nothingis known of the potential of other avian paramyx-oviruses to cause disease in people.
DOMESTIC AND CAPTIVE ANIMAL HEALTH CONCERNSNewcastle Disease is one of the most important dis-eases of poultry around the world. Historically, epi-zootic ND has caused high mortality and massiveexpenditures on eradication in developed countries(Walker et al. 1973; Alexander 1988a). Vaccinationand biosecurity measures required because of ND
Infectious Diseases of Wild Birds12

virus are a constant cost to industrial poultry production.However, the most important economic impact of NDmay be on small poultry ocks in Asia, Africa, CentralAmerica, and parts of South America because of themajor signi cance of these ocks to local economiesand nutrition, and because of the regular high mortal-ity caused by ND in these settings (Alexander 2000a).For example, in Nepal it has been estimated that asmany as 90% of chickens in small village ocks dieeach year from ND. Small rural ocks are the mostlikely source for infections in birds that enter the petbird trade.
In addition to effects on traditional domestic species,Newcastle Disease has been a cause of economic loss incaptive ratites and domestic pigeons and is a constanthazard to zoos and wild animal rehabilitation facilities,which can suffer losses of birds to disease and, poten-tially, depopulation orders imposed on healthy speci-mens that were exposed to ND virus by infected birdsbrought into these facilities for medical care. Captivebreeding programs for endangered bird species experi-ence similar risks (Kaleta and Baldauf 1988; Vindevogeland Duchatel 1988; Bailey et al. 1996).
Strains of ND virus highly pathogenic to domesticpoultry are enzootic in free-living populations of Dou-ble-crested Cormorants and Rock Pigeons, and bothspecies have been sources of infection for commercialpoultry ocks (Alexander et al. 1984; Heckert et al.1996). Alexander and others (1998) considered itlikely that wild ducks, geese, and swans carried strainsof ND virus pathogenic to poultry among sites of out-breaks in commercial poultry in Europe in 1997.
WILDLIFE POPULATION IMPACTS ANDMANAGEMENT IMPLICATIONSFew data are available to evaluate the potential effect ofND on wild bird populations. In Double-crested Cor-morants, ND appears to cause high mortality, but onlyamong young-of-the-year. Such mortality in the pre-edging period,even if it occurs frequently and recur-rently, may have little or no impact on overall populationsize or structure (Kuiken 1999). Population effects ofND in wild Rock Pigeons have not been studied. Theremay be an indirect negative impact of ND on popula-tions of tropical birds that are popular in the pet birdtrade. Stringent import regulations for such birds,imposed to prevent importation of ND viruses, probablyare one stimulus for illegal trading practices, whichoften result in high mortality rates from diseases andhandling. This may result in additional captures of wildbirds and further reductions in wild populations.
Wildlife managers should be aware that strains ofND highly pathogenic to domestic poultry are enzooticin Double-crested Cormorants and Rock Pigeons but,
as far as is currently known, only in these species.Other species of wild birds also are capable of carryingsuch pathogenic strains of ND for short periods oftime. Where vaccination of poultry against ND is notroutinely carried out, prevention of ND in poultryrequires nonporous physical barriers between domes-tic and wild birds to ensure that transmission of NDviruses (and several other important pathogens) doesnot occur. Wherever possible, wildlife managersshould work with local poultry enterprises to reducethe risk of having ND enter these premises from wildbird sources and the risk of wild birds becomingexposed to ND virus strains present in poultry.
TREATMENT AND CONTROLThere is no treatment for ND. Vaccination with bothlive and killed vaccines is used in domestic birds as anadjunct to biosecurity procedures, which are the pri-mary means of prevention of ND in the commercialpoultry industry (Alexander 2000a). Vaccination maybe an effective method of ND prevention in gallina-ceous species in rare or endangered species recoveryprograms, but is not a practical solution for wild birdpopulations. ND virus may be eliminated by pasteur-ization of table eggs and egg products for 4.5 minutesat 64¡C; rendering for several minutes at 100¡C; pro-cessing of meat for 30 minutes at 30¡C or one minuteat 80¡C. The rate at which the virus is destroyeddepends on the strain of virus, the quantity of virus,the time of treatment and the media in which the treat-ment occurs (Beard and Hanson 1984). In the com-mercial poultry industry of most developed countries,outbreaks of ND trigger stamping-out, or eradica-tion, responses whereby all infected and potentialcontact birds are killed, there is sanitary disposal of allcarcasses and bird products, and affected premises aredisinfected and left without birds for a period of time.
ACKNOWLEDGEMENTSThijs Kuiken and Gary Wobeser contributed substan-tially to the authorsunderstanding of ND through dis-cussion, collaborative research and review.
UNPUBLISHED DATAF. A. Leighton, Canadian Cooperative Wildlife HealthCentre, Saskatoon, Saskatchewan, Canada.
LITERATURE CITEDAdair, B.M., M.S. McNulty, D. Todd, T.J. Connor, and
K. Burns. 1989. Quantitative estimation of Newcastledisease virus antibody levels in chickens and turkeysby ELISA. Avian Pathology 18:175—192.
Aldous, E.W., M.S. Collins, A. McGoldrick, andD.J. Alexander. 2001. Rapid pathotyping of Newcastle
Newcastle Disease 13

disease virus (NDV) using uorogenic probes in aPCR assay. Veterinary Microbiology 80:201—212.
Alexander, D.J. 1988a. Historical aspects. In NewcastleDisease, 1st Ed., D.J. Alexander (ed.). Kluwer Acade-mic Publishers, Boston, MA, U.S.A., pp. .
Alexander, D.J. 1988b. Newcastle disease Virus AnAvian Paramyxovirus. In Newcastle Disease, 1st Ed.D.J. Alexander (ed.). Kluwer Academic Publishers,Boston, MA, U.S.A., pp. 11—22.
Alexander, D.J. 1997. Newcastle disease and other avianParamyxoviridae infections. In Diseases of Poultry,10th Ed., B. W. Calnek (ed.). Iowa State UniversityPress, Ames, IA, U.S.A., pp. 541—569.
Alexander, D.J. 2000a. Newcastle disease and other avianparamyxoviruses. Revue Scienti que et Technique del Office International des Epizooties 19:443—462.
Alexander, D.J. 2000b. Newcastle disease in ostriches(Struthio camelus) a re view. Avian Pathology29:95—100.
Alexander, D.J. 2001. Gordon Memorial Lecture:Newcastle disease. British Poultry Science 42:5—22.
Alexander, D.J. 2004. Newcastle disease. In Manual ofStandards for Diagnostic Tests and Vaccines for Ter-restrial Animals (mammals, birds and bees), 5th Ed.World Organization for Animal Health (OIE), Paris,France, pp. 270—282.
Alexander, D.J., R.J. Manvell, J.P. Lowings, K.M. Frost,M.S. Collins, P.H. Russell, and J.E. Smith. 1997. Anti-genic diversity and similarities detected in avianparamyxovirus type 1 (Newcastle disease virus) iso-lates using monoclonal antibodies. Avian Pathology26:399—419.
Alexander, D.J., H.T. Morris, W.J. Pollitt, C.E. Sharpe,R.L. Eckford, R.M.Q. Sainsbury, L.M. Mansley,R.E. Gough, and G. Parsons. 1998. Newcastle diseaseoutbreaks in domestic fowl and turkeys in GreatBritain during 1997. Veterinary Record 143:209—212.
Alexander, D.J., G. Parsons, and R. Marshall. 1984.Infection of fowls with Newcastle disease virus byfood contaminated with pigeon faeces. VeterinaryRecord 115:601—602.
Alexander, D.J., G.W.C. Wilson, P.H. Russell, S.A. Lister,and G. Parsons. 1985. Newcastle disease outbreaks infowl in Great Britain during 1984. Veterinary Record117:429—434.
Allan, W.H., and R.E. Gough. 1974. A standard hemag-glutination inhibition test for Newcastle disease (1).A comparison of macro and micro methods. Veteri-nary Record 95:120—123.
Artois, M., R. Manvell, E. Fromont, and J-B. Schweyer.2002. Serosurvey for Newcastle disease and avianin uenza A virus antibodies in great cormorants fromFrance. Journal of Wildlife Diseases 38:169—171.
Ashton, W.L. G. 1984. The risks and problems connectedwith the import and export of captive birds. BritishVeterinary Journal 140:317—327.
Bailey, T.A., P.K. Nicholls, J.H. Samour, J. Naldo,U. Wernery, and J. C. Howlett. 1996. Postmortem nd-ings in bustards in the United Arab Emirates. AvianDiseases 40:296—305.
Barton, J.T., A.A. Bickford, G.L. Cooper, B.R. Charlton,and C.J. Cardona. 1992. Avian paramyxovirus Type 1infections in racing pigeons in California. 1. ClinicalSigns, Pathology, and Serology. Avian Diseases 36:463—468.
Beard, C.W. 1980. Serologic procedures. In Isolation andIdenti cation of Avian Pathogens, S.B. Hitchner,C.H. Domermuth, H.G. Purchase, and J.E. Williams(eds.). American Association of Avian Pathologists,Kennet Square, PA, U.S.A., pp. 129—135.
Beard, C.W. and R.P. Hanson. 1984. Newcastle disease. InDiseases of Poultry, 8th Ed., M.S. Hofstad, H.J. Barnes,B.W. Calnek, W.M. Reid, and H.W. Yoder (eds.). IowaState University Press, Ames, IA, U.S.A., pp. 452—470.
Beard, C.W. and W.J. Wilkes. 1985. A comparison ofNewcastle disease hemagglutination-inhibition testresults from diagnostic laboratories in the south east-ern United States. Avian Diseases 29:1048—1056.
Blaxland, J.D. 1951. Newcastle disease in shags and cor-morants and its signi cance as a factor in the spread ofthis disease among domestic poultry. VeterinaryRecord 63:731—733.
Bradshaw, J.E. and D.O. Trainer. 1966. Some infectiousdiseases of waterfowl in the Mississippi. Journal ofWildlife Management 30:570—576.
Brown, C., D.J. King, and B.S. Seal. 1999. Pathogenesisof Newcastle disease in chickens experimentallyinfected with viruses of different virulence. VeterinaryPathology 36:125—132.
Brown, J., R.S. Resurreccion, and T.G. Dickson. 1990.The relationship between the hemagglutination-inhibition test and enzyme-linked immunosorbentassay for detection of antibody to Newcastle disease.Avian Diseases 34:585—587.
Brugh, M. Jr., C.W. Beard, and W.J. Wilkes. 1978. Thein uence of test conditions on Newcastle diseasehemagglutination-inhibition titres. Avian Diseases22:320—328.
Chu, H.P., G. Snell, D.J. Alexander, and G.C. Schild.1982. A single radial immunodiffusion test for anti-bodies to Newcastle disease virus. Avian Pathology11:227—234.
Clavijo, A., Y. Robinson, T. Booth, and F. Munroe. 2000.Velogenic Newcastle disease in imported caged birds.Canadian Veterinary Journal 41:404—408.
Collins, M.S., S. Franklin, I. Strong, G. Meulemans, andD.J. Alexander. 1998. Antigenic and phylogenetic stud-ies on a variant Newcastle disease virus using anti-fusionprotein monoclonal antibodies and partial sequencing ofthe fusion protein gene. Avian Pathology 27:90—97.
Cvelic-Cabrilo, V.,H. Mazija, Z. Bidin, and W.L. Ragland.1992. Correlation of hemagglutination-inhibition and
Infectious Diseases of Wild Birds14

enzyme-linked immunosorbent assays for antibodiesto Newcastle disease virus. Avian Pathology 21:509—512.
Czifra, G., and M. Nilsson. 1996. Detection of PMV-1speci c antibodies with a monoclonal antibody block-ing enzyme-linked immunosorbent assay. AvianPathology 25:691—703.
De Leeuw, O., and B. Peeters. 1999. Completenucleotide sequence of Newcastle disease virus: evi-dence for the existence of a new genus within the sub-family Paramyxovirinae. Journal of General Virology80:131—136.
Doyle, T.M. 1927. A hitherto unrecorded disease of fowlsdue to a ltre-passing virus. Journal of ComparativePathology and Therapeutics 40:144.
Farley, J.M., S.H. Romero, M.G. Spalding, M.L. Avery, andD.J. Forrester. 2001. Newcastle disease in double-crested cormorants in Alabama, Florida and Mississippi.Journal of Wildlife Diseases 37:808—812.
Gelb, J. Jr., and C.G. Cianci. 1987. Detergent-treatedNewcastle disease viruses as an agar gel precipitin testantigen. Poultry Science 66:845—853.
Ghumman, J.S., and R.A. Bankowski. 1976. In vitroDNA synthesis in lymphocytes from turkeys vacci-nated with LaSota, TC, and inactivated Newcastle dis-ease vaccines. Avian Diseases 20:18—31.
Gohm, D.S., B. Thus, and M.A. Hofmann. 2000. Detec-tion of Newcastle disease virus in organs and faeces ofexperimentally infected chickens using RT-PCR.Avian Pathology 29:143—152.
Goodman, B.B. and R.P. Hanson. 1988. Isolation ofavian paramyxovirus-2 from domestic and wild birdsin Costa Rica. Avian Diseases 32:713—717.
Hari Babu, Y. 1986. The use of a single radial haemolysistechnique for the measurement of antibodies to New-castle disease virus. Indian Veterinary Journal63:982—984.
Heckert, R.A., M.S. Collins, R.J. Manvell, I. Strong,J.E. Pearson, and D.J. Alexander. 1996. Comparisonof Newcastle Disease viruses isolated from cormorantsin Canada and the U.S.A. in 1975, 1990 and 1992.Canadian Journal of Veterinary Research 60:50—54.
Holmes, H.C. 1979a. Resistance of the respiratory tractof the chicken to Newcastle disease virus infection fol-lowing vaccination: the effect of passively acquiredantibody on its development. Journal of ComparativePathology 89:11—19.
Holmes, H.C. 1979b. Virus-neutralizing antibody in seraand secretions of the upper and lower respiratory tractof chickens inoculated with live and inactivated New-castle disease virus. Journal of Comparative Pathol-ogy 89:21—29.
Jestin, V., and A. Jestin. 1991. Detection of Newcastledisease virus RNA in infected allantoic uids by invitro enzymatic ampli cation (PCR). Archives ofVirology 118:151—161.
Johnson, D.C., C.E. Couvillion, and J.E. Pearson. 1986.Failure to demonstrate viscerotropic velogenic New-castle disease in psittacine birds in the Republic of thePhilippines. Avian Diseases 30:813—815.
Johnston, K.M. and D.W. Key. 1992. Paramyxovirus-1 inferal pigeons (Columba livia) in Ontario. CanadianVeterinary Journal 33:796—800.
Kaleta, E.F. and C. Baldauf. 1988. Newcastle disease inFree-Living and Pet Birds. In Newcastle Disease, 1stEd. D.J. Alexander (ed.). Kluwer Academic Publishers,Boston, MA, pp. 197—246.
Khan, M.I. 1994. Newcastle Disease. In Handbookof Zoonoses, Section B: Viral Diseases, 2nd. Ed.,G.W. Beran (ed.). CRC Press, United States,pp. 473—481.
Kho, C.L., M.L. Mohd-Azmi, S.S. Arshad, andK. Yusoff. 2000. Performance of an RT-nested PCRELISA for detection of Newcastle disease virus. Jour-nal of Virological Methods 86:71—83.
Kuiken, T. 1999. Review of Newcastle Disease inCormorants: Waterbirds 22:333—347.
Kuiken, T., R.A. Heckert, J. Riva, F.A. Leighton, andG. Wobeser. 1998a. Excretion of pathogenic Newcas-tle disease virus by double-crested cormorants (Pha-lacrocorax auritus) in absence of mortality or clinicalsigns of disease. Avian Pathology 27:541—546.
Kuiken, T., F.A. Leighton, G. Wobeser, K.L. Danesik,J. Riva, and R.A. Heckert. 1998b. An epidemic ofNewcastle disease in Double-Crested Cormorants fromSaskatchewan. Journal of Wildlife Diseases 34:457—471.
Kuiken, T., G. Wobeser, F.A. Leighton, D.M. Haines,B. Chelack, J.B. Bogdan, L. Hassard, R.A. Heckert,and J. Riva. 1999. Pathology of Newcastle Disease inDouble-Crested Cormorants from Saskatchewan, withcomparison of diagnostic methods. Journal of WildlifeDiseases 35:8—23.
MacPherson, L.W. 1956. Some observations on the epi-zootiology of Newcastle disease. Canadian Journal ofComparative Medicine 10:55—168.
Meteyer, C.U., D.E. Docherty, L.C. Glaser, J.C.Franson, D.A. Senne, and R. Duncan. 1997. Diagnos-tic ndings in the 1992 epornitic of neurotropicvelogenic Newcastle disease in Double-crested Cor-morants from the upper midwestern United States.Avian Diseases 41:171—180.
Miers, L.A., R.A. Bankowski, and Y.C. Zee. 1983. Opti-mizing the enzyme-linked immunosorbent assay forevaluating the immunity of chickens to Newcastle dis-ease. Avian Diseases 27:1112—1125.
Morgan, I.R., and H.A. Westbury. 1981. Virological stud-ies of Adelie penguins (Pygoscelis adeliae) in Antarc-tica. Avian Diseases 25:1019—1026.
Moses, H.E., C.A. Brandly, and E.E. Jones. 1947. ThepH stability of viruses of Newcastle disease and fowlplague. Science 105:477—479.
Newcastle Disease 15

Nagai, Y., H.D. Klenk, and R. Rott. 1976. Proteolyticcleavage of the viral glycoproteins and its signi cancefor the virulence of Newcastle disease virus. Virology72:494—508.
Olesiuk, O.M. 1951. In uence of environmental factorson viability of Newcastle disease virus. AmericanJournal of Veterinary Research 12:152—155.
Palmer, S.F., and D.O. Trainer. 1971. Newcastle Disease.In Infectious and Parasitic Diseases of Wild Birds, 1sted., J.W. Davis, R.C. Anderson, L. Karstad, andD.O. Trainer (eds.). The Iowa State University Press,Ames, IA, U.S.A., pp. 3—16.
Parry, S.H., and I.D. Aitken. 1977. Local immunity in therespiratory tract of the chicken. II. The secretoryimmune response to Newcastle disease virus and therole of IgA. Veterinary Microbiology 2:143—165.
Pearson, J.E., D.A. Senne, D.J. Alexander, W.D. Taylor,L.A. Peterson, and P.H. Russell. 1987. Characterizationof Newcastle disease virus (avian paramyxovirus-1) iso-lated from pigeons. Avian Diseases 31:105—111.
Reynolds, D.L., and A.D. Maraqa. 2000a. Protectiveimmunity against Newcastle disease: the role of cell-mediated immunity. Avian Diseases 44:145—154.
Reynolds, D.L., and A.D. Maraqa. 2000b. Protectiveimmunity against Newcastle disease: the role of anti-bodies speci c to Newcastle disease polypeptides.Avian Diseases 44:138—144.
Rima, B., D.J. Alexander, M.A. Billeter, P.L. Collins,D.W. Kingsbury, M.A. Lipkind, Y. Nagai, C. Orvell,C.R. Pringle, and V. ter Meulen. 1995. Paramyxoviri-dae. In Virus Taxonomy: Sixth Report of the Interna-tional Committee on Taxonomy of Viruses, F.A.Murphy, C.M. Fauquet, D.H.L. Bishop, S.A. Ghabrial,A.W. Jarvis, G.P. Martelli, M.A. Mayo, andM.D. Summer (eds.). Vienna: Springer-Verlag, Austria,pp. 268—274.
Rivetz, B., W. Weisman, M. Ritterband, F. Fish, andM. Herzberg. 1985. Evaluation of a novel rapid kit forthe visual detection of Newcastle disease antibodies.Avian Diseases 29:929—942.
Russell, P.H. 1988. Monoclonal antibodies in research,diagnosis and epizootiology of Newcastle disease. InNewcastle Disease, 1st Ed., D.J. Alexander (ed.).Kluwer Academic Publishers, Boston, MA, U.S.A.,pp. 131—146.
Russell, P.H. 1993. Newcastle disease virus: virus repli-cation in the harderian gland stimulates lacrimal IgA;the yolk sac provides early lacrimal IgG. VeterinaryImmunology and Immunopathology 37:151—163.
Russell, P.H., and G.O. Ezeifeka. 1995. The Hitchner B1strain of Newcastle disease virus induces high levelsof IgA, IgG and IgM in newly hatched chicks. Vaccine13:61—66.
Snyder, D.B., W.W. Marquart, E.T. Mallinson, andE. Russek. 1983. Rapid serological pro ling byenzyme-linked immunosorbent assay. I. Measurementof antibody activity titre against Newcastle disease virusin a single serum dilution. Avian Diseases 27:161—170.
Sousa, R.L., T.C. Cardoso, A.C. Paulillo, H.J. Montassier,and A. A. Pinto. 1999. Antibody response to Newcastledisease vaccination in a ock of young partridges(Rhynchotus rufescens). Journal of Zoo and WildlifeMedicine 30:459—461.
Stallknecht, D.E., D.A. Senne, P.J. Zwank, S.M. Shane,and M.T. Kearney. 1991. Avian paramyxoviruses frommigrating and resident ducks in coastal Louisiana.Journal of Wildlife Diseases 27:123—128.
Timms, L. and D.J. Alexander. 1977. Cell-mediatedimmune response of chickens to Newcastle diseasevaccines. Avian Pathology 6:51—59.
Vickers, M.L. and R.P. Hanson. 1980. Experimental infec-tion and serologic survey for selected paramyxovirusesin red-winged blackbirds (Agelaius phoeniceus). Jour-nal of Wildlife Diseases 16:125—130.
Vickers, M.L. and R.P. Hanson. 1982. Newcastle diseasevirus in waterfowl in Wisconsin. Journal of WildlifeDiseases 18:149—158.
Vindevogel, H. and J.P. Duchatel. 1988. Panzootic New-castle Disease Virus in Pigeons. In Newcastle Disease,1st Ed., D.J. Alexander (ed.). Kluwer Academic Pub-lishers, Boston, MA, U.S.A., pp. 184—196.
Walker, J.W., B.R. Heron, and M.A. Mixson. 1973.Exotic Newcastle disease eradication program in theUnited States. Avian Diseases 17:486—503.
Westbury, H. 2001. Newcastle disease virus: an evolvingpathogen. Avian Pathology 30:5—11.
Wilson, J.E. 1950. Newcastle disease in a Gannet (Sulabassana). Veterinary Record 62:33—34.
Wilson, R.A., C. Perotta Jr., and R.J. Eckroade. 1984. Anenzyme-linked immunosorbent assay that measuresprotective antibody levels to Newcastle disease virusin chickens. Avian Diseases 28:1079—1085.
Wise M.G., D.L. Suarez, B.S. Seal, J.C. Pedersen,D.A. Senne, D.J. King, D.R. Kapczynski, andE. Spackman. 2004. Development of a real-timereverse-transcription PCR for detection of Newcastledisease virus RNA in clinical samples. Journal ofClinical Microbiology 42:329—338.
Wobeser, G., F.A. Leighton, R. Norman, D.J. Myers,D. Onderka, M.J. Pybus, J.L. Neufeld, G.A. Fox, andD.J. Alexander. 1993. Newcastle disease in wild waterbirds in western Canada, 1990. Canadian VeterinaryJournal 34:353—359.
World Organization for Animal Health (OIE). 2004.International Animal Health Code: mammals, birdsand bees. Paris: World Organization for Animal Health.
Infectious Diseases of Wild Birds16