induction of inhibitory factor kbamrna in the central ... · induction of inhibitory factor kbamrna...
TRANSCRIPT

Proc. Natl. Acad. Sci. USAVol. 94, pp. 10985–10990, September 1997Neurobiology
Induction of inhibitory factor kBa mRNA in the central nervoussystem after peripheral lipopolysaccharide administration:An in situ hybridization histochemistry study in the rat
(endotoxinycytokineynuclear factor kByblood–brain barrieryglia)
NING QUAN*, MICHAEL WHITESIDE, LEOPOLD KIM, AND MILES HERKENHAM
Section on Functional Neuroanatomy, National Institute of Mental Health, Building 36, Room 2D15, Bethesda, MD 20892-4070
Edited by S. M. McCann, Pennington Biomedical Research Center, Baton Rouge, LA, and approved July 23, 1997 (received for reviewNovember 14, 1996)
ABSTRACT In this study we investigate the mRNA ex-pression of inhibitory factor kBa (IkBa) in cells of the ratbrain induced by an intraperitoneal (i.p.) injection of lipo-polysaccharide (LPS). IkB controls the activity of nuclearfactor kB, which regulates the transcription of many immunesignal molecules. The detection of IkB induction, therefore,would reveal the extent and the cellular location of brain-derived immune molecules in response to peripheral immunechallenges. Low levels of IkBa mRNA were found in the largeblood vessels and in circumventricular organs (CVOs) ofsaline-injected control animals. After an i.p. LPS injection(2.5 mgykg), dramatic induction of IkBa mRNA occurred infour spatio-temporal patterns. Induced signals were firstdetected at 0.5 hr in the lumen of large blood vessels and inblood vessels of the choroid plexus and CVOs. Second, at 1–2hr, labeling dramatically increased in the CVOs and choroidplexus and spread to small vascular and glial cells throughoutthe entire brain; these responses peaked at 2 hr and declinedthereafter. Third, cells of the meninges became activated at 2hr and persisted until 12 hr after the LPS injection. Finally,only at 12 hr, induced signals were present in ventricularependyma. Thus, IkBa mRNA is induced in brain afterperipheral LPS injection, beginning in cells lining the bloodside of the blood–brain barrier and progressing to cells insidebrain. The spatiotemporal patterns suggest that cells of theblood–brain barrier synthesize immune signal molecules toactivate cells inside the central nervous system in response toperipheral LPS. The cerebrospinal f luid appears to be aconduit for these signal molecules.
One way that the central nervous system (CNS) responds toperipheral immune challenges is by generating its own immunesignal molecules (1). The mechanisms by which this phenom-enon is induced have not been elucidated. A key regulatorycomponent of intracellular signal pathways in cells of theimmune system is the nuclear factor kB (NF-kB)yrel family oftranscription factors. These factors respond to numerous im-mune challenges by activating gene transcription for a widevariety of immune signal molecules (2). Tracing the activationof NF-kB in the CNS after peripheral immune challenges maytherefore reveal the extent and neuroanatomical location ofthe processes that are involved in responding to peripheralimmune signals and, in turn, generating centrally derivedimmune molecules.
The mechanisms by which NF-kB regulates gene expressionhave recently been reviewed (3–5). Briefly, NF-kB is normallypresent in the cytoplasm as a dormant complex with an
inhibitor, IkB. Upon extracellular stimulation, IkB is phos-phorylated and degraded. This leads to the release of freeNF-kB, which translocates to the nucleus and binds to specifickB sequences of the DNA to initiate transcription of relatedgenes including immunoreceptors, cytokines, and, interest-ingly, its own inhibitor, IkB.
Two unique features of the NF-kByIkB system result fromits feedback regulation. First, the activation of transcription byNF-kB also triggers the synthesis of IkB (4). Second, NF-kB-activated transcription is maintained by continuous degrada-tion of IkB (5), which is sustained by ongoing extracellularstimulation (6). Thus, the expression of IkB mRNA parallelsboth the NF-kB activity and the duration of the activatingextracellular stimulation. This temporal parallelism betweenIkB mRNA expression and the effective external stimulationis different from other transcription factors (e.g., c-Fos) thatare transiently expressed only at the onset of cellular stimu-lation (7). These features may be exploited to observe thelocalization of cells undergoing NF-kB-regulated gene tran-scription and infer the presence and duration of extracellularstimuli.
In this study, we measured IkBa mRNA expression by in situhybridization histochemistry in brain sections of rats after theywere injected intraperitoneally (i.p.) with lipopolysaccharide(LPS) or sterile saline to trace the activation of responsive cellsthat potentially synthesize immune signal molecules in theCNS. Because LPS is a large molecule that is generally thoughtnot to diffuse significantly across the intact blood–brainbarrier (BBB) (8), we specially examined the brain levelscontaining areas with a leaky BBB, such as the circumven-tricular organs (CVOs) (9). We selected IkBa as a marker ofNF-kB activation instead of the other members of the IkBfamily because it is ubiquitous and responds to most forms ofextracellular stimulation (10). Our results show profoundLPS-induced elevation of IkBa mRNA levels in selected cellsin the CNS. The apparent migration of the observed IkBamRNA labeling across the BBB into deep cells of the brainsuggests pathways by which peripheral LPS may activate thesynthesis of immune molecules in CNS.
MATERIALS AND METHODS
Animals and LPS Injection. Male Sprague–Dawley rats(175–200 gm; Taconic Farms) were group housed and handleddaily before experimentation. They were injected i.p. witheither LPS (2.5 mgykg) dissolved in 0.9% saline or 0.9% sterile
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked ‘‘advertisement’’ inaccordance with 18 U.S.C. §1734 solely to indicate this fact.
© 1997 by The National Academy of Sciences 0027-8424y97y9410985-6$2.00y0PNAS is available online at http:yywww.pnas.org.
This paper was submitted directly (Track II) to the Proceedings office.Abbreviations: AP, area postrema; BBB, blood–brain barrier; CSF,cerebrospinal f luid; CVO, circumventricular organ; IkBa, inhibitoryfactor kBa; LPS, lipopolysaccharide; NF-kB, nuclear factor kB; NTS,nucleus of the solitary tract; OVLT, organum vasculosum of laminaterminalis; SFO, subfornical organ; CNS, central nervous system.*To whom reprint requests should be addressed.
10985
saline alone. Animals were then killed by decapitation at 0.5,1, 2, 4, 8, 12, and 24 hr after the injection (n 5 3–5 per timepoint). Five other groups of animals were injected intrave-nously (i.v.) with 1, 5, 25, or 125 mgykg or with 2.5 mgykg ofLPS (n 5 3–5 per dose) and killed 3 hr after the injection. Tocontrol for the effects due to saline injection alone, controlanimals that received no injection were also killed (n 5 3).Injections were timed so that the time of sacrifice fell between09:00 and 13:00 hr.
Brain Section Collection. Brains were removed immediatelyafter decapitation, frozen by immersion in 2-methyl butane at230°C, and stored at 270°C before sectioning. They were thencryostat-cut to 15-mm-thick coronal sections and thaw-mounted onto gelatin-coated slides, dried, and stored at 240°Cuntil further processing. Levels collected were organum vas-culosum of the lamina terminalis (OVLT) (20.02 mm relativeto bregma); subfornical organ (SFO) (20.92 mm); centralnucleus of amygdala containing also the arcuate nucleus andmedian eminence (23.3 mm); and area postrema (AP) con-taining the nucleus of the solitary tract (NTS) (213.7 mm)(11).
In Situ Hybridization. The in situ hybridization protocolswere performed as described previously for ribonucleotide(cRNA) probes (12). First, tissue sections were processed byfixation with 4% formaldehyde solution, acetylation with0.25% acetic anhydride in 0.1 M triethanolamine-HCl (pH 8.0)solution, dehydration with ethanol, and delipidation withchloroform.
Second, the antisense probes directed against the full-length(1.05 kb) rat IkBa cDNA inserted into the pBluescript plasmid(generously provided by Rebecca Taub, University of Penn-sylvania) was transcribed using the Riboprobe System (Pro-mega) with T7 RNA polymerase and [a-35S]UTP (specificactivity . 1,000 Ciymmol; New England Nuclear; 1 Ci 5 37GBq) after linearization with BamHI restriction enzyme (Pro-mega). To control for the specificity of the probe, sense probesof rat IkBa were also generated by transcribing the sameplasmid with T3 RNA polymerase after linearization withHindIII restriction enzyme (Promega). Finally, radiolabeledprobes were diluted in the riboprobe hybridization buffer andapplied to brain sections (500,000 cpmysection). After over-night incubation at 55°C in a humidified chamber, slidescontaining brain sections were washed first in 20 mgyml RNasesolution and then in 23 standard saline citrate (SSC) and 0.23SSC (55°C and 60°C) solutions to reduce nonspecific bindingof the probe. The slides were then dehydrated with ethanol andair-dried for autoradiography.
Autoradiography. Slides and 14C plastic standards contain-ing known amounts of radioactivity (American Radiochemi-cals, St. Louis) were placed in x-ray cassettes, apposed to film(BioMax MR; Kodak) for 4 days, and developed in an auto-matic film developer (X-Omat; Kodak). To determine ana-tomical localization of hybridized probes at the cellular level,sections were dipped in nuclear track emulsion (NTB-2;Kodak), exposed for 2 weeks, developed (D19; Kodak) for 2min at 16°C, and counterstained with cresyl violet.
Combined Immunohistochemistry and Hybridization His-tochemistry. Four cell type-specific antibodies were used todetermine the phenotypes of IkBa mRNA-producing cells:anti-PECAM (platelet and endothelial cell adhesion molecule,generously donated to us by W. F. Hickey, Dartmouth MedicalSchool); anti-GFAP (glial fibrillary acidic protein; ICN);OX-42 (PharMingen), and ED2 (Serotec). These antibodiesspecifically mark endothelial cells (13), astrocytes, microglia,and perivascular monocytes (14), respectively. The doublelabeling was carried out as follows. Fresh-frozen slide-mounted brain sections were fixed in 4% formaldehyde for 15min and rinsed three times in PBS; they were then incubatedwith the above primary antibodies diluted in PBS (1:400 foranti-PECAM, anti-GFAP, and ED2; 1:2000 for OX-42) for 2
hr. After three rinses in PBS, sections were incubated with thebiotinylated secondary antibody (goat anti-mouse IgG, 1:200)for 1 hr, and antibody bindings were visualized by the con-ventional avidin-biotin immunoperoxidase protocol. Thesesections were then re-fixed in 4% formaldehyde, treated with1 mlyml proteinase K for 15 min, acetylated, and dehydrated.Subsequent localization of IkBa mRNA by in situ hybridiza-tion was then carried out as described above.
Data Analysis. Autoradiographic film images of brain sec-tions and standards were digitized on a Macintosh computer-based image analysis system with IMAGE software (WayneRasband, National Institute of Mental Health). Light trans-mittance through the film was measured by outlining thestructure on the TV monitor. A density-slice function wasapplied to each structure to select densities greater than filmbackground and thus measured transmittance confined to thecellular sources of the radioactivity. The density so obtainedwas used to represent the relative amount of mRNA expressionof IkBa.
RESULTS
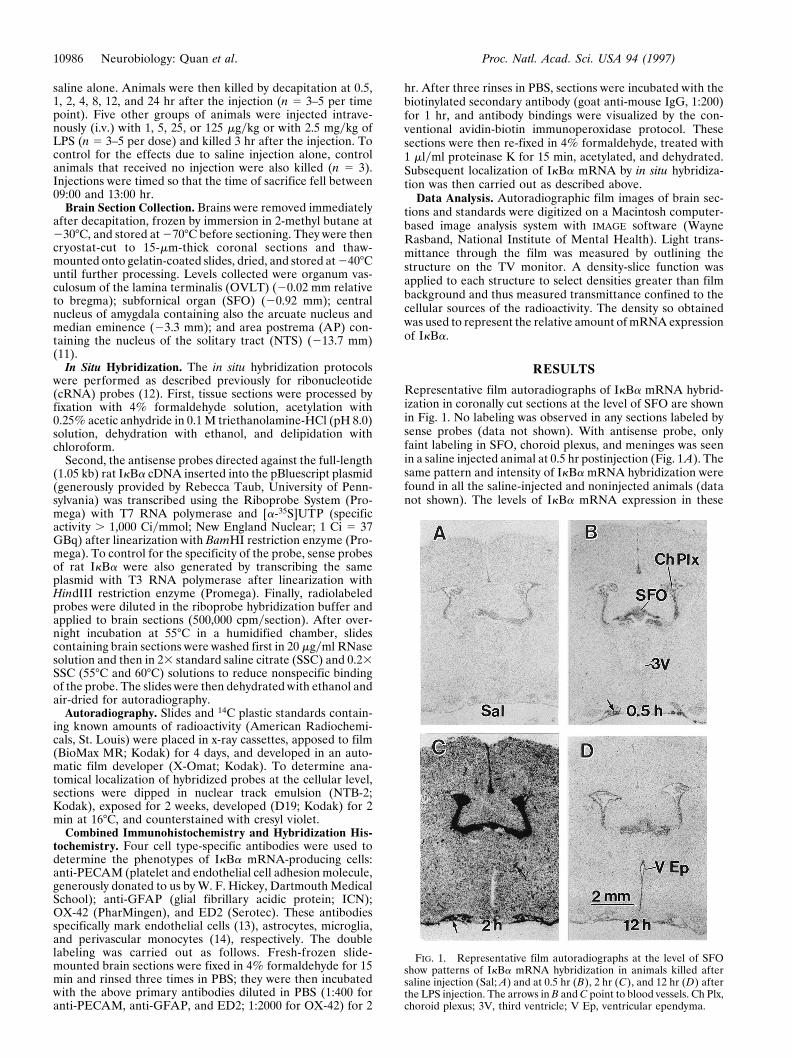
Representative film autoradiographs of IkBa mRNA hybrid-ization in coronally cut sections at the level of SFO are shownin Fig. 1. No labeling was observed in any sections labeled bysense probes (data not shown). With antisense probe, onlyfaint labeling in SFO, choroid plexus, and meninges was seenin a saline injected animal at 0.5 hr postinjection (Fig. 1A). Thesame pattern and intensity of IkBa mRNA hybridization werefound in all the saline-injected and noninjected animals (datanot shown). The levels of IkBa mRNA expression in these
FIG. 1. Representative film autoradiographs at the level of SFOshow patterns of IkBa mRNA hybridization in animals killed aftersaline injection (Sal; A) and at 0.5 hr (B), 2 hr (C), and 12 hr (D) afterthe LPS injection. The arrows in B and C point to blood vessels. Ch Plx,choroid plexus; 3V, third ventricle; V Ep, ventricular ependyma.
10986 Neurobiology: Quan et al. Proc. Natl. Acad. Sci. USA 94 (1997)
animals were measured by densitometry and designated asbasal levels. The density of labeling measured in any otherconditions will be expressed in parentheses as the percentageof these basal levels. The SEM was typically ,10% of the meanlevels of radioactivity.
After LPS injection, induction of IkBa was found at severaltime points. At 0.5 hr, induction of IkBa mRNA was apparentin SFO (122%), choroid plexus (287%), and large blood vesselsat the base of the brain (Fig. 1B). At 2 hr, the LPS injectioninduced maximal levels of IkBa mRNA observed in thisexperiment (Fig. 1C) in SFO (924%) and choroid plexus(362%). In addition, meninges at the base of the brain, smallpenetrating blood vessels, and scattered cells inside the brainwere now strongly expressing IkBa mRNA. At 4 and 8 hr,expression levels in all the previously stimulated regions exceptmeninges had declined to near the basal levels (data notshown). At 12 hr, IkBa message in SFO (119%), choroid plexus(117%), and blood vessels was still just slightly above the basallevels (Fig. 1D). A previously unaffected structure, the ven-tricular ependyma, by contrast, was now strongly labeled (Fig.1D). There was a 97% increase of hybridization signal over the
basal level in these cells. The induction of IkBa in themeningeal cells at the base of the brain persisted until this time;i.e., the labeling density was still three times the basal level, andit was three fourths of the maximal level seen at 2 hr. At 24 hr,the IkBa expression in all the brain regions declined towardthe basal levels (data not shown).
The global pattern of the induction of IkBa mRNA at otherlevels, such as OVLT and central nucleus of amygdala con-taining median eminence, was similar to the pattern seen atSFO level; i.e., IkBa mRNA was first induced in blood vessels,CVOs, and choroid plexus, then in cells of the meninges andbrain parenchyma, and finally in ventricular ependyma whilepreviously activated regions declined. However, two interest-ing differences were observed at the level of AP.
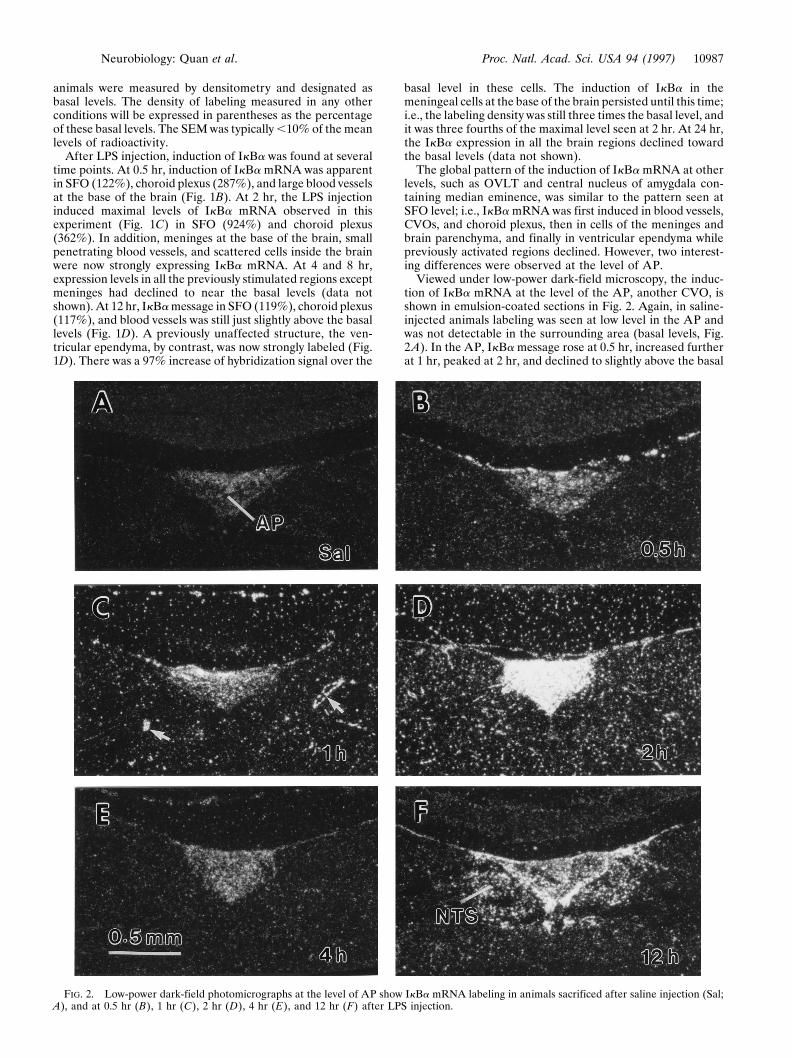
Viewed under low-power dark-field microscopy, the induc-tion of IkBa mRNA at the level of the AP, another CVO, isshown in emulsion-coated sections in Fig. 2. Again, in saline-injected animals labeling was seen at low level in the AP andwas not detectable in the surrounding area (basal levels, Fig.2A). In the AP, IkBa message rose at 0.5 hr, increased furtherat 1 hr, peaked at 2 hr, and declined to slightly above the basal
FIG. 2. Low-power dark-field photomicrographs at the level of AP show IkBa mRNA labeling in animals sacrificed after saline injection (Sal;A), and at 0.5 hr (B), 1 hr (C), 2 hr (D), 4 hr (E), and 12 hr (F) after LPS injection.
Neurobiology: Quan et al. Proc. Natl. Acad. Sci. USA 94 (1997) 10987
level by 4 hr (Fig. 2 B–E). At 12 hr, however, strong IkBalabeling reappeared once again (Fig. 2F) before it subsidedtoward the basal level at 24 hr (data not shown). In the tissuesurrounding the AP, induced IkBa message was just detectable
at 0.5 hr, became apparent in small penetrating blood vesselsand in scattered small cells of the brain parenchyma at 1 hr,intensified to the maximal level in evenly distributed small cellsof the brain parenchyma at 2 hr, and declined at 4 hr. At 12 hr,however, strong labeling also reappeared, but this time re-stricted only to small cells in the area of the NTS.
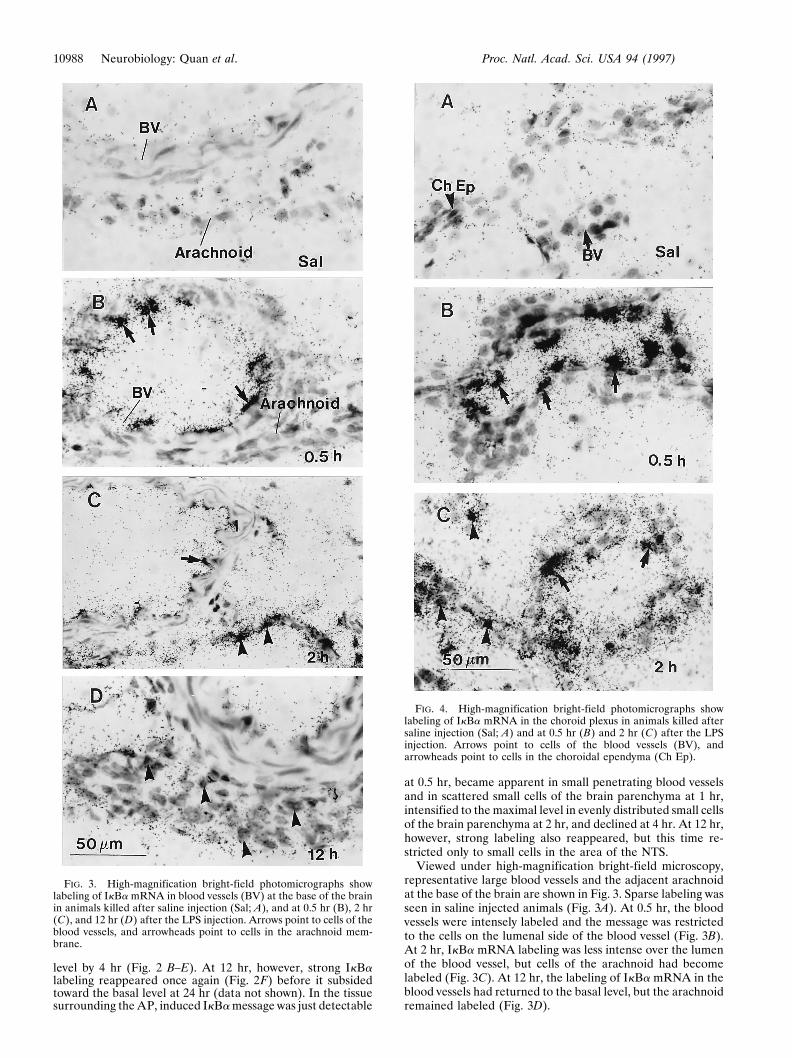
Viewed under high-magnification bright-field microscopy,representative large blood vessels and the adjacent arachnoidat the base of the brain are shown in Fig. 3. Sparse labeling wasseen in saline injected animals (Fig. 3A). At 0.5 hr, the bloodvessels were intensely labeled and the message was restrictedto the cells on the lumenal side of the blood vessel (Fig. 3B).At 2 hr, IkBa mRNA labeling was less intense over the lumenof the blood vessel, but cells of the arachnoid had becomelabeled (Fig. 3C). At 12 hr, the labeling of IkBa mRNA in theblood vessels had returned to the basal level, but the arachnoidremained labeled (Fig. 3D).
FIG. 3. High-magnification bright-field photomicrographs showlabeling of IkBa mRNA in blood vessels (BV) at the base of the brainin animals killed after saline injection (Sal; A), and at 0.5 hr (B), 2 hr(C), and 12 hr (D) after the LPS injection. Arrows point to cells of theblood vessels, and arrowheads point to cells in the arachnoid mem-brane.
FIG. 4. High-magnification bright-field photomicrographs showlabeling of IkBa mRNA in the choroid plexus in animals killed aftersaline injection (Sal; A) and at 0.5 hr (B) and 2 hr (C) after the LPSinjection. Arrows point to cells of the blood vessels (BV), andarrowheads point to cells in the choroidal ependyma (Ch Ep).
10988 Neurobiology: Quan et al. Proc. Natl. Acad. Sci. USA 94 (1997)
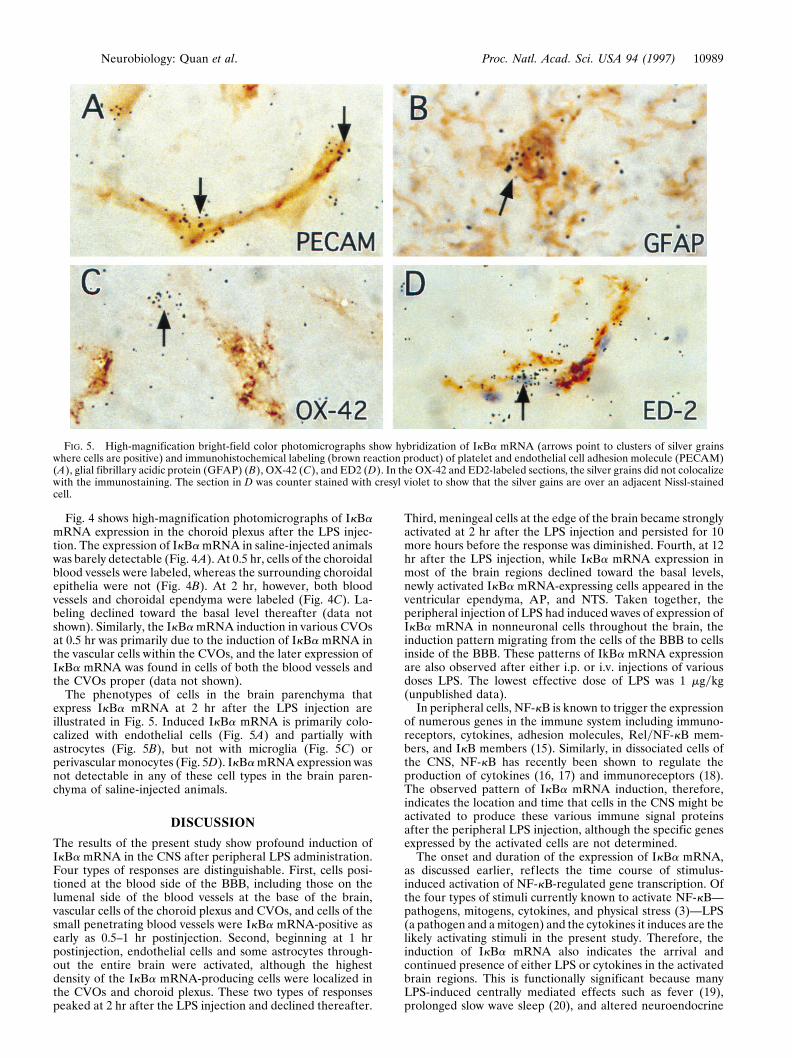
Fig. 4 shows high-magnification photomicrographs of IkBamRNA expression in the choroid plexus after the LPS injec-tion. The expression of IkBa mRNA in saline-injected animalswas barely detectable (Fig. 4A). At 0.5 hr, cells of the choroidalblood vessels were labeled, whereas the surrounding choroidalepithelia were not (Fig. 4B). At 2 hr, however, both bloodvessels and choroidal ependyma were labeled (Fig. 4C). La-beling declined toward the basal level thereafter (data notshown). Similarly, the IkBa mRNA induction in various CVOsat 0.5 hr was primarily due to the induction of IkBa mRNA inthe vascular cells within the CVOs, and the later expression ofIkBa mRNA was found in cells of both the blood vessels andthe CVOs proper (data not shown).
The phenotypes of cells in the brain parenchyma thatexpress IkBa mRNA at 2 hr after the LPS injection areillustrated in Fig. 5. Induced IkBa mRNA is primarily colo-calized with endothelial cells (Fig. 5A) and partially withastrocytes (Fig. 5B), but not with microglia (Fig. 5C) orperivascular monocytes (Fig. 5D). IkBa mRNA expression wasnot detectable in any of these cell types in the brain paren-chyma of saline-injected animals.
DISCUSSION
The results of the present study show profound induction ofIkBa mRNA in the CNS after peripheral LPS administration.Four types of responses are distinguishable. First, cells posi-tioned at the blood side of the BBB, including those on thelumenal side of the blood vessels at the base of the brain,vascular cells of the choroid plexus and CVOs, and cells of thesmall penetrating blood vessels were IkBa mRNA-positive asearly as 0.5–1 hr postinjection. Second, beginning at 1 hrpostinjection, endothelial cells and some astrocytes through-out the entire brain were activated, although the highestdensity of the IkBa mRNA-producing cells were localized inthe CVOs and choroid plexus. These two types of responsespeaked at 2 hr after the LPS injection and declined thereafter.
Third, meningeal cells at the edge of the brain became stronglyactivated at 2 hr after the LPS injection and persisted for 10more hours before the response was diminished. Fourth, at 12hr after the LPS injection, while IkBa mRNA expression inmost of the brain regions declined toward the basal levels,newly activated IkBa mRNA-expressing cells appeared in theventricular ependyma, AP, and NTS. Taken together, theperipheral injection of LPS had induced waves of expression ofIkBa mRNA in nonneuronal cells throughout the brain, theinduction pattern migrating from the cells of the BBB to cellsinside of the BBB. These patterns of IkBa mRNA expressionare also observed after either i.p. or i.v. injections of variousdoses LPS. The lowest effective dose of LPS was 1 mgykg(unpublished data).
In peripheral cells, NF-kB is known to trigger the expressionof numerous genes in the immune system including immuno-receptors, cytokines, adhesion molecules, RelyNF-kB mem-bers, and IkB members (15). Similarly, in dissociated cells ofthe CNS, NF-kB has recently been shown to regulate theproduction of cytokines (16, 17) and immunoreceptors (18).The observed pattern of IkBa mRNA induction, therefore,indicates the location and time that cells in the CNS might beactivated to produce these various immune signal proteinsafter the peripheral LPS injection, although the specific genesexpressed by the activated cells are not determined.
The onset and duration of the expression of IkBa mRNA,as discussed earlier, reflects the time course of stimulus-induced activation of NF-kB-regulated gene transcription. Ofthe four types of stimuli currently known to activate NF-kB—pathogens, mitogens, cytokines, and physical stress (3)—LPS(a pathogen and a mitogen) and the cytokines it induces are thelikely activating stimuli in the present study. Therefore, theinduction of IkBa mRNA also indicates the arrival andcontinued presence of either LPS or cytokines in the activatedbrain regions. This is functionally significant because manyLPS-induced centrally mediated effects such as fever (19),prolonged slow wave sleep (20), and altered neuroendocrine
FIG. 5. High-magnification bright-field color photomicrographs show hybridization of IkBa mRNA (arrows point to clusters of silver grainswhere cells are positive) and immunohistochemical labeling (brown reaction product) of platelet and endothelial cell adhesion molecule (PECAM)(A), glial fibrillary acidic protein (GFAP) (B), OX-42 (C), and ED2 (D). In the OX-42 and ED2-labeled sections, the silver grains did not colocalizewith the immunostaining. The section in D was counter stained with cresyl violet to show that the silver gains are over an adjacent Nissl-stainedcell.
Neurobiology: Quan et al. Proc. Natl. Acad. Sci. USA 94 (1997) 10989

activity (21) have been attributed to the pro-inflammatorycytokines that LPS induces. Therefore, the following discus-sion will focus on the pathways, suggested by the presentresults, by which cytokines might arrive in the CNS after theLPS injection.
A salient finding of the present study is the initial responseafter the LPS injection in cells of the cerebrovasculature. Theearly responsiveness of these cells may be expected becausethey are readily accessible by circulating LPS and cytokines.Cells inside the BBB, by contrast, are not freely accessible byblood-borne LPS (8) and cytokines [although small amounts ofcirculating cytokines have been reported to be transportedacross the BBB (22)]. In addition, receptors for LPS (23, 24),tumor necrosis factor a (25), and interleukin 1 (26) have beenlocalized in the endothelial cells of brain vasculature. More-over, the activation of the NF-kB system in these cells suggeststhat they respond to the peripheral stimulation by producingtheir own signal molecules, probably cytokines. This is sup-ported by our recent finding that interleukin 1b mRNA israpidly synthesized in the CVOs and in brain vascular cellsafter peripheral LPS injection (27).
At 2 hr after the LPS injection, many astrocytes throughoutthe entire brain were seen to express IkBa mRNA. Thissuggests that cytokines signals have now reached brain paren-chyma, and, importantly, the responsive astrocytes are prob-ably producing centrally derived cytokines. It is noteworthythat the activation of cells in the brain proper is uniform,independent of the distance from the CVOs (comparing Fig.2 D to F). This is significant because it has been shown that iflarge molecules excluded by the BBB were to enter the brain,they will first do so at the CVOs and then diffuse slowly to thevicinity of these structures (9), displaying concentration gra-dients moving away from the CVOs, similar to that seen in Fig.2F. The activation pattern we observed in the brain paren-chyma at 2 hr after the LPS injection (Figs. 1C and 2D),therefore, is not likely to be produced by LPS and cytokinesthat arrived by this pathway. The pattern may be best explainedby cytokines that were either generated in the cells of brainblood vessels or transported from circulation across the entireBBB. Because only small amounts of certain cytokines (i.e.,less than 5% of the blood levels) have been shown to betransported across the BBB (22), it is possible that the majorityof the cytokines present in CNS at 2 hr was synthesized by theearlier observed IkBa mRNA-expressing vascular cells. Pre-sumably, these cells secrete cytokines on their ablumenal side.Indeed, we found numerous endothelial cells expressing IkBamRNA.
The highest local concentrations of IkBa mRNA-expressingcells were seen in the CVOs, choroid plexus, and meninges at2 hr after the LPS injection. This is probably due to theconcentration of cytokine-responsive cells in these sites. Forexample, it has been shown that type 1 interleukin 1 receptorsare highly concentrated in AP, choroid plexus, and meninges(28). In addition, these sites may also receive stimulation bysubstantial amounts of peripheral cytokines because theypossess a leaky BBB (9). The present results suggest that at 2hr after the LPS injection, the main cytokine-producing cellsare located in these sites. Because the CVOs, choroid plexus,and meninges are also bathed by interstitial cerebrospinal f luid(CSF), molecules secreted from these cells will be transportedby CSF flow. The persistent activation of the meningeal cellsat the base of the brain after this time may result from thestimulation by cytokines flowing in the exiting subarachnoidCSF. On the other hand, cytokines secreted from these cellsmay also be delivered by the ventricular CSF to brain paren-chyma, especially to structures in the close vicinity of theventricle.
The finding that IkBa mRNA was induced in cells of theventricular ependyma at 12 hr after the peripheral LPS
injection while most of the CVOs and blood vessels were nolonger active is very interesting, but puzzling. Because thesecells were not stimulated earlier, the extracellular stimuli thatare present at 12 hr may be different from those present atearlier time points. It is not clear, however, whether thesestimuli arrived from peripheral circulation or from late re-sponsive cells of the CNS. The close vicinity of these cells tothe ventricles suggests that ventricular CSF provides a func-tional conduit for these signal molecules.
Beyond suggesting pathways for the transduction of periph-eral immune signals into the brain, the present results may haveimportant implications for the pathogenesis of viral infectionsof the CNS. This is because the replication of many viruses,including HIV, are regulated by NF-kB (3). It is conceivablethat viral proliferation in the CNS could be facilitated whenperipheral infections induce NF-kB activation in the cells ofthe CNS similar to that seen in the present study.
1. Hopkins, S. J. & Rothwell, N. J. (1995) Trends Neurosci. 18,83–88.
2. Muller, J. M., Ziegler-Heitbrock, H. W. & Baeuerle, P. A. (1993)Immunobiology 187, 233–256.
3. Miyamoto, S. & Verma, I. M. (1995) Adv. Cancer Res. 66,255–292.
4. Cheng, Q., Cant, C. A., Moll, T., Hofer-Warbinek, R., Wagner,E., Birnstiel, M. L., Bach, F. H. & de Martin, R. (1994) J. Biol.Chem. 269, 13551–13557.
5. Henkel, T., Machleidt, T., Alkalay, I., Kronke, M., Ben-Neriah,Y. & Baeuerle, P. A. (1993) Nature (London) 365, 182–185.
6. Beg, A. A., Finco, T. S., Nantermet, P. V. & Baldwin, A. S., Jr.(1993) Mol. Cell. Biol. 13, 3301–3310.
7. Kruijer, W., Cooper, J. A., Hunter, T. & Verma, I. M. (1984)Nature 312, 711–6.
8. Schmahl, F. W., Schlote, W. & Reinhard, U. (1976) Curr. Top.Crit. Care Med. 1, 171–178.
9. Broadwell, R. D. & Sofroniew, M. V. (1993) Exp. Neurol. 120,245–263.
10. Verma, I. M., Stevenson, J. K., Schwarz, E. M., Van Antwerp, D.& Miyamoto, S. (1995) Genes Dev. 9, 2723–2735.
11. Paxinos, G. & Watson, C. (1986) The Rat Brain in StereotaxicCoordinates (Academic, Orlando).
12. Whitfield, H. J., Jr., Brady, L. S., Smith, M. A., Mamalaki, E., Fox,R. J. & Herkenham, M. (1990) Cell. Mol. Neurobiol. 10, 145–157.
13. Williams, K. C., Zhao, R. W., Ueno, K. & Hickey, W. F. (1996)J. Neurosci. Res. 45, 747–757.
14. Angelov, D. N., Neiss, W. F., Streppel, M., Walther, M., Gun-tinas-Lichius, O. & Stennert, E. (1996) Glia 16, 129–139.
15. Grilli, M., Chiu, J. J. & Lenardo, M. J. (1993) Int. Rev. Cytol. 143,1–62.
16. Fisher, S. N., Kim, Y. U., Shin, M. L., Sparacio, S. M., Zhang, Y.,Vilcek, J. & Benveniste, E. N. (1994) J. Immunol. 153, 3210–3217.
17. Moynagh, P. N., Williams, D. C. & O’Neill, L. A. (1994)J. Immunol. 153, 2681–2690.
18. Massa, P. T., Ozato, K. & McFarlin, D. E. (1993) Glia 8, 201–207.19. Blatteis, C. M. (1990) Yale J. Biol. Med. 63, 133–146.20. Krueger, J. M. & Majde, J. A. (1995) Int. Arch. Allergy Immunol.
106, 97–100.21. McCann, S. M., Lyson, K., Karanth, S., Gimeno, M., Belova, N.,
Kamat, A. & Rettori, V. (1995) Ann. N.Y. Acad. Sci. 771,386–395.
22. Banks, W. A., Kastin, A. J. & Broadwell, R. D. (1995) Neuro-immunomodulation 2, 241–248.
23. Noel, R. F., Jr., Sato, T. T., Mendez, C., Johnson, M. C. &Pohlman, T. H. (1995) Infect. Immun. 63, 4046–4053.
24. Read, M. A., Cordle, S. R., Veach, R. A., Carlisle, C. D. &Hawiger, J. (1993) Proc. Natl. Acad. Sci. USA 90, 9887–9891.
25. Bebo, B. F., Jr., & Linthicum, D. S. (1995) J. Neuroimmunol. 62,161–167.
26. Van Dam, A. M., De Vries, H. E., Kuiper, J., Zijlstra, F. J., DeBoer, A. G., Tilders, F. J. & Berkenbosch, F. (1996) FASEB J. 10,351–356.
27. Quan, N., Whiteside, M. & Herkenham, M. (1997) Neuroscience,in press.
28. Ericsson, A., Liu, C., Hart, R. P. & Sawchenko, P. E. (1995)J. Comp. Neurol. 361, 681–698.
10990 Neurobiology: Quan et al. Proc. Natl. Acad. Sci. USA 94 (1997)