incompatibility groups the classification f- …vol. 112, 1972 r factor incompatibility groups 667...
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, Nov. 1972, p. 666-675Copyright 0 1972 American Society for Microbiology
Vol. 112, No. 2Printed in U.S.A.
Incompatibility Groups and the Classification off- Resistance Factors
Y. A. CHABBERT, M. R. SCAVIZZI, J. L. WITCHITZ, G. R. GERBAUD, AND D. H. BOUANCHAUD
Service de Bacteriologie Medicale, Institut Pasteur, Paris 75015, France
Received for publication 2 June 1972
Incompatibility between R factors has been reported by several authors, andfour incompatibility groups have already been described by Datta and Hedgesamong Rfi- factors. The stability of 12 plasmids in pairs was studied after 116crosses, and five new groups were found, designated 5, 6, 7, 8, and 9. Eachplasmid studied belongs to one single group. Incompatibility between plasmidsin pairs is a clear-cut phenomenon, is easy to observe, and can provide a reli-able method for recognizing and classifying resistance factors, and for tracingtheir spread among bacterial species.
Previous studies on drug resistance factorshave shown that these plasmids can be classi-fied in two groups, Rfi- and Rfi-, according totheir effects on the fertility of male strains ofEscherichia coli (Watanabe et al., 26). Rfi+factors, when present in an Hfr strain, are alsoable to prevent the lysis of this strain by male-specific phages. This property is frequentlyused to distinguish Rfi- and Rfi - factors, andit was shown by Egawa and Hirota (9) thatRfi' factors repress the synthesis of F pili bymale strains of E. coli.Bacteria carrying transmissible R factors
synthetize external appendages called R pili.On the basis of morphological studies of thesepili, it was suggested that Rfi- pili were re-lated to F pili and Rfi- pili to I pili (i.e., pilisynthetized by bacteria carrying a ColI factor).Consequently Rfi - factors were also called "F-like" or "R(f)," and Rfi- factors "I-like" or"R(i)." These facts have been extensively dis-cussed by Meynell, Meynell and Datta (15)and by Novick (17). However, it was observedlater that some Rfi+ factors govern the syn-thesis of "I-like" pili (10, 22), and it becameclear that these different properties cannot beused indiscriminately to recognize and classifyR factors.
Independently, it was observed by Watanabeet al. (26) that two plasmids cannot stablycoexist in bacteria if both are Rfi+ or Rfi -,probably because they compete for the samereplication site. This phenomenon, called "in-compatibility," can be easily observed withsome pairs of R factors; but in fact many Rfi-factors are compatible with one another in
naturally occuring strains or in E. coli K-12 (2,4, 12, 21). So far, incompatibility between Rfactors in pairs seems the best way to classifythese plasmids because gene(s) involved in thisphenomenon might be essential to ensure thereplication and the apportionment of thesereplicons in daughter cells (16). Such incom-patibility groups among R factors could be com-pared to classes I and II described by Peyru,Wexler, and Novick among penicillinase plas-mids in Staphylococcus aureus (19).
Recently, Datta and Hedges studied the sta-bility of R factors in pairs and their frequencyof transfer (8). By these criteria, three groupswere found and called I, N, and P. Later, afourth group was discovered, and referred to asW (11). At the same time, we were engaged instudying the incompatibility phenomenonamong a large set of plasmids of clinical origin.Additional crosses were performed to checkwhether the groups we observed were compa-rable with groups I, N, W, and P.The present paper will confirm the existence
of groups I and N, and demonstrate that manyother classes exist among plasmids. Theseclasses can be separated by using the incom-patibility phenomenon as the sole criterion.We will discuss the possibility of identifyingand classifying R factors on this basis, and ofdeveloping an epidemiology of these geneticelements.
MATERIALS AND METHODSBacterial strains and R factors. Twelve natu-
rally occuring strains and their R factors are de-scribed in Table 1. All R factors were transferred by
666
on March 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

VOL. 112, 1972 R FACTOR INCOMPATIBILITY GROUPS 667
TABLE 1. Naturally occurring strains and plasmids
Origin of strains R factorsa References
Salmonella paratyphi B 1965, feces, Tours R, amp 3Providencia stuartii 1966, abscess, Paris (St-An- R,, kanP sul 3
toine Hospital)Shigella flexneri 1966, feces, Strasbourg R22 amp str chi tetB sul 5Pseudomonas aeruginosa 1969, urine, Paris (Necker R40a amp kanP sul 1
Hospital)Klebsiella pneumoniae 1969, urine, Paris (Claude- R55 amp chl sul genK 27, 28Bernard Hospital)
Salmonella paratyphi B 1970, feces, Creteil R., amp kanP tetBEscherichia coli 1969, feces, Kremlin-Bicetre Hos- R7, amp str chl tetB sul 24
pital
Klebsiella pneumoniae 1970, feces, Creteil R,5 amp kanP tetB _bSalmonella panama 1968, blood, Kremlin-Bicetre 4
Hospital R,,, ampR,,2kanP 4R,1tetA 4
Escherichia coli 1958, feces, Quebec R ,c str kanP _c
a See key for resistance markers in Materials and Methods.b Obtained from Dr. Sole, Creteil Hospital.c Obtained from Dr. Poty, Laval University, Quebec.
conjugation to mutants of E. coli K-12 strains (C600from E. Wollman, J5 from N. Datta, IP 54117 fromL. Le Minor), resistant to sodium azide (Azir) or ri-fampin (Rifr).
Strains carrying plasmids representative of groupsN, W, and P were kindly furnished by N. Datta.These factors were RN3 str tetA sul (group N), R.-chl kanA str sul (group W), and R,, ampC kanAtetA (group P). R64 str tetA (drd mutant), obtainedfrom E. Meynell, is representative of group I.
Symbols for resistance characters are: amp, ampi-cillin; ampC, ampicillin and carbenicillin; chl, chlor-amphenicol; kanA, kanamycin; kanP, kanamycinand paromomycin; genK, gentamicin and kana-mycin (27, 28); tetA, tetracycline; tetB, tetracyclineand minocycline (23); str, streptomycin; sul, sulfona-mides.
Transfer, transduction, and stability of R fac-tors in pairs. Routine procedure for transfer of Rfactors was as described by Watanabe and Fukasawa(25). Concentrations and origin of drugs were de-scribed in a previous paper by Bouanchaud andChabbert (4). When the frequency of transfer was
very low, donor and recipient bacteria were growntogether on cellophane membranes lying on nutrientagar for 24 hr before selection of resistant recipients(6). Frequency of transfer was expressed as thenumber of resistant recipients per number of donors.Selection was carried out for plasmid-plasmid or
plasmid-chromosome markers; the stability of pairsof plasmids was scored after 10 to 30 generations indrug-free medium, by checking the resistance of theprogeny of five to ten colonies by the replica-platingmethod. If the bacteria were still resistant to alldrugs, an additional cross was systematically carried
out, to make sure that both plasmids were separatelytransferable and had not recombined.
All R factors studied here were transduced "enbloc" by phage Plkc (obtained from F. Jacob) be-tween E. coli K-12 strains. Transduced R factorswere transferred to E. coli recA - (from E. Dubnau)to check their replicative autonomy, and then trans-ferred to an Hfr strain (from F. Jacob) to test theactivity of male-specific phages f2 and MS2 (ob-tained from L. Le Minor and E. Meynell). Thestrains remained sensitive to these phages, and all Rfactors, according to this indirect test, are fi-. Thisseries of rather complicated controls was done for allR factors before their compatibility in pairs wasstudied. Transductions also provided derivatives ofR factors with deletions of various resistancemarkers; these derivatives are listed in Table 2. De-rivative R7,1l was obtained after nitrosoguanidinetreatment of R7,. Symbol "del" refers to deletedmarkers.
RESULTSThe stability of 116 pairs of plasmids, in-
cluding plasmids representative of groups I, N,W, and P (8, 11), was studied as describedabove. Only nine pairs were found unstable.Three pairs of isogenic plasmids with deletionsaffecting convenient resistance markers werealso studied and were found to be unstable. Allother pairs of plasmids were stable; we reportthe details of the results below.Incompatibility between isogenic plas-
mids. Derivatives of three plasmids, R22 amp
on March 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

~~~~~CHABBERT ETAL.J.Bc
str chl tetB sul, R69~amp kanP tetBl, and R,5amp chl sul genK, were obtained sponta-neously or during transduction by phage Plkc(see Table 2). These derivatives, which hadundergone deletions of varying numbers of re-
TABLE~2. Derivatives of wild-type R factorSa
Wild-type R factors Derivatives
R22 amp str chi tetB sul R22, amp tetB Strde,chldel SUldel
R,22- amp str chi sultetBdlel
R,, amp chi sul genK R,,-, chi sul aMpdelgenKdel
R,, amp kanP tetB R69-, amp tetB kanplelR6-2 amp kanP tetBdel
R7, amp str chi tetB sul R,,-, amp str chi sultetBdel
R,, amp kanP tetB R,,-, amp tetB kanFpdelR,,S str tetA sul RN3l str sul tetAdel
aSee key for resistance markers in Materials andMethods. Symbol "del" refers to deleted markers.
.sistance markers, were R22 1 amp tetB and
R22 2amp str chi sul; R,691 amp tetB and R,,-2
amp kanP; and R,,-, chl sul. Results of crosses
between strains harboring derivatives of plas-mids R22, R69,, and R,, can be seen in Table 3.
For pairs R22- /R22-2 and R69-/R692I, selection
for a marker of the entering plasmid and a
chromosomal marker of the recipient led to an
exclusion of the resident plasmid in 98 to 100%
of the host bacteria. For pair R,,/R,,-,, no ex-
clusion of the resident plasmid (R,,-,) was ob-
served; on the contrary, the entering plasmid
(R,,) was lost in all bacteria grown in drug-free
medium. R,,-, chl sul remained transferable by
conjugation, so its possible integration in the
host chromosome can be excluded.
The percentage of bacteria carrying both
plasmids after subculture in drug-free medium
was 0 for derivatives of R22and R,,; it was 1%
for plasmids derived from R69.
An additional plasmid-plasmid selection,
not reported in Table 3, was carried out for
pair R69-, amp tetB/RI,9~2 amp kanP. In that
TABLE 3. Incompatibility between isogenic plasmids
FrequencyDonors and recipientS" Selectors of lb 2C 3d Phenotypes 4e 5f
transfer
D: R22, amp tetB tet + rif 2 x 10-iR: Plasmid-negative
D: R,22, amp tetBR: R,22 amp str chi sul tet + rif 8 x 10-" 3 10 580 Amp Str Chl Sul TetB 0 0
Amp Str Chl Sul 0 0Amp TetB 580 100
30 1,100 Amp Str Chl Sul TetB 0 0Amp Str Chl Sul 0 0Amp TetB 1,100 100
D: R55 amp chi sul genK amp + rif 6 x 10-iR: Plasmid-negative
D: R55 amp chi sul genK amp + rif 5 x 10, 2 20 181 Amp Chl Sul GenK 0 0R: R55_, chi sul Chl Sul 181 100
D: R.,, amp hanP kan + rif 2 x 10'1R: Plasmid-negative
D: R.,, amp kanP kan + rif 1 X 10- 4 3 14 599 Amp KanP TetB 7 1R: R69 amp tetB Amp KanP 588 98
Amp TetB 4 0.60
a"Donors: E. coli J5; recipients: E. coli J5 Rifr. See Materials and Methods for key to resistance markers.bNumber of recombinant colonies studied.cNumber of generations in drug-free.d Total number of colonies studied after subculture in drug-free medium.eNumber of colonies of each type.f'Percentage of colonies of each type.
668 J. BACTERIOL.
on March 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

R FACTOR INCOMPATIBILITY GROUPS
case, it was possible to obtain and maintain an
ampicillin-kanamycin-tetracycline phenotypefor a indefinite number of generations, as longas the subcultures were grown on agar platescontaining kanamycin and tetracycline. How-ever, after one subculture in drug-free me-
dium, all bacteria tested carried only one
plasmid. Exclusion even seems to occur at a
high rate on plates containing both selectors.Isolated colonies were picked from these platesand suspended in broth, and samples were
immediately inoculated in agar plates con-
taining kanamycin or tetracycline, or both, or
no drug. Only 20 to 30% of the bacteria were
able to grow on both selectors, and 30 to 40%on a single selector. Therefore, a large numberof bacteria present in colonies grown on bothselectors are likely to carry only one plasmid,but could remain phenotypically resistant longenough to survive and eventually to grow for a
few generations under these conditions. Ofcourse, segregation occurring during the enu-
meration procedure makes the phenomenondifficult to quantitate.
Incompatibility with I and N resistancefactors. Only three R factors were found to beincompatible with R64 and RN3 (or RN3,-) rep-
resentative of groups I and N. The incompat-ible pairs were R, 2/R64, R113/RN 3- , andR P,3/RN 3. Experimental conditions, resistancecharacters, and results can be seen in Table 4.The "dislodgement" of the resident plasmidby the entering one occurred rapidly and was
observed in 97 to 100% of the recipients testedafter subculture(s) in drug-free medium. Thisphenomenon occurred even when a plasmid-plasmid selection was done for R,,2/R64, butin that case, about 43% of the recipients were
still carrying both plasmids after five genera-
tions in drug-free medium; after 30 genera-
tions, 3% of the recipients carried both plas-mids, 97% carried the entering plasmid and 0%carried the initial resident plasmid. The rea-sons that the resident plasmid is excludedfrom its host strain are not clear, but compa-
rable phenomena have also been observed withF-like colicin factors in an F strain of E. coli(14).From the results reported in Table 4, it can
be concluded that R,13 and RP,3 belong to thesame group than RN 3, and that R,12 belongs tothe same group than R64. No incompatibilitywas found between R factors listed in Table 1and R factors representative of groups P andW.Incompatibility among other factors.
Among plasmids listed in Table 1, nine werefound compatible with plasmids R64, RN3, R8<,
and R,, so they do not belong to the corre-sponding groups I, N, W, and P. However, se-ries of crosses between strains harboring theseplasmids (or derivatives obtained as previouslydescribed) demonstrated the existence of in-compatible pairs among them. These incom-patible pairs were R,I3/RP C3, R22-1/R9R55/R16, R95 /R692, and R40a/R16. Experimen-tal conditions, resistance markers, and resultscan be seen in Table 5. Selection for markersof the entering plasmic and of the chromosomeof the recipient led to an exclusion of the resi-dent plasmid, which was found in less than 1%of the recipients in drug-free medium for pairsR 13/RP3 R55/R4oaR and R55/R16 . The residentplasmid was still present in 45% and 65% of therecipients with pairs R22 J/R and R.5 /R69 2,respectively. An additional compatibility ex-periment (not reported in Table 5) betweenR951 and R,9 2 but with a plasmid-plasmidselection led to similar results. In that case,plasmid-negative recipients were also found,and less than 1% of recipients carried bothplasmids in drug-free medium.When no plasmid markers were available to
check the presence of both genetic elements(R40,, amp kanP sul/R16 kanP sul), additionaltransfers into plasmid-negative E. coli K-12were carried out using recombinant cells asdonors and kanamycin as a selector. The re-sistance of a large number of new recipientswas tested after subculture in drug-free me-dium; all were resistant to ampicillin and kan-amycin, so it can be supposed that plasmidR,6 was no longer present in the recombinantcells, because cotransfer of both R4,a and R,6into all new recipients is very unlikely.Among the progeny of recombinant cells
carrying R22 1 amp tetB/R9 amp, 45% of thebacteria were resistant to ampicillin and car-ried R.; 55% were resistant to ampicillin andtetracycline and could carry R22 l (hypothesis1) or both R22, and R9 (hypothesis 2). Theprogeny of a large number of colonies resistantto ampicillin and tetracycline was studied; allbacteria were found to be resistant to bothdrugs with no exception, and able to transfersimultaneously amp and tetB characters insubsequent crosses. Therefore hypothesis 2 isruled out, and R22, and R, are incompatible.Study of stable pairs of R factors. The
crosses leading to incompatibility have beendescribed in previous sections. All other pairstested in 104 crosses were found to be able tocoexist stably in the recipient host for at least30 generations in drug-free medium. An av-erage of 1,000 colonies were tested after eachtransfer by the replica-plating method. The
669VOL. 112, 1972
on March 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
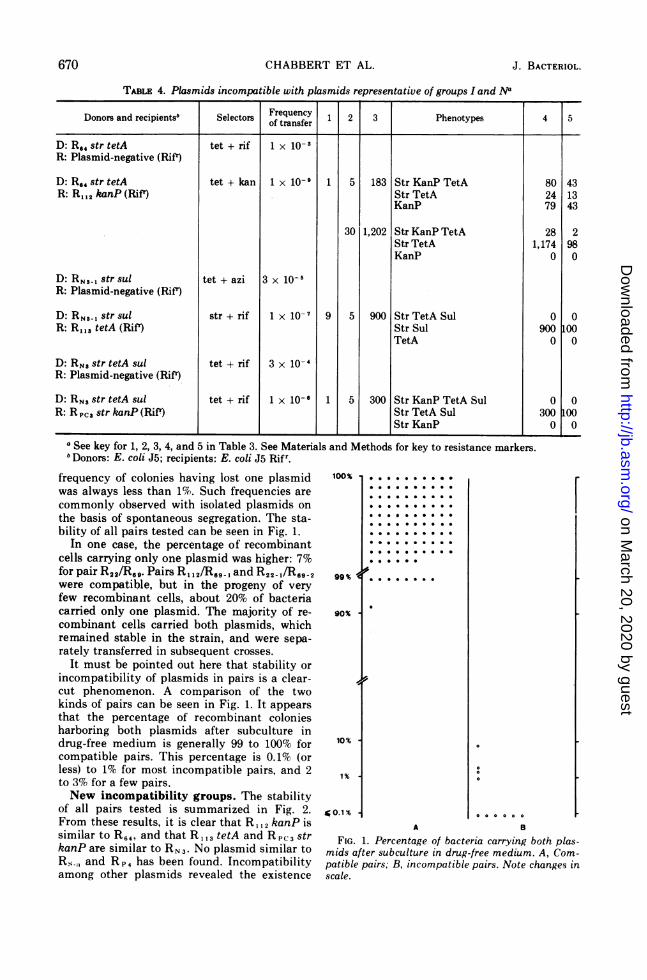
TABLE 4. Plasmids incompatible with plasmids representative of groups I and Na
Donors and recipientsb Selectors Frequency 1 2 3 Phenotypes 4 5of transfer
D: R.4 str tetA tet + rif 1 x 10-3R: Plasmid-negative (Rifr)
D: R.4 str tetA tet + kan 1 x 10-9 1 5 183 Str KanP TetA 80 43R: R112 kanP (RifT) Str TetA 24 13
KanP 79 43
30 1,202 Str KanP TetA 28 2Str TetA 1,174 98KanP 0 0
D: RNS3- str sul tet + azi 3 x 10-6R: Plasmid-negative (Rifr)
D: RNS-I str sul str + rif 1 x 10-7 9 5 900 Str TetA Sul 0 0R: R 13s tetA (Rift) Str Sul 900 100
TetA 0 0
D: RNS str tetA sul tet + rif 3 x 10-4R: Plasmid-negative (RifP)
D: RNS str tetA sul tet + rif 1 x 10-6 1 5 300 Str KanP TetA Sul 0 0R: R PCi str kanP (Rifr) Str TetA Sul 300 100
Str KanP 0 0
aSee key for 1, 2, 3, 4, and 5 in Table 3. See Materia,Donors: E. coli J5; recipients: E. coli J5 Rifr.
frequency of colonies having lost one plasmidwas always less than 1%. Such frequencies arecommonly observed with isolated plasmids onthe basis of spontaneous segregation. The sta-bility of all pairs tested can be seen in Fig. 1.
In one case, the percentage of recombinantcells carrying only one plasmid was higher: 7%for pair R2/R,,6. Pairs R,,2/R.91, and R221JRfI 2were compatible, but in the progeny of veryfew recombinant cells, about 20% of bacteriacarried only one plasmid. The majority of re-combinant cells carried both plasmids, whichremained stable in the strain, and were sepa-rately transferred in subsequent crosses.
It must be pointed out here that stability orincompatibility of plasmids in pairs is a clear-cut phenomenon. A comparison of the twokinds of pairs can be seen in Fig. 1. It appearsthat the percentage of recombinant coloniesharboring both plasmids after subculture indrug-free medium is generally 99 to 100% forcompatible pairs. This percentage is 0.1% (orless) to 1% for most incompatible pairs, and 2to 3% for a few pairs.New incompatibility groups. The stability
of all pairs tested is summarized in Fig. 2.From these results, it is clear that R,12 kanP issimilar to R54, and that R,,3 tetA and R pc3 strkanP are similar to RN3. No plasmid similar toRs,, and Rp4 has been found. Incompatibilityamong other plasmids revealed the existence
als and Methods for key to resistance markers.
100%
99% I
90%
10%
1% I
4 0.1z I I . . . . . .
A BFIG. 1. Percentage of bacteria carrying both plas-
mids after subculture in drug-free medium. A, Com-patible pairs; B, incompatible pairs. Note changes inscale.
670 CHABBERT ET AL. J. BACTERIOL.
0 0 : 0 0 0
0 * 0 0 a 0 a *
.- A I w
on March 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

R FACTOR INCOMPATIBILITY GROUPS
TABLE 5. Incompatibility between other R factorsa
Donors and recipientsb [ Selectors [ transfer J1J 2 3 ] Phenotypes | I i.5
D: R,,, tetAR: Plasmid-negative (Rifr)
D: R,,3 tetAR: Rpc, str kanP(Rifr)
D: R22 1 amp tetBR: Plasmid-negative (Rifr)
D: R,2,- amp tetBR: R, amp (Rifr)
D: R,, amp chi sul genKR: Plasmid-negative (Azir)
D: R,, amp chli sul genKR: R1, kanP sul (Azir)
D: R55 amp chi sul genKR: Plasmid-negative (Azir)
D: R,, amp chl sul genKR: R40,, amp kanP sul (Azir)
D: R9.1 amp tetB (Rifr)R: Plasmid-negative (Azir)
D: R,,-1 amp tetB (Rifr)R: R.9,2 amp kanP (Azir)
D: R,,-1 amp tetB (Rif')R: R,9,2 amp kanP (Azir)
D: R4, amp kanP sulR: Plasmid-negative (Rifr)
D: R4,,, amp kanP sulR: R1,, kanP sul (rif)
tet + rif
tet + rif
1 x 10-4
1 x 10-6
amp + rif 1 x 10- 4
tet + rif
chl + azi
chl + azi
chl + azi
chl + azi
tet + azi
tet + azi
kan + rif
amp + rif
amp + rif
6 x 10-6
1 x 10-i
5 x 10-6
2 + 10-4
8 x 10-6
2 x 10-5
1 x 10-6
2 x 10-7
2 x 10-'
2 x 10-6
1 15
3 130
3
2
3
3
188
663
10 11,106
10 1,981
10
10
518
634
TetAStr KanP TetAStr KanP
Amp TetBAmp
Amp Chl Sul GenK KanPKanP SulAmp Chl Sul GenK
Amp Chl Sul GenK KanPAmp KanP SulAmp Chl Sul GenK
Amp KanP TetBAmp KanPAmp TetBSensitive
Amp KanP TetBAmp KanPAmp TetBSensitive
170180
367296
00
1,106
7143
1,857
434110964
0634
00
9190
5545
00
100
32
93
0.7652112
0100
00
aSee key for 1, 2, 3, 4, and 5 in Table 3, and see Results for pairs R22 J/R, and R40a/R16. See Materials andMethods for key to resistance markers.
b Donors: E. coli J5; recipients: E. coli J5 Rif or Azir.
of new incompatibility classes, and we suggestcalling these classes 5, 6, 7, 8, and 9 (groups I,N, W, and P being the first four groups of thisclassification). We also suggest using thesymbol com to refer to incompatibility class;the plasmids belonging to new classes shouldbe called R22 com5 amp str chl tetB sul; R,com5 amp; R,5 com6 amp chl sul genK; R40acom6 amp kanP sul, etc. All plasmids studied
here and their incompatibility class are listedin Fig. 2.Frequency of transfer. The frequency of
transfer (as defined in Materials and Methods)into a plasmid-negative recipient was deter-mined and compared to the frequency oftransfer into a recipient already carrying aplasmid. Results in the cases where the recip-ient already contained a plasmid incompatible
VOL. 112, 1972 671
on March 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

CHABBERT ET AL.
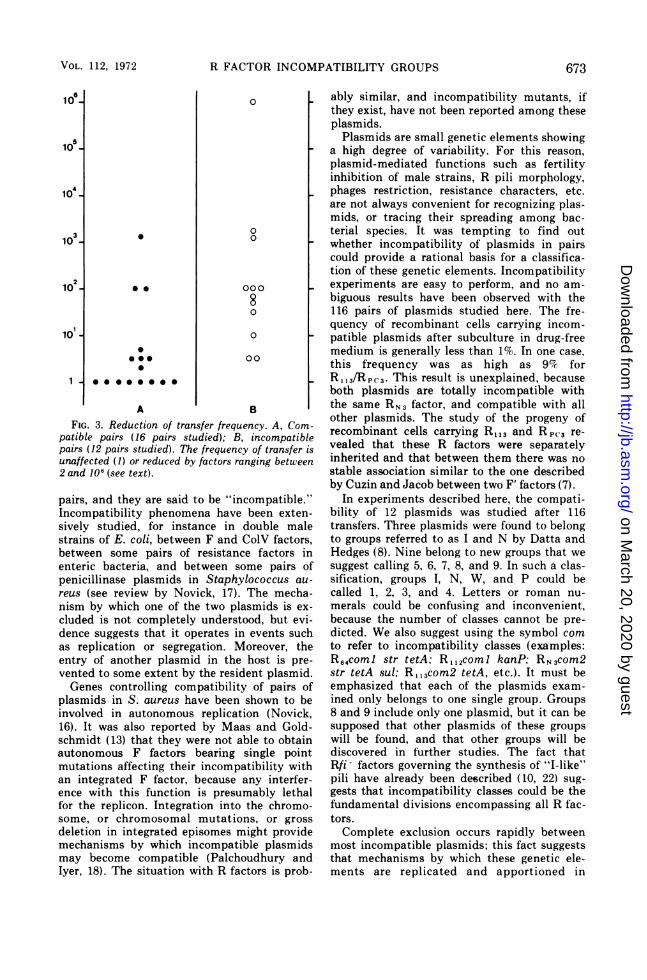
with the entering one can be seen in Tables 3,4, and 5 and have been summarized in Fig. 3.In that case, the frequency of transfer was re-duced by factors ranging between 2 (R.9 1/R69-2)and 1,000,000 (R112/R14), with an average of100 in most transfers.The reduction of the frequency of transfer
when the recipient already carried a plasmidcompatible with the entering one was deter-mined in 16 crosses and can also be seen inFig. 3. There was no reduction in eight crosses;the frequency was reduced by factors rangingbetween 2 and 10 in five crosses, and by fac-tors ranging between 100 and 1,000 in threecrosses.
It can easily be se, n in Fig. 3 that frequen-cies of transfer and their reduction when therecipients are plasmid-positive strains are (i)highly variable, (ii) unpredictable, and (iii) notclearly related to incompatibility. Moreover,the segregation that occurs in the progeny ofrecipients carrying incompatible plasmids cer-tainly causes great changes in the apparentfrequency of transfer.
DISCUSSIONIt is now well known that different types of
plasmids or episomes can coexist in a bacterialcell. However, some plasmids cannot coexist in
com I R 64 + _ _ _ _ _ 11111
R112
la,RN3 + + *|com2 RNH3T t m
R113 ~0 +
RpC3 0-- - -
com 3 RS_S
com4 Rp4
com 5R22(_+ +
Rg 0
com 6 R55 + + + - _ _
R40a ~~~~0 + - - - -
R16 0 - - - -
com 7 R6(a + + _
com8 R111 0 _
com9 R71( 0
FIG. 2. Incompatibility of plasmids in pairs. +, Incompatible pairs; -, compatible pairs; *, studied byDatta and Hedges (8, 11); 0, not done; (a), a derivative listed in Table 2 was used in some crosses.
672 J. BACTERIOL.
on March 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
R FACTOR INCOMPATIBILITY GROUPS
10_
2
10 -
2
10 -
1-
S.
0 00
.0000
0
0
0
000
80
0
00
A B IFIG. 3. Reduction of transfer frequency. A, Com-
patible pairs (16 pairs studied); B, incompatiblepairs (12 pairs studied). The frequency of transfer isunaffected (1) or reduced by factors ranging between2 and 106 (see text).
pairs, and they are said to be "incompatible."Incompatibility phenomena have been exten-sively studied, for instance in double malestrains of E. coli, between F and ColV factors,between some pairs of resistance factors inenteric bacteria, and between some pairs ofpenicillinase plasmids in Staphylococcus au-
reus (see review by Novick, 17). The mecha-nism by which one of the two plasmids is ex-
cluded is not completely understood, but evi-dence suggests that it operates in events suchas replication or segregation. Moreover, theentry of another plasmid in the host is pre-vented to some extent by the resident plasmid.Genes controlling compatibility of pairs of
plasmids in S. aureus have been shown to beinvolved in autonomous replication (Novick,16). It was also reported by Maas and Gold-schmidt (13) that they were not able to obtainautonomous F factors bearing single pointmutations affecting their incompatibility withan integrated F factor, because any interfer-ence with this function is presumably lethalfor the replicon. Integration into the chromo-some, or chromosomal mutations, or grossdeletion in integrated episomes might providemechanisms by which incompatible plasmidsmay become compatible (Palchoudhury andIyer, 18). The situation with R factors is prob-
ably similar, and incompatibility mutants, ifthey exist, have not been reported among theseplasmids.Plasmids are small genetic elements showing
a high degree of variability. For this reason,
plasmid-mediated functions such as fertilityinhibition of male strains, R pili morphology,phages restriction, resistance characters, etc.are not always convenient for recognizing plas-mids, or tracing their spreading among bac-terial species. It was tempting to find outwhether incompatibility of plasmids in pairscould provide a rational basis for a classifica-tion of these genetic elements. Incompatibilityexperiments are easy to perform, and no am-biguous results have been observed with the116 pairs of plasmids studied here. The fre-quency of recombinant cells carrying incom-patible plasmids after subculture in drug-freemedium is generally less than 1%. In one case,this frequency was as high as 9% forRI I3/RPC3. This result is unexplained, becauseboth plasmids are totally incompatible withthe same RN3 factor, and compatible with allother plasmids. The study of the progeny ofrecombinant cells carrying R,13 and Rp3 re-
vealed that these R factors were separatelyinherited and that between them there was no
stable association similar to the one describedby Cuzin and Jacob between two F' factors (7).
In experiments described here, the compati-bility of 12 plasmids was studied after 116transfers. Three plasmids were found to belongto groups referred to as I and N by Datta andHedges (8). Nine belong to new groups that we
suggest calling 5, 6, 7, 8, and 9. In such a clas-sification, groups I, N, W, and P could becalled 1, 2, 3, and 4. Letters or roman nu-
merals could be confusing and inconvenient,because the number of classes cannot be pre-dicted. We also suggest using the symbol comto refer to incompatibility classes (examples:R64coml str tetA; R 12COml kanP; RN3com2str tetA sul; R,13COm2 tetA, etc.). It must beemphasized that each of the plasmids exam-
ined only belongs to one single group. Groups8 and 9 include only one plasmid, but it can besupposed that other plasmids of these groupswill be found, and that other groups will bediscovered in further studies. The fact thatRfi factors governing the synthesis of "I-like"pili have already been described (10, 22) sug-gests that incompatibility classes could be thefundamental divisions encompassing all R fac-tors.Complete exclusion occurs rapidly between
most incompatible plasmids; this fact suggeststhat mechanisms by which these genetic ele-ments are replicated and apportioned in
673VOL. 112, 1972
on March 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

CHABBERT ET AL.
daughter cells are under stringent control. Italso suggests that there is probably one copy or
a few copies of each R factor, and that theyconsist of one replicon with one region similarto the mcr-seg region described by Novick inpenicillinase plasmids (16). It must be pointedout that incompatibility phenomena were ob-served after transduction and transfer of R fac-tors into E. coli K-12, and that the resultscould be different in other bacterial hosts or
with other plasmids.It seems that incompatibility experiments
can provide a reliable method for recognitionand classification of R factors. Such a methodis very much needed in epidemiological stud-ies, where it is sometimes difficult to deter-mine whether newly isolated plasmids are de-rived from already described plasmids or not.It is possible to draw some conclusions of epi-demiological interest from results reportedhere. There are many incompatibility groups
among Rfi- factors. The same group can in-clude plasmids present in totally different spe-
cies. For instance, group 5 includes R22 iso-lated from Shigella flexneri (Strasbourg, 1966),and R9 isolated from Salmonella paratyphi B(Tours, 1965); group 6 includes R55 isolatedfrom Klebsiella pneumoniae (Paris, Claude-Bernard Hospital, 1969), R40a isolated fromPseudomonas aeruginosa (Paris, Necker Hospi-tal, 1969), and R,6 isolated from Providenciastuartii (Paris, Saint-Antoine Hospital, 1966),etc. R factors from the same group may havetotally different resistance markers, such as
R64coml str tetA, and Rl,,coml kanP, or
RN,3com2 str tetA sul, R,13com2 tetA, andR pc3com2 str kanP.The classification proposed here confirms
the coexistence of three factors in Salmonellapanama described by Bouanchaud and Chab-bert (4). There three compatible plasmids be-long to three different groups and are
R1lcom8 amp, Rll 2com1 kanP, and R,,3com2tetA. Other S. panama strains carrying similarR factors have been isolated in Creteil Hos-pital (Pietkiewitch et al., 19). A Salmonellaparatyphi B strain and a K. pneumoniaestrains resistant to the same drugs (ampicillin,kanamycin, and tetracycline) were also iso-lated from one patient in the same hospital. S.paratyphi B carried R69com7 amp kanP tetBand K. pneumoniae carried R95com7 amp
kanP tetB. Such a result leads to the conclu-sion that no transfer of drug resistance factorsoccurred in vivo from S. panama (R,,,) (R112)(R1,3) into S. paratyphi (R69) or K. pneumo-niae (R9,). On the contrary, such a transfermay have occurred between S. paratyphi B and
K. pneumoniae: these strains were isolatedfrom the same sample, their R factors belongto the same class (com7) and mediate resist-ance to the same drugs (ampicillin, kana-mycin-paromomycin, tetracycline-minocyc-line). For all the above-mentioned reasons, itseems desirable to continue studying incom-patibility phenomena between R factors as abasis for their recognition and classification.
ACKNOWLEDGMENTSWe are indebted to G. Bieth and C. Carlier for their ex-
cellent technical assistance.
LITERATURE CITED
1. Acar, J. F., J. G. Baudens, and Y. A. Chabbert. 1970.Clinical and laboratory evaluation of cephalexin inurinary-tract infections. Antimicrob. Ag. Chemother.1969, p. 398-404.
2. Bannister, D. 1970. Analysis of an R+ strain carryingtwo fi - sex factors. J. Gen. Microbiol. 61:273-281.
3. Baudens, J. G., and Y. A. Chabbert. 1967. Analyse desfacteurs de resistance transferables isoles en France.Ann. Inst. Pasteur 112:565-580.
4. Bouanchaud, D. H., and Y. A. Chabbert. 1969. Stablecoexistence of three fi - resistance factors in S.panama and E. coli. J. Gen. Microbiol. 58:107-113.
5. Bouanchaud, D. H., M. R. Scavizzi, and Y. A. Chabbert.1968. Elimination by ethidium bromide of antibioticresistance in enterobacteria and staphylococci. J. Gen.Microbiol. 54:417-425.
6. Chabbert, Y. A., and J. C. Patte. 1960. Cellophantransfer: application to the study of activity of combi-nations of antibiotics. Appl. Microbiol. 8:193-197.
7. Cuzin, F., and F. Jacob. 1967. Association stable dedeux 6pisomes F diff6rents dans un clone d'-Escherichia coli. Ann. Inst. Pasteur 113:145-155.
8. Datta, N., and R. W. Hedges. 1971. Compatibilitygroups among fi - R factors. Nature (London) 234:222.
9. Egawa, R., and Y. Hirota. 1962. Inhibition of fertility bymultiple drug-resistance factor (R) in E. coli K12.Jap. J. Genet. 37:66-69.
10. Grindley, J. N., and E. S. Anderson. 1971. I-like resist-ance factors with the fi character. Genet. Res. 17:267-271.
11. Hedges, R. W., and N. Datta. 1971. fi- R factors givingchloramphenicol resistance. Nature (London) 234:220-221.
12. Khatoon, H., and R. V. Iver. 1971. Stable coexistence offi - factors in E. coli. Can. J. Microbiol. 17:669-675.
13. Maas, W. K., and A. D. Goldschmidt. 1969. A mutant ofEscherichia coli permitting replication of two F fac-tors. Proc. Nat. Acad. Sci. U.S.A. 62:873-880.
14. MacFarren, A. C., and R. C. Clowes. 1967. A compara-tive study of two F-like colicin factors ColV2 andColV3 in Escherichia coli K-12. J. Bacteriol. 94:365-377.
15. Meynell, E., G. G. Meynell, and M. Datta. 1968. Phylo-genetic relationships of drug-resistance factors andother transmissible bacterial plasmids. Bacteriol. Rev.32:55-83.
16. Novick, R. P. 1967. Penicillinase plasmids of Staphylo-coccus aureus. Fed. Proc. 26:29-38.
17. Novick, R. P. 1969. Extrachromosomal inheritance inbacteria. Bacteriol. Rev. 33:210-235.
18. Palchoudhury, S. R., and U. N. Iyer. 1971. Compati-bility between two F' factors in an E. coli strainbearing a chromosomal mutation affecting DNA syn-thesis. J. Mol. Biol. 57:319-333.
J. BACTERIOL.674
on March 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

R FACTOR INCQMPATIBILITY GROUPS
19. Peyru, G., L. Wexler, and R. P. Novick. 1969. Naturallyoccuring penicillinase plasmids in Staphylococcusaureus. J. Bacteriol. 98:215-221.
20. Pietkiewitch, K., Y. Hamon, L. Le Minor, and Y. A.Chabbert. 1969. Correlation entre les bacteriocines etles facteurs R de Salmonella panama. Ann. Inst. Pas-teur 117:645-655.
21. Romero, E. 1970. Two classes of fi- R factors. Boll. Ist.Sieroter. Milan. 49:484-487.
22. Romero, E., and E. Meynell. 1969. Covert fi- R factorsin fi+ R+ strains of bacteria. J. Bacteriol. 97:780-786.
23. Scavizzi, M. R. 1972. Existence de deux caracteres deresistance aux tetracyclines. Ann. Inst. Pasteur 122:3-17.
24. Scavizzi, M. R., and Y. A. Chabbert. 1970. Etude de latransferabilite des facteurs de resistance heberges par
675
les Colibacilles enteropathogenes lors des epidemiesde creches. Pathol. Biol. 18:1079-1087.
25. Watanabe, T., and T. Fukasawa. 1961. Episome-medi-ated transfer of drug resistance in Enterobacteriaceae.I. Transfer of resistance factors by conjugation. J.Bacteriol. 81:669-678.
26. Watanabe, T., H. Nishida, C. Ogata, T. Arai, and S.Sato. 1964. Two types of naturally occuring R factors.J. Bacteriol. 88:716-726.
27. Witchitz, J. L., and Y. A. Chabbert. 1971. High leveltransferable resistance to gentamicin. J. Antibiot.(Tokyo) 24:137-139.
28. Witchitz, J. L., and Y. A. Chabbert. 1971. Rasistancetransferable a la gentamicine. I. Expression ducaractere de resistance. Ann. Inst. Pasteur 121:733-742.
VOL. 112, 1972
on March 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from