in vitro effect of recombinant interferon gamma in combination with lps on amoebicidal activity of...
TRANSCRIPT

Immunobiol., vol. 188, pp. 203-219 (1993)
Department of Biochemistry and Microbiology, University of Victoria, Victoria, British Columbia, Canada
In vitro Effect of Recombinant Interferon Gamma in Combination with LPS on Amoebicidal Activity of Murine Kupffer Cells
ESFANDIAR GHADIRIAN and AFsAR SALIMI
Received June 11, 1992 . Accepted in Revised Form December 15, 1992
Abstract
The present study examines the role of liver macrophages (Kupffer cells), of C57BLl6 mice, as effector cells responsible for the killing of Entamoeba histolytica trophozoites in vitro. It was shown that unstimulated Kupffer cells were inefficient in the killing of E. histolytica trophozoites in vitro. Interferon gamma (IFN-y) alone was not able to activate Kupffer cells to amoebicidal state. However, Interferon gamma and lipopolysaccharide (LPS) acted synergistically in this phenomenon. It seems that the acquisition of amoebicidal activity is associated with the involvement of hydrogen peroxide, because the addition of catalase partially decreases the killing of this parasite by Kupffer cells. In addition, it appears that the amoebicidal activity of IFN -y-treated Kupffer cells is contact-dependent.
Our results indicate that the immunologic production of IFN-y is important in the activation of Kupffer cells for controlling this parasite and that Kupffer cells are strong effector cells against the amoebae.
Introduction
Amoebiasis is a major health problem in Third World countries. Entamoeba bistolytica is present worldwide, but is most frequently encountered in tropical climates in association with poor sanitary living conditions. It is the third most common cause of death in areas where parasitic diseases are endemic, after malaria and schistosomiasis (1). The mechanisms of the host defense against this parasite have been investigated by various laboratories, but are still not well understood (2-5). E. bistolytica induces both humoral and cellular immune response in humans and animals (6). Although high titers of anti-amoebic antibody develop over the course of invasive disease, its presence is not associated with resistance to infection (7, 8). By excluding the humoral response for acquisition of effective immunity to invasive disease, investigators from various laboratories have looked at the effector cells responsible for protective immune mechanisms against this parasite. In a series of investigations in hamsters and in mice, it was found that depression of cell-mediated immunity such as splenectomy (9), immunosuppression (10), treatment of

204 . E. GHADIRIAN and A. SALIMI
animals with silica (11, 12), or with anti-macrophage serum (13) reduced host resistance to intestinal and extraintestinal amoebiasis. In contrast, treatment of the animals with macrophage activating factors (BeG, Corynebacterium parvum, or Listeria monocytogens) had a protective effect (11,14). SALATA and his co-workers (2) demonstrated that monocytederived macrophages from patients suffering from amoebiasis and treated with lymphokines were able to kill amoebic trophozoites in vitro. In another study, GHADIRIAN and BOUT (5) found that peritoneal macrophages treated with both IFN-y and LPS were able to kill E. histolytica trophozoites in vitro. They also demonstrated that in vitro macrophage amoebicidal activity was due to both oxygen-dependent and-independent mechanisms. Because of the importance of the liver in the formation of amoebic liver abscesses (15), it is of interest to study the role of the hepatic macrophages or Kupffer cells in amoebiasis. The role of Kupffer cells in host defense against tumor growth, microbial and parasitic infections has been investigated by many researchers (16-19). However, their role against amoebiasis has not yet been determined.
The purpose of the present study was to characterize the in vitro interactions between virulent axenic E. histolytica trophozoites and Kupffer cells, and to study the role of recombinant IFN-y in the in vitro generation of Kupffer cells capable of killing of this parasite. This study demonstrates that Kupffer cells, treated with recombinant IFN-y and LPS in vitro, are effective in killing amoebic trophozoites. Activated Kupffer cells appear to kill amoebae through a contact and partly oxidativedependent mechanism.
Materials and Methods
Mice
CS7BL!6 male mice were purchased (Charles River, St.-Constant, Quebec, Canada) and used at 6-7 weeks of age in all experiments.
Parasite
Strain IP:0682: 1
The IP:0682: 1 strain of E. histolytica used in all experiments was isolated by Dr. MEEROVITCH in 1982 from the dysentric stool of a l-year-old Amerindian child in the amoebiasis endemic area of Saskatchewan, Canada (20). The initial isolate was established, and thereafter grown, in Robinson's medium (21) for 30 3-day passages before axenization in TYIS-33 medium. This strain is highly virulent in the gerbil (Meriones unguculatus) (20).
Interferon gamma
Recombinant murine interferon gamma, with a specific activity of 6.8 x 107 units per mg protein, was generously supplied by Dr. H. M. SHEPARD (Genentech Inc., South SanFrancisco, CA, USA). The approach to IFN-y production is basically that which has been reported for the production of cloned human IFN-y (22).

Effect of IFN-y on Amoebae' 205
Monoclonal anti-TNF antibody
Monoclonal antibody against TNF was provided by Dr. M. BROKHAUS (F. Hoffmann-La Roche, Basel, Switzerland). The liquid form was diluted in PBS solution to the required dosage.
Preparation of E. histolytica trophozoites for in vitro study
The amoebae were grown in axenic TYI-S-33 medium in 50 ml screw-capped flasks (Corning, NY, USA) and incubated at 37°C. The culture media was transferred three times weekly. Three-day-old axenic culture containing trophozoites were used in the experiments. The cultures were ice-chilled, transferred into 50 ml disposable plastic centrifuge tubes (Corning), and centrifuged at 1500 r.p.m. for 10 min. the supernatant was decanted, and the viability of amoebae was assessed by phase-contrast microscopy and by trypan blue staining as described previously (2, 5). In all experiments, the amoebic trophozoites viability was consistently between 95 to 97 %. The amoebae were diluted in tissue culture medium 199 (Gibco laboratories, Grand Island, NY, USA) supplemented with 5 mM L-cystein, 10 % heatinactivated fetal bovine serum (Gibco) and 25 mM Hepes (Gibco) at pH 6.8.
Preparation of Kupffer cells
Kupffer cells were obtained using modified protocols of DIESSELHOFF-DEN DULK et a!. (23) and LEPAY et a!. (24,25). Briefly, mice were sacrificed with CO2 and the abdomen opened. The liver was perfused with a butterfly needle (Abbott laboratories, USA), through the portal vein using Hanks' Balanced Salt Solution (HBSS) (Gibco) containing 0.05 % collagenase (Sigma Chemical Co., St. Louis, MO, USA), 5 % fetal bovine serum (Gibco), 100 U of penicillin per ml at flow rate of 3-4 ml!min and at 3rC. Once perfusion was established, the inferior vena cava was cut, following which the liver became pale in color. The liver was then removed, separated into pieces, and the fragments were digested for 45 min under constant stirring in HBSS containing 0.05 % collagenase at 37°C. After destruction of parenchymal cells by enzymes, the cell suspension was filtered through a stainless steel mesh to remove indigestible materials, and centrifuged once for 10 min at 1500 r.p.m. within Dulbecco's Modified Eagle Medium (DMEM) (Gibco). The remaining red blood cells were lysed with hemolytic Gey's solution (26). The cells were diluted in DMEM and centrifuged three times for 10 min at 1500 r.p.m. with Ca- and Mg-free HBSS. This procedure resulted in removal of parenchymal cells (24). At this stage, cytocentrifuge (Cytospin centrifuge, Shandon Southern Instruments, Camberly, England) preparations of the suspension were prepared for morphological studies. The slides preparations were dried rapidly in air, and stained with Wright stain (27). After three washes, the total number of nonparenchymal cells were counted with a hemacytometer. The average yield of nonparenchymal cells obtained by collagenase digestion was approximately 3.8 X 107 cells/mouse liver. The number of viable nonparenchymal cells was determined by staining cell populations with trypan blue (16, 24, 28). The cell viability was consistantly between 90 to 95 %. Greater than 95 % of adherent cells were able to phagocytose the latex beads particles and therefore characterized as Kupffer cells. Each Kupffer cell phagocytoses an average of 10-15 latex bead particles (0.8 flm diameter; Sigma). On the basis of morphology and phagocytosis of latex beads particles, 23-26 % of nonparenchymal cells were Kupffer cells. These results were consistant with those of CROFTON et a!. (29). Kupffer cells plated at desired density according to each experiment.
Cell culture
After counting, the nonparenchymal cells were cultured in 24-well culture plate at 4 x 107
cells/ml!wel!. The culture medium consisted of DMEM supplemented with 10 % heatinactivated fetal calf serum, 5 % antibiotic (streptomycin), and was incubated at least 2 h at 37°C in 5 % COb until all Kupffer cells were tightly adherent. Repeated washing of the wells removed all non-adherent cells. The total volume of media in each well was 1 m!. Adherent cells were incubated for 24 b in the same media.

206 . E. GHADIRIAN and A. SALIMI
Treatment of Kupffer cells with IFN-y and LPS
Kupffer cell monolayers incubated overnight in 24-well plates were washed three times with HBSS, and then they were preincubated with IFN-y (500 D/ml) andlor LPS (20 ng/ml) for various periods of time as indicated in the text. IFN-y and LPS used in this study were appropriately diluted with DMEM, as indicated in the text.
Cytotoxicity assay
The assay for in vitro amoebicidal activity of Kupffer cells was carried out as described previously (5). Briefly, E. histolytica trophozoites were suspended in medium 199 containing 5.7 mM L-cystein (Gibco), 10 % heat-inactivated fetal bovine serum and 25 mM Hepes at pH 6.8. Amoebic suspensions containing 5 x 105 trophozoite simi, were added to Kupffer cells monolayers. Initial studies were carried out at a Kupffer cell to parasite ratio of 20: 1. The total volume of each well was 1 m!. The plates were incubated at 3rC in 5 % CO2 atmosphere for 4 h, the standard assay time. The assay was done in triplicate. Control cultures were treated with medium without IFN-y or without LPS. Experiments were repeated three to four times, with results representing the average of three to four observations.
The effect of catalase on amoebicidal activity of Kupffer cells
Kupffer cells were prepared according to the procedure described above. Kupffer cells, preincubated for 24 h with both IFN-y and LPS, were incubated for 30 min with 4000 D/ml of catalase (from bovine liver, Sigma), following which the desired concentrations of amoebic trophozoites were added to the wells for 4 h at 3rC (5). The viability of trophozoites was determined as described previously. The experiments were repeated two to three times.
Role of monoclonal anti-TNF antibody on TNF-mediated killing of E. histolytica trophozoites
Kupffer cells were harvested and incubated with IFN-y (500 D/ml) and LPS (20 ng/ml) for 24 h. After incubation time, the cells were washed once with HBSS, 500 units of anti-TNF antibody and amoebic trophozoites were added to the cells simultaneously. After 4 h of incubation, the viability of amoebae was determined by trypan blue. Experiments were done in triplicate culture and repeated 4 times.
Statistical analysis
Student's t-test was used to analyze the data. A probability level of P < 0.05 was considered significant for all experiments.
Results
In vitro amoebicidal activity of Kupffer cells treated with either IFN-y or LPS
In the first set of experiments, the ability of IFN-y to activate Kupffer cells for amoebicidal activity without a second signal was tested. In this experiment, Kupffer cells were treated with 6 different concentrations of IFN-y (from 50 to 1000 Vlml) for 24 h, without the addition of LPS. The results demonstrate that IFN -y alone does not have the capacity to activate Kupffer cells for amoebicidal activity (Table 1). Although the treatment of Kupffer cells with 750 and 1000 units of IFN-y showed slight increase in the number of dead trophozoites, this was not significant as compared to the untreated group.

Effect of IFN-y on Amoebae· 207
Table 1. In vitro amoebicidal activity of Kupffer cells treated either with LPS or IFN-y"
LPS treatment % Dead trophozoites IFN -y treatment % Dead trophozoites concentrations of E. histolytica concentrations 01 E. histolytica (ng/ml) (Mean± SD) (U/ml) (Mean± SD)
5 20.7 ± 5.8 50 27 ± 5.8 10 20± 6 250 26± 7 20 26 ± 5.3 500 29 ± 5 40 28 ± 5.5 750 33 ± 7.1 80 27.3 ± 4.2 1000 31 ± 6.5 untreated 25.3 ± 6.1 untreated 23.3 ± 5.7
". The data are represented as the mean of 4 observations ± SD.
Kupffer cells function for amoebicidal activity in the absence of IFN-y was also examined. In this experiment, Kupffer cells were incubated with different concentrations of LPS (from 5 to 80 ng/ml) for 24 h. These results showed that different concentrations of LPS alone did not induce Kupffer cells to become cytotoxic against E. histolytica trophozoites as compared to control group (Table 1).
In vitro amoebicidal activity Kupffer cells treated with both IFN-y and LPS
In the next experiment, the ability of macrophages to be activated by IFN -y and LPS for amoebic killing was investigated. Kupffer cells were treated with various concentrations of IFN-y (from 50 to 1000U/ml) in the presence of a constant concentration of LPS (20 ng/ml).
The results presented in Figure lA demonstrate the in vitro amoebicidal activity of Kupffer cells treated with six different concentrations of IFN-y. In the presence of a constant amount of LPS, there was an inverse doseresponse relationship between the concentration of IFN-y used and the viability of E. histolytica trophozoites. There was no significant cytotoxic effect on amoebae when Kupffer cells were treated with 50, 125 or 250 units of IFN-y in the presence of 20 ng LPS. In contrast, when the larger concentrations of 500, 750 or 1000 units of IFN-y were used, the cytotoxicity of Kupffer cells was highly significant (P < 0.001) as compared to those groups treated with lower concentrations of IFN-y. The results of this experiment suggest that IFN-y concentrations of less than 500 U/ml are not sufficient to trigger Kupffer cells for killing E. histolytica trophozoites. Concentrations of IFN-y of 750 and 1000 U/ml, respectively, did not increase the cytotoxicity of Kupffer cells against E. histolytica trophozoites as compared to those treated with 500 units of IFN-y. Therefore, for the next set of experiments, 500 units of IFN -y were used.
The amoebicidal activity of Kupffer cells was also examined with various concentrations of LPS in the presence of fixed concentration of IFN -y. In this study, Kupffer cells were treated with 6 different concentrations of LPS (5, 10, 15, 20, 40, and 80 ng/ml) in the presence of a constant amount of
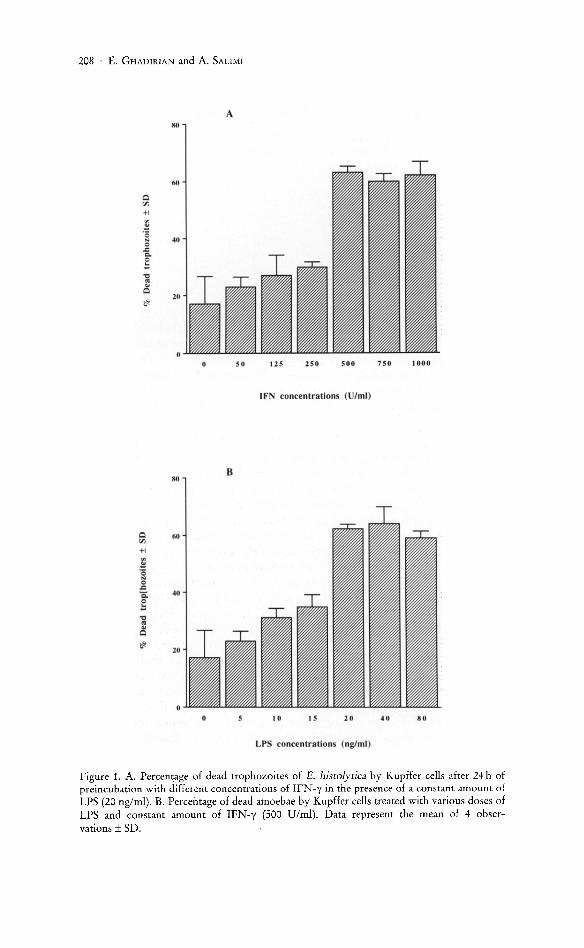
208 . E . GHADIRIAN and A. SALIMI
A Mil
1141
Q 'Jl
+ • ., ~ '0 N 411 0 &: Co
E .., " " Q
211 c"
50 125 250 500 750 1000
I F concent rations (U/ml)
B 80
Q
'" 60
+I
~ '0 ... 0 &: c;: 40
E .., .. .. Q
t'< 20
10 15 20 411 KO
LPS concentrations ( nglml )
Figure 1. A. Percentage of dead trophozoites of E. histolytica by Kupffer cells after 24 h of preincubation w ith different concentrations of IFN -y in the presence of a constant a mount of LPS (20 n g/ml). B. Percentage of dead amoebae by Kupffer cells treated with various doses of LPS and constant amount of IFN-y (500 Vlml). Data represent the mean of 4 observations ± SD.
90
80
70 Q r/l
+1 60 til
~ ·0 N Q SO -= =-'" ... 1 40
~ ~ 30
20
10
i 1: 1
Effect of IFN-y on Amoebae' 209
o NOII·treated Kupffer ceUs
• 500 VIm! IFN & 20 ng LPS
1: 10 1 : 20
Ratio of Kupffer cells to amoebae
1 : 40
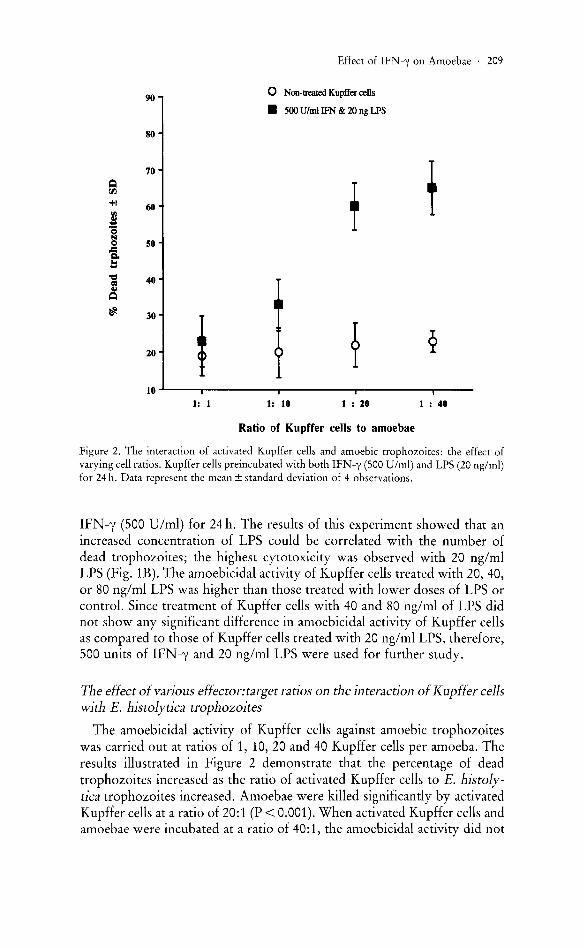
Figure 2. The interaction of activated Kupffer cells and amoebic trophozoites: the effect of varying cell ratios. Kupffer cells preincubated with both IFN-y (500 U/ml) and LPS (20 ng/ml) for 24 h. Data represent the mean ± standard deviation of 4 observations.
IFN-y (500 U/ml) for 24 h. The results of this experiment showed that an increased concentration of LPS could be correlated with the number of dead trophozoites; the highest cytotoxicity was observed with 20 ng/ml LPS (Fig. lB). The amoebicidal activity of Kupffer cells treated with 20,40, or 80 ng/ml LPS was higher than those treated with lower doses of LPS or controL Since treatment of Kupffer cells with 40 and 80 ng/ml of LPS did not show any significant difference in amoebicidal activity of Kupffer cells as compared to those of Kupffer cells treated with 20 ng/ml LPS, therefore, 500 units of IFN-y and 20 ng/ml LPS were used for further study.
The effect of various effector: target ratios on the interaction of Kupffer cells with E. histolytica trophozoites
The amoebicidal activity of Kupffer cells against amoebic trophozoites was carried out at ratios of 1, 10, 20 and 40 Kupffer cells per amoeba. The results illustrated in Figure 2 demonstrate that the percentage of dead trophozoites increased as the ratio of activated Kupffer cells to E. histolytica trophozoites increased. Amoebae were killed significantly by activated Kupffer cells at a ratio of 20:1 (P < 0.001). When activated Kupffer cells and amoebae were incubated at a ratio of 40:1, the amoebicidal activity did not
210 . E. GHADIRIAN and A. SALIMI
80 --0-- Control
-+- 500 U/ml IFN & 20 ng/ml LPS
60
Q rn +I
! ·S .. = ..c: 40 c. = !::
~ ~
20
o+-------~------._------._------r_----__,
o 6 12 18 30
Hours of pre-Incubation with IFN and LPS
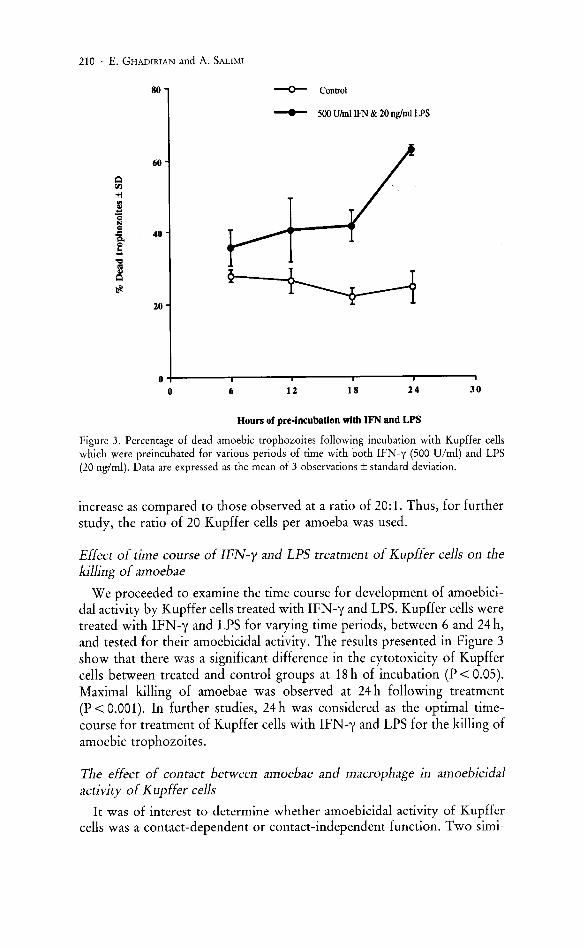
Figure 3. Percentage of dead amoebic trophozoites following incubation with Kupffer cells which were preincubated for various periods of time with both IFN-y (500 Vlml) and LPS (20 nglml). Data are expressed as the mean of 3 observations ± standard deviation.
increase as compared to those observed at a ratio of 20:1. Thus, for further study, the ratio of 20 Kupffer cells per amoeba was used.
Effect of time course of IFN-y and LPS treatment of Kupffer cells on the kl1ling of amoebae
We proceeded to examine the time course for development of amoebicidal activity by Kupffer cells treated with IFN-y and LPS. Kupffer cells were treated with IFN -y and LPS for varying time periods, between 6 and 24 h, and tested for their amoebicidal activity. The results presented in Figure 3 show that there was a significant difference in the cytotoxicity of Kupffer cells between treated and control groups at 18 h of incubation (P < 0.05). Maximal killing of amoebae was observed at 24 h following treatment (P < 0.001). In further studies, 24 h was considered as the optimal timecourse for treatment of Kupffer cells with IFN-y and LPS for the killing of amoebic trophozoites.
The effect of contact between amoebae and macrophage in amoebicidal activity of Kupffer cells
It was of interest to determine whether amoebicidal activity of Kupffer cells was a contact-dependent or contact-independent function. Two simi-
Effect of IFN-y on Amoebae' 211
lar procedures were performed. In the first procedure, after incubation of Kupffer cells with both IFN-y and LPS for 24 h, the supernatant was removed and placed in separate wells E. histolytica trophozoites were added to wells containing both cell-free supernatant and Kupffer cells. After 4 h of incubation the viability of amoebae was determined by trypan blue exclusion. Control wells were treated identically except IFN-y and
LPS were not added. In the second procedure, after incubation of Kupffer cells with both
IFN-y and LPS for 24 h, the supernatant was decanted and fresh medium was added to the cells. Kupffer cells were incubated for 4 h. Following this incubation period, the supernatant was removed and aliquoted into separate wells and amoebic trophozoites were added to wells containing either the cell-free supernatants or Kupffer cells. After 4 h of incubation, the viability of amoebae was determined by trypan blue exclusion.
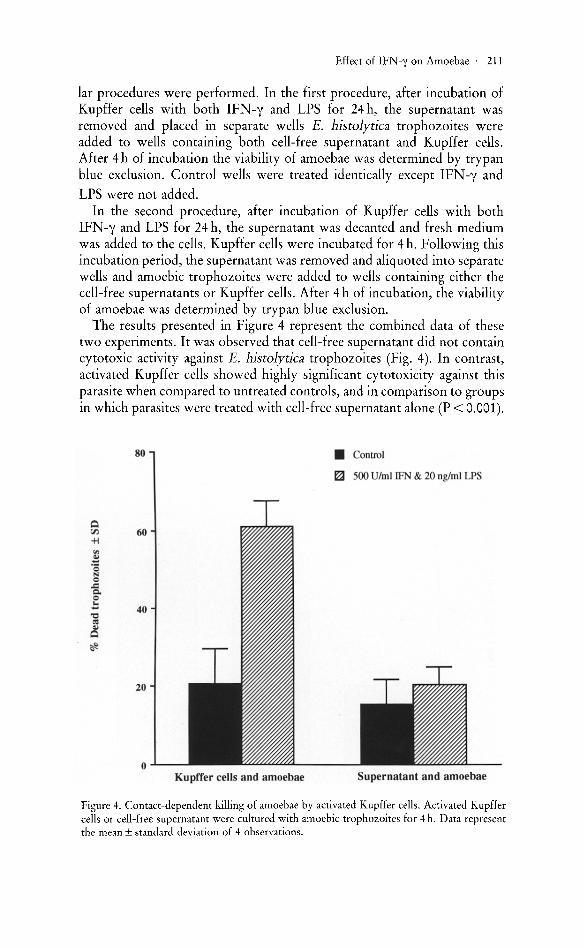
The results presented in Figure 4 represent the combined data of these two experiments. It was observed that cell-free supernatant did not contain cytotoxic activity against E. histolytica trophozoites (Fig. 4). In contrast, activated Kupffer cells showed highly significant cytotoxicity against this parasite when compared to untreated controls, and in comparison to groups in which parasites were treated with cell-free supernatant alone (P < 0.001) .
80 • Control
~ 500 Ulml IFN & 20 ng/ml LPS
Q rJ) 60 +i II>
~ '0 .... 0 .::: c. 0 !: ~
40 <IS OJ
Q
~
20
o Kupffer cells and amoebae Supernatant and amoebae
Figure 4. Contact-dependent killing of amoebae by activated Kupffer cells. Activated Kupffer cells or cell-free supernatant were cultured with amoebic trophozoites for 4 h. Data represent the mean ± standard deviation of 4 observations.
212 . E. GHADIRIAN and A. SALIMI
We concluded that IFN-y and LPS simultaneously allowed the murine Kupffer cells to become cytotoxic against amoebic trophozoites. Our data strongly suggests that contact between Kupffer cells and E. histolytica trophozoites was required for this killing.
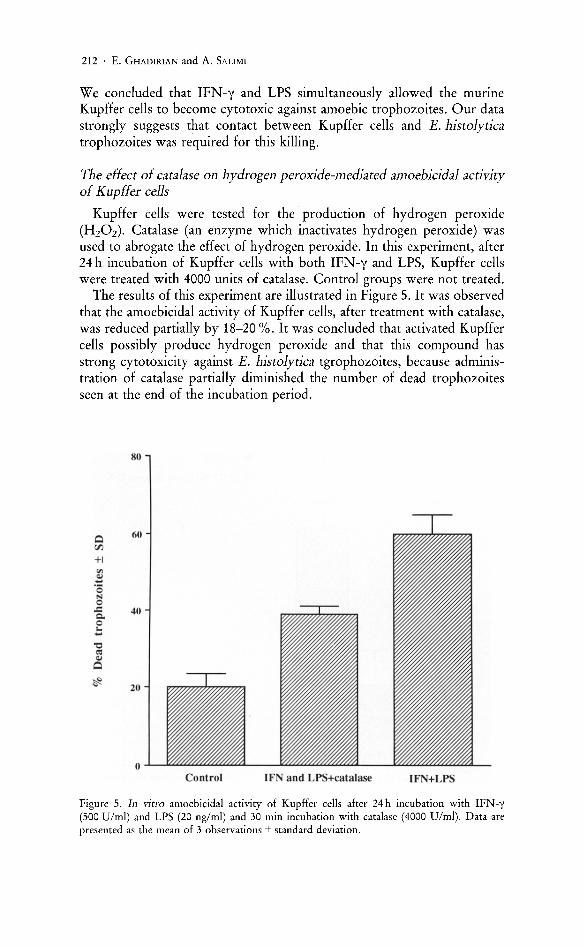
The effect of catalase on hydrogen peroxide-mediated amoebicidal activity of Kupffer cells
Kupffer cells were tested for the production of hydrogen peroxide (HzOz). Catalase (an enzyme which inactivates hydrogen peroxide) was used to abrogate the effect of hydrogen peroxide. In this experiment, after 24 h incubation of Kupffer cells with both IFN-y and LPS, Kupffer cells were treated with 4000 units of catalase. Control groups were not treated.
The results of this experiment are illustrated in Figure 5. It was observed that the amoebicidal activity of Kupffer cells, after treatment with catalase, was reduced partially by 18-20 %. It was concluded that activated Kupffer cells possibly produce hydrogen peroxide and that this compound has strong cytotoxicity against E. histolytica tgrophozoites, because administration of catalase partially diminished the number of dead trophozoites seen at the end of the incubation period.
Q 60 00
+1
'" ~ '0
p,J 0 ..c 40 Co 0 ... -'0 eo: ~
Q
<f- 20
Control IFN and LPS+catalase IFN+LPS
Figure 5. In vitro amoebicidal activity of Kupffer cells after 24 h incubation with IFN-y (500 Vlml) and LPS (20 ng/ml) and 30 min incubation with catalase (4000 Vlml). Data are presented as the mean of 3 observations ± standard deviation.

Effect of IFN-y on Amoebae' 213
The effect of monoclonal anti-TNF antibody on amoebicidal activity of Kupffer cells
The involvement of TNF in the killing of amoebic trophozoites by activated Kupffer cells was investigated by treating the Kupffer cell-parasite cultures with monoclonal anti-TNF antibody.
Following treatment of Kupffer cells with IFN -y and LPS for 24 h, the supernatant was discarded. One group of activated Kupffer cells received 500 units of monoclonal anti-tumor necrosis factor (anti-TNF) antibody. The second group remained untreated. Kupffer cells and amoebae were incubated for 4 h, and the number of dead trophozoites was determined by trypan blue exclusion. The results of this experiments show that treatment of Kupffer cells with anti-TNF antibody partially reduced the cytotoxic activity of activated Kupffer cells (Fig. 6). Therefore, it was concluded that TNF produced by activated Kupffer cells had a strong effect on the ameobicidal activity of Kupffer cells, and this was reflected by the reduction in the number of dead trophozoites following the addition of anti-TNF antibody to cultures containing activated Kupffer cells.
70
60
Q rJ)
SO +I III
~ '0 40 N 0 ~ 0-0 !: "0 30 :Il ~ Q
~ 20
10
0
Figure 6. The effect of monoclonal anti-TNF antibody on the amoebicidal activity of activated Kupffer cells. After 24 h of incubation with IFN-y (500 Vim I) and LPS (20 ng/ml), Kupffer cells were treated with anti-TNF (500 Vlml) antibody. Data are expressed as the mean of 4 observations ± standard deviation.

214 . E. GHADIRIAN and A. SALIMI
Discussion
It has been established that murine peritoneal macrophages (5), and bone marrow-derived macrophages (30) acquire amoebicidal activity following cytokine treatment. The role of Kupffer cells in the destruction of amoebic trophozoites has not yet been investigated.
The purpose of the present study was to investigate the role of IFN-y, a cytokine with known macrophage activating potential, in the amoebicidal activity of Kupffer cells.
Kupffer cells are the largest family of the so-called tissue macrophages. These cells have a remarkable capacity to ingest exogenous particles including bacteria, and therefore play a major role in host defense mechanisms (31, 32). CROCKER and his co-workers (28) have demonstrated that liver macrophages are important in host defense in a murine model of infection with Leishmania donovani. In another study (16), it was shown that resident murine Kupffer cells performed neither spontaneous extracellular lysis of tumor cells, nor killing of intracellular parasitic Leishmania microorganisms; however, these cells were readily activated with lymphokines and recombinant IFN-y. Such activated Kupffer cells were able to efficiently kill tumor cells and intracellular Leishmania spp. parasites. Later, FILICE (33) found that recombinant IFN -y increased or at least maintained the microbicidal activity of Kupffer cells. These observations emphasized the role of lymphokines, particularly IFN-y, on the activation of Kupffer cells in host defense mechanisms against tumors and parasites.
Since liver plays an important role in hepatic amoebiasis, it was of interest to study the role of Kupffer cells in the killing of E. histolytica trophozoites m vltro.
The results presented in this study demonstrate that Kupffer cells activated with both IFN -y and LPS were able to kill amoebic trophozoites in vitro. Our results further show that high concentrations of either IFN-y or LPS alone were insufficient to induce activation of murine Kupffer cells to kill E. histolytica trophozoites in vitro (Table 1). Although Kupffer cells treated with 500,750 and 1000 units of IFN-y alone had a greater cytotoxic activity against amoebic trophozoites, the killing was not statistically higher than those untreated Kupffer cells. DECKER et al. (18) have also shown that activation of the murine Kupffer cells was obtained only by a combination of IFN-y and LPS. However, in some systems IFN-y alone could induce Kupffer cells to cytotoxicity against all strains of Leishmania (16) and could maintain (33) the ability of Kupffer cells to partially inhibit T. gondii multiplication. Another study has shown that LPS alone activates Kupffer cells to increase tumor cytotoxic activities (34).
A close correlation was found to exist between the concentrations of IFN-y used to activate Kupffer cells and the level of amoebicidal activity observed (Fig. 1A). A concentration of 500 D/ml IFN-y induced maximal (55-65 %) killing of amoebae when the concentration of LPS was hold constant at 20 ng/ml. Values of less than 500 D/ml induced an insignificant

Effect of IFN-y on Amoebae· 215
level of cytotoxicity when compared to those achieved by untreated Kupffer cells. Concentrations higher than 500 units of IFN-y induced similar levels of cytotoxicity as compared to Kupffer cells treated with 500 U/ml of IFN-y.
A close association has also been noted between the does of LPS used in this experiment and the percentage of dead trophozoites (Fig. lB). It was found that LPS concentrations of less than 20 ng/ml were ineffective in triggering Kupffer cells for cytotoxic activity against E. histolytica trophozoites.
We conclude from these data (Table 1, Fig 1A and Fig. lB) that activation of Kupffer cells for amoebicidal activity requires two signals: IFN-y and LPS confirming the findings of GHADIRIAN and BOUT (5) with peritoneal macrophages. These studies conclusively demonstrate that IFN -y and LPS can induce Kupffer cells activation for E. histolytica killing.
The ability of IFN-y and LPS to activate Kupffer cells and develop cytotoxicity was time-dependent. The peak response developed following a 24 h incubation period with the activating factors (Fig. 3). Similar kinetics have also been reported for the time course of treatment of peritoneal macrophages with IFN-y and LPS (5).
Activated macrophages have previously been reported to kill target cells by contact-dependent mechanisms (36). Our results suggest that contact between amoebic trophozoites and Kupffer cells were required for maximal killing (Fig. 4). In this study, it was demonstrated that Kupffer cells showed highly significant amoebicidal activity as compared to groups of amoebae treated with supernatant alone. Other authors (2, 30) have also shown that contact between activated macrophages and amoebae is necessary to initiate the killing of this parasite.
Our studies indicate that both oxidative-dependent and -independent mechanisms are important for the killing of amoebae by the activated Kupffer cells. Although some investigators have reported that Kupffer cells are incapable of oxidative respiratory burst activity (24), others have demonstrated that Kupffer cells, stimulated in vitro, are able to exhibit oxidative respiratory burst activity (36-38). Kupffer cells infected with Plasmodium berghei produced reactive oxygen species (39) and were able to effectively kill parasites. Also, stimulation of Kupffer cells by injection of Lactobacillus casei augmented oxygen radical production (40). It has been shown that there is a close correlation with Kupffer cells capacity to release oxygen intermediates and their ability to kill intracellular organisms (33, 39). In this study, it was found that catalase, which inactivates hydrogen peroxide, partially inhibits the amoebicidal ability of activated Kupffer cells (Fig. 5). The results of this study also confirmed other observations that elicited Kupffer cells were able to exhibit oxidative respiratory burst (36-38, 40). In yet another study (41) it was found that amoebic trophozoites were susceptible to exogenous hydrogen peroxide. The oxidative-dependent and -independent mechanisms of activated macrophages against amoebae in humans and animals was previously reported by some investigators (2, 3, 5,

216 . E. GHADIRIAN and A. SALIMI
30). Our results suggest that activated Kupffer cells are able to produce hydrogen peroxide and other molecules and these components are able to kill E. histolytica trophozoites.
The results presented in Figure 6 show that Kupffer cells, activated by IFN -y and LPS, possibly produce TNF and that this TNF mediates the killing of E. histolytica trophozoites; The administration of anti-TNF antibody to treated Kupffer cells reduced the number of dead trophozoites significantly. Other authors (17, 42, 43) have also shown that activated Kupffer cells were able to produce TNF. It has been demonstrated that TNF plays an important role in host defense against certain tumors (17, 44-46) and parasites (47-50) in vitro. The role of TNF as an autocrine molecule that is able to trigger its own production was reported by some investigators (31, 46, 51). In our study, it seems that TNF produced by activated Kupffer cells is able to trigger the same Kupffer cells to produce more TNF as well as other cytotoxic molecules which are capable of killing E. histolytica trophozoites in vitro (Fig. 6).
In a recent study (52), the effect of TNF on the amoebicidal activity of Kupffer cells has been described. In this study, it has been shown that TNF alone is able to induce Kupffer cells to acquire amoebicidal activity. It was demonstrated that both oxygen-dependent and -independent mechanisms are important for the killing of amoebae by the TNF-treated Kupffer cells.
Another report by DENIS and GHADIRIAN (53) examined the importance of reactive nitrogen intermediates in macrophage amoebicidal function. In this study, it is shown that reactive nitrogen species play an important role in the killing of E. histolytica by activated macrophages. They confirmed other investigations that reactive nitrogen species are an important component of the microbicidal armamentarium of activated macrophages. Of considerable interest are that recent findings which show that the mono kine TNF acts as an endogenous trigger for the production of reactive nitrogen intermediates. This is in keeping with the role of TNF as an important mono kine involved in stimulating the amoebicidal activity of mouse macrophages. IFN-y may act as a priming agent which stimulates TNF production and its associated nitric oxide production.
In summary, we have shown that murine Kupffer cells activated by IFNy and LPS are capable of killing the virulent parasite, E. histolytica in vitro. Our data also indicates that following appropriate activation and establishment of contact, both oxygen-dependent and oxygen-independent mechanisms are probably involved in the killing of the parasite by host Kupffer cells.
Acknowledgement
This work was supported by a grant from the Thrasher Research Fund.

Effect of IFN-y on Amoebae· 217
References
1. WALSH, J. A. 1986. Problems in recognition and diagnosis of amoebiasis. Estimation of the global magnitude for morbidity and mortality. Rev. Infect. Dis. 8: 228.
2. SALATA, R. A., R. D. PEARSON, and J. I. RAVDIN. 1985. Interaction of human leukocytes and Entamoeba histolytica: killing of virulent amoeba by the activated macrophages. J. Clin. Invest. 76: 491.
3. SALATA, R. A., H. W. MURRAY, B. Y. RUBIN, and J. I. RAVDIN. 1987. The role of gamma interferon in the generation of human macrophages cytotoxic for Entamoeba histolytica trophozoites. Am. Trop. Med. Hyg. 37: 72.
4. SALATA, R. A. and J. I. RAVDIN. 1986. Review of the human mechanisms directed against Entamoeba histolytica. Rev. Infect. Dis. 8: 261.
5. GHADIRIAN, E. and D. BOUT. 1988. In vitro killing of Entamoeba histolytica trophozoites by interferon-y-activated mouse macrophages. Immunobiol. 176: 341.
6. TRISSL, D. 1982. Immunology of Entamoeba histolytica in human and animal hosts. Rev. Infect. Dis. 4: 1154.
7. KRUPP, I. M. and S. J. POWELL. 1971. Antibody response to invasive amoebiasis in Durban South Africa. Am. J. Trop. Med. Hyg. 20: 414.
8. KRUPP, I. M. 1970. Antibody response in intestinal and extraintestinal amoebiasis. Am. J. Trop. Med. Hyg. 19: 57.
9. GHAHDIRIAN, E. and E. MEEROVITCH. 1981a. Effect of splenectomy on the size of amoebic liver abscesses and metastatic foci in hamsters. Infec. Immun. 31: 329.
10. GHADIRIAN, E. and E. MEEROVITCH. 1981b. Effect of immunosuppression on the size and metastasis of amoebic liver abscesses in hamsters. Parasite Immunol. 3: 329.
11. GHADIRIAN, E. and E. MEEROVITCH. 1982. Macrophage requirement for host defence against experimental hepatic amoebiasis in the hamster. Parasite Immunol. 4: 219.
12. GHADIRIAN, E. and P. A. L. KONGSHAVN. 1984. Genetic control of susceptibility of mice to infection with E. histolytica. Parasite Immunol. 6: 349.
13. GHADIRIAN, E., E. MEEROVITCH, and P. A. L. KONGSHAVN. 1983. Role of macrophages in host defense against hepatic amoebiasis in hamsters. Infec. Immun. 42: 1017.
14. GHADIRIAN, E. and P. A. L. KONGSHAVN. 1986. Protection of mice against intestinal amoebiasis with BCG, Corynebacterium parvum and Listeria monocytogenes. Parasite Immuno!. 8: 663.
15. KATZENSTEIN, D., V. RICKERSON, and A. BRAUDE. 1982. New concepts of amebic liver abscess derived from hepatic imaging, serodiagnosis, and hepatic enzymes in 67 consecutive cases in San Diego. Medicine 61: 237.
16. DECKER, T., A. KIDERLEN, and M. L. LOHMANN-MATTHES. 1985. Liver macrophages (Kupffer cells) as cytotoxic effector cells in extracellular and intracellular cytotoxicity. Infect. Immun. 50: 358.
17. DECKER, T., M. L. LOHMANN-MATTHES, and G. E. GIFFORD. 1987. Cell-associated tumor necrosis factor (TNF) as a killing mechanism of activated cytotoxic macrophages. J. Immuno!. 138: 957.
18. DECKER, T., M. L. LOHMANN-MATTHES, U. KARCK, T. PETERS, and K. DECKER. 1989. Comparative study of cytotoxicity, tumor necrosis factor, and prostaglandin release after stimulation of rat Kupffer cells, murine Kupffer cells, and murine inflammatory liver macrophages. J. Leuk. Bio!. 45: 139.
19. SHERWOOD, E., D. L. WILLIAMS, R. B. McNAMME, E. L. JONES, I. B. BROWDER, and N. R. DILUZIO. 1987. In vitro tumoricidal activity of resting and glucan-activated Kupffer cells. J. Leuk. Bio!. 42: 69.
20. CHADEE, K. and E. MEEROVITCH. 1984. The Mongolian gerbil (Meriones unguculatus) as an experimental host for Entamoeba histolytica. Am. J. Trop. Med. Hyg. 33: 47.
21. ROBINSON, G. L. 1968. The laboratory diagnosis of human parasitic amoebae. Trans. Roy. Soc. Trop. Med. Hyg. 62: 285.
22. PACE. J. L., S. W. RUSSELL, B. A. TORRES, H. M. JOHNSON, and P. W. GARY. 1983. Recombinant mouse Interferon gamma induces the primary step in macrophage activation for tumor cell killing . .T. Immunol. 130: 2011.

218 . E. GHADIRIAN and A. SALIMI
23. DIESSELHOFF-DEN DULK, M. M. c., R. W. CROFTON, and R. VAN FURTH. 1979. Origin and kinetics of Kupffer cells during an acute inflammatory response. Immunology 37: 7.
24. LEPAY, D. A., C. F. NATHAN, R. M. STEINMAN, H. W. MURRAY, and Z. A. COHN. 1985. Murine Kupffer cells: mononuclear phagocytes deficient in the generation of reactive oxygen intermediates. J. Exp. Med. 161: 1079.
25. LEPAY, D. A., R. M. STEINMAN, C. F. NATHAN, H. W. MURRAY, and A. C. ZANVIL. 1985 Liver macrophages in murine listeriosis: cell mediated immunity is correlated with an influx of macrophages capable of generating reactive oxygen intermediates. J. Exp. Med. 161: 1503.
26. MISHELL, B. B. and S. M. SHIIGI (eds.). 1980. Selected methods in cellular immunology. W. H. Freeman and Company, San Francisco. 23 pp.
27. OLIVIER, M. and C. E. TANNER, 1987. Susceptibilities of macrophage populations to infection in vitro by Leishmania donovani. Infect. Immun. 55: 467.
28. CROCKER, P. R., J. M. BLACKWELL, and D. J. BRADLEY. 1984. Expression of the natural resistance gene lsh in resident liver macrophages. Infect. Immun. 43: 1033.
29. CROFTON, R. W., M. M. C. DIESSELHOFF-DEN DULK, and R. V. FURTH. 1978. The origin, kinetics, and characteristic of Kupffer cells in the normal steady state. J. Exp. Med. 148: 1.
30. DENIS, M. and K. CHADEE. 1989. Cytokine activation of murine macrophages for in vitro killing of Entamoeba histolytica trophozoites. Infect. Immun. 57: 1750.
31. DECKER, K. 1990. Biologically active products of stimulated liver macrophages (Kupffer cells). Eur. J. Biochem. 192: 245.
32. JONES, E. A. and J. A. SUMMERFIELD. 1988. Kupffer cells. In: The liver: Biology and pathobiology, second edition, eds. I. M. ARIAS, W. B. JAKOBY, H. POPPER, D. SCHACHTER, and D. A. SHAFRITZ. Raven Press, Ltd., New York. 683 pp.
33. FILICE, G. A. 1988. Antibacterial properties of Kupffer cells. Infec. Immun. 56: 1430. 34. VAN BossuYT, H., C. DESMARTEZ, B. RAMBOUT, and E. WISSE. 1988. Response of
cultured rat Kupffer cells to lipopolysaccharide. Arch. Toxico!. 62: 316. 35. SHARMA, S. D. and W. F. PIESENS. 1978. Tumor cell killing by macrophages activated in
vitro with lymphocytes mediators. III. Inhibition by cytochalasins, cholchicine, and vinblastine. Cel!. Immuno!. 38: 276.
36. ARTHUR, M. J. P., P. KOWALSKI-SAUNDERS, and R. WRIGHT. 1988. Corynebacterium parvum-elicited hepatic macrophages demonstrate enhanced respiratory burst activity compared with resident Kupffer cells in the rat. Gastroenterology 91: 174.
37. MATSUO, S., A. NAKAGAWARA, K. IKEDA, M. MITSUYAMA, and K. NUMOTO. 1985. Enhanced release of reactive oxygen intermediates by immunologically activated rat Kupffer cells. Clin. Exp. Immunol. 59: 203.
38. BHATNAGER, R., R. SCHIRMER, M. ERNST, and K. DECKER. 1981. Superoxide release by zymosan-stimulated rat Kupffer cells in vitro. Eur. J. Biochem. 119: 171.
39. PRASAD, R. N., K. J. VIRK, R. C. MAHAJAN, and N. K. GANGULY. 1990. Reactive oxygen species generation by Kupffer cells and blood monocytes of mice infected with Plasmodium berghei and Chloroquine treatment. Jap. J. Exp. Med. 160: 1.
40. HASHIMOTO, S., K. NOMOTO, T. MATSOZAKI, T. YOKOKURA, and M. MUTAI. 1984. Oxygen radical production by peritoneal macrophages and Kupffer cells elicited with Lactobacillus casei. Infect. Immun. 44: 61.
41. GHADIRIAN, E., S. D. SOMMERFIELD, and P. A. L. KONGSHAVN. 1986. Susceptibility of Entamoeba histolytica to oxidants. Infect. Immun. 51: 263.
42. BUSAM, K. J., A. HOMFEID, R. ZAWATZKY, S. KASTNER, J. BAUER, W. GEROK, and K. DECKER. 1990. Virus vs endotoxin-induced activation of liver macrophages. Eur. J. Biochem. 191: 577.
43. PETERS, T., U. KRACK, and K. DECKER. 1990. Interdependence of tumor necrosis factor, prostaglandin E2, and protein synthesis in lipopolysaccharide-exposed rat Kupffer cells. Eur. J. Riochem. 191: <;83.
44. CARSWELL, E. A., L. J. OLD, R. L. KASSEL, S. GREEN, N. FOIRE, and B. WILLIAMSON. 1975. An endotoxin-induced serum factor that causes necrosis of tumors. Proc. Nat!. Acad. Sci. USA 72: 3666.
45. OLD, L. J. 1985. Tumor necrosis factor (TNF). Science 230: 630.

Effect of IFN-y on Amoebae· 219
46. MACE, K. F., M. J. EHRKE, K. HORl, D. L. MACCUllBlN, and E. MIHICH. 1988. Role of tumor necrosis factor in macrophage activation and tumoricidal activity. Cancer Res. 48: 5427.
47. TAVERN, J., J. TAVERNIER, W. FIERS, and J. H. L. PLAYFAIR. 1984. Recombinant tumor necrosis factor inhibits malaria parasites in vivo but not in vitro. Clin. Exp. Immunol. 67: l.
48. DETITTo, E. H., J. R. CATTERALL, and J. S. REMINGTON. 1986. Activity of recombinant tumor necrosis factor on Toxoplasma gondii and Trypanosoma cruzi. J. Immunol. 134: 1342.
49. JAMES, S. L., J. GLAVEN, S. GOLDBERG, M. S. MELTZER, and E. PEARCE. 1990. Tumor necrosis factor (TNF) as a mediator of macrophage helminthotoxic activity. Parasite Immunol. 12: 1.
50. KONGSHAVN, P. L. A. and E. GHADIRIAN. 1988. Enhancing and suppressive effects of tumor necrosis factorlcachectin on growth of Trypanosoma musculi. Parasite Immunol. 10: 581.
51. PHILIP, R. and L. B. EpSTEIN. 1986. Tumor necrosis factor as immunomodulator and mediator of monocyte cytotoxicity induced by itself, y-interferon and interleukin-1. Nature 323: 86.
52. SALIMI, A. and E. GHADIRIAN. 1992. Effect of recombinat tumor necrosis factor alpha on in vitro amoebicidal activity of murine Kupffer cells. Microbial Pathogen. Submitted for publication.
53. DENIS, M. and E. GHADIRIAN. 1992. Activated macrophages kill Entamoeba histolytica trophozoites by releasing reactive nitrogen intermediates. Microbial Pathogen. 12: 193.
Dr. ESFANDIAR GHADIRIAN, Department of Biochemistry and Microbiology, University of Victoria, P.O. Box 3055, Victoria, British Columbia, Canada V8W 3P6