il movimento - dottorunk.files.wordpress.com · in questo caso di co-contrazione . la presenza di...
TRANSCRIPT

Il movimento
June 23, 2011
I movimenti possibili vengono divisi in tre tipologie a seconda della circuiteria neuronale sottostante: i movimentivolontari, i movimenti ri�essi ed i movimenti ritmici. I movimenti volontari non presentano i caratteri diobbligatorietà degli altri due e inoltre migliorano con l'esperienza; una de�nizione alternativa può infatti essere quelladi movimenti �ne appreso con l'esperienza. I movimenti ri�essi sono privi invece dello stadio della consapevolezza.In generale per qualsiasi tipo di movimento sono importanti le informazioni snesoriali, che non necessariamentedevono diventare coscienti per poter essere sfruttate; l'informazione visiva è sicuramente la più importante, tanto èvero che un cieco si muove in modo molto diverso da un vedente. Il SNC sfrutta i segnali a�erenti dalla periferia indue modi: fornisce sensazioni da un lato, ma dall'altro guida il movimento.
1 I ri�essi
I movimenti più semplici sono quelli ri�essi, che sono risposte involontarie relativamente stereotipate aduno stimolo. Qualsiasi ri�esso per essere de�nito tale deve nascere da uno stimolo esterno; la risposta può essererappresentata da una contrazione muscolare (liscia o striata) o da una secrezione ghiandolare (endocrina o esocrina).La risposta deve essere sempre totalmente involontaria. La risposta ad uno stimolo è in generale sempre simile, perquesto si parla di sterotipia relativa; esistono movimenti stereotipati non ri�essivi, ad esempio la scrittura, quindinon è un attributo su�ciente a de�nire un ri�esso. I ri�essi molto spesso sono molto stereotipati in certe situazioni,ma pochissimo in altre: questo attributo dipende molto dal contesto, tuttavia è presente alla nascita. La stereotipiadei ri�essi non richiede dunque alcun tipo di apprendimento prima di poter essere messa in atto. In generale esistepoi una relazione invariante tra la sede dello stimolo e la sede della risposta, rapporto che non esiste nei movimentivolontari; in maniera simile esiste anche un rapporto tra intensità dello stimolo e intensità della risposta.
Alla base di un ri�esso esiste una serie di cinque componenti fondamentali che prende il nome di arco ri�esso.Il primo elemento è uno stimolo speci�co (I) in grado di attivare un recettore che invia le informazioni tramiteuna via a�erente (II) al SNC. All'interno del SNC (tronco dell'encefalo o midollo spinale) esiste una popolazionedi neuroni che costituisce il centro ri�esso (III) che elabora le informazioni in arrivo e le reinvia tramite una viae�erente (IV) all'e�ettore (V).
1.1 I ri�essi miotatici
Lo stiramento di un muscolo risulta in una contrazione del muscolo stesso e questo comportamento prende il nomedi ri�esso miotatico da stiramento o diretto. Lo stimolo adeguato è dunque un allungamento passivo del muscoloe viene recepito dal fuso neuromuscolare. All'interno del fuso le �bre a�erenti 1a danno informazione sullalunghezza del muscolo e sulla velocità di allungamento mentre le �bre 2 danno informazioni sulla sola lunghezza.Le �bre 1a penetrano nel midollo spinale e formano un ramo ascendente per il sistema delle colonne dorsali e unramo anteriore che si dirige al corno ventrale dove sono contenuti i motoneuroni; queste �bre si portano ad eccitaredirettamente i motoneuroni del muscolo omonimo, ed è questo l'unico esempio di circuito monosinaptico conosciutonell'uomo. La mediazione di vari interneuroni produce invece l'eccitazione dei motoneuroni dei muscoli sinergici el'inibizione di quelli dei muscoli antagonisti (in questo secondo caso l'interneurone prende il nome di interneuroneinibitorio 1a). Il risultato dell'attivazione delle �bre a�erenti 1a è dunque la contrazione del muscolo omonimoinsieme agli antagonisti ed il rilassamento degli antagonisti.
1
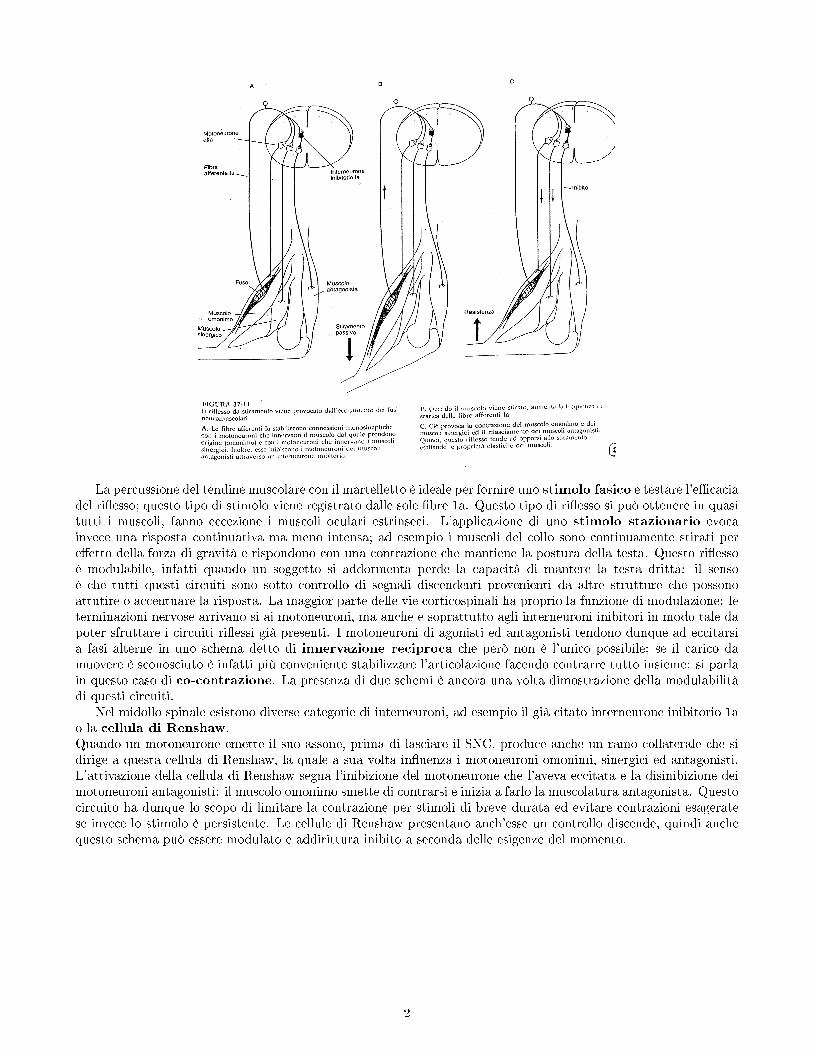
La percussione del tendine muscolare con il martelletto è ideale per fornire uno stimolo fasico e testare l'e�caciadel ri�esso; questo tipo di stimolo viene registrato dalle sole �bre 1a. Questo tipo di ri�esso si può ottenere in quasitutti i muscoli, fanno eccezione i muscoli oculari estrinseci. L'applicazione di uno stimolo stazionario evocainvece una risposta continuativa ma meno intensa; ad esempio i muscoli del collo sono continuamente stirati pere�etto della forza di gravità e rispondono con una contrazione che mantiene la postura della testa. Questo ri�essoè modulabile, infatti quando un soggetto si addormenta perde la capacità di mantere la testa dritta: il sensoè che tutti questi circuiti sono sotto controllo di segnali discendenti provenienti da altre strutture che possonoattutire o accentuare la risposta. La maggior parte delle vie corticospinali ha proprio la funzione di modulazione: leterminazioni nervose arrivano si ai motoneuroni, ma anche e soprattutto agli interneuroni inibitori in modo tale dapoter sfruttare i circuiti ri�essi già presenti. I motoneuroni di agonisti ed antagonisti tendono dunque ad eccitarsia fasi alterne in uno schema detto di innervazione reciproca che però non è l'unico possibile; se il carico damuovere è sconosciuto è infatti più conveniente stabilizzare l'articolazione facendo contrarre tutto insieme: si parlain questo caso di co-contrazione. La presenza di due schemi è ancora una volta dimostrazione della modulabilitàdi questi circuiti.
Nel midollo spinale esistono diverse categorie di interneuroni, ad esempio il già citato interneurone inibitorio 1ao la cellula di Renshaw.Quando un motoneurone emette il suo assone, prima di lasciare il SNC, produce anche un ramo collaterale che sidirige a questa cellula di Renshaw, la quale a sua volta in�uenza i motoneuroni omonimi, sinergici ed antagonisti.L'attivazione della cellula di Renshaw segna l'inibizione del motoneurone che l'aveva eccitata e la disinibizione deimotoneuroni antagonisti: il muscolo omonimo smette di contrarsi e inizia a farlo la muscolatura antagonista. Questocircuito ha dunque lo scopo di limitare la contrazione per stimoli di breve durata ed evitare contrazioni esageratese invece lo stimolo è persistente. Le cellule di Renshaw presentano anch'esse un controllo discende, quindi anchequesto schema può essere modulato e addirittura inibito a seconda delle esigenze del momento.
2

L'organo tendineo del Golgi riconosce come stimolo adeguato la tensione posta a carico di un muscolo ed èalla base del ri�esso miotatico inverso; in questo caso le risposte sono opposte: i muscoli antagonisti si contraggonoe gli agonisti e sinergici si rilasciano. L'aumento di tensione è segnato dall'aumento di scarica del recettore el'informazione raggiunge il midollo spinale per tramite delle �bre a�erenti 1b che raggiungono gli interneuroniinibitori 1b. Gli interneuroni inibitori formano sinapsi con i vari motoneuroni, eccitando gli antagonisti ed inibendogli altri: il ri�esso miotatico inverso poggia dunque su un circuito sempre disinaptico. Questo ri�esso è moltosfruttato dal sistema nervoso nell'adattamento muscolare alla fatica. Quando la corteccia attua uno schema motoriofa contrarre dei muscoli i quali accorciandosi fanno scaricare l'organo tendineo e quindi riducono la loro forza totale:l'intensità di una contrazione è data dunque dall'intensità dello schema motorio al netto dell'inibizione miotatica.Quando una data posizione viene mantenuta per un certo intervallo di tempo il muscolo si a�atica e produce menoforza; lo schema motorio non deve però essere ricalibrato alla nuova situazione in quanto meno forza signi�caanche meno inibizione e quindi il crollo è meno repentino. Il circuito miotatico inverso può inoltre essere attivato daa�erenze cutanee o articolari. Le a�erenze articolari esistono solo per angoli articolari estremi e quindi pericolosi:si inibiscono così contrazioni che rischierebbero di danneggiare l'articolazione stessa. Le a�erenze cutanee sono legateinvece alla possibilità di maneggiare con cura gli oggetti: appena entro in contatto con un oggetto fragile la miacontrazione è inibita e posso così avere una presa �soft�.
Il ri�esso miotatico da stiramento è di fatto un feedback negativo sulla lunghezza del muscolo: quando questaaumenta il sistema si oppone. La prima ipotesi sul senso di questo sistema fu dunque quella di controllore dellalunghezza muscolare. Il guadagno del ri�esso miotatico è in realtà molto basso, cioè l'entità della correzioneè sempre molto minore di quella della perturbazione, quindi non è in grado di compensare e�cacemente un al-lungamento errato: il senso di questo ri�esso deve dunque essere un altro. Il guadagno è molto basso in quantoper necessità �siche questi schemi neuronali si portano dietro una certa latenza non eliminabile; un sistema adalto guadagno e alta latenza è molto instabile in quanto nell'arco di tempo necessario a correggere la lunghezza lesituazioni ambientali potrebbero essere già cambiate.
3

Il vero signi�cato del ri�esso è più evidente analizzando il comportamento anche dei motoneuroni gamma. Quandoil muscolo ha un carico da sostenere le vie motorie discendenti attivano i motoneuroni gamma che stirano il fuso equesto sollecita i motoneuroni alfa a per la contrazione. Se il carico aumenta la scarica del fuso cresce e quindi lacontrazione muscolare si fa più intensa; se invece il carico diminuisce il fuso smette di scaricare in quanto sensibileal solo allungamento e il muscolo si allunga �ntanto che lo stiramento è allineato allo stato di attivazione deimotoneuroni gamma. Questo meccanismo è detto regolazione gamma del ri�esso da stiramento: in sostanzail ri�esso è un meccanismo di servoassistenza al mantenimento di una posizione. Una seconda interpretazione delri�esso è quella delmantenimento della rigidezza in accoppiata con il ri�esso inverso. La rigidezza di un muscoloè de�nita come
K =∆T
∆L
Una variazione di lunghezza induce dunque una variazione di tensione proporzionale alla rigidezza. La forza svilup-pata dipende dunque dalla lunghezza del muscolo, ma se ogni comando motorio dovesse tenere conto della lunghezzail controllo sarebbe complicatissimo: mantenendo costante la rigidezza i comandi corticali possono fare a meno ditenere conto dello stato di allungamento muscolare, sempli�cando di molto il sistema. In sostanza l'adattamento infunzione dei carichi e della lunghezza viene operato dall'insieme dei ri�essi miotatici, sollevando la corteccia da talicompiti.
1.2 Il ri�esso �essorio
Il ri�esso �essorio coinvolge tutta la muscolatura �essoria di un arto o di un segmento corporeo e ha la funzione diallontanare una porzione somatica da uno stimolo. Lo stimolo più importante nell'evocare un ri�esso �essorio è dinatura nociva, cioè quello in grado di attivare i nocicettori.
4

Le scariche a�erenti dai nocicettori vengono trasportate dalle �bre nocicettive A delta o C e raggiungono il cornodorsale del midollo spinale, dove si impegnano in un circuito polisinaptico; questo circuito per tramite di una seriedi interneuroni raggiunge i motoneuroni dei muscoli �essori attivandoli e di quelli estensori inibendoli. Quandol'arto viene allontanato con successo la contrazione rimane comunque dopo la �ne dello stimolo: questo indica che ilcircuito deve essere riverberante, per non ricadere nello stesso stimolo. Nel ri�esso da retrazione sono ben evidentisia il rapporto con la sede dello stimolo che con la sua intensità. Alla �essione di un intero arto si accompagnapoi l'estensione crociata dell'arto opposto; questo comportamento è particolarmente utile nei quadrupedi inquanto permette di redistribuire il peso dell'animale in modo da non cadere per l'assenza di un punto d'appoggio.Nell'uomo questo tipo di risposta è presente per gli arti inferiori, ma è meno e�cace per via della postura bipededa noi assunta.
Un circuito simile a quello del ri�esso �essorio viene sfruttato anche da altre due tipologie di ri�essi importantiin clinica:
• I ri�essi addominali vengono evocati dalla stimolazione dell'addome con una punta smussa. Le contrazionisono organizzate secondo la topogra�a dell'addome.
• I ri�essi plantari richiedono un intervento centrale per manifestarsi. In un adulto sano la stimolazione dellapianta del piede produce una �essione plantare delle dita; in un neonato o in un soggetto con danni allevie corticospinali la risposta è invece un'estensione a ventaglio delle dita che prende il nome di segno di
Babinski.
2 I movimenti ritmici e la locomozione
La locomozione è una capacità posseduta da tutti gli animali, e nel nostro caso è divisa in una fase di oscillazionee in una fase di appoggio. Negli anni passati questo schema motorio era di origine incerta: da un lato losi credeva talmente complesso da dover essere volontario, dall'altro appariva tanto stereotipato da poter essereri�esso. Un movimento di origine volontaria non dovrebbe essere possibile in un preparato spinalizzato, cioècon una lesione mediotoracica: in realtà un animale in questa condizione posto su un tappeto mobile è in gradodi compiere movimenti di locomozione, quindi non si tratta di movimento volontario. Un esperimento analogosi può condurre su un preparato dea�erentato, cioè con radici spinali sezionate: in questo caso non dovrebbeessere possibile realizzare movimenti ri�essi in quanto i recettori periferici non possono condurre informazioni alSNC. In animali dea�erentati si dimostrò però che iniezioni di L-dopa a livello spinale producevano movimenti dilocomozione: non si può dunque parlare nemmeno di un ri�esso. Un terzo esperimento sfrutta la paralisi indotta dalcuraro: in un animale curarizzato i muscoli non si possono muovere, ma andando a registrare l'attività dei nervi
5

dei muscoli estensori e �essori si trova un'attività alternata. In sostanza la locomozione non può essere volontariae nemmeno ri�essa: si tratta di un movimento ritmico la cui genesi è legata a circuiti di neuroni detti generatoricentrali di schemi motori. I generatori centrali sono attivati da segnali tonici in arrivo dalla corteccia o daaltre regioni del SNC e si occupano di trasformare questi segnali da tonici a fasici. Nel caso della locomozione igeneratori centrali si trovano a livello del midollo spinale se sono tutti innati: un bambino non riesce a camminarenon perchè privo di questi centri, ma perchè incapace di controllarli. In un animale da laboratorio è evidente chea�errando un arto gli altri tre continuano a potersi muovere: esistono dunque quattro generatori di schemi motori,uno per arto. La locomozione può essere indotta sperimentalmente andando a stimolare certe regioni corticali, nelgatto la più importante è la regione motoria mesencefalica che dunque deve essere in collegamento ai centrimotori. Variando l'intensità degli stimoli alla regione mesencefalica non cambia l'ampiezza dei potenziali d'azionedei movimenti locomotori, ma cambia l'accoppiamento dei generatori: si passa da un accoppiamento alternato abasse intensità ad uno sincrono ad intensità elevate.
Un animale spinalizzato è in grado di compiere movimenti locomotori, ma questi sono molto semplici ed im-poveriti rispetto a quelli di un animale sano. Le a�erenze periferiche hanno infatti il ruolo di adattare i generatoricentrali alle situazioni reali del mondo esterno; in conclusione esistono si dei generatori centrali, ma questi pro-ducono solamente l'essenziale per la locomozione, tutto il resto è frutto di un'elaborazione che richiede l'apporto dia�erenze periferiche.
Un uomo spinalizzato non cammina assolutamente. Qualche risultato è stato ottenuto con stimoli molto piùcomplessi rispetto a quelli su�cienti per un animale, e comunque solo in individui giovani. I generatori centrali dischemi motori esistono anche nell'uomo, solo che nell'evoluzione i controlli dei centri superiori sono diventati moltopiù marcati, al punto da limitare molto la possibilità di attivarli direttamente.
6