how do animals sense the environment
TRANSCRIPT

BIOL 3520: Cell Physiology
1
Lecture 25: How do animals sense their
environment?
Tiffany A. Timbers, Ph.D.http://www.slideshare.net/ttimbers/how-do-animals-sense-the-environment

2
http://smallbusinessbonfire.com/do-your-services-pass-the-sniff-test

http://wabikes.org/2010/11/15/stop-signs-the-kudzu-of-american-bike-paths/3

https://s-media-cache-ak0.pinimg.com/736x/90/13/4c/ 90134cdcb39ab99d7485a34f199f615c.jpg
4

Learning Objectives
You will be able to:
• Describe the mechanism for sensation in olfactory sensory neurons.
• Describe how the components of cilia contribute to sensation.
• Name 3 major systems affected in patients with cilia disorders.
• Interpret results from an experiment with regards to a given hypothesis about sensation.
5

6http://www.wikicell.org/eightSystemImage/sensorySystem/Sensory%20System.jpg
touch temperature pain
Human Sensory Systems

Cell Physiology Source Book, 4th Edition
Chapter 36, p. 633-646 - Sensory Receptors and Mechanotransduction Chapter 38, p. 669-678 - Visual Transduction Chapter 39, p. 690-695 - Gustatory and Olfactory Sensory Transduction
Assigned Readings:
7

Reminder: Action Potential Generation
extracellular
intracellular
Na+
Ca2+
Na+
Na+
Na+
Ca2+Ca2+Ca2+
Na+
Na+
Ca2+
Na+
Resting state
8
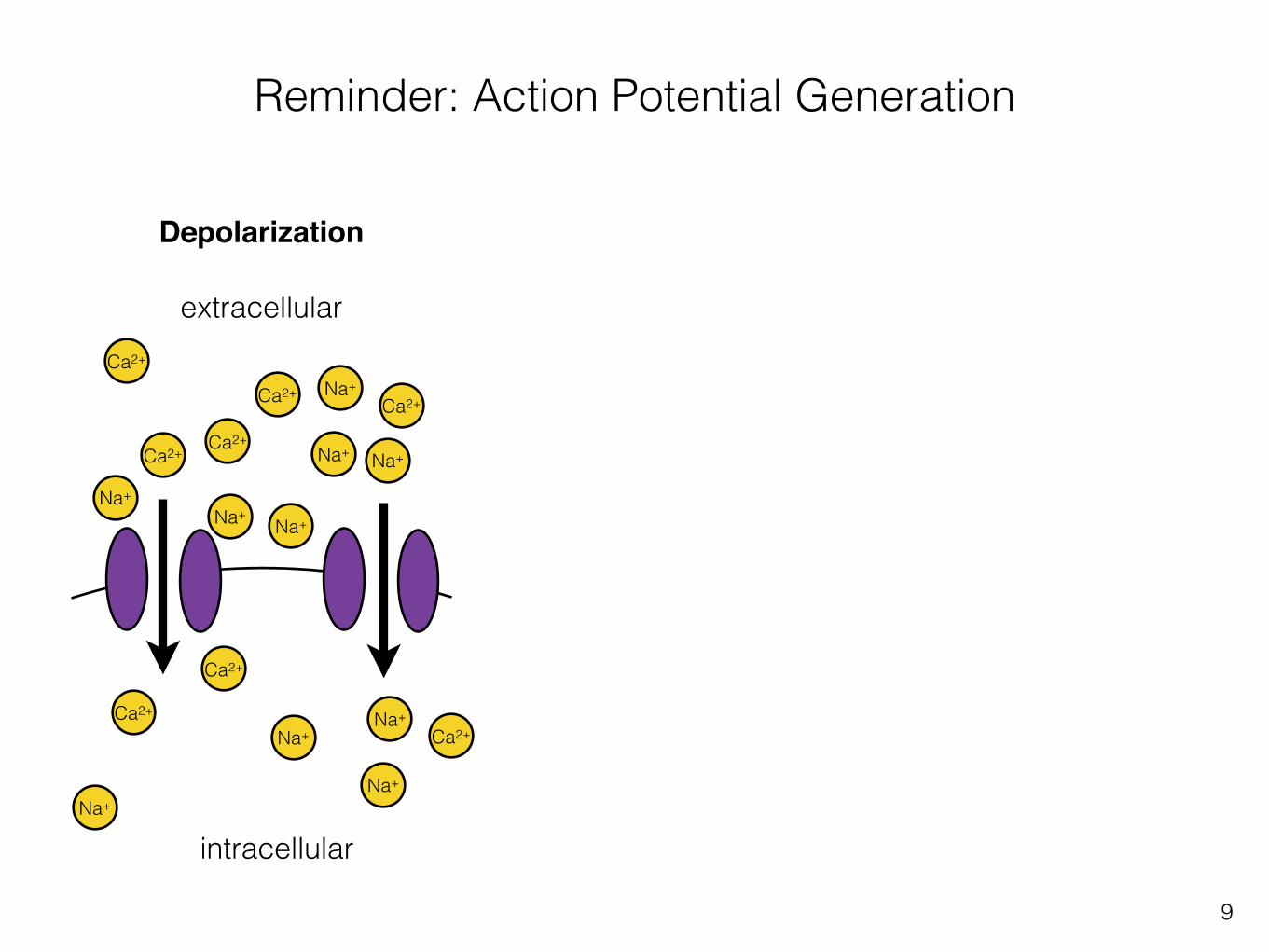
Reminder: Action Potential Generation
extracellular
intracellular
Na+
Ca2+
Na+
Na+ Na+
Ca2+Ca2+
Ca2+
Na+
Na+
Ca2+
Na+
Na+
Ca2+
Na+
Ca2+Na+
Ca2+
Depolarization
9

Reminder: Action Potential Generation
extracellular
intracellular
Na+
Ca2+
Na+
Na+ Na+
Ca2+Ca2+
Ca2+
Na+
Na+
Ca2+
Na+
Na+
Ca2+
Na+
Ca2+Na+
Ca2+
Depolarization
10

11
projection to brain
olfactory receptor neuron
cilia
olfactory bulb
inhaled air
Olfactory Receptor Neurons
2007 Wolfers Kluwer Health | Lippincott Williams & Wilkins
12
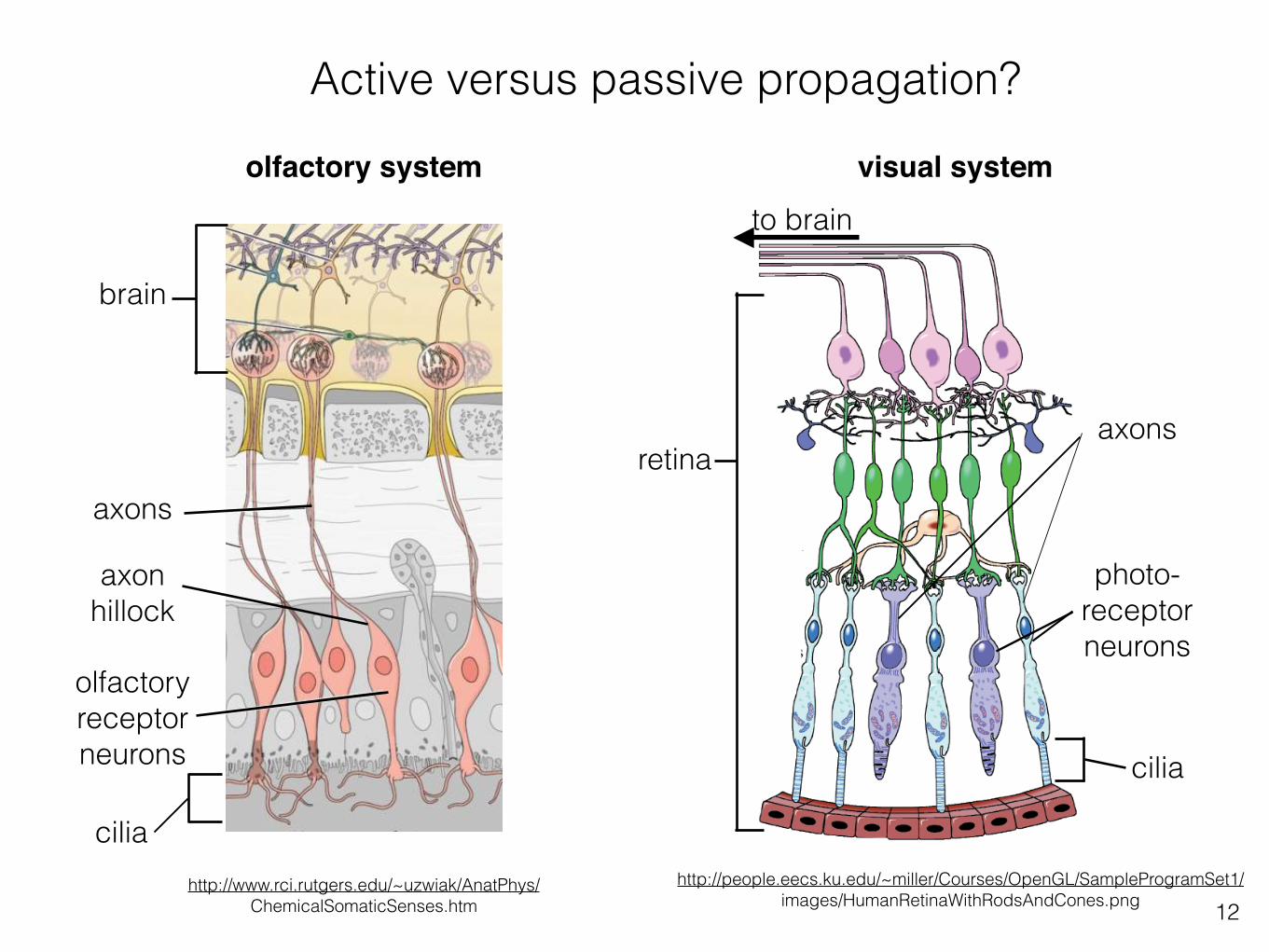
Active versus passive propagation?
http://www.rci.rutgers.edu/~uzwiak/AnatPhys/ ChemicalSomaticSenses.htm
axons
olfactory receptor neurons
cilia
axon hillock
brain
olfactory system visual system
photo- receptor neurons
axons
to brain
cilia
retina
http://people.eecs.ku.edu/~miller/Courses/OpenGL/SampleProgramSet1/ images/HumanRetinaWithRodsAndCones.png

13
Modality - cells have specialized receptors to sense external stimuli
Cell Physiology Source Book 4th Edition, Figure 36.1

14
represent odor intensity, and which represent other infor-mation, such as odor quality?
We consider several models of odor intensity coding atthis level. Are they compatible with physiological data? Dothey predict stable perception of odor quality over a rangeof concentrations?
Although the neural code in the olfactory bulb mustrepresent both odor concentration and identity it is crucialfor the brain to disambiguate the two kinds of information.For olfactory navigation tasks, stimulus concentrationvaries with distance from a target odor source; animals
must be able to maintain a concentration-invariant repre-sentation of odor quality over biologically relevant concen-tration ranges to track the source. Although odors aregenerally thought to retain their quality over a range ofconcentrations, concentration changes greater than twoorders of magnitude may yield changes in odor qualityfor some odorants [3,31] but not others [32].
Spike rate codingGiven that odorant concentration is correlated with spikerates of OSN inputs to glomeruli, we may ask if this rate
Golfα Golfα AC3
GDP GTP
ATP
1.0
0.5
0.0
1.0
0.5
0.0
1E-3 0.01 0.1 1
1E-3 0.01 0.1 1
cAMPCa2+
Cl–
R
OH
Odorant
Transduc!oncurrent
R*
Transduc!oncurrent
Spike firing
Firin
g ra
teTr
ansd
uc!o
ncu
rren
tAc
!vat
ed re
cept
or(R
*)
Log [odorant]
Log [odorant]
[Odorant]
(A) (B)
(C)
(D)
Low
High
[Odorant]
βγ
TRENDS in Neurosciences
Figure 1. Odorant concentration coding in olfactory sensory neurons (OSNs). During sensory transduction (A), odorant molecules bind and stabilize the active states ofolfactory receptors (R) in ciliary membranes of OSNs. The activated receptors (R*) couple to G proteins (Golf) and increase synthesis of cyclic AMP (cAMP) by type IIIadenylyl cyclase (AC3). The cAMP opens cyclic nucleotide-gated channels that conduct calcium ions into the cilia and, in turn, open a channel (ANO2) mediating adepolarizing efflux of chloride ions. The resulting transduction current is passed to the OSN cell body, where it drives a train of action potentials (spikes). The concentrationof detected odorant is encoded nonlinearly at each step of transduction: by a hyperbolic dependence of the number of activated receptors (R*) in the cilia (B), a stronglycooperative variation in amplitude of the transduction current (C), and similar sigmoidal variation of spike firing rate relayed by OSN axons (D). Data from [113] (C,D):response of normalized currents and firing rates of frog OSN to cineole; mammalian OSNs exhibit similar dose–response profiles.
Review Trends in Neurosciences August 2014, Vol. 37, No. 8
446
represent odor intensity, and which represent other infor-mation, such as odor quality?
We consider several models of odor intensity coding atthis level. Are they compatible with physiological data? Dothey predict stable perception of odor quality over a rangeof concentrations?
Although the neural code in the olfactory bulb mustrepresent both odor concentration and identity it is crucialfor the brain to disambiguate the two kinds of information.For olfactory navigation tasks, stimulus concentrationvaries with distance from a target odor source; animals
must be able to maintain a concentration-invariant repre-sentation of odor quality over biologically relevant concen-tration ranges to track the source. Although odors aregenerally thought to retain their quality over a range ofconcentrations, concentration changes greater than twoorders of magnitude may yield changes in odor qualityfor some odorants [3,31] but not others [32].
Spike rate codingGiven that odorant concentration is correlated with spikerates of OSN inputs to glomeruli, we may ask if this rate
GolfαGolfαAC3
GDPGTP
ATP
1.0
0.5
0.0
1.0
0.5
0.0
1E-3 0.01 0.1 1
1E-3 0.01 0.1 1
cAMPCa2+
Cl–
R
OH
Odorant
Transduc!oncurrent
R*
Transduc!oncurrent
Spike firing
Firing rate
Transduc!on
current
Ac!vated receptor
(R*)
Log [odorant]
Log [odorant]
[Odorant]
(A) (B)
(C)
(D)
Low
High
[Odorant]
βγ
TRENDS in Neurosciences
Figure 1. Odorant concentration coding in olfactory sensory neurons (OSNs). During sensory transduction (A), odorant molecules bind and stabilize the active states ofolfactory receptors (R) in ciliary membranes of OSNs. The activated receptors (R*) couple to G proteins (Golf) and increase synthesis of cyclic AMP (cAMP) by type IIIadenylyl cyclase (AC3). The cAMP opens cyclic nucleotide-gated channels that conduct calcium ions into the cilia and, in turn, open a channel (ANO2) mediating adepolarizing efflux of chloride ions. The resulting transduction current is passed to the OSN cell body, where it drives a train of action potentials (spikes). The concentrationof detected odorant is encoded nonlinearly at each step of transduction: by a hyperbolic dependence of the number of activated receptors (R*) in the cilia (B), a stronglycooperative variation in amplitude of the transduction current (C), and similar sigmoidal variation of spike firing rate relayed by OSN axons (D). Data from [113] (C,D):response of normalized currents and firing rates of frog OSN to cineole; mammalian OSNs exhibit similar dose–response profiles.
ReviewTrends in Neurosciences August 2014, Vol. 37, No. 8
446
represent odor intensity, and which represent other infor-mation, such as odor quality?
We consider several models of odor intensity coding atthis level. Are they compatible with physiological data? Dothey predict stable perception of odor quality over a rangeof concentrations?
Although the neural code in the olfactory bulb mustrepresent both odor concentration and identity it is crucialfor the brain to disambiguate the two kinds of information.For olfactory navigation tasks, stimulus concentrationvaries with distance from a target odor source; animals
must be able to maintain a concentration-invariant repre-sentation of odor quality over biologically relevant concen-tration ranges to track the source. Although odors aregenerally thought to retain their quality over a range ofconcentrations, concentration changes greater than twoorders of magnitude may yield changes in odor qualityfor some odorants [3,31] but not others [32].
Spike rate codingGiven that odorant concentration is correlated with spikerates of OSN inputs to glomeruli, we may ask if this rate
GolfαGolfαAC3
GDPGTP
ATP
1.0
0.5
0.0
1.0
0.5
0.0
1E-3 0.01 0.1 1
1E-3 0.01 0.1 1
cAMPCa2+
Cl–
R
OH
Odorant
Transduc!oncurrent
R*
Transduc!oncurrent
Spike firing
Firing rate
Transduc!on
current
Ac!vated receptor
(R*)
Log [odorant]
Log [odorant]
[Odorant]
(A) (B)
(C)
(D)
Low
High
[Odorant]
βγ
TRENDS in Neurosciences
Figure 1. Odorant concentration coding in olfactory sensory neurons (OSNs). During sensory transduction (A), odorant molecules bind and stabilize the active states ofolfactory receptors (R) in ciliary membranes of OSNs. The activated receptors (R*) couple to G proteins (Golf) and increase synthesis of cyclic AMP (cAMP) by type IIIadenylyl cyclase (AC3). The cAMP opens cyclic nucleotide-gated channels that conduct calcium ions into the cilia and, in turn, open a channel (ANO2) mediating adepolarizing efflux of chloride ions. The resulting transduction current is passed to the OSN cell body, where it drives a train of action potentials (spikes). The concentrationof detected odorant is encoded nonlinearly at each step of transduction: by a hyperbolic dependence of the number of activated receptors (R*) in the cilia (B), a stronglycooperative variation in amplitude of the transduction current (C), and similar sigmoidal variation of spike firing rate relayed by OSN axons (D). Data from [113] (C,D):response of normalized currents and firing rates of frog OSN to cineole; mammalian OSNs exhibit similar dose–response profiles.
ReviewTrends in Neurosciences August 2014, Vol. 37, No. 8
446
transductioncurrent
represent odor intensity, and which represent other infor-mation, such as odor quality?
We consider several models of odor intensity coding atthis level. Are they compatible with physiological data? Dothey predict stable perception of odor quality over a rangeof concentrations?
Although the neural code in the olfactory bulb mustrepresent both odor concentration and identity it is crucialfor the brain to disambiguate the two kinds of information.For olfactory navigation tasks, stimulus concentrationvaries with distance from a target odor source; animals
must be able to maintain a concentration-invariant repre-sentation of odor quality over biologically relevant concen-tration ranges to track the source. Although odors aregenerally thought to retain their quality over a range ofconcentrations, concentration changes greater than twoorders of magnitude may yield changes in odor qualityfor some odorants [3,31] but not others [32].
Spike rate codingGiven that odorant concentration is correlated with spikerates of OSN inputs to glomeruli, we may ask if this rate
Golfα Golfα AC3
GDP GTP
ATP
1.0
0.5
0.0
1.0
0.5
0.0
1E-3 0.01 0.1 1
1E-3 0.01 0.1 1
cAMPCa2+
Cl–
R
OH
Odorant
Transduc!oncurrent
R*
Transduc!oncurrent
Spike firing
Firing rate
Transduc!on
current
Ac!vated receptor
(R*)
Log [odorant]
Log [odorant]
[Odorant]
(A) (B)
(C)
(D)
Low
High
[Odorant]
βγ
TRENDS in Neurosciences
Figure 1. Odorant concentration coding in olfactory sensory neurons (OSNs). During sensory transduction (A), odorant molecules bind and stabilize the active states ofolfactory receptors (R) in ciliary membranes of OSNs. The activated receptors (R*) couple to G proteins (Golf) and increase synthesis of cyclic AMP (cAMP) by type IIIadenylyl cyclase (AC3). The cAMP opens cyclic nucleotide-gated channels that conduct calcium ions into the cilia and, in turn, open a channel (ANO2) mediating adepolarizing efflux of chloride ions. The resulting transduction current is passed to the OSN cell body, where it drives a train of action potentials (spikes). The concentrationof detected odorant is encoded nonlinearly at each step of transduction: by a hyperbolic dependence of the number of activated receptors (R*) in the cilia (B), a stronglycooperative variation in amplitude of the transduction current (C), and similar sigmoidal variation of spike firing rate relayed by OSN axons (D). Data from [113] (C,D):response of normalized currents and firing rates of frog OSN to cineole; mammalian OSNs exhibit similar dose–response profiles.
Review Trends in Neurosciences August 2014, Vol. 37, No. 8
446
synaptic transmission
axon hillock
cellbody
dend
rite
cilia
cilia
axon
synapse(glutamate)
represent odor intensity, and which represent other infor-mation, such as odor quality?
We consider several models of odor intensity coding atthis level. Are they compatible with physiological data? Dothey predict stable perception of odor quality over a rangeof concentrations?
Although the neural code in the olfactory bulb mustrepresent both odor concentration and identity it is crucialfor the brain to disambiguate the two kinds of information.For olfactory navigation tasks, stimulus concentrationvaries with distance from a target odor source; animals
must be able to maintain a concentration-invariant repre-sentation of odor quality over biologically relevant concen-tration ranges to track the source. Although odors aregenerally thought to retain their quality over a range ofconcentrations, concentration changes greater than twoorders of magnitude may yield changes in odor qualityfor some odorants [3,31] but not others [32].
Spike rate codingGiven that odorant concentration is correlated with spikerates of OSN inputs to glomeruli, we may ask if this rate
Golfα Golfα AC3
GDP GTP
ATP
1.0
0.5
0.0
1.0
0.5
0.0
1E-3 0.01 0.1 1
1E-3 0.01 0.1 1
cAMPCa2+
Cl–
R
OH
Odorant
Transduc!oncurrent
R*
Transduc!oncurrent
Spike firing
Firin
g ra
teTr
ansd
uc!o
ncu
rren
tAc
!vat
ed re
cept
or(R
*)
Log [odorant]
Log [odorant]
[Odorant]
(A) (B)
(C)
(D)
Low
High
[Odorant]
βγ
TRENDS in Neurosciences
Figure 1. Odorant concentration coding in olfactory sensory neurons (OSNs). During sensory transduction (A), odorant molecules bind and stabilize the active states ofolfactory receptors (R) in ciliary membranes of OSNs. The activated receptors (R*) couple to G proteins (Golf) and increase synthesis of cyclic AMP (cAMP) by type IIIadenylyl cyclase (AC3). The cAMP opens cyclic nucleotide-gated channels that conduct calcium ions into the cilia and, in turn, open a channel (ANO2) mediating adepolarizing efflux of chloride ions. The resulting transduction current is passed to the OSN cell body, where it drives a train of action potentials (spikes). The concentrationof detected odorant is encoded nonlinearly at each step of transduction: by a hyperbolic dependence of the number of activated receptors (R*) in the cilia (B), a stronglycooperative variation in amplitude of the transduction current (C), and similar sigmoidal variation of spike firing rate relayed by OSN axons (D). Data from [113] (C,D):response of normalized currents and firing rates of frog OSN to cineole; mammalian OSNs exhibit similar dose–response profiles.
Review Trends in Neurosciences August 2014, Vol. 37, No. 8
446
represent odor intensity, and which represent other infor-mation, such as odor quality?
We consider several models of odor intensity coding atthis level. Are they compatible with physiological data? Dothey predict stable perception of odor quality over a rangeof concentrations?
Although the neural code in the olfactory bulb mustrepresent both odor concentration and identity it is crucialfor the brain to disambiguate the two kinds of information.For olfactory navigation tasks, stimulus concentrationvaries with distance from a target odor source; animals
must be able to maintain a concentration-invariant repre-sentation of odor quality over biologically relevant concen-tration ranges to track the source. Although odors aregenerally thought to retain their quality over a range ofconcentrations, concentration changes greater than twoorders of magnitude may yield changes in odor qualityfor some odorants [3,31] but not others [32].
Spike rate codingGiven that odorant concentration is correlated with spikerates of OSN inputs to glomeruli, we may ask if this rate
Golfα Golfα AC3
GDP GTP
ATP
1.0
0.5
0.0
1.0
0.5
0.0
1E-3 0.01 0.1 1
1E-3 0.01 0.1 1
cAMPCa2+
Cl–
R
OH
Odorant
Transduc!oncurrent
R*
Transduc!oncurrent
Spike firing
Firin
g ra
teTr
ansd
uc!o
ncu
rren
tAc
!vat
ed re
cept
or(R
*)
Log [odorant]
Log [odorant]
[Odorant]
(A) (B)
(C)
(D)
Low
High
[Odorant]
βγ
TRENDS in Neurosciences
Figure 1. Odorant concentration coding in olfactory sensory neurons (OSNs). During sensory transduction (A), odorant molecules bind and stabilize the active states ofolfactory receptors (R) in ciliary membranes of OSNs. The activated receptors (R*) couple to G proteins (Golf) and increase synthesis of cyclic AMP (cAMP) by type IIIadenylyl cyclase (AC3). The cAMP opens cyclic nucleotide-gated channels that conduct calcium ions into the cilia and, in turn, open a channel (ANO2) mediating adepolarizing efflux of chloride ions. The resulting transduction current is passed to the OSN cell body, where it drives a train of action potentials (spikes). The concentrationof detected odorant is encoded nonlinearly at each step of transduction: by a hyperbolic dependence of the number of activated receptors (R*) in the cilia (B), a stronglycooperative variation in amplitude of the transduction current (C), and similar sigmoidal variation of spike firing rate relayed by OSN axons (D). Data from [113] (C,D):response of normalized currents and firing rates of frog OSN to cineole; mammalian OSNs exhibit similar dose–response profiles.
Review Trends in Neurosciences August 2014, Vol. 37, No. 8
446
low
high[Odorant]
Information flow in olfactory receptor neurons
1. Sensory transduction to generate a graded receptor potential via cyclic nucleotide signalling.
2. Action potential generated at axon hillock if receptor potential is large enough.
3. Signal is transmitted to higher level neurons via synaptic release.
1.
2.
3.
Mainland et al., 2014

15
Transduction in the cilia of olfactory receptor neurons
http://sites.sinauer.com/neuroscience5e/animations15.01.html

16
Cilia organize channels, receptors and signalling machinery
ciliumdendrite
to cell body
odorantsreceptorion channel
+
depolarization spreads throughout dendrite+++ +
+
++ +
+ +
+

17
Cilia are microtubule-based organelles
dendriteto cell body
basal body(microtubules)
axoneme (microtubules)
cilium

18
dendriteto cell body
cilium
The transition zone regulates what enters and leaves the cilium
transition zone

19
Molecular motors transport proteins along the cilium
dendriteto cell body
cilium
kinesin
dynein

20
Name the numbered components
4
3
1 7
65
2

21
What function do these components serve?
1. transition zone
2. kinesin
3. cilium

22
Cilia have a broad biomedical relevance

Hua Jin
central nervous system
Rosenbaum & Witman, 2002
epithelial cells from kidney collecting tubule
Alamy
sperm
Science Photo Library
respiratory epithelium
olfactory receptor neuronsbrain
Motile cilia
Primary sensory cilia
Alamy
sperm
Science Photo Library
respiratory epithelium
Alamy
sperm
Science Photo Library
respiratory epitheliumMotile cilia
Alamy
sperm
Science Photo Library
respiratory epithelium
23

Cilia disorders affect most systems in the body
blindness
deafness
chronic respiratory infection
situs inversus
heart disease
infertility
obesity
cognitive dysfunction
polydactyly
kidney disease
24

25
How do we study sensory systems?

Zeynep F. Altunwww.wormatlas.org
André Karwath/Wikimedia Commons
oregonstate.edu/terra/2013/07/from-zebrafish-to-you/
http://www.healthyhomescoalition.org/mice-and-rats
26

1) Known and reproducible neural anatomy
2) Short-lifespan
3) Freeze at -80 C
4) Small, sequenced genome
5) Easy to manipulate genetics and make mutants
6) Transparent (ease of imaging)
7) Inexpensive to work with
wormatlas.org
Caenorhabditis elegans
27

28
C. elegans 60 ciliated sensory neurons sense chemical, thermal and mechanical stimuli
head tail
sensory neuron cell bodies
sensory neuron cell bodies
Michel Leroux & Tiffany Timbers
axoneme
axoneme
basalbody
basalbody

General approach to study the mechanism of sensation in C. elegans
1. Screen for abnormal sensory neuron development and function in mutants
2. Determine cellular and sub-cellular localization to infer function
3. Assess specific ciliated sensory neuron defects using synaptic and cilia markers in mutants
29

30
What genes are important for sensation?

31
Kwangjin Park & Tiffany Timbers
Socket cell
Cuticle
Sheath cell
Cilium
Dendrite
WT cilia mutant DiI
DiI
head
tail
Assay sensory neuron development and function in mutants: Dye-filling
ciliated sensoryneurons

32
0.00
0.25
0.50
0.75
1.00
0.00 0.25 0.50 0.75 1.00Proportion of amphid defects
Pro
porti
on o
f pha
smid
def
ects
20x
a b
wild-type
defective
c
Timbers et al., Figure 1
VC20615
VC20628
amphid ciliated neurons
wild-typedye-filling
defectivedye-filling
wild-typedye-filling
defectivedye-filling
phasmid ciliated neurons
480 deep-sequencedC. elegans strains fromthe multi-mutationMillion Mutation Project(MMP) collection
mixed-stageculture
each strain testedseparately (in duplicate)
x 480
staining withfluorescent diI
microscopy analysis:score amphids andphasmids separatelyfor dye-filling
plot results
amphid andphasmiddye-fill defect
amphid onlydye-fill defect
phasmid onlydye-fill defect
wild-typedye-filling
mutant C. elegans
soak in a lipophilic dye
examine under a microscope
Timbers et al., under revision at Genome Research, 2015
tail neurons
wild-type
mutant
Dye-filling procedure
Assay sensory neuron development and function in mutants: Dye-filling
head neurons
wild-type
mutant
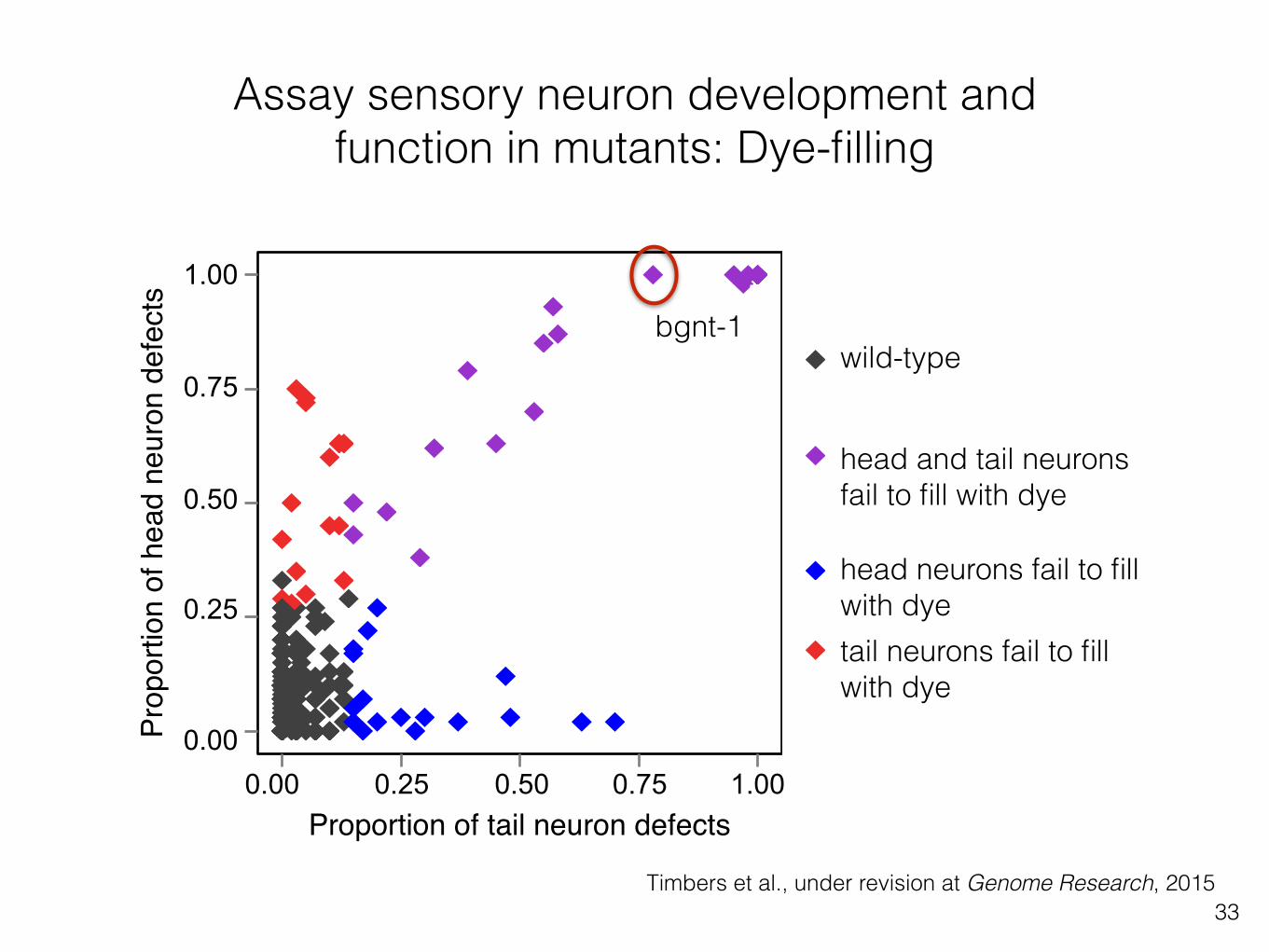
33Timbers et al., under revision at Genome Research, 2015
0.00
0.25
0.50
0.75
1.00
0.00 0.25 0.50 0.75 1.00Proportion of amphid defects
Pro
porti
on o
f pha
smid
def
ects
20x
a b
wild-type
defective
c
Timbers et al., Figure 1
VC20615
VC20628
amphid ciliated neurons
wild-typedye-filling
defectivedye-filling
wild-typedye-filling
defectivedye-filling
phasmid ciliated neurons
480 deep-sequencedC. elegans strains fromthe multi-mutationMillion Mutation Project(MMP) collection
mixed-stageculture
each strain testedseparately (in duplicate)
x 480
staining withfluorescent diI
microscopy analysis:score amphids andphasmids separatelyfor dye-filling
plot results
amphid andphasmiddye-fill defect
amphid onlydye-fill defect
phasmid onlydye-fill defect
wild-typedye-fillingwild-type
head and tail neurons fail to fill with dye
head neurons fail to fill with dyetail neurons fail to fill with dye
Proportion of tail neuron defects
Prop
ortio
n of
hea
d ne
uron
def
ects
Assay sensory neuron development and function in mutants: Dye-filling
bgnt-1

C. elegans bgnt-1 is homologous to Mammalian B3gnt1
• hydrocephalus
• Dandy-Walker malformation
• seizures
• encephalocele
• retinal dysplasia
• severe hypotonia (“floppy”)
• increased creatine kinase (CK) levels
• micropenis
• multicystic kidneys
• Mutations in human B3gnt1 lead to Walker Warburg syndrome, a disorder affecting the muscle, brain and eyes.
• Walker Warburg syndrome symptoms in patients with B3gnt1 mutations:
34Cilia disorder-related symptoms

C. elegans detect CO2 via ciliated BAG sensory neurons
Assay sensory neuron development and function in mutants: CO2 avoidance
wormatlas.org35

C. elegans avoid CO2
36
Hallem et al., 2008

The Multi-worm Tracker
Timbers et al., in preparation for PLoS Genetics, 2015
CO2 stimulus delivery
image extraction
post-experimentanalysis
37

Timbers et al., in preparation for PLoS Genetics, 2015
Assay sensory neuron development and function in mutants: CO2 avoidance
38

Timbers et al., in preparation for PLoS Genetics, 2015
CO2
Assay sensory neuron development and function in mutants: CO2 avoidance
39

Hypothesis: gcy-9 and bbs-8 are required for detecting CO2
Timbers et al., in preparation for PLoS Genetics, 2015
CO2
40

41
What role do these genes play in sensation?

42
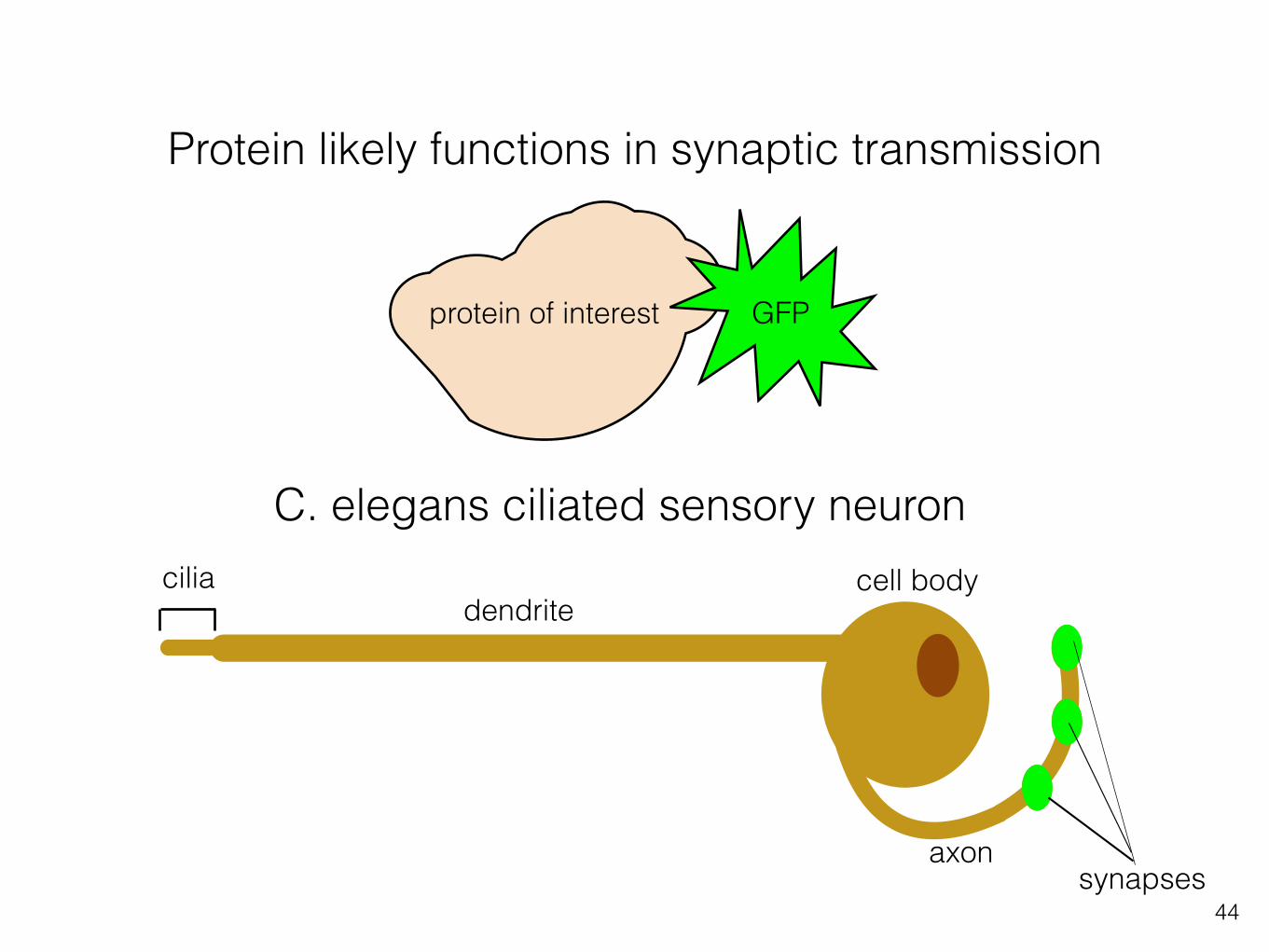
Fluorescent proteins and in vivo imaging can infer function
ciliadendrite
cell body
axonsynapses
C. elegans ciliated sensory neuron
GFPprotein of interest

43
Protein likely has functions in transduction
ciliadendrite
cell body
axonsynapses
C. elegans ciliated sensory neuron
GFPprotein of interest
44
Protein likely functions in synaptic transmission
ciliadendrite
cell body
axonsynapses
C. elegans ciliated sensory neuron
GFPprotein of interest

45
head cilia
BBS-7
BBS-7 localizes to cilia
Mohan et al., 2013
motors
cilia axoneme
basalbody

This protein localizes to cilia and synaptic endings of sensory neurons
Chunmei Li
cilia synapses
CEP neurons
cell bodiesGFP-tagged protein
46

47
What role does the protein play in the cilia and/or synapse?

48
DAF-25::GFP is cilia localized
Jensen et al., 2013

49
Olfactory transduction in C. elegans
Guanylate cyclasewild-type
basal body
Guanylate cyclasedaf-25 mutant
Dynein motor protein
Jensen et al., 2013
Ciliabasal body
wormbook.org
G-protein
GFP

What C. elegans can tell us about sensation
• What molecules participate in this process (e.g. dye-filling assay, sensory behaviour assays)
• How these molecules contribute to sensation via their expression patterns (e.g. cilia versus synapse)
• 30-40% genes in C. elegans have homologues (related genes) in humans, therefore many genes identified as important for sensation in C. elegans likely play similar roles in humans.
50

51
Summary:
How do animals sense the environment

projection to brain
olfactory receptor neuron
cilia
olfactory bulb
inhaled air
2007 Wolfers Kluwer Health | Lippincott Williams & Wilkins
Olfactory neurons sense the odourants via sensory transduction in the cilia.
This is propagated via action potentials and synaptic release to higher centres in the brain.
52

Cilia contributes to sensation by organizing molecules necessary for signal transduction in close approximation.
53dendrite
to cell body
cilium
kinesin
dynein
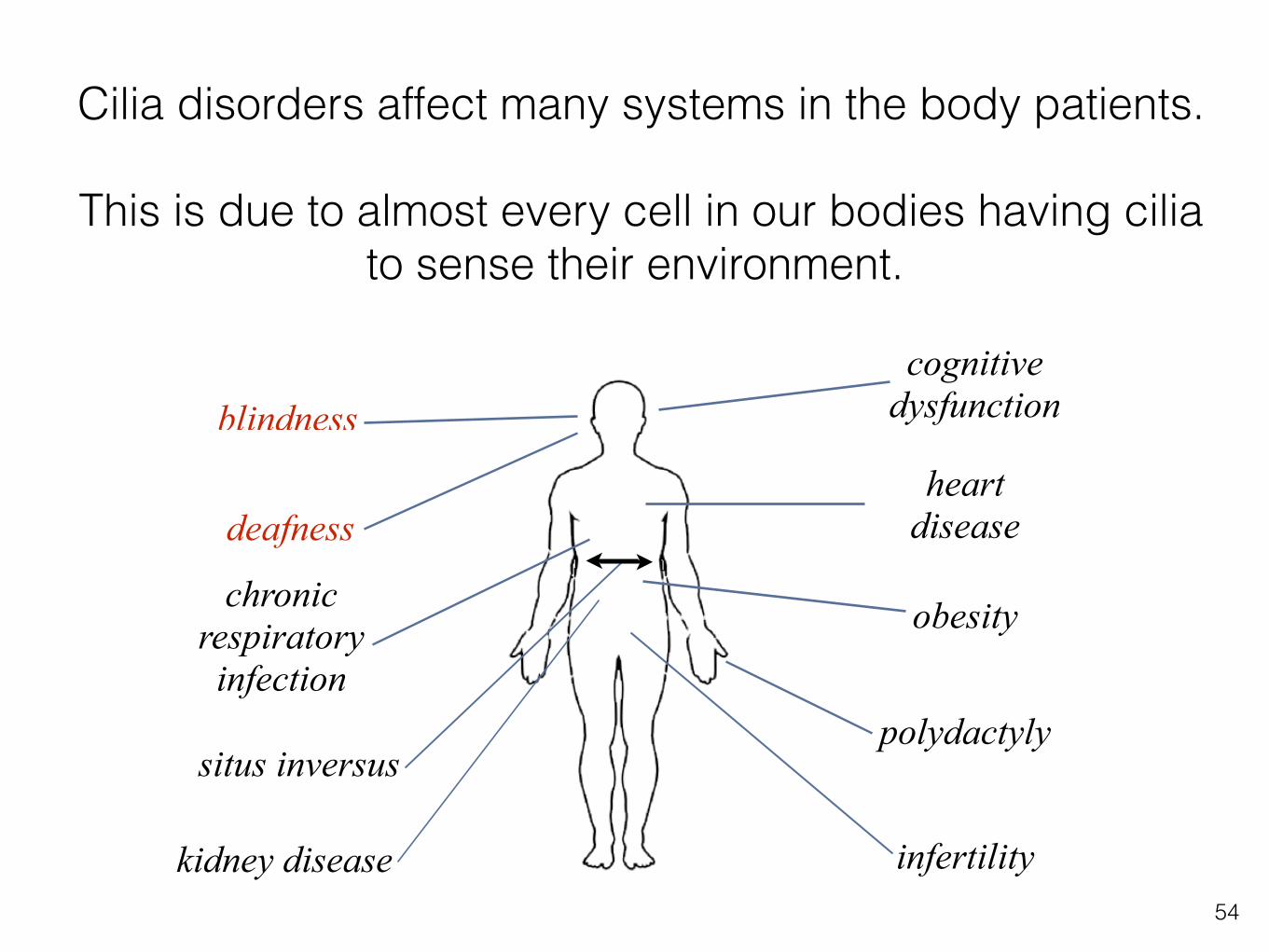
Cilia disorders affect many systems in the body patients.
This is due to almost every cell in our bodies having cilia to sense their environment.
54
blindness
deafness
chronic respiratory infection
situs inversus
heart disease
infertility
obesity
cognitive dysfunction
polydactyly
kidney disease
C. elegans is a fantastic experimental model to study how animals sense the environment.
55
Slavica Berber
Niels Ringstad/MITNiharb