hla-a and dpb1 loci confer susceptibility to grave's disease
TRANSCRIPT

HLA-A and DPB 1 Loci Confer Susceptibility to Graves' Disease
Rui-Ping Dong, Akinori Kimura, Ryoko Okubo, Hirotoshi Shinagawa, Hajime Tamai, Yasuharu Nishimura, and Takehiko Sasazuki
ABSTRACT: To investigate HLA-linked genetic factors involved in the pathogenesis of Graves' disease, 76 pa- tients and 317 healthy controls in the Japanese population were examined for HLA-A, B, C, DR, and DQ specificities by serologic typing and for HLA-DPB1 alleles by DNA typing by using the PCR-SSOP method. The frequencies of HLA-A2, B46, Cw11, and DPBI*0501 were in- creased and those of HLA-A24, B7, Bw52, and DR1 were decreased in the patients. The increased frequencies of HLA-A2 and DPB 1"0501 in the patients were statistically
significant when the corrected p value (Pc) was applied (Pc ~ 0.02 and p, ~ 0.002, respectively). ORs for a risk to develop the disease were calculated among individuals positive for DPB 1"0501 and/or HLA-A2, and the highest OR (10.5) was observed in individuals possessed both DPBI*0501 and HLA-A2. This observation suggests a synergic involvement of a HLA class II allele (DPBI*0501) and an HLA class I allele (HLA-A2) in the pathogenesis of Graves' disease. Human Immunology 35, 165-172 (1992)
ABBREVIATIONS AHVR allelic hypervariable region IDDM insulin-dependent diabetes mellitus OR odds ratio PCR polymerase chain reaction
RA RR RFLP SSOP
rheumatoid arthritis relative risk restriction fragment length polymorphism sequence-specific oligonucleotide probe
I N T R O D U C T I O N
Graves' disease is a typical organ-specific autoimmune disease and genetic factors are suggested to be involved in its pathogenesis, based on the existence of multiplex families [ 1 - 3 ] and the difference in concordant rate of the disease between monozygotic and dizygotic twins [4]. The major histocompatibility complex has been shown to control both immune responsiveness to natural antigens and susceptibility to some diseases in humans and in experimental animals [5-7] , and the association between the disease and HLAs has been reported in various ethnic groups, e.g., HLA-A1, B8, and DR3 in
From the Department of Genetics (R.P.D., A.K., R.O., H.S., Y.N., T.S.), Medical Institute of Bioregulation; and the Department of Psychoso- matic Medicine (H. T.), Faculty of Medicine, Kyushu University, Fukuoka, Japan.
Address reprint requests to Dr. T. Sasazuki, Department of Genetics, Medical Institute of Bioregulation, Kyushu University, 3-1-1 Maidashi, Higashi-ku, Fukuoka 812, Japan.
Received July 14, 1992," accepted September 18, 1992.
Caucasians [8], B46 and DR9 in Chinese [9], and A10, B8, and DQw2 in Asian Indians [10]. In the Japanese population, HLA-B35, B5 [11], B46 [12], and A l l , B46, B48, DR8, DQ4, and DPw2 [13] were re- ported to be associated with the disease. On the other hand, we have previously reported that the susceptibility to Graves' disease is controlled by at least two major genes linked to HLA and Gm, respectively [2, 14, 15]. These observations indicate the involvement of HLA- linked genetic factor(s) in the pathogenesis of Graves' disease. The HLA genes have been analyzed by serologic typing and it is known that the typing of class II HLAs, especially HLA-DP, is sometimes difficult due to the limited availability of well-defined serologic or cellular reagents, the presence of autoantibodies, and medica- tions. This difficulty can now be overcome by HLA typing at the D N A level by using the polymerase chain reaction (PCR)[16-18]. Because PCR-sequence-spe- cific oligonucleotide probe (SSOP) analysis reveals not
Human Immunology 35, 165-172 (1992) 165 © American Society for Histocompatibility and Immunogenetics, 1992 0198-8859/92/$5.00

166 R.-P. Dong et al.
only that two alleles are different, but how they differ, specific amino acid residues of the HLAs, which are critical for the susceptibility to Graves' disease, will be identified by this method.
We report here the analysis of 76 Japanese patients with Graves' disease for HLA-A, B, C, DR, and DQ by serologic typing and for the HLA-DPB1 gene by the PCR-SSOP typing to investigate further the HLA- linked genetic factor(s).
MATERIALS AND METHODS
Patients. A total of 76 unrelated Japanese patients with Graves' disease (11 men and 65 women; mean age, 32.4 years; range, 11-58 years) were the subjects of this study. Graves' disease was diagnosed on the basis of clinical symptoms, thyroid function tests, and I23I up- take. As the control population, 317 healthy unrelated Japanese were used.
Serologic typing. A panel of highly selected alloantisera in the Ninth Japan HLA Workshop or the Tenth Interna- tional Histocompatibility Workshop [19] were used to identify HLA-A, B, C, DR, and DQ specificities by using a standard complement-dependent microcytotoxicity assay [20].
D N A typing of DPBI alleles by PCR-SSOP ana- lysis. DNA was extracted from peripheral granulocytes of each subject as described previously [21]. Genomic DNA was subjected to 30 cycles of PCR in a thermal cycler (Perkin Elmer Cetus, Norwalk, CT) to amplify the second exon of DPB1 gene, using thermostable DNA
polymerase (Ampli Taq, Perkin Elmer Cetus, obtained via Takara, Kyoto). The primers and SSOPs were those used in the DNA component of the 11th International HLA Workshop [22]. Conditions of PCR and proce- dures of hybridization with SSOPs were described pre- viously [21]. The allele designations are those from WHO HLA Nomenclature Committee [23].
Statistical analysis. The frequencies of HLA-A, B, C, DR, DQ, and DPB1 alleles in 76 patients with Graves' disease were compared with those in 317 healthy unre- lated Japanese controls. Strength of the statistical associ- ation between Graves' disease and a genetic marker was expressed by the relative risk (RR) as given by Woolf [24] or by the odds ratio (OR) [25], and the statistical significance was examined by the chi-squared test with Yates' correction. The corrected p value (Pc) was calcu- lated by multiplying the p value by 108, the number of alleles tested at class I and class II loci. Linkage disequi- librium between given HLA alleles was tested by the method of Mittal [26].
RESULTS
Serologic HLA typing. Among the HLA-A, B, C, DR, and DQ specificities tested, the frequencies of A2(67.1% vs 41.6%, phenotype frequency in the patients versus that in the controls; X 2 = 15.98, p < 0.0001, RR = 2.86), B46 (21.1% vs 10.7%; X 2 -- 5.89, p < 0.02, RR = 2.22), and Cwl l (15.8% vs 7.3%; X 2 -- 5.50, p < 0.03, RR = 2.94) showed significant positive association with the disease (Table 1). The increased frequency of HLA- B46 is consistent with the previous report [12, 13],
TABLE 1 Association between Graves' disease and HLA determined by serologic typing
% Patients % Controls Relative HLA type a (n = 76) (n = 317) risk X 2 pb
A2 67.1 41.6 2.86 15.98 <0.0001 c A24 47.4 61.5 0.56 5.06 <0.03
B7 1.3 11.4 0.10 6.12 <0.02 B46 21.1 10.7 2.22 5.89 <0.02 B52 11.8 24.0 0.43 5.09 <0.03
Cwl 1 15.8 7.3 2.94 5.50 <0.03
DR1 2.6 11.4 0.21 4.39 <0.05 DR8 28.9 23.7 1.31 0.92 NS DR9 31.6 31.2 1.02 0 NS
DQ1 61.8 71.9 0.63 2.97 NS DQ3 60.5 59.0 1.07 0.06 NS
a Allele designations are those from the WHO HLA Nomenclature Committee [23]. Only the HLAs described in the text are listed.
b The statistical significance of the association between HLA and disease was determined by chi-squared analysis with Yates' correction. NS, not significant. c pC < 0.02.
HLA and Graves' Disease 167
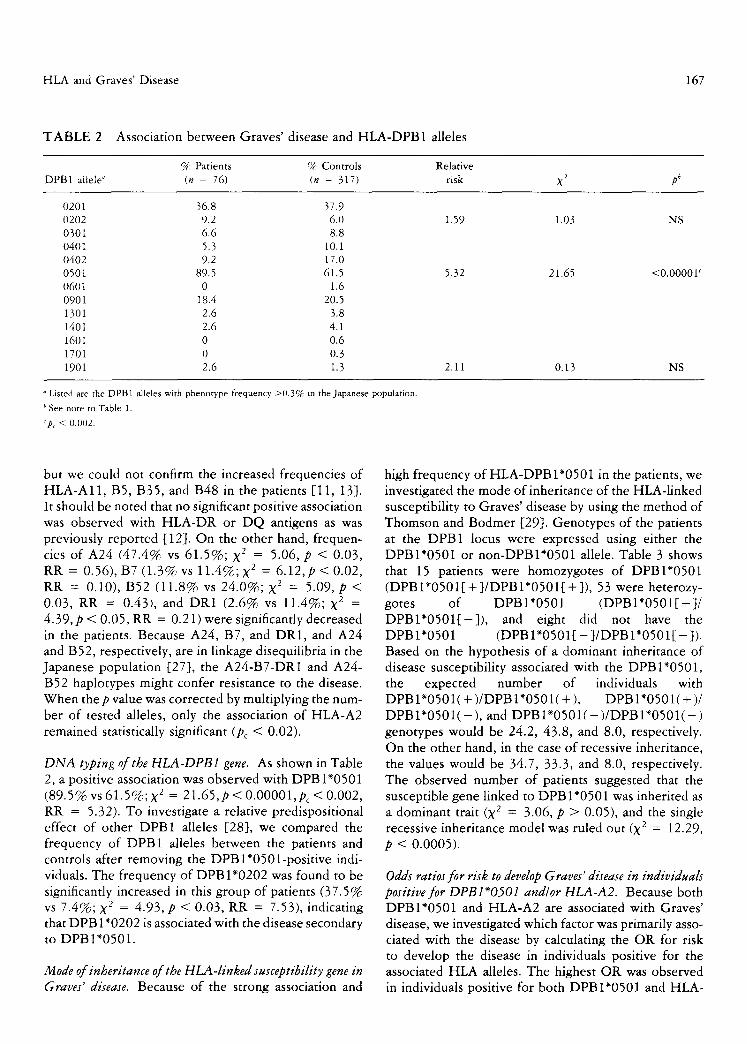
T A B L E 2 Association between Graves' disease and HLA-DPB1 alleles
% Patients % Controls Relative DPB1 allele ~ (n = 76) (n = 317) risk X 2 pb
0201 36.8 37.9 0202 9.2 6.0 1.59 1.03 NS 0301 6.6 8.8 0401 5.3 10.1 0402 9.2 17.0 0501 89.5 61.5 5.32 21.65 <0.0000Y 0601 0 1.6 0901 18.4 20.5 1301 2.6 3.8 1401 2.6 4.1 1601 0 0.6 1701 0 0.3 1901 2.6 1.3 2.11 0.13 NS
o Listed are the DPB1 alleles with phenotype frequency >0.3% in the Japanese population.
b See note to Table 1.
'p~ < 0.002.
but we could not confirm the increased frequencies of HLA-A11, B5, B35, and B48 in the patients [11, 13]. It should be noted that no significant positive association was observed with HLA-DR or DQ antigens as was previously reported [12]. On the other hand, frequen- cies of A24 (47.4% vs 61.5%; X 2 = 5.06, p < 0.03, RR = 0.56), B7 (1.3% vs 11.4%; X 2 = 6.12,p < 0.02, RR -- 0.10), B52 (11.8% vs 24.0%; X 2 -- 5.09, p < 0.03, RR -- 0.43), and DR1 (2.6% vs 11.4%; X 2 = 4.39, p < 0.05, RR -- 0.21) were significantly decreased in the patients. Because A24, B7, and DR1, and A24 and B52, respectively, are in linkage disequilibria in the Japanese population [27], the A24-B7-DR1 and A24- B52 haplotypes might confer resistance to the disease. When the p value was corrected by multiplying the num- ber of tested alleles, only the association of HLA-A2 remained statistically significant (Pc < 0.02).
DNA typing of the HLA-DPB1 gene. As shown in Table 2, a positive association was observed with DPBI*0501 (89.5% vs 61.5%; X 2 = 21.65,p < 0.00001,pc < 0.002, RR = 5.32). To investigate a relative predispositional effect of other DPB1 alleles [28], we compared the frequency of DPB1 alleles between the patients and controls after removing the DPBl*0501-posi t ive indi- viduals. The frequency of DPBI*0202 was found to be significantly increased in this group of patients (37.5% vs 7.4%; X 2 = 4.93, p < 0.03, RR = 7.53), indicating that DPB 1"0202 is associated with the disease secondary to DPBI*0501.
Mode of inheritance of the HLA-linked susceptibility gene in Graves' disease. Because of the strong association and
high frequency of HLA-DPB 1"0501 in the patients, we investigated the mode of inheritance of the HLA-linked susceptibility to Graves' disease by using the method of Thomson and Bodmer [29]. Genotypes of the patients at the DPB1 locus were expressed using either the DPBI*0501 or non-DPBl*0501 allele. Table 3 shows that 15 patients were homozygotes of DPBI*0501 ( D P B I * 0 5 0 1 [ + ] / D P B I * 0 5 0 1 [ + ] ) , 53 were heterozy- gotes of DPBI*0501 (DPBI*0501[ + ]/ D P B I * 0 5 0 1 [ - ] ) , and eight did not have the DPBI*0501 (DPB 1 " 0 5 0 1 [ - ] /DPB 1 " 0 5 0 1 [ - ]). Based on the hypothesis of a dominant inheritance of disease susceptibility associated with the DPBI*0501, the expected number of individuals with D P B I * 0 5 0 1 ( + ) / D P B 1"0501(+ ), D P BI* 0 5 0 1 (+ ) / DPBI*0501( - ), and DPB 1"0501( - ) /DPB 1"0501( - ) genotypes would be 24.2, 43.8, and 8.0, respectively. On the other hand, in the case of recessive inheritance, the values would be 34.7, 33.3, and 8.0, respectively. The observed number of patients suggested that the susceptible gene linked to DPB 1"0501 was inherited as a dominant trait (X 2 --- 3.06, p > 0.05), and the single recessive inheritance model was ruled out (X ~ -- 12.29, p < 0.0005).
Odds ratios for risk to develop Graves' disease in individuals positive for DPBl*0501 and~or HLA-A2. Because both DPBI*0501 and HLA-A2 are associated with Graves' disease, we investigated which factor was primarily asso- ciated with the disease by calculating the OR for risk to develop the disease in individuals positive for the associated HLA alleles. The highest OR was observed in individuals positive for both DPBI*0501 and HLA-

168 R.-P. Dong et al.
TABLE 3 Mode of inheritance of DPBl*0501-associated susceptibility to Graves' disease
D P B 1 " 0 5 0 1 ( + )/
D P B 1 " 0 5 0 1 ( + )
D P B 1 " 0 5 0 1 ( + )/
D P B 1 " 0 5 0 1 ( - )
D P B 1 " 0 5 0 1 ( - )/
D P B I * 0 5 0 1 ( - ) X 2 p
O b s e r v e d (n = 76) 15 53 8
E x p e c t e d Reces s ive 34 .7 33 .3 8 .0
D o m i n a n t 24 .2 43 .8 8 .0
1 2 . 2 9 < 0 . 0 0 0 5
3 .06 > 0 . 0 5
Mode of inheritance is analyzed according to Thomson and Bodmer [29] using DPB 1"0501 as the disease-linked genetic marker.
A2 (OR -- 10.5), whereas DPBl*0501-positive A2- negative or DPB 1 *0501 -negative A2-positive individu- als exhibited an OR of 4.4 or 2.2, respectively (Table 4). This observation suggests that the combination of DPBI*0501 and HLA-A2 confers the susceptibility to Graves' disease.
DISCUSSION
This study was undertaken to evaluate the possible impli- cation of HLA class I and class II genes in the susceptibil- ity to Graves' disease. It has been reported that a strong association of the disease with the HLAs was observed mainly for HLA class I and only to a lesser extent for HLA class II (HLA-DR or DQ) by serologic typing in the Japanese population [11-13]. Because the HLA class II genes can now be analyzed at the DNA level, we first examined the HLA-DQA1 and DQB1 genes in 67 pa- tients by using the PCR-SSOP method. However, we found only a weak negative association and no significant positive association between the HLA-DQ alleles and the disease (manuscript in preparation). Because there is a strong linkage disequilibrium between HLA-DR and DQ alleles [22, 27], it was suggested that the major gene controlling the susceptibility to the disease might not be within the HLA-DQ and/or DR regions. Then we
TABLE 4 High risk for Graves' disease in individuals positive for both HLA-A2 and DPBI*0501
H L A alleles Pa t i en t s ~ C o n t r o l s ~ O d d s
H L A - A 2 D P B I * 0 5 0 1 (n = 76) (n = 317 ) ra t io ~
- - 4 ( 5 . 3 % ) 84 ( 2 6 . 5 % )
+ - 4 ( 5 . 3 % ) 38 ( 1 2 . 0 % ) 2.2 - + 21 ( 2 7 . 6 % ) 101 ( 3 1 . 9 % ) 4 .4
+ + 4 7 ( 6 1 . 8 % ) 94 ( 2 9 . 7 % ) 10.5
a Individuals who have ( + ) or do not have ( - ) the HLA alleles are counted.
b Odds ratio for the risk of disease is calculated in each group by using the HLA- A2-negative and DPB 1"0501-negative group as reference.
analyzed the HLA-DPB 1 gene because it had not been tested by using the PCR-SSOP method in Graves' dis- ease. From the DPB 1 analysis, we demonstrated a strong positive association with HLA-DPB 1"0501 in the Japa- nese patients. Inoue et al. have recently analyzed the DPB1 gene of Japanese patients by Southern blot hy- bridization (restriction fragment length polymorphism [RFLP] method) and reported a weak negative associa- tion with DPw2 [13]. The HLA-DPB genes were dis- criminated by RFLP into only five subtypes [30], whereas PCR-SSOP defined virtually all DPB 1 alleles [22], which included more than 30 alleles [23]. RFLP- defined DPw2 may correspond to DPBI*0201, *0202, and "1601 [22, 30], but we could not confirm the de- creased frequency of these DPB1 alleles in this study. Inoue et al. [ 13] have not noted the change in frequency of other DPB subtypes, but RFLP-based DPB1 typing might not be able to demonstrate the positive association with DPBI*0501 because it depends on the detection of polymorphisms in the noncoding region of the DPB 1 and DPB2 genes [30], which is quite different from direct analysis of the coding region of the DPB1 gene itself by the PCR-SSOP typing reported here.
Among the associated HLA class I alleles, HLA-A2, B46, and Cwl i are known to be in linkage disequilibria in the Japanese population [27, 31]. We found that DPB 1"0501 was in linkage disequilibria with DR8-DQ 1 and DR9-DQ3 haplotypes in the Japanese population (t = 5.29 and 2.07, respectively). Because the frequen- cies of DR8, DR9, DQ1, and DQ3 did not significantly deviate among the patients, DR and DQ alleles by them- selves may not be primarily involved in the susceptibil- ity. Moreover, after the p value was corrected by the number of tested alleles (n -- 108), only the increased frequencies of HLA-A2 and DPBI*0501 remained sta- tistically significant (Pc < 0.02 and Pc < 0.002, respec- tively). These data suggest that HLA-A2 and DPBI*0501 are primarily associated with the disease, and that the association with HLA-B46 and Cwl 1 might reflect the linkage disequilibrium with HLA-A2 [27, 31]. The mode of inheritance of the susceptibility linked to DPBI*0501 was investigated by the method of

HLA and Graves' Disease 169
DPBI*0101
DPBI*0201
DPBI*0202 DPBI*0301
DPBI*0401
DPBI*0402
DPBI*0501 DPBI*0601
DPBI*0801
DPBI*0901
DPBI*I001
DPBI*II01
DPBI*I301
DPBI*I401
DPBI*I501
DPBI*I601
DPBI*I701
DPBI*I801
DPBI*I901
DPBI*2001
DPBI*2101
DPBI*2201
DPBI*2301
DPBI*2401
DPBI*2501
DPBI*2601
DPBI*2701
DPBI*2801
DPBI*2901
DPBI*3001
DPBI*3101
DPBI*3201
DPBI*3301
DPBI*3401
DPBI*3501
DPBI*3601
1o 20 30 40 5o 60 70 80 90
NYYVQGRQECYAFNGTQRFLERYIYNREEYARFDSDVGEFRAVTELGRPAAEYWNSQKDILEEK{~AVPDRVCRHNYELDEAVTLQRR
--LF ......................... FV .................. DE ............ E ...... M ....... GGPM .....
LF ......................... LV .................. E ............. E ...... M ....... GGPM---
..... L ....................... FV .................. DED ....... L ...........................
--LF ......................... F ........................................ M ....... GGPM .....
--LF ......................... FV .................. DE ................... M ....... GGPM .....
LF ......................... LV .................. E . . . . . . . . . . . . . . . . . . . . M ..............
---L ....................... FV .................. DED ....... L---E ...... M ..............
LF ......................... FV .................. DE ............ E .....................
-H-L ....................... FV .................. DED ........... E .....................
-H-L ....................... FV .................. DE ............ E .....................
---L ..................... Q ............................... L---R ...... M ..............
---L ......................................................... E ...... I ..............
-H-L ....................... FV .................. DED ....... L .........................
......................... Q ............................... L---R ...... M ....... VGPM---
LF ......................... FV .................. DE ............ E ...... M ..............
-H-L ....................... FV .................. DED ........... E ...... M ..............
........................... FV .................. DE ................... M ....... VGPMI--
LF ......................... FV .................. E ............. E . . . . . . I ..............
..... L ....................... FV .................. DED ....... L .......... M ................
..... L ....................... LV .................. E ............. E ...... M ................
--LF ......................... LV .................. E ............. E ...... M ................
--LF ......................... FV ....................................... M ....... GGPM .....
--LF ......................... F ................... E .................... M ....... GGPM .....
..... L ....................... FV .................. DE ........ L ...........................
---h .................................................................................
..... n ................................................................ M ................
--LF ......................... F ................... DE ........ L .......... M ....... VGPM---
..... L ....................... FV .................. DED ....... L---E .......................
---H-L ....................... FV .................. E ............. E ...... M ...............
--LF ......................... F ............................. L ...... L---M ................
--LF ......................... FV .................. DEV ........... E ...... M ....... GGPM .....
--LF ......................... F ................................. E ...... M ....... GGPM---
--LF ......................... LV ............................ L ...... L---M ....... VGPM---
---H-L ....................... FV .................. DED .................................
..... L ....................... LV .................. E .................... M ................
A B C D E F
FIGURE 1 Deduced amino acid sequence of the HLA- DPB 1 allele. The six allelic hypervariable regions are indicated by letters A-F. The allele designations are according to the recommendation by the WHO Nomenclature Committee [23].
Thomson and Bodmer [29], and the HLA-linked suscep- tibility was suggested to be inherited as a dominant trait. This observation is consistent with the previous reports [1-3, 14, 15].
When the highly sensitive PCR-based DNA-typing technique is used, a single base substitution could be detected and then amino acid residues at a particular position of HLA molecules associated with insulin-de- pendent diabetes mellitus (IDDM) [32] and rheumatoid arthritis (RA) [33] could be identified. We looked for the HLA-DPB 1 gene in the patients, asking whether a particular epitope (or amino acid sequence) of the HLA- DP molecule was associated with the disease, and DPBI*0501 was identified as the predominant associ-
ated allele because it conferred the highest RR and was found in 89.5% of the patients. In addition, the in- creased frequency of DPBI*0202 was observed in DPB 1"0501-negative patients, suggesting that this was the second associated allele. Figure 1 shows the deduced amino acid sequences of the first domain of the DP~ chain, including six allelic hypervariable regions (AHVRs, Fig. 1) [34]. Amino acid residue leucine (Leu) at codon 35 in the second AHVR (AHVR-B, Fig. 1) is unique to the disease-associated DPBI*0501 and DPBI*0202, in addition to the newly defined DPB1*2101, "2201, "3401, and "3601 [23]. Because the charge and hydrophobicity of Leu is not so different from that of tyrosine (Tyr) or phenylalanine (Phe) of other DPB 1 alleles, it might not confer a gross difference in the structure and peptide-binding capacity of the DP 3 chain. On the other hand, the combination of residues ofglutamic acid (Glu) and alanine (Ala) at codons 55 and 56, respectively, in the third AHVR (AHVR-C, Fig. 1) is unique to DPBI*0501, *0202, "1901, "2101, "2201, "2401, "3001, and "3601, of which only DPBI*0501,

170 R.-P. Dong et al.
*0202, and "1901 were found in the healthy Japanese individuals, and showed an RR of ~ 1.0 in the patients. Because Glu-Ala residues differ in charge from the Ala- Ala and aspartic acid (Asp)-Glu residues of the other DPB1 alleles that are not associated with the disease, Glu-Ala at codons 55 -56 may be a critical epitope for disease susceptibility via conferring the binding of dis- ease-associated peptides. It should be noted that the frequency of patients having the critical epitope reached 93.4%, whereas it was 65.0% in healthy individuals. Because DPBI*0501 had a higher incidence than DPBI*0202 and DPB1*1901, there should be an addi- tional critical epitope on the DP/3 chain. DPBI*0501 has a basic amino acid lysine (Lys) residue at codon 69 in the fourth A H V R (AHVR-D, Fig. 1), whereas DPB 1"0202 and DPB 1" 1901 have an acidic amino acid Glu residue at this position, implying that Lys at codon 69 may also be important for susceptibility. Therefore, the combination of Glu-Ala at codons 55-56 and Lys at codon 69 of the DPB1 molecule may primarily deter- mine susceptibility to Graves' disease because these amino acid residues are predicted to be pointed into the groove of HLA class II molecules and involved in bind- ing of antigenic peptides [35]. It is also interesting to note that residues 55 and 69 of the DP~ chain corre- spond to residue 57 of the DQfl chain that is suggested to be a critical residue for susceptibility to IDDM [32], and residue 70 of the DR/~ chain that may be involved in the pathogenesis of RA [33], respectively.
The increased frequency of HLA-A2 and DPBI* 0501 might reflect the linkage disequilibrium between HLA-A2 and DPBI*0501 (t = 3.2). However, DPBI* 0501 showed higher OR and incidence (OR = 4.4, phenotype frequency = 89.5%) than did HLA-A2 (OR -- 2.2, phenotype frequency -- 67.1%), and the combination of DPBI*0501 and HLA-A2 showed the highest OR (10.5) (Table 4). In addition, the frequencies of the H L A - D R / D Q haplotypes that are also in linkage disequilibrium with HLA-A2 and DPBI*0501 [27], e.g., DR8-DQ1 and DR9-DQ3, were not significantly increased in the patients. These observations imply that DPB 1"0501 is the main allele that confers susceptibility and the risk could be greatly increased in combination with HLA-A2. Although the function of the DP mole- cule has not been fully elucidated, some diseases have been found to be associated with specific DP alleles, such as juvenile chronic arthritis with DPBI*0201 [36], celiac disease with DPBI*0402 and DPBI*0301 [37], and severe aplastic anemia with DPw3 [38]. In addition, the DP molecule can present antigenic peptide to T cells [39] and may induce rejection of transplanted tissue [40] and acute graft-versus-host disease in bone marrow transplants [41]. These findings indicate that the HLA-DP molecule, like other class II molecules, plays
an important role in immune response and in susceptibil- ity to disease. Recent reports describe T-cell clones (CD4 +) reactive to an HLA-A 1-derived peptide [42] or DR3-derived peptide [43] presented by DPw3, sug- gesting that the DP molecule can bind specific HLA peptides. It may be possible that HLA-A2 is presented to CD4 + T cell as an allele-specific self-peptide in the context of the DPB 1"0501 molecule and is involved in the disease.
We have previously reported that HLA-A2, B46, Cwl 1, and DR53 were associated with susceptibility to Hashimoto's thyroiditis [44]. The associated class I HLAs are common to that found in the Graves' disease. We report here that susceptibility to Graves' disease is strongly associated with DPBI*0501 in addition to HLA-A2 and B46. Our preliminary analysis of the DPB 1 gene in Hashimoto's thyroiditis revealed an increased frequency of DPBI*0202 (12.8% vs 6.0%; RR = 2.75, p ~ 0.05) and no significant deviation in frequency of DPBI*0501 (63.8% vs 61.5%; RR -~ 1.10, p ~ 0.7) (48 patients were analyzed). Because Graves' disease and Hashimoto's thyroiditis are autoimmune diseases affecting the thyroid organ and associated with the same class I HLAs, a gene in the HLA class I loci may deter- mine the affected tissue while a gene in HLA class II loci, HLA-DPB1 itself, or a nearby gene for Graves' disease and the HLA-DRB4 (DR53) gene for Hashi- moto's thyroiditis, may differentiate the diseases.
ACKNOWLEDGMENTS
This work was supported in part by grants from the Ministry of Education, Science and Culture, Japan, and research grants from the Ministry of Health and Welfare, Japan.
REFERENCES
1. Farid NR, BarnardJM, Marshall WH, Woolfrey I, O'Dris- call RF: Thyroid autoimmune disease in a large Newfound- land family: the influence of HLA. J Clin Endocrinol Metab 45:1165, 1977.
2. Uno H, Sasazuki T, Tamai H, Matsumoto H: Two major genes, linked to HLA and Gm, control susceptibility to Graves' disease. Nature 292:768, 1981.
3. Hawkins BR, Ma JT, Lain KS, Wang CC, Yeung RT: Analysis of linkage between HLA haplotype and suscepti- bility to Graves' disease in multiple-case Chinese families in Hong Kong. Acta Endocrinol 110:66, 1985.
4. Harvald B, Hauge M: A catamnestic investigation of Dan- ish twins. Dan Med Bull 3:150, 1956.
5. McDevitt HO: Current concepts in immunology: regula- tion of the immune response by the major histocompatibil- ity system. N EnglJ Med 303:1514, 1980.
6. Todd JA, Acha-Orbea H, Bell JI, Chao N, Fronek N,

HLA and Graves' Disease 171
Jacob CO, McDermott M, Sinha AA, Timmerman L, Steinman L, McDevitt HO. A molecular basis for MHC class II-associated autoimmunity. Science 240:1003, 1988.
7. Sasazuki T, Nishimura N, Muto M, Ohta N: HLA-linked genes controlling the immune response and disease sus- ceptibility. Immunol Rev 70:51, 1983.
8. Tiwai JL, Terasaki PL: HLA and Disease Associations. New York, Springer-Verlag, 1986.
9. Yeo PPB, Chan SH, Thai AC, Ng WY, Lui KF, Wee GB, Tan SH, Lee BW, Wong HB, Cheah JS: HLA Bw46 and DR9 associations in Graves' disease of Chinese patients are age-and sex-related. Tissue Antigens 34:179, 1989.
10. Tandon N, Mehra NK, Taneja V, Vaidya MC, Kochupillai N: HLA antigens in Asian Indian patients with Graves' disease. Clin Endocrinol 33:21, 1990.
11. Kawa A, Nakamura S. Nakazawa M, Sakaguchi S, Kawa- bata T, Maeda Y, Kanehisa T: HLA-Bw35 and B5 in Japanese patients with Graves' disease. Acta Endocrinol 86:754, 1977.
12. Naito S, Sasaki H, Arakawa K: Japanese Graves' disease association with HLA-Bw46. Endocrinol Jpn 34:685, 1987.
13. Inoue D, Sato K, Enomoto T, Sugawa H, Maeda M, Inoko H, Tsuji K, Mori T, Imura H: Correlation of HLA types and clinical findings in Japanese patients with hyperthyroid Graves' disease: evidence indicating the existence of four subpopulations. Clin Endocrinol 36:75, 1992.
14. Sasazuki T, Uno H, Yasuda N, Tamai H, Matsumoto H: Evidence for HLA-linked and Gm-linked genes in Graves' disease. In Tamir BB (ed): Human Genetics, Part B. New York, Alan R Liss, 1982.
15. Tamai H, Uno H, Hirota Y, Matsubayashi S, Kuma K, Matsumoto H, Kumagai LF, Sasazuki T, Nagataki S: Im- munogenetics of Hashimoto's and Graves' disease. J Clin Endocrinol Metab 60:62, 1985.
16. Mullis KB, Faloona F: Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. Methods Enzy- mol 55:335, 1987.
17. Saiki PK, Gelfand DH, Stoffel S, Scharf SJ, Higuchi R, Horn GT, Mullis KB, Erlich HA: Primer-directed enzy- matic amplification of DNA with thermostable DNA poly- merase. Science 239:487, 1988.
18. Erlich HA, Bugawan TL: HLA DNA typing. In Innis MA, Gelfand DH, Sninsky JJ, White TJ (eds): PCR Protocols. New York, Academic, 1990.
19. Dupont B: Immunobiology of HLA: Histocompatibility Testing 1987. New York, Springer-Verlag, 1989.
20. Terasaki PI, McClellandJD: Microdroplet assay of human serum cytotoxins. Nature 204:998, 1964.
21. Dong RP, Kimura A, Numano F, Yajima M, Hashimoto Y, Kishi Y, Nishimura Y, Sasazuki T: HLA-DP antigen and Takayasu arteritis. Tissue Antigens 39:106, 1992.
22. Kimura A, Dong RP, Harada H, Sasazuki T: DNA typing
of HLA class II genes in B-lymphoblastoid cell lines homo- zygous for HLA. Tissue Antigens 40:5, 1992.
23. BodmerJG, Marsh SGE, Albert ED, Bodmer WF, Dupont B, Erlich HA, Mach B, Mayr WR, Parham P, Sasazuki T, Schreuder GMTh, Strominger JL, Svejgaard A, Terasaki PI: Nomenclature for factors of the HLA system, 1991. Tissue Antigens 39:161, 1992.
24. WoolfB: On estimating the relation between blood group and disease. Ann Hum Genet 19:251, 1955.
25. Svejgaard A, Ryder LP: HLA-genotype distribution and genetic models of insulin-dependent diabetes mellitus. Ann Hum Genet 45:293, 1981.
26. Mittal KK: The HLA polymorphism and susceptibility to disease. Vox Sang 31:161, 1976.
27. Imanishi T, Akaza T, Kimura A, Tokunaga K, Gojobori T: Allele frequencies and haplotype frequencies for HLA and complement loci in various ethnic groups. In Tsuji K, Aizawa M, Sasazuki T (eds): HLA 1991, vol 1. Oxford, Oxford University Press, 1992 (in press).
28. Payami H, Joe S, Farid NR, Stenszky V, Chan SH, Yeo PPB, Cheah JS, Thomson G: Relative predispositional effects (RPEs) of marker alleles with disease: HLA-DR alleles and Graves' disease. Am J Hum Genet 45:541, 1989.
29. Thomson G, Bodmer W: The genetic analysis of HLA and disease association. In DaussetJ, Svejgaard A (eds): HLA and Disease. Copenhagen, Munksgaard, 1977.
30. Simons MJ, Erlich HA: RFLP: sequence interrelations at the DPA and DPB loci. In Dupont B (ed): Immunobiology of HLA: Histocompatibility Testing 1987. New York, Springer-Verlag, 1989.
31. Chiewsilp P., Sujirachato K, Mayr WR, Perrier P, Hasek- ura H, Ota M, Aizawa M, Wakisaka A, Ikeda H, Naito S, Akaza T, Jeannet M, Zachary A, Braun W: Antigen soci- ety #16 report (Cwl, Cw3, Cwl0, Cw11). In Dupont B (ed): Immunobiology ofHLA: Histocompatibility Testing 1987. New York, Springer-Verlag, 1989.
32. ToddJA, BellJI, McDevitt HO: HLA-DQ 3 gene contri- butes to susceptibility and resistance to insulin-dependent diabetes mellitus. Nature 329:599, 1987.
33. Rcnningen KS, Spurkland A, Egeland T, Iwe T, Munthe E, Vartdal F, Erik E: Rheumatoid arthritis may be primarily associated with HLA-DR4 molecules sharing a particular sequence at residues 67-74. Tissue Antigens 36:235, 1990.
34. Bugawan TL, Begovich AB, Erich HA: Rapid HLA-DPB typing using enzymatically amplified DNA and nonradio- active sequence-specific oligonucleotide probes. Immuno- genetics 32:231, 1990.
35. BrownJH, Jardetzky T, Saper M, Samraoui B, Bjorkman PJ, Wiley DC: A hypothetical model for the foreign anti- gen binding site of class II histocompatibility molecules. Nature 332:845, 1988.
36. Begovich AB, Bugawan TL, Nepom BS, Klitz K, Nepom GT, Erlich HA: A specific HLA-DP/3 allele is associated

172 R.-P. Dong et al.
with pauciarticular juvenile rheumatoid arthritis but not adult rheumatoid arthritis. Proc Natl Acad Sci USA 86:9489, 1989.
37. Bugawan TL, Angelini G, Larrick J, Auricchio S, Ferrara GB, Erlich HA: A combination of a particular HLA-DP/~ allele and an HLA-DQ heterodimer confers susceptibility to coeliac disease. Nature 339:470, 1989,
38. Odum N, Platz P, Morling N, Jakobsen N, Jakobsen BK, Ryder LP, Svejgaard A: Increased frequency of HLA- DPw3 in severe aplastic anemia (AA). Tissue Antigens 29:184, 1987.
39. Celis E, Karr RW: Presentation of an immunodominant T cell epitope of hepatitis B surface antigen by the HLA- DPw4 molecule. J Virol 63:747, 1989.
40. Bonneville M, Moreau JF, Blokland E, Pool J, Mosian JP, Goulmy E, Soullilou JP: T lymphocyte cloning from
rejected kidney allograft: recognition repertoire of allore- active T cell clone. J Immunol 141:4187, 1988.
41. Odum N, Platz P, Jakobsen BK, Petersen CM, Jakobsen N, Moiler J, Ryder LP, Lamm L, Svejgaard A: HLA-DP and bone marrow transplantation: DP incompatibility and severe acute graft versus host disease. Tissue Antigens 30:213, 1987.
42. Essaket S, Fabron J, de Preval C: Corecognition of HLA- A1 and HLA-DPw3 by a human CD4 + alloreactive T lymphocyte clone. J Exp Med 172;387, 1990.
43. De Koster HS, Anderson DC, Termijtelen A: T cells sensitized to synthetic HLA-DR3 peptide give evidence of continuous presentation of denatured HLA-DR3 mole- cules by HLA-DP. J Exp Med 169:1191, 1989.
44. Honda K, Tamai H, Morita T, Kuma K, Nishimura Y, Sasazuki T: Hashimoto's thyroiditis and HLA in Japanese. J Clin Endocrinol Metab 69:1268, 1989.