hip1null_final revised ms_hmg_2-04
TRANSCRIPT

Huntingtin Interacting Protein 1 mutationslead to abnormal hematopoiesis,spinal defects and cataracts
Katherine I. Oravecz-Wilson1, Mark J. Kiel1,2, Lina Li1, Dinesh S. Rao1, Djenann Saint-Dic1,
Priti D. Kumar1, Melissa M. Provot1, Kurt D. Hankenson3, Venkat N. Reddy4,
Andrew P. Lieberman5, Sean J. Morrison1,2 and Theodora S. Ross1,*
1Department of Internal Medicine, 2Howard Hughes Medical Institute, 3Orthopaedic Research Laboratory,4Kellogg Eye Center and 5Department of Pathology, University of Michigan Medical School, Ann Arbor,
MI 48109, USA
Received December 31, 2003; Revised and Accepted February 16, 2004
Huntingtin Interacting Protein 1 (HIP1) binds clathrin and AP2, is overexpressed in multiple human tumors,and transforms fibroblasts. The function of HIP1 is unknown although it is thought to play a fundamentalrole in clathrin trafficking. Gene-targeted Hip1�/� mice develop premature testicular degeneration andsevere spinal deformities. Yet, although HIP1 is expressed in many tissues including the spleen and bonemarrow and was part of a leukemogenic translocation, its role in hematopoiesis has not been examined. Inthis study we report that three different mutations of murine Hip1 lead to hematopoietic abnormalitiesreflected by diminished early progenitor frequencies and resistance to 5-FU-induced bone marrow toxicity.Two of the Hip1 mutant lines also display the previously described spinal defects. These observationsindicate that, in addition to being required for the survival/proliferation of cancer cells and germlineprogenitors, HIP1 is also required for the survival/proliferation of diverse types of somatic cells, includinghematopoietic progenitors.
INTRODUCTION
HIP1 was originally isolated by yeast two-hybrid screening as ahuntingtin-associated protein (1,2). Since the original isolation ofHIP1, it has also been found to be part of a chromosomaltranslocation with the PDGFbR in leukemia (3). HIP1 and itsclose relative HIP1-related (HIP1r) (4) share ANTH/ENTHdomains, leucine zipper domains and TALIN homology domainsand interact with clathrin as well as other clathrin coat com-ponents (5–10). The yeast ortholog of Hip1, sla2, is essentialfor yeast cellular growth, is involved in assembly of thecytoskeleton and is required for endocytosis (11). The similaritiesin amino acid sequence and predicted domains between HIP1,HIP1r and SLA2P suggest an analogous role for HIP1 and HIP1rin the regulation of cytoskeletal and endocytic processes.
Although by homology and its interaction with clathrin, HIP1is thought to play a role in endocytosis, its actual biochemicaland physiologic role(s) are unknown. For example, expressionof HIP1 has been reported to be pro-apoptotic (12,13). Incontrast, we have found that HIP1 is overexpressed in multiple
primary human tumors (14) and that HIP1 protein is necessaryfor the survival of mouse germline progenitors (9) and manycell lines (14). Overexpression of full-length HIP1 transformsfibroblasts, and cell lines with HIP1 overexpression haveincreased levels of multiple growth factor receptors (15,16).This promotion of cell survival, proliferation and receptorsignaling maybe due to decreased degradation of growth factorreceptors as a result of altered trafficking. However, thephysiological requirement for HIP1 in normal somatic cells hasreceived only limited study.
Given that HIP1 is widely expressed, we were interested indetermining whether it was required for proliferation andsurvival in somatic cells beyond the spine (17) and testis (9). Tobegin to test this we evaluated hematopoiesis in distinct Hip1mutant mouse lines and consistently found hematopoietic aswell as spermatogenic and spine defects. All of the phenotypesmust be a result of either increased cell death, altereddifferentiation or diminished cell proliferation, suggesting thatHIP1 is necessary for maintenance of cellularity in multipletissue types including the bone marrow.
*To whom correspondence should be addressed. Tel: þ1 7346155509; Email: [email protected]
Human Molecular Genetics, 2004, Vol. 13, No. 8 851–867DOI: 10.1093/hmg/ddh102Advance Access published on March 3, 2004
Human Molecular Genetics, Vol. 13, No. 8 # Oxford University Press 2004; all rights reserved
Downloaded from https://academic.oup.com/hmg/article-abstract/13/8/851/2355752by gueston 14 February 2018

RESULTS
Targeted inactivation of Hip1
Since stable overexpression of HIP1 alters the levels of multipledifferent receptor tyrosine kinases, we wanted to test if thetargeted mutation of Hip1 in the mouse would result in pheno-typic effects in multiple different tissues. Because the Hip1 genehas a complex structure (220 kb and contains at least 30 exons),gene targeting has a chance of generating hypomorphic allelesand the phenotypic evaluation of the effects of multiple knockoutalleles is valuable. We have therefore analyzed the phenotypes ofmice that resulted from the homozygous presence of two newHip1 mutant alleles and describe hematopoietic and ophthalmicabnormalities that have not been reported previously.
First, a conditional knockout allele and its germline Crerecombined allele were generated. The latter allele has adeletion of exons 3–5 of Hip1 (see Supplementary MaterialFig. S1 for construction of targeting vector and Fig. 1 forresultant allele structures). Importantly, the neomycin cassettewas deleted along with exons 3–5. From our first targetingevent we had several correctly targeted ES cells confirmed by50, 30 and neo probes (right-hand panels of Fig. 2A and data notshown). The conditional allele was designated Hip1loxp. Therecombined allele was designated Hip1D3-5. The latter mutation
was used to generate homozygous mice to compare theirphenotypes to phenotypes observed in the other Hip1‘null’ micethat we generated (see below).
The Hip1‘null’ allele that was evaluated in greatest detail forthis report was generated serendipitously as follows. Whileattempting to knock the HIP1/PDGFbR fusion protein cDNA(3) into the Hip1 locus (see Supplementary Material Fig. S2for construction of targeting vector and Fig. 3 for resultantallele structures), we found that some of our correctly targetedES cells (Fig. 2A) did not express the HIP1/PDGFbR protein(Fig. 2B). Expression of the HIPl/PDGFbR fusion protein wastested with three different antibodies that detect the fusionprotein: anti-HIP1, anti-PDGFbR and the anti-phosphotyrosineantibody, 4G10. The last antibody recognizes the constitutivelyactive HIP1/PDGFbR (18). This analysis was facilitated by ourobservation that the Hip1 promoter is strongly active in EScells (see the detection of the endogenous murine HIP1 proteinexpressed from the non-targeted allele in panel I of Fig. 2B).One of the HIP1/PDGFbR non-expressing ES cells wasinjected into blastocysts and chimeric mice were generated.Since the targeted allele from the HIP1/PDGFbR targeted cellline failed to express HIP1/PDGFbR at the protein level, itoffered the prospect of an independent Hip1‘null’ allele.
To determine why a HIP1/PDGRbR fusion protein was notexpressed from the targeted allele in some of the targeted ES cell
Figure 1. Schematic of the structure and probes for Hip1loxp and Hip1D3-5 alleles. The screening strategy involved digesting the genomic DNA with EcoRI orBamH1 and Southern blotting with either the 50 probe, to yield a 16.5 kb wt allele and a 15.7 kb recombinant allele, or the 30 probe to yield a 12.0 kb wild typeallele or a 7.0 kb recombinant allele, respectively. To determine if there was further recombination in the CMV-cre genetic background, the genomic DNA was alsoprobed for the neomycin cassette and its absence was diagnostic of recombination as confirmed by western blot (Fig. 2D). Abbreviations: Ap¼ApaI; B¼BamHI;E¼EcoRI; H¼HindIII; Hp¼HpaI; K¼KpnI; RV¼EcoRV; S¼ SacI; Sp¼ SpeI; Sph¼ SphI; X¼XbaI; Xh¼XhoI.
852 Human Molecular Genetics, 2004, Vol. 13, No. 8
Downloaded from https://academic.oup.com/hmg/article-abstract/13/8/851/2355752by gueston 14 February 2018
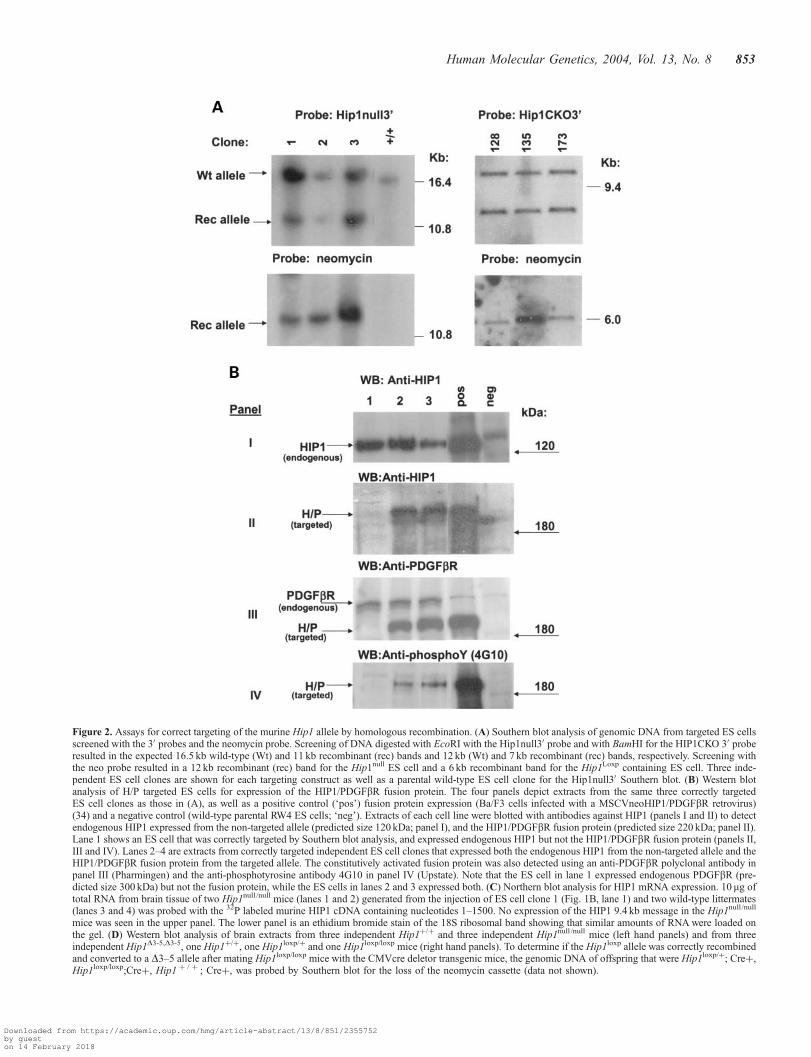
Figure 2. Assays for correct targeting of the murine Hip1 allele by homologous recombination. (A) Southern blot analysis of genomic DNA from targeted ES cellsscreened with the 30 probes and the neomycin probe. Screening of DNA digested with EcoRI with the Hip1null30 probe and with BamHI for the HIP1CKO 30 proberesulted in the expected 16.5 kb wild-type (Wt) and 11 kb recombinant (rec) bands and 12 kb (Wt) and 7 kb recombinant (rec) bands, respectively. Screening withthe neo probe resulted in a 12 kb recombinant (rec) band for the Hip1null ES cell and a 6 kb recombinant band for the Hip1Loxp containing ES cell. Three inde-pendent ES cell clones are shown for each targeting construct as well as a parental wild-type ES cell clone for the Hip1null30 Southern blot. (B) Western blotanalysis of H/P targeted ES cells for expression of the HIP1/PDGFbR fusion protein. The four panels depict extracts from the same three correctly targetedES cell clones as those in (A), as well as a positive control (‘pos’) fusion protein expression (Ba/F3 cells infected with a MSCVneoHIP1/PDGFbR retrovirus)(34) and a negative control (wild-type parental RW4 ES cells; ‘neg’). Extracts of each cell line were blotted with antibodies against HIP1 (panels I and II) to detectendogenous HIP1 expressed from the non-targeted allele (predicted size 120 kDa; panel I), and the HIP1/PDGFbR fusion protein (predicted size 220 kDa; panel II).Lane 1 shows an ES cell that was correctly targeted by Southern blot analysis, and expressed endogenous HIP1 but not the HIP1/PDGFbR fusion protein (panels II,III and IV). Lanes 2–4 are extracts from correctly targeted independent ES cell clones that expressed both the endogenous HIP1 from the non-targeted allele and theHIP1/PDGFbR fusion protein from the targeted allele. The constitutively activated fusion protein was also detected using an anti-PDGFbR polyclonal antibody inpanel III (Pharmingen) and the anti-phosphotyrosine antibody 4G10 in panel IV (Upstate). Note that the ES cell in lane 1 expressed endogenous PDGFbR (pre-dicted size 300 kDa) but not the fusion protein, while the ES cells in lanes 2 and 3 expressed both. (C) Northern blot analysis for HIP1 mRNA expression. 10 mg oftotal RNA from brain tissue of two Hip1null /null mice (lanes 1 and 2) generated from the injection of ES cell clone 1 (Fig. 1B, lane 1) and two wild-type littermates(lanes 3 and 4) was probed with the 32P labeled murine HIP1 cDNA containing nucleotides 1–1500. No expression of the HIP1 9.4 kb message in the Hip1null /null
mice was seen in the upper panel. The lower panel is an ethidium bromide stain of the 18S ribosomal band showing that similar amounts of RNA were loaded onthe gel. (D) Western blot analysis of brain extracts from three independent Hip1þ/þ and three independent Hip1null /null mice (left hand panels) and from threeindependent Hip1D3-5,D3-5, one Hip1þ/þ, one Hip1loxp/þ and one Hip1loxp/loxp mice (right hand panels). To determine if the Hip1loxp allele was correctly recombinedand converted to a D3–5 allele after mating Hip1loxp/loxp mice with the CMVcre deletor transgenic mice, the genomic DNA of offspring that were Hip1loxp/þ; Creþ,Hip1loxp/loxp;Creþ, Hip1 þ / þ ; Creþ, was probed by Southern blot for the loss of the neomycin cassette (data not shown).
Human Molecular Genetics, 2004, Vol. 13, No. 8 853
Downloaded from https://academic.oup.com/hmg/article-abstract/13/8/851/2355752by gueston 14 February 2018

Figure 2. Continued.
Figure 3. Schematic of the structures and probes for the H/P knockin targeting vector and actual recombinant Hip1null allele. Exons 2–7 were deleted in the target-ing vector as well as the deletion ‘null’ mutant. ‘bGHpA’ is the bovine growth hormone polyadenylation signal. Abbreviations: B¼BamHI; E¼EcoRI;H¼HindIII; Hp¼HpaI; K¼KpnI; RV¼EcoRV; S¼ SacI; Sph¼ SphI; X¼XbaI; Xh¼XhoI.
854 Human Molecular Genetics, 2004, Vol. 13, No. 8
Downloaded from https://academic.oup.com/hmg/article-abstract/13/8/851/2355752by gueston 14 February 2018

lines (Fig. 2B), the sequence of the genomic locus for themutant Hip1 allele was analyzed. To do this a genomic libraryfrom the blastocyst injected ES cell DNA (represented in lanes 1of Fig. 2A and B) was created and two clones were isolated thatcontained the human HIP1/PDGRbR sequences. The human–mouse junction in both clones was sequenced and comparedwith mouse genomic sequence surrounding exon 2. A 498 bpdeletion of mouse genomic sequences was found in the regiontargeted by the knockin vector (Supplementary MaterialFig. S3). This placed the human HIP1/PDGFbR cDNA inmurine intron 2 instead of being fused in frame with murineexon 2 (Fig. 3). The rest of the Hip1 allele was intact. Thisdeletion predicts a frameshift mutation in the Hip1 proteinproduct, as murine exon 1 is fused to the human HIP1/PDGFbRcDNA out of frame. Henceforth we refer to the previouslydescribed Hip1 allele (9) that replaced exons 2–8 with theneomycin cassette as ‘�’ and the targeted Hip1 allele describedin detail here as ‘null’.
Chimeric mice generated from the ES cells with theserendipitous Hip1‘null’ allele as well as the Hip1loxp allelewere mated with C57BL/6 females and F1 agouti pups weregenotyped by Southern blot analysis. The F1 heterozygotes forthe different alleles were then intercrossed to generate F2
animals. The numbers of Hip1null/null mice from this particularintercross were significantly decreased compared with theexpected Mendelian ratios (55% hets, 27% wts and 17%
homozytgotes; n¼ 395; P< 0.001). Genetic and pathologicevaluation of embryos from days 11–18.5 post coitum showednormal Mendelian ratios and normal appearing embryos,indicating that the partial lethality may be perinatal (data notshown). The extent of perinatal lethality increased as Hip1targeted mice were back-crossed onto a C57/BL6 background.There were no differences noted in growth rates or young adultweights among surviving mice of the F2 generation.
The brains of Hip1null/null mice did not express detectableHip1 mRNA by northern blot analysis using a pair of probesthat spanned the entire cDNA (Fig. 2C). Given that Hip1null/null
mice generated from the HIP1/PDGFbR-targeted allele exhibitmore severe or penetrant phenotypes than the originallydescribed Hip1�/� mice (9), as described herein, we suggestthat the original mice may have expressed an undetectedhypomorphic allele. Consistent with the lack of a detectableHip1 mRNA, there was also no detectable HIP1 protein inthe Hip1null/null brains (Fig. 2D; left hand panel) or in multipleother tissues from both male and female Hip1null/null mice(Fig. 4). It is also noteworthy in light of our observedhematopoietic phenotype (see below) that HIP1 protein waspresent in the whole bone marrow of wild-type mice and notHip1null/null mice (Fig. 5A inset).
The F1 heterozygotes for the Hip1loxp allele, as mentionedabove, have also been intercrossed to generate F2 animals. Sofar, the expected number of Hip1loxp/loxp homozygotes has been
Figure 4. mHip1 expression in various mouse tissues from littermate Hip1null /null and Hip1þ/þ males (upper two panels) and females (lower two panels). Using 6%SDS–PAGE 100mg of protein per lane were separated and transferred to nitrocellulose. The polyclonal antibody against the 50 end of mHIP1 (UM354) was used ata dilution of 1 : 5000 with an overnight incubation at 4�C. Donkey anti-rabbit HRP secondary antibody used at 1 : 5000. For these tissues marked with asterisks,limited amount of protein was available so only 10 mg/lane were loaded on the gel. Several extracts from mouse colons had no detectable Hip1 expression. Thelatter is consistent with its lack of HIP1 expression in normal human colonic epithelium (14). The small amount of expression in some colon extracts on westernanalysis is likely to be due to the presence of non-epithelial portions of the colon such as the endothelium of blood vessels and enteric nervous tissue.
Human Molecular Genetics, 2004, Vol. 13, No. 8 855
Downloaded from https://academic.oup.com/hmg/article-abstract/13/8/851/2355752by gueston 14 February 2018

born and brains from the homozygous mice showed only aslight reduction in the levels of HIP1 protein (right hand panelof Fig. 2D). When these mice were then mated with thetransgenic CMVcre deletor mice obtained from JacksonLaboratories [TgN(hCMV-cre)140Sau; stock number 002471],we were fortunate to obtain some Fl heterozygous mice withgermline recombination to generate the Hip1D3–5 allele (seeFig. 1 for structure; this allele was identified by Southern blotanalysis for loss of the neomycin cassette). F2 mice withhomozygous recombined alleles (Hip1D3–5/D3–5) have been
generated and have no detectable murine HIP1 protein (righthand panel of Fig. 2D). Our expansion of the Hip1D3–5 colonyhas allowed us to accumulate phenotypic data for comparisonto the observed phenotypes in the Hip1null/null mice (see belowand Table 1). Interestingly, in contrast to the Hip1null/null mice,to date the Hip1D3–5/D3–5 mice have been born in Mendelianratios (26% wt, 49% hets and 24%; n¼ 119; P¼ 0.8) with noevidence of the predicted partial lethality of the Hip1null/null
mice. Potential reasons for this difference include geneticbackground effects, the possibility that the Hip1D3–5 allele is
Figure 5. Response of Hip1null /null mice to 5-FU treatment in vivo demonstrates diminished sensitivity to toxic injury. (A) Western blot analysis of mHip1 expres-sion in whole bone marrow extracts and survival after sequential 5-FU treatment. Whole bone marrow from three wild-type and three Hip1null /null mice were com-bined and run on 6% PAGE as in Figure 2 and analyzed for HIP1 expression using the HIPl/lB11 monoclonal antibody (inset). 5-FU was administered weekly at adose of 150 mg/kg, and survival was monitored (n¼ 10 per genotype). Results were analyzed with a log-rank non-parametric test and expressed as Kaplan–Meiersurvival curves (P< 0.003). At the end of the experiment, 67% of Hip1null /null mice survived as compared with none of the wild-type littermates. This experimentwas repeated twice with similar results. (B) Complete blood counts (CBC) during 5-FU treatment. ‘Pre-bleed’ was done prior to the first 5 FU treatment, andthereafter at day 14 and 28 before each weekly treatment. At the ‘pre-bleed’ time point all mice had normal WBC counts (11 000–12 000/ml). By 14 days post5-FU treatment, all the animals developed leukopenia with counts ranging from 2000–7000/ml. By 21 days, all the Hip1þ/þ mice had WBC counts less then1000/ml. The Hip1null /null blood counts remained normal. The platelet counts followed the same pattern as the WBC counts with respect to genotype. Mice ofall genotypes had platelet counts in the normal range at the pre-bleed time point (606 000–797 000/ml). The platelet count from the Hip1þ/þ mice fell dramaticallyat 14 days, while the Hipnull /null maintained almost normal platelet counts throughout the entire time course.
Table 1. Phenotypic comparison of the Hip1 mutant mice
Phenotype Hip1 mutation
Metzler et al. (17) �/� (9) null/null D3–5/D3–5
Testicular degeneration ND þ þþ þþMale fertility ND Fertile Infertile InfertileHematopoietic defects ND 50% 85% ND5-FU resistant ND ND Yes YesHunchback 85% ND 100% 80%Cataracts ND No Yes No
ND, no data available. %¼ percentage mice with phenotype by 1 year of age.
856 Human Molecular Genetics, 2004, Vol. 13, No. 8
Downloaded from https://academic.oup.com/hmg/article-abstract/13/8/851/2355752by gueston 14 February 2018

hypomorphic or that the Hip1null allele, but not the Hip1D3–5
allele, affects expression of neighboring genes that are partiallynecessary for perinatal survival of mice.
Hip1 mutant mice exhibit lower numbers of primitivebut not more mature hematopoietic progenitors in vitro
In addition to finding HIP1 expressed at the protein level inwhole bone marrow, we have found that Hip1 mRNA isexpressed at similar levels in both bone marrow andhematopoietic stem cells (HSCs; Supplementary MaterialFig. S4). These expression data together with the fact that thehuman Hip1 allele was identified as part of a oncogenictranslocation in leukemia (3) and Hip1 is located at chromo-some 7q11.2, a region frequently deleted in hematopoieticmalignancies (19), suggested that HIP1 may have a role inhematopoiesis. Because of this, we originally evaluatedperipheral blood counts and in vitro hematopoiesis in the
original group of Hip1�/� mice and found significanthematopoietic abnormalities in vitro (Fig. 6). We thereforealso evaluated hematopoiesis in the new Hip1null/null mice.Neither the Hip1�/� nor the Hip1null/null mice had differencesin peripheral blood cell counts relative to control littermates(data not shown). To evaluate whether Hip1 deficiency affectedhematopoietic progenitors, bone marrow cells obtained fromHip1�/� mice, HIP1null/null mice as well as heterozygous andwild-type littermates corresponding to each line were culturedin methylcellulose. Since different types of hematopoieticprogenitors form distinct colonies in methylcellulose, we wereable to assay whether there was a difference between these micein the frequencies of various types of progenitors (20,21).There was no difference between either type of HIP1-deficientmice and control heterozygous or wild-type littermates in theoverall frequency of clonogenic hematopoietic progenitors or inthe frequencies of BFU-E (erythroid progenitors) or CFU-GM(myeloid progenitors that give rise to both granulocytes and/or
Figure 6. Hip1�/� bone marrow and HSCs generate fewer primitive hematopoietic progenitor colonies in methylcellulose cultures. Data were obtained from fourindependent experiments with mice ranging from 6 weeks to 5 months of age. The frequencies of colony forming progenitors did not change with age. (A) Theoverall frequency of clonogenic progenitors and frequency of CFU-GM as a percentage of the number of cells plated (mean�SEM). Bone marrow cells from11–12 individual animals of each gentoype were plated onto methycellulose, and colonies were scored as described in the Materials and Methods. There was nosignificant difference in the overall cloning frequency (black bars) or in the frequency of CFU-GM colonies (white bars) between Hip1þ/þ, Hip1þ/� and Hip1�/
� mice. (B) The frequency of CFU-GEMM was significantly reduced in Hip1�/� mice, shown as a percentage of bone marrow cells added to culture(mean�SEM). The frequencies of CFU-GEMM in Hip1þ/þ and Hip1þ/þ mice were not statistically different. Also shown are the frequencies of CFU-GEMMamong the two subsets of Hip1�/� mice as a percentage of cells plated (mean�SEM). �/�LOW were those animals with severely reduced numbers of CFU-GEMM, constituting 50% of all �/� mice. *P< 0.05 for comparison between þ/þ and �/� animals; **P< 0.001 for comparison between þ/þ and �/�LOW. Unlike Hip1�/� mice, CFU-GEMM frequencies were not bimodally distributed among control littermates.
Human Molecular Genetics, 2004, Vol. 13, No. 8 857
Downloaded from https://academic.oup.com/hmg/article-abstract/13/8/851/2355752by gueston 14 February 2018

macrophages; Figs 6A and 7A–C). There were also nodifferences in the frequencies of a number of phenotypicallydefined lymphoid progenitor populations (including thymocytesubpopulations, pro-B, and pre-B cells) that were compared byflow-cytometry in Hip1 deficient and control littermates (datanot shown).
In contrast, there was a significant difference in the frequencyof the most primitive progenitor population that can be detectedin the methylcellulose assay, the CFU-GEMM (an earlierprogenitor with the potential to make granulocytes, erythrocytes,macrophages and megakaryocytes). In each of four independentexperiments, 50% of Hip1�/� bone marrow samples yieldednormal numbers of CFU-GEMM, while the remaining 50%exhibited severely reduced numbers of CFU-GEMM relative tolittermate controls (P< 0.0001; Fig. 6B). These data demon-strate that half of Hip1�/� mice exhibit a defect in the frequencyof primitive hematopoietic progenitors, without exhibiting anydefect in the frequencies of more mature hematopoietic pro-genitors or in peripheral blood counts. In comparison, althoughHip1null/null bone marrow produced CFU-GEMM colonies ofnormal size, the frequency of such colonies was severely reducedin 85% of mice analyzed (P< 0.0011; Fig. 7D). It is possiblethat the diminished penetrance of this phenotype in the Hip1�/�
mice compared with the Hip1null/null mice was secondary todiffering effects of background (both were mixed 129Svj/C57BL6). The other possibility is that the Hip1�/� mice werehypomorphic at the Hip1 locus and the Hip1null/null mice werenot. These data demonstrate that HIP1-deficient mice exhibit adefect in the frequency or differentiation of primitive hemato-poietic progenitors in vitro, without exhibiting any defect inthe frequency of more mature hematopoietic progenitors or inperipheral blood cell counts.
By adding the growth factors thrombopoietin and Flt-3 ligandto the methylcellulose medium, we were able to partially correctthe reduced frequency of CFU-GEMM in bone marrow from thesame Hip1null/null mice (P¼ 0.l7; Fig. 7E compared with D).One possibility is that the loss of Hip1 expression led to anincreased requirement for growth factors to promote the survivalof CFU-GEMM. This would be consistent with the observationthat stable overexpression of HIP1 increases the levels of growthfactor receptors and confers the ability to grow in reducedserum concentrations (16). An alternate possibility is thatthrombopoietin and Flt-3 allowed a distinct HIP1-independentprogenitor to form CFU-GEMM colonies.
To test whether HIP1 deficiency affects the differentiation ofHSCs in culture, HSCs were isolated by flow-cytometry fromadult bone marrow as Thy-1.1 1oSca-1þLineage�c-kitþ cells (22)and sorted into methylcellulose cultures at a density of one cellper well. The frequency of Thy-1.1 1oSca-1þLineage�c-kitþ cellsdid not differ between Hip1null/null and wild-type mice. In each oftwo independent experiments comparing three null mice withthree wild-type mice (all littermates), Hip1null/null HSCs exhibiteda similar clonogenic capacity to wild-type HSCs, with 74–80% ofHSCs forming colonies in methylcellulose. However, Hip1null/null
HSCs formed a significantly higher proportion of CFU-GMmyeloid colonies (P< 0.05) and a significantly lower proportionof more primitive CFU-GEMM colonies (P< 0.05; Fig. 7F andG). The CFU-Meg difference was not statistically significant.This suggests that Hip1null/null HSCs were less able to undergomultilineage differentiation in culture upon stimulation by the
cytokines present in the methylcellulose medium. Together, thedata suggest that growth and differentiation of HSCs, and certainother hematopoietic progenitors are regnlated by HIP1.
HIP1-deficient mice are resistant to myeloablation
As described above, Hip1null/null mice exhibited normal bloodcell counts and normal numbers of HSCs. Although Hip1null/null
bone marrow cells formed fewer CFU-GEMM colonies inculture, these data did not tell us whether this reflected areduction in the number of these progenitors in vivo or simplytheir inability to survive and form normal colonies in culture.Because of this and because HIP1 regulates growth factorreceptor signaling, we decided to further test whether HIP1deficiency alters hematopoiesis in vivo by stressing thehematopoietic system. Treatment with the cytotoxic compound5-fluorouracil (5-FU) kills many hematopoietic cells, particularlydividing progenitors. Quiescent HSCs are recruited into cycleafter 5-FU treatment to replace the blood cells that are killed(22,23). Since proliferating HSCs are more sensitive to 5-FUtoxicity, sequential treatment with 5-FU eventually leads tohematopoietic failure due to depletion of the HSC pool and theinability to regenerate lost blood cells.
To stress the mice, we treated Hip1null/null mice and wild-typelittermates with sequential weekly doses of 5-FU. Hip1null/null
mice were significantly more resistant to 5-FU-inducedhematopoietic failure than wildtype mice (Fig. 5A; P< 0.003;log rank test). As expected, 5-FU treatment strongly reducedwhite blood cell (WBC) counts in wild-type mice, and the extentof the reduction increased with successive 5-FU treatments untilall of the mice died (Fig. 5B). In the Hip1null/null mice, WBCcounts dropped after the first two 5-FU treatments, but thenrebounded to normal levels on day 28, 6 days after the third5-FU treatment. Only three of 10 Hip1null/null mice died. Thusthe WBC counts and survival data indicate that Hip1null/null micewere resistant to myeloablation by serial administration of 5-FU.It should be noted that all mice in each experiment were litter-matched males to control for effects of sex and background. Thisdemonstrates that HIP1 deficiency does alter hematopoiesisin vivo in response to stress.
To add evidence in support of the hypothesis that thehematopoietic phenotype is a direct result of HIP1 deficiency,rather than affects that are secondary to the complexity of the‘null’ allele or presence of the neomycin cassette, we have alsochallenged Hip1D3–5/D3–5 mice with 5-FU. In these HIP1-deficient mice we found a similar resistance to 5-FU-inducedbone marrow failure and death (Supplementary MaterialFig. S5).
The mechanism by which HIP1 deficiency makes hemato-poietic cells more resistant to serial treatment with 5-FU is notclear. One possibility is that reduced sensitivity of HIP1-deficient HSCs to certain growth factors precluded some ofthese cells from being recruited into cycle after 5-FU treatment.This would make the HIP1-deficient mice more resistant toserial 5-FU treatment because the reduced proliferation ofHIP1-deficient HSCs after 5-FU treatment would makesubsequent rounds of 5-FU treatment less toxic to the HSCpool. Owing to the increased lethality of HIP1-deficiency on aC57/BL6 background, bone marrow reconstitution and com-petitive repopulation experiments have not yet been possible.
858 Human Molecular Genetics, 2004, Vol. 13, No. 8
Downloaded from https://academic.oup.com/hmg/article-abstract/13/8/851/2355752by gueston 14 February 2018

Figure 7. Hip1null /null bone marrow and HSCs generate fewer primitive hematopoietic progenitor colonies in methylcellulose cultures. No statistically significantdifferences were observed between wild-type and Hipnull /null bone marrow cells in total clonogenicity (A), generation of myeloid colonies (CFU-GM; B) or gen-eration of erythroid colonies (BFU-E; C) In contrast, a marked reduction was observed in the generation of primitive colonies containing granulocytes, erythroidcells, macrophages, and megakaryocytes (CFU-GEMM; D); *P¼ 0.0011). The difference was partially reversed by addition of the early acting cytokines Flt-3 andthrombopoietin to the methylcellulose culture (E; P¼ 0.17). HSCs were isolated from whole bone marrow as Thy-1.1loSca-1þLineage c-kitþ cells and directlyplated into methylcellulose. HIP1-deficient HSCs showed a reduced ability to form primitive CFU-GEMM colonies as compared to wild-type littermate controls(F; **P< 0.05). There was a corresponding increase in the proportion of CFU-GM and CFU-Meg colonies formed from HSCs (G) but no overall difference inclonogenicity (74–80% of cells formed colonies irrespective of genotype). This suggests that HIP1 deficiency may alter the response of HSCs to growth factors,changing the way they differentiate in culture. Data for (A–D) were derived from four representative experiments (Hip1null /null, n¼ 7; Hip1þ/þ; n¼ 8). Data for (E)were derived from three representative experiments (Hip1null /null, n¼ 5; Hip1þ/þ, n¼ 6). Data for (F and G) were derived from two representative experiments(Hip1null /null, n¼ 6; Hip1þ/þn¼ 6).
Human Molecular Genetics, 2004, Vol. 13, No. 8 859
Downloaded from https://academic.oup.com/hmg/article-abstract/13/8/851/2355752by gueston 14 February 2018

Hip1-deficient male mice are infertile
The original Hip1�/� mice showed testicular degeneration (9).The new Hip1null/null mice also showed testicular degenerationwith apoptosis at the postmeiotic spermatid stage and most ofthe males were infertile. The scoring system for testiculardegeneration that was described in Table 3 of our previouswork on testicular degeneration in the HIP1-deficient back-ground was used for our evaluation of testicular degeneration(9). Consistent with the possibility that the original Hip1mutant allele was hypomorphic, the degree of testiculardegeneration in the Hip1null/null mice was qualitatively increasedcompared with the Hip1�/� mice.
To quantitate the reduced fertility, five Hip1null/null male miceand male five wild-type male mice were mated with twoproven-breeder females each (one male per cage) over a3-month period. Only 0.8 litters/female were generated in theHip1null/null cages whereas the wild-type cages generated2.4 litters/female (P< 0.001; Pearson’s chi-square test). Of theeight total litters produced from Hip1null/null males, five werederived from just one male. Hence, most Hip1null/null maleshave profoundly reduced fertility but a minority of Hip1null/null
males may have normal fertility. This suggests that there is alow level of incomplete penetrance of this phenotype. Inaddition, we have not yet observed any fertile Hip1D3–5/D3–5
male mice despite multiple attempts at mating.
Other abnormalities in the HIP1-deficient mice
As early as 4 months of age many of the Hip1null/null andHip1D3–5/D3–5 mice developed a hunched posture resulting froma severe curvature of the spine (kypholordosis) that couldreadily be observed on X-rays (Fig. 8). This phenotype hasbeen observed in another Hip1 knockout mouse that wasrecently published (17) but was not observed in the Hip1�/�
mice (9). The mechanism for this phenotype has beenhypothesized to be due to a central nervous system defectassociated with diminished AMPA receptor trafficking in theHip1 mutant background. However, it remains unclear exactlyhow this CNS defect might translate into the hunchbackphenotype. In addition, despite a vigilant search, there havebeen no morphologic abnormalities observed in the CNSassociated with Hip1 deficiency in the mice described here orin the previous reports of Hip1 knockout phenotypes (9,17).
We have found that this phenotype is more penetrant infemales. By 4 months of age, 38% (n¼ 12/33) of the observedHip1null/null females had the phenotype compared with only2.7% (n¼ 1/37) of the Hip1null/null males. There were no wild-type mice with this phenotype (n¼ 113). Approximately half ofthe females (n¼ 17) had been pregnant and were breeding atthe time of developing this back deformity, showing anincreased trend for development of this phenotype in pregnant(47%, n¼ 8/17) compared with non-pregnant female mice(28%, n¼ 5/18). This phenotype was also associated withsevere weight loss as it progressed. By one year of age, 100%of Hip1null/null mice and 0% Hip1þ/þ mice in a separateobservation group that consisted of nine litter-matched mutantand control paired mice (total n¼ 18) had the kypholordoticspine phenotype.
Histologic examination of the spines or long bones (femursand tibia) from the affected mice did not show severeosteoporosis, osteoarthritis or neurologic pathology that wouldaccount for this phenotype (data not shown). In addition, wehave stained the skeletons of Hip1null/null mice and their wild-type littermates at various ages (ranging from 2 weeks to 6months) with alizarin red (bone) and Alcian blue (cartilage)and, interestingly, found that after digesting away the soft tissueand soaking the skeletons in glycerol as part of the stainingprocedure, the curvature of the spine was partially relieved. Inaddition, the bone to cartilage ratio in these mice was notaffected by HIP1-deficiency, indicating that there was nodevelopmental or degenerative skeletal or cartilage defect. Theradiographic density of the thoracic spine, femurs and skullappeared lower in the nulls compared to the wild types (forexamples see arrows in Fig. 8A). Yet microcomputed tomo-graphy of femurs, vertebrae and ribs in nine littermate pairs ofHip1null/null (with an obvious huntchback phenotype) and wild-type mice showed no significant difference in bone densitybetween genotypes (24). This indicates that, although the bonesof Hip1null/null mice may be slightly osteopenic, they were notosteoporotic. It remains to be determined if this mild decreasein bone density is directly due to the intrinsic loss of HIP1expression or is due to a secondary effect of the hunchbackphenotype. HIP1 is highly expressed in peripheral nerves so thekypholordosis could be secondary to peripheral nerve defects,although it remains unknown whether there are primary defectsin peripheral nerve function in HIP1-deficient mice.
A variety of other mutant mice display the kypholordoticphenotype (25–29). However, these studies have shed little lighton the mechanism of the spinal abnormalities. There has beenspeculation that the phenotype represents acceleration of normalprocess associated with aging such as degenerative osteoarthritisof the intervertebral facet joints, osteoporosis, connective tissuedefects, musculoskeletal and/or neurologic defects. In thisregard, a recent report showed that activation of the tumorsuppressor gene p53 also led to a hunchback phenotype (29). Inlight of HIP1 acting as an oncoprotein (14,16), it is interestingthat the hunchback phenotype in the Hip1null/null mice isconsistent with a similar phenotype observed in a mouse withactivation of a tumor suppressor gene, such as p53.
Finally, 71% of the Hip1null/null mice had externally visiblemicro-ophthalmia (Fig. 9A). Interestingly, our Hip1�/� andHip1D3–5/D3–5 mice have not developed this phenotype. Uponcloser examination, all (100%) of the Hip1null/null mice hadsmall eyes when evaluated based on darkfield illumination ofthe dissected lens (Fig. 9B), histology (Fig. 9C) or eye weight(Fig. 9E). In addition 100% of the Hip1null/null mice had micro-lens with nuclear cataracts and associated cortical abnor-malities (Fig. 9B and C). Although the cornea, iris and retina ofHip1null/null mice were also significantly smaller than in wild-type littermates, their histological organization was normal.The eye abnormalities were observed even in mice at 3 weeksof age, suggesting that this phenotype is developmental ratherthan degenerative (Fig. 9E and data not shown). As expectedfrom this phenotype, the HIP1 protein was expressed in eyesfrom wild-type mice but not in Hip1null/null mice (Fig. 4). Thereduced size of the mutant eyes is consistent with a potentialrole for HIP1 in the survival or proliferation of cells in multipleregions of the developing eye.
860 Human Molecular Genetics, 2004, Vol. 13, No. 8
Downloaded from https://academic.oup.com/hmg/article-abstract/13/8/851/2355752by gueston 14 February 2018

A close up view of hematoxylin stained nuclei from theHip1null/null versus the wild-type lens is shown in Figure 9D. Inaddition to there being an abnormal presence of nuclei in thecells that form the central portion of the lens, we also foundthat the nuclei were condensed (see arrows) and there wasvacuolization of the cytoplasm. In comparison, the nuclei fromthe wild-type lens were only found in the epithelium and werecharacterized by intact, non-pyknotic nuclei. To work towardsan understanding of the mechanism of the lens degeneration,TUNEL assays were performed to assess if the nuclei found inthe abnormal, pyknotic lens cells (see arrows of Fig. 9D forexamples) were apoptotic. These cells were indeed TUNEL-positive (Fig. 9F), suggesting that HIP1 is not only necessary
for the appropriate differentiation of the lens epithelial cellsinto the well-organized, light-transmitting lens fibers, but alsofor their survival. However, since the homozygous presence ofthe other alleles of Hip1 does not lead to cataracts, presumablybecause those alleles are to some degree hypomorphic, itremains possible that this phenotype is related to the alterationof an undefined neighboring gene(s).
DISCUSSION
Previous studies have demonstrated that a variety of cancersoverexpress HIP1 (14) and that HIP1 is capable of trans-forming fibroblasts (16). The observed pro-growth properties
Figure 8. Kypholordotic spinal curvature of Hip1-deficient mice compared with Hip1þ/þ mice. (A) Representative photographs of 5-month-old Hip1null /null micecompared with Hip1þ/þ mice. The upper panel shows pronounced kypholordosis and reduced body mass in the Hip1null /null mice. The radiograph of the Hip1null /
null mouse in the upper panel shows the severe spinal deformity as well as a reduction of bone density in the curvature of the spine (arrow), ribs, and skull (arrow) ascompared to the Hip1þ/þ mice. (B) Radiographs of the HiplD3-5/D3-5 mice. The Hip1D3-5/D3-5 in the right hand panel shows a kyphotic deformity (45� angle for thecervical and thoracic lines) at the age of 4 months. Its wild-type littermate (left hand panel) does not have kyphosis (90� angle).
Human Molecular Genetics, 2004, Vol. 13, No. 8 861
Downloaded from https://academic.oup.com/hmg/article-abstract/13/8/851/2355752by gueston 14 February 2018

of overexpressed HIP1 probably arise from increased levels ofreceptor tyrosine kinases (15). One would therefore expect thatregulation of HIP1 levels and function would be critical in themaintenance of cellular homeostasis. In fact, analysis of whatmay have been a Hip1 hypomorphic mouse demonstrated thatfull-length HIP1 expression is required for the survival ofspermatogenic progenitors (9).
To follow this up with additional in vivo data we havegenerated two new Hip1 mutant alleles that are distinct fromthe original allele, each with their own advantages anddisadvantages. Metzler et al. (17) has constructed a distinctHip1 mutation, as well. In the latter report they show that HIP1deficiency leads to a spinal phenotype that we also describe inthis report. They describe it as a neurologic phenotype as themice develop an associated gait abnormality with muscle
wasting. On the other hand, we think that the gait abnormalityand progressive wasting may be secondary to the thoracolum-bar defects compressing on peripheral nerves and note thatHIP1-deficient mice, like the mice described here andpreviously (9,17), do not have any abnormalities in the brainor spinal cord morphology. Since our mice may have a trendtowards a worsening of their hunchback phenotype withthe physical stress of pregnancy, this also suggests that thevertebral column defects may be the primary abnormality andthe gait problems secondary. However, the actual mechanismof how the phenotype develops remains to be elucidated.
What is very interesting and relevant to our report is thatMetzler et al. (17) show evidence that Hip1 deficiency mayalter the levels of glutamate receptors in neurons cultured fromHip1 knockout mice. They show evidence that there is a
Figure 9. Hip1null /null mice have micro-ophthalmia and cataracts. (A) Representative photograph of the small eyes in the Hip1null /null mice compared with the Hip1þ/
þ mice. (B) Darkfield illumination of lenses dissected from representative Hip1null /null and wild-type mice demonstrating a cataract in the Hip1null /null eye. Note howthe lens from the null mouse has increased nuclear as well as cortical light scattering compared with the lens from the control animal. (C) Histology of the Hip1null /null
and Hip1þ/þ eyes. H/E stained cross-sections of the Hip1null /null eye showed a distinct difference in size of the entire contents of the globe with disruption in theorganization of the lens (arrows). The other structures of the eye (optic nerve, retina, cornea, and iris), although smaller in size, have a normal organization.(D) High power view of the areas of the lens boxed in (C). Note the pyknotic nuclei (arrows) in the area that forms the central portion of the lens and vacuolizationof the cytoplasm in the Hip1null /null lens. In comparison, the wild-type lens is well organized with a single layer of epithelium containing non-pyknotic nucleisurrounding the evenly placed lens fibers. (E) Weights of the eyes from Hip1null /null and Hip1þ/þ mice. Eyes were dissected from the mice, fixed in Bouins and thenweighed prior to obtaining histology. From 3 weeks of age to 5 months of age there was no significant growth in the weights of the Hip1null /null eyes. The weights ofthe Hip1þ/þ eyes were significantly larger than the Hip1null /null eyes at all ages and did grow significantly between 3 weeks and 4 months of age. (F) TUNEL assays oflens from 6-week old Hip1þ/þ and Hip1þ/þ mice, stained with NBT-BCIP and counterstained with Nuclear Fast Red. The lens of the Hip1þ/þ mice had no deepviolet-black cells. In contrast, there were large numbers of TUNEL-positive cells seen in the Hip1null /null lens (white arrows).
862 Human Molecular Genetics, 2004, Vol. 13, No. 8
Downloaded from https://academic.oup.com/hmg/article-abstract/13/8/851/2355752by gueston 14 February 2018

relatively diminished level of intracellular glutamate receptorsin the HIP1-deficient background (17). This is consistent withHip1 having a role in trafficking of multiple receptors and addsAMPA receptors to the list of receptors that may be regulatedby HIP1 (in addition to the EGFR, FGFR and PDGFbRs) (15).
In contrast to our current observations, the report by Metzleret al. (17) as well as our previous knockout report (9) did notdescribe the hematopoietic or ophthalmic defects in the HIP1-deficient background. These phenotypes may not have beenobserved because they are subtler, or the previously character-ized alleles may be hypomorphic. There could also bedifferences in background effects (all of our observations havebeen on a mixed B6/129 background) or neighboring geneeffects that could result from the various mutations.
Despite the differences between the various knockout mice,the collective data indicate that HIP1 expression is necessaryfor the normal proliferation, survival or differentiation of cellsfrom a number of somatic tissues, including the hematopoieticsystem. Analyses of the response of Hip1null/null bone marrowto growth factors in culture suggest that these HIP1-deficientcells required higher levels of cytokines that are present oradditional cytokines than wild-type cells for appropriatesurvival and differentiation. Together, with data that implicateHIP1 in the regulation of clathrin-mediated receptor trafficking,these results suggest that a variety of somatic cells, as well asgermline cells (9), and cancer cells (16), depend on HIP1 forthe efficient transduction of survival and proliferation signalsfrom ligand stimulated receptors.
Within the hematopoietic system, HIP1-deficient primitivebut not more mature hematopoietic progenitors were lessefficient at forming multilineage (CFU-GEMM) colonies inmethylcellulose. However, Hip1null/null mice appeared to havenormal numbers of HSCs based on the frequency of Thy-1.1loSca-1 þ Lineage7c-kit þ cells. Hip1null/null Thy-1.1loSca-1þ
Lineage7c-kit þ cells also formed normal numbers of coloniesin methylcellulose, but these colonies included an increasedfrequency of CFU-GM colonies and a decreased frequencyof CFU-GEMM colonies. Adding additional cytokines to themethylcellulose medium increased the frequency of CFU-GEMMcolonies formed by Hip1null/null bone marrow cells. These datasuggest that hematopoietic progenitors require HIP1 in order toexhibit normal sensitivities to cytokines and to efficiently undergomultilineage differentiation in culture.
In order to test whether hematopoiesis was altered in vivo byHIP1 deficiency, the mice were serially treated with 5-FU.Although Hip1null/null and Hip1D3-5/D3-5 mice exhibited normalblood cell counts under normal conditions, they differed fromwild-type mice in their response to weekly 5-FU treatment. TheHip1 mutant mice exhibited higher WBC and platelet counts atday 21 compared with wild-type mice and most of the Hip1mutant mice survived the sequential 5-FU treatments. Sinceserial, weekly 5-FU treatments leads to hematopoietic failureby killing the HSCs that are recruited into cycle as a result of5-FU treatment, it is likely that Hip1 mutant mice bettertolerate 5-FU treatment because their HSCs are not recruitedinto cycle in the same way as wild-type HSCs. This would alsobe consistent with Hip1null/null whole bone marrow exhibitingreduced cytokine sensitivity, as cytokine stimulation aftermyeloablation is presumably what triggers HSC activation.Taken together, the data indicate that HIP1 is required in astage-specific manner by primitive hematopoietic progenitorsin order to respond normally to cytokines in vitro and in vivo.
The selective effect of HIP1 deficiency on multipotent pro-genitors is reminiscent of the functions of the cyclin-dependentkinase inhibitors p27 (30) and p21 (31), which are also requiredby hematopoietic progenitors in a stage-specific manner. HIP1,p21 and p27 are unusual in that most proteins that have beenfound to affect hematopoiesis have widespread effects on both
Figure 9. Continued.
Human Molecular Genetics, 2004, Vol. 13, No. 8 863
Downloaded from https://academic.oup.com/hmg/article-abstract/13/8/851/2355752by gueston 14 February 2018

primitive and restricted progenitors. It will be interesting todetermine whether HIP1 selectively regulates the signaling ofreceptors that regulate p21 function in hematopoietic progenitors.
A requirement for HIP1 in cell survival, differentiation orproliferation could also explain the other phenotypes observed inthis study. The Hip1null/null mice that exhibited the most severespinal deformity had associated generalized weight loss andabsence of subcutaneous fat (Fig. 8). Moreover, the Hip1null/null
mice were consistently micro-ophthalmic and cataracts wereobserved in 100% of the Hip1null/null mice compared with afrequency of 0.5% in wild-type littermates. Interestingly, thecataract phenotype was not observed in the other Hip1 mutantmouse lines, suggesting that there is either a more completeknockout with this allele, or that there are neighboring genesthat are affected by this relatively complex allele. Since Hip1 issuch a large gene with at least 30 exons and its first intron ismore than 100 kb in size, it is expected that there could bedifficulties in generating completely null mutant alleles. Inaddition, mutations leading to neighboring gene effects resultingin a phenotype related to two or more gene alterations are alwaysa possibility, especially in a gene-rich area such as the area ofchromosome 5 that is syntenic to human 7q11.
It is unlikely that the complex HIP1 phenotype is attributableto neighboring gene effects caused by the presence of a residualneomycin cassette in some of the Hip1 mutant alleles. First, thepresence of the neomycin cassette in the Hip1loxp/loxp mice doesnot cause any of the observed phenotypes in the absence ofHIP1 deletion. Second, the Hip1D3–5/D3–5 mice exhibited thetestis, back and hematopoietic phenotypes, despite lacking aresidual neomycin cassette. The incomplete penetrance origin-ally observed in the Hip17 /7 mice could be due to the effectsof a hypomorphic allele or a background effect.
Two of the phenotypes related to homozygous Hip1mutations, hematopoietic and spinal defects, together with thechromosomal location of the human HIP1 gene to 7q11,suggests that there may be autosomal recessive mutations ofHIP1 in human patients with genetic syndromes that includehematopoietic and skeletal abnormalities. The chromosomallocus of 7q11 and its syntenic 5q region in mice is a gene-richregion and is somatically deleted in acute and chronicleukemias (19). In addition, germline mutations in patientswith Shwachman–Diamond syndrome have been mapped tohuman chromosome 7q11 (32), where HIP1 is located. Thissyndrome is autosomal recessive, and has hematopoietic andskeletal defects. Germline mutations in an open reading framedesignated SBDS (that does not correspond to HIP1) on 7q11have been described in some of these individuals (33). It will beimportant to determine if individuals that do not exhibitmutations in SBDS have germline mutations in HIP1.
MATERIALS AND METHODS
Antibodies
The 30 polyclonal HIP1 antibody, pcHIP1 and the monoclonalantibodies to HIP1, HIP1/4B10 and HIP1/1B11 have pre-viously been described (14,34). The anti-mHIP1–50 endantibody was created by injecting a glutathione sepharose-purified GST-mHIP1–50 antigen into rabbits using standard
procedures. The GST-mHIP1–50 was constructed by subcloningthe 50 end of murine HIP1 (included all sequences up to thesingle EcoRI site in murine HIP1) into pGEX 4T3.
Generation of Hip1 mutant mice
The conditional Hip1 knockout vector (pHIP1CKO) wasconstructed from two mouse genomic subclones designatedEcoB/E and Bam187. These are shown in SupplementaryMaterial Figure S1. Using these two clones, the targetingvector, pHip1CKO, was completed as follows. The most 50
subclone (subclone EcoB/E) was digested with KpnI andBamHI (50 homologous arm) and subcloned into (p98TB/KpnI-BamHI to create ‘step 1’ plasmid (5.2 kb). Subclone Bam187was then digested with EcoRV and HindIII (30 homologousarm) and subcloned into EcoRV/HindIII-digested ‘step 1’plasmid to obtain ‘step 2’ plasmid (8.7 kb). Subclone Bam187was then digested with BamHI, filled in, digested with EcoRV(conditional knockout region) and sub-cloned into HindIIIdigested and blunted 38LoxpNeo and screened for correctorientation by sequencing. This was called the ‘step 3’ plasmid(9.7 kb). The latter plasmid was digested with XhoI and SfiI,filled in with Klenow. The released insert was sub-cloned into‘step 2’ plasmid that had been digested with AscI and filled inwith Klenow. The correct orientation was confirmed bysequencing to obtain the pHip1CKO (15.5 kb).
To construct the HIP1/PDGFbR targeting vector(Supplementary Material Fig. S2), the original Hip1 knockoutvector (pHIP1KO) (9) was sequenced with a reverse primer inthe neomycin resistance cassette to choose a sense oligo 50 ofthe XhoI site in intron 1 (oligo 1). Antisense oligo 2 wasdesigned from the human HIP1 cDNA starting at nucleotide156. Sense oligo 3 was also derived from the human HIP1cDNA and started at nucleotide 157 (50-GCT GTA AAG GAAAAA CAC GCC-30) and reverse oligo 4 was derived from thehuman HIP1 cDNA starting at position 588. Subclone EcoB/E(Supplementary Material Fig. S2) was then used as a templatefor PCR using oligos 1 and 2. The 2.5 kb PCR fragment wasthen digested with XhoI. The pcDNA3H/P template (18) wasthen used as a template for generation of a 0.4 kb PCR fragmentusing oligos 3 and 4. Avector designated pGL-1/polyA/loxPneowas then digested with SalI (compatible with XhoI) and Ecl1361(blunt). A three-fragment ligation was then completed with thissticky-blunt vector and the two PCR fragments described above.The resultant vector was designated pGL-1/step5 with an insertof 5 kb. The 30 end of the H/P cDNA as an insert was thencreated by digesting pcDNA3H/P with NotI, filling in theoverhangs with Klenow and then digesting with HpaI to obtainthe 3.6 kb fragment. The pGL1/step5 vector was next digestedwith HpaI and ligated with the 3.6 kb HpaI/NotI (blunt) H/P 30
end. This was designated pGL-1/step6 with an insert size of8.6 kb. This vector was then digested with NotI and AscI andfilled in with Klenow. This generated an 8.6 kb fragment thatcontained the H/P cDNA fused at its 50 end with Hip1 murinegenomic sequence that included the 50 portion of exon 2and fused at its 30 end to a polyadenylation signal andneomycin resistance cassette (Hip1_H/P_polyA_loxPneo).The p38LoxPNeo/HIP1 (9) was digested with XhoI and SfiIto excise the already existing neomycin resistance cassette,
864 Human Molecular Genetics, 2004, Vol. 13, No. 8
Downloaded from https://academic.oup.com/hmg/article-abstract/13/8/851/2355752by gueston 14 February 2018

filled in with Klenow and then ligated with the 8.6 kb (Hipl_H/P_polyA_loxPneo) insert. The final targeting vector (18.9 kb)was designated H/P-KI/LoxPNeo. The H/P-KI/LoxPNeo vectorwas electroporated into 129SvJ RW1 ES cells, selected forG418 resistance and screened by Southern blotting for correctlytargeted clones (Fig. 3A). Generations of chimeric mice andgerm line transmission of the mutant alleles were achieved usingstandard techniques.
Preparation of genomic DNA
Mouse tails were resuspended in Lysis buffer containing 10 mM
Tris (pH 7.5–7.9), 5 mM EDTA and 0.4 M NaCl, followed byaddition of 10% SDS to the final concentration of 0.2% andproteinase K to 500 mg/ml. The cell lysis samples were shakenat 55�C overnight. The samples were then subjected to organicextraction, twice with phenol-chloroform and twice withchloroform. Cellular DNA was precipitated with 2.5 vols of100% ethanol and washed with 70% ethanol once. DNApellets, after drying, were resuspended in TE buffer and DNAconcentrations were measured by spectrophotometer.
Southern bolt analysis
For analysis of the Hip1 ‘null’ or ‘loxp’ and ‘D3–5’ alleles,10 mg of genomic DNA was digested with EcoRI or BamHIovernight and run on a 0.8% agarose Tris borate EDTA gel at80 V for 16 h. The gel, after acidic solution treatment andalkaline neutralization, was blotted onto Hybond-N filter. TheDNA bound filter was then blocked in prehybridization buffercontaining 5� SSC, 5� Denhardt’s solution, 1% SDS and100 mg/ml denatured salmon sperm DNA for 3 h at 65�C. Thehybridization was carried out overnight at 65�C with theneomycin cassette probe, the Hip1CKO 30 probe (created byPCR amplification of sequences 0.3 kb 50 of the BamHI sitethat delineates the 30 end of the Bam187 subclone;Supplementary Material Fig. S1). The forward primer sequence(RSS3CONDF) was 50-GAG GGA GCA GGC TCC TCC-30
and the reverse primer sequence (RSS3CONDR) was 50-TGGATT CAC CAT GTC GCC-30), the Hip1CKO 50 probe(generated by digestion and release of a 0.5 kb fragment fromthe zeocin resistant subclone EcoB/E with EcoRI and XbaI),Hip1 knockin 30 probe (generated by digestion of the zeocinresistant BamH/B subclone in pZero-1 with HindIII and XhoIto release a 1.3 kb fragment) or the Hip1 knockin 50, probe(generated by digestion of the EcoB/E subclone with EcoRIand XhoI to release a 0.5 kb fragment; see Figs 1 and 3 forpositions of the probes).
To prepare the probes for labeling the probe fragments wereexcised from a 1% agarose–TBE gel and DNA was extractedfrom the gel using QIAEX II agarose gel as directed by thesupplier (Qiagen). The purified 50 and 30 probes were thenlabeled with 32P using a random-primed labeling kit (Roche).After hybridization, the blot was washed stringently, twice at65�C with 2� SSC for 20 min and twice at 65�C with 1� SSCfor 10 min and twice at 65�C with 0.1�SSC for 10 min. Theblot was exposed to Kodak Biomax film overnight.
Northern blot analysis
Total RNA was isolated from mouse brain using TRIZOL(Invitrogen). RNA was electrophoresed on 1% agarose gel(Invitrogen) with 6% formaldehyde (Sigma). The gel wasstained with ethidium bromide for 30 min, and then destainedfor 3 h. The presence of sharp bands corresponding to the 18and 28 s ribosomal RNAs were confirmed by ultravioletillumination. RNA was transferred overnight to Nytran(Schleicher and Schuell) by capillary action, and the blot wasUV cross-linked and prehybridized in buffer containing5�SSC, 5�Denhardt’s solution, 1% SDS (w/v), 100 mg/mldenatured salmon sperm DNA for 3 h at 65�C. 32P labeling ofthe mHip1 (24396) probe was made by random primed labeling(Roche) with 32P-dCTP (NEN). The blot was then hybridizedovernight at 65�C, washed twice in 2� SSC for 20 min, twice in1�SSC for 10 min, twice in 0.1� SSC for 10 min, and imagedon Kodak Biomax film. The film was exposed for 4 days.
Isolation of bone marrow cells for functionalcharacterization
Bones were dissected and overlying blood vessels, muscle andfascia were thoroughly excised. Cells were flushed from eachmarrow cavity with Hank’s Buffered Salt Solution withoutcalcium or magnesium, supplemented with 2% heat-inactivatedcalf serum (HBSSþ) using a 3 ml syringe and 27G needle.Cells were triturated into single cell suspension and filteredthrough nylon screen prior to antibody staining.
Flow-cytometric isolation of hematopoietic stem cells
HSCs were isolated as previously described (22). Briefly,whole bone marrow cells from mice that had been mated withC57BL/Ka-Thy1.1 mice and selected to have the Thy1.1surface marker were incubated with a panel of unconjugatedmonoclonal antibodies to lineage specific surface markersincluding B220 (6B2), CD3 (KT31.1), CD5 (53–7.3), CD8(53–6.7), Gr-1 (8C5) and Ter119. Following dilution, pelletedcells were resuspended in anti-rat IgG specific F(ab)2 fragmentconjugated to phycoerythrin (PE). Cells were subsequentlystained with directly conjugated antibodies to Sca-1 (Ly6A/E;allophycocyanin (APC), c-kit [2B8; biotinylated (bio)],Thy-1.1 [19EX5; fluorescein-5-isothiocyanate (FITC)] Mac-1(M1/70; PE) and CD4 (GK1.5; PE). Streptavidin conjugatedto PharRed (APC-Cy7) was used to visualize c-kit. HSCswere often pre-enriched by using paramagnetic microbeads(Miltenyi Biotec) to select Sca-1þ or c-kitþ cells. Prior toflow-cytometry, cells were resuspended in 2 mg/ml 7-AAD todiscriminate live from dead cells. Only live (7-AAD negative)cells were included in the sorts and analyses. HSCs wereisolated as Thy-1.1 1oScaþ lineage�Mac- 1�CD4�c-kitþ. Allflow-cytometry was performed on a FACS Vantage (BecktonDickinson).
Lineage analysis of whole bone marrow cells byflow cytometry
Briefly, directly conjugated antibodies to B220 (6B2), Mac-1(M1/70), Gr-1 (8C5), CD3 (KT3 1.1), IgM, CD4 (GK1.5),
Human Molecular Genetics, 2004, Vol. 13, No. 8 865
Downloaded from https://academic.oup.com/hmg/article-abstract/13/8/851/2355752by gueston 14 February 2018

Thy-1.1 (19EX5) and Sca-1 (Ly6A/E) were added at appro-priate dilution to whole bone marrow cells suspended at�l� 108 cells/ml. Cells were resuspended in 2 mg/ml 7-AAD inHBSSþ prior to flow-cytometry.
Methylcellulose culture
Approximately 750–1000 live whole bone marrow cells orsingle resorted HSCs were plated per well of 96-well plates.Each well contained 100 ml 1.0% methylcellulose medium with20% charcoal absorbed fetal bovine serum, 1% BSA, 50 or150 ng/ml stem cell factor (SCF), 10 ng/ml interleukin-6 (IL-6),3 U or 9 U/ml erythropoietin (Epo) and 10 ng/ml interleukin-3(IL-3), plus or minus 10 ng/ml Flt-3 and 10 ng/ml thrombo-poietin (Tpo). Cultures were maintained at 37�C at fullyhumidified conditions in 5% CO2. BFU-E and CFU-GM werescored on days 8–10 and verified on day 12. CFU-GEMM wasscored on day 12 and verified on day 14.
Treatment of mice with 5-FU
Littermate mice of varying genotypes in the B6/129 mixedbackground (12–18 weeks of age) were injected intraperitone-ally with 5-FU (Adrucil, Pharmacia and Upjohn Co.,Kalamazoo, MI, USA; 150 mg/kg) once a week. The survivalof the mice was recorded daily and peripheral blood countswere obtained every 14 days. Mice were bled using a needlestick in the saphenous vein of the hind leg. Approximately 50–100 m1 of blood were collected from each animal. Blood countswere measured on a HEMAVET Multispecies HematologyAnalyzer (CDC Technologies, Oxford, CT, USA).
Preparation and histologic analysis of the eye
Animals were euthanized with isoflurane (Aerrane, BaxterPharmaceutical, Deerfield, II, USA) and the eyes removedintact with the optical nerve. The eyes were then fixed inBouin’s Fixative (LabChem Inc., Pittsburgh, PA, USA) for 24 hand stored in 70% ethanol. The fixed eyes were weighed(Fig. 5E), embedded in paraffin, sectioned and stained withhematoxylin and eosin (Fig. 5C and D). The TUNEL assay wasperformed after deparaffinization of the eye sections using thein situ cell death detection kit (Roche), using an alkalinephosphatase-conjugated anti-fluorescein dUTP antibody.
Methods for quantitative real time-PCR (qPCR)
Approximately 2000–10 000 cells were directly sorted into400 ml Trizol (Ambion, Austin, TX, USA) containing250 m/ml glycogen (Roche, Indianapolis, IN, USA). RNAwas extracted according to the manufacturer’s instructions. Theextracted RNA (30 ml volume) was treated for 20 min at 37�Cwith 2 ml RNase-free DNase-1 (2 U/ml; Ambion) in thepresence of 2 ml RNase-inhibitor (10 U/ml; Invitrogen). TheRNA was then purified using an RNeasy Mini Kit (Qiagen,Valencia, CA, USA) according to the manufacturer’s instruc-tions and washed three times with RNase-free water. The RNAwas used for making cDNA by reverse transcription withrandom hexamer. The cDNA was extracted with phenol-chloroform and precipitated with 20 mg glycogen. After
dissolving the cDNA with RNase-free water, cDNA equivalentto 200 cells was used for each PCR reaction. Primers weredesigned to generate short amplicons (100–150 bp). The PCRreactions were performed using a LightCycler (RocheDiagnostic Corporation) according to the manufacturer’sinstructions. The RNA content of samples compared byqPCR was normalized based on the amplification of hypox-anthine phosphoribosyl transferase (HPRT). In addition toconfirming the specificity of the qPCR reactions by examiningthe melting curves of the products, qPCR products wereseparated in 2% agarose gels to confirm the presence of a singleband of the expected size. To estimate the difference in theexpression levels of individual RNAs between samples, weassumed that one cycle difference in the timing of amplificationby qPCR was equivalent to a 1.9-fold difference in expressionlevel (90% amplification efficiency), a typical value (35).
The primers used for each qPCR assay were as follows.HPRT forward primer sequence was 50-CCTCATGGACTGA-TTATGGACA-30 and reverse primer sequence was 50-ATGT-AATTCCAGCAGGTCAGCAA-30; HIP1 forward primersequence was 50-CGGACTCAAGAGCAACAGGATG-30 andreverse primer sequence was 50-AGCCATTTCGCTTCTGAC-TGGG; HIP1r forward primer sequence was 50-GCTTACCGT-GGAGATGTTTGACTAC-30 and reverse primer sequence wasTCCTGAATGACCTGGATGAGCG-30; huntingtin forwardprimer sequence was 50-ATGGGCACACATCTCTGGAAAC-30 and reverse primer sequence was 50-TTCAGCAGGGAT-ACGGTTGACC-30.
SUPPLEMENTARY MATERIAL
Supplementary Material is available at HMG Online.
ACKNOWLEDGEMENTS
We would like to thank members of the Ross Laboratory forcritical review of the manuscript. This work was supported bya postdoctoral fellowship grant from the Huntington’s DiseaseSociety of America (D.R.), the grants R0l CA82363-01A1 andRO1 CA098730-02, ASH and the Damon Runyon Foundation(T.S.R.).
REFERENCES
1. Kalchman, M.A., Koide, H.B., McCutcheon, K., Graham, R.K., Nichol, K.,Nishiyama, K., Kazemi-Esfarjani, P., Lynn, F.C., Wellington, C.,Metzler, M. et al. (1997) HIP1, a human homologue of S. cerevisiae Sla2p,interacts with membrane-associated huntingtin in the brain. Nat. Genet.,16, 44–53.
2. Wanker, E.E., Rovira, C., Scherzinger, E., Hasenbank, R., Walter, S.,Tait, D., Colicelli, J. and Lehrach, H. (1997) HIP-I: a huntingtin interactingprotein isolated by the yeast two-hybrid system. Hum. Mol. Genet., 6,487–495.
3. Ross, T.S., Bernard, O.A., Berger, R. and Gilliland, D.G. (1998) Fusion ofHuntingtin interacting protein 1 to platelet-derived growth factor betareceptor (PDGFbetaR) in chronic myelomonocytic leukemia witht(5;7)(q33;q11.2). Blood, 91, 4419–4426.
4. Seki, N., Muramatsu, M., Sugano, S., Suzuki, Y., Nakagawara, A.,Ohhira, M., Hayashi, A., Hori, T. and Saito, T. (1998) Cloning, expressionanalysis, and chromosomal localization of HIP1R, an isolog of huntingtininteracting protein (HIP1). J. Hum. Genet., 43, 268–27l.
866 Human Molecular Genetics, 2004, Vol. 13, No. 8
Downloaded from https://academic.oup.com/hmg/article-abstract/13/8/851/2355752by gueston 14 February 2018

5. Engqvist-Goldstein, A.E., Kessels, M.M., Chopra, V.S., Hayden, M.R. andDrubin, D.G. (1999) An actin-binding protein of the S1a2/Huntingtininteracting protein 1 family is a novel component of clathrin-coated pitsand vesicles. J. Cell Biol., 147, 1503–1518.
6. Engqvist-Goldstein, A.E., Warren, R.A., Kessels, M.M., Keen, J.H.,Heuser, J. and Drubin, D.G. (2001) The actin-binding protein Hip1Rassociates with clathrin during early stages of endocytosis and promotesclathrin assembly in vitro. J. Cell Biol., 154, 1503–1518.
7. Mishra, S.K., Agostinelli, N.R., Brett, T.J., Mizukami, I., Ross, T.S. andTraub, L.M. (2001) Clathrin- and AP-2-binding sites in HIP1 uncover ageneral assembly role for endocytic accessory proteins. J. Biol. Chem.,276, 46,230–46,236.
8. Metzler, M., Legendre-Guillemin, V., Gan, L., Chopra, V., Kwok, A.,McPherson, P.S. and Hayden, M.R. (2001) HIP1 functions in clathrin-mediated endocytosis through binding to clathrin and adaptor protein 2.J. Biol. Chem., 276, 39,271–39,276.
9. Rao, D.S., Chang, J.C., Kumar, P.D., Mizukami, I., Smithson, G.M.,Bradley, S.V., Parlow, A.F. and Ross, T.S. (2001) Huntingtin interactingprotein 1 Is a clathrin coat binding protein required for differentiation oflate spermatogenic progenitors. Mol. Cell. Biol., 21, 7796–7806.
10. Waelter, S., Scherzinger, B., Hasenbank, R., Nordhoff, B., Lurz, R.,Goehler, H., Gauss, C., Sathasivam, K., Bates, G.P., Lehrach, H. et al.(2001) The huntingtin interacting protein HIP1 is a clathrin andalpha-adaptin-binding protein involved in receptor-mediated endocytosis.Hum. Mol. Genet., 10, 1807–1817.
11. Holtzman, D.A., Yang, S. and Drubin, D.G. (1993) Synthetic-lethalinteractions identify two novel genes, SLA1 and SLA2, that controlmembrane cytoskeleton assembly in Saccharomyces cerevisiae. J. Cell Biol.,122, 635–644.
12. Hackam, A.S., Yassa, A.S., Singaraja, R., Metzler, M., Gutekunst, C.A.,Gan, L., Warby, S., Wellington, C.L., Vaillancourt, J., Chen, N. et al. (2000)Huntingtin interacting protein 1 induces apoptosis via a novel caspase-dependent death effector domain. J. Biol. Chem., 275, 41,299–41,308.
13. Gervais, F.G., Singaraja, R., Xanthoudakis, S., Gutekunst, C.A.,Leavitt, B.R., Metzler, M., Hackam, A.S., Tam, J., Vaillancourt, J.P.,Houtzager, V. et al. (2002) Recruitment and activation of caspase-8 by theHuntingtin-interacting protein Hip-1 and a novel partner Hippi. Nat. CellBiol., 4, 95–105.
14. Rao, D.S., Hyun, T.S., Kumar, P.D., Mizukami, I.F., Rubin, M.A.,Lucas, P.C., Sanda, M.G. and Ross, T.S. (2002) Huntingtin-interactingprotein 1 is overexpressed in prostate and colon cancer and is critical forcellular survival. J. Clin. Invest., 110, 351–360.
15. Hyun, T.S., Rao, D.S., Saint-Dic, D., Evan Michael, L., Kumar, P.D.,Bradley, S.V., Mizukami, I.F., Oravecz-Wilson, K.I. and Ross, T.S. (2004)HIP1 and HIP1r stabilize receptor tyrosine kinases and bind3-phosphoinositides via ENTH domains. J. Biol. Chem., 279, in press.
16. Rao, D.S., Bradley, S.V., Kumar, P.D., Hyun, T.S., Saint-Dic, D.,Oravecz-Wilson, K.I., Kleer, C.G. and Ross, T.S. (2003) Altered receptortrafficking in huntingtin interacting protein 1-Transformed cells. CancerCell, 3, 471–482.
17. Metzler, M., Li, B., Gan, L., Georgiou, J., Gutekunst, C.A., Wang, Y.,Torre, E., Devon, R.S., Oh, R., Legendre-Guillemin, V. et al. (2003)Disruption of the endocytic protein HIP1 results in neurological deficitsand decreased AMPA receptor trafficking. EMBO J., 22, 3254–3266.
18. Ross, T.S. and Gilliland, D.G. (1999) Transforming properties of thehuntingtin interacting protein 1/platelet-derived growth factor beta receptorfusion protein. J. Biol. Chem., 274, 22,328–22,336.
19. Le Beau, M.M., Espinosa, R., 3rd, Davis, E.M., Eisenbart, J.D.,Larson, R.A. and Green, E.D. (1996) Cytogenetic and moleculardelineation of a region of chromosome 7 commonly deleted in malignantmyeloid diseases. Blood, 88, 1930–1935.
20. McNiece, I.K., Langley, K.E. and Zsebo, K.M. (1991) Recombinant humanstem cell factor synergises with GM-CSF, G-CSF, IL-3 and epo to stimulatehuman progenitor cells of the myeloid and erythroid lineages.Exp. Hematol., 19, 226–231.
21. Bernstein, I.D., Singer, J.W., Smith, F.O., Andrews, R.G., Flowers, D.A.,Petersens, J., Steinmann, L., Najfeld, V., Savage, D., Fruchtman, S. et al.(1992) Differences in the frequency of normal and clonal precursors ofcolony-forming cells in chronic myelogenous leukemia and acutemyelogenous leukemia. Blood, 79, 1811–1816.
22. Morrison, S.J. and Weissman, I.L. (1994) The long-term repopulatingsubset of hematopoietic stem cells is deterministic and isolatable byphenotype. Immunity, 1, 661–673.
23. Harrison, D.E. and Lerner, C.P. (1991) Most primitive hematopoietic stemcells are stimulated to cycle rapidly after treatment with 5-fluorouracil.Blood, 78, 1237–1240.
24. Hankenson, K.D., Bain, S.D., Kyriakides, T.R., Smith, E.A., Goldstein, S.A.and Bornstein, P. (2000) Increased marrow-derived osteoprogenitor cellsand endosteal bone formation in mice lacking thrombospondin 2. J. BoneMiner. Res., 15, 851–862.
25. Iba, K., Durkin, M.E., Johnsen, L., Hunziker, E., Damgaard-Pedersen, K.,Zhang, H., Engvall, E., Albrechtsen, R. and Wewer, U.M. (2001) Mice witha targeted deletion of the tetranectin gene exhibit a spinal deformity.Mol. Cell. Biol., 21, 7817–7825.
26. Brady, J.P., Garland, D.L., Green, D.E., Tamm, E.R., Giblin, F.J. andWawrousek, E.F. (2001) AlphaB-crystallin in lens development and muscleintegrity: a gene knockout approach. Invest. Ophthal. Visual Sci., 42,2924–2934.
27. Dabovic, B., Chen, Y., Colarossi, C., Zambuto, L., Obata, H. andRifkin, D.B. (2002) Bone defects in latent TGF-beta binding protein(Ltbp)-3 null mice; a role for Ltbp in TGF-beta presentation. J. Endocrinol.,175, 129–141.
28. Lawler, J., Sunday, M., Thibert, V., Duquette, M., George, E.L., Rayburn, H.and Hynes, R.O. (1998) Thrombospondin-1 is required for normal murinepulmonary homeostasis and its absence causes pneumonia. J. Clin. Invest.,101, 982–992.
29. Tyner, S.D., Venkatachalam, S., Choi, J., Jones, S., Ghebranious, N.,Igelmann, H., Lu, X., Soron, G., Cooper, B., Brayton, C. et al. (2002) p53mutant mice that display early ageing-associated phenotypes. Nature,415, 45–53.
30. Cheng, T., Rodrigues, N., Dombkowski, D., Stier, S. and Scadden, D.T.(2000) Stem cell repopulation efficiency but not pool size is governed byp27(kip1). Nat. Med., 6, 1235–1240.
31. Cheng, T., Rodrigues, N., Shen, H., Yang, Y., Dombkowski, D., Sykes, M.and Scadden, D.T. (2000) Hematopoietic stem cell quiescence maintainedby p21cip1/waf1. Science, 287, 1804–1808.
32. Popovic, M., Goobie, S., Morrison, J., Ellis, L., Ehtesham, N., Richards, N.,Boocock, G., Durie, P.R. and Rommens, J.M. (2002) Fine mapping of thelocus for Shwachman-Diamond syndrome at 7q11, identification of shareddisease haploypes, and exclusion of TPST1 as a candidate gene. Eur. J.Hum. Genet., 10, 250–258.
33. Boocock, G.R., Morrison, J.A., Popovic, M., Richards, N., Ellis, L.,Durie, P.R. and Rommens, J.M. (2003) Mutations in SBDS areassociated with Shwachman-Diamond syndrome. Nat. Genet., 33,97–101.
34. Saint-Dic, D., Chang, S.C., Taylor, G.S., Provot, M.M. and Ross, T.S.(2001) Regulation of the Src homology 2-containing inositol 5-phosphataseSHIP1 in HIP1/PDGFbeta R-transformed cells. J. Biol. Chem., 276,21,192–21,198.
35. Fink, L., Seeger, W., Ermert, L., Hanze, J., Stahl, U., Grimminger, F.,Kummer, W. and Bohle, R.M. (1998) Real-time quantitative RT–PCR afterlaser-assisted cell picking. Nat. Med., 4, 1329–1333.
Human Molecular Genetics, 2004, Vol. 13, No. 8 867
Downloaded from https://academic.oup.com/hmg/article-abstract/13/8/851/2355752by gueston 14 February 2018