high-yield production of alpha-galactosidase … · keywords: alpha-galactosidase, fungi, soy meal...
TRANSCRIPT

1620 Biotechnol. & Biotechnol. eq. 24/2010/1
Article DOi: 10.2478/v10133-010-0015-5 FB
Keywords: alpha-galactosidase, fungi, soy meal extract
Biotechnol. & Biotechnol. eq. 2010, 24(1), 1620-1623
IntroductionThe α-galactosidases (α-D-galactoside galactohydrolase, EC 3.2.1.22) are carbohydrases which catalyse hydrolysis of α-1,6-linked α-galactoside residual from simple oligosaccharides such as melibiose, raffinose, stachyose and from polymeric galactomannans (1). these enzymes are used in the hydrolysis of raffinose and stachyose present in soy beans and other leguminous food and feed that cause intestinal discomfort, flatulence and low feed utilization in monogastrites (12). In beet sugar industry, α-galactosidases are used to remove raffinose from beet molasses increasing the sucrose yield (23). α-Galactosidase is also used to improve the gelling properties of galactomannans to be used as food thickeners (5). the enzymic conversion of raffinose family oligosaccharides (RO) in soymilk may be a rational alternative to improve the nutritional quality of this low-cost, high-quality protein supplement for humans and animals (20). in the pulp and paper industry α-galactosidase could enhance the bleaching effect of β-mannanases on softwood pulp (7). In human medicine this enzyme can be used for the treatment of Fabry’s disease (3) or for the blood type conversion (25). Weignerova et al. (2009) summarizes recent advances in the use of α-galactosidases in synthetic and biotransformation applications (28).
α-Galactosidase is widely distributed in microorganisms, plants and animals. Microorganisms have the advantage of being highly active producers, and among them the α-galactosidases from filamentous fungi are the most suitable for technological
applications because of their extracellular localization, acidic pH optimum and broad stability profiles (9).
Abundant information is available on the biosynthesis of α-galactosidase from filamentous fungi belonging to the genera Penicillium and Aspergillus (1, 4, 6, 8, 9, 16, 18, 21, 23, 25, 26) during submerged fermentation.
There are a few investigations about α-galactosidase production in stationary condition of cultivation (solid-state fermentation) (22, 24, 27).
The aim of the present paper is to investigate the capacity of both fungal strains Penicillium chrysogenum ln33 and Aspergillus niger M47 to produce highly active extracellular α-galactosidases in submerged shake flask and stationary cultures as well as in laboratory bioreactor experiments using soya meal extract as nutrient medium. to our knowledge, this is the first report on production of α-galactosidase from filamentous fungus P. chrysogenum.
Materials and MethodsFungi, medium and culture conditionstwo fungal strains Penicillium chrysogenum ln33 and Aspergillus niger M47 from the national Bank for industrial Microorganisms and Cell Cultures (№ 8390 and № 2334, respectively) were used in this study. The fungal cultures were maintained on beer agar slants. Five ml of spore suspension (approximately 2×109-1010 spores/ml) were added to Erlenmeyer flasks (500 ml capacity) containing 50 ml of soya meal extract (SMe) with 5% dry content used as nutrient medium (2). The cultivation was carried out stationary as well as on a rotary shaker (220 rpm) at 30°c. in a laboratory bioreactor
HIGH-YIELD PRODUCTION OF ALPHA-GALACTOSIDASE EXCRETED FROM PENICILLIUM CHRYSOGENUMAND ASPERGILLUS NIGER
P. Aleksieva1, B. Tchorbanov2, L. Nacheva1
1Bulgarian Academy of Sciences, Institute of Microbiology, Sofia, Bulgaria2Bulgarian Academy of Sciences, institute of organic chemistry with centre of Phytochemistry,Sofia, BulgariaCorrespondence to: Lilyana NachevaE-mail: [email protected]
ABSTRACTα-Galactosidase production by the fungal strains Penicillium chrysogenum LN33 and Aspergillus niger M47 cultivated in shaking and stationary conditions using soya meal extract as nutrient medium was investigated. Submerged shake flask fermentation was preferred over stationary cultivation for P. chrysogenum LN33 as it gave a high α-galactosidase yield (4200U/l). Because of pellet formation the semicontinuous shake-flask fermentation was carried out for P. chrysogenum LN33 α-galactosidase production during 32 days. The maximum α-galactosidase activity by A. niger M47 (7000U/l) was obtained in stationary conditions of cultivation which is 27-28% greater as compared with the enzyme titer in shaking cultures. Air-lift laboratory bioreactor cultivation of both P. chrysogenum LN33 and A. niger M47 resulted in high-yield production of α-galactosidase (8700U/l and 11200U/l, respectively), which is of particular interest for biotechnological application in food processing.
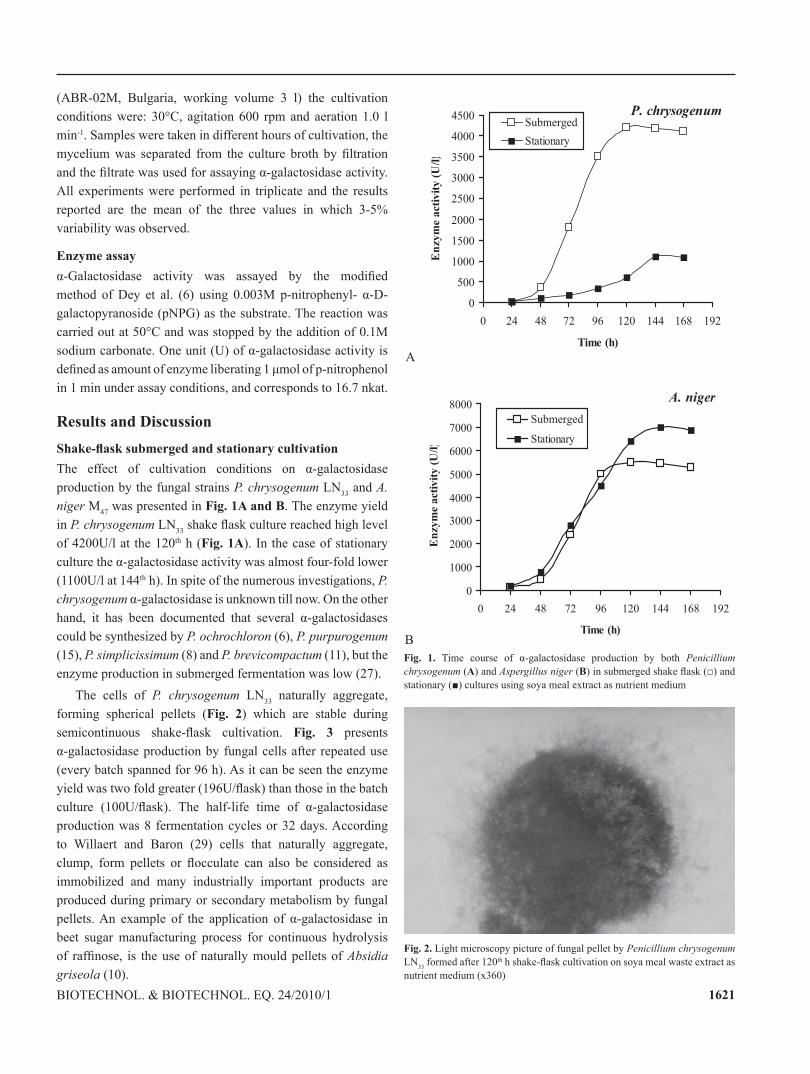
1621Biotechnol. & Biotechnol. eq. 24/2010/1
(ABR-02M, Bulgaria, working volume 3 l) the cultivation conditions were: 30°c, agitation 600 rpm and aeration 1.0 l min-1. Samples were taken in different hours of cultivation, the mycelium was separated from the culture broth by filtration and the filtrate was used for assaying α-galactosidase activity. All experiments were performed in triplicate and the results reported are the mean of the three values in which 3-5% variability was observed.
Enzyme assayα-Galactosidase activity was assayed by the modified method of Dey et al. (6) using 0.003M p-nitrophenyl- α-D-galactopyranoside (pNPG) as the substrate. The reaction was carried out at 50°c and was stopped by the addition of 0.1M sodium carbonate. One unit (U) of α-galactosidase activity is defined as amount of enzyme liberating 1 μmol of p-nitrophenol in 1 min under assay conditions, and corresponds to 16.7 nkat.
Results and DiscussionShake-flask submerged and stationary cultivationThe effect of cultivation conditions on α-galactosidase production by the fungal strains P. chrysogenum ln33 and A. niger M47 was presented in Fig. 1A and B. the enzyme yield in P. chrysogenum ln33 shake flask culture reached high level of 4200U/l at the 120th h (Fig. 1A). in the case of stationary culture the α-galactosidase activity was almost four-fold lower (1100U/l at 144th h). In spite of the numerous investigations, P. chrysogenum α-galactosidase is unknown till now. On the other hand, it has been documented that several α-galactosidases could be synthesized by P. ochrochloron (6), P. purpurogenum (15), P. simplicissimum (8) and P. brevicompactum (11), but the enzyme production in submerged fermentation was low (27).
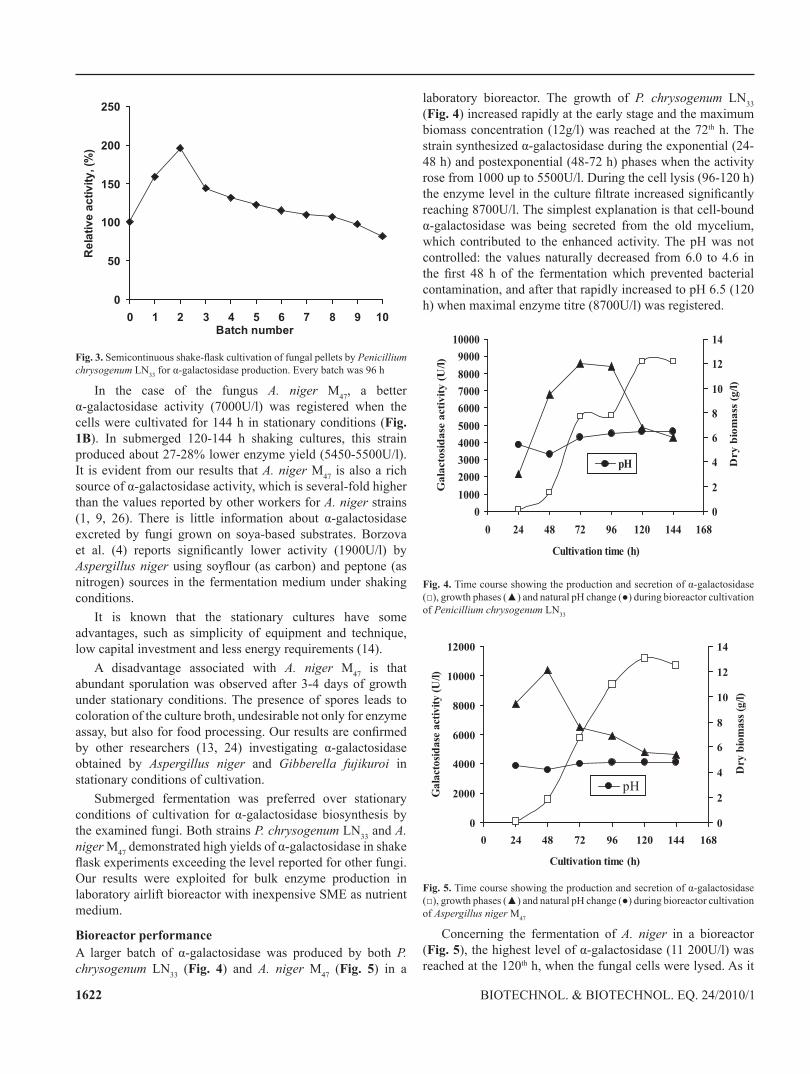
the cells of P. chrysogenum ln33 naturally aggregate, forming spherical pellets (Fig. 2) which are stable during semicontinuous shake-flask cultivation. Fig. 3 presents α-galactosidase production by fungal cells after repeated use (every batch spanned for 96 h). As it can be seen the enzyme yield was two fold greater (196U/flask) than those in the batch culture (100U/flask). The half-life time of α-galactosidase production was 8 fermentation cycles or 32 days. According to Willaert and Baron (29) cells that naturally aggregate, clump, form pellets or flocculate can also be considered as immobilized and many industrially important products are produced during primary or secondary metabolism by fungal pellets. An example of the application of α-galactosidase in beet sugar manufacturing process for continuous hydrolysis of raffinose, is the use of naturally mould pellets of Absidia griseola (10).
A
P. chrysogenum
0
500
1000
1500
2000
2500
3000
3500
4000
4500
0 24 48 72 96 120 144 168 192
Time (h)
En
zym
e a
cti
vit
y (
U/l
)
Submerged
Stationary
B
A. niger
0
1000
2000
3000
4000
5000
6000
7000
8000
0 24 48 72 96 120 144 168 192
Time (h)
En
zym
e a
cti
vit
y (
U/l
)
Submerged
Stationary
Fig. 1. Time course of α-galactosidase production by both Penicillium chrysogenum (A) and Aspergillus niger (B) in submerged shake flask (□) and stationary (■) cultures using soya meal extract as nutrient medium
Fig. 2. light microscopy picture of fungal pellet by Penicillium chrysogenum ln33 formed after 120th h shake-flask cultivation on soya meal waste extract as nutrient medium (x360)
1622 Biotechnol. & Biotechnol. eq. 24/2010/1
0
50
100
150
200
250
0 1 2 3 4 5 6 7 8 9 10Batch number
Rel
ativ
e ac
tivity
, (%
)
Fig. 3. Semicontinuous shake-flask cultivation of fungal pellets by Penicillium chrysogenum ln33 for α-galactosidase production. Every batch was 96 h
in the case of the fungus A. niger M47, a better α-galactosidase activity (7000U/l) was registered when the cells were cultivated for 144 h in stationary conditions (Fig. 1B). in submerged 120-144 h shaking cultures, this strain produced about 27-28% lower enzyme yield (5450-5500U/l). It is evident from our results that A. niger M47 is also a rich source of α-galactosidase activity, which is several-fold higher than the values reported by other workers for A. niger strains (1, 9, 26). There is little information about α-galactosidase excreted by fungi grown on soya-based substrates. Borzova et al. (4) reports significantly lower activity (1900U/l) by Aspergillus niger using soyflour (as carbon) and peptone (as nitrogen) sources in the fermentation medium under shaking conditions.
It is known that the stationary cultures have some advantages, such as simplicity of equipment and technique, low capital investment and less energy requirements (14).
A disadvantage associated with A. niger M47 is that abundant sporulation was observed after 3-4 days of growth under stationary conditions. the presence of spores leads to coloration of the culture broth, undesirable not only for enzyme assay, but also for food processing. Our results are confirmed by other researchers (13, 24) investigating α-galactosidase obtained by Aspergillus niger and Gibberella fujikuroi in stationary conditions of cultivation.
Submerged fermentation was preferred over stationary conditions of cultivation for α-galactosidase biosynthesis by the examined fungi. Both strains P. chrysogenum ln33 and A. niger M47 demonstrated high yields of α-galactosidase in shake flask experiments exceeding the level reported for other fungi. our results were exploited for bulk enzyme production in laboratory airlift bioreactor with inexpensive SME as nutrient medium.
Bioreactor performanceA larger batch of α-galactosidase was produced by both P. chrysogenum ln33 (Fig. 4) and A. niger M47 (Fig. 5) in a
laboratory bioreactor. the growth of P. chrysogenum ln33 (Fig. 4) increased rapidly at the early stage and the maximum biomass concentration (12g/l) was reached at the 72th h. the strain synthesized α-galactosidase during the exponential (24-48 h) and postexponential (48-72 h) phases when the activity rose from 1000 up to 5500U/l. During the cell lysis (96-120 h) the enzyme level in the culture filtrate increased significantly reaching 8700U/l. The simplest explanation is that cell-bound α-galactosidase was being secreted from the old mycelium, which contributed to the enhanced activity. The pH was not controlled: the values naturally decreased from 6.0 to 4.6 in the first 48 h of the fermentation which prevented bacterial contamination, and after that rapidly increased to ph 6.5 (120 h) when maximal enzyme titre (8700U/l) was registered.
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
0 24 48 72 96 120 144 168
Cultivation time (h)
Ga
lacto
sid
ase
acti
vit
y (
U/l
)
0
2
4
6
8
10
12
14
Dry
bio
ma
ss (
g/l
)
pH
Fig. 4. Time course showing the production and secretion of α-galactosidase (□), growth phases (▲) and natural pH change (●) during bioreactor cultivation of Penicillium chrysogenum ln33
0
2000
4000
6000
8000
10000
12000
0 24 48 72 96 120 144 168
Cultivation time (h)
Ga
lact
osi
da
se a
ctiv
ity
(U
/l)
0
2
4
6
8
10
12
14
Dry
bio
ma
ss (
g/l
)
pH
Fig. 5. Time course showing the production and secretion of α-galactosidase (□), growth phases (▲) and natural pH change (●) during bioreactor cultivation of Aspergillus niger M47
concerning the fermentation of A. niger in a bioreactor (Fig. 5), the highest level of α-galactosidase (11 200U/l) was reached at the 120th h, when the fungal cells were lysed. As it

1623Biotechnol. & Biotechnol. eq. 24/2010/1
can be seen (Fig. 5) the biomass concentration was reduced almost twice from its maximal value of 12.1 g/l in the 48th h to 5.6 g/l in the 120th h. Maximum α-galactosidase activity occurred at ph 4.8, also in the absence of ph control. the best enzyme yields of α-galactosidase by P. chrysogenum ln33 and A. niger M47 were several-fold higher than the activities reported for other fungi belonging to the genera Aspergillus, Penicillium, Thermomyces, Humicola and Trichoderma (1, 4, 7, 8, 9, 11, 16, 19, 26, 30).
Conclusionsthe present study of the fungal species P. chrysogenum as a source of extracellular α-galactosidase has not been investigated previously. Moreover, to our knowledge, for the first time we have demonstrated the semicontinuous shake-flask cultivation of fungal pellets of P. chrysogenum ln33 for α-galactosidase production during one month.
the results of the present paper indicate the potential of the fungal strains P. chrysogenum ln33 and A. niger M47 for production of high yields of α-galactosidase under bioreactor fermentation conditions. this might be employed for enzymatic hydrolysis of soymilk in order that commercially viable galacto-oligosaccharides are removed.
AcknowledgementsThe authors thank the National Foundation for Scientific Research, Bulgarian Ministry of education and Sciences for financial support of Project CC 1509/05.
REFERENCES1. Ademark P., Larsson M., Tjerneld F., Stalbrand H.
(2001) enzyme Microb. technol., 29, 441-448.2. Aleksieva P., Nacheva L., Bratovanova E., Tchorbanov
B. (2007) Compt. Rend. Acad. Bulg. Sci., 60, 691-696.3. Asano N., Ishii S., Kizu H., Ikeda H., Yasuda K., Kato
A., Martin O.R., Fan J.-Q. (2000) eur. J. Biochem., 267, 4179-4186.
4. Borzova N.V., Malanchuk V.M., Varbanets L.D., Seifullina I.I., Zubkov S.T. (2001) Mikrobiol. Z., 63, 27-36.
5. Bulpin P.V., Gidley M.J., Jeffcoat R. (1990) Carbohyd. Polym., 12, 155-168.
6. Dey P.M., Patel S., Brownleader M.D. (1993) Biotechnol. Appl. Biochem., 17, 361-371.
7. Kotwal S.M., Khan M.I., Khire J.M. (1995) J. Ind. Microbiol. Biotechnol., 15, 116-120.
8. Luonteri E., Alatalo E., Siika-aho M., Penttila M., Tenkanen M. (1998) Biotechnol. Appl. Biochem., 28, 179-188.
9. Manzanares P., de Graaff L.H., Visser J. (1998) Enzyme Microb. technol., 22, 383-390.
10. Masoud F., Nasrin M., Saeed M., Reza M., Manouchehr V., Reza B.M. (2002) World J. Microbiol. Biotechnol., 18, 649-653.
11. Mckay A.M. (1991) J. Food Sci., 56, 1749-1750.12. Mulimani V.H. and Ramaligam (1995) Biochem. Mol.
Biol. int., 36, 897-905. 13. Mulimani V.H., Ramaligam (1997) World J. Microbiol.
Biotechnol., 13, 583-585. 14. Nigam P., Singh D. (1994) J. Basic Microbiol., 34, 405-
423.15. Park G.G., Lee S.Y., Park B.K., Ham S.S., Lee J.H.
(1991) J. Microbiol. Biotechnol., 1, 90-95.16. Puchart V., Vrsanska M., Bhat M.K., Biely P. (2000)
Biochim. Biophys. Acta, 1524, 27-37.17. Rättö M., Siika-aho M., Buchert J., Valkeajarvi A.,
Viikari L. (1993) Appl. Microbiol. Biotechnol., 40, 449-454.
18. de Rezende S.T., Guimaraes V.M., Rodrigues M.C., Felix C.R. (2005) Braz. Arch. Biol. technol., 48, 195-202.
19. Rios S., Pedgregosa A.M., Monistrol I.F., Laborda F. (1993) FEMS Microbiol. Lett., 112, 5-42.
20. Sanni A.I., Onilude A.A., Ogundoye O.R. (1997) nahrung., 41,18-21.
21. Sinitsyna O.A., Fedorova E.A., Vakar I.M., Kondratieva E.G., Rozhkova A.M., Sokolova L.M., Bubnova T.M., Okunev O.N., Chulkin A.M., Vinetsky Y.P., Sinitsyn A.P. (2008) Biochemistry (Moscow), 73, 97-106.
22. Shankar SK., Mulimani V.H. (2007) Biores. Technol., 98, 958-961.
23. Shibuya H., Kobayashi H., Park G.G., Komatsu Y., Sato T., Kaneko R., Nagasaki H., Yoshida S., Kasamo K., Kusakabe I. (1995) Biosci. Biotechnol. Biochem., 59, 2333-2335.
24. Somiari R.I., Balogh E. (1995) Enzyme Microb. Technol., 17, 311-316.
25. Varbanets L.D., Malanchuk V.M., Buglova T.T., Kuhlmann R.A. (2001) carbohyd. Polym., 44, 357-363.
26. Wallis G.L.F., Easton R.L., Jolly K., Hemming F.W., Peberdy J.F. (2001) eur. J. Biochem., 268, 4134-4143.
27. Wang C.L., Li D.F., Lu W.Q., Wang Y.H., Lai C.H. (2004) lett. Appl. Microbiol., 39, 369-375.
28. Weignerova L., Simerska P., Kren V. (2009) Biocatal. Biotransfor., 27, 79-89.
29. Willaert R.G. and Baron G.V. (1996) In: Immobilized Living Cell Systems, modeling and experimental methods (R.G. Willaert, G.V. Baron, L.D. Backer, Eds.), p. 12. West Sussex.
30. Zeilinger S., Kristufek D., Arisan-atact I., Hodits R., Kubicek C.P. (1993) Appl. Environ. Microbiol., 59, 1347-1353.