health and oral inflammatory diseases: a ... their advantageous effects maintaining health of...
TRANSCRIPT

EUROPEAN JOURNAL OF INFLAMMATION
1721-727X (2009)Copyright © by BIOLIFE, s.a.s.
This publication and/or article is for individual use only and may not be furtherreproduced without written permission from the copyright holder.
Unauthorized reproduction may result in financial and other penalties53
THE ROLE OF MOLECULAR CHAPERONES (HSPAs/HSP70s) IN ORAL HEALTH AND ORAL INFLAMMATORY DISEASES: A REVIEW
T.K. FÁBIÁN, L. GÓTAI, A. BECK, G. FÁBIÁN1 and P. FEJÉRDY
Semmelweis University Budapest, Faculty of Dentistry, Clinic of Prosthetic Dentistry, Budapest; 1Semmelweis University Budapest, Faculty of Dentistry, Clinic of Pediatric Dentistry and
Orthodontics, Budapest, Hungary, EU
Received December 4, 2008 - Accepted May 28, 2009
EDITORIAL
Mailing address: Dr. Tibor Károly Fábián, Semmelweis University Budapest, Clinic of Prosthetic Dentistry, Szentkirályi utca 47,1088 Budapest, Hungary, EUTel: ++36 1338 4380 Fax: ++36 1317 5270e-mail: [email protected]
Key words: HSPA, HSP70, saliva, inflammation, periodontal, mucosal, bone, tooth
Heat shock proteins of the 70kDa family (HSPAs/HSP70s) are major molecular chaperones and cytokines of most cells and microbes, extracellular and interstitial fluids, blood, synovial fluids and secretory body fluids like saliva. The induction of human HSPAs plays an important role at cellular level under most stress conditions; whereas microbial HSPAs improve microbial tolerance to environmental changes, and improve virulence and resistance against antimicrobial peptides. Extracellular HSPAs reveal cytoprotective properties and are involved in numerous physiological and pathological events, including modulation of cytokine release and immunity. Accordingly, HSPAs play a role in the maintenance of pulpal health, and the repair of injured dental hard tissues. HSPAs also play a role in stress adaptation of periodontal tissues, and in the maintenance of periodontal and mucosal health including defense against microbes, prevention of mucosal allergic reactions, and facilitation of healing of ulcers and wounds. Despite their advantageous effects maintaining health of several oral tissues, HSPAs are likely to play a role in the disadvantageous amplification of pulpal inflammatory response to bacteria, and in the formation of several periapical inflammatory lesions. HSPAs may also induce gingivitis under certain conditions, and play a role in the progression of periodontal bone defects. HSPAs may also play a role in atopic-type allergic reactions, autoimmune disorders, and haptenation in certain cases. Based on the above data, it can be assumed that HSPAs play an important role in oral defense under healthy conditions; however, their role is somewhat “Janus-faced” under pathological conditions.
The HSPA (HSP70) chaperone familyHeat shock proteins (HSPs) of the 70kDa family
(HSPAs) are major molecular chaperones and cytokine chaperokines (1) of most cells and tissues, extracellular and interstitial fluids, blood, synovial fluids and also secretory body fluids like saliva (2-3). There are at least thirteen homologous proteins in the human HSPA chaperone family. These proteins are either stress inducible or constitutively expressed
in most tissues. There are family members specific to endoplasmic reticulum or mitochondria. Other family members reside mainly in the cytosol and nucleus, and some are also present in lysosomes (4). Homologues have also been identified in bacteria, fungi and parasites.
Main intracellular functions of HSPAsHSPA type molecules have rather far-reaching
Vol. 7, no. 2, 0-0 (2009)

54 55Eur. J. Inflamm.
intracellular functions (2, 4). Some of these proteins are involved in multiple chaperoning and housekeeping functions like folding and refolding proteins, and coping with harmful aggregations of denatured proteins (2, 4). They also stabilize lysosomal membranes during stress. Others influence signal transduction pathways, cell cycle and senescence regulation, or inhibit apoptotic pathways (2, 4). They also facilitate the transport of newly synthesized proteins into the ER and mitochondrial lumen, and participate in their subsequent folding. Some are also involved in chaperone-mediated autophagy, and the disassemble of clastrin coated vesicles (2, 4).
Induction and inhibition of HSPAs in cells and microbes
As mentioned above, several HSPAs can be induced in the cells; however, there are great stimulation-specific, tissue-specific and species-specific variations in relation to induced HSPA expression (5). The majority of physical, chemical, biological, psychological or pathophysiological stressors and several medicamentous or medicinal herbal agents induce the expression of HSPAs in numerous tissues. Microbial HSPAs may also be induced under several stressful (micro)environmental changes. Increased expression of HSPAs in microbes improves their tolerance to environmental changes (6), and may further improve their virulence (6) and their resistance to antimicrobial peptides of innate immunity (7). Although the majority of publications are dedicated to the induction of HSPAs, there are studies available in the scientific literature related to the inhibition of HSPAs. HSPAs may be inhibited (or at least downregulated) by several proinflammatory cytokines such as interferon (IFN)-γ and tumor necrosis factor (TNF)-α; indicating that there is inhibitory feedback on HSPA induction under inflammation (8). The expression of inducible HSPAs may also be inhibited via certain benzylidene lactame compounds.
Main extracellular function of HSPAs In addition to their intracellular function, HSPAs
are frequently released into the extracellular spaces from cells (and also from microbes) under several stress conditions or when undergoing lysis or
necrosis (1-2). Besides these, release of extracellular HSPAs may also be triggered by body exercises or psychological stress (3, 9). Extracellular HSPAs reveal cytoprotective properties through cell surface association, which may be followed by internalization. Extracellular HSPAs (including both human and microbial) are also involved in numerous physiological and pathological events, including the modulation of cytokine release and immunity (1-2). Importantly, the increased level of extracellular HSPAs may induce both the activation and inhibition of the immune and inflammatory response (1, 10). HSPAs have both stimulating and inhibitory epitopes (10) which either activate or inhibit cytokine and chemokine production of dendritic cells or monocytes and maturation of dendritic cells. Besides the above, extracellular HSPAs modulate neuronal function (2), and entering the blood stream they also possess the ability to act at distant sites of the body as ancestral danger signals. Besides being present in the extracellular (interstitial) fluid and blood, the presence of HSPAs in human saliva has also been reported (9, 11) indicating that extracellular HSPAs may also play an important role on the oral surfaces.
HSPAs AND ORAL HEALTH
Health of the dental pulp and tooth repairThere is a relatively high level of HSPAs in
odontoblasts and pulp fibroblast of the normal dental pulp. HSPAs may be induced in dental pulp cells in vitro due to reoxygenation following hypoxia of hypoxia-reoxygenation stress models (12). HSPA expression may also be induced using heat stress of in vitro cultured pulp cells (13). Furthermore, an animal study on rats indicated that the level of HSPAs increases in the odontoblast process within three days following dentin cavity preparation („drilling of tooth”) (14); and the level of HSPAs is comparatively high during reparative dentinogenesis in newly formed odontoblast-like cells approximately fifteen days after the cavity preparation injury (14). Again, HSPAs are likely to inhibit secondary apoptosis occurring 1 day after a cavity preparation injury (15), but not primary apoptosis 1 hour after injury (15) of pulp cells, due to transient translocation from the cytoplasm to the
T.K. FÁBIÁN ET AL.

54 55Eur. J. Inflamm.
nuclei and the inhibition of c-Jun N-terminal kinase (JNK) pathway (15). Similarly, HSPAs are likely to be involved in the inhibition of pulp cell apoptosis induced by long-lasting elastic lateral forces of teeth in an animal study (16) using a classical model for disadvantageous mechanical load and orthodontic tooth movement. Premised data strongly indicate that HSPAs play an important role in maintaining pulpal health, and in repairing injured dental hard tissues (i.e. dentin) of teeth (14). HSPAs are also likely to play an important role in the differentiation of newly formed odontoblast-like cells (14), which play a major role in premised reparative dentinogenesis.
Tooth surface defense against bacteriaBesides the aforementioned effects of HSPAs
on the dental pulp, data indicate an important defense function of HSPAs also on tooth surfaces. Extracellular HSPAs (17), including also salivary HSPAs (18), bind several oral microbes including a major cariogen bacteria Streptococcus mutans (17-18). Moreover, salivary HSPAs bind hydroxylapatite, the major inorganic component of tooth surfaces (18). Based on the above results, it is likely that salivary HSPAs may play a role in the acquired pellicle formation followed by bacterial adhesion on tooth surfaces (18). Since the pH optimum of bacterial binding of salivary HSPAs is rather acidic, it is around pH 5.0-5.5 (18), and such a low pH is not typical for tooth surfaces at the early stage of bacterial colonization, therefore salivary HSPAs are likely to inhibit rather than facilitate bacterial adhesion to the acquired pellicle (18). Interestingly, binding of salivary HSPAs to bacteria and to hydroxylapatite was partially inhibited by ATP (18), although the meaning of ATP dependency of this binding property is not yet clear.
Immune surveillance and periodontal healthHSPAs may be induced in periodontal ligament
cells in vitro due to reoxygenation following hypoxia of hypoxia-reoxygenation stress models (12) in gingival epithelial cells with local application of low-intensity pulsed ultrasound application (19), and via heat shock in osteoblasts, (20) indicating that HSPAs play an important role in the stress adaptation of most periodontal tissues. Furthermore, the higher serum level of specific human antibodies against
microbial HSPAs, but importantly not against HSPAs of human origin, tends to positively correlate with healthier periodontal tissues (21), indicating that microbial heat shock proteins are immunodominant antigens triggering adaptive immunity of the organism to contribute to protection against oral microorganisms, and to maintain gingival health (21). Furthermore, the peptide binding C-terminal portion of microbial HSPAs strongly enhances serum IgG response triggered by certain (microbial) antigenic peptides following binding of C-terminal portion to the antigen at issue (10), which may also contribute to improved gingival health. Moreover, bacterial HSPAs (especially their premised C-terminal portion) also stimulate the generation of pro-inflammatory cytokines, c-c chemokines, and the maturation of dendritic cells (10), which strongly improves innate immunity and also drives adaptive immunity.
Cytoprotection and mucosal defenseThe increased level of cellular HSPAs was
brought into connection with improved protection against heat and alkaline exposure of the oral mucosa. Besides premised intracellular induction of HSPAs, important mechanisms for the maintenance of mucosal health are based on the cytoprotective effects of extracellular HSPAs (3, 11). Extracellular cytoprotective effects seem to be based on three different mechanisms: 1) aspecific binding of HSPAs on mucosal cell surfaces leading to surface defense against toxins most likely through protection and repair (chaperoning) of mucosal cell surface proteins (3, 11); 2) a more specific adhesin-type binding to sulfoglycolipid structures of mucosal cells preventing bacterial colonization of mucosal surfaces through occupying mucosal binding sites of HSPA related bacterial adhesins (3, 18); 3) surface receptor binding of HSPAs mostly followed by internalization, leading to the decrease of the cells’ apoptotic and necrotic liability and release of several cytokines (1-2).
Immune/inflammatory defense of mucosaBesides surface protection, extracellular
HSPAs also participate in several immunological mechanisms (1-2) responsible for mucosal defense. The appearance of intracellular HSPAs on the

56 57Eur. J. Inflamm.
surface of spontaneously developing tumor cells leads to the lysis of these cells by natural killer (NK) cells (1-2). Surface expression of HSPAs on tumor cells is frequently accompanied by HSPA release (via lipid rafts and/or exosomes) leading to the activation of NK cells, Langerhans cells and dendritic cells that are present in the environment (1-2). Premised activation leads to increased uptake and cross-presentation of tumor-derived HSPA-peptide complexes for T cell recognition, which is a highly important mechanism for the maintenance of mucosal health via proper immunological elimination of appearing mucosal malignant cells (1-2). Furthermore, cellular or microbial damage (or release) induced extracellular appearance of both uncomplexed „free” HSPAs and membrane bound HSPAs act as ancestral danger signals, which leads to (1-2, 4): 1) the release of proinflammatory cytokines from several immune cells like monocytes, dendritic cells, macrophages, T lymphocytes; 2) the release of NO from macrophages; 3) the activation of NK cells; 4) the activation of complement via antibody-independent alternative pathway; 5) the induction of local sIgA responses against microbial HSPA homologues, which blocks adherence and prevents transmucosal invasion of the microbe at issue (3, 18). Moreover, because of their chaperoning ability, uncomplexed HSPAs bind other peptides; which results in complexes inducing receptor-mediated uptake into antigen-presenting cells to cross-present these complexes as antigens (coupled with MHC-I or MHC-II molecules) to cytotoxic T cells and NK cells (1-2). Furthermore, HSPAs are likely to exert an opsonizing effect on bacteria, which activates the killing activity of polymorphonuclear neutrophil (PMN) leukocytes (17).
Defense function of salivary HSPAsMost of the above-mentioned immunological
functions are also expected for salivary HSPAs covering the surface of oral mucosa (3, 18), especially as oral mucosa (particularly non-keratinized parts) is extensively populated by antigen presenting Langerhans and dendritic cells (22), and Langerhans cells are properly oriented to „sample” the oral fluids with their dendrites toward the mucosal surface. It is also likely that salivary HSPAs entrap and agglutinate bacteria (18), especially because they are
likely to be capable of forming dimers and oligomers. The opsonizing effect of salivary HSPAs similar to those of HSPAs in blood (17) may also be taken into consideration, as the bacterial binding property of salivary HSPAs is very similar to those of HSPAs in blood (18). Salivary HSPAs via expected salivary HSPA/Histatin-5 complex (which would be able to enter and destroy Candida albicans) was assumed to have an antifungal effect (18) based on previous Histatin-5 and Ssa1p, Ssa2p (HSPAs of Candida albicans) related studies.
Oral tolerance and prevention of allergyBesides surface protection and immunological
defense, HSPAs may also play a role in the prevention of mucosal (and/or generalized) allergic reactions. A recent article (22) indicated that the most efficient tools used in various vector systems for sublingual immunotherapeutic approaches target innate immunity receptors such as Toll-like receptors of oral mucosal dendritic cells (22), and that the internalization of the vector (and vector-bound antigen) into the immune cells may also increase efficiency (22). Furthermore, optimization of mucosal adhesion of antigens with the help of mucoadhesive additives may also enhance the efficiency of such systems (22). In this study, we would like to point out that HSPAs are able to bind antigenic peptides forming HSPA-antigen complexes (1-2), and that HSPAs fulfill all of the premised criteria of expected optimal vectors as well (1-3, 18). Consequently, HSPAs of mucosal cell origin (18), as well as salivary HSPAs (18), are likely to play a role as „natural occurring vectors” for physiological desensitization processes, similarly as described in relation with „oral tolerance”-based sublingual immunotherapeutic approaches (22).
Stress adaptation and HSPA excretion of salivary glands
Heat shock efficiently induces HSPAs in salivary gland cells in several animal models (23), and the increased level of cellular HSPAs was brought into connection with the prevention of irradiation-induced cellular damage of salivary glands (24). Accordingly, the level of HSPAs in human saliva could also be increased by several stressors including psychological stress (9) and local heat or mechanical stress (massage)
T.K. FÁBIÁN ET AL.

56 57Eur. J. Inflamm.
of the major salivary glands (18). Moreover, stress-induced increase of HSPAs in the blood seems to be regulated via sympathetic activation-coupled release of norepinephrine targeting α1-adrenergic receptors of not yet known target cells (25). Importantly, functional α1-adrenergic receptors were also reported in human salivary glands, and elevation of salivary HSPA values were found to be accompanied by sympathetic upregulation (9). All these findings strongly indicate that HSPAs play an important role in the stress adaptation of salivary glands; and increased excretion of HSPAs into the saliva belongs to the salivary glands’ functional answer to various stressors. Accordingly, salivary HSPAs can also be induced via chewing and taste stimuli (11), indicating that regularly occurring stress of forced fluid secretion (parasympathetic stimuli) is also coupled with an increased excretion of salivary HSPAs.
HSPAs IN ORAL INFLAMMATORY DISEASES
Inflammation of dental pulpDespite their advantageous effects maintaining
health of dental pulp (described above), HSPAs are likely to play an important role in the disadvantageous amplification of pulpal inflammatory response to bacteria (6) from carious dentin and to several restorative dental materials (26). Although HSPA-dependent inflammatory pathways of dental pulp have not yet been investigated in detail, it is likely that the majority of immune stimulatory and also immune modulatory functions of human and microbial HSPAs come into play (6) with a shifted balance toward immune activation and inflammation. Interestingly, methacrylate-based dental adhesives used for deep dentin cavity or direct pulp-capping may advantageously influence such HSPA-dependent pulpal inflammatory processes for two reasons: 1) such adhesives do not trigger cytotoxicity-induced (26) HSPA expression of human monocytes (27); 2) they down-regulate stress-induced HSPA expression of monocytes playing a major role in such reactions (27). Importantly, the blue light used for curing adhesives also does not induce the expression of HSPAs in monocytes.
Periapical inflammationHSPAs also play a role in the formation of
several periapical inflammatory lesions (28). The expression of HSPAs was increased in lymphocytes and endothelial cells of inflammatory periapical granulation tissues (28). There was also a tendency towards the increase of HSPA expression in lining epithelium of periapical inflammation-induced radicular and residual cysts comparing to Malassez’ epithelial rests of control areas (28), which may indicate a possible role of HSPAs in the activation and proliferation of lining epithelium (28). Please also note that surprisingly the Malassez’ epithelial rests of control non-cystic areas also express a relatively high amount of HSPAs (28). Furthermore, HSPAs (both microbial and mammalian) stimulate bone resorption (29), presumably via the induction of proinflammatory cytokines, which are also known activators of osteoclasts (8). Premised bone resorptive effect is another strong indication of an important role of HSPAs in the progression of periapical lesions.
Inflammation of gingiva and periodontal tissues Although the role of HSPAs is likely to be
protective against gingivitis in general (as described above), extracellular HSPAs may nevertheless induce gingivitis under certain conditions. Data clearly indicated that there are antibodies against human HSPs of the 60 kDa family which may serve as autoantigens and could initiate an autoimmune response that contributes to the initiation of gingivitis (30). Since antibodies against HSPAs are also likely to be present (31), it may not be excluded that HSPAs also induce gingivitis via such autoimmune mechanisms (18). Furthermore, the bone-resorptive effect of (microbial and mammalian) HSPAs (29) are likely to play an important role in the progression of periodontal bone defects after the turning of gingivitis into a more severe inflammation (i.e. periodontitis) with pocket formation and the irreversible destruction of the periodontal bone (8, 18). Similarly, HSPAs are to likely play a role in the accelerated loss of periodontal bone under spondyloarthropathies (8). Another aspect of periodontal inflammatory disorders is coupled with Porhyromonas gingivalis (P. gingivalis), a major pathogen of periodontitis in relation with the finding that live P. gingivalis downregulates HSPAs in monocytes (32). Premised „calming

58 59Eur. J. Inflamm.
behavior” of P. gingivalis related to monocyte HSP70 response (32) could be a reason for the high (periodonto)pathogenity of premised microbe as the initiation of effective humoral response against P gingivalis is strongly dependent on the function of antigen-presenting cells including monocytes (33). Considering that the level of species specific IgG coupled humoral immunity against P. gingivalis in periodontitis-stable patients is higher than those with active periodontitis (34-35), premised depressed responsiveness of peripheral blood mononuclear cells (monocytes) is likely to play a major role in the pathomechanism of P. gingivalis infection-coupled periodontitis (33). Interestingly, extracted P. gingivalis lipopolysacharides (LPS) upregulate HSPAs in monocytes (32); therefore lysis of P. gingivalis via any pathways would presumably restore responsiveness and the antigen presenting function of monocytes.
Oral ulcers and wound healingThe increased HSPA level of mucosal cells
has been demonstrated in the case of non-specific oral ulcerations and gingival wound healing (19). Although oral tissues have not been particularly investigated, it is very likely that the role of HSPAs in the pathomechanism and healing of oral ulcers and oral wounds is similar to those described in relation to gastric ulcers, and wound healing of the skin (3, 18). In the case of gastric ulcers, HSPA is markedly overexpressed in cells located at the ulcer base, the level decreasing with ulcer healing, and the extent of HSPA induction in mucosal cells inversely correlates with the severity of newly induced ulcers. Similarly, the level of intracellular HSPAs positively correlates with the efficiency of wound healing of the skin, and the level of HSPAs decreases with the progress of the healing process. Besides premised intracellular effects of HSPAs, a coupled extracellular HSPA related defense (3, 18) is also likely because: 1) forced intracellular expression of HSPAs seems to increase the secretion of HSPAs into the extracellular space (36); 2) the regulation of intracellular HSPA level of gastrointestinal cells seems to be mediated by alpha 1A-adrenoceptors, the same receptor type which seems to be responsible for extracellular release (increase of blood level) of HSPAs (25); 3) extracellular HSPAs are released
by white blood cells in wound fluid 4) in vivo delivery of HSPAs increase the efficiency of wound healing by the stimulation of macrophage-mediated phagocytosis of wound debris (37), 5) extracellular HSPAs activate epidermal growth factor (EGF) receptors and related signaling pathways (37), which are likely to speed up the healing process by angiogenetic and cell proliferating effects of salivary EGF. Based on these data, it is likely that HSPAs play an important role in the healing of ulcers and wounds of oral mucosa, similarly to those of other gastrointestinal mucosal surfaces and the skin (18). However, the important distinctive features of saliva like hyposmolarity, and several special components including salivary defense proteins or proteases should also be considered in the case of oral lesions. It may also be considered that expected coupling of intra and extracellular surface defense in case of oral ulcers and wounds is primarily given by the fact that salivary HSPAs are present on the oral ulcer and wound surfaces covered by saliva (18).
Oral lichen planusIn contrast to early data indicating increased
expression of HSPAs there is a very likely slight decrease of HSPA level in oral mucosal ceratinocytes under oral lichen planus which is a chronic inflammatory mucosal disease of uncertain etiology. Importantly, topical treatment of this mucosal disorder with tacrolimus, an agent inducing HSPAs and having immunomodulatory properties, moderately increases the expression of HSPAs in oral ceratinocytes (38) and leads to rapid improvements in the majority of oral lichen planus cases (38). Although changes of HSPAs are „moderate” in this case only (under both disease and improvement), an important role of HSPAs in oral lichen planus may be expected based on premised findings.
Mucosal allergic reactions and autoimmunityDespite the expected role of HSPAs in the „oral
tolerance” (see above), some data in the literature suggest that overexpression of HSPAs may also play a role in the appearance of atopic-type (IgE mediated) allergic reactions, in autoimmune disorders, and in haptenation inducing T cell immunity and sensitization. However, it is not yet clear to what extent extracellular HSPAs are involved in such
T.K. FÁBIÁN ET AL.

58 59Eur. J. Inflamm.
immune surveillance and cellular stress tolerance. The local use of several physiotherapies to induce HSPAs seems to be reasonable as a pretreatment before presumable local injuries (including operative dental and oral surgery). Similarly, the overcoming of initial inflammatory reactions may be promoted in this way. Induction of HSPAs could also be useful to „reactivate” and promote the healing of certain chronic inflammations, especially those coexisting with a decreased HSPA level. In case of acute inflammation at an advanced stage, theoretically both induction and inhibition of HSPAs could be useful, depending on the properties of a given immune/inflammatory reaction; however, our knowledge is not sufficient to decide which choice (i.e. induction or inhibition) would be better in a concrete case.
It should also be considered that many of the oral aspects discussed above still remain hypotheses, as a lot of the data is not particularly related to oral tissues, therefore, the possible effects of oral environment and specificities of the oral mucosa could not be taken into consideration in all cases. Consequently, besides summarizing the possible role of HSPAs in the oral cavity and oral inflammatory disorders, another goal of the present review is to stimulate research activities leading to clear related evidence.
REFERENCES
1. Asea A. Stress proteins and initiation of immune response: Chaperokine activity of Hsp70. Exerc Immunol Rev 2005; 11:34-35.
2. Calderwood SK, Mambula SS, Gray PJ jr, Theriault JR. Extracellular heat shock proteins in cell signaling. (Minireview). FEBS Letters 2007; 581:3689-94.
3. Fábián TK, Fejérdy P, Nguyen MT, Sőti Cs, Csermely P. Potential immunological functions of salivary Hsp70 in mucosal and periodontal defense mechanisms. Arch Immunol Ther Exp 2007; 55:91-8.
4. Kampinga HH, Hageman J, Vos MJ, et al. Guidelines for the nomenclature of the human heat shock proteins. Cell Stress Chaperones 2009; 14:105-11.
5. Singh AK, Lakhotia SC. Tissue-specific variations in the induction of Hsp70 and Hsp64 by heat shock in
processes in relation to the oral mucosa, which considerably differs from other mucosal surfaces in this relation (22). Cross-reactivity of specific antibodies against microbial HSPAs with human HSPAs may occur similarly to other types of HSPs (3, 18). However, the occurrence of such phenomenon is likely to be somewhat more improbable in the case of HSPAs, because of a somewhat lower degree of homology compared to other HSPs. Autoantibodies against human HSPAs also occur in the blood of normal healthy subjects (31) with a frequency comparable to those of patients suffering from certain autoimmune diseases. Interestingly, a highly frequent occurrence of autoantibodies against human HSPAs was found in the serum of metal-allergic and atopic patients, whereas atopic patients without any metal allergy did not show this property (39). The latter finding may indicate the possibility that metals bind and affect HSPAs of such patients, resulting in structural alteration triggering autoimmune reaction (39). Evaluating the role of HSPAs in allergic and autoimmune reactions, it should also be considered that HSPAs also exert immune regulatory and anti-inflammatory functions (1, 10). The administration of recombinant HSPAs may also result in the attenuation of experimental autoimmune diseases. It has also been shown that low-affinity T cells are reactive against autologous heat shock proteins (40), which might lead to the generation of Th2 (IL-4- and IL-10-producing), Th3 (transforming growth factor-β-producing), or Tr1 (IL-10-producing) regulatory T cell responses and consequent release of premised regulatory cytokines. Taking together all these data, it is likely that HSPAs may take part in both the release and control of allergic and autoimmune reactions of the oral mucosa under certain conditions (3, 18).
CONCLUSIONS
Based on the above data, it can be assumed that HSPAs play an important role in oral defense under both healthy and pathological conditions; however, their role is somewhat „Janus-faced” exerting both immune stimulating and immune regulatory properties. Nevertheless, it may be concluded that slight stimulation of HSPA expression (i.e. via body exercises or psychological training) seems to be advantageous even in healthy conditions to improve

60 61Eur. J. Inflamm.
insects. Cell Stress Chaperones 2000; 5:90-7. 6. Henderson B, Allan, E, Coates ARM. Stress wars:
the direct role of host and bacterial molecular chaperones in bacterial infection. Infect Immunity 2006; 74:3693-706.
7. Shelburne CE, Coulter WA, Olguin DA, Lantz MS, Lopatin DE. Induction of β-defensin resistance in the oral anaerobe Porphyromonas gingivalis. Antimicr Agent Chemother 2005; 49:183-7.
8. Fábián TK, Csermely P, Fábián G, Fejérdy P. Spondyloarthropathies and bone resorption. Possible role of heat shock protein 70 (Hsp70). Acta Physiol Hung 2009; 96:149-55.
9. Fábián TK, Tóth Zs, Fejérdy L, Kaán B, Csermely P, Fejérdy P. Photo-acoustic stimulation increases the amount of 70 kDa heat shock protein (Hsp70) in human whole saliva. A pilot study. Int J Psychophysiol 2004; 52:211-6.
10. Wang Y, Whittall T, McGowan E, Younson J, Kelly C, Bergmeier LA, Singh M, Lehner T. Identification of stimulating and inhibitory epitopes within the heat shock protein 70 molecule that modulate cytokine production and maturation of dendritic cells. J Immunol 2005; 174:3306-16.
11. Fábián TK, Gáspár J, Fejérdy L, Kaán B, Bálint M, Csermely P, Fejérdy P. Hsp70 is present in human saliva. Med Sci Monitor 2003; 9:BR62-BR65.
12. Amemiya H, Matsuzaka K, Kokubu E, Ohta S, Inoue T. Cellular responses of rat periodontal ligament cells under hypoxia and reoxygenation conditions in vitro. J Periodontal Res 2008; 43:322-7.
13. Amano T, Muramatsu T, Amemiya K, Kubo K, Shimono M. Responses of rat pulp cells to heat stress in vitro. J Dent Res 2006; 85:432-5.
14. Chen Z, Fan M, Bian Z, Zhang Q, Zhu Q, Lu P. Immunolocalization of heat shock protein 70 during reparative dentinogenesis. Chin J Dent Res 2000; 3:50-5.
15. Kitamura C, Kimura K, Nakayama T, Toyoshima K, Terashita M. Primary and secondary induction of apoptosis in odontoblasts after cavity preparation of rat molars. J Dent Res 2001; 80:1530-4.
16. Shigehara S, Matsuzaka K, Inoue T. Morphohistological change and expression of HSP70, osteopontin and osteocalcin mRNAs in rat dental pulp cells with orthodontic tooth movement.
Bull Tokyo Dent Coll 2006; 47:117-24.17. Nguyen MT, Fábián TK, Singh M, Csermely P,
Sőti Cs. Bacterial binding and opsonizing effect of extracellular Hsp70. FEBS J, 2008; 275(S):460.
18. Fábián TK, Sőti Cs, Nguyen MT, Csermely P, Fejérdy P. Expected functions of salivary HSP70 in the oral cavity. In Heat shock proteins: New research. E. Morel, C. Vincent eds. Nova Science Publishers, Inc. New York, 2008; pp. 321-40.
19. Ikai H, Tamura T, Watanabe T, Itou M, Sugaya A, Iwabuchi S, Mikuni-Takagaki Y, Deguchi S. Low-intensity pulsed ultrasound accelerates periodontal wound healing after flap surgery. J Periodontal Res 2008; 43:212-6.
20. Li S, Chien S, Brånemark PI. Heat shock-induced necrosis and apoptosis in osteoblasts. J Orthop Res 1999; 17:891-9.
21. Lopatin DE, Shelburne CE, Van Poperin N, Kowalski CJ, Bagramian RA. Humoral immunity to stress proteins and periodontal disease. J Periodontol 1999; 70:1185-93.
22. Novak N, Haberstok J, Bieber T, Allam JP. The immune privilege of the oral mucosa. Trends Mol Med 2008; 14:191-8.
23. Yao J, Munson KM, Webb WW, Lis JT. Dynamics of heat shock factor association with native gene loci in living cells. Nature 2006; 442:1050-3.
24. Lee HJ, Lee YJ, Kwon HC, Bae S, Kim SH, Min JJ, Cho CK, Lee YS. Radioprotective effect of heat shock protein 25 on submandibular glands of rats. Am J Pathol 2006; 169:1601-11.
25. Johnson JD, Fleshner M. Releasing signals, secretory pathways, and immune function of endogenous extracellular heat shock protein 72. J Leukoc Biol 2006; 79:425-34.
26. Oshima H, Hatayama T, Nakamura M. A possibility for new evaluating method of cytotoxicity by using heat shock protein assay. J Material Sci 1997; 8:143-7.
27. Noda M, Wataha JC, Kaga M, Lockwood PE, Volkmann KR, Sano H. Components of dentinal adhesives modulate heat shock protein 72 expression in heat-stressed THP-1 human monocytes at sublethal concentration. J Dent Res 2002; 81:265-9.
28. Suzuki T, Kumamoto H, Ooya K, Motegi K. Expression of inducible nitric oxide synthase and heat shock proteins in periapical inflammatory
T.K. FÁBIÁN ET AL.

60 61Eur. J. Inflamm.
lesions. J Oral Pathol Med 2002; 31:488-93.29. Nair SP, Meghji S, Reddi K, Poole S, Miller AD,
Henderson B. Molecular chaperones stimulate bone resorption. Calcif Tissue Int 1999; 64:214-8.
30. Schett G, Metzler B, Kleindienst R, Moschén I, Hattmannsdorfer R, Wolf H, Ottenhoff T, Xu Q, Wick G. Salivary anti-hsp65 antibodies as a diagnostic marker for gingivitis and a possible link to atherosclerosis. Int Arch Allergy Immunol 1997; 114:246-50.
31. Pockley AG, Shepherd J, Corton JM. Detection of heat shock protein 70 (Hsp70) and anty-Hsp70 antibodies in the serum of normal individuals. Immunol Investig 1998; 27:367-77.
32. Saba JA, McComb ME, Potts DL, Costello CE, Amar S. Proteomic mapping of stimulus-specific signaling pathways involved in THP-1 cells exposed to Porhyromonas gingivalis or its purified components. J Proteome Res 2007; 2211-21.
33. Petit MDA, Wassenaar A, van der Velden U, van Eden W, Loos BG. Depressed responsiveness of peripheral blood mononuclear cells to heat-shock proteins in periodontitis patients. J Dent Res 1999; 78:1393-400.
34. Shelburne CE, Shelburne PS, Dhople VM, et al. Serum antibodies to Porphyromonas gingivalis chaperone HtpG predict health in periodontitis susceptible patients. PLoS ONE 2008; 3:e1984.
35. Rams TE, Listgarten MA, Slots J. Actinobacillus
actinemicetemcomittans and Porphyromonas gingivalis subgingival presence, species-specific serum immunoglobulin G antibody levels, and periodontitis disease recurrence. J Periodontal Res 2006; 41:228-34.
36. Wang MH, Grossmann ME, Young CYF. Forced expression of heat-shock protein 70 increases the secretion of Hsp70 and provides protection against tumor growth. British J Cancer 2004; 90:926-31.
37. Kovalchin JT, Wang R, Wagh MS, Azoulay J, Sanders M, Chandawarkar RY. In vivo delivery of heat shock protein 70 accelerates wound healing by up-regulating macrophage-mediated phagocytosis. Wound Repair Regen 2006; 14:129-37.
38. Tavassol F, Starke OF, Völker B, Kokemüller H, Eckardt A. Heat-shock protein expression and topical treatment with tacrolimus in oral lichen planus: an immunohystochemical study. Int J Maxillofac Surg 2008; 37:66-9.
39. Jin GB, Nakayama H, Shmyhlo M, Inoue S, Kondo M, Ikezawa Z, Ouchi Y, Cyong JC. High positive frequency of antibodies to metallothionein and heat shock protein 70 in sera of patients with metal allergy. Clin Exp Immunol 2003; 131:275-9.
40. Munk ME, Schoel B, Modrow S, Karr RW, Young RA, Kaufmann SH. T lymphocytes from healthy individuals with specificity to self-epitopes shared by the mycobacterial and human 65-kilodalton heat shock protein. J Immunol 1989; 143:2844-9.

EUROPEAN JOURNAL OF INFLAMMATION Vol. 7, no. 2, 0-0 (2009)
1721-727X (2009)Copyright © by BIOLIFE, s.a.s.
This publication and/or article is for individual use only and may not be furtherreproduced without written permission from the copyright holder.
Unauthorized reproduction may result in financial and other penalties63

EUROPEAN JOURNAL OF INFLAMMATION Vol. 7, no. 2, 0-0 (2009)
1721-727X (2009)Copyright © by BIOLIFE, s.a.s.
This publication and/or article is for individual use only and may not be furtherreproduced without written permission from the copyright holder.
Unauthorized reproduction may result in financial and other penalties63
It is well known that the chemical mediators of inflammatory response, such as histamine, C3a, C5a
(complement components), bradykinin, leukotrienes (LTC4, LTD4, LTE4), PAF, cytokines and substance
INFLAMMATORY COMPOUNDS: NEUROPEPTIDE SUBSTANCE P AND CYTOKINES
M.L. CASTELLANI, P. FELACO1, F. PANDOLFI2, V. SALINI3, D. DE AMICIS3, J. VECCHIET4, S. TETÈ5, C. CIAMPOLI5, F. CONTI6, G. CERULLI7, A. CARAFFA7, P.
ANTINOLFI7, C. CUCCURULLO8, A. PERRELLA9, T.C. THEOHARIDES10, M.A. DE LUTIIS11, D. KEMPURAJ10 and Y.B. SHAIK12
Immunology Division, Medical School, University of Chieti-Pescara, Italy; 1Division of Nephrology, University of Chieti, Italy; 2Catholic University, Rome, Italy; 3Department of Human Dynamic,
University of Chieti-Pescara, Italy; 4Clinic of Infectious Diseases, Medical School, University of Chieti-Pescara, Italy; 5Dental School, University of Chieti-Pescara, Italy; 6Gynecology Division,
University of Chieti, Italy; 7Orthopeadic Division, University of Perugia, Italy; 8Division of Medical Pathology, University of Chieti, Italy; 9Department of Infectious Diseases, Cotugno Hospital, Napoli,
Italy; 10Department of Pharmacology and Experimental Therapeutics, Biochemistry and Internal Medicine Tufts University School of Medicine, Tufts-New England Medical Center, Boston, MA, USA;
11Department of Biology, University of Chieti, Chieti, Italy; 12Department of Medicine, Boston University School of Medicine, Boston, MA, USA
Received April 30, 2009 – Accepted June 3, 2009
Inflammatory diseases represent one of the major causes of morbidity and mortality throughout the world and they affect the functions of several tissues. The pathophysiology of these diseases involves release of many pro-inflammatory mediators such as cytokines/chemokines, histamine, C3a, C5a (complement components), bradykinin, leukotrienes (LTC4, LTD4, LTE4), PAF, and substance P, in addition to anti-inflammatory molecules. Recently, it has been demonstrated that neuroimmune interactions are important in the initiation and progress of inflammatory processes. Substance P is an 11-amino acid neuropeptide that is released from nerve endings in many tissues. It acts via membrane-bound NK1 receptors (NK1R). Inflammatory and neuropeptides such as substance P stimulate the release of chemokines, in particular IL-8, a potent neutrophil chemoattractant. Expression of IL-8 is regulated mainly by the transcription factors NF-kappaB, activating protein-1. Substance P plays an important role in immunological and inflammatory states, and it is a mediator of tissue injury, asthma, arthritis, allergy and autoimmune diseases. In this article, our studies revisited the interrelationship between these two powerful inflammatory compounds: substance P and cytokines. These observations suggest that these inflammatory molecules may represent a potential therapeutic target to treat several inflammatory states.
EDITORIAL
Mailing address: Dr. Maria Luisa Castellani,Department of Medicine and Aging, Medical School,University of Chieti-Pescara,Via dei Vestini,66100 Chieti, ItalyTel: ++39 3286122802e-mail:[email protected]
Key words: inflammation, neuropeptide, cytokine, chemokine

64 65Eur. J. Inflamm.
P increase vascular permeability (1-2). Injury to a tissue provokes the release of inflammatory mediators that dilate arterioles and post–capillary venules and increase capillary permeability (3). In this case, the cells of the immune system cross the endothelial barrier and migrate to the specific site of injury (4). This migration is accomplished by the process of chemical signaling (chemotaxis) mediated by specific chemokines (5). However, previously discovered, there are also non-specific chemo-attractant compounds such as C3a, LTB4, lipoxins (A and B), PAF, bacterial products, cytokines (6).
Cytokines are proteins that act in an autocrine/paracrine manner to regulate leukocyte activity and most of them are elaborated as part of inflammatory response (7-8). Chemokines are a subset of cytokines that promote immune cell trafficking and localization to sites of inflammation (9). One of the most powerful inflammatory cytokines is tumor necrosis factor-α (TNF-α) which is derived predominantly from activated macrophages and acts via cell membrane-bound receptors (10-12). Raised levels of cytokines have been described in a number of acute conditions such as arthritis, allergy, sepsis, burns and major surgery (13-16), and administration of IL-1, TNFα, IL-6, and other pyrogenic cytokines induce, directly or indirectly, fever in vivo (17-20). Administration of certain cytokines such as IL-4, IL-10, IL-1RA (IL-1 receptor antagonist) or corticosteroids provoke the inhibition of inflammation in vitro (21-24) and subsequent reduction of mortality in septic shock in vivo (25).
Hemokines control mast cell infiltration in several inflammatory diseases, including stress and neurological dysfunctions (26-28).
Thus, cytokines and chemokines play several roles in regulating and amplifying immunity and inflammation (29-30). These effects include leukocyte recruitment and/or retention in the central nervous system (CNS) (31). Chemokines are the key modulators of inflammation, acting through G-protein-coupled receptors, and they encourage migration of cells to the site of inflammation (32-33).
The release of specific chemokines such as RANTES from activated macrophages is a crucial step in T cell recruitment necessary for establishing local inflammatory responses (34). In addition,
RANTES, a member of the intercrine beta subfamily and a C-C chemokine has been reported to act as a selective chemoattractant for monocytes, T cells, eosinophils and mast cells rather than neutrophils (35-36).
RANTES and various other chemokines play an active role in recruiting leukocytes into inflammatory sites and are potent chemokines involved in macrophage activation (37-38). It has been reported that chemokines, including ubiquitous RANTES, mediate and stimulate the release of inflammatory products in several cells including mast cells (39-40).
Activation of cytokine receptors and alterations in cytokines in the CNS are thought to play important roles in neuronal dysfunction and in the pathogenesis of the nervous system diseases (41).
Much evidence suggests a cross-talking between nerve fibers and the immunity system (42). Neuropeptides are capable of directly or indirectly inducing neurogenic inflammation and cause an increasing of induced cytokine/chemokines, vasodilatation, plasma extravasation, and cellular adhesion molecule expression required for activation and trafficking of inflammatory cells into the inflamed tissue (43-44). Neuropeptides may also play a role in the repair of tissue injury (45). Neuropeptide modulation of immune cell function is an important mechanism of neuro-immune system cross-talk, and substance P (SP) is one such key neuropeptide involved in this field (46-47).
Substance P is an 11-amino acid neuropeptide that is released from nerve endings in many tissues, and belongs to a family of related peptides called tachykinins and plays critical roles in immune-regulation in human and animal models (48). Other three related tachykinins are known, substance K, neuromedin K and hemokinin, which also have some biological activity in immunity and inflammation (48-50).
SP is a product of the sensory ganglion cells, and it is transported to peripheral sites where it is stored and released on noxious stimulation, and exerts a key role in the pathogenesis of inflammation. It is relevant to note that SP is found in abundant concentrations in the brain, gut, and lungs, where it plays an important part in immunoregulation. It is evident that SP can play a role in augmenting
M.L. CASTELLANI ET AL.

64 65Eur. J. Inflamm.
leukocytes such as macrophages, dendritic cells, T lymphocytes, B lymphocytes and mast cells, suggesting that leukocytes may be transiently innervated.
Many reports suggest that SP/NK-1R activates two convergent proinflammatory signaling pathways, PKCs and PI3K-Akt, resulting in ERK1/2 and NF-kappaB activation and chemokine production. The SP-preferring receptor neurokinin-1 receptor has two forms: a full-length NK1R (NK1R-F) isoform and a truncated NK1R (NK1R-T) isoform, which lacks the terminal cytoplasmic 96-aa residues (53-54).
Activation of NK1R-T elicited serine phosphorylation of CCR5, indicating that the communication between CCL5 and SP may occur at the level of the receptor. These observations support an important pro-inflammatory role for substance P, acting via NK1 receptors in acute inflammatory diseases and associated tissue injury (67). It has been also reported that knockout mice deficient in the preprotachykinin-A gene, the precursor gene for substance P, are also protected against acute inflammation and tissue injury. Blockade of the tachykinin NK1 receptor may therefore represent an important strategy in the treatment of patients with signs of severe neuro-inflammatory diseases.
Substance P (SP) is a potent modulator of monocyte/macrophage function and causes transcription and translation of several different cytokines/chemokines such as tumor necrosis factor-alpha (TNF-alpha), macrophage inflammatory protein-1 (MIP-1) and GM-CSF, RANTES, MCP-1, CXCL8, along with other proinflammatory compounds, proteases (chymase and tryptase), histamine, leukotrienes and prostaglandin D2 (6, 68-70).
As a mediator of pain, SP has been shown to play an important role in inflammatory states such as asthma, immune complex-mediated lung injury, experimental arthritis, and inflammatory bowel disease (71-73).
Src family kinases (SFKs) are known to be involved in cytokine signaling. It has been reported that SFKs is involved in substance P-induced chemokine production and inflammation. SFKs, specifically Src, have been widely studied also in tumorigenesis (53-54). However, recent evidence has revealed that SFKs are among the most important
both the innate and adaptive immune systems (48, 51-52).
Human lymphocytes express SP receptor and SP enhances human T cell proliferative responses (53). It has been reported that SP is capable of enhancing concanavalin A activated mononuclear cells, resulting in a strong increase in the production of antibody IgA (53-56). Moreover, SP increases the generation of this antibody in IL-5 or TGF-beta co-stimulated cultures of human T and B lymphocytes (54).
The immunomodulation by substance P includes human cell activation and proliferation, with cytokine and chemokine generation and release (57). Substance P was first isolated by Leeman S. et al. as an undecapeptide with important neurotransmitter-neuromodulator effects. In addition, substance P was shown to induce and mediate inflammation, angiogenesis, infections, intestinal mucosal immunity and stress. Substance P is able to activate several immune cells, such as CD4+ and CD8+ T lymphocytes, mast cells, NK cells and macrophages (47, 58). In addition, SP and neurokinin A appear to be chemotactic for human lymphocytes rather than monocytes (59).
In the light of these studies, we believe that substance P is important in understanding the pathophysiology of inflammation and immunity.
It has been shown that substance P by itself causes an increase of synthesis of CC and CXC chemokines in inflammatory cells. CXCL8 (IL-8) is a CXC chemokine with chemotactic and inflammatory properties. We recently found that SP is able to stimulate the release of IL-8 (CXCL8) in mast cells (26, 60-61). Since CXCL8 is a member of the CXC chemokine subfamily with potent chemotactic activity on neutrophils (62), and is a primary inflammatory cytokine, we conclude that these observations, obtained from human derived cord blood mast cell cultures (HDCBMC), a good and valid model in vitro, support the concept that the neurogenic system modulates inflammatory events by substance P and chemokine CXCL8 release (26, 63-64).
Neuropeptide SP is also released from innervating the skin sensory nerves upon exposure to several stimulants and acts via membrane-bound NK1 receptors (NK1R). This receptor is expressed by

66 67Eur. J. Inflamm.
families for the intracellular signal transduction
related to acute inflammatory responses (74). Inhibition of SFKs attenuate chemokine production and may prevent ischemia-reperfusion-induced injury, and attenuate sepsis, acute lung damage, and other organ injuries (74-77).
However, there is still much to clarify on the mechanisms utilized by the nervous system to regulate immune functions, and understanding the interrelation ship between the nervous system and the immune system will provide new information for future treatment of inflammatory and immunological diseases.
In conclusion, these studies are focused on the role of SP and cytokines, and their receptors in playing and modulating inflammatory response, and considering the possibility of potential cross-talk between a product of the central nervous system (CNS) and the immune system.
REFERENCES
1. Feuerstein G, Hallenbeck JM. Prostaglandins, leukotrienes, and platelet-activating factor in shock. Annu Rev Pharmacol Toxicol 1987; 27:301-13.
2. Cinque B, Fanini D, Di Marzio L, et al. Involvement of cPLA2 inhibition in dexamethosone-induced thymocyte apoptosis. In J Immunpathol Pharmacol 2008; 21:539-51.
3. Saria A. Neuroimmune interactions in the airways: implications for asthma, allergy, and other inflammatory airway diseases. Brain Behav Immun 1988; 2:318-21.
4. Liu D, Jiang LS, Dai LY. Substance P and its receptors in bone metabolism. Neuropeptides 2007; 41:271-83.
5. Rittner HL, Brack A, Stein C. Pain and the immune system. Br J Anaesth 2008; 101:40-4.
6. Castellani ML, Conti P, Felaco M, Vecchiet J, Ciampoli C, Cerulli G, Boscolo P, Theoharides TC. Substance P upregulates LTB in rat adherent macrophages from granuloma induced by KMnO. Neurotox Res 2009; 15:49-56.
7. Shephard RJ. Cytokine responses to physical activity, with particular reference to IL-6: sources, actions, and clinical implications. Crit Rev Immunol 2002; 22:165-82.
8. Tarantino G, Coppola A, Conca P, Cimino E, Di Minno G. Can serum TGF-beta be used to evaluate the response to antiviral therapy of haemophilic patients with HCV-related chronic hepatitis? In J Immunopathol Pharmacol 2008; 21:1007-12.
9. Martino G, Grohovaz F, Brambilla E, et al. Proinflammatory cytokines regulate antigen-independent T-cell activation by two separate calcium-signaling pathways in multiple sclerosis patients. Ann Neurol 1998; 43:340-9.
10. Calcagni E, Elenkov I. Stress system activity, innate and T helper cytokines, and susceptibility to immune-related diseases. Ann NY Acad Sci 2006; 1069:62-76.
11. Sisto M, D’Amore M, Scagliusi P, Mitolo V, Lisi S. Selective TNF-alpha gene silencing attenuates apoptosis in human salivary gland epithelial cells. Int J Immunopathol Pharmacol 2008; 21:1045-7.
12. Mitola S, Sorbello V, Ponte E, et al. Tumor necrosis factor-alpha in airway secretions from cystic fibrosis patients upregulate endothelial adhesion molecules and induce airway epithelial cell apoptosis: implications for cystic fibrosis lung disease. Int J Immunopathol Pharmacol 2008; 21:851-65.
13. Clark N, Keeble J, Fernandes ES, Starr A, Liang L, Sugden D, de Winter P, Brain SD. The transient receptor potential vanilloid 1 (TRPV1) receptor protects against the onset of sepsis after endotoxin. FASEB J 2007; 21:3747-55.
14. Piancatelli D, Bellotta L, Del Beato T, Duse M, Della Penna MR. Total IL-12 levels are increased in paediatric atopic dermatitis: correlations with age and disease severity. Int J Immunopathol Pharmacol 2008; 21:359-65.
15. Moschese V, Graziani S, Avanzini MA, et al. A prospective study on children with initial diagnosis of transient hyopogammaglobulinemia of infancy: results from the Italian Primary Immunodeficiency Network. Int J Immunopathol Pharmacol 2008; 21:343-52.
16. Bernardini R, Pucci N, Rossi ME, et al. Allergen specific nasal challenge to latex in children with latex allergy: clinical and immunological evaluation. In J Immunopathol Pharmacol 2008; 21:333-41.
17. Calcagni E, Elenkov I.Stress system activity, innate and T helper cytokines, and susceptibility to
M.L. CASTELLANI ET AL.

66 67Eur. J. Inflamm.
immune-related diseases. Ann NY Acad Sci 2006; 1069:62-76.
18. Calvisi V, Lupparelli S. C-reactive protein changes in the uncomplicated course of arthroscopic anterior cruciate ligament reconstruction. Int J Immunopathol Pharmacol 2008; 21:603-7.
19. Vivas JR, Regnault B, Michel V, et al. Interferon gamma-signature transcript profiling and IL-23 upregulation in response to Helicobacter pylori infection. Int J Immunopathol Pharmacol 2008; 21:515-26.
20. Fulgenzi A, Ticozzi P, Gabel CA, Dell’Antonio G, Quattrini A, Franzone JS, Ferrero ME. Periodate oxidized ATP (oATP) reduces hyperalgesia in mice: involvement of P2X7 receptors and implications for therapy. Int J Immunopathol Pharmacol 2008; 21:61-71.
21. Ishizuka K, Sugimura K, Homma T, et al. Influence of interleukin-10 on the interleukin-1 receptor antagonist/interleukin-1 beta ratio in the colonic mucosa of ulcerative colitis. Digestion 2001; 63:22-7.
22. Guerriero C, Manco S, Paradisi A, Capizzi R, Fossati B, Fabrizi G. Extragenital lichen sclerosus and atrophicus treated with topical steroids and retinoids in a child with vitiligo. Int J Immunopathol Pharmacol 2008; 21:757-9.
23. Cordiali-Fei P, Ardigò M, Mastroianni A, et al. Serum cytokines and vioumoral immunological characterization of psoriatic patients in long term etanercept treatment. In J Immunopathol Pharmacol 2008; 21:643-9.
24. Stratta P, Marcuccio C, Campo A, et al. Improvement in relative survival of patients with vasculitis: study of 101 cases compared to the general population. Int J Immunopathol Pharmacol 2008; 21:631-42.
25. Kasai T, Inada K, Takakuwa T, et al. Anti-inflammatory cytokine levels in patients with septic shock. Res Commun Mol Pathol Pharmacol 1997; 98:34-42.
26. Castellani ML, Shaik YB, Perrella A, et al. Expression and secretion of CXCL8 (IL-8), release of tryptase and transcription of histidine decarboxylase mRNA by anti-IgE-activated human umbilical cord blood-derived cultured mast cells. Neuroimmunomodulation 2007; 14:97-104.
27. Marcucci F, Incorvaia C, Sensi L, et al. Lack of
inflammatory cells in the oral mucosa of subjects undergoing sublingual immunotherapy. Int J Immunopathol Pharmacol 2008; 21:609-13.
28. Galliera E, Locati M, Mantovani A, Corsi MM. Chemokines and bone remodelling. Int J Immunopathol Pharmacol 2008; 21:585-91.
29. Castellani ML, Ciampoli C, Felaco M, et al. Neuropeptide substance P induces mRNA expression and secretion of CXCL8 chemokine, and HDC in human umbilical cord blood mast cells. Clin Invest Med 2008; 31:E362-72.
30. Di Gioacchino M, Perrone A, Petrarca C, et al. Early cytokine modulation after the repid induction phase of sublingual immunotherapy with monomeric allergoids. Int J Immunopathol Pharmacol 2008; 21:969-76.
31. Bost KL. Tachykinin-mediated modulation of the immune response. Front Biosci 2004; 9:3331-2.
32. Koon HW, Zhao D, Zhan Y, Simeonidis S, Moyer MP, Pothoulakis C. Substance P-stimulated interleukin-8 expression in human colonic epithelial cells involves protein kinase Cdelta activation. J Pharmacol Exp Ther 2005; 314:1393-400.
33. Inoue K, Takano H, Yanagisawa R, et al. Antioxidative role of interleukin-6 in septic lung injury in mice. Int J Immunopathol Pharmacol 2008; 21:501-7.
34. Kulka M, Sheen CH, Tancowny BP, Grammer LC, Schleimer RP. Neuropeptides activate human mast cell degranulation and chemokine production. Immunology 2008; 123:398-410.
35. Dunzendorfer S, Kaser A, Meierhofer C, Tilg H, Wiedermann CJ. Cutting edge: peripheral neuropeptides attract immature and arrest mature blood-derived dendritic cells. J Immunol 2001; 166:2167-72.
36. Ciardelli L, Garofoli F, Stronati M, et al. Human colostrum T lymphocytes and their effector cytokines actively aid the development of the newborn immune system. Int J Immunopathol Pharmacol 2008; 21:781-6.
37. Kulka M, Sheen CH, Tancowny BP, Grammer LC, Schleimer RP. Neuropeptides activate human mast cell degranulation and chemokine production. Immunology 2008; 123:398-410.
38. Vojdani A. Antibodies as predictors of complex

68 69Eur. J. Inflamm.
49. Papahariadou M, Kalogeromitros D, Athanasiadis GI, et al. Autoimmunity and parasites. Int J Immunopathol Pharmacol 2008; 21:5-9.
50. Theoharides TC, Makris M, Kalogeromitros D. Allergic inflammation and adipocytokines. Int J Immunopathol Pharmacol 2008; 21:1-4.
51. Gelardi M, Maselli del Giudice A, Fiorella ML, Fiorella R, Russo C, Soleti P, Di Gioacchino M, Ciprandi G. Non-allergic rhinitis with eosinophils and mast cells constitutes a new severe nasal disorder. Int J Immunopathol Pharmacol 2008; 21:325-31.
52. De Berardis D, Conti CM, Campanella D, et al. Evaluation of C-reactive proetin and total serum cholesterol in adult patients with bipolar disorder. Int J Immunopathol Pharmacol 2008; 21:319-24.
53. Bost KL. Tachykinin-mediated modulation of the immune response. Front Biosci 2004; 9:3331-2.
54. Stanisz AM, Scicchitano R, Dazin P, Bienenstock J, Payan DG. Distribution of substance P receptors on murine spleen and Peyer’s patch T and B cells. J Immunol 1987; 139:749-54.
55. Izzi V, Chiurchiù V, D’Aquilio F, Martino A, Tresoldi I, Modesti A, Baldini PM. Endomorphin-1 inhibits the activation and the development of a hyporesponsive-like phenotype in lipopolysaccharide-stimulated THP-1 monocytes. Int J Immunopathol Pharmacol 2008; 21:833-43.
56. Chan CP, Lei HY. Autophagy induction in T cell-independent acute hepatitis induced by concanavalin A in SCID/NOD mice. Int J Immunopathol Pharmacol 2008; 21:817-26.
57. Minardi D, Ghiselli R, Lucarini G, et al. Activity and expression of nitrix oside sythase in rat bladder after sacral neurmodulation. Int J Immunopathol Pharmacol 2008; 21:129-35.
58. Caorsi C, Cappello P, Ceruti P, Amici A, Marchini C, Novelli F, Forni G, Giovarelli M. CCL 16 enhances the CD8+ and CD4+ cell reactivity to human HER-2 elicited by dendritic cells loaded with rat ortholog HER-2. Int J Immunopathol Pharmacol 2008; 21:867-77.
59. Lalle E, Sacchi A, Abbate I, et al. Activation of interferon response genes and of plasmacytoid dendritic cells in HIV-1 positive subjects with GB virus C co-infection. Int J Immunopathol Pharmacol
autoimmune diseases and cancer. Int J Immunopathol Pharmacol 2008; 21:553-66
39. Nitschke M, Sohn K, Dieckmann D, Gibbs BF, Wolff HH, Amon U. Effects of basophil-priming and stimulating cytokines on histamine release from isolated human skin mast cells. Arch Dermatol Res 1996; 288:463-8.
40. Hauswith AW, Excribano L, Prados A, et al. CD203c is overexpressed on neoplastic mast cells in systemic mastocytosis and is upregulated upon IgE receptor cross-linking. Int J Immunopathol Pharmacol 2008; 21:797-806.
41. Fiebich BL, Schleicher S, Butcher RD, Craig A, Lieb K. The neuropeptide substance P activates p38 mitogen-activated protein kinase resulting in IL-6 expression independently from NF-kappa B. J Immunol 2000; 165:5606-11.
42. Misery L. Skin, immunity and the nervous system. Br J Dermatol 1997; 137:843-50.
43. Noguchi K, Yamashiro S, Matsuzaki T, Sakanashi M, Nakasone J, Miyagi K, Sakanashi M. Effect of 1-week treatment with erythropoietin on the vascular endothelial function in anaesthetized rabbits. Br J Pharmacol 2001; 133:395-405.
44. Speranza L, Franceschelli S, Pesce M, Vinciguerra I, De Lutiis MA, Grilli A, Felaco M, Patruno A. Phosphodiesterase type-5 inhibitor and oxidative stress. Int J Immunopathol Pharmacol 2008; 21:879-89.
45. Peters EM, Ericson ME, Hosoi J, Seiffert K, Hordinsky MK, Ansel JC, Paus R, Scholzen TE. Neuropeptide control mechanisms in cutaneous biology: physiological and clinical significance. J Invest Dermatol 2006; 126:1937-47.
46. Levite M. Neurotransmitters activate T-cells and elicit crucial functions via neurotransmitter receptors. Curr Opin Pharmacol 2008; 8:460-71.
47. Putignani L, Antonucci G, Paglia MG, Vincenzi L, Festa A, De Mori P, Loiacono L, Visca P. Cryptococcal lymphadenitis as a manifestation of immune reconstitution inflammatory sindrome in an HIV-positive patient: a case report and review of the literature. Int J Immunopathol Pharmacol 2008; 21:751-6.
48. Pascual DW. The role of tachykinins on bacterial infections. Front Biosci 2004; 9:3209-17.
M.L. CASTELLANI ET AL.

68 69Eur. J. Inflamm.
2008; 21:161-71.60. Kalogeromitros C, Makris M, Chliva C, Aggelides X,
Kempuraj D, Theoharides TC. A quercetin containing supplement reduces niacin-induced flush in humans. Int J Immunopathol Pharmacol 2008; 21:s509-14.
61. Patriarca G, Schiavino D, Lombardo C, Altomonte G, De Cinti M, Buonuomo A, Numera E. Tolerability of aztreonam in patients with IgE-mediated hypersenitivity to beta-lactams. Int J Immunopathol Pharmacol 2008; 21:375-9.
62. Capodicasa E, Cornacchine P, Natalizi B, Bartolo A, Coaccioli S, Marconi P, Scaringi L. Omeprazole induces apoptosis in normal human polymorphonuclear leucocytes. Int J Immunopathol Pharmacol 2008; 21:73-85.
63. Pifferi M, Caramella D, Ragazzo V, Cangiotti AM, Macchia P, Boner AL. Bronchiectasis in children with recurrent pneumonia: an immunopathological damage associated with secondary cilary dysmotility Int J Immunopathol Pharmacol 2008; 21:215-9.
64. De Vita G, Patti R, Sparacello M, Balistreri CR, Candore G, Caruso C. Impact of different texture of polypropylene mesh on the inflammatory response. Int J Immunopathol Pharmacol 2008; 21:207-14.
65. Katsanos GS, Anogianaki A, Castellani M, et al. Biology of neurotensin: revisited study. Int J Immunopathol Pharmacol 2008; 21:255-9.
66. Sonneck K, Baumgartner C, Rebuzzi I, et al. Recomvinant allergens promote expression of aminopeptidase-n (CD13) on basophils in allergic patients. Int J Immunopathol Pharmacol 2008; 21:11-21.
67. Ciccocioppo F, Lanuti P, Marchisio M, et al. Expression and phosphorylation of protein kinase C isoforms in Abeta(1-42) activated T lymphocytes from Alzheimer’s disease. Int J Immunopathol Pharmacol 2008; 21:23-33.
68. Minardi D, Lucarini G, Filoa A, Milanese G, Zizzi A, Di Primio R, Montironi R, Muzzonigro G. Prognostic role of tumor necrosis, microvessel density, bascular endothelial growth factory and hypoxia inducile factor-1alpha in patients with clear cell renal carcinoma after radical nephrectomy in a longterm
follow-up. Int J Immunopathol Pharmacol 2008; 21:447-55.
69. Raffaelli L, Serini S, Piccioni E, Manicone PF, Berardi D, Perfetti G, Calviello G. N-3 plyunsaturated fatty acid effect in periodontal disease: sstate of art and possibile mechanisms involved. Int J Immunopathol Pharmacol 2008; 21: 261-6.
70. Colanardi MC, Nettis E, Traetta P, Delle Donne P, Ferrannini A, Vacca A. Parecoxib a san alternative in COX-2 hypersensitivity. Int J Immunopathol Pharmacol 2008; 21:233-5.
71. Capsoni F, Ongari AM, Frigerio E, Taglioni M, Altomare GF. Effect of Efalizumab on neutrophil and monocyte functions in patients with psoriasis. Int J Immunopathol Pharmacol 2008; 21:437-45
72. Ferdinande L, Demetter P, Perez-Novo C, Waeytens A, Taildeman J, Rottiers I, Rottiers P, De Vos M, Cuvelier CA. Inflamed intestinal mucosa features a specific epthelial expression pattern of indoleamine 2,3-dioxygenase. Int J Immunopathol Pharmacol 2008; 21:289-95.
73. Vojdani A. Antibodies as predictors of complex autoimmune diseases. Int J Immunopathol Pharmacol 2008; 21:267-78.
74. Balthasar N, Coppari R, McMinn J, et al. Leptin receptor signalling in POMC neurons is required for normal body weight homeostasis. Neuron 2004; 42:983-91.
75. Inoue K, Takano H, Ohnuki M, Yanagisawa R, Sakurai M, Shimada A, Mizushima K, Yoshikawa T. Size effects of nanomaterials on lung inflammation and coagulatory disturbance. Int J Immunopathol Pharmacol 2008; 21:197-206
76. Capoluongo E, Vwento G, Lulli P, et al. Epithelial lining fluid neutrophil-gelatinase-associated lipocalin levels in premature newborns with bronchopulmonary dysplasia and patency of ductus arteriosus. Int J Immunopathol Pharmacol 2008; 21:173-9.
77. Cardelli P, Ferraironi M, Amodeo R, et al. Evaluation of neutrophil CD64 expression and preocalcitonin as useful markers in early diagnosis of sepsis. Int J Immunopathol Pharmacol 2008; 21:43-9.

EUROPEAN JOURNAL OF INFLAMMATION Vol. 7, no. 2, 0-0 (2009)
1721-727X (2009)Copyright © by BIOLIFE, s.a.s.
This publication and/or article is for individual use only and may not be furtherreproduced without written permission from the copyright holder.
Unauthorized reproduction may result in financial and other penalties71

EUROPEAN JOURNAL OF INFLAMMATION Vol. 7, no. 2, 0-0 (2009)
1721-727X (2009)Copyright © by BIOLIFE, s.a.s.
This publication and/or article is for individual use only and may not be furtherreproduced without written permission from the copyright holder.
Unauthorized reproduction may result in financial and other penalties71
Dopamine (DA) is a monoamine neurotransmitter which plays a crucial role in the development and maintenance of the nervous system. In adults, dopamine regulates motor functions, is involved in emotional response and is central to the reward system. DA exerts its effects via G-protein coupled receptors (1).
The family of dopamine receptors consists of five members: D1, D2, D3, D4, and D5 (1). They are divided into two main categories: D1-like (D1,
D5) and D2-like (D2, D3, D4) receptors (2-3). These proteins are expressed throughout the brain but each subtype has a unique pattern of expression.
In spite of all the pharmacological treatments for schizophrenia, the proportion of treatment-resistant schizophrenia has been estimated as 20–40% in the schizophrenic patients, and this unfortunate situation in the clinical psychiatry field still remains unchanged, even after the introduction of several atypical antipsychotic agents (4).
DOPAMINE RECEPTOR GENE EXPRESSION CHANGES IN PERIPHERAL BLOOD MONONUCLEAR CELLS FROM SCHIZOPHRENIC PATIENTS TREATED WITH
HALOPERIDOL AND OLANZAPINE
G.H. SHARIATI1,2, G. AHANGARI1, M.R. ASADI3, F. POYAFARD3 and H.R. AHMADKHANIHA4
1Department of Medical Genetics, National Institute for Genetic Engineering and Biotechnology(NIGEB), Tehran; 2Department of Genetic & Biology, Jundishahpour Medical
Science University, Ahwaz; 3Department of Psychiatric, Rozbeh Hospital, Tehran Medical University, Tehran; 4Tehran Psychiatric Institute, Iran University of Medical Sciences, Tehran;
World Health Organization Collaborating Center for Mental Health, Tehran, Iran
Received January 12, 2009 – Accepted May 27, 2009
We investigated dopamine receptor gene expression in peripheral blood mononuclear cells of schizophrenic patients before and after treatment. Also dopamine receptor genes expression profile was compared in two treatment groups including haloperidol and olanzapine. The peripheral blood mononuclear cells were separated from whole blood by Ficoll-hypaque; the total cellular RNA was extracted and the cDNA was synthesized. This process was followed by real-time polymerase chain reaction using primer pairs specific for five dopamine receptor mRNAs and β-actin as internal control. The results show the presence of all types of dopamine receptor in lymphocytes. Dopamine receptor gene expression profile in dopamine receptor D2 gene and dopamine receptor D4 gene showed significant changes that were correlated with the type of treatment and Clinical Global Impressions score improvement. In conclusion, the present study shows that human lymphocytes express dopamine receptor D1-D5 genes. Moreover, investigated dopamine receptors gene expression in peripheral blood mononuclear cells of schizophrenic patients correlated with clinical symptom improvement.
Mailing address: G. Ahangari, MT, Ph.D., Department of Molecular Medicine and Immunology, National Institute for Genetic engineering and Biotechnology, P.O. Box 14155, 6343 Tehran, IranTel: ++98 214580384 Fax: ++98 2144580399e-mail: [email protected]
Key words: dopamine gene receptor, gene expression, schizophrenia

72 73Eur. J. Inflamm.
Clinical studies have shown that antipsychotics can improve symptoms of schizophrenia, but the molecular mechanism of these drugs has remained elusive. One of the important problems of this investigation is simple non-invasive model designing to study gene expression during treatments.
Blood products and peripheral blood mononuclear cells (BPMC) are readily accessible and may reflect molecular processes in the central nervous system of schizophrenic patients (5-6), although the validity of peripheral markers is still under debate. Substances in serum and PBMC that have been previously studied in schizophrenia include members related to the therapy, to receptors for major neurotransmitters and G-protein subunits which regulate intracellular signal transduction (6-7).
This study seeks to examine whether human PBMC could be useful in the study of the mechanism of antipsychotic treatment in schizophrenia patients. Based on this model we compared the effects of haloperidol and olanzapine on schizophrenia symptom improvements and also determined the changes of dopamine receptor gene expression profiles.
MATERIALS AND METHODS
Twenty schizophrenia patients (age 20-35 years) before and after medication took part in this study. A reduction of at least one category on the Clinical Global Impressions (CGI) scale may be considered a response to treatment. Written informed consent was obtained from each individual. Peripheral blood samples (4 ml) were obtained from the cubital vein and collected in cell preparation tubes containing an anticoagulant. Peripheral blood mononuclear cells were isolated by Ficoll-Hypaque density centrifugation (Pharmacia. Uppsala, Sweden). The lymphocyte layer was collected and washed three times in Phosphate buffer saline (PBS). The total mRNA was isolated from lymphocytes by RNA blood minikit (Roach, Germany), and the amount and purity of RNA were determined by spectrophotometry (8). Dopamine receptor mRNA expression was determined by the 3’ 5’ fluorogenic Taqman approach (Roach, Germany). Total (RNA of 1500 ng was reverse transcribed into first-strand cDNA by using oligonucleotide dT primers and 2.5 units of multiscribe (recombinant Maloney murine leukemia virus) reverse transcriptase in a final volume of 40 μL (9). Primers for DR1, DR2, DR3, DR4,DR5 and housekeeping gene β-actin were designed using primer
express software to exclude amplification of genomic DNA and pseudo genes (Table I). cDNA of 75 ng was used for PCR amplification in a final volume of 25 μl with 1 unit of Taq DNA polymerase (Roach, Germany). PCR was carried out in a Real-time-PCR (Roach, Germany) with a Cyber green fluorogenic nucleotide to monitor cDNA amplification by the increase in fluorescence intensity. Each PCR product of dopamine receptor (DRD1-DRD5) was sequenced by DNA sequencer ABI 3700 capillary system (Applied Biosystem, USA). The statistical analyses were performed with the software package SPSS 16. The Student T test was used to compare relative gene expression and GCI score in two treatment groups. Statistical significance was defined as p<0.05.
RESULTS
In this study we examined dopamine receptor gene expression changes in the PMC of schizophrenic patients during two treatment periods lasting 4 weeks. Blood samples were taken and the clinical state was assessed at baseline, and after 4 weeks of treatment, so that each patient served as his own control. We focused on all subtypes of dopamine receptors (DRD1-DRD5).The experiments were performed using peripheral blood lymphocytes. Expression of the different dopamine receptors gene segments was studied by analyzing total RNA extracted from the samples. In order to detect dopamine gene receptor expression on the RNA level, Real time PCR was performed for the regions of different dopamine receptors.
The results presented here provide direct evidence that human lymphocytes express dopamine D1 and D2-like receptors belonging to DRD1 to DRD5 receptor subtypes, respectively, in PBL of schizophrenia patients. The taqman assays were tested by blasting against the entire human genome (NCBI-national Center for Biotechnology Information 2008) to exclude sequencing at unwarranted sites (10). The specificities of the obtained PCR products for the respective dopamine receptor fragments were confirmed by a capillary sequenced analysis ABI 3700 machine.
Gene expression changes with treatment were determined per patient, relative to his own baseline mRNA level. The inter-subject comparison reduction is due to inter individual variability and illness heterogeneity factors and focuses on
G.H. SHARIATI ET AL.

72 73Eur. J. Inflamm.
treatment-related changes. In follow-up treatments, significant changes were observed with mean CGI score (p=0.01). Extra pyramidal side effects were absent in all patients. No significant differences were found in the CGI scales between the olanzapine and haloperidol treated sub-groups of patients. Dopamine receptor gene expressions were shown in Table II. Changes of dopamine receptor genes based on the type of treatments are shown in Figs. 1 and 2. No statistically significant differences were found in the dopamine receptor gene expression before treatment between patients assigned to olanzapine and haloperidol groups. However, mean endpoint of D4 and D5 receptors genes were significantly higher in haloperidol treated patients than in olanzapine treated patients (P=0.04). Also, the mean endpoint of D2 receptor genes were significantly lower in haloperidol treated patients than in olanzapine treated patients (P=0.01).
The mean CGI score changes in haloperidol
treated patients were correlated with changes of D1 receptor gene expression (r=0.76, p=0.04) but no statistically significant differences were found in the other dopamine receptors gene expressions in this group. The mean of CGI score changes in olanzapine treated patients was correlated with changes of D4 and D5 receptor gene expressions (D4; r=0.8, p=0.005, D5; r=0.8, p=0.005) but no statistically significant differences were found in the other dopamine receptor gene expressions in this group.
DISCUSSION
We found consistent gene expression changes in PMC of two schizophrenic patient treatment groups. This supports our suggestion that PBMC may be useful in investigating the mechanism of action of these drugs in clinical settings and is in accordance with the literature (5-7). Moreover, the strength of
Table I. Primer sequences and product size.
Primer ReversePrimer ForwardProductACCESSIONLOCUS
GAT GAA TTA GCC CAC CCA AAC CAAACCCACAAGC CCC TCT GAT G471 bpNM_000794DRD1
AAGGGCACG TAG AAG GAG ACGCGGAC AGA CCC CAC TAC AA521 bpNM_016574DRD2
AGTGGCACTCCCCGAGGTTGGCCGCATTTGCTGTGATGTTTTT670 bpU32499DRD3
GGAAGGCCCCGACCACCACCCTGCGGCTCCAACTGTGCT153 bpNM_000797DRD4
GCAGATCCATGAGGGGGTTTAACCTGTGCGTCATCAGCGT107 bpNM_000798DRD5
Table I. Primer sequences and product size.
Table II. Gene expression of dopamine receptor in two treatments groups.
Haloperidol** Olanzapine** Drug
Dopamine receptor gene Baseline* After treatment* P Value Baseline* After treatment* P ValueD1 3.55 ± 6.22 5.38 ± 8.44 0.63 9.67 ±1.48 3.43 ±3.75 0.08
D2 0.11± 0.08 0.01 ± 0.01 0.04 0.07 ±0.01 0.07 ±0.06 0.98
D3 92.88 ± 39.33 56.22 ± 48.45 0.10 29.82 ±39.37 41.70 ±43.07 0.51
D4 1.44 ± 3.69 2.10 ± 3.57 0.35 7.62 ±1.43 1.34 ±3.09 0.18
D5 3.18 ± 1.99 2.86 ± 3.77 0.85 2.44 ±1.63 1.27 ±2.84 0.17
* Gene expression of dopamine receptor that calculated as target gene concentration divided to beta actin concentration. ** Antipsychotic therapy for 4 weeks
Table II. Gene expression of dopamine receptor in two treatments groups.
* Gene expression of dopamine receptor that calculated as target gene concentration divided to beta actin concentration.** Antipsychotic therapy for 4 weeks
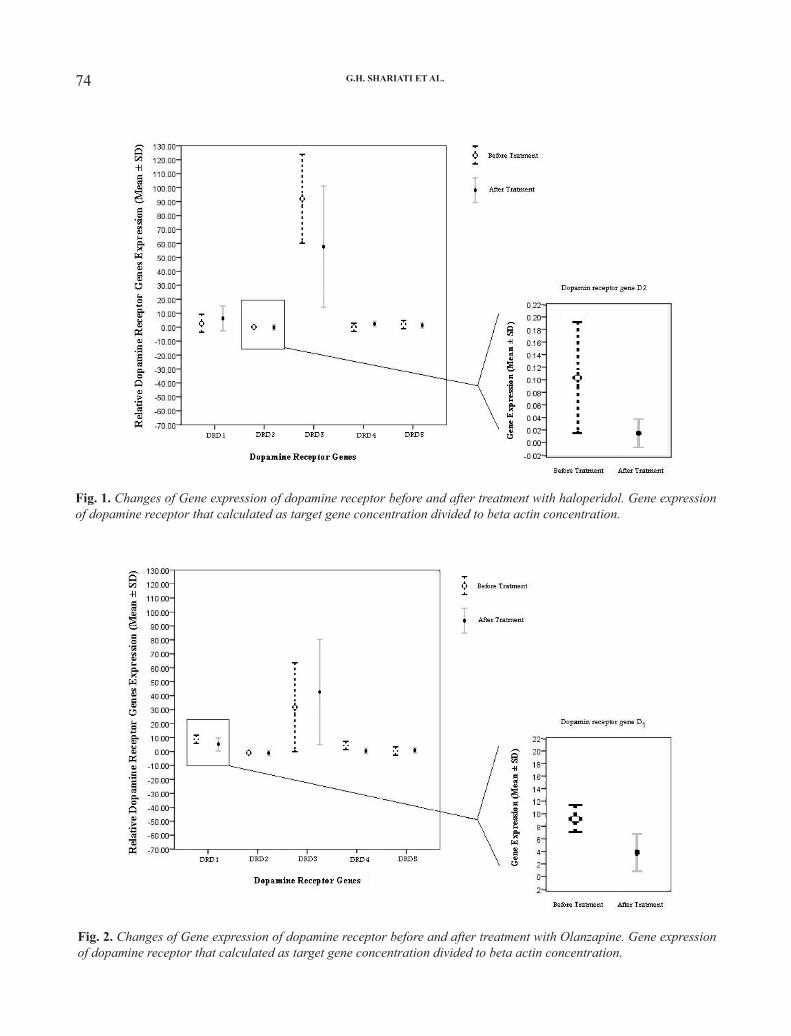
74 75Eur. J. Inflamm.
Fig. 1. Changes of Gene expression of dopamine receptor before and after treatment with haloperidol. Gene expression of dopamine receptor that calculated as target gene concentration divided to beta actin concentration.
Fig. 2. Changes of Gene expression of dopamine receptor before and after treatment with Olanzapine. Gene expression of dopamine receptor that calculated as target gene concentration divided to beta actin concentration.
G.H. SHARIATI ET AL.

74 75Eur. J. Inflamm.
the procedure established in this work was the within subject design, such that each of the patients served as their own controls. This reduced the potential problems due to the heterogeneity of schizophrenic illness and highlights the treatment-related changes.
While the validity of peripheral changes in gene expressions as indicators of brain processes is still debated, there is evidence of crosstalk between neurotransmitters and immune-related proteins in brain and blood (5-7, 12), and serotonin receptors (13) can be synthesized and function within the central nervous system, as well as in lymphocytes. This fulfills a fundamental condition for correlation between brain and periphery, i.e. the criterion of expression of gene in both compartments (7, 14). Taken together, it is plausible to consider that the peripheral gene changes observed in this study following treatment may reflect, at least in part, relevant brain processes.
Subsequently, we investigated the changes in dopamine receptor gene transcript profiles in two treatment groups in relation to clinical symptom improvements. In this study the relationship between PMC gene expression changes and clinical improvement was determined. Previous controlled studies demonstrated this relationship especially in improvement in primary negative symptoms (15). This suggests that PMC molecular markers may be useful in investigation of the mechanism of action of treatment over time. The PMC gene expression changes occurred in parallel to clinical improvements in schizophrenic patients consistent with the study of Kirilova et al. (16). Also, the profile of dopamine receptor genes may be specific with type of treatments that report similar results in other studies (17-18). Of course, larger and more controlled studies with a broader range of schizophrenia patients to clarify the significance of these changes for clinical efficacy and understanding of the molecular mechanism of antipsychotic treatment is recommend.
ACKNOWLEDGEMENTS
We are indebted to Professor K. Smith (Boston University) for his scientific support and helpful suggestions and Dr Arash Hossein-nezhad Shirvani for his help. The author gratefully acknowledges the support of the National Institute for Genetic
Engineering and Biotechnology for grant number: 0249, RAZI Psychiatric Hospital, Tehran Medical Science University, Tehran Psychiatric Institute, Iran Medical Science University and all the participants in this study.
REFERENCES
1. AR Katritzky, Dobchev DA, Stoyanova-Slavova IB, et al. Novel computational models for predicting dopamine interactions. Exp Neurol 2008; 211:150-71.
2. Grandy DK, Marchionni MA, Makam H, et al. Cloning of the cDNA and Gene for a human D2 dopamine receptor. Proc Natl Acad Sci USA 1989; 86:9762-6.
3. Van Tol HH, Bunzow JR, Guan HC, Sunahara RK, Seeman P, Niznik HB, Civelli O. Cloning of the gene for a human dopamineD4 receptor with high affinity for the antipsychotic clozapine. Nature 1991; 350:610-4.
4. Weiss EL, Longhurst JG, Bowers MB, Mazure CM. Olanzapine for treatment-refractory psychosis in patients responsive to clozapine. J Clin Psychopharmacol 1999; 19:378-80.
5. Kronfol Z, Remick DG. Cytokines and the brain: implications for clinical psychiatry. Am J Psychiatry 2000; 157:683-94.
6. Bowden NA, Weidenhofer J, Scott RJ, et al. Preliminary investigation of gene expression profiles in peripheral blood lymphocytes in schizophrenia. Schizophr Res 2006; 82:175-83.
7. Chertkow Y, Weinreb O, Youdim MB, Silver H. Gene expression changes in peripheral mononuclear cells from schizophrenic patients treated with a combination of antipsychotic with fluvoxamine. Prog Neuropsychopharmacol Biol Psychiatry 2007; 31:1356-62.
8. Ahangari G, Tehrani M, Halapi E, Hammar H, Wigzell H. RT-PCR based analysis of T-cell receptor B variable region gene usage in normal human SKIN. Scand J Immunol 1996; 44:330-4.
9. Sambrook J, Fritsch EF, Maniatis T. Molecular cloning. A laboratory manual, Cold Spring Harbor Laboratory Press USA 2003; 433-527.
10. NCBI-National center for biotechnology information, NCBI Blast home page, Http://

76 EUROPEAN JOURNAL OF INFLAMMATION Vol. 7, no. 2, 0-0 (2009)
1721-727X (2009)Copyright © by BIOLIFE, s.a.s.
This publication and/or article is for individual use only and may not be furtherreproduced without written permission from the copyright holder.
Unauthorized reproduction may result in financial and other penalties77
www.ncbi.nlm.nih.gov/Blast11. Gladkevich A, Nelemans SA, Kauffman HF, Korf
J. Microarray profiling of lymphocytes in internal diseases with an altered immune response: potential and methodology. Mediators Inflamm 2005; 6:317-30.
12. Miquel Bernardo, Eduard Parellada, Francisco Lomen, et al. Double-blind olanzapine vs. haloperidol D2 dopamine receptor blockade in schizophrenic patients: a baseline-endpoint I IBZM SPECT study. Psychiatry Research: Neuroimaging Section 2001; 107:87-97.
13. Grimaldi B, Fillion G. 5-HT-moduline controls serotonergic activity: implication in neuroimmune reciprocal regulation mechanisms. Prog Neurobiol 2000; 60:1-12.
14. Sullivan PF, Fan C, Perou CM. Evaluating the comparability of gene expression in blood and brain. Am J Med Genet B Neuropsychiatr Genet 2006; 141:261-8.
15. Silver H, Nassar A, Aharon N, Kaplan A. The onset and time course of response of negative symptoms to add-on fluvoxamine treatment. Int Clin Psychopharmacol 2003; 18:87-92.
16. Kirillova GP, Hrutkay RJ, Shurin MR, Shurin GV, Tourkova IL, Vanyukov MM. Dopamine receptors in human lymphocytes: Radioligand binding and quantitative RT-PCR assays. J Neurosci Methods 2008; 174:272-80.
17. Bertran-Gonzalez J, Bosch C, Maroteaux M, Matamales M, Hervé D, Valjent E, Girault JA. Opposing patterns of signaling activation in dopamine D1 and D2 receptor-expressing striatal neurons in response to cocaine and haloperidol. J Neurosci 2008; 28:5671-85.
18. Padín JF, Rodríguez MA, Domínguez E, et al. Parallel regulation by olanzapine of the patterns of expression of 5-HT2A and D3 receptors in rat central nervous system and blood cells. Neuropharmacology 2006; 51:923-32.
G.H. SHARIATI ET AL.

76 EUROPEAN JOURNAL OF INFLAMMATION Vol. 7, no. 2, 0-0 (2009)
1721-727X (2009)Copyright © by BIOLIFE, s.a.s.
This publication and/or article is for individual use only and may not be furtherreproduced without written permission from the copyright holder.
Unauthorized reproduction may result in financial and other penalties77
The dual roles of heat-shock proteins (HSPs) in the immune system are still a matter of scientific debate
(1). Due to their potential use as immunomodulators in a variety of pathological processes, special attention
NO EVIDENCE OF PATHOLOGICAL AUTOIMMUNITY FOLLOWING MYCOBACTERIUM LEPRAE HEAT-SHOCK PROTEIN 65-DNA VACCINATION
IN MICE
D.S. LIMA, C.R. ZÁRATE-BLADÉS, P.R.M. SOUZA, A.P. TROMBONE,R.R. SANTOS-JUNIOR1, I.T. BRANDÃO, A.P. MASSON, V.L. BONATO,
A.A.M. COELHO-CASTELO, A. SARTORI2, M. VENDRAMINI3, E.G. SOARES4,L.A. BENVENUTTI3, C.L. SILVA and V. COELHO5,6
The Centre for Tuberculosis Research, Department of Biochemistry and Immunology, Medicine School of Ribeirão Preto, University of São Paulo, São Paulo; 1Department of Clinical Analyses,
Faculty of Pharmaceutical Sciences, São Paulo State University, Araraquara, São Paulo;2Department of Microbiology and Immunology, Biosciences Institute, São Paulo State University,
Botucatu, São Paulo; 3Department of Pathology, University of São Paulo Medical School, São Paulo; 4Department of Pathology, School of Medicine of Ribeirão Preto, University of São Paulo, Ribeirão Preto, São Paulo; 5Heart Institute (InCor), University of São Paulo Medical School, São Paulo; 6Institute for Investigation in Immunology, National Institute of Science and Technology -
INCT, São Paulo, Brazil
Received March 18, 2009 – May 26, 2009
Heat-shock proteins (HSPs) are currently one of the most promising targets for the development of immunotherapy against tumours and autoimmune disorders. This protein family has the capacity to activate or modulate the function of different immune system cells. They induce the activation of monocytes, macrophages and dendritic cells, and contribute to cross-priming, an important mechanism of presentation of exogenous antigen in the context of MHC class I molecules. These various immunological properties of HSP have encouraged their use in several clinical trials. Nevertheless, an important issue regarding these proteins is whether the high homology among HSPs across different species may trigger the breakdown of immune tolerance and induce autoimmune diseases. We have developed a DNA vaccine codifying the Mycobacterium leprae Hsp65 (DNAhsp65), which showed to be highly immunogenic and protective against experimental tuberculosis. Here, we address the question of whether DNAhsp65 immunization could induce pathological autoimmunity in mice. Our results show that DNAhsp65 vaccination induced antibodies that can recognize the human Hsp60 but did not induce harmful effects in 16 different organs analysed by histopathology up to 210 days after vaccination. We also showed that anti-DNA antibodies were not elicited after DNA vaccination. The results are important for the development of both HSP and DNA-based immunomodulatory agents.
Mailing address: Verônica Coelho,Instituto do Coração, Laboratório de Imunologia,Faculdade de Medicina da Universidade de São Paulo, Av Dr Enéas de Carvalho Aguiar, 44 bloco II, 9 andar -Cerqueira César 05403-001 São Paulo, SP, BrasilTel: ++55 11 30695905 Fax: ++55 11 30829350 e-mail: [email protected]
Key words: autoantibodies, DNA vaccine, heat-shock protein, autoimmunity

78 79Eur. J. Inflamm.
has recently been focused on HSP immunobiology. Accordingly, HSPs have been tested in prophylactic and therapeutic experimental models of different types of diseases (2-4), and some of these proteins, or derived peptides, have been tested in clinical trials (5-6). In addition to their well-known functions as molecular chaperones, HSPs have been shown to act as immunodominant antigens and to play a role in the activation of different cell types, cytokine secretion and cross-presentation of tumour antigens (6-8). Nonetheless, various HSPs were related to the development of autoimmune diseases (9-10). It is thought that peptides derived from foreign HSPs (e.g., bacterial HSPs) could induce cross-reactivity against the human HSP homolog leading to breakdown of tolerance, thereby triggering autoimmune diseases (11). Therefore, any attempt to use HSPs as immunomodulators must consider this and should include the evaluation of the harmful potentiality of these molecules.
Several reports, from our group and others, have described the use of the Mycobacterium leprae Hsp65 gene in a DNA vaccine for the prevention and treatment of tuberculosis in different animal models (12-14). Moreover, the use of this vaccine in combination with chemotherapy was effective in the treatment of multi-drug resistant tuberculosis. This association is particularly relevant as an alternative to circumvent the limitations of traditional therapies (15-17).
Despite the success of DNAhsp65 immunization in different animal models and the lack of major toxicity detected in either the pre-clinical studies or in a phase I clinical trial (18-23), it has been suggested that this vaccine could cause necrosis in the lungs of vaccinated animals (24-26). These results have not been observed by other groups and, on the contrary, several subsequent reports have provided increasing evidence in favour of the potential therapeutic use of Hsp65 against tuberculosis (13-14, 16). However, it is mandatory to thoroughly exclude any possible harmful effects mediated by this vaccine, including the development of autoimmune diseases. In light of this, we analysed the development of potentially pathological autoimmune responses, as well as tissue damage, following DNAhsp65 immunization in healthy mice. We observed that the DNAhsp65 vaccine induced antibodies that can recognize both
mycobacterium Hsp65 and human Hsp60. However, no anti-DNA antibodies were induced. Additionally, the animals did not present clinical features of autoimmune diseases and no significant tissue lesions were detected following the histopathological analysis of 16 different organs, up to 210 days post-vaccination.
MATERIALS AND METHODS
Animals and immunizationSix- to eight-week-old, specific pathogen-free female
BALB/c mice were obtained from the local breeding facility of the University of São Paulo at Ribeirão Preto, Brazil. Mice were manipulated according to the Brazilian Committee for Animal Care and Use (COBEA) guidelines. Mice were divided into three groups: 1) pVAX1-hsp65 (vaccine group), 2) pVAX1 (vector group), and 3) phosphate buffered saline (control group). Mice were intramuscularly immunized with three doses of 100 µg of plasmid, given at two-week intervals. We analysed three animals per group in three independent experiments, totalizing nine mice per group. The mice were euthanized 45, 75 or 210 days following the start of immunization.
Detection of anti-HSP antibodiesThe levels of the anti-Hsp65 and anti-Hsp60 antibodies
were determined in mouse sera using ELISA (BD Biosciences, San Diego, CA). Plates (Maxisorp Nunc-Immuno; Roskilde, Denmark) were coated with Hsp65 or Hsp60 (5 µg/ml) in a coating solution (14.3 mM Na2CO3, 10.3 mM NaHCO3, 0.02% NaN3, pH 9,6), incubated at 4°C overnight and then blocked with 10% FBS in PBS for 60 min at 37°C. The collected sera were added to the plates and incubated for 2 h at 37°C. Following this incubation, biotin-conjugated anti-mouse IgG1 and IgG2a (A85-1 and R19-15; Sigma-Aldrich, St Louis, MO) were added for detection of specific antibodies. After washing, plates were incubated at room temperature for 30 min with the StreptAB Kit (Dako, Carpinteria, CA, USA). In order to detect the bound antibodies, the OPD substrate (Sigma-Aldrich, St Louis, MO) was added. The reaction was stopped by the addition of 50 µl of a 16% solution of sulfuric acid. The optical density was measured at 490 nm.
Detection of anti-DNA antibodiesThe anti-dsDNA ELISA was performed according to
the method described by Eaton et al. (27). For Indirect Immunofluorescence, a commercially available Crithidia luciliae anti-dsDNA assay was used according to the manufacturer’s instructions (INOVA Diagnostics, San Diego, CA, USA). Briefly, 50 µl of diluted sera (1:50
D.S. LIMA ET AL.

78 79Eur. J. Inflamm.
in PBS) was added to the Crithidia luciliae slides. After incubation for 30 minutes, the slides were washed and 50 µl of FITC-conjugated anti-IgG (INOVA Diagnostics, San Diego, CA, USA) were added. After a 30-minute incubation, the slides were washed again and examined with a fluorescent microscope.
Histological analysisSixteen different organs were collected: lungs, heart,
brain, eye, skin, intestine, muscle, thyroid, kidney, suprarenal gland, liver, pancreas, thymus, spleen, ankle joint and lymph nodes. Tissues were fixed in 10% formalin, embedded in paraffin blocks, prepared routinely and then sectioned for light microscopy. Tissues were stained with hematoxylin and eosin and analysed by two different pathologists at University of São Paulo at Ribeirão Preto, School of Medicine and Heart Institute of the University of São Paulo, São Paulo, Brazil.
StatisticsAll values are expressed as mean ± SEM. Data were
analysed using analysis of variance (ANOVA). When significant differences were observed, the Tukey test was also used. p values less than 0.05 were considered significant.
RESULTS
Anti-Hsp65 and anti-Hsp60 antibodies elicited by DNAhsp65 vaccination
IgG1 and IgG2a anti-Hsp65 antibodies were detected in mice immunized with the DNAhsp65 vaccine. While both isotypes of anti-Hsp65 antibodies were detected in all vaccinated mice, antibodies were detected in neither the vector group nor the PBS control group. High levels of anti-HSP65 IgG2a antibodies were maintained up to 210 days after the first dose of the vaccine. In contrast, anti-Hsp65 IgG1 antibodies showed a slight decrease at this time point (Fig. 1A).
We also tested whether the DNAhsp65 vaccination also elicited antibodies that can recognize the human Hsp60. Indeed, antibodies to Hsp60 were also elicited following vaccination, though at a much lower level. Additionally, these antibodies were almost undetectable 210 days after vaccination (Fig. 1B).
DNAhsp65 vaccination does not elicit anti-DNA antibodies
In order to determine if anti-DNA antibodies were generated after plasmid vaccination or if the
autoimmune diseases were induced by vaccination, the serum from mice immunized with DNAhsp65 was evaluated by ELISA and immunofluorescence. Serum samples from NZB/NZW F1 animals were used as a positive control. As shown in Fig. 2 (A and B), no anti-DNA antibodies were detected 210 days after plasmid immunization.
Histological analysis In order to identify any histological sign of
tissue damage compatible with an autoimmune inflammatory process following vaccination, we examined 16 different organs. This evaluation was made at 45, 75 and 210 days following the first dose of the DNAhsp65 vaccine. Nine animals per group were analysed at each time point. The histopathological analysis did not show any evidence of inflammation that would be suggestive of an autoimmune disease. Tissue samples from control groups and vaccinated animals demonstrated essentially the same features, with a conservation of tissue characteristics at all experimental time points (data available on line at http://www.fmrp.usp.br/iba/lvg/). As shown in Table I, sporadic alterations, including hemorrhage, lymphocyte infiltration and congestion, were observed in some areas of the lung tissue in all experimental groups. However, given the limited extent of these lesions, they are likely to be of no clinical relevance. Importantly, these analyses were performed in two different laboratories in a single-blinded fashion.
DISCUSSION
The use of HSPs has emerged as a promising approach for the development of new immunotherapeutic resources due to their role in inducing both pro-inflammatory and regulatory immune responses in different settings (28).
The main bottleneck in this development is the lack of consistent knowledge that allows for the manipulation of the immunomodulatory effects of HSPs for specific applications (29). Moreover, as HSPs have been implicated also in the pathogenesis of autoimmune diseases (11), it is of special importance to determine the potential of these molecules to induce pathogenic autoimmunity.
Due to its prophylactic and therapeutic efficacy against tuberculosis in mice and guinea pigs (12-17),

80 81Eur. J. Inflamm.D.S. LIMA ET AL.
Table I. Frequency of mice displaying tissue lesions*.
The numbers correspond to the number of animals displaying each type of lesion. Congestion, infiltration and hemorrhagic foci were observed in lungs. PBS: control group; Vector: pVAX1 alone; Vaccine: DNAhsp65 vaccine. Day: days after the beginning of immunization.*No structural or statistically significant differences were observed in the lesions among the groups.
Fig. 1. Anti-HSP antibodies: BALB/c mice were immunized intramuscularly with 100 µg of DNAhsp65 or vector (DNAv) in three doses, with a two-week interval between each dose. The control group received PBS. The anti-HSPs antibody serum levels were detected in the pre-immune condition and at 45, 75 and 210 days following the start of vaccination, by ELISA. A) IgG1 and IgG2a anti-Hsp65 antibodies.B) Total IgG anti-Hsp60 antibodies. The results are presented by the average ± standard error of the mean of three independent experiments, totalling nine animals in each group. The dotted line represents the cut-off for the assay. *p<0.05 related to vector and control groups
Day 45 Day 75 Day 210 PBS Vector Vaccine PBS Vector Vaccine PBS Vector VaccineMinorcongestion
1/9 3/8 3/9 2/9 4/9 4/9 3/9 1/9 2/9
Minorinfiltration
2/9 4/8 5/9 4/9 3/9 5/9 5/9 2/9 4/9
Hemorrhagic foci
1/9 1/8 3/9 2/9 2/9 1/9 3/9 2/9 3/9
Others 0/9 1/8 2/9 2/9 1/9 2/9 1/9 2/9 1/9 Total 3/9 5/8 5/9 7/9 7/9 6/9 7/9 2/9 5/9
Anti-hsp65 Antibodies
Anti-hsp60 Antibodies

80 81Eur. J. Inflamm.
the DNAhsp65 vaccine is presently considered one of the most promising anti-tuberculosis candidates for clinical use (30-31). Moreover, its potential applications have been extended to other infectious models, arthritis, diabetes and cancer (20-23, 32). In the present study, our objective is to determine whether DNAhsp65 vaccination elicits any tissue inflammation suggestive of an autoimmune disease and/or potentially pathogenic immune responses.
For this purpose, we looked for tissue damage potentially induced by the vaccine in 16 different organs and we analysed the humoral immune response against both mycobacterial Hsp65 and human Hsp60.
The DNAhsp65 vaccine clearly stimulated the production of high levels of anti-Hsp65 antibodies. This is indirect evidence for the effective transfection of cells after DNAhsp65 vaccination. We also detected anti-human Hsp60 antibodies following DNAhsp65 immunization, though at a lower level. Our data indicate that this vaccine was able to induce an anti-Hsp65 antibody response that possibly displayed some cross-reactivity with the human Hsp60 and, most likely, with the endogenous murine Hsp60. Although we did not test antibody reactivity specifically to murine Hsp60, as the human and mouse Hsp60 sequences share more than 95% sequence identity, it is very likely that
Fig. 2. Anti-DNA antibodies. A) The serum anti-DNA antibody levels were determined by ELISA (diluted 50-fold). Serum from NZB/NZW mice was used as positive control. The results are presented as the average ± standard error of the mean of three independent experiments totalling nine animals in each group. *p<0.001. B) Serum anti-DNA antibodies were measured by indirect immunofluorescence for reactivity with Crithidia luciliae DNA. (I) Positive control for anti-DNA antibodies (NZB/NZW mice); (II) negative control for anti-DNA antibodies (pre-immune BALB/c serum) and (III) a negative result for serum anti-DNA antibodies in a DNAhsp65 vaccinated mouse.

82 83Eur. J. Inflamm.
disease. No evidence of significant tissue damage was observed in vaccinated mice at any of the three different time points.
Our results are in contrast to those of other investigators, who have reported that subunit vaccines based on the recombinant M. tuberculosis Hsp65 and Hsp70, as well as DNA vaccines encoding the Ag85 and M. leprae Hsp65 genes, may have harmful effects in mice and guinea pigs (24-25). These investigators observed no protection
some anti-Hsp65 antibodies elicited by the vaccine were cross-reactive to the endogenous Hsp60, as previously reported (33). However, despite the generation of potentially cross-reactive antibodies in vaccinated animals, no sign of pathogenic autoimmunity was detected, even 210 days after the first dose of immunizations. Sixteen different organs were evaluated histopathologically in a blinded manner in two different laboratories for any sign of tissue damage suggestive of an autoimmune
D.S. LIMA ET AL.
Fig. 3. Histological evaluation of tissues. Hematoxylin and eosin staining of tissue sections of six different organs at 210 days after the initial DNAhsp65 immunization. No significant inflammatory infiltrate or other signs of tissue damage were detected. The different experimental groups are indicated in each photo.

82 83Eur. J. Inflamm.
with these vaccines and reported that a subsequent challenge with M. tuberculosis lead to necrotizing bronchointerstitial pneumonia and bronchiolitis. Moreover, when Hsp65 and Ag85 were used in an immunotherapeutic model, classical Koch reactions with necrosis in the interior of granulomas were observed (25-26). Since both antigens (Hsp65 and Ag85) have been frequently used by different laboratories around the world with repeated success in different models (12-21), it is possible that the type of lesions observed by Turner (24) and Taylor (25-26) et al. could be an isolated phenomenon. This could possibly be due to the contamination of the vaccines with endotoxin (30), a problem that we excluded through the production of plasmids with endotoxin levels that are in accordance with US and European guidelines (34).
As anti-DNA antibodies are important diagnostic and nephritogenic factors in systemic lupus erythematosus (SLE), we determined whether our vaccine induced the production of anti-DNA antibodies. These could be triggered by the breakdown of immunoregulatory mechanisms induced by vaccination or even stimulated by the vaccine DNA itself. The methylation state and CpG sequences on bacterial DNA could activate B and T cells leading to the production of anti-DNA antibodies (35-36). However, using two different techniques, we detected no anti-DNA antibody production in vaccinated mice in our present study. In contrast, high levels of anti-DNA antibodies were detected in the serum samples of F1 NZB/NZW mice, a model of SLE that was used as a positive control in the assays.
Our present data are in accordance with previous results from our group, in which DNAhsp65 vaccination in experimental models of autoimmune diseases, such as arthritis and diabetes, resulted in a significant reduction in the tissue injury of vaccinated animals. This suggests that this vaccine may also have immunoregulatory properties (20, 32). In fact, some HSPs have been shown to be recognized specifically by regulatory T cells (33). This is a major point to be elucidated for M. leprae Hsp65.
In this study, we show that the DNAhsp65 vaccine did not induce autoimmune diseases or significant histopathological lesions in healthy BALB/c mice. Nevertheless, further characterization of the effects
of DNAhsp65 vaccination is necessary due to the broad distribution of the vaccine in different organs following intramuscular inoculation, as we previously reported (37). Moreover, considering the therapeutic properties of this vaccine against chronic tuberculosis in murine models, and its potential use in the clinic, it is also necessary to investigate whether this vaccine induces autoimmune diseases in models of tuberculosis treatment in normal and immune-compromised individuals.
ACKNOWLEDGEMENTS
We are grateful to Ana Maria Rocha, Giovanna Santos, Juliana Hori, Tatiana Schneider and Thiago Malardo, for assistance in histological procedures.
REFERENCES
1. Coelho V, Broere F, Binder RJ, Shoenfeld Y, Moudgil KD. Heat-shock proteins: Inflammatory versus regulatory attributes. Cell Stress Chaperones 2008; 13:119-25.
2. Zhang X, Yu C, Zhao J, et al. Vaccination with a DNA vaccine based on human PSCA and HSP70 adjuvant enhances the antigen-specific CD8+ T-cell response and inhibits the PSCA+ tumors growth in mice. J Gene Med 2007; 9:715-26.
3. Quintana FJ, Carmi P, Cohen IR. DNA vaccination with heat shock protein 60 inhibits cyclophosphamide-accelerated diabetes. J Immunol 2002; 169:6030-5.
4. Lowrie DB, Silva CL, Colston MJ, Ragno S, Tascon RE. Protection against tuberculosis by a plasmid DNA vaccine. Vaccine 1997; 15:834-38.
5. van Eden W, van der Zee R, Prakken B. Heat-shock proteins induce T-cell regulation of chronic inflammation. Nat Rev Immunol 2005; 5:318-30.
6. Takakura Y, Takemoto S, Nishikawa M. Hsp-based tumor vaccines: state-of-the-art and future directions. Curr Opin Mol Ther 2007; 9:385-91.
7. Li Y, Subjeck J, Yang G, Repasky E, Wang XY. Generation of anti-tumor immunity using mammalian heat shock protein 70 DNA vaccines for cancer immunotherapy. Vaccine 2006; 24:5360-70.
8. van Eden W, Wick G, Albani S, Cohen I. Stress, heat shock proteins, and autoimmunity: how immune

84 85Eur. J. Inflamm.
responses to heat shock proteins are to be used for the control of chronic inflammatory diseases. Ann NY Acad Sci 2007; 1113:217-37.
9. Caldas C, Luna E, Spadafora-Ferreira M, et al. Cellular autoreactivity against heat shock protein 60 in renal transplant patients: peripheral and graft-infiltrating responses. Clin Exp Immunol 2006; 46:66-75.
10. Riganò R, Profumo E, Buttari B, et al. Heat shock proteins and autoimmunity in patients with carotid atherosclerosis. Ann NY Acad Sci 2007; 1107:1-10.
11. van Eden W, Koets A, van Kooten P, Prakken B, van der Zee R. Immunopotentiating heat shock proteins: negotiators between innate danger and control of autoimmunity. Vaccine 2003; 21:897-901.
12. Paula L de, Silva CL, Carlos D, et al. Comparison of different delivery systems of DNA vaccination for the induction of protection against tuberculosis in mice and guinea pigs. Genet Vaccines Ther 2007; 5:2.
13. Nuermberger E, Tyagi S, Williams KN, Rosenthal I, Bishai WR, Grosset H. Rifapentine, moxifloxacin, or DNA vaccine improves treatment of latent tuberculosis in a mouse model. Am J Respir Crit Care Med. 2005; 11:1452-6.
14. Shi C, Yuan S, Zhang H, Zhang T, Wang L, Xu Z. Cell-mediated immune responses and protective efficacy against infection with Mycobacterium tuberculosis induced by Hsp65 and hIL-2 fusion protein in mice. Scand J Immunol 2009; 69:140-9.
15. Lowrie DB, Tascon RE, Bonato VL, et al. Therapy of tuberculosis in mice by DNA vaccination. Nature 1999; 400:269-71.
16. Silva CL, Bonato VL, Coelho-Castelo AA, et al. Immunotherapy with plasmid DNA encoding mycobacterial hsp65 in association with chemotherapy is a more rapid and efficient form of treatment for tuberculosis in mice. Gene Ther 2005; 12:281-7.
17. Zárate-Bladés CR, Bonato VL, da Silveira EL, et al. Comprehensive gene expression profiling in lungs of mice infected with Mycobacterium tuberculosis following DNAhsp65 immunotherapy. J Gene Med 2009; 11:66-78.
18. Lukacs KV, Lowrie DB, Stokes RW, Colston MJ. Tumor cells transfected with a bacterial heat-shock gene lose tumorigenicity and induce protection
against tumors. J Exp Med 1993; 178:343-8.19. Bonato VL, Goncalves ED, Soares EG, et al.
Immune regulatory effect of pHSP65 DNA therapy in pulmonary tuberculosis: activation of CD8+ cells, interferon-gamma recovery and reduction of lung injury. Immunology 2004; 113:130-8.
20. Santos-Junior RR, Sartori A, De Franco M, et al. Immunomodulation and protection induced by DNA-hsp65 vaccination in an animal model of arthritis. Hum Gene Ther 2005; 16:1338-45.
21. Ribeiro AM, Bocca AL, Amaral AC, et al. DNAhsp65 vaccination induces protection in mice against Paracoccidioides brasiliensis infection. Vaccine 2009; 27:606-13.
22. Michaluart P, Abdallah KA, Lima FD, et al. Phase I trial of DNA-hsp65 immunotherapy for advanced squamous cell carcinoma of the head and neck. Cancer Gene Ther 2008; 10:676-84.
23. Victora GD, Socorro-Silva A, Volsi EC, et al. Immuno-response to vaccination to DNA-hsp65 in a phase I clinical trial with head and neck cancer patients. Cancer Gene Ther 2009; in press.
24. Turner OC, Roberts AD, Frank AA, et al. Lack of protection in mice and necrotizing bronchointerstitial pneumonia with bronchiolitis in guinea pigs immunized with vaccines directed against the hsp60 molecule of Mycobacterium tuberculosis. Infect Immun 2000; 68:3674-9.
25. Taylor JL, Turner OC, Basaraba RJ, Belisle JT, Huygen K, Orme IM. Pulmonary necrosis resulting from DNA vaccination against tuberculosis. Infect Immun 2003; 71:2192-8.
26. Taylor JL, Ordway DJ, Troudt J, Gonzalez-Juarrero M, Basaraba RJ, Orme IM. Factors associated with severe granulomatous pneumonia in Mycobacterium tuberculosis-infected mice vaccinated therapeutically with hsp65 DNA. Infect Immun 2005; 73:5189-93.
27. Eaton RB, Schneider G, Schur PH. Enzyme immunoassay for antibodies to native DNA. Specificity and quality of antibodies. Arthritis Rheum 1983; 26:52-62.
28. Rajaiah R, Moudgil KD. Heat-shock proteins can promote as well as regulate autoimmunity. Autoimmun Rev 2009; 8:388-403.
29. Bolhassani A, Rafati S. Heat-shock proteins as powerful weapons in vaccine development. Expert
D.S. LIMA ET AL.

84 85Eur. J. Inflamm.
Rev Vaccines 2008; 8:1185-99.30. Lowrie DB. DNA vaccines for therapy of
tuberculosis: where are we now? Vaccine 2006; 24:1983-9.
31. Rook GA, Lowrie DB, Hernandez-Pando R. Immunotherapeutics for tuberculosis in experimental animals: is there a common pathway activated by effective protocols? J Infect Dis 2007; 196:191-8.
32. Santos-Junior RR, Sartori A, Bonato VLD, et al. Immune modulation induced by tuberculosis DNA vaccine protects NOD mice from diabetes progression. Clinical Experimental Immunology 2007; 149:570-8.
33. Quintana FJ, Cohen IR. DNA vaccines coding for heat-shock proteins (HSPs): tools for the activation of HSP-specific regulatory T cells. Expert Opin Biol
Ther 2005; 5:545-54.34. Tuomela M, Stanescu I, Krohn K. Validation
overview of bio-analytical methods. Gene Ther 2005; 12(S):1318.
35. Cornelie S, Poulain-Godefroy O, Lund C, et al. Methylated CpG-containing plasmid activates the immune system. Scand J Immunol 2004; 59:143-51.
36. Krieg AM, Yi AK, Matson S, et al. CpG motifs in bacterial DNA trigger direct B-cell activation. Nature 1995; 374:546-9.
37. Coelho-Castelo AA, Trombone AP, Rosada RS, Santos RR Jr, Bonato VL, Sartori A, Silva CL. Tissue distribution of a plasmid DNA encoding Hsp65 gene is dependent on the dose administered through intramuscular delivery. Genet Vaccines Ther 2006; 4:1.

EUROPEAN JOURNAL OF INFLAMMATION Vol. 7, no. 2, 0-0 (2009)
1721-727X (2009)Copyright © by BIOLIFE, s.a.s.
This publication and/or article is for individual use only and may not be furtherreproduced without written permission from the copyright holder.
Unauthorized reproduction may result in financial and other penalties87

EUROPEAN JOURNAL OF INFLAMMATION Vol. 7, no. 2, 0-0 (2009)
1721-727X (2009)Copyright © by BIOLIFE, s.a.s.
This publication and/or article is for individual use only and may not be furtherreproduced without written permission from the copyright holder.
Unauthorized reproduction may result in financial and other penalties87
THE EFFECTS OF DARBEPOETIN ON PERITONEAL FIBROSIS INDUCED BY CHEMICAL PERITONITIS AND ON PERITONEAL TISSUE MMP-2 AND TIMP-2
LEVELS IN RATS
A. BORAZAN, T. CAMSARİ1, Z. CAVDAR2, S. SARİOGLU3, O. YİLMAZ4, G. OKTAY2, A. SİFİL1, A. CELİK1, C. CAVDAR1, A. AYSAL3 and E. KOLATAN4
Department of Nephrology, Faculty of Medicine, Mustafa Kemal University, Hatay; 1Department of Nephrology, 2Department of Biochemistry, 3Department of Pathology, 4Department of Animal
Research Center, Faculty of Medicine, Dokuz Eylul University, Izmir, Turkey
Received February 16, 2009 – Accepted May 5, 2009
The aim of this study is to assess the influence of darbepoetin on the development of peritoneal fibrosis in rats induced by chlorhexidine gluconate (0.1%) and ethanol (15%) and to determine the effect on peritoneal tissue levels of MMP–2 and TIMP–2, possible important factors in progression of peritoneal fibrosis. Twenty-four female Wistar albino rats were divided into three groups. The first group (CH group) received 3 ml/200g daily intraperitoneal injections of chlorhexidine gluconate (0.1%) and ethanol (15%) dissolved in saline to induce chemical peritonitis; group 2 (ESA group) received 3 ml/200g daily injections of chlorhexidine gluconate (0.1%) and ethanol (15%) dissolved in saline and also darbepoetin 12.5 microgr/ per kilogram/ day subcutaneously on the first and seventh days; group 3 (Control group) received intraperitoneal 0.9% saline (3 ml/200g/d) through the right lower quadrant by 21 gauge needle. The study duration was fourteen days. On the fifteenth day rats were sacrificed, parietal peritoneum samples were obtained from the left anterior abdominal wall. Pathological samples were examined using Hematoxyline & Eosin (HE) stains. The thickness, vasculpathy, and inflammation were determined by light microscopy. MMP–2 and TIMP–2 were studied immunohistochemically by monoclonal antibody staining. The activity of MMP–2 on peritoneal tissue was studied by gelatin zymography and TIMP–2 protein level was analysed by ELISA, biochemically. The decrease in thickness of parietal peritoneum in group ESA was statistically significant when compared to CH group (p<0.05). Inflammation scores, and vascularization score surfaces were not statistically different between these groups (p>0.05). Immunohistochemically, darbepoetin was shown to decrease MMP–2 expression on parietal peritoneum in CH group (p<0.05), but had no effect on TIMP–2 (p>0.05). Biochemically the ratio of active MMP–2 to proMMP–2 was more significantly increased in the ESA group than in the CH group (p<0.001), however, TIMP- 2 levels in both groups were decreased compared to the control group (p<0.05). Darbepoetin histopathologically reduced peritoneal fibrosis induced by chlorhexidine gluconate. We can suggest that Darbepoetin does not cause peritoneal fibrosis and may prevent peritoneal fibrosis in rats possibly related to an effect on MMP-2 expression. Further research regarding the utility and dosage should be considered.
Mailing address: Dr Ali Borazan,Mustafa Kemal Universitesi, Tıp Fakültesi,Nefroloji BD 31100 Hatay, TurkeyTel: ++90 0326 2140649 Fax: ++90 0326 2148214e-mail: [email protected]
Key words: peritoneal fibrosis, MMP-2, TIMP-2

88 89Eur. J. Inflamm.
Peritoneal dialysis (PD) is an efficient and reliable model of renal replacement therapy for patients with end-stage renal failure. The changes developing in peritoneal membrane over time eventually leads to termination of peritoneal dialysis. Peritoneal fibrosis is an important cause of reduced peritoneal dialysis efficiency (1-2).
There are many important factors playing a role in the development of peritoneal fibrosis (PF) in peritoneal dialysis (PD) patients. The most important factors leading to PF are bioincompatible dialysates, high glucose concentrations in dialysates, glucose degradation products resulting from overheated dialysates, and advanced glycation end products (3-4). Uremia alone has been shown to be a causative factor of PF in studies comparing peritoneal thickness between end stage renal patients not on dialysis and healthy control groups (5).
These factors, together with the influence of other peritoneal cells, i.e peritoneal fibroblasts, macrophages, monocytes and neutrophils, and of secreted cytokines from these cells lead peritoneal mesothelial cells to develop peritoneal fibrosis. The principal pathogenetic mechanism in the development of PF is increased production of peritoneal mesothelial cells and/or peritoneal fibroblasts, and increased extracellular matrix accumulation (1-2). These pathological changes may lead to sclerosing encapsulated peritonitis (SEP) which is a serious complication of PD. The membrane changes occurring in long-term PD are severe submesothelial thickening, decrease in mesothelial cells, and neovascularization. The change in peritoneal membrane solute transport capacity is postulated to be due to increased vascular surface area. Revascularization and vasculopathy have a pivotal effect on peritoneal permeability for fluid and solutes (6).
Recent studies have demonstrated higher matrix metalloproteinase (MMP) levels especially that of MMP-2 in peritoneal tissue and drained dialysates of experimentally induced PF models and SEP cases in humans (2, 7). MMP-2 is produced mainly by fibroblasts, epithelial cells and macrophages, whereas MMP-9 is produced by inflammatory cells, mainly neutrophils (2, 5). The release of MMPs from mesothelial cells is stimulated by interleukin-1 (IL-1), transforming growth factor–β (TGF-β)
and hepatocyte growth factor (HGF). Stimulated epithelial cells also contribute to angiogenesis induced by release of MMP-2 and MMP-9. MMP suppressor agents were used to prevent inflammation, fibrosis and tumor invasion (8). The activation of MMPs is blocked by tissue enzyme specific and native tissue inhibitors of metalloproteinases (TIMPs) and α-macroglobulin. Four different types of TIMPs were defined; TIMP-1, TIMP-2, TIMP-3 and TIMP-4. The equilibrium between TIMPs and MMPs plays a key role also in equilibrium between extracellular matrix accumulation and degradation (8).
Darbepoetin, as an erythropoiesis stimulating agent (ESA), has been used in the treatment of anemia of chronic renal failure. Recent studies have also shown its cell protective functions (9). In traumatic head injuries it was shown to reduce cerebral infarction area by 75%, in coronary ischemia it was also shown to improve left ventricular function and volume, as well as loss of cardiomyocytes (10-11). Furthermore, it has been demonstrated to prevent kidneys from inflammatory and fibrotic insult in ongoing cyclosporine toxicity (12).
It has been demonstrated that ESAs decrease expression of TGF-ß mRNA and cell death by apoptosis, increase proliferation of renal tubular cells, and have no effect on hemoglobin and hematocrit levels at low doses (9, 13-14). Darbepoetin was shown to prevent tubulointerstitial injury, glomerulosclerosis and vascular sclerosis in 5/6 nephrectomized test animals (15). Currently there is no study proving the effectiveness of Darbepoetin in peritoneal fibrosis. In this study we investigated the effect of darbepoetin on peritoneal fibrosis induced by chemical peritonitis made by 15% ethyl alcohol and 0.1% chlorhexidine dissolved in saline in rats, and on peritoneal tissue MMP-2 and TIMP-2 expressions detected immunohistochemically and biochemically.
MATERIALS AND METHODS
This study was undertaken after approval by the Ethical Committee of Experimental Animal Investigations at Dokuz Eylul University, Faculty of Medicine. The study group was composed of a total of 24 female Wistar albino rats, weighing between 200-230 gr. The rats were housed in a multidisciplinary laboratory for fourteen days
A. BORAZAN ET AL.

88 89Eur. J. Inflamm.
in standard hutches, four rats to each hutch, with standard food and water provided. Exclusion criteria included development of infections and systemic reactions other than peritoneal sclerosis, or any other reason for morbidity. The rats included in the study were weighed before the study and divided into three groups (n: eight rats per group). The duration of the study was fourteen days. To induce peritoneal fibrosis we used a model described by Ishiii and et al.(16) that was composed of 0.1% CH and 15% ethyl alcohol dissolved in saline prepared in aseptic area. Darbepoetin ( Aranesp, Amgen Europe B.V Breda, Holland) was used as 10 µg /0.4 ml (25 µg/ml).
The first group of rats (Chlorhexidine gluconate=CH group) were given daily intraperitoneal injections of 15% ethyl alcohol / 0.1 % CH in saline ( 3 ml/200g) through a 21 gauge needle for fourteen days. The second group of rats (Erythropoiesis stimulating agent=ESA group) were given the same treatment as the first group plus darbepoetin 12.5 µg/kg, as a subcutaneous injection on day 0 and the 7th day using a 21 gauge needle. The third group of rats (Control group) were given intraperitoneal injection of 0.9% saline (3 ml/200g) for fourteen days, using a 21 gauge needle. All intraperitoneal applications were performed in the right lower quadrant of the abdomen.
On the fifteenth day the rats were sacrificed by using a toxic dose of ether. After they were weighed and a sample of the parietal peritoneum from the left anterior abdominal wall was taken. Samples were divided into two; and the samples for pathological examination were fixed with 10% formaldehyde. The samples for biochemical examination were kept in tubes at -80°C.
Histopathologic analysisVertical sections of 3 mm thickness were prepared
from formaldehyde fixed samples, embedded in paraffin and stained with hematoxylin-eosin (H&E). The thickness of parietal peritoneum, vasculopathy, and inflammation were examined by light microscopy.
Light microscopic examinationTo measure peritoneal thickness at sections stained with
H&E, microscopic images were transferred to a computer through a camera (Olympus BX50, Olympus Optical Co, Tokyo, Japan). Median of thicknesses of parietal peritoneum at ten sites was measured using image analysis software. Inflammation was scored semi-quantitatively in H&E stained sections accordingly: 0: no inflammation, 1: mild, 2: moderate, 3: severe. Vasculopathy was scored as follows, modified from Williams and coll. (5); 0: normal, 1: subendothelial hyalinization, 2: luminal irregularity and stenosis, 3: luminal obstruction (17-19).
Immunohistochemical analysisSections prepared on slides with poly-L-lysine were
stained with MMP-2 Neomarkers (72 kDa collagenase IV) Ab-1 (Clone CA-4001) (1: 100.200 mcg/ml), and TIMP-2 Ab-5 Neomarkers (prediluted, Clone 3A4) (Cat MS-1485-R7 7 ml) immunohistochemical stains. Placental tissue for MMP-2 and colonic tumoral tissue for TIMP-2 were used as positive controls. The expression was evaluated by light microscopy, by giving scores for the density and intensity from 0 (none) to 4 (extensive-strong). The multiplication of these gave a score from 1 to 16. Values equal and less than four represented weak staining and higher values represented intense staining (20).
Preparation of tissue samples for biochemical analysisFrozen parietal peritoneum samples (100 mg) were
pulverized by liquid nitrogen. Then they were transferred to 1.5 ml microcentrifuge tubes, on ice, containing 1.0 ml a extraction medium (50 mM tris-HCl pH7.4, 1% NP-40. 0.25% deoxycholate, 150 mM NaCl, 1 mM EDTA) Then, the samples were homogenized by 10 seconds of sonification on ice by sonicator (Sonics&Materials Inc, Danbury Connecticut, USA). After 15 min centrifugation at 10,000 x g supernatants were collected and stored at –80°C until analysis (21).
Protein analysisProtein levels were determined by using Bicinconinic
acid protein kit (BCA) (Sigma, Germany). Bovine serum albumin was used as a standard (22).
Gelatin zymographyGelatin zymography was used to determine pro- and
active forms of MMP-2 (23). Tissue homogenates were diluted 1:1 with non-reducing sample buffer (0.5 M Tris/HCl pH 6.8, SDS, glycerol and bromophenol blue), and samples were separated within 7.5% SDS polyacrylamide gel composed of 0.1 % gelatin as a substrate at 125 voltage for 4 h at 4°C. MMP-2 marker (Chemicon) that includes both pro- and active forms was used as a positive control. After electrophoresis, gels were washed with 2.5% Triton X-100 twice for 15 minutes at room temperature to remove SDS and incubated for 18 h at 37°C in activation buffer ( 0.05 M Tris-HCl, 0.05 M NaCl, 10 mM CaCl2,
0.05% Brij 35, pH 7.6) to digest the gelatin substrate by denatured MMPs. After incubation, the gels were stained with 0.5% Coomassie Blue R-250 for 30 mins for staining and destained twice in 40% methanol and 10% acetic acid in distilled water. Pro- and active forms of MMP-2 were seen as lytic bands on a blue background. Gels were semi-quantified by using computerized densitometry (UVP Bio Imaging System with a Labworks 4.6 Image Acquisition Software, Cambrige, UK). Area and optic density of each band was calculated. Gelatin substrate digestion levels
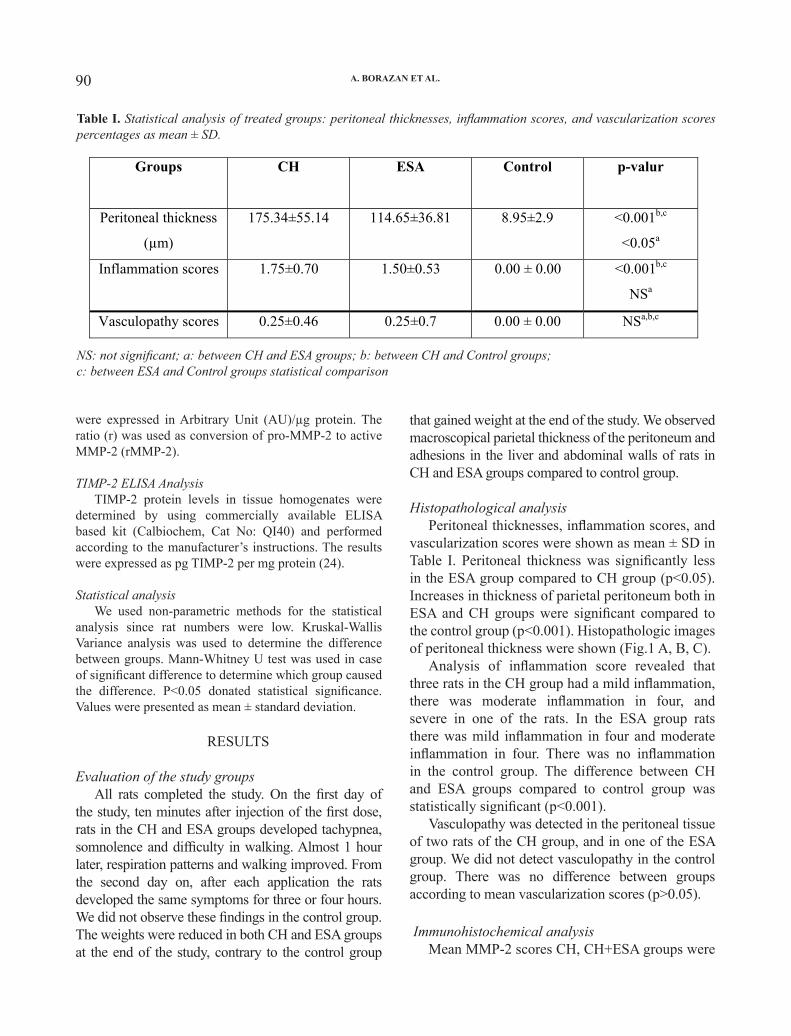
90 91Eur. J. Inflamm.
were expressed in Arbitrary Unit (AU)/µg protein. The ratio (r) was used as conversion of pro-MMP-2 to active MMP-2 (rMMP-2).
TIMP-2 ELISA AnalysisTIMP-2 protein levels in tissue homogenates were
determined by using commercially available ELISA based kit (Calbiochem, Cat No: QI40) and performed according to the manufacturer’s instructions. The results were expressed as pg TIMP-2 per mg protein (24).
Statistical analysisWe used non-parametric methods for the statistical
analysis since rat numbers were low. Kruskal-Wallis Variance analysis was used to determine the difference between groups. Mann-Whitney U test was used in case of significant difference to determine which group caused the difference. P<0.05 donated statistical significance. Values were presented as mean ± standard deviation.
RESULTS
Evaluation of the study groupsAll rats completed the study. On the first day of
the study, ten minutes after injection of the first dose, rats in the CH and ESA groups developed tachypnea, somnolence and difficulty in walking. Almost 1 hour later, respiration patterns and walking improved. From the second day on, after each application the rats developed the same symptoms for three or four hours. We did not observe these findings in the control group. The weights were reduced in both CH and ESA groups at the end of the study, contrary to the control group
that gained weight at the end of the study. We observed macroscopical parietal thickness of the peritoneum and adhesions in the liver and abdominal walls of rats in CH and ESA groups compared to control group.
Histopathological analysisPeritoneal thicknesses, inflammation scores, and
vascularization scores were shown as mean ± SD in Table I. Peritoneal thickness was significantly less in the ESA group compared to CH group (p<0.05). Increases in thickness of parietal peritoneum both in ESA and CH groups were significant compared to the control group (p<0.001). Histopathologic images of peritoneal thickness were shown (Fig.1 A, B, C).
Analysis of inflammation score revealed that three rats in the CH group had a mild inflammation, there was moderate inflammation in four, and severe in one of the rats. In the ESA group rats there was mild inflammation in four and moderate inflammation in four. There was no inflammation in the control group. The difference between CH and ESA groups compared to control group was statistically significant (p<0.001).
Vasculopathy was detected in the peritoneal tissue of two rats of the CH group, and in one of the ESA group. We did not detect vasculopathy in the control group. There was no difference between groups according to mean vascularization scores (p>0.05).
Immunohistochemical analysis Mean MMP-2 scores CH, CH+ESA groups were
1
Table I. Statistical analysis of treated groups: peritoneal thicknesses, inflammation scores, and vascularization scores percentages as mean ± SD.
Groups CH ESA Control p-valur
Peritoneal thickness
(µm)
175.34±55.14 114.65±36.81 8.95±2.9 <0.001b,c
<0.05a
Inflammation scores 1.75±0.70 1.50±0.53 0.00 ± 0.00 <0.001b,c
NSa
Vasculopathy scores 0.25±0.46 0.25±0.7 0.00 ± 0.00 NSa,b,c
NS: not significant; a: between CH and ESA groups; b: between CH and Control groups;c: between ESA and Control groups statistical comparison
Table I. Statistical analysis of treated groups: peritoneal thicknesses, inflammation scores, and vascularization scores percentages as mean ± SD.
NS: not significant; a: between CH and ESA groups; b: between CH and Control groups; c: between ESA and Control groups statistical comparison
A. BORAZAN ET AL.
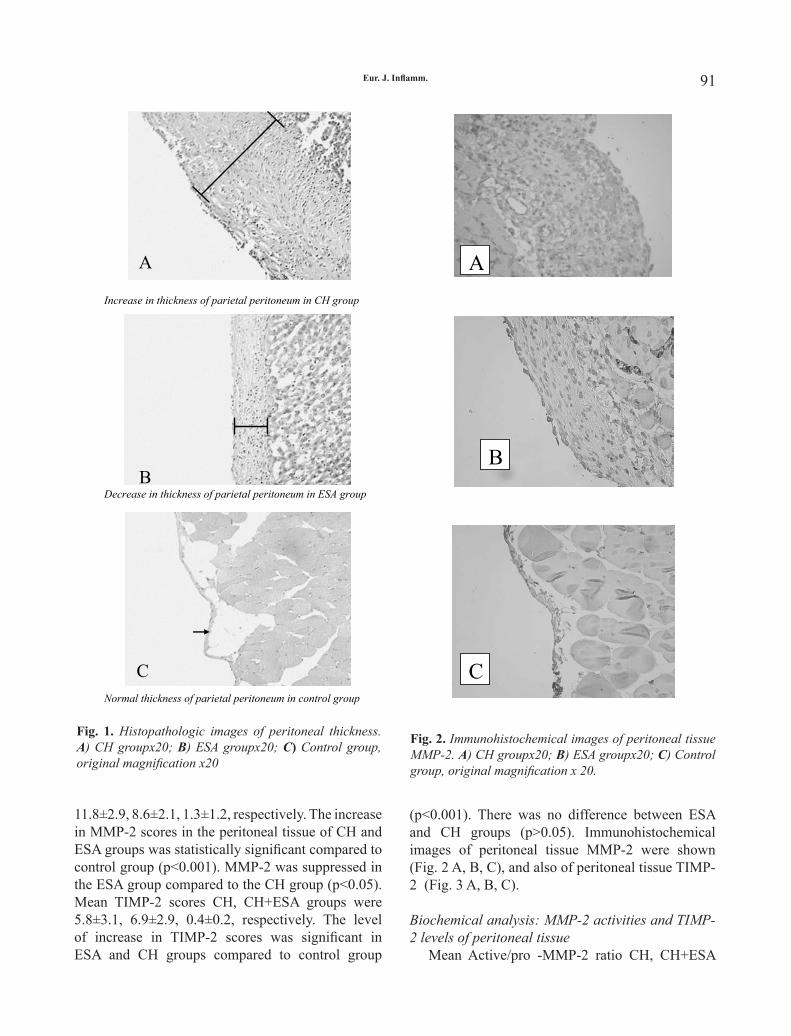
90 91Eur. J. Inflamm.
11.8±2.9, 8.6±2.1, 1.3±1.2, respectively. The increase in MMP-2 scores in the peritoneal tissue of CH and ESA groups was statistically significant compared to control group (p<0.001). MMP-2 was suppressed in the ESA group compared to the CH group (p<0.05). Mean TIMP-2 scores CH, CH+ESA groups were 5.8±3.1, 6.9±2.9, 0.4±0.2, respectively. The level of increase in TIMP-2 scores was significant in ESA and CH groups compared to control group
(p<0.001). There was no difference between ESA and CH groups (p>0.05). Immunohistochemical images of peritoneal tissue MMP-2 were shown (Fig. 2 A, B, C), and also of peritoneal tissue TIMP-2 (Fig. 3 A, B, C).
Biochemical analysis: MMP-2 activities and TIMP-2 levels of peritoneal tissue
Mean Active/pro -MMP-2 ratio CH, CH+ESA
A
Increase in thickness of parietal peritoneum in CH group
B Decrease in thickness of parietal peritoneum in ESA group
C Normal thickness of parietal peritoneum in control group
Fig. 1. Histopathologic images of peritoneal thickness. A) CH group x20; B) ESA group x20;
C) Control group, original magnification x20. Fig. 1. Histopathologic images of peritoneal thickness. A) CH groupx20; B) ESA groupx20; C) Control group, original magnification x20
1
Fig. 2. Immunohistochemical images of peritoneal tissue MMP-2. A) CH group x20; B) ESA group x20; C) Control group, original magnification x20
A
B
C
Fig. 2. Immunohistochemical images of peritoneal tissue MMP-2. A) CH groupx20; B) ESA groupx20; C) Control group, original magnification x 20.

92 93Eur. J. Inflamm.
groups were 2.5± 2.4, 7.1±3.4, 0.6±0.4 respectively. Active MMP-2/pro-MMP2 ratio was significantly higher in the ESA group compared to the CH group (p<0.01). A zymogram representing all groups was presented.
Mean TIMP-2 scores CH, CH+ESA groups were 1800±1200, 1700±1300, 3400±2300 pg/mg protein, respectively. Decrease in TIMP-2 was statistically significant in ESA and CH groups compared to
control group (p<0.05). There was no difference between ESA and CH groups (p>0.05).
DISCUSSIONIn our study we demonstrate that darbepoetin
decreased parietal peritoneal thickness histopathologically compared to a chlohexidine group, but had no effect on inflammation and vascularization scores. Although darbepoetin decreased the thickness of the peritoneum, it had no effects on inflammation and vascularization scores which can be explained by the findings that occurred later on in the course. In this study the changes seen in parietal peritoneum showed no difference histopathologically or biochemically.
Darbepoetin was not a pure molecule in our study. Darbepoetin alpha is used as an adjunct to some ingredients, such as monobasic sodium phosphate, dibasic sodium phosphate, sodium chloride, polysorbate 80, and pure water. In conclusion, since darbepoetin caused lesser peritoneal fibrosis we suggest that neither darbepoetin, nor other adjunctive compounds had any effect on the development of peritoneal fibrosis. Darbepoetin is introduced as a compound and used clinically subcutaneously, so we could not use darbepoetin in its pure form, therefore the effect of adjunctive substances on peritoneal fibrosis could not be estimated.
In the present study we induced experimental peritoneal fibrosis by using 15% ethyl alcohol and 0.1% chlorhexidine gluconate dissolved in saline. Experimental peritoneal fibrosis can be induced by intraperitoneal injection of chlorhexidine gluconate (2). It is a chemical agent that is not used clinically but used to generate experimental peritoneal fibrosis. There are other methods to induce experimental peritoneal fibrosis, and studies have been ongoing to prevent peritoneal fibrosis (7, 25).
Junor and colleagues first described the role of gluconate and alcohol in pathogenesis of peritoneal fibrosis in 1985 (16). They reported the development of clinical ESP when peritoneal catheters are sterilized by chlorhexidine gluconate in alcohol. Upon this finding Ishii and colleagues showed development of peritoneal fibrosis in test animals by using 0.1% chlorhexidine gluconate and 15% alcohol dissolved in saline (16). Symptoms such as tachypnea, somnolence and difficulty in walking observed in
2
Fig. 3. Immunohistochemical images of peritoneal tissue TIMP-2. A) (CH group x20); B)ESA group x20; C) Control group, original magnification x20.
A
B
C
Fig. 3. Immunohistochemical images of peritoneal tissue TIMP-2. A) CH groupx20, B) ESA groupx20; C) Control group, original magnification x20.
A. BORAZAN ET AL.

92 93Eur. J. Inflamm.
peritoneal fibrosis (2, 28, 30).In this study MMP-2 levels in peritoneum were
shown to be suppressed in the darbepoetin group compared to the CH group, but increased compared to the saline group in immunohistochemical analysis. On the other hand, the ratio of active to pro-MMP-2 in peritoneum was high in the darbepoetin group compared to the CH group in a biochemical analysis. Immunohistochemically, MMP-2s in peritoneum represent both pro- and active forms of the molecule. Accordingly, we can conclude that darbepoetin increases total MMP-2 level, and also accelerates pro-MMP-2 conversion to active MMP-2. We thought that the effect of darbepoetin in decreasing development of peritoneal fibrosis induced by CH in rats was due to increases in active MMP-2 level. Immunohistochemically, peritoneal TIMP-2 levels were significantly high in both darbepoetin and CH groups compared to the saline group, but no difference was observed between the darbepoetin and CH groups. Biochemically, however, it was reduced significantly in both darbepoetin and CH groups compared to the saline group. These controversies may be related to equal tissue volumes of biochemically-measured TIMP-2 in all groups, but rather thin samples of peritoneum in the saline group of immunohistochemically measured TIMP-2 causing low TIMP-2 scores. Also, we did not suggest that this resulted from sampling of peritoneum biochemically. If this occurred, this would reflect that a TIMP-2 level at
test animals after induction of peritoneal fibrosis by 15% ethyl alcohol and 0.1% chlorhexidine gluconate had not been previously reported. We suggested that high dose ethyl alcohol and chlorhexidine gluconate induced acute metabolic acidosis, and neurological effects of alcohol or hypotension might have contributed to these symptoms. At the end of the study we observed peritoneal adhesions over liver and anterior abdominal wall in rats with chemical peritonitis.
Recently, it has been demonstrated that ESAs have effects other than chronic renal failure related anemia. They inhibited apoptosis in experimental acute renal failure model made by bilateral ischemic reperfusion model, increased tubular epithelial regeneration and accelerated gain of renal functions (9). Furthermore, they decreased interstitial fibrosis and tubular injury.
Matrix metalloproteinases and their natural inhibitors play a role in the ongoing process of peritoneal fibrosis (26-27). MMPs and TIMPs are produced by peritoneal mesothelial cells, fibroblasts and macrophages in peritoneum, and also in peritoneal tissue cultures MMP-2, MMP-3 and MMP-9, and TIMP-1 and TIMP-2 activity levels were shown to be increased. TIMPs not only suppress MMPs but also increase their activity at lower tissue concentrations (26, 28-29). Recent experimental animal studies have shown that use of MMP suppressors prevent neovascularization and inflammatory cell accumulation and thus hinder
3
PC CH ESA Control
ProMMP-2
ActiveMMP-2
Fig. 4. Gelatin zymography of groups representative parietal peritoneum samples. PC: Positive Control
Fig. 4. Gelatin zymography of groups representative parietal peritoneum samples. PC: Positive Control

94 95Eur. J. Inflamm.
inflammatory peritoneum was lower than that of subperitoneal muscle and other tissues, yet this is contrary to current knowledge. According to these findings, peritoneal TIMP-2 levels were suppressed biochemically in CH-induced peritoneal fibrosis. In the darbepoetin group compared to control group, the TIMP-2 level increase due to inflammation might be related to decreased peritoneal fibrosis development. In our findings TIMP-2 levels were non-significantly high in the darbepoetin group compared to the CH group. This could be due to ongoing inflammation or low sample numbers in groups.
In the light of these findings we conclude that darbepoetin has its action on activation and increase of peritoneal TIMP-2, and acceleration of conversion of pro-MMP-2 to active MMP-2 to prevent peritoneal fibrosis. Other mechanisms also seem to play role in preventing peritoneal fibrosis on CH-induced peritoneal fibrosis in rats. We need further prospective studies to elucidate mechanisms responsible for the development of peritoneal fibrosis.
In conclusion, in this study we achieved parietal peritoneal fibrosis by injecting 15% ethyl alcohol and 0.1% CH dissolved in saline. Darbepoetin caused a histopathologic decrease of peritoneal fibrosis induced chemically by CH. Immunohistochemically, it suppressed peritoneal tissue MMP-2 compared to CH group and had no effect on TIMP-2. Biochemically, it augmented the ratio of pro- to active MMP-2 on the parietal peritoneum, but had no effect on TIMP-2 levels. The decrease in peritoneal fibrosis by darbepoetin can be attributed to its stimulatory effect on TIMP-2 activity, and on the conversion of pro- to active MMP-2. Other pathways may also play a role. We can say that darbepoetin does not cause peritoneal fibrosis even if it prevents it, but further clinical and experimental studies concerning dose and duration are needed.
ACKNOWLEDGEMENTS
This research was partially carried out at Dokuz Eylul University Medical School Learning Resources Center Research Laboratory (R-LAB).
This study was granted by Committee of Aid and Support for Dokuz Eylul Renal Patients with Foundation of Support for Investigations. No grants
were provided for darbepoetin alpha (Aranesp) supplied by Amgen Europe B.V.-Netherlands, nor from the Eczacibasi Drug Corporation.
REFERENCES
1. Krediet RT. 30 years of peritoneal dialysis development: the past and the future. Perit Dial Int 2007; 27:S35-41.
2. Hirahara I, Inoue M, Okuda K, Ando Y, Muto S, Kusano E. The potential of matrix metalloproteinase-2 as a marker of peritoneal injury, increased solute transport, or progression to encapsulating peritoneal sclerosis during peritoneal dialysis--a multicentre study in Japan. Nephrol Dial Transplant 2007; 22:560-7.
3. Margetts PJ, Bonniaud P. Basic mechanisms and clinical implications of peritoneal fibrosis. Perit Dial Int 2003; 23:530-41.
4. Miyata T, Devusyst O, Kurukowa K, Van Ypersele de Strihou C. Toward better dialysis compatibility advances in the biochemistry and pathophysiology of the peritoneal membranes. Kidney Int 2002; 61:375-86.
5. Williams JD, Craig KJ, Topley N, et al. Morphologic changes in the peritoneal membrane of patients with renal disease. J Am Soc Nephrol 2002; 13:470-9.
6. Heimburger O, Waniewski J, Werynski A, Tranaeus A. Peritoneal transport in CAPD patients with permanent loss of ultrafiltration capacity. Kidney Int 1990; 38:495-506.
7. Hirahara I, Umeyama K, Shofuda K, et al. Increase of matrix metalloproteinase-2 in dialysate of rat sclerosing encapsulating peritoneal model. Nephrology 2002; 7:161-9.
8. Nagase H, Visse R, Murphy G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovascular Research 2006; 69:562-73.
9. Johnson DW, Pat B, Vesey DA, Guan Z, Endre Z, Gobe GC. Delayed administration of darbepoetin or erythropoietin protects against ischemic acute renal injury and failure. Kidney Int 2006; 69:1806-13.
10. Brines ML, Ghezzi P, Keenan S, et al. Erythropoietin crosses the blood-brain barrier to protect against experimental brain injury. Proc Natl Acad Sci USA 2000; 97:10526-31.
A. BORAZAN ET AL.

94 95Eur. J. Inflamm.
11. Calvillo L, Latini R, Kajstura J, et al. Recombinant human erythropoietin protects the myocardium from ischemia-reperfusion injury and promotes beneficial remodeling. Proc Natl Acad Sci USA 2003; 100:802-6.
12. Lee SH, Li C, Lim SW, et al. Attenuation of interstitial inflammation and fibrosis by recombinant human erythropoietin in chronic cyclosporine nephropathy. Am J Nephrol 2005; 25:64-76.
13. Park SH, Choi MJ, Song IK, et al. Erythropoietin decreases renal fibrosis in mice with ureteral obstruction: role of inhibiting TGF-beta-induced epithelial-to-mesenchymal transition. J Am Soc Nephrol 2007; 18:1497-507.
14. Nishiya D, Omura T, Shimada K, et al. Effects of erythropoietin on cardiac remodeling after myocardial infarction. J Pharmacol Sci 2006; 101:31-9.
15. Bahlmann FH, Song R, Boehm SM, et al. Low-dose therapy with the long-acting erythropoietin analogue darbepoetin alpha persistently activates endothelial Akt and attenuates progressive organ failure. Circulation 2004; 110:1006-12.
16. Ishii Y, Sawada T, Shimizu A, Tojimbara T, et al. An experimental sclerosing encapsulating peritonitis model in mice. Nephrol Dial Transplant 2001; 16:1262-6.
17. Sis B, Sarioglu S, Sokmen S, et al. Desmoplasia measured by computer assisted image analysis: an independent prognostic marker in colorectal carcinoma. J Clin Pathol 2005; 5:32-8.
18. Sarioglu S, Sis B, Celik A, et al. Quantitative digital histochemistry with methenamine silver staining in renal allograft biopsies excluding pure chronic allograft nephropathy cases. Transplant Proc 2006; 38:490-1.
19. Sis B, Sarioglu S, Celik A, et al. Renal medullary changers in renal allograft recipients with raised serum creatinine. J Clin Pathol 2006; 59:377-81.
20. Sokmen S, Lebe B, Sarioglu S, et al. Prognostic value of CD44 expression in colorectal carcinomas.
Anticancer Res 2001; 21:4121-6.21. Cohen PA, Gower AC, Stucchi AF, Leeman
SE, Becker JM, Reed KL. A neurokinin-1 receptor antagonist that reduces intraabdominal adhesion formation increases peritoneal matrix metalloproteinase activity. Wound Repair Reg 2007; 15:800-8.
22. Wiechelman KJ, Braun RD, Fitzpatrick JD. Investigation of the bicinchoninic acid protein assay: identification of the groups responsible for color formation. Anal Biochem 1988; 175:231-7.
23. Kleiner DE, Stetler-Stevenson WG. Quantitative Zymography: detection of picogram quantities of gelatinases. Anal Biochem 1994; 21:325-9.
24. Clark IM, Powell LK, Wright JK, Cawston TE. Polyclonal and monoclonal antibodies against human tissue inhibitor of metalloproteinases (TIMP) and the design of an enzyme-linked immunosorbent assay to measure TIMP. Matrix 1991; 11:76-85.
25. Ersoy R, Celik A, Yilmaz O, et al. The effects of irbesartan and spironolactone in prevention of peritoneal fibrosis in rats. Perit Dial Int 2007; 27:424-31.
26. Martin J, Yung S, Robson RL, Steadman R, Davies M. Production and regulation of matrix metalloproteinases and their inhibitors by human peritoneal mesothelial cells. Perit Dial Int 2000; 20:524-33.
27. Margetts PJ, Bonniaud P. Basic mechanisms and clinical implications of peritoneal fibrosis Perit Dial Int 2003; 23:530-41.
28. Dollery CM, McEwan JR, Henney AM. Matrix metalloproteinases and cardiovascular disease. Circ Res 1995; 77:863-8.
29. Galis ZS, Khatri JJ. Matrix metalloproteinases in vascular remodeling and atherogenesis: The good, the bad, and the ugly. Circ Res 2002; 90:251-62.
30. Nagase H, Woessner JF Jr. Matrix metalloproteinases. J Biol Chem 1999; 274: 21491-4.

EUROPEAN JOURNAL OF INFLAMMATION Vol. 7, no. 2, 0-0 (2009)
1721-727X (2009)Copyright © by BIOLIFE, s.a.s.
This publication and/or article is for individual use only and may not be furtherreproduced without written permission from the copyright holder.
Unauthorized reproduction may result in financial and other penalties97

EUROPEAN JOURNAL OF INFLAMMATION Vol. 7, no. 2, 0-0 (2009)
1721-727X (2009)Copyright © by BIOLIFE, s.a.s.
This publication and/or article is for individual use only and may not be furtherreproduced without written permission from the copyright holder.
Unauthorized reproduction may result in financial and other penalties97
Inflammatory bowel disease (IBD), including Crohn’s disease and ulcerative colitis, are important problems in industrialized countries and have been associated with increased colon cancer risk (1). The complete aetiology of both diseases is still unknown but it is likely that it involves genetic, environmental and immunological factors. IBD is a chronic immune-mediated disease in which endogenous bacteria are thought to play an important role, as suggested by
numerous clinical observations and experimental studies. There is a growing body of evidence that supports the role of intestinal microbiota in the initiation and progression of IBD (2).
Experimental models of colitis are used because they imitate human IBD, ulcerative colitis, Crohn’s disease and chronic relapsing diseases of unknown aetiology (3). These models showed that probiotics can confer beneficial effects on
EFFECT OF YOGHURT ON THE CYTOKINE PROFILE USING A MURINE MODEL OF INTESTINAL INFLAMMATION
A. DE MORENO DE LEBLANC1, S. CHAVES1,2 and G. PERDIGÓN1,2
1Centro de Referencia para Lactobacilos (CERELA-CONICET), San Miguel de Tucumán; 2Cátedra de Inmunología, Facultad de Bioquimíca, Química y Farmacia,
Universidad Nacional de Tucumán, Argentina. Received November 12, 2008 – Accepted April 2, 2009
Mailing address: Gabriela Perdigón, CERELA-CONICET Chacabuco 145, San Miguel de Tucumán,Tucumán, T4000ILC, ArgentinaTel: ++54 381 4310465 ext. 129Fax: ++54 381 4005600e-mail: perdigon @cerela.org.ar
Key words: inflammation, yoghurt, cytokines, intestinal microbiota
Inflammatory bowel disease (IBD), including Crohn’s disease and ulcerative colitis, are important problems in industrialized countries. The complete aetiology of both diseases is still unknown but likely involves genetic, environmental and immunological factors. The aim of this work is to study the anti-inflammatory mechanisms reported for yoghurt in a colon cancer model in order to evaluate its usefulness in the treatment of intestinal inflammation such as Crohn’s disease. A trinitrobenzenesulfonic acid (TNBS)-induced colitis model was used. The influence of yoghurt feeding was studied before and after TNBS inoculation. The effect on the intestinal microbiota and on the host immune response was evaluated. IgA-producing cells and cells positive for specific cytokines (IL-12, IL-17, IFNγ and IL-10) were analyzed. Yoghurt administration diminished the severity of inflammation in the TNBS-inoculated mice. This effect occurred mainly through IL-10, which was increased in the intestinal tissues throughout the study, and by the decrease observed in IL-17 and IL-12 levels. In addition to this immunomodulatory capacity, another mechanism by which yoghurt could exert the anti-inflammatory activity observed in our model would be through beneficial changes in the intestinal microbiota (increases in the bifidobacteria and lactobacilli populations). These changes in the intestinal microbiota could also be considered one of the causes of the intestinal inflammation reduction. These results show that yoghurt administration modulated the immune response, inducing down regulation of the inflammatory cytokines produced by the immune cells involved in the inflammatory process. The protective effect of yoghurt could also be mediated through beneficial changes in the intestinal microbiota favouring lactobacilli and bifidobacteria population.

98 99Eur. J. Inflamm.
anti-inflammatory mechanisms previously reported for yoghurt in a colon cancer model to evaluate whether this product could prevent or be useful for the treatment of intestinal inflammation. In order to meet this objective, a TNBS induced colitis model was used and the influence of yoghurt feeding on the intestinal microbiota and on the anti-inflammatory gut immune response was evaluated.
MATERIALS AND METHODS
Experimental groupsFive-week-old female BALB/c mice weighing 25-28
g were obtained from the random-bred colony maintained at CERELA and divided into five experimental groups. 1) TNBS group: The mice received inoculation of trinitrobenzenesulfonic acid (TNBS) to induce inflammation; 2) yoghurt-TNBS-yoghurt group: The mice were fed with yogurt for ten consecutive days (basal yogurt), treated with TNBS, and then fed again with yogurt; 3) yoghurt-TNBS group: The mice were given yogurt for 10 consecutive days and then inoculated with TNBS to study the effect of yoghurt on the prevention of the inflammation; 4) Control group: The mice were inoculated with the TNBS vehicle (phosphate buffered saline, PBS in ethanol 50% without TNBS); 5) yogurt control group: Mice were fed with yogurt during all the experiment but without TNBS inoculation.
All groups were fed ad libitum with a balanced diet and were maintained in a room with a 12-h light/dark cycle at 18±2ºC. Each experimental group consisted of 20-25 mice.
All animal protocols were preapproved by the Animal Protection Committee of CERELA and all experiments complied with the current laws of Argentina.
Yoghurt preparationSimulated commercial yogurt was freshly prepared
and controlled every day to keep the number of bacteria constant and to avoid variations due to storage. Yoghurt was prepared from cultures of L. delbrueckii subsp. bulgaricus and S. thermophilus (strain pools were used from the CERELA culture collection). The total number of bacteria in the fermented product was 2×109 CFU/ml.
Induction of intestinal inflammation and feeding procedure
For induction of the intestinal inflammation, mice (which were anesthetized intraperitoneally using a mix of ketamine hydrocholoride (Hollyday-Scott S.A., Buenos Aires, Argentina), 100 µg/g body weight, and xylazine hydrochloride (Rompun, BAYER, Division Sanidad
intestinal inflammation partially because of their immunomodulatory effects. Probiotics are defined as live microorganisms which, when administered in adequate amounts, confer a health benefit to the consumer (4). Probiotics can be useful in the prevention or treatment of IBD because they can improve the intestinal microbial balance and the intestine immunologic barrier, particularly through immunoglobulin (Ig) A response and alleviation of intestinal inflammatory responses, which produce gut-stabilizing effects (5).
Lactic acid bacteria (LAB) are present in many foods, including yoghurt, and are frequently used as probiotics to favour biological functions in the host. Yoghurt has been defined in the Codex Alimentarius (2002) as a coagulated milk product that results from the lactic acid fermentation of milk by Lactobacillus (L.) delbrueckii subsp. bulgaricus and Streptococcus (S.) thermophilus. In this definition, in addition to the yoghurt starter cultures, the peptides and other substances released during the fermentation process can also be important. Many researchers have studied the therapeutic effects of yogurt and LAB commonly used in yogurt production against diseases such as cancer, infection, and gastrointestinal disorders. The immunomodulating and immunostimulating properties of yogurt have also been well documented (6). The regulation of the inflammatory immune response is one of the mechanisms by which yoghurt could prevent the risk of colon cancer in an experimental model, and could be effective in models of intestinal inflammatory disease.
Modification of the Th1 and Th2 subpopulations in favour of the Th1 population seems to be one of the mechanisms that can produce the development of chronic intestinal inflammatory diseases. This concept is reinforced by different studies that demonstrated increases in pro-inflammatory cytokines produced by this subpopulation. In this way, the suppression or regulation of Th1 response can be an alternative for the treatment of IBD in animals and humans. Numerous works have shown the importance of regulating IL-10 cytokine in the modulation of the immune response at intestinal level, and mice deficient in this gene that encodes this cytokine have been used as models of intestinal inflammation (7).
The aim of the present work is to study the
A. DE MORENO ET AL.

98 99Eur. J. Inflamm.
Animal, Buenos Aires, Argentina), 5 µg/g body weight) received by intrarectal administration 100 µl of a solution of TNBS (Sigma, St Louise; 2mg/mouse) dissolved in PBS 0.01 M, pH 7.4 and mixed with an equal volume of ethanol (50% ethanol). Control mice received PBS mixed with ethanol (without TNBS) using the same technique.
Body weight of mice was controlled every day until the end of the experiment.
Sampling procedures and colon histology The following samples were obtained from each group:
basal sample (day 0, the same day of TNBS inoculation and previous to it), and 3, 7 or 14 days after TNBS inoculation. Ten mice for each group were sacrificed at each sampling period. Five mice were sacrificed for histological and immunohistochemical evaluations of the large intestine. The experiments were repeated 3 times. Serial paraffin sections of 4 µm were made and stained with hematoxilin-eosin for light microscopy examination. For the colon histology, the microscopic slides were reviewed and the extent of colonic damage and inflammation was assessed using the histopathological grading system of Ameho et al (8) (Table I). The other five mice (N=5) of each group were analyzed for the liver translocation and to study the microbiota in the large intestine.
Study of the intestinal microbiotaThe large intestine of mice were aseptically removed,
weighed and placed into sterile tubes containing 5 ml of peptone water (0.1%). The samples were immediately homogenized under sterile conditions using a homogenizer (MSE, England). Serial dilutions of the homogenized samples were obtained and aliquots (0.1 ml) of the appropriate dilution were spread onto the surface of the following agarized media: Reinforced Clostridial agar (RCA, Britania, Buenos Aires, Argentina) for total anaerobic bacteria; RCA containing 0.2% LiCl, colistin 4mg/L, 1% aniline blue and after sterilization adjusted to pH 5 with acetic acid (RCA-pH5) for isolation of bifidobacteria; Mann-Rogosa-Sharp (MRS Britania, Buenos Aires, Argentina) for total lactobacilli and MacConkey (Britania, Buenos Aires, Argentina) for Enterobacteriaceae. This last culture media was aerobically incubated at 37ºC for 24 h, all others plates were anaerobically incubated at 37ºC for 72-96 h.
Determination of bacterial translocation to liverLivers were aseptically removed, weighed and placed
into sterile tubes containing 5 ml of peptone water and mechanically homogenized under sterile conditions using a homogenizer (MSE, England). The samples were homogenized, and cultures were performed on Mann-Rogosa-Sharp Agar, MacConkey agar and LAPTg
agar (yeast extract, peptone, tryptone and glucose). The cultures were aerobically incubated at 37°C for 24-72 h.
Immunofluorescence assay for IgA-secreting cells in the large intestine
Large intestine tissue sections (4 µm) from each mouse were used for the immunofluorescence assays. The numbers of IgA(+) cells were determined by direct immunofluorescence assay. Slides were incubated with α-chain monospecific antibody conjugated with fluorescein isothiocyanate (FITC, Sigma, St Louis, USA). The number of fluorescent cells was counted in 30 fields of vision, as seen at 1,000x magnification using a fluorescent light microscope. The results were expressed as positive cells in ten fields of vision.
Cytokine-producing cell determination in histological sections
Cytokine positive cells were detected by indirect immunofluorescence on large intestine tissues, following the technique described by de Moreno de LeBlanc and Perdigon (6). Rabbit anti-mouse IFNγ and IL-10, or goat anti-mouse IL-12 polyclonal antibodies (Peprotech, Inc. Rocky Hill, NJ, USA) and goat anti-mouse IL-17 polyclonal antibody (R&D System, Minneapolis, USA) (diluted in saponin-PBS) were applied to the sections for 75 min at room temperature (21ºC). The sections were then treated with a dilution of goat anti-rabbit antibody conjugated with fluorescein isothiocyanate (FITC, Jackson Immuno Research, Labs. Inc., West Grove, USA). The number of fluorescent cells was counted in thirty fields of vision as seen at 1,000x magnification using a fluorescence light microscope and expressed as number of positive cells in ten fields of vision.
Statistical analysisStatistical analyses were performed using MINITAB
14 software (Minitab, Inc., State College, PA) by ANOVA GLM followed by a Tukey’s posthoc test, and P < 0.05 was considered significant. Unless otherwise indicated, all values (N=15) were the means of 3 independent trials (no significant differences were observed between individual replicates) ± standard deviation.
RESULTS
Effect of yoghurt in TNBS-treated mice. Analysis of mortality, body weight and intestinal histology
Mice treated with TNBS (2mg/mouse) showed 25-30% mortality rate which was associated with a significant loss of body weight (Fig. 1) and high

100 101Eur. J. Inflamm.
levels of colitis (intestinal histological alterations). The TNBS control group showed the highest weight loss that became significant 3 days post inoculation (27.4 ± 1.5 and 22.5 ± 1.4 for 1 and 3 days, respectively); severe mucosal damage that included a loss of crypts, necrosis, and focal influx of inflammatory cells in the mucosa and submucosa were observed (Fig. 2E). The higher score of colonic damage was obtained 7 days post-TNBS (4.7±1.2, Table II). Decrease in the goblet cell counts was another characteristic of this group. Premature death resulted mainly from an excessive inflammatory
reaction, as assessed by post-mortem autopsy and considering the histology and other parameters studied in the mice sacrificed during the study, this group should have a greater mortality rate.
The greater number of mice that received exclusively the vehicle had similar macroscopic and microscopic appearance of their colons compared to the normal mice (Fig. 2A). A small number of these animals had mucosal or submucosal infiltrates focalized in regions of the colon.
The administration of yoghurt increased mononuclear cell numbers in the lamina propria of
Table I. Histopathological grading scale of chemically induced colitis.
Grade Microscopic findings
0 Histological findings identical to normal mice 1 Mild mucosal and/or submucosal inflammatory infiltrate (admixture of
neutrophils) and oedema. Punctate mucosal erosions often associated with capillary proliferation. Muscularis mucosae intact
2 Grade 1 changes involving 50% of the specimen. 3 Prominent inflammatory infiltrate and oedema (neutrophils usually
predominating) frequently with deeper areas of ulceration extending through the muscularis mucosae into the submucosa. Rare inflammatory cells invading the muscularis propriae but without muscle necrosis
4 Grade 3 changes involving 50% of the specimen 5 Extensive ulceration with coagulative necrosis bordered inferiorly by
numerous neutrophils and lesser numbers of mononuclear cells. Necrosis extends deeply into the muscularis propria
6 Grade 5 changes involving 50% of the specimen
Table I. Histopathological grading scale of chemically induced colitis.
Table II. Colon microscopic scores of mice.
Days post-TNBS Group
3 7 13
Control 0.8±0.7a 0.6±0.4a 0
Y-TNBS 1.6±1a,b 2.9±0.9b,c 1.5±1a,b
Y-TNBS-Y 1.4±0.9a,b 2.5±0.6b 1.2±0.7a,b
TNBS 2.3±0.7b 4.7±1.2c 3.3±0.6c
Each value represents the mean of N = 15 ± SD. Means for each value without a common letter differ significantly (p < 0.05). Y = yoghurt
Table II. Colon microscopic scores of mice.
Each value represents the mean of N = 15 ± SD. Means for each value without a common letter differ significantly (p < 0.05). Y = yoghurt
A. DE MORENO ET AL.

100 101Eur. J. Inflamm.
the large intestine as was reported previously by de Moreno de LeBlanc (9) but these were focalized near other histological unaltered areas (Fig. 2B).
Ten days of yoghurt feeding previous to the TNBS inoculation did not significantly decrease the mortality rate, but the body weight did not decrease significantly compared with the basal data. Mice from this group showed an important reduction of the colonic damage caused by the inflammatory agent (Table II). Although some infiltrates in mucosa and submucosa were present, most of the tissues of the large intestine were similar to those observed in the alcohol control animals (Fig. 2C).
The group of mice that received continuous administration of yoghurt (before and after TNBS) showed decreased mortality rates compared to the inflammation control group, however these animals did not significantly lose body weight ,and the histological observations showed similar characteristics to the last described group with an
important increase in the number of goblet cells (Fig. 2D and Table II).
Changes in the intestinal microbiota by yoghurt administration and bacterial translocation to liver
Mice fed 10 days with yoghurt showed a significant increase in the bifidobacteria counts (mean log CFU of 6.5 ± 1.0) compared to the control group that did not receive fermented milk (3.1 ± 0.2, Fig. 3A).
Another bacterial population that increased significantly in mice receiving yoghurt was total lactobacilli (8.0 ± 0.3 and 7 ± 0.1 for yoghurt and control group, respectively; (Fig. 3A).
These increases (bifidobacteria and lactobacilli) were maintained in mice that received yoghurt after TNBS inoculation but not in mice that stopped yoghurt administration (yoghurt-TNBS group, Fig. 3B). Significant differences between groups were not observed for the other culture media assayed
Table III. Bacterial translocation to the liver.
Group Sample MacConkey LAPTg MRS Control Basal WG 2.4±1.0 a WG 3d post TNBS 2.1±0.9 a,b 3.8±1.3 a 2.4±0.8 a,b
7d post TNBS WG 0.9±0.7 a 1.6±0.1 a
13d post TNBS WG WG WG Yoghurt Basal WG 2.5±1.5 a WG 3d post TNBS 1.7±0.3 a 2.3±1.8 a 1.7±0.1 a
7d post TNBS 1.6±0.9 a,b 1.9±1 a,b 1.6±0.2 a
13d post TNBS WG 1.5±0.9 a WG Y-TNBS Basal WG 2.5±1.5 a WG 3d post TNBS 3.9±1.6 b,c 4.9±1.1 a 2.4±1.2 a,b
7d post TNBS WG 1.4±0.6 a 2.9±2.0 a
13d post TNBS WG WG 3±1.6 a
Y-TNBS-Y Basal WG 2.5±1.5 a WG 3d post TNBS 3.0±1.0 a,b 4.0±1.5 a 2.2±0.6 a,b
7d post TNBS 1.3±0.7 a 1.7±0.7 a 1.5±0.1 a
13d post TNBS WG WG 2.7±1.0 a
TNBS Basal WG 2.4±1.0 a WG 3d post TNBS 5.2±0.5 c 3.6±0.9 a 3.9±2.0 b
7d post TNBS 2.9±0.8 b 4.0±1.4 b 5.1±1.4 b
13d post TNBS WG 0.3±0.2 b 2.9±0.2 a
Colony counts are expressed as log10 numbers of bacteria per gram of liver. Each value represents the mean of N = 15 ± SD. For each time point, comparing all the groups, means for each culture medium without a common letter differ significantly (p < 0.05). WG = without growth in the sample without dilution, Y = yoghurt
Table III. Bacterial translocation to the liver.
Colony counts are expressed as log10 numbers of bacteria per gram of liver. Each value represents the mean of N = 15 ± SD. For each time point, comparing all the groups, means for each culture medium without a common letter differ significantly (p < 0.05). WG = without growth in the sample without dilution, Y = yoghurt

102 103Eur. J. Inflamm.
results were compared to the yoghurt control group.
Study of IgA+ cells in the large intestine Yoghurt feeding increased the number of IgA+
cells in the large intestine of mice; this was observed in the yoghurt control group in the second and third samples. When mice were treated with TNBS, yoghurt administration increased the number of IgA+ cells compared to the TNBS control group. When the fermented milk was administered continuously; this effect was significant in the last sample (74 ± 10 for yoghurt-TNBS-yoghurt group and 57 ± 6 for TNBS group, Fig. 4).
Study of cytokine positive cells in the large intestinesTNBS-induced inflammation increased the
(data not shown). Bacterial translocation in the liver showed that
mice treated with TNBS (TNBS group) increased the number of CFU in the three growth media assayed (MacConkey, LAPTg and MRS). This increase was higher on days 3 and 7 post-induction of the inflammation and decreased afterwards, when the mice that survived improved their inflammatory condition (Table III). Mice from the group that received yoghurt 10 days before TNBS inoculation showed lower translocation than the TNBS group, and values returned to basal numbers of CFU in liver at day 7 post-TNBS. Similar results to this last group were obtained in mice that received continuous yoghurt administration (before and after TNBS). No significant differences were observed when these
days after TNBS inoculation
0 1 3 5 7 9 11 13
Bod
y w
eigh
t (g)
16
18
20
22
24
26
28
30
32
34
Mor
talit
y (p
erce
ntag
e)
0
20
40
60
80
100
*
Fig. 1.
Fig. 1. Changes in the body weight, mortality percentage and histological parameters. Weight changes of normal control mice treated with 50% ethanol (black whole line and circle) and the test groups: yoghurt control (black dotted line and empty square); yoghurt-TNBS (black long dash line and gray triangle); yoghurt-TNBS-yoghurt (black whole line and gray square); TNBS (black long dash line and diamond). Weight data from all experiments are shown. Each point represents average ± SD of weight data pooled from 45 mice (15 mice from each separate trial). * significant difference (p < 0.05) is observed only for TNBS group 3 days after TNBS inoculation, compared with the basal data for the same group. Mortality was only observed for the groups inoculated with TNBS: yoghurt-TNBS (gray bars); yoghurt-TNBS-yoghurt (diagonally lined bars) and TNBS (black bars). Results are expressed as means ± SD of the mortality of 45 mice (15 mice in each three different experimental trials) for each group during the period of the experiment.
A. DE MORENO ET AL.
102 103Eur. J. Inflamm.
number of IFNγ and IL-12 positive cells in the two first samples (29 ± 8 and 24 ± 3 for IFNγ, and 25 ±3 and 20 ± 2 for IL-12, 3 and 7 days post TNBS, respectively (Fig. 5). Yoghurt feeding decreased the number of positive cells for both cytokines, IFNγ in the two same time points and IL-12 in the second sample taken 7 days post-TNBS (16 ± 5 and 17 ± 2 for previous and continuous yoghurt feeding), showing a regulation of the inflammatory immune response. For IL-12+ cells, mice given yoghurt (yoghurt control group) did not show significant differences compared to the control group without inflammation (alcohol group). IL-17 was another cytokine studied because it has been reported in several inflammatory pathologies. In our model, IL-17+ cells increased in all the groups treated with TNBS, 3 days after the inoculation (32 ± 6, 23 ± 10 and 22 ± 8 for TNBS, yoghurt-TNBS and yoghurt-TNBS-yoghurt groups, respectively) compared with the control group without inflammation (12 ± 1). In the other two samples taken 7 and 13 days post-TNBS, the increased number of these cells was maintained for the TNBS group, but not for the mice given yoghurt (Fig. 5).
Yoghurt feeding by itself did not increase IL-12 or IL-17 secreting cells, compared to the control without inflammation, but in the assay for IFNγ+ cells it was observed that yoghurt administration significantly increased the number of the cells positive for this cytokine in the basal sample and 3 days post-TNBS.
IL-10 was another cytokine studied and it was observed that mice fed with yoghurt increased numbers of IL-10+ cells compared to the TNBS control (Fig. 5). These increases were higher in mice that received continuous yoghurt administration, including after TNBS inoculation.
DISCUSSION
Previous works showed that yoghurt can prevent the growth of a chemically induced colon cancer in mice. This effect was attributed to its anti-inflammatory activity. Yoghurt can modulate the immune response by 1) stimulating cytokine production when this is required, or 2) inducing down-regulation of the immune cells to avoid an exacerbated immune response. This effect would
Fig. 2. Fig. 2. Histological study comparing the different experimental groups. Slices from large intestine of mice were studied after staining with hematoxilin-eosin. All the samples were obtained 7 days post-TNBS with a magnification of 100x. A) Alcohol control group. B) Yoghurt group. Tissue maintained the typical structure of the large intestine but it is possible to observe immune cell infiltration in specific areas (arrows). C) and D) Yoghurt-TNBS and yoghurt TNB-yoghurt group: In both groups there are cells infiltrating mucosa and submucosa (arrows) but most of the tissues of the large intestine were similar to those observed in the alcohol control animals. E) TNBS group: Severe mucosal damage is observed with loss of crypts (encircled), focal influx of inflammatory cells in the mucosa and submucosa (arrows). Decrease in the goblet cell was another characteristic of this group.

104 105Eur. J. Inflamm.
administration of yoghurt showed decreased mortality rates compared to the inflammation control (Fig. 1).
These results led us to study the possible mechanisms by which yoghurt feeding can exert its beneficial effects on the TNBS-treated mice.
Most of the models in which animals develop spontaneous or chemically-induced colitis are influenced by the microbiota present in the intestinal lumen. This fact is supported by the reduction or absence of intestinal inflammation in TNBS or dextran sulphate sodium (DSS) models of colitis using antibiotic-treated and germ-free animals (13-14). In addition, many studies have shown
occur mainly through IL-10, which was increased in the tissue of the mice receiving yoghurt (10).
In the present study, the effect of yoghurt was evaluated using a murine model of intestinal inflammation. The inflammation induced by TNBS was selected as a model because it shares important similarities with human Crohn’s disease such as transmural inflammation, lymphocyte infiltration and Th1-dominated cytokine profiles (11-12).
In the present work it was observed that yoghurt feeding previous to the TNBS inoculation or continuously (before and after TNBS), reduced the colonic damage caused by the inflammatory agent (Table II); the group of mice that received continuous
Mac Conkey LAPTg MRS RCA-pH5 RCA
Log1
0 C
FU /
g of
LI
0
2
4
6
8
10
aa
a
aa b
a
ba
b
Days post TNBS
-4 -2 0 2 4 6 8 10 12 14 16 18
Log
CFU
/ g
of L
I
0
2
4
6
8
10
A)
B)
Fig. 3.
Fig. 3. Effect of yoghurt feeding and TNBS inoculation on the microbiota of the large intestine. Results are expressed as means ± SD of the log10 CFU/g large intestine (LI). A) Represents the basal data, previous to the TNBS inoculation, for control mice (black bars) and mice fed 10 days with yoghurt (gray bars). B) Represents the results obtained for bifidobacteria (RCA-pH5 medium) in all the groups assayed. Mice from yoghurt-TNBS group showed growth in this culture medium only in the first sample (3 days post TNBS). Mice fed continuously with yoghurt [yoghurt (diagonal bars) and yoghurt-TNBS-yoghurt (horizontal bars) groups] showed growth in all the samples assayed. For the other groups (50% ethanol control and TNBS) no colonies were present in the lowest dilution assayed (10-2). Each mean represents data from five animals. a–bMeans for each medium without a common letter differ significantly (p < 0.05).
A. DE MORENO ET AL.

104 105Eur. J. Inflamm.
that not all bacterial species have equal activities in promoting or reducing intestinal inflammation.
In the present study, when the basal sample, taken after 10 days of yoghurt feeding (the day of TNBS inoculation), was compared to mice that did not receive yoghurt previously, changes in the intestinal microbiota were observed. Yoghurt feeding increased the bifidobacteria and lactobacilli counts. The relationship between the administration of fermented milks containing lactobacilli and the increase in Bifidobacterium counts has been previously reported (15). This fact can be related with recent findings that some LAB strains have metabolic pathways needed for the synthesis and release of molecules that selectively stimulate the growth of endogenous bifidobacteria. It has been suggested that the increase of these bacteria could confer a beneficial effect on the stability of the intestinal microbiota (16). The increase in lactobacilli count was in agreement with other publications where the intake of probiotic bacteria or fermented milk caused the same effects (17).
Bifidobacteria and lactobacilli are known to be
desirable microbiota inhabitants of the intestines and are related with many beneficial effects in the gastrointestinal tract such as the prevention of inflammation.
The TNBS model is associated with the absence of LAB and an increase in other aerobic isolates such as Escherichia coli and Staphylococcus spp. (18). Similarly, decreased levels of faecal lactobacilli and bifidobacteria have also been reported in Crohn’s disease (19).
The mechanisms whereby probiotics exert their effects in general, and in IBD in particular, are not yet fully understood. Some LAB and bifidobacteria can produce and release specific low molecular weight anti-microbial substances (bacteriocins) (20). Some probiotic bacteria are capable of normalizing increased intestinal permeability to potentially antigenic macromolecules (21); another characteristic of intestinal inflammation is the increase in the permeability of the colonic mucosa barrier and a decrease in IgA levels. All these parameters contribute to bacterial translocation to
Days post TNBS
0 3 7 13
N°
of p
ositi
ve c
ells
/ 10
file
ds
0
20
40
60
80
100
120
140
a,b,c
a,c
a,b,d
a,b b
c c c
d b,d
a,b,c
a,c
c
a,b,da,b
a,ba,c
Fig. 4.Fig. 4. Effect of TNBS-induced inflammation and yoghurt feeding on IgA+ cells in the large intestine. Positive cells were counted in histological sections from large intestine of alcohol control (black bars), yoghurt (diagonally lined bars), yoghurt-TNBS (light gray bars), yoghurt-TNBS-yoghurt (horizontal lined bars) and TNBS (dark gray bars) groups. Data correspond to the means ± SD of results of 15 animals from three separate experiments. a,b,c,dMeans for each value without a common letter differ significantly (p < 0.05).

106 107Eur. J. Inflamm.
other organs. In this study, the lesser extent of colonic damage in
the intestine from mice which received yoghurt was related with a decrease in the bacterial translocation to the liver, compared to the mice from the TNBS control group (Table III). All these observations confirm that yoghurt feeding diminished the severity of inflammation in the mice and that changes in the intestinal microbiota at the colitis induction site could be a contributing factor to this beneficial effect.
Considering previous results where yoghurt feeding showed anti-inflammatory properties by modulating the immune response (22), the study of its effect on the intestinal immunity was evaluated.
Probiotics can modify the immune functions of
the host, although the specific mechanisms remain unclear. Enhanced intestinal IgA production, which provides defence for the mucosal surface (23), has been reported as one of the beneficial effects associated to probiotic consumption.
In the present study, yoghurt feeding increased the number of IgA+ cells in the large intestine of mice; this was observed in the yoghurt control group and also in mice treated with TNBS, in which yoghurt administration increased the number of IgA+ cells compared to the TNBS control group (Fig. 4).
These results agree with the increase of IgA reported in connection with the administration of L. rhamnosus GG in Crohn’s disease (24) and with our previous results where the administration of yoghurt
IFN�
Days post-TNBS
0 3 7 13
N°
cells
/ 10
fiel
ds
0
10
20
30
40
IL-12
Days post TNBS
0 3 7 13
N°
cells
/ 1
0 fie
lds
0
5
10
15
20
25
30
IL-17
Days post TNBS
0 3 7 13
N°
cells
/ 1
0 fil
eds
0
10
20
30
40
IL-10
Days post TNBS
0 3 7 13
N°
cells
/ 10
fiel
ds
0
5
10
15
20
25
30
a,b
a,c
b
a,b,ca,c
c
a,b,c
a,b
a,ca,c
a,c
a,b,c
a,b
a,b
b
a
a,ca,c
c
a,ca,c
a,b
a,c a,c
a,b
aa,b,c
a
b
a,ba,b
a,b,c,dc,d
c
d
a,b,c
a,c
a,b
a,b,ca
a
b
a,b
a,c
a,c
c
a
a,ba,b
a
c
a
aa,b
c
a,c
Fig. 5. Fig. 5. Effect of TNBS inoculation and yoghurt feeding on the cytokine+ cells of the large intestine. Positive cells for each cytokine were counted in histological sections from large intestine of alcohol control group (black bars), yoghurt (diagonally lined bars), yoghurt-TNBS (light gray bars), yoghurt-TNBS-yoghurt (horizontal lined bars) and TNBS (dark gray bars) groups. Data correspond to the means ± SD of results from 15 animals of three independent experiments. a,b,c,dMeans for each cytokine without a common letter differ significantly (P < 0.05).
A. DE MORENO ET AL.

106 107Eur. J. Inflamm.
increased the number of IgA+ cells in a colon cancer model in mice. The increase in this cell population but not in IgG secreting cells in the large intestine of mice fed with yoghurt could limit the inflammatory response, since IgA is considered an important barrier in colonic neoplasia (10).
An effective inflammatory immune response initially requires recruitment of immunocompetent cells to the site of inflammation and subsequently appropriate activation and regulation. Cytokines play a critical role in this setting, since they regulate the proliferation and differentiation of T cells and determine the course of an inflammatory process by releasing pro- and anti-inflammatory cytokines. The chronic intestinal inflammation model has been presumably driven by a Th1-predominant immune response which results in accumulation and activation of T lymphocytes and macrophages. By orchestrating the inflammatory process, pro-inflammatory cytokines such as interleukin IL-12 and IFNγ are thought to promote the development of these diseases (25).
IL-12 plays a pivotal role in Thl T cell differentiation and induces naive T cells to produce IFNγ. Mannon et al. (26) prompted a successful clinical trial where intestinal inflammation was treated with a monoclonal antibody against IL-12, considering that previous studies demonstrated that the expression of IL-12, especially the IL- 12p40 subunit is greatly enhanced in patients with Crohn’s disease. In our model, yoghurt feeding decreased the number of positive cells for both IFNγ and IL-12 in the large intestine of mice treated with TNBS after 13 days. This effect was observed with the previous or continuous yoghurt administration. With regard to IFNγ, increases in the number of cells that produce this cytokine were observed in mice given yoghurt in the basal sample and three days post-TNBS. This observation agrees with previous results where yoghurt feeding increased the number of IFNγ+ cells, but this fact was not related with inflammation in the intestine. It has been suggested that the large number of these positive cells in mice fed yoghurt could be related with the increase in immune cell numbers observed in the intestine. IFNγ would also be regulated by other cytokines such as IL-10 (10).
The recently described IL-17 expressing T helper cells (Th-17) may play a central role in T
cell-mediated diseases including IBD (27). IL-17 has been found to be elevated in a variety of inflammatory conditions including IBD (28). In this sense, it is now a general concept that IL-17-stimulated production of inflammatory mediators is a key element in the inflammatory cascade in a variety of pathological conditions.
The study of the IL-17+ cells confirmed the anti-inflammatory effect of yoghurt in our model. Yoghurt feeding by itself did not increase this cytokine. In contrast to mice treated with TNBS which maintained the IL-17+ cells increased during all the experiment, the mice that received yoghurt decreased the cells positive for this cytokine in the samples taken after 7 days.
In addition, considering the results obtained with the pro-inflammatory cytokines where yoghurt administration induced a down regulation of the inflammatory response, IL-10 was studied as a regulatory cytokine. Berg et al. (7) showed the role of IL-10 in intestinal inflammation and carcinogenesis. Mice with a disruption of the IL-10 gene showed inflammatory changes in ceacum, colon and rectum with a high incidence of colorectal adenocarcinomas. IL-10 also participates in the normal tolerance to indigenous microbiota and its deficiency is related to inflammation (29).
In this study, the number of IL-10+ cells increased significantly in the mice fed with yoghurt compared to the TNBS group: This increase was more pronounced in continuous feeding than when yoghurt administration was suspended.
This last observation showed that IL-10 could be one of the mechanisms by which yoghurt can exert its anti-inflammatory effect. Interestingly, it was reported that the induction of colitis can be prevented by co-transfer of syngeneic CD4+CD45RBlow T cells (30). These CD4+CD45RBlow T cells are shown to exert their inflammatory effect via production of IL-10 and transforming growth factor-β (TGF-β).
The results obtained suggest that yoghurt could modulate the inflammatory cytokines IL-12 and IL-17 inducing down regulation of the immune cells involved in the inflammatory process and favouring IL-10 anti-inflammatory cytokine production.
In addition to this immunomodulatory capacity, another mechanism by which yoghurt could exert

108 109Eur. J. Inflamm.
the anti-inflammatory activity observed in our model would be through beneficial changes in the intestinal microbiota with increases in the bifidobacteria and lactobacilli populations.
The results obtained also showed the importance of continuous administration of yoghurt, even after the inflammation. This observation was related with the beneficial changes in the intestinal microbiota and the regulation of the immune system maintained with the continuous yoghurt consumption.
Studies of other mechanisms such as cells apoptosis and the innate immune cells involved in the anti-inflammatory effect of yoghurt are currently under investigation.
ACKNOWLEDGEMENTS
This work was financially supported by Agencia Nacional de Promoción Científica y Tecnológica (ANPyCT-FONCYT, PICT 05/33213), Consejo de Investigación de la Universidad Nacional de Tucumán (CIUNT D351) and Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICET, PIP 5445), Argentina.
REFERENCES
1. Gillen CD, Walmsley RS, Prior P, Andrews HA, Allan RN. Ulcerative colitis and Crohn’s disease: a comparison of the colorectal cancer risk in extensive colitis. Gut 1994; 35:1590-2.
2. Linskens RK, Huijsdens XW, Savelkoul PH, Vandenbroucke-Grauls CM, Meuwissen SG. The bacterial flora in inflammatory bowel disease: current insights in pathogenesis and the influence of antibiotics and probiotics. Scand J Gastroenterol Suppl 2001; 234:29-40.
3. Fiocchi C. Inflammatory bowel disease: etiology and pathogenesis. Gastroenterology 1998; 115:182-205.
4. FAO/WHO. Evaluation of health and nutritional properties of powder milk and live lactic acid bacteria. Food and Agriculture Organization of the United Nations and World Health Organization Expert Consultation Report 2001; Available from www.fao.org/es/ESN/probio/probio.htm .
5. Isolauri E. Probiotics in human disease. Am J Clin Nutr 2001; 73:1142S-46.
6. de Moreno de Leblanc A, Perdigon G. Yogurt feeding inhibits promotion and progression of experimental colorectal cancer. Med Sci Monit 2004; 10:BR96-104.
7. Berg DJ, Davidson N, Kuhn R, Muller W, Menon S, Holland G, Thompson-Snipes L, Leach MW, Rennick D. Enterocolitis and colon cancer in interleukin-10-deficient mice are associated with aberrant cytokine production and CD4(+) TH1-like responses. J Clin Invest 1996; 98:1010-20.
8. Ameho CK, Adjei AA, Harrison EK, et al. Prophylactic effect of dietary glutamine supplementation on interleukin 8 and tumour necrosis factor alpha production in trinitrobenzene sulphonic acid induced colitis. Gut 1997; 41:487-93.
9. de Moreno de LeBlanc A, Perdigon G. Antitumour activity of yoghurt. In: Progress in Colorectal Cancer Research. J. D. Martinez ed. Nova Science Publisher Hauppauge, NY, USA 2005, p. 115-40.
10. de Moreno de LeBlanc A, Matar C, Perdigon G. The application of probiotics in cancer. Br J Nutr 2007; 98:S105-10.
11. Neurath M, Fuss I, Strober W. TNBS-colitis. Int Rev Immunol 2000; 19:51-62.
12. te Velde AA, Verstege MI, Hommes DW. Critical appraisal of the current practice in murine TNBS-induced colitis. Inflamm Bowel Dis 2006; 12:995-9.
13. Rath HC, Wilson KH, Sartor RB. Differential induction of colitis and gastritis in HLA-B27 transgenic rats selectively colonized with Bacteroides vulgatus or Escherichia coli. Infect Immun 1999; 67:2969-74.
14. Chandran P, Satthaporn S, Robins A, Eremin O. Inflammatory bowel disease: dysfunction of GALT and gut bacterial flora (I). Surgeon 2003; 1:63-75.
15. Garrido D, Suau A, Pochart P, Cruchet S, Gotteland M. Modulation of the fecal microbiota by the intake of a Lactobacillus johnsonii La1-containing product in human volunteers. FEMS Microbiol Lett 2005; 248:249-56.
16. Mitsuoka T. Bifidobacteria and their role in human health. J Ind Microbiol 1990; 81:131-36.
17. Tannock GW, Munro K, Harmsen HJ, Welling GW, Smart J, Gopal PK. Analysis of the fecal microflora of human subjects consuming a probiotic product
A. DE MORENO ET AL.

108 109Eur. J. Inflamm.
containing Lactobacillus rhamnosus DR20. Appl Environ Microbiol 2000; 66:2578-88.
18. Videla S, Vilaseca J, Guarner F, Salas A, Treserra F, Crespo E, Antolin M, Malagelada JR. Role of intestinal microflora in chronic inflammation and ulceration of the rat colon. Gut 1994; 35:1090-7.
19. Favier C, Neut C, Mizon C, Cortot A, Colombel JF, Mizon J. Fecal beta-D-galactosidase production and Bifidobacteria are decreased in Crohn’s disease. Dig Dis Sci 1997; 42:817-22.
20. Delgado S, O’Sullivan E, Fitzgerald G, Mayo B. Subtractive screening for probiotic properties of lactobacillus species from the human gastrointestinal tract in the search for new probiotics. J Food Sci 2007; 72:M310-5.
21. Madsen K, Cornish A, Soper P, McKaigney C, Jijon H, Yachimec C, Doyle J, Jewell L, De Simone C. Probiotic bacteria enhance murine and human intestinal epithelial barrier function. Gastroenterology 2001; 121:580-91.
22. de Moreno de LeBlanc A, Valdez J, Perdigón G. Inflammatory immune response. Eur J Inflamm 2004; 2:21-31.
23. Lamm ME. Current concepts in mucosal immunity. IV. How epithelial transport of IgA antibodies relates to host defense. Am J Physiol 1998; 274:G614-7.
24. Malin M, Suomalainen H, Saxelin M, Isolauri E. Promotion of IgA immune response in patients
with Crohn’s disease by oral bacteriotherapy with Lactobacillus GG. Ann Nutr Metab 1996; 40:137-45.
25. Fuss IJ, Neurath M, Boirivant M, Klein JS, de la Motte C, Strong SA, Fiocchi C, Strober W. Disparate CD4+ lamina propria (LP) lymphokine secretion profiles in inflammatory bowel disease. Crohn’s disease LP cells manifest increased secretion of IFN-gamma, whereas ulcerative colitis LP cells manifest increased secretion of IL-5. J Immunol 1996; 157:1261-70.
26. Mannon PJ, Fuss IJ, Mayer L, et al. Anti-interleukin-12 antibody for active Crohn’s disease. N Engl J Med 2004; 351:2069-79.
27. Dong C. Diversification of T-helper-cell lineages: finding the family root of IL-17-producing cells. Nat Rev Immunol 2006; 6:329-33.
28. Zhang Z, Zheng M, Bindas J, Schwarzenberger P, Kolls JK. Critical role of IL-17 receptor signaling in acute TNBS-induced colitis. Inflamm Bowel Dis 2006; 12:382-8.
29. Sydora BC, Tavernini MM, Wessler A, Jewell LD, Fedorak RN. Lack of interleukin-10 leads to intestinal inflammation, independent of the time at which luminal microbial colonization occurs. Inflamm Bowel Dis 2003; 9:87-97.
30. Singh B, Read S, Asseman C, et al. Control of intestinal inflammation by regulatory T cells. Immunol Rev 2001; 182:190-200.

EUROPEAN JOURNAL OF INFLAMMATION Vol. 7, no. 2, 0-0 (2009)
1721-727X (2009)Copyright © by BIOLIFE, s.a.s.
This publication and/or article is for individual use only and may not be furtherreproduced without written permission from the copyright holder.
Unauthorized reproduction may result in financial and other penalties111

EUROPEAN JOURNAL OF INFLAMMATION Vol. 7, no. 2, 0-0 (2009)
1721-727X (2009)Copyright © by BIOLIFE, s.a.s.
This publication and/or article is for individual use only and may not be furtherreproduced without written permission from the copyright holder.
Unauthorized reproduction may result in financial and other penalties111
The monochromatic excimer light 308 nm (MEL 308 nm) is a UVB radiation generated by a light source emitted by excitation energy decrease of a molecule (excimer) formed by the combination of two atoms: the noble gas xenon and chlorine (1). It has already been used with good results in the treatment of psoriasis, vitiligo and, less frequently, lichen cutaneous (2-5). Oral Lichen planus (OLP) is a cytotoxic autoimmune disease of unknown aetiology. OLP can cause considerable inconvenience to the patient and is often resistant to the various therapeutic options used, tending to become chronic (6-7).
The purpose of this study is to assess the clinical efficacy of monochromatic excimer light in the treatment of symptomatic oral lichen planus (OLP).
MATERIALS AND METHODS
Twenty-two patients were recruited, aged between 34 and 81 years (mean age 63.6 years), from October 2005 to August 2006: 13 women and 9 men, affected by oral lichen planus. The duration of OLP was between 2 and 20 years. Written informed consent was obtained from each subject.
All patients had suspended topical and systemic therapies for OLP for at least 2 months. Participants in this study underwent mucosal biopsy to confirm diagnosis, Dental Materials Series patch-test (F.I.R.M.A, Italy), and blood tests to research antinucleus antibodies (ANA) and markers of hepatitis B and C virus (6, 8-10).
Patients with ANA> 1:40 and pregnant or breastfeeding women were excluded from therapy.
The individual lesions were processed through an
THERAPY OF ORAL LICHEN PLANUS WITH MONOCHROMATIC EXCIMER LIGHT
L. GERMI, M. LAZZARO and C. VELLER-FORNASA
Dermatology Unit, San Bortolo Hospital, Vicenza, Italy
Received November 13, 2008 – Accepted March 30, 2009
Mailing address: Dr Germi Lerica, Unità Operativa Dermatologia,Ospedale San Bortolo,Via Rodolfi 37 ,36100 Vicenza, ItalyTel: ++39 0444 753606 Fax: ++39 0444 753167e-mail: [email protected]
Key words: oral lichen planus, monochromatic excimer light 308 nm, therapy
Oral lichen planus can cause considerable inconvenience to the patient and is often resistant to the various therapeutic options used, tending to become chronic. The monochromatic excimer light 308 nm is UVB radiation generated and has already been used with good results in the treatment of psoriasis, vitiligo and, less frequently, lichen cutaneous. The purpose of this study is to assess the clinical efficacy of monochromatic excimer light in the treatment of symptomatic oral lichen planus. Twenty patients were treated weekly with single phototherapic sessions. The therapy was suspended on achieving a total score of gravity at least 35% or less than the score of departure. At the end of the study scoring on the size of lesions, intensity of white, intensity of erythema, grade of sick oral gingiva, for both gingival and non-gingival lichen was improved. The statistical processing found a highly significant reduction in post-therapy (p< 0002) of all subjective manifestations tested. Side effects were not observed. Our study shows that monochromatic excimer light 308 nm would appear to be an effective and well-tolerated treatment for oral lichen planus and could represent a new additional therapeutic strategy.

112 113Eur. J. Inflamm.
accessory of an MEL 308 nm device (Excilite TM Deka Medical Lasers, Florence, Italy): a narrow fiberoptic handpiece, circular in shape with a diameter of 8 mm, which produces a power density of 48mW/cm2. The patients were treated weekly with a single phototherapic session, lasting 5 seconds, corresponding to an energy dose of 0.25 J/cm2. The therapy was suspended on achieving a total score of gravity at least 35% or less than the score at the beginning.
A number of sessions comprised between 7 and 30 per lesion was needed to achieve this value.
At the beginning and the end of therapy the clinical evaluation of disease severity was made by a clinical index based on various parameters, as described by G. Bethke and P.A. Reichart referring to J.O. Andreasen’s original work published in 1968 (11-12). The clinical index of OLP severity distinguishes OLP in gingival and non-gingival lesions; the scores of severity result from developing two tables, one for gingival lesions and one for non-gingival lesions (Table I).
The evaluation of clinical evolution was made considering the individual lesions and not the patients because of the contemporary presence of lesions with different clinical forms (reticular, plaque, erosive, atrophic) in the gingival and non-gingival localizations in some patients and the possible time difference in achieving clinical response to treatment of various forms of lesions present in the same patient.
At the beginning and the end of therapy the patients also had to fill in a card for self-assessment of pain: Visual Analogue Scale (VAS) of pain as index of subjective symptoms. The VAS of patients’ pain, valued pre- and post-therapy, was required for the worst and weakest experienced pain intensity, on average, at the time of completing the card. The data on pain symptoms were subjected to statistical analysis with the mini tab program and t-Paired test.
RESULTS
Two patients were withdrawn from the study because of non-compliance: they stopped therapy after 2 and 4 treatments. Two patients were hepatitis B positive without signs of active liver disease. No patient was Dental Materials Series patch-test positive.
The total number of treated lesions was 25 out of 20 patients who completed the therapy (12 patients with only non-gingival lesions, 3 with only gingival lesions, 5 with both non-gingival and gingival lesions). In Tables II, III and IV respectively, the clinical aspect of non-gingival and gingival lesions, the
scores before and after treatment, the number of treatments carried out per lesion, and the self-assessment of pain before and after treatment are summarised. At the end of the study the scores on the size of lesions, intensity of white, intensity of erythema, and the grade of sick oral gingiva for both gingival and non-gingival lichen improved (Tables II-IV). The regression of the disease was linked to the clinical form and size of the lesion. On average, the reticular, atrophic forms and plaque healed more easily than the erosive forms and also the symptoms related to them decreased proportionately.
The statistical processing through the t-PAIRED test found a highly significant reduction in post therapy (p< 0002) of all subjective manifestations tested. Side effects such as erythema, pain, appearance of bullous lesions and hyperpigmentation were not observed. The therapy was well- tolerated by all patients. Follow-up visits were made every month after the suspension of excimer light therapy for four months and the disease did not re-emerged during this period.
DISCUSSION
The pathogenesis of OLP is characterized by apoptosis of basal keratinocytes, triggered by contact between CD8+-activated lymphocytes and an unknown antigen expressed on the surface of the basal cells. Basement membrane degradation, which allows lymphocytes to migrate, involves proteolytic enzymes called matrix metalloproteinases (MMPs) (13).
Our study shows that MEL 308 nm would appear to be an effective and well-tolerated treatment for oral lichen planus and could represent a new additional therapeutic strategy. Its effectiveness is likely linked to immunosuppressive activity at mucous-cutaneous levels, inhibiting or destroying the cutaneous lymphocytic infiltrate by apoptosis induction. It has been demonstrated that early MEL results in a reduction of T-lymphocytes infiltrating the skin, with a drastic decrease in inflammatory cytokine expression. On one hand, it leads to a decreased expression of antiapoptotic protein Bcl-2, while on the other it induces an increase of apoptotic mediator p53 (2- 5,14-15).
It is also interesting to note that the erosive OLP appears to be responsive to MEL, though
L. GERMI ET AL.

112 113Eur. J. Inflamm.
more slowly than other clinical forms of OLP, and this is particularly promising because conventional therapies are usually inadequate for the erosive forms (16).
We believe that the Betchke-Reichart’s Index is the best method for the clinical evaluation of OLP and of treatment results, because this index, compared to other previous indices, presents: separate evaluation of gingival and non-gingival OLP; determination of grade of illness on the basis of subjective and objective parameters, high sensitivity to small differences with different grades of severity
assignable to prefixed parameters (e.g. size of lesions, intensity of white colour and erythema); possibility to assess the results of therapy also in patients who have not had a complete remission; independence between the valuation made by the doctor and by the patient (11, 17-21).
On the other hand, for the evaluation of subjective symptoms, VAS scale (visual analog scale) is considered an ideal scale in the literature, the best method of judgement and universally accepted, because it is more independent, because of the language used, compared to verbal scales (22).
1
Table I. Summary and values of all indices.
Score of indices Indices of localization Grade 0
zeroGrade 1 light
Grade 2 middle
Grade 3 severe
Index of size (Basic Index) Score
Reticular/papular form 0 1 2 3 Plaque/ atrophic form 0 1.5 3 4.5
Erosive and bullous form 0 2 4 6
Supplementary indices
Index of intensity of whitish pattern 0 0.5 1 1.5 Index of erythema (mucosal erythema for gingival form) 0 0.5 1 1.5
Index of oral gingiva (for gingival form) 0 1 - -
Table I. Summary and values of all indices.
2
Table II. Clinical aspect of non-gingival lesions and scores on the assessment on the basis of G. Bethke and P.A. Reichart�s criteria at the beginning and the end of therapy. In the last column the number of treatment sessions/lesion.
Non-gingival lesions
Index of size
Pre Post
Score
Pre Post
Index of white
intensity
Pre Post
Score
Pre Post
Index of erythema intensity
Pre Post
Score
Pre Post
Total
Pre Post
N° sessions
Plaque Gr 1 Gr 0 1.5 0 Gr 3 Gr 0 1.5 0 Gr 0 Gr 0 0 0 3 0 30Erosion Gr 3 Gr 1 6 2 Gr 2 Gr 1 1 0.5 Gr 3 Gr 1 1.5 0.5 8.5 3 22Atrophy Gr 3 Gr 1 4.5 1.5 Gr 2 Gr 1 1 0.5 Gr1 Gr 0 0.5 0 6 2 12Erosion Gr 3 Gr 1 6 2 Gr 2 Gr 0 1 0 Gr 3 Gr 1 1.5 0.5 8.5 2.5 22Erosion Gr 3 Gr 1 6 2 Gr 3 Gr 1 1.5 0.5 Gr 2 Gr 0 1 0 8.5 2.5 12Atrophy Gr 3 Gr 1 4.5 1.5 Gr 2 Gr 0 1 0 Gr 2 Gr 1 1 0.5 6.5 2 15Plaque Gr 1 Gr 0 1.5 0 Gr 1 Gr 0 0.5 0 Gr 1 Gr 0 0.5 0 2.5 0 7Atrophy Gr 2 Gr 0 3 0 Gr 3 Gr 0 1.5 0 Gr 2 Gr 0 1 0 5.5 0 15Erosion Gr 3 Gr 1 6 2 Gr 2 Gr 1 1 0.5 Gr 3 Gr 1 1.5 0.5 8.5 3 12Erosion Gr 2 Gr 0 4 0 Gr 2 Gr 0 1 0 Gr 3 Gr 0 1.5 0 6.5 0 15Erosion Gr 3 Gr 1 6 2 Gr 3 Gr 1 1.5 0.5 Gr 3 Gr 1 1.5 0.5 9 3 26Erosion Gr 3 Gr 1 6 2 Gr 2 Gr 1 1 0.5 Gr 1 Gr 0 0.5 0 7.5 2.5 7Erosion Gr 3 Gr 1 6 2 Gr 3 Gr 1 1.5 0.5 Gr 2 Gr 0 1 0 8.5 2.5 30Erosion Gr 2 Gr 0 4 0 Gr 3 Gr 0 1.5 0 Gr 1 Gr 0 0.5 0 6 0 7Atrophy Gr 3 Gr 1 4.5 1.5 Gr 2 Gr 1 1 0.5 Gr 1 Gr 0 0.5 0 6 2 15Erosion Gr 3 Gr 1 6 2 Gr 2 Gr 1 1 0.5 Gr 2 Gr 0 1 0 8 2.5 22Erosion Gr 3 Gr 1 6 2 Gr 2 Gr 0 1 0 Gr 2 Gr 1 1 0.5 8 2.5 15
Table II. Clinical aspect of non-gingival lesions and scores on the assessment on the basis of G. Bethke and P.A. Reichart’s criteria at the beginning and the end of therapy. In the last column the number of treatment sessions/lesion.

114 115Eur. J. Inflamm.
3
3
Table III. Clinical aspect of gingival lesions and scores on the assessment on the basis of G. Bethke and P.A. Reichart�s criteria at the beginning and the end of therapy. In the last column the number of treatment sessions/lesion.
Gingival lesions
Index of size
Pre Post
Score
Pre Post
Index of white
intensity
Pre Post
Score
Pre Post
Index of erythema intensity
Pre Post
Score
Pre Post
Index of oral
gingiva
Pre Post
Score
Pre Post
Total
Pre Post
N°sessions
Reticular Gr 1 Gr 0 1 0 Gr 2 Gr 0 1 0 Gr 0 Gr 0 0 0 Gr 0 Gr 0 0 0 2 0 7Reticular Gr 2 Gr 0 2 0 Gr 2 Gr 0 1 0 Gr 1 Gr 0 0.5 0 Gr 1 Gr 0 1 0 4.5 0 15Atrophy Gr 3 Gr 1 4.5 1.5 Gr 1 Gr 0 0.5 0 Gr 2 Gr 1 1 0.5 Gr 1 Gr 0 1 0 7 2 26Atrophy Gr 2 Gr 1 3 1.5 Gr 2 Gr 0 1 0 Gr 2 Gr 1 1 0.5 Gr 1 Gr 0 1 0 6 2 15Plaque Gr 1 Gr 0 1.5 0 Gr 2 Gr 0 1 0 Gr 2 Gr 0 1 0 Gr 0 Gr 0 0 0 3.5 0 15Reticular Gr 2 Gr 1 2 1 Gr 3 Gr 1 1.5 0.5 Gr 2 Gr 0 1 0 Gr 0 Gr 0 0 0 4.5 1.5 7Erosion Gr 3 Gr 1 6 2 Gr 2 Gr 1 1 0.5 Gr 3 Gr 1 1.5 0.5 Gr 1 Gr 0 1 0 9.5 3 15Reticular Gr 1 Gr 0 1 0 Gr 1 Gr 0 0.5 0 Gr 0 Gr 0 0 0 Gr 0 Gr 0 0 0 1.5 0 7
Table III. Clinical aspect of gingival lesions and scores on the assessment on the basis of G. Bethke and P.A. Reichart’s criteria at the beginning and the end of therapy. In the last column the number of treatment sessions/lesion.
4
4
Table IV. Data on the perception of pain in its various expressions found in patients using self-assessment (VAS Pain) at the beginning and at the time of suspension of therapy.
Pain self-assessment card (VAS - pain) at first visit
Patients Pain intesity Pain intesity Pain intesity Pain intesity Total N° stronger form
Pre Post
weaker form
Pre Post
average
Pre Post
moment of the visit
Pre Post
Score
Pre Post 1 3 1 0 0 1 0 1 0 5 1 2 9 3 7 1 6 1 7 1 29 6 3 8 3 4 1 5 1 7 1 24 6 4 7 3 4 1 5 1 6 0 22 5 5 8 4 4 1 5 1 7 2 24 8 6 8 3 4 1 5 1 7 1 24 6 7 4 1 1 0 2 1 3 0 10 2 8 6 2 2 0 3 1 4 0 15 3 9 9 5 7 2 6 3 8 2 30 12
10 8 3 4 1 6 2 8 2 26 8 11 10 6 8 4 7 5 10 4 35 19 12 10 6 7 2 6 3 10 6 33 17 13 8 3 6 0 6 1 8 0 28 4 14 6 0 1 0 3 0 5 0 15 0 15 5 1 1 1 2 1 4 0 12 3 16 6 3 2 1 3 1 4 1 15 6 17 6 1 2 1 3 1 5 0 16 3 18 9 4 7 2 6 2 8 3 30 11 19 4 1 1 1 2 1 3 0 10 3 20 8 4 4 1 5 2 7 1 24 8
Table IV. Data on the perception of pain in its various expressions found in patients using self-assessment (VAS Pain) at the beginning and at the time of suspension of therapy.
L. GERMI ET AL.

114 115Eur. J. Inflamm.
Dermatol Venereol 2006; 20:523-6.6. Thornhill MH, Pemberton MN, Simmons RK,
Theaker ED. Amalgam-contact hypersensitivity lesions and oral lichen planus. Oral Surg Oral Med Oral Pathol Oral Radiol Endod 2003; 95:291-9.
7. Trehan M, Taylor CR. Low-dose excimer 308-nm laser for the treatment of oral lichen planus. Arch Dermatol 2004; 140:415-20.
8. Koch P. Oral Lichenoid Lesions: induction by exogenous factors? Dermatosen 1998; 46:196.
9. Eisen D, Carrozzo M, Bagan JV, Thongprasom S. Oral lichen planus: clinical features and management. Oral Diseases 2005; 11:338-49.
10. Deris E, Serez K. The prevalence of dermatologic manifestations related to chronic Hepatitis C virus infection in a study from a single centre in Turkey. Acta Dermatovenerol Alp Panonica Adriat 2005; 14:93-8.
11. Bethke G, Reichart PA. Assessment of severity of oral lichen planus using a new clinical index. Mund Kiefer Gesichtschir 2005; 9:152-60.
12. Andreasen JO. Oral lichen planus: a clinical evaluation of 115 cases. Oral Surg Oral Med Oral Pathol 1968; 25:31-42.
13. Mazzarella N, Femino F, Gombos F, De Rosa A, Giuliano M. Matrix metalloproteinase gene expression in oral lichen planus: Erosive vs Reticular Forms. J Eur Acad Dermatol Venereol 2006; 20:953-57.
14. Bianchi B, Campolmi P, Mavilia L, Danesi A, Rossi R, Cappugi P. Monochromatic excimer light (308 nm): an immunohistochemical study of cutaneous T cell and apoptosis-related molecules in psoriasis. J Eur Acad Dermatol Venereol 2003; 17:408-13.
15. Bascones-Ilundain C, Gonzalez-Moles MA, Esparza-Gómez G, Gil-Montoya JA, Bascones-Martínez A. Importance of apoptotic mechanisms in inflammatory infiltrate of oral lichen planus lesions. Anticancer Res 2006; 26:357-62.
16. Trehan M, Taylor CR. Low-dose excimer 308-nm laser for the treatment of oral lichen planus. Arch Dermatol 2004; 140:415-20.
17. Olivier V, Lacour JP, Mousnier A, Garraffo R, Monteil RA, Ortonne JP. Treatment of chronic erosive oral lichen planus with low concentrations of topical tacrolimus. An open prospective study. Arch Dermatol 2002; 138:1335-38.
In our study, the VAS scale was used to quantify the critical moments of suffering due to the disease, showing that patients indicated a good comfort after this therapy. Furthermore, it is interesting to note how the VAS scale corresponded to lower values that tended to zero in cases of clinical remission, giving a highly significant value to the statistical analysis.
Although we did not compare our patients with a control group, the data described in our study and valuated with Betchke-Reichart’s index and VAS scale seem to be encouraging and indicates that the monochromatic excimer light, with its immunosuppressive action at cutaneous levels, can be considered with due right among the therapies of choice of OLP as a new, safe, well-tolerated and very promising strategy. In fact, being focused on the lesions, it saves the healthy mucosa from unnecessary and/or harmful applications; it does not involve taking drugs; normally consists of a limited number of treatments executed for a few seconds, once a week; it improves the quality life of the patient from the earliest applications.
The optimal dose of 308 nm, the frequency of treatment and possible combination with other therapies should be established in future studies.
REFERENCES
1. Brazzini B, Mavilia L, Hercogova J, Lotti TM. Excimer light: pearls from around the world Int J Immunopathol Pharmacol 2002; 13:41-4.
2. Campolmi P, Mavilia L, Lotti T M, Rossi R, Cappugi P. 308 nm monochromatic excimer light for the treatment of palmoplantar psoriasis. Int J Immunopathol Pharmacol 2002; 13:32-4.
3. Cappugi P, Mavilia L, Mavilia C, Brazzini B, Rossi R, Hercogova J, Lotti TM, Campolmi P, Andreassi L. 308 nm monochromatic excimer light in psoriasis: clinical evaluation and study of cytokine levels in the skin. Int J Immunopathol Pharmacol 2002; 13:14-19.
4. Hofer A, Hassan AS, Kerl FJ, Wolf P. The efficacy of excimer laser (308 nm) for vitiligo at different body sites. J Eur Acad Dermatol Venereol 2006; 20:558-64.
5. Nistico SP, Saraceno R, Stefanescu S, Chimenti S. A 308-nm monochromatic excimer light in the treatment of palmoplantar psoriasis. J Eur Acad

116 EUROPEAN JOURNAL OF INFLAMMATION Vol. 7, no. 2, 0-0 (2009)
1721-727X (2009)Copyright © by BIOLIFE, s.a.s.
This publication and/or article is for individual use only and may not be furtherreproduced without written permission from the copyright holder.
Unauthorized reproduction may result in financial and other penalties117
Mayer P, Watson JJ. Oral lichen planus patient profile, disease progression and treatment responses. J Am Dent Assoc 2001; 132:901-09.
21. Rode M, Kogoj-Rode M. Malignant potential of the reticular form of oral lichen planus over a 25-year observations period in 55 patients from Slovenia. J Oral Sci 2002; 44:109-11.
22. Price DD, Bush FM, Long S, Harkins SW. A comparison of pain measurement characteristics of mechanical visual analogue and simple numerical rating scale. Pain 1994; 56:217-26.
18. Thongprasom K, Luengvisut P, Wongwatanakij A, Boonjatturus C. Clinical evaluation in treatment of oral lichen planus with topical Fluocinolone acetonide: a 2-year follow-up. J Oral Pathol Med 2003; 32:315-22.
19. Rodstrom PO, Hakeberg M, Jontell M, Nordin P. Erosive oral lichen planus treated with clobetasol propionate and triamcinolone acetonide in Orabase: a double-blind clinical trial. J Dermatol Treat 1994; 5:7-10.
20. Chainani-Wu N, Silverman SJr, Lozada-Nur F,
L. GERMI ET AL.

116 EUROPEAN JOURNAL OF INFLAMMATION Vol. 7, no. 2, 0-0 (2009)
1721-727X (2009)Copyright © by BIOLIFE, s.a.s.
This publication and/or article is for individual use only and may not be furtherreproduced without written permission from the copyright holder.
Unauthorized reproduction may result in financial and other penalties117
Behçet’s disease (BD) is a chronic, relapsing inflammatory syndrome that involves multiple organs and shows four primary features: recurrent aphthous ulcers of oral mucosa, skin lesions such as erythema nodosum and pseudofolliculitis, uveitis and genital ulcers. There are also articular, gastrointestinal, vascular and central nervous system manifestations. One of the most important disorders connected with BD is uveoretinitis that can lead to blindness. Treatment of BD is aimed at modifying the inflammatory response, especially the hyperactivity of neutrophils and T cell functions (1).
Treatment usually includes corticosteroids (up to 1 mg/kg/day), azathioprine, or ciclosporin. Other agents may include colchicine, mycophenolate mofetil, cyclophosphamide and chlorambucil. Therapeutic administration of anti-TNF-α blockers
has been reported in patients with BD refractory to conventional immunosuppressive drugs (4-15).
Clinical caseWe report the case of a 54-year-old female
patient with a long history (since 1997) of painful, periodically recurring oral and genital ulcers and recurrent phlebitis and thrombosis. She was diagnosed for BD in 2000 after an episode of optic nerve ischemia of the right eye. HLAB51 was performed and tested positive. At that time, besides heparin, colchicine and low dose aspirin, the immunosuppressive regimen consisted of prednisone 25 mg/daily, azathioprine 200 mg/daily, ciclosporin 200 mg/daily, and hydroxychloroquine 200 mg/daily. Despite such therapy she had persistent oral and genital ulcers, erythema nodosum and asimmetrical
LETTER TO THE EDITOR
BEHÇET’S DISEASE AND ETANERCEPT: EIGHTY WEEKS OF EXPERIENCE
B. KROEGLER, G. DI MUZIO, F. GIOVANNANGELI, M. DE MATTIA, C. PERRICONE and R. PERRICONE
University of Rome Tor Vergata, Unit of Rheumatology, Department of Internal Medicine,Rome, Italy
Mailing address: Prof. Roberto Perricone, M.D., Chief of Rheumatology, University of Rome Tor Vergata, Department of Internal Medicine, Via Montpellier 1, 00133 Rome, Italy Tel/Fax: ++39 06 72596287 e-mail: [email protected]
Key words: Behçet’s disease, anti-TNF, Etanercept, Enbrel
Behçet’s disease (BD) is a chronic, relapsing inflammatory disease that involves multiple organs. Besides standard therapy, some patients with severe, non-responding disease experienced therapeutic blockage of the activity of TNF with encouraging results. We report the case of a patient affected with BD refractory to ciclosporin, azathioprine and corticosteroid therapy with orogenital ulcerations, optic nerve ischemia, arthritis and erythema nodosum. The patient was treated with etanercept, a TNF-α blocker, 25 mg twice a week. From the second administration of etanercept, the patient showed an improvement in oral and genital ulcers, of erythema nodosum, and in visual acuity. ESR and CRP also showed a dramatic reduction returning to normal, and after eight weeks immunosuppressive therapy could be discontinued because of complete remission of oral-genital ulcers and of erythema nodosum. From week 24 to the last control carried out at week 80, the patient showed clinical and laboratory disease remission. Etanercept long-term therapy was demonstrated to be safe and effective in our patient with refractory BD, and allowed to spare steroids and other immunosuppressive drugs, thus preventing the related side-effects.

118 119Eur. J. Inflamm.
arthritis.In July 2006, laboratory examinations showed
high C-reactive protein (CRP, 13.2 mg/l; normal value 0-0.5 mg/l) and erythrocyte sedimentation rate (ESR, 46 mm/h; normal value 0-20 mm/h). Anti-nuclear antibodies (ANA) were positive (1:160, speckled). Anti-extractable nuclear antigen, anti-double strand-DNA, anticardiolipin IgG and IgM, anti-β2GPI IgG and IgM and LAC were also performed and tested negative. Echocardiography showed mitral and tricuspid valve prolapse. All the assays were repeated during the course of the therapy at weeks 12, 24, 36, 54 and 80. Autoantibodies (except ANA) always tested negative during the follow-up.
Since laboratory and clinical features did not ameliorate, despite the above-mentioned treatment, we decided to start anti-TNF therapy with etanercept (Enbrel, Wyeth Lederle S.p.A, Italy), at the dosage of 25 mg twice a week. Concomitant treatments were prednisone 7.5 mg/daily, azathioprine 150 mg/daily and ciclosporin 200 mg/daily that were discontinued after two months of follow-up.
From the second administration of etanercept, the patient showed an improvement of oral and genital ulcers, of erythema nodosum, and of visual acuity. ESR and CRP also showed a dramatic reduction, returning to normal as reported in Table I. After eight weeks, immunosuppressive therapy could be discontinued as there was complete remission of oral-genital ulcers and of erythema nodosum. After 24 weeks the patient maintained the remission of mucosal and genital ulcers and of erythema
nodosum and showed also the remission of other BD manifestations such as arthritis. At the last control performed at week 80, the patient still showed clinical and laboratory disease remission.
DISCUSSION
TNF-α is a potent pro-inflammatory cytokine whose importance in the pathogenesis of several inflammatory disorders, including BD, has been widely demonstrated. Levels of circulating TNF-α and soluble TNF receptor are increased in the peripheral blood of patients with active BD, and T-cell-mediated immune responses are thought to play a major role in the pathogenesis of BD (2-3). Recent findings suggest that the TNF-α blocker infliximab is beneficial in patients with BD with mucocutaneous (4-5), ocular (6-7, 11-12), central nervous system (13) and gastrointestinal involvement (8-9).
Melikoglu et al. found in a double blind placebo controlled study that etanercept was effective in suppressing most of the mucocutaneous and arthritic manifestations of BD, and Scheinberg reported the potential beneficial use of etanercept in two patients with severe recurrent aphtous ulcers (14-15). However, long-term follow-up studies have not yet been reported.
In this study, we show that etanercept was beneficial both at short and long term in a patient affected with BD. Indeed, we observed a dramatic and prompt response after the second etanercept administration, which was still evident after 80 weeks of follow-up, and remained positive during
Time (weeks) ANA C3 C4 CH50 ESR CRP
Injection → 0 1:160 speckled 146 27 72 46 13.4
12 1:320 speckled 119 19 121 28 2.4
24 1:160 speckled 139 25 116 7 0.8
36 1:80 speckled 155 32 136 24 0.6
54 1:80 speckled 130 22.4 87 16 1.180 1:80 speckled 140 21.4 149 17 2.1
Table I. Laboratory findings during 80 weeks of Etanercept treatment.
B. KROEGLER ET AL.

118 119Eur. J. Inflamm.
the course of the disease. The presence of ANA in BD patients is not a frequent event, and infliximab treatment was reported to be associated with an increase of the titres of such antibodies (16). In some other cohorts of patients no ANA induction was reported even after one year of etanercept treatment (17). Since in our patient ANA were positive (1:160) at baseline, etanercept was chosen, and from 36 to 80 weeks ANA titres were even decreased (1:80).
Several mechanisms may underlie the rapid and persistent clinical response that was observed, such as an efficient and persistent neutralization of TNF-α at the dermal level, probably due to the binding properties of etanercept to TNF-α (18), and/or a decrease in the levels of proinflammatory cytokine secretion by T cells (19) avoiding a proinflammatory cytochine-driven immune response (20). Whatever the mechanism, the rapid therapeutic effect is important in BD in order to prevent ocular and neurological complications. Moreover, the safety and efficacy of etanercept, when given in a long-term administration, may encourage the use of this drug in chronic BD. This therapeutic approach may be useful not only for treating and preventing relapses in patients with BD, but also for allowing to spare steroids and other immunosuppressive drugs, thus avoiding all the related side effects.
Although we realize that this successful outcome in one single patient does not provide the ultimate proof of efficacy of this treatment in the disease, we feel that the results are rather encouraging.
REFERENCES
1. Yamashita N. Hyperreactivity of neutrophils and abnormal T-cell Homeostasis: a new insight for pathogenesis of Behçet’s disease. Int Rev Immunol 1997; 14: 11-20.
2. Turan B, Gallati H, Erdi H, et al. Systemic levels of the T cell regulatory cytokines IL-10 and IL-12 in Behçet’s disease: soluble TNFR-75 as a biological marker of disease activity. J Rheumatol 1997; 24:128-32.
3. Evereklioglu C, Er H, Turkoz Y, et al. Serum levels of TNF-α, sIL-2R, IL-6 and IL-8 are increased and associated with elevated lipid peroxidation in patients with Behçet’s disease. Mediators Inflamm 2002; 11:87-93.
4. Robertson LP, Hickling P. Treatment of recalcitrant orogenital ulceration of Behcet’s syndrome with infliximab. Rheumatology Oxford 2001; 40:473-4.
5. Goossens PH, Verburg RJ, Breedveld FC. Remission of Behcet’s syndrome with tumor necrosis factor alpha blocking therapy. Ann Rheum Dis 2001; 60:637.
6. Sfikakis PP, Theodossiadis PG, Katsiari CG, Kaklamanis P, Markomichelakis NN. Effect of infliximab on sight-threatening panuveitis in Behcet’s disease. Lancet 2001; 358:295-6.
7. Triolo G, Vadala M, Accardo-Palumbo A, et al. Anti-tumor necrosis factor monoclonal antibody treatment for ocular Behcet’s disease. Ann Rheum Dis 2002; 61:560-1.
8. Travis SP, Czajkowski M, McGovern DP, Watson RG, Bell AL. Treatment of intestinal Behcet’s syndrome with chimeric tumor necrosis factor alpha antibody. Gut 2001; 49:725-8.
9. Hassard PV, Binder SW, Nelson V, Vasiliauskas EA. Anti-tumor necrosis factor monoclonal antibody therapy for gastrointestinal Behcet’s disease: a case report. Gastroenterology 2001; 120:995-9.
10. Yamashita N, Kaneoka H, Kaneko S, et al. Role of the γδ T lymphocytes in the development of Behçet’s disease. Clin Exp Immunol 1997; 107:241-7.
11. Lindstedt EW, Baarsma GS, Kuijpers RW, van Hagen PM. Anti-TNF-alpha therapy for sight threatening uveitis. Br J Ophthalmol 2005; 89:533-6.
12. Ohno S, Nakamura S, Hori S, et al. Efficacy, safety, and pharmacokinetics of multiple administration of infliximab in Behçet disease with refractory uveoretinitis. J Rheumatol 2004; 31:1362-8.
13. Sarwar H, McGrath Jr H, Espinoza LR. Successful treatment of long-standing Neuro-Behçet’s disease with infliximab. J Rheumatol 2005; 32:181-3.
14. Melikoglu M, Fresko I, Mat C, et al. Short-term trial of Etanercept in Behçet’s disease: a double blind, placebo controlled study. J Rhematol 2005; 32:98-105.
15. Scheinberg MA. Treatment of recurrent oral aphtous ulcers with Etanercept. Clin Exp Rheumatol 2002; 20:733-4.
16. Elezoglou A, Kafasi N, Kaklamanis PH, Theodossiadis PG, Kapsimali V, Choremi E, Vaiopoulos G, Sfikakis PP. Infliximab treatment-induced formation of

120
2002; 301:418-26.19. Zou JX, Braun J, Sieper J. Immunological basis for
the use of TNF alpha-blocking agents in ankylosing spondylitis and immunological changes during treatment. Clin Exp Rheumatol 2002; 20:S34-7.
20. Chapman PT, Yarwood H, Harrison AA, et al. Endothelial activation in monosodium urate monohydrate crystal-induced inflammation: in vitro and in vivo studies on the role of tumor necrosis factor alpha and interleukin 1. Arthritis Rheum 1997; 128:443-50.
autoantibodies is common in Behçet’s disease. Clin Exp Rheumatol 2007; 25:S65-9.
17. De Rycke L, Baeten D, Kruithof E, Van den Bosch F, Veys EM, De Keyser F. Infliximab, but not etanercept, induces IgM anti-double-stranded DNA autoantibodies as main antinuclear reactivity: biologic and clinical implications in autoimmune arthritis. Arthritis Rheum 2005; 52:2192-201.
18. Scallon B, Cai A, Solowski N, et al. Binding and functional comparisons of two types of tumor necrosis factor antagonists. J Pharmacol Exp Ther
B. KROEGLER ET AL.