generalizing from experiments: is predation strong or weak in the new england rocky intertidal?
TRANSCRIPT

Oecologia (1991) 88:1 8 Oecologia �9 Springer-Verlag 1991
Original papers
Generalizing from experiments: is predation strong or weak in the New England rocky intertidal? Bruce A. Menge
Department of Zoology, Oregon State University, Corvallis, OR, 97331-2914, USA
Received November 20, 1990 / Accepted in revised form May 10, 1991
Summary. Petraitis (1990) recently critized previous generalizations regarding the effects of predation in the New England rocky intertidal region (e.g., Menge 1976; Lubchenco and Menge 1978). Contrary to Lubchenco's and my conclusions, Petraitis concluded that (1) barna- cles and not musseis are the favored prey of dogwhelks and (2) barnacles and not dogwhelks control mussel abundances in the mid and low rocky intertidal region. I provide evidence that these criticisms are unwarranted. First, Lubchenco and I never claimed that diet com- position reflected prey preference. Moreover, predators can influence prey abundance without preferring the prey. Hence, claims regarding preferences have no bear- ing on our conclusions. Second, Petraitis' experiments do not invalidate Lubchenco's and my experimental results. Reanalyses of our experimental data support the earlier conclusion that at wave-sheltered sites, whelks reduce the abundance of mussels independently of barnacle abun- dances. Further, at all but one of Lubchenco's and my study sites, predator densities were higher than at Petrait- is' site. Thus, the absence of a predator effect in Petraitis's study was most likely due to low predator density rather than a lack of generality of our earlier results. This re- evaluation therefore suggests that within a broader con- ceptual framework, Petraitis' apparently divergent re- sults are actually consistent with ours.
Key words: Community regulation - Experiments - New England - Predation - Rocky intertidal
The ultimate goal in ecology is to discover general truths about how populations, communities, and ecosystems are regulated. Although in the 1960's there was a wave of enthusiasm for something approaching a universal generalization (i.e., competition regulated populations and communities), subsequent developments made it clear that whatever generalizations are possible will be
Off'print requests to: B.A. Menge
contingent on specific conditions. The primary develop- ment that forced us into a mode of seeking "contingent" rather than "'universal" generalizations was the growing body of evidence that factors such as predation and disturbance could regulate populations or communities, and that competition did not always occur where expect- ed, or have the effects predicted by theory. Such dis- coveries were based largely on field experimentation, which has gained wide acceptance as the most powerful tool available to ecologists in identifying the processes which regulate populations and communities.
With the emphasis on experimentation, however, there arose a new dilemma in the search for generaliza- tion. As argued by Diamond (1986), experiments are often limited in spatial and temporal extent, replication is sometimes difficult and often low, and habitat heterogeneity can make matching of sites questionable. In the face of such difficulties, how is field experiment- based generalization possible?
Although these and related problems are undeniably important, inventive approaches and increasing sophis- tication in knowledge of statistics and experimental de- sign have spurred further increases in the use of experi- mentation in ecology. Novel approaches have permitted the development of greater power in analysis of large- scale but unreplicated experiments (e.g., Carpenter 1990). Another approach that has been advocated in the investigation of community regulation over large spatial scales is what might be called the "comparative experi- mental" approach (e.g., Menge and Sutherland 1976, 1987). This involves the performance of replicated field experiments at different sites along environmental gra- dients. Use of identical experimental designs at each site provides insight into the site-specific operation of such processes as predation and competition, while com- parisons among sites provides insight into how the strengths of these processes are modified by environ- mental conditions (e.g., Dayton 1971 ; Menge 1976; Lub- chenco and Menge 1978; Lubchenco 1980, 1986; Louda 1982). Along the rocky coast of New England, for in- stance, this approach was used as the basis for the

generalization that control of the abundances of barna- cles and mussels by whelk predation could vary with the degree of wave exposure (Menge 1976). Field experi- ments indicated that, in general, prey abundances were controlled by whelks at sites of relatively low, but not at sites of relatively high wave force. Within this fairly broad generalization, however, the experiments also showed some local variation in details. Algal canopies both suppressed prey abundance through a whiplash effect and enhanced predation rates by providing shelter, increases in mussel abundance were slower on vertical than on horizontal substrata, mussel recruitment was enhanced by the prior presence of barnacles or sub- stratum rugosity, and prey recruitment varied among years, sites, and with latitude (Menge 1976, 1978a, b, 1991).
The general issue addressed here is the validity of this "comparative experimental" approach. Sutherland and I used these and similar results in other habitats to sug- gest a general model of community structure (Menge and Sutherland 1976). We proposed that trophic structure complexity and the importance of predation decreased, and the importance of competition increased with in- creased environmental stress. The model has withstood the test of time largely intact (Sih et al. 1985; Schoener 1989; Menge and Farrell 1989), although it has been both extended and made more contingent (Menge and Sutherland 1987; Menge and Olson 1990). Recently, however, Petraitis (1990) questioned some aspects of the original basis of this model. Here, I address his criticisms, and the broader questions they raise regarding the role of empirical knowledge in developing generalizations in ecology.
Point: a summary of Petraitis' claims
Petraitis summarized my work (and some of that of Jane Lubchenco) as follows: "Menge and Lubchenco con- cluded that the occurrence of macroalgae, barnacles and mussels depended on the ability of the dogwhelk [Nucella lapillus] to control mussels. They showed that the dogwhelk preys primarily on the blue mussel Mytilus edulis and that mussels can overgrow and smother barna- cles and several species of common macroalgae..." "Menge... argued that wave surge hampered the preda- tory activities of the dogwhelk. Thus macroalgae occur on protected shores because the dogwhelks destroy near- ly all mussels, which are a competitive dominant. Mus- sels escape predation on wave swept shores and suppress macroalgae, which cannot successfully compete for space against mussels." (p. 405, Petraitis 1990).
Petraitis then states that "there are two flaws with this generalization". The first is that mussels are common in some protected bays "where dogwhelks are often found (Petraitis 1987)". The second is that we failed to control "for the effects of barnacles". By this he meant that since mussel recruitment is enhanced by prior occupancy of space by barnacles (and rugose rock surfaces: Menge 1976; Seed 1976), mussels may occur in a habitat because of the presence of a barnacle substratum, not because of the absence of Nucella (Petraitis 1990).
Petraitis then presents the results of two experiments in which these supposed flaws were addressed. The study site was in the low zone of a protected cove (Mackerel Cove; hereafter MC) on Swans Island, Maine, where mussels have covered 20-38% of the shore since 1983 (Petraitis 1987, 1990). Experiments included or excluded Nucella in cages which also included a barnacle-covered rock, and included either of two experimental surfaces; a fiberglass resin casting of barnacles, or a smooth, bare granite rock (1985 and 1987 experiments) or a resin "rock" (1987 experiment). The two experiments were initiated in late May, 1985 and 1987, respectively, and were terminated in early-mid August of the same year. Number of mussels settling on the surface was the depen- dent variable of interest. Analysis indicated that Nucella had no effect on mussel recruitment, but that the type of surface had statistically significant effects in both experi- ments. Petraitis concluded (p. 410) that "the results of this [his] study are at odds with previous work..." which claimed that predators, not substratum controlled the distribution of mussels in New England (e.g., Lubchenco and Menge 1978; Menge 1976). Although Petraitis ac- knowledges "that habitat and methodological differences between this study" and ours may be the basis for his different results, he suggests "that this is not the case..." (p. 412, Petraitis 1990).
Petraitis (1990) generalizes from his low zone experi- ments to question the conclusion (Menge 1976) that Nucella is a controlling predator in the mid zone. He argues that mussel abundance is not controlled directly by dogwhelks, but by variation in the availability of suitable substratum. He found higher numbers of mus- sels recruiting to resin casts of Semibalanus balanoides compared to smooth granite rocks in the 1985 experi- ment, and compared to smooth resin rocks in the 1987 experiment.
I also documented enhanced mussel recruitment on rugose surfaces (Menge 1976, Fig. 18). Petraitis argues, however, that the direct interaction between barnacles and mussels is the primary factor affecting mussels in the mid zone at our study areas, and that my earlier sugges- tion (Menge 1976) that Nucella negatively directly affect- ed mussel abundance through predation was erroneous. Further, he questions "the premise that dogwhelks regu- late the density of mussels...", arguing that two assump- tions were made in this premise. These were that Nucella was the only mid zone predator abundant enough to control mussel abundances and that mussels are the preferred prey of dogwhelks (Petraitis 1990, p. 410-411).
Counterpoint: reconsideration of the role of Nucella
Petraitis' claim that I assumed that Nucella was the only mid zone predator abundant enough to control mussel abundances was discussed in an earlier interchange (Ed- wards et al. 1982; Menge 1982), in which the possible role offish and crabs relative to that of whelks in the mid zone was debated. Petraitis cited the Edwards et al. critique but not my response, thus evidently accepting the view that other predators were in fact important in the mid

zone. This may be the case, but, as noted in Menge (1982), this view is not consistent with evidence available at the time regarding the effects of fish and crabs on sessile prey relative to the effects of whelks. Moreover, Petraitis did not work in the mid zone, suggesting that any conclusion he makes from his experiments regarding this particular issue is unwarranted.
Petraitis' claim that we assumed that mussels are the preferred prey of whelks is a misstatement and is evident- ly based on a misinterpretation of our data on field diets (Petraitis, pers. comm.). Consider four points. First, nowhere have we stated or assumed that "mussels are the preferred prey of dogwhelks, and dogwhelks will consis- tently eat mussels regardless of their size and the avail- ability of alternative prey". Nor did we "state that Nucella... prefers... Mytilus... over Semibalanus..." (both quotes from Petraitis 1990, p. 411). Second, although Nucella eats both barnacles and mussels, and sometimes the proportion of mussels in the diet is high (e.g., Table 3, Menge 1976; Table 3, Lubchenco and Menge 1978), diet percentages in the field indicate nothing about preference. Preferred prey are those selected when the predator is given a choice; field diets (or "apparent" diets; Peterson and Bradley 1978; Fairweather and Un- derwood 1983) simply reflect the prey eaten in the field (e.g., Menge 1972). Since the preferred prey may be scarce and thus rarely encountered, however, the field diet may consist primarily of non-preferred prey. For instance, the small predatory seastar Leptasterias hexac~ tis prefers to eat small herbivorous gastropods but the dominant prey in its field diet is the non-preferred barna- cle Balanus 9landula (Menge 1972). The large predatory seastar Pisaster ochraceus prefers mussels (Landenberger 1968; Mauzey et al. 1968 ; Paine 1969) but non-preferred and energetically unattractive barnacles are usually the dominant prey in its field diet (Paine 1980). There may thus be no correspondence between a predator's field diet and its prey preference.
Third, a predator can have significant effects on even non-preferred prey, as long as the predator consumes the prey species in substantial numbers (e,g., Caswell 1978; Hastings 1980). For instance, mussels are preferred and barnacles non-preferred prey of the seastar Pisaster och~ raceus (see above), yet Pisaster can control barnacle abundance (Dayton 1971). In New England, field ob- servations (Menge 1976; Lubchenco and Menge 1978) indicated that mussels and barnacles were both major items in the diet of dogwhelks. Although whelks eat barnacles faster than mussels (data from Connell 1961; Menge 1983; and others cited in Petraitis 1990), direct observation in field experiments indicated that both prey were regularly consumed in treatments with predators present.
Fourth, in some predator exclusions in experiments in the mid zone and in most of those in the low zone, mussels became abundant despite a low cover of barna- cles (e.g., Figs. 3, 11 in Lubchenco and Menge 1978). If barnacles controlled mussels, mussels would not have increased in these cases. Hence, whether or not whelks prefer mussels is largely irrelevant to the question of whether whelks control mussel abundance.
Why the difference?
Experimental Design and Execution
Petraitis' primary claim is that by failing to control for barnacle abundance in Lubchenco's and my experiments, we mistook a barnacle effect on mussels for a predation effect on mussels. Thus, Petraitis postulates that lower abundance of mussels in the presence than in the absence of Nucella (e.g., Fig. 1) was due to the higher cover of Semibalanus in the absence than the presence of Nucella, and not to the difference in predation.
To evaluate this claim, I reanalyzed the data from my New England experiments. Since both Petraitis and I have shown that barnacles enhance mussel abundance, it is desirable to determine if there is a predation effect on mussels that is independent of this barnacle enhance- ment effect. I used ANCOVA to determine the signifi- cance of the relationship between mussel abundance (de- pendent variable: percent cover) and predation (indepen- dent variable: presence or absence) with initial barnacle abundance (percent cover) as the covariate. Initial barna- cle abundance was used as the covariate in an effort to avoid possible confounding of barnacle abundance with different postrecruitment sources of mortality in different treatments. Data from both wave-exposed and wave- sheltered sites in August (when Petraitis' experiments were terminated) and October (when our short-term ex- periments were usually terminated) were analyzed. If our previous conclusion was true, then the results of this analysis would indicate a significant treatment (preda- tion) effect after the effect of the covariate (barnacle cover) was removed.
The results of this analysis generally support the hypothesis that predation has an effect on mussel abun- dance at relatively wave-sheltered (but not wave- exposed) sites (Table 1). At all but one site (Chamber- lain), the assumption of homogeneous slopes of mussel- barnacle regressions in the presence and absence of Nucella in August and October is accepted (Table 1; Predation • Initial Barnacle cover interactions in Tests for Parallel Slopes are not significant). At the exposed sites, the Covariance Analysis (Table 1) indicated that the effect of predation after removal of the effect of the covariate (Initial Barnacle cover) was not significant. At two protected sites, Little Brewster Cove, Massachusetts and Grindstone Neck, Maine, slopes were also homoge- neous (Table 1). In both cases, the Covariance Analysis indicated that the effect of predation in both August and October was highly significant independent of initial bar- nacle cover. For the results at Chamberlain, Maine, a somewhat more exposed but still relatively protected site, the assumption of homogeneous slopes is not met and renders invalid further analysis of the effect of predation on mussel cover using ANCOVA (Sokat and Rohtf 1981). Hence, this analysis suggests that predation had a strong effect on mussels at two of three wave-sheltered sites at least, and had no effect at wave-exposed sites. Both results are consistent with our earlier conclusions (e.g., Menge 1976).
Further analysis suggests that the results at Chamber- lain may also be consistent with our earlier conclusions.
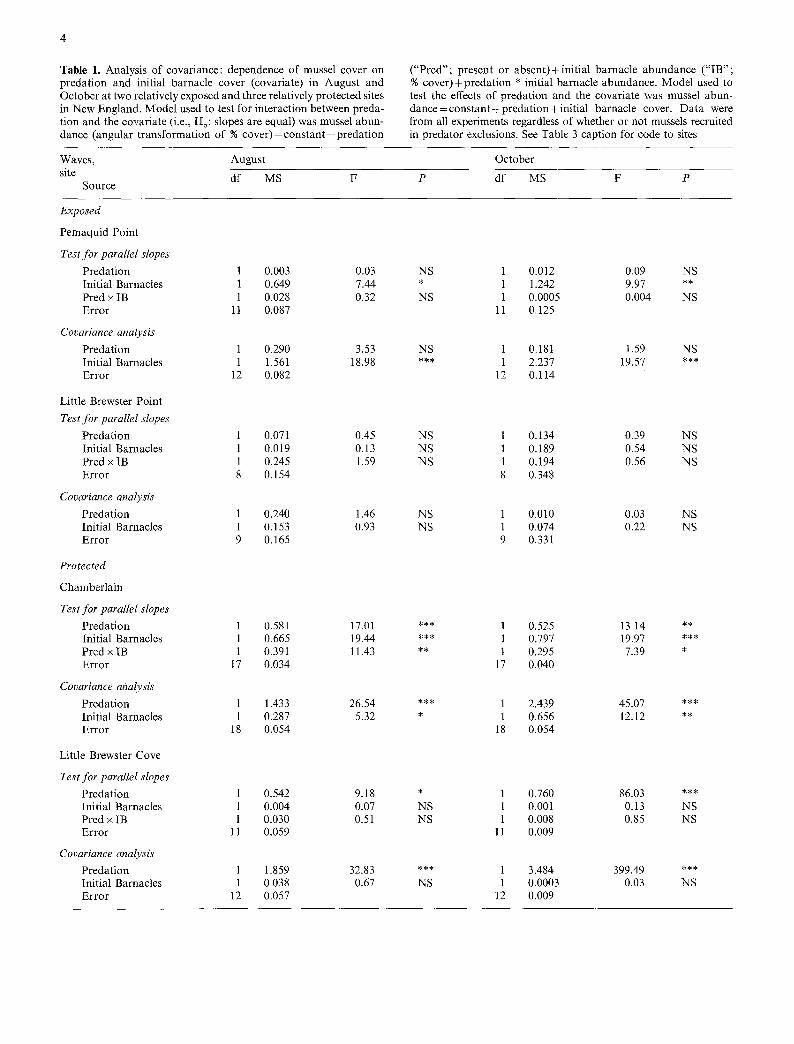
Table 1. Analysis o f covariance: dependence o f mussel cover on predat ion and initial barnacle cover (covariate) in August and October at two relatively exposed and three relatively protected sites in New England. Mode l used to test for interact ion between preda- t ion and the covariate (i.e., Ho: slopes are equal) was mussel abun- dance (angular t ransformat ion o f % c o v e r ) = c o n s t a n t + p r e d a t i o n
( "P red" ; present or absent) + initial barnacle abundance ( " IB" ; % cover) + predat ion * initial barnacle abundance. Model used to test the effects o f predat ion and the covariate was mussel abun- d a n c e = c o n s t a n t + p r e d a t i o n + i n i t i a l barnacle cover. Da ta were from all experiments regardless of whether or not mussels recruited in predator exclusions. See Table 3 caption for code to sites
Waves, August
site d f MS Source
October
P d f MS F P
Exposed
Pemaquid Point
Test for parallel slopes Predat ion 1 0.003 0.03 Initial Barnacles 1 0.649 7.44 Pred • IB 1 0.028 0.32 Error 11 0.087
Covarianee analysis
Predat ion 1 0.290 3.53 Initial Barnacles 1 1.561 18.98 Error 12 0.082
Little Brewster Point
Test for parallel slopes Predat ion 1 0.071 0.45 Initial Barnacles 1 0.019 0.13 Pred • IB 1 0.245 1.59 Error 8 0.154
Covariance analysis Predat ion 1 0.240 1.46 Initial Barnacles 1 0.153 0.93 Error 9 0.165
Protected
Chamber la in
Test for parallel slopes Predat ion 1 0.581 17.01 Initial Barnacles 1 0.665 19.44 Pred x IB 1 0.391 11.43 Error 17 0.034
Covariance analysis
Predat ion 1 1.433 26.54 Initial Barnacles 1 0.287 5.32 Error 18 0.054
Little Brewster Cove
Test for parallel slopes Preda t ion 1 0.542 9.18 Initial Barnacles 1 0.004 0.07 Pred x IB 1 0.030 0.51 Error l 1 0.059
Covariance analysis' Predat ion 1 1.859 32.83 Initial Barnacles 1 0.038 0.67 Error 12 0.057
NS 1 0.012 0.09 NS * 1 1.242 9.97 ** NS 1 0.0005 0.004 NS
11 0.125
NS 1 0.181 1.59 NS *** 1 2.237 19.57 ***
12 0.114
NS 1 0.134 0.39 NS NS 1 0.189 0.54 NS NS 1 0.194 0.56 NS
8 0.348
NS 1 0.010 0.03 NS NS 1 0.074 0.22 NS
9 0.331
*** 1 0.525 13.14 ** *** 1 0.797 19.97 *** ** 1 0.295 7.39 *
17 0.040
*** 1 2.439 45.07 *** * 1 0.656 12.12 **
18 0.054
* 1 0.760 86.03 *** NS 1 0.001 0.13 NS NS 1 0.008 0.85 NS
11 0.009
*** 1 3.484 399.49 *** NS 1 0.0003 0.03 NS
12 0.009
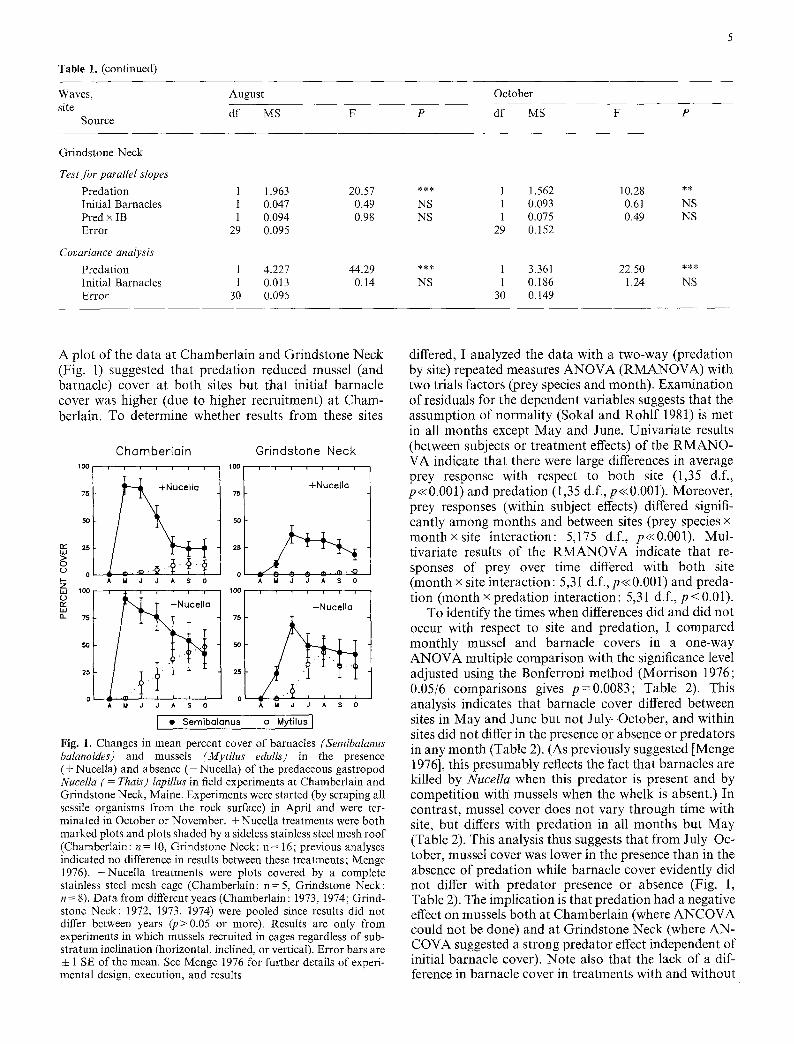
Table 1. (continued)
Waves, August
site df MS Source
F
October
P df MS F P
Grindstone Neck
Test for parallel slopes Predation 1 1.963 Initial Barnacles 1 0.047 Pred x IB 1 0.094 Error 29 0.095
Covariance analysis Predation 1 4.227 Initial Barnacles 1 0.013 Error 30 0.095
20.57 0.49 0.98
44.29 0.14
NS NS
NS
1 1.562 10.28 ** 1 0.093 0.61 NS 1 0.075 0.49 NS
29 0.152
1 3.361 22.50 *** 1 0.186 1.24 NS
30 0.149
A plot of the data at Chamberlain and Grindstone Neck (Fig. 1) suggested that predation reduced mussel (and barnacle) cover at both sites but that initial barnacle cover was higher (due to higher recruitment) at Cham- berlain. To determine whether results from these sites
n," h i > 0 0
I - Z w 0 n4 bJ 0-
Chamberlain 1 ~ l l , v l l =
2 5
M J J A S O
2 5
0 M J J A $ O
l O 0
7 5
5 0
2 5
0
I 0 0 u
I
:f 2,[ o
Grindstone Neck u i i i i v r
+Nucella
A M J J A $ O
, r l l , ~ l
-NucePla
A M J J A S O
I �9 Semibalanus o Mytilus I
Fig. 1. Changes in mean percent cover of barnacles (Semibalanus balanoides) and mussels (Mytilus edulis) in the presence ( + Nucella) and absence ( - Nucella) of the predaceous gastropod Nucella (= Thais) lapillus in field experiments at Chamberlain and Grindstone Neck, Maine. Experiments were started (by scraping all sessile organisms from the rock surface) in April and were ter- minated in October or November. + Nucella treatments were both marked plots and plots shaded by a sideless stainless steel mesh roof (Chamberlain: n = i0, Grindstone Neck: n = 16; previous analyses indicated no difference in results between these treatments; Menge 1976). - N u c e l l a treatments were plots covered by a complete stainless steel mesh cage (Chamberlain: n = 5, Grindstone Neck: n = 8). Data from different years (Chamberlain: 1973, 1974; Grind- stone Neck: 1972, 1973, 1974) were pooled since results did not differ between years (p>0.05 or more). Results are only from experiments in which mussels recruited in cages regardless of sub- stratum inclination (horizontal, inclined, or vertical). Error bars are ! 1 SE of the mean. See Menge 1976 for further details of experi- mental design, execution, and results
differed, I analyzed the data with a two-way (predation by site) repeated measures ANOVA (RMANOVA) with two trials factors (prey species and month). Examination of residuals for the dependent variables suggests that the assumption of normality (Sokal and Rohlf 1981) is met in all months except May and June. Univariate results (between subjects or treatment effects) of the RMANO- VA indicate that there were large differences in average prey response with respect to both site (1,35 d.f., p<<0.001) and predation (1,35 d.f., p<<0.001). Moreover, prey responses (within subject effects) differed signifi- cantly among months and between sites (prey species x month x site interaction: 5,175 d.f., p<<0.001). Mul- tivariate results of the RMANOVA indicate that re- sponses of prey over time differed with both site (month x site interaction: 5,31 d.f., p<<0.001) and preda- tion (month x predation interaction: 5,31 d.f., p < 0.01).
To identify the times when differences did and did not occur with respect to site and predation, I compared monthly mussel and barnacle covers in a one-way ANOVA multiple comparison with the significance level adjusted using the Bonferroni method (Morrison 1976; 0.05/6 comparisons gives p=0.0083; Table 2). This analysis indicates that barnacle cover differed between sites in May and June but not JulyOctober , and within sites did not differ in the presence or absence or predators in any month (Table 2). (As previously suggested [Menge 1976], this presumably reflects the fact that barnacles are killed by Nucella when this predator is present and by competition with mussels when the whelk is absent.) In contrast, mussel cover does not vary through time with site, but differs with predation in all months but May (Table 2). This analysis thus suggests that from July-Oc- tober, mussel cover was lower in the presence than in the absence of predation while barnacle cover evidently did not differ with predator presence or absence (Fig. 1, Table 2). The implication is that predation had a negative effect on mussels both at Chamberlain (where ANCOVA could not be done) and at Grindstone Neck (where AN- COVA suggested a strong predator effect independent of initial barnacle cover). Note also that the lack of a dif- ference in barnacle cover in treatments with and without
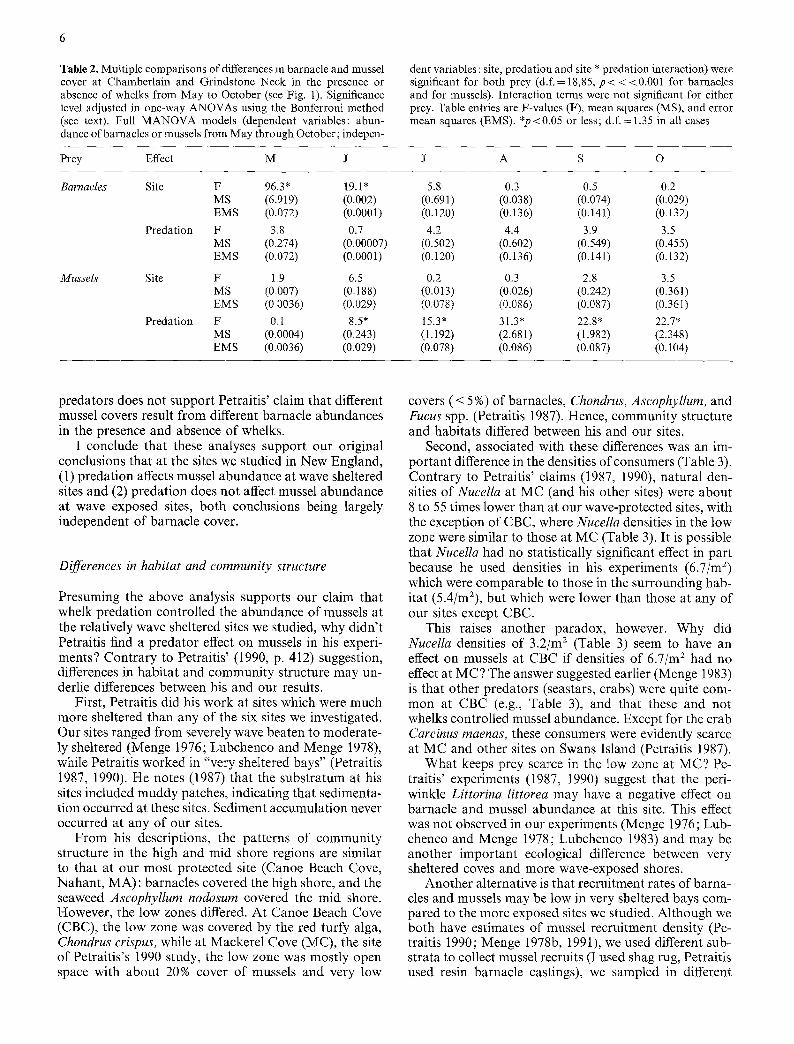
Table 2. Multiple comparisons of differences in barnacle and mussel cover at Chamberlain and Grindstone Neck in the presence or absence of whelks from May to October (see Fig. 1). Significance level adjusted in one-way ANOVAs using the Bonferroni method (see text). Full MANOVA models (dependent variables: abun- dance of barnacles or mussels from May through October; indepen-
dent variables : site, predation and site * predation interaction) were significant for both prey (d.f. = 18,85, p< < <0.001 for barnacles and for mussels). Interaction terms were not significant for either prey. Table entries are F-values (F), mean squares (MS), and error mean squares (EMS). *p<0.05 or less; d.f. = 1,35 in all cases
Prey Effect M J J A S O
Barnacles Site F 96.3" 19.1" 5.8 0.3 0.5 0.2 MS (6.919) (0.002) (0.691) (0.038) (0.074) (0.029) EMS (0.072) (0.0001) (0.120) (0.136) (0.141) (0.132)
Predation F 3.8 0.7 4.2 4.4 3.9 3.5 MS (0.274) (0.00007) (0.502) (0.602) (0.549) (0.455) EMS (0.072) (0.0001) (0.120) (0.136) (0.141) (0.132)
Mussels Site F 1.9 6.5 0.2 0.3 2.8 3.5 MS (0_007) (0.188) (0.013) (0.026) (0.242) (0.361) EMS (0_0036) (0.029) (0.078) (0.086) (0.087) (0.36I)
Predation F 0.1 8.5* 15.3" 31.3" 22.8* 22.7* MS (0.0004) (0.243) (1.192) (2.681) (1.982) (2.348) EMS (0.0036) (0.029) (0.078) (0.086) (0.087) (0.104)
predators does not support Petraitis' claim that different mussel covers result from different barnacle abundances in the presence and absence of whelks.
I conclude that these analyses support our original conclusions that at the sites we studied in New England, (1) predation affects mussel abundance at wave sheltered sites and (2) predation does not affect mussel abundance at wave exposed sites, both conclusions being largely independent of barnacle cover.
Differences in habitat and community structure
Presuming the above analysis supports our claim that whelk predation controlled the abundance of mussels at the relatively wave sheltered sites we studied, why didn't Petraitis find a predator effect on mussels in his experi- ments? Contrary to Petraitis' (1990, p. 412) suggestion, differences in habitat and community structure may un- derlie differences between his and our results.
First, Petraitis did his work at sites which were much more sheltered than any of the six sites we investigated. Our sites ranged from severely wave beaten to moderate- ly sheltered (Menge 1976; Lubchenco and Menge 1978), while Petraitis worked in "very sheltered bays" (Petraitis 1987, 1990). He notes (1987) that the substratum at his sites included muddy patches, indicating that sedimenta- tion occurred at these sites. Sediment accumulation never occurred at any of our sites.
F rom his descriptions, the patterns of community structure in the high and mid shore regions are similar to that at our most protected site (Canoe Beach Cove, Nahant, MA) : barnacles covered the high shore, and the seaweed Ascophyllum nodosum covered the mid shore. However, the low zones differed. At Canoe Beach Cove (CBC), the low zone was covered by the red turfy alga, Chondrus crispus, while at Mackerel Cove (MC), the site of Petraitis's 1990 study, the low zone was mostly open space with about 20% cover of mussels and very low
covers (< 5 %) of barnacles, Chondrus, Ascophyllum, and Fucus spp. (Petraitis 1987). Hence, community structure and habitats differed between his and our sites.
Second, associated with these differences was an im- portant difference in the densities of consumers (Table 3). Contrary to Petraitis' claims (1987, 1990), natural den- sities of Nucella at MC (and his other sites) were about 8 to 55 times lower than at our wave-protected sites, with the exception of CBC, where Nucella densities in the low zone were similar to those at MC (Table 3). It is possible that Nucella had no statistically significant effect in part because he used densities in his experiments (6.7/m 2) which were comparable to those in the surrounding hab- itat (5.4/m2), but which were lower than those at any of our sites except CBC.
This raises another paradox, however. Why did Nucella densities of 3.2/m 2 (Table 3) seem to have an effect on mussels at CBC if densities of 6.7/m 2 had no effect at MC? The answer suggested earlier (Menge 1983) is that other predators (seastars, crabs) were quite com- mon at CBC (e.g., Table 3), and that these and not whelks controlled mussel abundance. Except for the crab Carcinus maenas, these consumers were evidently scarce at MC and other sites on Swans Island (Petraitis 1987).
What keeps prey scarce in the tow zone at MC? Pe- traitis' experiments (1987, 1990) suggest that the peri- winkle Littorina littorea may have a negative effect on barnacle and mussel abundance at this site. This effect was not observed in our experiments (Menge 1976; Lub- chenco and Menge 1978; Lubchenco 1983) and may be another important ecological difference between very sheltered coves and more wave-exposed shores.
Another alternative is that recruitment rates of barna- cles and mussels may be low in very sheltered bays com- pared to the more exposed sites we studied. Although we both have estimates of mussel recruitment density (Pe- traitis 1990; Menge 1978b, 1991), we used different sub- strata to collect mussel recruits (I used shag rug, Petraitis used resin barnacle castings), we sampled in different
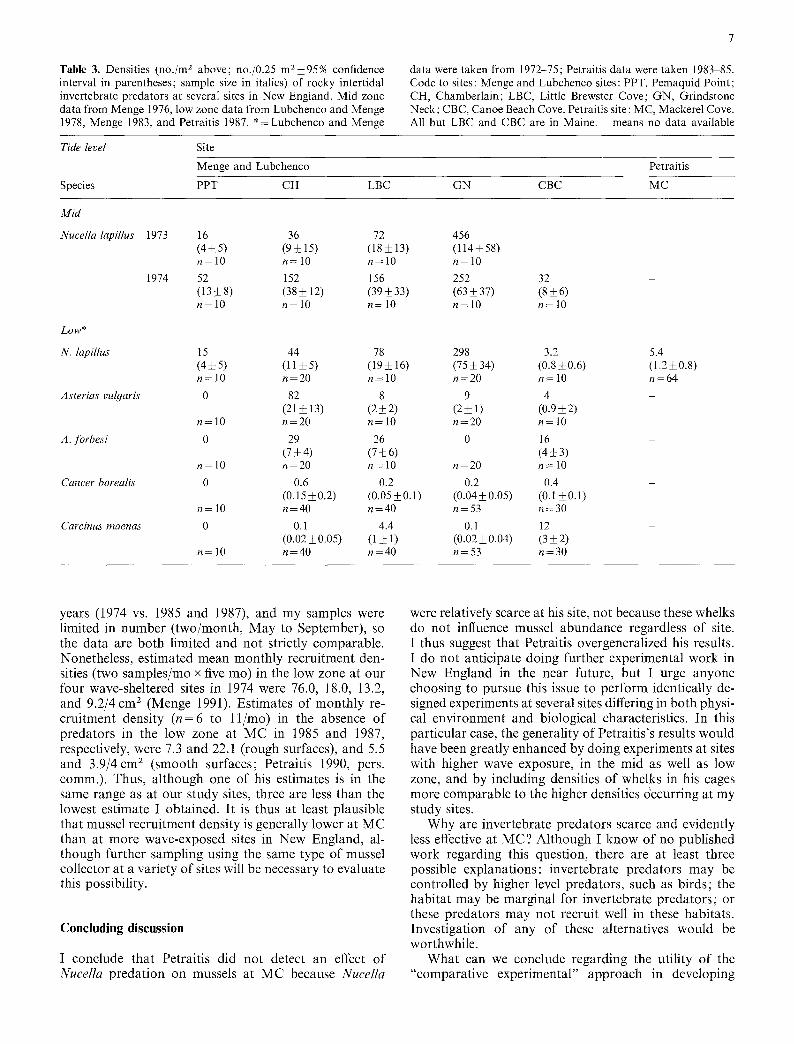
Table 3. Densities (no. /m 2 above; no./0.25 m 2_+95% confidence interval in parentheses; sample size in italics) of rocky intertidal invertebrate predators at several sites in New England. Mid zone data from Menge 1976, low zone data from Lubchenco and Menge 1978, Menge 1983, and Petraitis 1987. *=Lubchenco and Menge
data were taken from 1972-75; Petraitis data were taken 1983-85. Code to sites: Menge and Lubchenco sites: PPT, Pemaquid Point; CH, Chamberlain; LBC, Little Brewster Cove; GN, Grindstone Neck; CBC, Canoe Beach Cove. Petraitis site : MC, Mackerel Cove. All but LBC and CBC are in Maine. - means no data available
Tide level Site
Menge and Lubchenco Petraitis
Species PPT CH LBC GN CBC MC
Mid
Nucella lapillus 1973
1974
16 36 72 456 (4-+5) (9,+15) (18+13) (114+58) n=lO n=lO n=lO n=lO 52 152 156 252 32 (13 ,+ 8) (38 ,+ 12) (39 ,+ 33) (63 + 37) (8 + 6) n=lO n=lO n=lO n=lO n=lO
Low*
N. lapillus 15 44 78 298 3.2 (4 + 5) (11 + 5) (19 _+ 16) (75 + 34) (0.8 • 0.6) n= 10 n=20 n= 10 n=20 n= 10
Aster• vulgaris 0 82 8 9 4 (21_+13) (2__2) (2_+1) (0.9_+2)
n= 10 n=20 n=10 n=20 n=10
A. forbes• 0 29 26 0 16 (7 ,+ 4) (7 ,+ 6) (4 _+ 3)
n= 10 n=20 n=10 n=20 n= 10 Cancer borealis 0 0.6 0.2 0.2 0.4
(0.15+0.2) (0 .05,+0.1) (0.04+0.05) (0.1,+0.1) n= 10 n=40 n=40 n=53 n=30
Carcinus maenas 0 O. 1 4.4 O. 1 12 (0.02_+0.05) (1,+ 1) (0 .02_+0.04) (3_+2)
n=10 n=40 n=40 n=53 n=30
5.4 (1.2+o.8) n=64
years (1974 vs. 1985 and 1987), and my samples were limited in number ( two/month, May to September), so the data are both limited and not strictly comparable. Nonetheless, estimated mean monthly recruitment den- sities (two samples/mo x five mo) in the low zone at our four wave-sheltered sites in 1974 were 76.0, 18.0, 13.2, and 9.2/4 cm 2 (Menge 1991). Estimates of monthly re- cruitment density ( n = 6 to l l / m o ) in the absence of predators in the low zone at MC in 1985 and 1987, respectively, were 7.3 and 22.1 (rough surfaces), and 5.5 and 3 .9/4cm 2 (smooth surfaces; Petraitis 1990, pers. comm.). Thus, al though one of his estimates is in the same range as at our study sites, three are less than the lowest estimate I obtained. It is thus at least plausible that mussel recruitment density is generally lower at MC than at more wave-exposed sites in New England, al- though further sampling using the same type of mussel collector at a variety of sites will be necessary to evaluate this possibility.
Concluding discussion
I conclude that Petraitis did not detect an effect of Nuce l la predat ion on mussels at MC because Nuce l la
were relatively scarce at his site, not because these whelks do not influence mussel abundance regardless of site. I thus suggest that Petraitis overgeneralized his results. I do not anticipate doing further experimental work in New England in the near future, but I urge anyone choosing to pursue this issue to per form identically de- signed experiments at several sites differing in both physi- cal environment and biological characteristics. In this particular case, the generality of Petraitis 's results would have been greatly enhanced by doing experiments at sites with higher wave exposure, in the mid as well as low zone, and by including densities of whelks in his cages more comparable to the higher densities 0ccurring at my study sites.
Why are invertebrate predators scarce and evidently less effective at MC? Although I know of no published work regarding this question, there are at least three possible explanations: invertebrate predators may be controlled by higher level predators, such as birds; the habitat may be marginal for invertebrate predators ; or these predators may not recruit well in these habitats. Investigation of any of these alternatives would be worthwhile.
What can we conclude regarding the utility of the "comparat ive experimental" approach in developing

genera l i za t ions in c o m m u n i t y eco logy? I suggest tha t the deba te cons ide red here ac tua l ly suppor t s , r a the r than unde rmines its usefulness. A l t h o u g h in our ear l ie r w o r k we conc luded tha t p r e d a t i o n con t ro l l ed p rey a b u n d a n c e in the low zone a t re la t ive ly she l te red sites (Lubchenco and M e n g e 1978), we also conc luded t ha t when scarce (as a t CBC), the effect o f whelks on musse ls was m i n o r in c o m p a r i s o n to tha t o f o the r p r e d a t o r s , , ( M e n g e 1983). Pe t ra i t i s also obse rved tha t whelks were scarce and ex- e r ted a m i n o r effect on mussels in his exper iments , which is prec ise ly w h a t we w o u l d have p r ed i c t ed on the basis o f ou r ear l ie r work . I thus conc lude tha t ou r ear l ier gene ra l i za t ion has b r o a d e r app l i cab i l i t y t han to ju s t our s tudy sites, a n d suggest t ha t the c o m p a r a t i v e exper imen- ta l a p p r o a c h can be a power fu l too l in the d e v e l o p m e n t o f gene ra l i za t ions in ecology.
Acknowledgements. I thank R.T. Paine for drawing my attention to Petraitis' paper shortly after its publication. Comments by E. Ber- low, C. Blanchette, D. Brosnan, M. Hixon, J. Lubchenco, S. Navar- rete, A. Olson, C. Peterson, P. Petraitis, J. Sutherland, P. van Tamelen, S. Yamada, and an anonymous reviewer were useful in the preparation of this response. P. Petraitis suggested the AN- COVA analysis and provided guidance in its use. My research in rocky intertidal communities has been supported by NSF, most recently through grant number OCE88 11369.
References
Carpenter SC (1990) Large-scale perturbations: opportunities for innovation. Ecology 71:2038-2043
Caswell H (1978) Predator-mediated coexistence: a nonequilibrium model. Am Nat 112 : 127 154
Connell JH (1961) Effects of competition, predation by Thais lapil- lus, and other factors on natural populations of the barnacle Balanus balanoides. Ecol Monogr 31:61-104
Dayton PK (1971) Competition, disturbance, and community or- ganization: the provision and subsequent utilization of space in a rocky intertidal community. Ecol Monogr 41:351-389
Diamond J (1986) Overview: laboratory experiments, field experi- ments, and natural experiments. In: Diamond J, Case TJ (eds) Community Ecology. Harper and Row, NY. pp 3-22
Edwards DC, Conover DO, Sutter ItI F (1982) Mobile predators and the structure of marine intertidal animals. Ecology 63:1175-1180
Fairweather PG, Underwood AJ (1983) The apparent diet of preda- tors and biases due to different handling times of their prey. Oecologia 56 : 169-179
Hastings A (1980) Disturbance, coexistence, history, and com- petition for space. Theor Pop Biol 18:363-372
Landenberger DE (1968) Studies on selective feeding in the Pacific starfish Pisaster in Southern California. Ecology 49:1062-1075
Lubchenco J (1980) Algal zonation in the New England rocky intertidal community: an experimental analysis. Ecology 61 : 333-344
Lubchenco J (1983) Littorina and Fueus." effects of herbivores, substratum heterogeneity, and plant escapes during succession. Ecology 64: 1116-1123
Lubchenco J (1986) Relative importance of competition and preda- tion: early colonization by seaweeds in New England. In: Dia- mond J, Case TJ (eds) Community Ecology. Harper and Row, NY. pp 537-555
Lubchenco J, Menge BA (1978) Community development and per- sistence in a low rocky intertidal zone. Ecol Monogr 48 : 67-94
Louda SM (1982) Distribution ecology: variation in plant recruit- ment over a gradient in relation to insect seed predation. Ecol Monogr 52:25-41
Mauzey KP, Birkeland C, Dayton PK (1968) Feeding behavior of asteroids and escape responses of their prey in the Puget Sound region. Ecology 49:603-619
Menge BA (1972) Foraging strategy of a starfish in relation to actual prey availability and environmental predictability. Ecol Monogr 42 : 25-50
Menge BA (1976) Organization of the New England rocky inter- tidal community: role of predation, competition and environ- mental heterogeneity. Ecol Monogr 46 : 355-393
Menge BA (1978a) Predation intensity in a rocky intertidal com- munity. Relation between predator foraging activity and en- vironmental harshness. Oecologia 34:1-16
Menge BA (1978b) Predation intensity in a rocky intertidal com- munity. Effect of an algal canopy; wave action and desiccation on predator feeding rates. Oecologia 34:17-35
Menge BA (1982) Reply to a comment by Edwards, Conover, and Sutter. Ecology 63 : 1180-1184
Menge BA (1983) Components of predation intensity in the low zone of the New England rocky intertidal region. Oecologia 58 : 141-155
Menge BA (1991) Relative importance of recruitment and other causes of variation in rocky intertidal community structure. J Exp Mar Biol Ecol 146:69-100
Menge BA, Lubchenco J (1981) Community organization in tem- perate and tropical rocky intertidal habitats: prey refuges in relation to consumer pressure gradients. Ecol Monogr 51 : 429-450
Menge BA, Sutherland JP (1976) Species diversity gradients: syn- thesis of the roles of predation, competition, and temporal heterogeneity. Am Nat 110:351 369
Menge BA, Sutherland P (1987) Community regulation: variation in disturbance, competition, and predation in relation to en- vironmental stress and recruitment. Am Nat 130:730-757
Menge BA, Farrell TM (1989) Community structure and interac- tion webs in shallow marine hard-bottom communities: tests of an environmental stress model. Adv Ecol Res 19:18%262
Menge BA, Olson AM (1990) Role of scale and environmental factors in regulation of community structure. Trends Ecol Evol 5 : 52-57
Morrison DF (1976) Multivariate statistical methods, 2nd edition. McGraw-Hill, NY
Paine RT (1969) The Pisaster-Tegula interaction: prey patches, predator food preference, and intertidal community structure. Ecology 50:950-961
Paine RT (1980) Food webs: linkage, interaction strength and community infrastructure. J Anim Ecol 49:667-685
Peterson CH, Bradley BP (1978) Estimating the diet of a sluggish predator from field observations. J Fish Res Board Can 35:136-141
Petraitis PS (1987) Factors organizing rocky intertidal communities of New England: herbivory and predation in sheltered bays. J Exp Mar Biol Ecol 109:112136
Petraitis PS (1990) Direct and indirect effects of predation, herbiv- ory and surface rugosity on mussel recruitment. Oecologia 83 : 405-413
Schoener TW (1989) Food webs from the small to the large. Ecol- ogy 70:1559-1589
Seed R (1976) Ecology In: Bayne BL (ed) Marine Mussels: Their Ecology and Physiology. Cambridge Univ Press, Cambridge. pp 13 65
Sih A, Crowley P, McPeek M, Petranka J, Strohmeier K (1985) Predation, competition, and prey communities: a review of field experiments. Ann Rev Ecol Syst 16:269-311
Sokal RR, Rohlf FJ (1981) Biometry, second edition. WH Free- man, San Francisco