gaba-mediated repulsive coupling between circadian clock ... · gaba-mediated repulsive coupling...
TRANSCRIPT

GABA-mediated repulsive coupling between circadianclock neurons in the SCN encodes seasonal timeJihwan Myunga,b, Sungho Hongc, Daniel DeWoskind, Erik De Schutterc, Daniel B. Forgerd,e, and Toru Takumia,b,f,1
aRIKEN Brain Science Institute, Wako, Saitama 351-0198, Japan; bGraduate School of Biomedical Sciences, Hiroshima University, Minami, Hiroshima734-8553, Japan; cComputational Neuroscience Unit, Okinawa Institute of Science and Technology, Onna, Okinawa 904-0495, Japan; dDepartment ofMathematics, University of Michigan, Ann Arbor, MI 48109; eDepartment of Computational Medicine and Bioinformatics, University of Michigan, AnnArbor, MI 48109; and fJapan Science and Technology Agency, Core Research for Evolutional Science and Technology, Chiyoda-ku, Tokyo 102-0076, Japan
Edited by Joseph S. Takahashi, Howard Hughes Medical Institute, University of Texas Southwestern Medical Center, Dallas, TX, and approved June 11, 2015(received for review November 5, 2014)
The mammalian suprachiasmatic nucleus (SCN) forms not only themaster circadian clock but also a seasonal clock. This neural network of∼10,000 circadian oscillators encodes season-dependent day-lengthchanges through a largely unknown mechanism. We show that re-gion-intrinsic changes in the SCN fine-tune the degree of networksynchrony and reorganize the phase relationship among circadianoscillators to represent day length. We measure oscillations of theclock gene Bmal1, at single-cell and regional levels in cultured SCNexplanted from animals raised under short or long days. Couplingestimation using the Kuramoto framework reveals that the networkhas couplings that can be both phase-attractive (synchronizing) and-repulsive (desynchronizing). The phase gap between the dorsal andventral regions increases and the overall period of the SCN shortenswith longer day length. We find that one of the underlying physio-logical mechanisms is the modulation of the intracellular chlorideconcentration, which can adjust the strength and polarity of the ion-otropic GABAA-mediated synaptic input. We show that increasing day-length changes the pattern of chloride transporter expression, yieldingmore excitatory GABA synaptic input, and that blocking GABAA sig-naling or the chloride transporter disrupts the unique phase andperiod organization induced by the day length. We test the con-sequences of this tunable GABA coupling in the context of exci-tation–inhibition balance through detailed realistic modeling.These results indicate that the network encoding of seasonal timeis controlled by modulation of intracellular chloride, which de-termines the phase relationship among and period difference be-tween the dorsal and ventral SCN.
day-length encoding | repulsive coupling | SCN | GABA | chloride
The physiological and behavioral rhythms of all life on earth arebound to the Earth’s rotational cycle of ∼24 h. This funda-
mental rhythm is also affected by the planet’s slanted rotationalaxis, which causes seasonal variations in the length of the day. Howlife has adapted to anticipate this yearly rhythm is still debated.The suprachiasmatic nucleus (SCN), the central circadian (∼24 h)
pacemaker in mammals, consists of ∼10,000 “clock” neurons. Thesesingle-cell clocks maintain endogenous rhythms by autoregulatoryfeedback loops of genes including period (Per) and brain and muscleArnt-like 1 (Bmal1) (1). Although it was speculated that seasonalrhythms might be encoded in a single cell (2), single-cell oscillationsremain similar regardless of the seasonal time that the SCN tissueencodes (3). Seasonal timing is thus proposed to be encoded in thenetwork of the SCN (4–7) through spatial reorganization of therelative phases of clocks within individual cells (8–10). Subsets ofSCN clocks form phase clusters (11) that map approximately todorsal (shell, D-SCN) and ventral (core, V-SCN) regions of theSCN. When measured through a luciferase reporter monitoringoscillations in the Bmal1 gene, the D-SCN and V-SCN clustersshow a phase gap, with the D-SCN leading the V-SCN. Althoughthe D-SCN/V-SCN cluster structure is preserved across a mod-erate range of day length (8–16 h) (12), the phase gap increaseswith increasing day length (13). The mechanism for this phasegap remains unknown.
Previous studies explain robust circadian timekeeping by in-phasesynchronization of the SCN (14, 15), analogous to the classical phasemodel of Kuramoto (16). This in-phase state, however, is insufficientto explain the day length-dependent phase gap that is stably seenexperimentally (12, 13). According to the phase model, a phase gapcan result from the difference in period between the D-SCN and theV-SCN; a larger phase gap emerges when the difference in period ofthe D-SCN and V-SCN increases. Recent techniques have beenproposed to estimate the coupling of biological oscillators from theirtime-course data using the Kuramoto formalism (17, 18). Usingthese techniques, we discover the phase clusters emerge throughbalancing of phase-repulsion (increasing the phase gap) and phase-attraction (decreasing the phase gap) in the SCN. This networkmotif is also predicted to decrease the period of the SCN as a wholeas the phase gap increases, something we observe experimentally.One difference between the D-SCN and V-SCN is through the
effects of the ionotropic γ-aminobutyric acid receptor (GABAA)(19–21). Contrary to the convention that the GABAergic inputs arehyperpolarizing, high intracellular chloride concentration in someSCN neurons can cause GABA to depolarize the cells (20). We findthat the effects of the phase-repulsive coupling disappear afterpharmacological blockade of GABAA, by a surgical cut that seversphysical connections between the clusters or by the chloride importerblocker, which decreases intracellular chloride and reverts the po-larity of GABAA signaling back to its normal inhibitory state. Oneexplanation for this, which we test here, is that there is a day length-dependent increase of the relative expression of the neuronal chlo-ride importer (Nkcc1) vs. the exporter (Kcc2), in line with the recentfinding of increased excitation by GABA under LD (22). However,how increasing intracellular chloride concentration leads to phaseattractive or repulsive coupling remains a mystery.
Significance
How animals track the seasons has long been amystery. We founda mechanism that explains how day length is encoded within theneuronal network of suprachiasmatic nucleus (SCN). Using an in-tegrated approach combining experiments and modeling, we findevidence for changes in the coupling in the SCN that divides theclock oscillations into two clusters as a function of day length. Weshow that asymmetric distribution of intracellular chloride acrossthe SCN causes this coupling change. Blocking GABA or chlorideimport erases the oscillator organization formed by day-lengthentrainment. These demonstrate that coupling through GABA is akey ingredient of day-length encoding in the SCN.
Author contributions: J.M. and S.H. designed research; J.M., S.H., and D.D. performed research;J.M., S.H., D.D., E.D.S., D.B.F., and T.T. analyzed data; and J.M., S.H., D.D., D.B.F., and T.T. wrotethe paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1421200112/-/DCSupplemental.
E3920–E3929 | PNAS | Published online June 30, 2015 www.pnas.org/cgi/doi/10.1073/pnas.1421200112

To answer how increasing intracellular chloride concentrationleads to phase attractive or repulsive coupling, we appeal to de-tailed physiological modeling of the SCN. In a companion paper,we argue that GABA signaling normally has little effect on the SCNmolecular clock (23). However, if GABA excites the D-SCN, a
tonic signal is released that synchronizes the molecular clock inthe D-SCN and can desynchronize and diminish oscillations theV-SCN, as well as increase this phase gap between the D-SCN andV-SCN. Here we show simulations that the day length-mediatedincreases in excitatory GABA in the D-SCN cause a shortening ofthe period in the D-SCN and a weakening of the synchronizingsignal from the V-SCN, both of which increase the phase gap. Wevalidate that these effects have a similar effect on the coupling inthe Kuramoto framework as estimated from the experimental data,validating chloride’s role in coding day length.
ResultsRepresentation of Entrained Day Length in the SCN by NetworkInteractions. To determine how day length is internally repre-sented, we performed time-lapse imaging of single-cell bio-luminescent Bmal1 reporter activity (Bmal1-ELuc) in SCN explantsfrom mice previously entrained under long-day (16:8 h light:dark;LD) and short-day (8:16 h light:dark; SD) light-dark cycles. After 30or more days in the light cycle (6), mice developed changes in thefree-run period under constant darkness (DD) (LD: 23.59 ± 0.05 h,mean ± SEM, n = 13; SD: 23.97 ± 0.04 h, n = 10; P = 0.00002,Student’s t test; SI Appendix, Fig. S1). The period of the Bmal1-ELuc reporter oscillation in the SCN changes similarly to thechange in behavioral period after day-length entrainment (12),similar to Per1 GFP rhythms in cultured SCNs after perinatalphotoperiodic imprinting (24). Bioluminescence imaging revealedthat the phase distribution of the cellular Bmal1-ELuc oscillations inthe SCN becomes narrower after SD and wider after LD entrain-ment (Fig. 1 A and B). The D-SCN and V-SCN (D-V) phase gapincreased linearly with increasing day length from SD to LD(Spearman’s ρ = 0.045, P < 0.05) and the increase was significant(SD vs. LD and 12:12 vs. LD, P < 0.05; Welch’s t test, SD SCNexplants n = 8; 12:12 n = 6; LD n = 6) (Fig. 1C).The initial phase distribution was latently retained in cultures
of isolated SCN subregions [dorsomedial (DM), ventromedial(VM), and ventrolateral (VL) “onigiri” sections] (Fig. 1D) tar-geted to dissect phase clusters (12). In the isolated SCN sub-regions, the first-day peak times (acrophases) of DM, VM, andVL were similar in the SD-entrained group (Fig. 1E, Upper; SD:DM n = 12, VM n = 17, VL n = 12), whereas a wider phase gapwas seen between the DM and ventral subregions (VM and VL)in the LD-entrained group (Fig. 1E, Lower; LD: DM n = 12, VMn = 13, VL n = 11). The intact SCN explant maintained thephase gap throughout culture (Fig. 1B), better than the isolatedsubregions, an effect likely due to coupling (Fig. 1E), althoughthe phase gap in the intact SCN did increase with time. The peaktime difference in the isolated subregion culture was significantlywider in the LD-entrained group than in the SD group (bothDM-VM and DM-VL comparisons, LD: P < 0.01, SD: P < 0.05;Mann–Whitney U test) (Fig. 1F).
Phase Model-Based Analysis Suggests Phase-Repulsive Coupling inthe SCN. Circadian Bmal1-ELuc oscillations in SCN neurons canbe approximated using the phase equations proposed by Kuramoto(12, 16), which assume that the dynamics of individual oscillatorsare close to limit cycles, that coupling between cells is weak, andthat the coupling only depends on the difference in phase betweenthe oscillators. In this model, the instantaneous angular frequencyof individual oscillators (dθi/dt) is linearly related to its interactionwith other oscillators, and estimating coupling coefficients (Kij) isessentially a multiple linear regression problem between the dy-namics of an oscillator (dθi/dt) and the coupling terms [sin(θj − θi)](Materials and Methods). Assuming only positive coupling (Kij > 0)would cause the oscillators to all come into phase (16). This all-attraction model, however, fit the data poorly (R2 = 0.55 ± 0.03, n =15), as did a model with all repulsive coupling (Kij < 0) (R2 = 0.56 ±0.03, n = 15). When the coupling coefficients were allowed to haveboth signs, the goodness of fit greatly improved (R2 = 0.98 ± 0.01,
A
C
E F
D
B
Fig. 1. Day length reorganizes the phases of Bmal1 oscillations. The length ofday regulates synchrony of the peak phases of circadian Bmal1 promoter ac-tivities in the SCN. (A) SD entrainment induces a narrow phase distribution on acoronal plane of the SCN (Upper), whereas LD widens the phase gap betweenthe dorsal (D-SCN) and the ventral subregion clusters (V-SCN) (Lower). The ar-rows on the scale bar indicate SD (1σ) of the period distribution. (B) The clusteraverages of the detrended Bmal1-ELuc oscillations show a narrow phase gapbetween D-SCN and V-SCN under SD (Upper) and a wider phase gap under LD(Lower). (C) The D-SCN and V-SCN (D-V) phase gap in the first day in cultureincreases nearly linearly with day length across SCNs (*P < 0.05; Welch’s t test).The dashed line indicates the linear fit and horizontal variation is for illustrationonly. (D) The isolated SCN microsection (onigiri section) of the dorsomedial (DM,red) subregion approximates D-SCN and ventromedial (VM, green) and ventro-lateral (VL, blue) subregions together approximate V-SCN (Upper) as di-agrammatically illustrated and color-coded (Lower) for E. The bilateral VLmicrosections were put immediately next to each other in actual culture. En-semble averages (thick line) of detrended, normalized Bmal1-ELuc oscillations inthe isolated SCN subregions are presented against plots of all samples (halftransparent) from SD (Upper) and LD (Lower) entrainments. The relative phasedistribution is retained in the isolated subregions in the first day in culture (Inset:ensemble average for first 18 h). (F) The phase gap as a median peak timedifference between DM and VM also increases with day length (*P < 0.05; **P <0.01; Mann–Whitney U test). See SI Appendix, Fig. S2 for RT-qPCR measurementof mRNA levels in D- and V-SCN.
Myung et al. PNAS | Published online June 30, 2015 | E3921
NEU
ROSC
IENCE
APP
LIED
MATH
EMATICS
PNASPL
US

n = 15), suggesting that all-attractive or all-repulsive couplingassumptions for the model are insufficient for describing theobserved oscillations.We next ask what the coupling estimation tells us about the
typical behavior of the SCN oscillators in each subregion. Av-eraging the couplings within the D-SCN and V-SCN, which givesus the average impact of one region on the average oscillation inthe other, we see that the V-to-D-SCN coupling is predicted tobe the opposite of the D-to-V-SCN coupling (Fig. 2A). Wesimulated many other possible mixtures of repulsive and attrac-tive couplings and found that the asymmetric structure, seen inour estimated coupling coefficients, promotes a metastable state(25) with partial synchrony (marked with #, Fig. 2B). Further-more, a stable phase gap (attained by an intrinsic period differ-ence between the D-SCN and V-SCN) with this coupling makesboth subregions oscillate faster (ΔFreq > 0 in all counts; Fig. 2C,Upper) in a day length-dependent manner (Fig. 2C, Lower).
Dynamics of the Oscillator Network with Asymmetric and TunableCouplings. We constructed a two-oscillator phase model of asym-metrically coupled fast and slow oscillators that represent the D-SCNand V-SCN clusters respectively. As estimated from the experimental
data, the fast oscillating D-SCN receives phase-repulsive couplingfrom the slow oscillating V-SCN and the V-SCN receives phase-attractive coupling from the D-SCN (Eq. 4 andMaterials andMethods).A stable phase gap exists in this model and can be adjusted byvarying the relative coupling strengths (Fig. 2D and Materials andMethods). Under LD, when the D-SCN leads the V-SCN in phase,the larger phase gap translates as a stronger repulsive coupling thatfurther increases the phase gap by pushing the D-SCN faster. Atthe same time, a weak attractive coupling on the V-SCN alsospeeds up the V-SCN to decrease the phase gap and both clustersreach a stable state with a faster oscillation than when uncoupled.This speeding up of both the dorsal and ventral SCN is similar tothe period-shortening aftereffect seen under DD after first beingexposed to a long photoperiod (12, 26). The network encoding canbe modified by adjusting the strength of the repulsive coupling onthe D-SCN (Fig. 3D) or, equivalently, by changing the D-SCN’speriod of oscillation (SI Appendix, Fig. S4).Although conceptually simple, this model replicates various
photic entrainment behaviors by adjusting the repulsive couplingstrength between the D-SCN and the V-SCN (SI Appendix, Fig. S7),suggesting that modulation of the D-SCN may hold the key to theencoding of day length.
GABAergic Connections Mediate Phase-Repulsive Coupling. GABA,the most predominantly expressed neurotransmitter in the SCN, canhave both inhibitory and excitatory effects (19, 20), making it diffi-cult to interpret its role in terms of circadian synchronization (14, 20,27, 28). Because the same GABAergic signal can have oppositeeffects depending on the intracellular chloride concentration indownstream cells (19), we tested GABA as a physiological source ofthe phase-repulsive coupling using a GABAA blocker, gabazine(GBZ, 10 μM). GBZ immediately removed the short-period after-effect in the cultured SCN from LD-entrained animals (Fig. 3A,Middle). The neuronal connection that mediated day-length encod-ing was likely to be synaptic because the surgical separation of thedorsal and ventral SCN also removed aftereffects (Fig. 3A, Lower).The effect of GBZ on period was not significant in the SD-entrainedSCN (Fig. 3B) but it was in the LD-entrained SCN (P < 0.05, Stu-dent’s t test; Fig. 3C). The phase model predicted period lengtheningand the narrowing of the phase gap after removal of the repulsivecoupling as under GBZ treatment (Fig. 3D). Because GABA canwork as a desynchronizing agent (28), blocking it with GBZ slightlyincreases the period (12) and the synchronization index comparedwith the control (SI Appendix, Fig. S5), making it a strong candidatefor the source of the repulsive coupling.
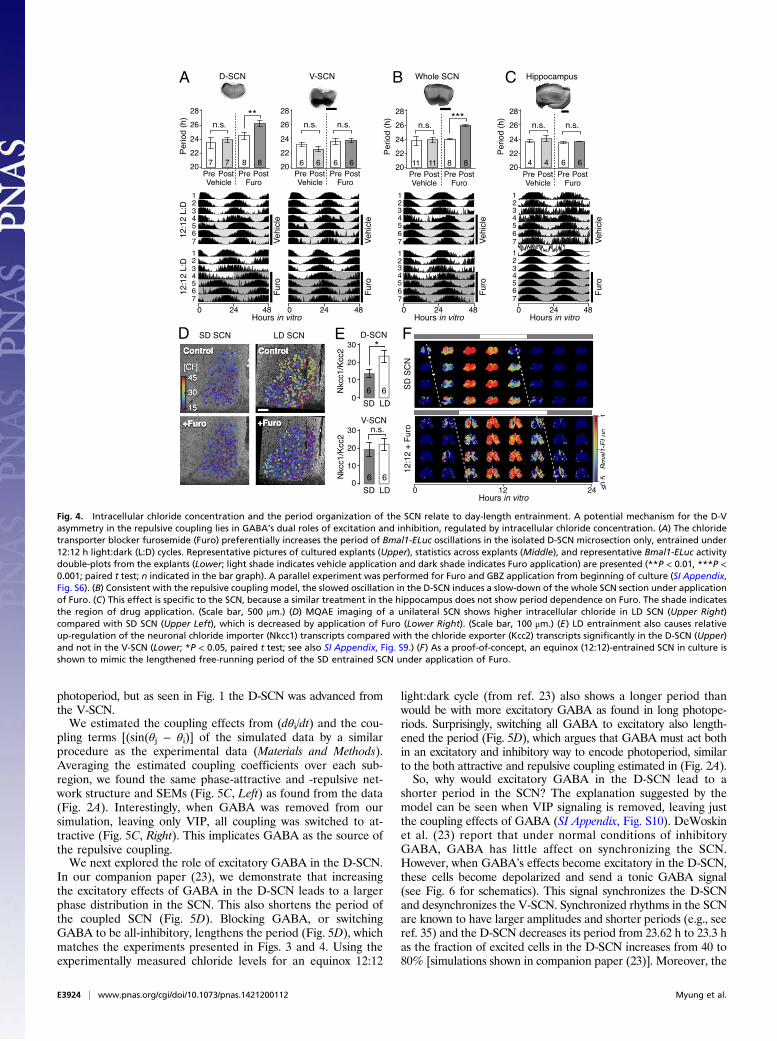
Day-Length Modulation of GABAA Connection by Adjustment ofChloride Homeostasis. Because the modulation of the D-SCN pe-riod was found to be a hallmark of day-length encoding, we soughtpharmacological agents that specifically modulate dorsal SCN os-cillations. We reasoned that preferential GABA excitation in theD-SCN (19) could contribute to the speed of the Bmal1 oscillationfrequency. The GABA excitation is caused by enhanced expressionof the neuronal chloride importer Nkcc1 compared with chlorideexporters, especially Kcc2 (29), leading to increased intracellularchloride and reversing the polarity of a cell’s response to GABAfrom inhibition to excitation in the SCN (20, 30). In isolated SCNsubregion cultures, chronic application of the chloride importer in-hibitor furosemide (Furo, 30 μM) increased the period of Bmal1-ELuc oscillations in the D-SCN, but not in the V-SCN, comparedwith the control condition before drug application (P < 0.01, paired ttest; Fig. 4A). The same was seen when Furo was applied at thebeginning of the culture. Coapplication of GBZ prevented this ef-fect, suggesting that the period lengthening is GABAA-mediated anda consequence of GABA excitation under high intracellular chlorideconcentration in the D-SCN (SI Appendix, Fig. S6). In the wholeSCN, Furo caused a period lengthening similar to that seen in theD-SCN, suggesting that the V-SCN follows the D-SCN and also
A B
C D
Fig. 2. Dynamics of SCN oscillators imply asymmetrically distributed repul-sive and attractive phase couplings. (A) The phase-model–based estimationof coupling finds a network motif of phase-repulsive coupling (blue) fromV- to D-SCN and phase-attractive coupling (red) from D- to V-SCN in thecross-sample mean (SEM ≤0.002, n = 6 12:12 SCNs). (B) Surrogate modelsunder various coupling schemes show that partial synchronization emergesunder the asymmetric attractive (+)–repulsive (−) coupling motif (markedwith ##) found in A, whereas desynchronization ensues under a randomlydistributed, structureless (marked with #) 50:50 mixture of attractive andrepulsive couplings (**P < 0.001, Student’s t test for triplicates; details inSI Appendix, Fig. S3). The synchronization index (order parameter, r) mea-sures the degree of synchronization at simulated day 30 (0 for completedesynchronization and 1 for complete synchronization). (C) Estimated shiftsin the intrinsic frequency due to the coupling effects (Upper) and averageintrinsic frequency in each subregion (Lower). (*P < 0.05, Student’s t test;number of explants indicated on the bar graph.) Note an increasing trend ofthe coupling contribution by day length in the D-SCN. (D) A two-oscillatorphase model explains the day length-modulated phase gap (Fig. 1A) with anasymmetric “push” from V- and “pull” from D-SCN as a consequence of thecoupling modulation, which can be explained by both coupling strength andintrinsic frequency (SI Appendix, Fig. S4).
E3922 | www.pnas.org/cgi/doi/10.1073/pnas.1421200112 Myung et al.

slows down its oscillation, causing the whole SCN to lengthen itsperiod (Fig. 4B). No such effect was seen when Furo was applied tothe hippocampus (Fig. 4C), which harbors a circadian clock but hasno known role in seasonal timekeeping.The corresponding relative bias of intracellular chloride con-
centration between the D-SCN and V-SCN was observed in ourmeasurement by N-(ethoxycarbonylmethyl)-6-methoxyquinoliniumbromide (MQAE) fluorescence. The chloride concentration wasfound to increase across the SCN after LD entrainment (Fig. 4D,Left) but was decreased by Furo preferentially in the D-SCN (Fig.4D, Right and SI Appendix, Fig. S8 D–I). At the transcript level,expression of all chloride transporters was up-regulated under LDcompared with SD (SI Appendix, Fig. S9 E, G, and H). However, inthe D-SCN, LD increased the relative expression ratio between theneuronal chloride importer Nkcc1 and the chloride exporter Kcc2(grand averages of six time points; P <0.05, paired t test), whereasthe increase was not significant in the V-SCN (Fig. 4E). In contrastto Kcc2, although the LD entrainment up-regulated the absoluteamount of transcripts, the change in the expression ratio betweenNkcc1 and Kcc1 was not significant (SI Appendix, Fig. S9I). Kcc2 iscritical for developmental EGABA shift in neurons and are expressedin nonarginine vasopressin (AVP) neurons in the SCN (31). An-other chloride exporter, Kcc1, is also neuronally expressed but,unlike Kcc2, is involved in osmotic cell volume regulation (31, 32).If indeed the intracellular chloride concentration modulates
the SCN network to encode the day length, the low intracellularchloride condition caused by chloride importer blockade should
switch a normal equinox (12:12 h light:dark)-entrained SCN intoan SD-entrained SCN. Consistent with this prediction, we foundthat application of Furo in the equinox (12:12)-entrained SCNnarrowed the internal phase gap and lengthened the period ofBmal1-ELuc oscillations in the entire explant, like what was seenin the SD-entrained SCN (Fig. 4F). These results suggest thatmodulation of chloride homeostasis can be a modus operandi forday-length encoding in the SCN.
Realistic Simulation Reveals the Mechanism of Day-Length Encoding.In a parallel study (23), we developed a multicellular and mul-tiscale SCN model (“realistic” model) comprising detailed elec-trophysiology (33), the biochemistry of intracellular circadiantimekeeping (34), as well as coupling through GABA and vaso-active intestinal polypeptide (VIP). This model tracks the bil-lions of action potentials and GABA postsynaptic currents in theSCN each day as well as over 1,000 chemical reactions in each ofover 1,000 cells that generate timekeeping. The model used ourmeasurements of intracellular chloride levels throughout the SCN,allowing for detailed study of the physiology of SCN coupling (seeMaterials and Methods and Fig. 6B for modeling schematics). Bymodulating the degree of GABA excitation in the D-SCN, as de-termined by our chloride measurements, and the intrinsic periods inthe cells of D-SCN, we found that the realistic model also predictedthe increasing phase gap with increasing day length (Fig. 5 A andB). Not only did the phase distribution increase with increasing
A B
C
D
Fig. 3. GABAA inhibition disturbs the phase and period organization caused by long or short day length. (A) Time-lapse images of normalized Bmal1-ELucoscillations in the LD-entrained SCNs are shown for every 3 h, color-coded to indicate the peak expression with dark red and lower-than-50% expression withdark blue (color bar on the bottom right). The clear D-V cluster formation and the short oscillation period can be seen in the control LD-entrained SCN(Upper). Application of the GABAA blocker gabazine (GBZ) lengthens the free-running period in culture while making the cluster separation less clear(Middle). Similarly, physical separation by a surgical cut (Inset) lengthens the free-running period in the LD SCN (Lower). (Scale bar, 100 μm.) (B and C) Theeffect on the free-running period is observed consistently in a larger number of samples through whole-field luminometry using a photomultiplier tube. GBZhas no effect on the SD SCN culture (B) but in the LD SCN culture (C) GBZ increases the period (*P < 0.05, Student’s t test; n indicated in the bar graph). (D) Theasymmetric repulsive coupling model can explain the phase and period organization under LD entrainment. With increased repulsive coupling in the D-SCN,the phase gap between D- and V- SCNs increases and the free-running period becomes shorter, as seen in the LD SCN. The light or dark shade indicates theregion of vehicle or drug application. Simulation parameters are described in Materials and Methods and SI Appendix, Table S2.
Myung et al. PNAS | Published online June 30, 2015 | E3923
NEU
ROSC
IENCE
APP
LIED
MATH
EMATICS
PNASPL
US
photoperiod, but as seen in Fig. 1 the D-SCN was advanced fromthe V-SCN.We estimated the coupling effects from (dθi/dt) and the cou-
pling terms [(sin(θj − θi)] of the simulated data by a similarprocedure as the experimental data (Materials and Methods).Averaging the estimated coupling coefficients over each sub-region, we found the same phase-attractive and -repulsive net-work structure and SEMs (Fig. 5C, Left) as found from the data(Fig. 2A). Interestingly, when GABA was removed from oursimulation, leaving only VIP, all coupling was switched to at-tractive (Fig. 5C, Right). This implicates GABA as the source ofthe repulsive coupling.We next explored the role of excitatory GABA in the D-SCN.
In our companion paper (23), we demonstrate that increasingthe excitatory effects of GABA in the D-SCN leads to a largerphase distribution in the SCN. This also shortens the period ofthe coupled SCN (Fig. 5D). Blocking GABA, or switchingGABA to be all-inhibitory, lengthens the period (Fig. 5D), whichmatches the experiments presented in Figs. 3 and 4. Using theexperimentally measured chloride levels for an equinox 12:12
light:dark cycle (from ref. 23) also shows a longer period thanwould be with more excitatory GABA as found in long photope-riods. Surprisingly, switching all GABA to excitatory also length-ened the period (Fig. 5D), which argues that GABA must act bothin an excitatory and inhibitory way to encode photoperiod, similarto the both attractive and repulsive coupling estimated in (Fig. 2A).So, why would excitatory GABA in the D-SCN lead to a
shorter period in the SCN? The explanation suggested by themodel can be seen when VIP signaling is removed, leaving justthe coupling effects of GABA (SI Appendix, Fig. S10). DeWoskinet al. (23) report that under normal conditions of inhibitoryGABA, GABA has little affect on synchronizing the SCN.However, when GABA’s effects become excitatory in the D-SCN,these cells become depolarized and send a tonic GABA signal(see Fig. 6 for schematics). This signal synchronizes the D-SCNand desynchronizes the V-SCN. Synchronized rhythms in the SCNare known to have larger amplitudes and shorter periods (e.g., seeref. 35) and the D-SCN decreases its period from 23.62 h to 23.3 has the fraction of excited cells in the D-SCN increases from 40 to80% [simulations shown in companion paper (23)]. Moreover, the
A B C
D E F
Fig. 4. Intracellular chloride concentration and the period organization of the SCN relate to day-length entrainment. A potential mechanism for the D-Vasymmetry in the repulsive coupling lies in GABA’s dual roles of excitation and inhibition, regulated by intracellular chloride concentration. (A) The chloridetransporter blocker furosemide (Furo) preferentially increases the period of Bmal1-ELuc oscillations in the isolated D-SCN microsection only, entrained under12:12 h light:dark (L:D) cycles. Representative pictures of cultured explants (Upper), statistics across explants (Middle), and representative Bmal1-ELuc activitydouble-plots from the explants (Lower; light shade indicates vehicle application and dark shade indicates Furo application) are presented (**P < 0.01, ***P <0.001; paired t test; n indicated in the bar graph). A parallel experiment was performed for Furo and GBZ application from beginning of culture (SI Appendix,Fig. S6). (B) Consistent with the repulsive coupling model, the slowed oscillation in the D-SCN induces a slow-down of the whole SCN section under applicationof Furo. (C) This effect is specific to the SCN, because a similar treatment in the hippocampus does not show period dependence on Furo. The shade indicatesthe region of drug application. (Scale bar, 500 μm.) (D) MQAE imaging of a unilateral SCN shows higher intracellular chloride in LD SCN (Upper Right)compared with SD SCN (Upper Left), which is decreased by application of Furo (Lower Right). (Scale bar, 100 μm.) (E) LD entrainment also causes relativeup-regulation of the neuronal chloride importer (Nkcc1) transcripts compared with the chloride exporter (Kcc2) transcripts significantly in the D-SCN (Upper)and not in the V-SCN (Lower; *P < 0.05, paired t test; see also SI Appendix, Fig. S9.) (F) As a proof-of-concept, an equinox (12:12)-entrained SCN in culture isshown to mimic the lengthened free-running period of the SD entrained SCN under application of Furo.
E3924 | www.pnas.org/cgi/doi/10.1073/pnas.1421200112 Myung et al.

V-SCN receives a tonic GABA signal at an earlier phase from thephase-advanced D-SCN, which, based on both experimental andsimulated phase-response curves (PRCs) (23), will delay theV-SCN. Thus, the V-SCN delays, increasing the phase gap betweenthe D-SCN and the V-SCN (SI Appendix, Fig. S10). In summary,excitatory GABA in the D-SCN speeds up the D-SCN and slowsthe V-SCN.If the D-SCN is phase-advanced from the V-SCN, experimental
and simulated PRCs both predict that the effect of VIP, which isreleased from the V-SCN, will delay the D-SCN, and lengthen itsperiod, in an effort to match its phase to that of the V-SCN.However, if the V-SCN is less synchronized and has a lower-amplitude rhythm, as would happen in the presence of excitatoryGABA, this VIP signal would become more diffuse, and the pe-riod-lengthening effect of the V-SCN on the D-SCN would di-minish. Our real-time quantitative PCR (RT-qPCR) data (SIAppendix, Fig. S2) suggest this because during long photoperiodsVIP transcripts in the V-SCN are present in significant amountsat all phases, matching this hypothesis. Thus, excitatory GABA inthe D-SCN diminishes the period lengthening effects of VIP onthe D-SCN, further speeding up the period.
DiscussionIn this paper, we propose asymmetrical repulsive coupling as anessential network mechanism that enables day-length encodingin the SCN. This type of coupling has been proposed previously,with the fundamental idea of phase repulsion implicit in themorning–evening oscillator interpretation of behavioral splittingunder LL (26, 36). Similarly, internal desynchronization has beenmodeled as two oscillators for sleep–wake cycles that are mu-tually repulsive of one other (37). Various scenarios with asym-metric coupling between two nonidentical oscillators have also
been considered using Pittendrigh–Pavlidis equations, whichshowed that a stable, but not antiphasic, phase gap is possiblewhen the positive (attractive) and negative (repulsive) couplinghave much different strengths (38).Our results point to modulation of GABA polarity as the
underlying physiological mechanism of tunable coupling (21, 22)that is responsible for day-length encoding via the phase gapbetween SCN oscillators (6). Although GABA is the mostabundant and ubiquitous neurotransmitter in the SCN, reportson its role for synchronization have been mixed. In neonatalpreparations, it is a synchronizer (14) or plays an unclear role(27), but in mature animals (>3 wk old) it can be a desynch-ronizer (28) (SI Appendix, Fig. S5) or can have both effectsdepending on the relative phase difference between SCN sub-regions (13). This is in contrast with VIP, which works as asynchronizer in the SCN (39) and plays a coherent role in thephase reorganization after entrainment under varied day lengths(13). GABA’s role in the SCN has been difficult to assess. Itspolarity can switch from inhibition to excitation depending ontime of the day (40) or the subregion of the SCN (19) throughmodulation of intracellular chloride (41). Chloride transporterscan switch the GABA-response polarity (20) and are expressedin a region-specific manner in the SCN (33) (Figs. 4E and 6A).Theoretically, we find two ways to increase the phase gap
between the D-SCN and V-SCN. First, the difference in intrinsicperiod between these two regions could be increased. Addi-tionally, the attractive coupling between the regions, likely me-diated by VIP signaling from the V-SCN, could be reduced. Ourresults point to both mechanisms working to modulate the phasegap. Excitatory GABA in the D-SCN causes greater synchronyand a shorter period in the D-SCN. It causes the opposite effecton the V-SCN, which releases the attractive pull of VIP.This is consistent with the higher synchrony in firing rate
rhythms in the D-SCN under LD (7) and dispersed phases in theV-SCN (5) seen experimentally. The weaker effective repulsivecoupling in the SD-entrained SCN is also consistent with highersynchrony (4) and molecular rhythmicity (42). The model pre-dicts that VIP provides the attractive coupling and that its re-moval markedly compromises the balance between the twoasymmetric couplings. GABA’s action is complex because itdepends on the intracellular chloride concentration, a fact thathas caused a great deal of confusion and debate in the literature(for example, see refs. 19, 20, and 40). Our experimental results,however, suggest that the modulation of GABA response through
A B
C D
Fig. 5. The realistic multiscale SCN model reproduces day length-dependentreorganization of phases. (A) A multiscale, multicellular SCN simulation thatmodels both electrophysiology and gene expression in each neuron faith-fully reproduces emergent separation of phases between D- and V-SCNsubregional oscillators under simulated LD, which is minimal under simu-lated SD. (B) The phase separation is replotted as the averages of the Bmal1transcript levels in D-SCN and V-SCN subregional clusters. (C) The estimatedmean phase coupling coefficients (K) from the simulation recovers theasymmetric coupling motif with a repulsive coupling from D-SCN to V-SCN(Left), which disappears when GABA coupling is removed from the modelparameter (Right) (SEM ≤0.001, n = 3 simulated SCNs). (D) The realisticmodel predicts both the shortened dorsal period owing to increased GABAexcitation during LD and the lengthened period in cultured SCN during GBZapplication (GABA KO) as consequences of the change in E/I ratio.
Cl-
Cl-A
Exc
itat
ory
G
AB
A
GABAA R
GABA
NKCC
KCC
[Cl-]i
Cl-High Cl-
Low Cl-Cl-
Cl-
Cl-
Inh
ibit
ory
G
AB
A
B
CaL/NMDARCl-
GABA
Ca2+
Depolarization
PKC/CREB
pCREB
CRE Per
Fig. 6. Model of day-length encoding by regulation of chloride homeo-stasis. (A) The GABA can introduce asymmetry in the SCN network couplingby the biased distribution of intracellular chloride, which can make GABAexcitatory (Upper) or inhibitory (Lower), driven by relative activation ofchloride importers (NKCC) vs. chloride exporters (KCC). (B) The GABA exci-tation under high intracellular chloride influences the clock gene expressioncycle through excitation–transcription coupling (21) and helps organize theoverall phase distribution of Bmal1 oscillations in SCN neurons, as the re-alistic model simulates.
Myung et al. PNAS | Published online June 30, 2015 | E3925
NEU
ROSC
IENCE
APP
LIED
MATH
EMATICS
PNASPL
US

the reorganization of chloride homeostasis makes a potent phys-iological pathway for day-length encoding. This interpretation isin line with the finding that the GABA-mediated excitatory inputincreases with the day length (22), given the increase of the chlo-ride importer over the exporter after LD entrainment. It is alsonotable that the short-period light–dark cycles of 22 h induce littlechange in methylation of clock genes but do affect other genesincluding chloride transporters (ref. 43; accession nos. GSE54124and GSE54021).In LD- and SD-entrained SCNs, we find little difference in the
amount of Bmal1 and Per2 transcripts, whereas LD up-regulatestranscripts of chloride transporters (Nkcc1, Kcc1, and Kcc2),their regulator (Wnk3), and major neuropeptides of the SCN(Avp and Vip) (SI Appendix, Fig. S9). We also find that theneuronal “with no lysine” kinase (Wnk3) is up-regulated in theD-SCN under LD (SI Appendix, Fig. S9F), because it increasesthe activity of NKCC1 and inhibits KCC1 and KCC2 (32).Synchronization is a powerful mechanism for bringing order
out of a number of neurons. However, if all neurons are lockedin the same state, they are unable to process diverse and complexinformation. The brain is, therefore, often observed to be in themetastable state between complete synchrony and desynchronythat generates spontaneously formed patterns such as, for ex-ample, those seen in ocular dominance columns (25). Thismetastable network state in the SCN is programmable by theseasonal day length and persists on a timescale much longer thanthe circadian timescale (44). The clock neurons in the SCN alsoexhibit spatiotemporal patterns in the oscillatory clock gene ex-pressions (13, 45). Metastability in the brain clock was noticedeven in the early days of circadian biology and has long beenthought to be relevant for encoding seasonal rhythms (26). Ourstudy identifies an active force resisting complete synchroniza-tion as a mechanism for metastability in the SCN, which enablesa richer encoding of day-length information. Because of thesefindings, we believe that the SCN potentially makes a powerfulplatform to study the brain’s metastable states.
Materials and MethodsDay-Length Entrainment of Bmal1-ELuc Transgenic Mice. We used Bmal1-Elucluciferase reporter mice, generated by Nakajima (46), on a C57BL/6J back-ground (SLC Japan) carrying a heterozygous transgene at an age aroundpostnatal 60–90 d. Animal experiment protocols were approved by theAnimal Research Committees of Hiroshima University and RIKEN Brain Sci-ence Institute. Conditions for breeding and day-length entrainment wereidentical to those in our previous study (12). Animals were housed in in-dividual cages with light controlled by a computer. The control day-lengthcondition was defined by the equinox light–dark ratio (12:12 h light:dark,L:D). The seasonal day-length conditions simulated summer and winterconditions approximately at the latitude of 45°N. In the SD paradigm, thelight phase lasted for 8 h and the dark phase for 16 h (8:16 L:D), and in theLD paradigm the light phase lasted for 16 h (16:8 L:D). Day-length entrain-ment was performed for minimum of 30 d before sampling (6). Control andmonitoring of the light conditions and general locomotor activities weredone with a white light-emitting diode and a periinfrared sensor through anArduino microprocessor (SmartProjects). The breeding room was maintainedat 25 °C at all times but inside the individual breeding chamber the ambienttemperature fluctuated slightly with the animal’s body temperature. Thisfluctuation was not exogenous and hence not a temperature cue for entrain-ment. Each chamber had a fan that generated constant environmental noise.
Continuous Bioluminescence Monitoring of SCN Explant Culture. Slice culture,imaging, and photomultiplier luminometry were performed as describedpreviously (12). Briefly, brains were isolated between 1600–2000 hours underdim light (<40 lx) after carefully dissecting optic nerves and coronally slicedat 250-μm thickness on a Microslicer (DSK) in ice-cold HBSS. SCNs were closelytrimmed immediately. All SCN slices were taken from the midposterior (MP)section of the SCN, defined as the region immediately posterior to thesudden breakpoint in the spatial distribution of phase clusters along theanterior–posterior axis, where the distribution becomes from mediolateralto dorsoventral (12). Each trimmed SCN explant was cultured on a mem-brane (Millicell-CM; Millpore) in a 35-mm dish containing 1 mL of Dulbecco’s
Modified Eagle Medium (Sigma) supplemented with B27 (Gibco/Invitrogen),penicillin–streptomycin (25 U/mL–25 μg/mL; Nacalai Tesque), and 100 μMbeetle luciferin (Promega). To prevent evaporation, each dish was sealedairtight with vacuum grease. Stock aliquots of 10 mM gabazine (Sigma) insterile distilled water and 30 mM furosemide (Sigma) in DMSO were addedto the 1-mL medium immediately before recording (1:1,000 dilution). Theslice culture was maintained at 37 °C (effective around the culture dish) andimaged continuously using three independent systems: a commercial in-cubator-microscope system (LV200, Olympus; Orca C4742-80–12AG, Hama-matsu; and a 40× objective, N.A. 0.6 LUCPlan FLN, Olympus), a custom-builtupright imaging system around a Nikon FN1 microscope (Orca R2 CCDcamera or ImagEM1K EM-CCD camera, Hamamatsu; a 20× objective or a 10×objective, and a relay lens, Nikon), and a custom-built microscope-free im-aging system (Hamamatsu Orca R2 directly coupled to a Nikon 20× objectiveand a Tokai Hit microincubator chamber). Parts of recordings publishedpreviously were reanalyzed for coupling estimation (12). Binning was set to4 × 4 and the exposure time was 15 min. For continuous bioluminescencemonitoring of the whole culture dish, we used a 24-dish-on-a-wheel pho-tomultiplier system (LM-2400; Hamamatsu) that counts photons for 1 minevery 15 min, per dish.
Subregional Isolation (Onigiri Section) of the SCN Explant. SCN explants fromthe MP section were microdissected into the dorsomedial (DM or D) regionand the ventral region (V), which was further dissected into ventromedial(VM), and ventrolateral (VL) regions. This scheme of microdissection was aspreviously described (onigiri sectioning, ref. 12). Sectioning was rapidlyperformed on explants after transferring onto the culture membrane. TheDM region was a V-shaped area from the dorsal tip of the explant extendingto the ventral tip of the third ventricle where the bilateral SCNs join. The cutregion was transferred on the blade to a separate dish. The sectioning andtransfer took about 30 s. The dishes were immediately sealed with vacuumgrease and incubated at 37 °C.
Measurement of Intracellular Free Chloride Concentration. We estimated thechloride concentration using quenching of MQAE (Molecular Probes) fluo-rescence by chloride. The MP sections of the SCN sliced at 100- to 150-μmthickness were transferred to culture membranes and incubated in culturemedium containing 2 mM MQAE for 1–2 h at 37 °C in the dark. Afterwashing and a brief incubation in artificial cerebrospinal fluid (ACSF; 124NaCl, 3 KCl, 1.2 CaCl2, 2.8 MgCl2, 25 NaHCO3, 1.2 NaH2PO4, 10 D-glucose, and10 Hepes, in millimolar, pH 7.4, with NaOH), fluorescence images werescanned at room temperature (23–25 °C) in z axis stacks on either a confocalmicroscope (FV1000; Olympus) under 405-nm excitation over 425–480 nm ora two-photon microscope (A1R; Nikon) under mode-locked 750-nm excita-tion over a 400- to 492-nm emission window. After imaging in ACSF (135 mMexternal chloride), the maximal fluorescence (F0) was determined by scanningthe sample after incubation in 0 mM chloride standard (0 KCl, 140 K-gluconate,10 Na-gluonate, 10 Hepes, in millimolar, pH 7.4, with KOH) and ionophoresnigericin (100 μM; Invitrogen) and tributyltin chloride (100 μM; Sigma). To de-termine the intracellular chloride concentration, we used the Stern–Volmer re-lation, ½Cl−�=K−1
SV ðF0=F − 1Þ, and calibrated using known chloride concentrationsat 10, 20, 30, and 40 mM by adjusting concentrations of KCl and K-gluconate ofthe 0 mM standard (SI Appendix, Fig. S8). Owing to bleaching, not all chloridestandards were used at the same time in one experiment. The Stern–Volmerconstant (KSV) was estimated to be 13 M−1 in aqueous solution (SI Appendix, Fig.S8 A–C) but was variable across the SCN samples in situ (10–25 M−1) owing tocellular exclusion and fluorescence bleaching of the MQAE dye. Images beforeand after drug treatments or chloride standards were flattened through z-stackaveraging on ImageJ (NIH) and automatically aligned using the imregister func-tion on MATLAB (The MathWorks). Regions-of-interest (ROIs) were manually se-lected to cover the round neuronal cell soma while avoiding areas around bloodvessels. ROI fluorescence averages and center-of-mass positions were exportedand sorted by center-of-mass position (SI Appendix, Fig. S8F) in Mathematica(Wolfram Research). The consistency of coronal slice plane between SD and LDcomparison groups was confirmed by comparable spatial AVP expression patternsin the post hoc immunofluorescence images (SI Appendix, Fig. S11), producedwith a polyclonal AVP antibody (1:1,000 dilution, Chemicon/Millipore) followingthe protocol described earlier (12).
Quantification of Chloride Transporter Transcripts by RT-qPCR. LD- and SP-entrained SCN samples were taken from six to eight animals after de-capitation under dim light (<40 lx) in 4-h time bins throughout a day (24 h)with an average of three independent samplings for each targeted time bin.The MP sections of the SCNs prepared at 300- to 350-μm thickness weretrimmed in RNAlater ICE solution (Life Technologies) to remove the optic
E3926 | www.pnas.org/cgi/doi/10.1073/pnas.1421200112 Myung et al.

chiasm and hypothalamic regions surrounding the SCN and dissected intoD-SCN and V-SCN microsections. The exact sampling time was determined asthe time that a slice was put into the RNAlater ICE solution. After dissection,samples were quickly transferred to 50 μL PicoPure extraction buffer (LifeTechnologies) and chilled in liquid nitrogen before being stored at −80 °C.RNA purification was performed using a PicoPure kit (10-μL elution) or anENZA total RNA kit (20-μL elution; Omega Bio-Tek). Total RNA from samplesper one time point was for D-SCN, 572 ± 31 ng (n = 50, SD and LD pooled;mean ± SEM) and for V-SCN, 384 ± 19 ng (n = 49), quantified using aspectrophotometer (NanoDrop; Thermo Scientific). The RNA quality assessedby RNA integrity number was 8.83 ± 0.17, estimated with 2100 BioAnalyzerNanoChip (Agilent Technologies) from randomly selected samples. After anRNA purity check, cDNA was synthesized with random primers. RT-qPCR wasperformed in three technical replicates using the cDNA equivalent of 7.5 ngmRNA in a 20-μL volume with SYBR Green DNA binding dye on 7900HT andStepOnePlus (all Applied Biosystems). The reaction steps were 50 °C for2 min, 95 °C for 10 min, and 40–45 cycles at 95 °C for 15 s and 60 °C for 1 min.PCR efficiencies of the primers were evaluated on SCN samples using sixdilution standards in triplicates. Two reference genes (Actb and Gapdh) wereused for standardization (47). Estimated Ct values were exported and ana-lyzed on Mathematica. We used the PCR efficiency (1 for 100%) and Ctvalues for quantification without weighting using the formula
�1+ PCR efficiencyActb
�CtActb+�1+ PCR efficiencyGapdh
�CtGapdh
2�1+ PCR efficiencysample
�Ctsample
and validated the formula using two dilutions of the cDNA template (7.5 ngvs. 15 ng) in select samples. The sequences of the forward and reverse primersare listed in SI Appendix, Table S1. All measurements were made ontechnical triplicates.
Analysis of Time-Lapse Bioluminescence Imaging Data. Two methods wereused for image preprocessing. In the ROI-based method, square pixels (15.4 ×15.4 μm2) containing bioluminescent cells were manually selected on ImageJand the mean luminosity and position of the ROIs were exported for sub-sequent period and phase analyses in Mathematica. In the grid-basedmethod, we first removed outliers from the raw images and performedconvolution with a single-cell-sized homogeneous kernel and reduced theimage size such that the kernel size corresponded to one pixel, effectivelymapping the images to ROI averages over all areas. The pixels in thosepreprocessed images were selected by a brightness criterion (12), sorted, andanalyzed in Mathematica.
Phase-Oscillator Model and Fitting of the Model to Data. In the phase-oscillatormodel-based approach, we assumed that the network of SCN neuronsfunctions as a system ofweakly coupled oscillators, as in other studies (12, 14).This led to a generalization to the canonical phase model (48), which de-scribes the temporal evolution of the phase θi of the i-th oscillator withperiod τi as
dθidt
=2πτi
+XNi=1
Hij
�θj − θi +φij
�. [1]
We assumed that the interaction function Hij is dominated by its firstFourier mode. We also assumed that the phase delay φij is vanishingly smallcompared with the circadian period. Therefore, the model was simplified tothe Kuramoto model (16):
dθidt
=2πτi
+XNi=1
Kij sin�θj − θi
�. [2]
Given our phase data {θi(t)} for individual oscillators, we can find Kij thatminimizes the sum of squared error (SSE) (see also ref. 18):
SSE=
������dθiðtÞ=dt−ÆdθiðtÞ=dtæt
�−XNj=0
Kij
�sin
�θjðtÞ−θiðtÞ
�−Æsin
�θjðtÞ−θiðtÞ
�æt������
2
2
,
[3]
where <x>t represents a time average of x. Note that we subtracted time-independent parts and only used the fluctuations in the data. In the cases fornegative or positive-only coefficients, we combined this SSE minimizationwith the corresponding constraint.
In all cases, we had more cells than the length of the data, making ourregression an ill-posed problem, particularly in the unconstrained estimationof negative and positive couplings. Inspired by the observation that couplingsin the SCN are sparse (49, 50), that is, only a small number of Kij’s are non-zero, we used simultaneous minimization of the L1 norm
��Kij
��1 =
PNj= 1 jKij j
for each i, which is known to efficiently recover the sparse coefficient matrix(51). By testing with simulated data in Fig. 2B and SI Appendix, Fig. S2, wefound that this procedure reduces the magnitude of the estimated couplingsbut maintains the sign of the couplings with significance. We also carriedout 10-fold cross-validation. For constrained and unconstrained cases, weused the convex optimization package CVX (52) and the glmnet package(53) in some unconstrained cases. All of the estimation procedures wereperformed in MATLAB 2012a.
For the simulated data used in Fig. 5, we were not limited by a smallnumber of time points. Therefore, we skipped the L1 norm minimization,performed a linear regression as described in the text, and verified that itgave results consistent with that found in Fig. 2. The longer and more stablesimulated data also allowed us to do the proper protophase-to-phasetransformation (18), which was done by extracting the protophases via aHilbert transform (where the first and last data points of the simulationwere not included) during one cycle for each of the oscillators individually,and combining them into one probability density function.
A Two-Oscillator Model for Day-Length Encoding. The coupling estimationinspired us to reduce Eq. 2 for the SCN network into two phase-oscillators,the dorsal (θD) and the ventral (θV) with respective intrinsic frequencies ωD
and ωV , which are related to the period τ by ω = 2π/τ:
dθDdt
=ωD −KVD sin�θV − θD
�
dθVdt
=ωV +KDV sin�θD − θV
�,
[4]
where KVD > 0 and KDV > 0 indicate the coupling contributions from V to Dand from D to V, respectively. KVD is preceded by a minus sign to representrepulsive coupling. Eqs. 4 are more easily analyzed by rewriting the variablesin terms of averages and differences: that is, phases as θ= ðθV + θDÞ=2 andΔθ= θV − θD, frequencies as ω= ðωV +ωDÞ=2 and Δω=ωV −ωD, and couplingconstants as K = ðKVD +KDVÞ=2 and ΔK =KDV −KVD. If the phase gap is small,that is, both oscillators operate in a state close to synchrony, we can ap-proximate sinΔθ∼Δθ and write
dΔθdt
=Δω−ΔK ·Δθ
dθdt
=ω−K ·Δθ:[5]
The oscillatorsmaintain a stable phase gap fΔθ=Δω=ΔKwhen dΔθ=dt = 0 andan effective frequency ~ω=ω−K · fΔθ= ~ω−ΔωK=ΔK. External light input can bemodeled to influence the average phase of the oscillator system (dθ/dt) so thatEq. 5 becomes
dθdt
=ω−K ·Δθ+ I�ϕlight − θ
�. [6]
Parameters for simulating consequences of various light inputs (SIAppendix, Fig. S7) are aD = 0.267 rad/h (τD = 23.5 h), ωV = 0.256 rad/h (τV = 24.5 h),KVD = 0.001 h−1, and KDV = 0.01 h−1. For rhythmic light input I = 0.01 h−1,and for constant conditions I(ϕlight − θ) = 0 rad/h, with other parametersunchanged for DD and ωD = 0.242 rad/h (τD = 26 h) for LL (SI Appendix,Table S2).
Realistic Multiscale, Multicellular SCN Model. Detailed simulations of SCNsignaling were performed using a multiscale, multicellular SCN model, de-veloped and given in full detail in refs. 23 and 35. Briefly, the model simulatesa network of 1,024 SCN neurons. Each neuron contains a biochemical model,based on mass action kinetics, of the negative feedback loops regulatingclock gene expression (34). Additionally, the electrical activities of eachneuron are also simulated, down to the timescale of individual action po-tentials, using a conductance-based model of SCN electrophysiology (33).These two models are coupled bidirectionally within each cell; the molecularclock phase controls two potassium channel conductances (gKCa and gK-leak)leading to circadian variation in the resting membrane voltage and electricalactivity of the cell. In the other direction, calcium from the electrophysiologymodel affects VIP release and can directly lead to activation of Per tran-scription as in ref. 35. Model cells in the network are coupled by VIP (some to
Myung et al. PNAS | Published online June 30, 2015 | E3927
NEU
ROSC
IENCE
APP
LIED
MATH
EMATICS
PNASPL
US

all connectivity) as in ref. 35, and through GABAA signaling (10% randomconnectivity), modified from ref. 54 to match the strength and timing ofGABAergic currents in the SCN (54, 55) (gsyn = 0.5, VT = −20 mV, KP = 3.0).The model enabled us to evaluate the overall network effect of differ-entially distributed chloride concentrations in single cells by varying theEGABA for different cells in the network (−32 mV for GABA-excited and −80mV for GABA-inhibited cells, respectively). All other electrophysiologyparameters were unchanged between the two populations and as in ref.23. SD SCN simulations used all original parameters as in ref. 23, with 40%of cells excited by GABA. For the LD SCN simulation, all molecular clockparameters were scaled to 102% of their original values in the 40% of SCNcells in the dorsal shell; ventral SCN parameters were unchanged. This ledto effective intrinsic periods of 23.7 h in the dorsal and 24.2 h in theventral SCN.
Statistics. For significance tests, we first checked the datasets for normalityand equal variance and when the two criteria were met we used Student’s ttest; when variance was unequal, we used Welch’s t test. For nonnormaldata, we used Mann–Whiney u test. For linear correlation of ordinal data
(day lengths), we used Spearman’s ρ. All statistical tests were performed inMathematica 9 and MATLAB 2012.
ACKNOWLEDGMENTS. We thank Michael Rosenblum for encouraging us topursue the idea of the phase-repulsive couplings in the beginning of thisproject. We also thank Hugh Piggins for editorial advice, Abdelhalim Azzi fordiscussion on genome-wide methylation and advice on real-time quantita-tive PCR of SCN samples, Heather Balance for advice on suprachiasmaticnucleus sampling for RNA, Pia Rose for critical reading of the manuscript,and all technicians of the T.T. laboratory for their assistance. This workwas supported by Human Frontiers of Science Program Grant RPG 24/2012.J.M. was supported by RIKEN Incentive Research Project Grant G1E-54500.T.T. was supported by Ministry of Education, Culture, Sports, Science andTechnology in Japan Grants-in-Aid for Scientific Research 25240277, 23111005,and 26670165, Core Research for Evolutional Science and Technology andthe Strategic International Cooperative Program from the Japan Scienceand Technology Agency, an intramural research grant for neurological andpsychiatric disorders from the National Center of Neurology and Psychiatry,the Takeda Science Foundation, and Mitsui Life Social Welfare Foundation.D.D. and D.B.F. were supported by Air Force Office of Scientific ResearchGrant FA 9550-14-1-0065.
1. Partch CL, Green CB, Takahashi JS (2014) Molecular architecture of the mammaliancircadian clock. Trends Cell Biol 24(2):90–99.
2. Daan S, et al. (2001) Assembling a clock for all seasons: Are there M and E oscillators inthe genes? J Biol Rhythms 16(2):105–116.
3. Inagaki N, Honma S, Ono D, Tanahashi Y, Honma K (2007) Separate oscillating cellgroups in mouse suprachiasmatic nucleus couple photoperiodically to the onset andend of daily activity. Proc Natl Acad Sci USA 104(18):7664–7669.
4. Schaap J, et al. (2003) Heterogeneity of rhythmic suprachiasmatic nucleus neurons:Implications for circadian waveform and photoperiodic encoding. Proc Natl Acad SciUSA 100(26):15994–15999.
5. Rohling J, Meijer JH, VanderLeest HT, Admiraal J (2006) Phase differences betweenSCN neurons and their role in photoperiodic encoding; a simulation of ensemblepatterns using recorded single unit electrical activity patterns. J Physiol Paris 100(5–6):261–270.
6. VanderLeest HT, et al. (2007) Seasonal encoding by the circadian pacemaker of theSCN. Curr Biol 17(5):468–473.
7. Brown TM, Piggins HD (2009) Spatiotemporal heterogeneity in the electrical activityof suprachiasmatic nuclei neurons and their response to photoperiod. J Biol Rhythms24(1):44–54.
8. Mrugala M, Zlomanczuk P, Jagota A, Schwartz WJ (2000) Rhythmic multiunit neuralactivity in slices of hamster suprachiasmatic nucleus reflect prior photoperiod. Am JPhysiol Regul Integr Comp Physiol 278(4):R987–R994.
9. Naito E, Watanabe T, Tei H, Yoshimura T, Ebihara S (2008) Reorganization of thesuprachiasmatic nucleus coding for day length. J Biol Rhythms 23(2):140–149.
10. Sumová A, Trávnícková Z, Peters R, Schwartz WJ, Illnerová H (1995) The rat supra-chiasmatic nucleus is a clock for all seasons. Proc Natl Acad Sci USA 92(17):7754–7758.
11. Foley NC, et al. (2011) Characterization of orderly spatiotemporal patterns of clockgene activation in mammalian suprachiasmatic nucleus. Eur J Neurosci 33(10):1851–1865.
12. Myung J, et al. (2012) Period coding of Bmal1 oscillators in the suprachiasmatic nu-cleus. J Neurosci 32(26):8900–8918.
13. Evans JA, Leise TL, Castanon-Cervantes O, Davidson AJ (2013) Dynamic interactionsmediated by nonredundant signaling mechanisms couple circadian clock neurons.Neuron 80(4):973–983.
14. Liu C, Weaver DR, Strogatz SH, Reppert SM (1997) Cellular construction of a circadianclock: Period determination in the suprachiasmatic nuclei. Cell 91(6):855–860.
15. Herzog ED, Aton SJ, Numano R, Sakaki Y, Tei H (2004) Temporal precision in themammalian circadian system: A reliable clock from less reliable neurons. J BiolRhythms 19(1):35–46.
16. Kuramoto Y (1984) Chemical Oscillations, Waves, and Turbulence (Springer, Berlin).17. Timme M (2007) Revealing network connectivity from response dynamics. Phys Rev
Lett 98(22):224101.18. Kralemann B, Cimponeriu L, Rosenblum M, Pikovsky A, Mrowka R (2008) Phase dy-
namics of coupled oscillators reconstructed from data. Phys Rev E Stat Nonlin SoftMatter Phys 77(6 Pt 2):066205.
19. Albus H, Vansteensel MJ, Michel S, Block GD, Meijer JH (2005) A GABAergic mecha-nism is necessary for coupling dissociable ventral and dorsal regional oscillators withinthe circadian clock. Curr Biol 15(10):886–893.
20. Choi HJ, et al. (2008) Excitatory actions of GABA in the suprachiasmatic nucleus.J Neurosci 28(21):5450–5459.
21. Meijer JH, Michel S, Vanderleest HT, Rohling JH (2010) Daily and seasonal adaptationof the circadian clock requires plasticity of the SCN neuronal network. Eur J Neurosci32(12):2143–2151.
22. Farajnia S, van Westering TL, Meijer JH, Michel S (2014) Seasonal induction of GA-BAergic excitation in the central mammalian clock. Proc Natl Acad Sci USA 111(26):9627–9632.
23. DeWoskin D, et al. (2015) Distinct roles for GABA across multiple time-scales in mammalian circadian timekeeping. Proc Natl Acad Sci USA 112:E3911–E3919.
24. Ciarleglio CM, Axley JC, Strauss BR, Gamble KL, McMahon DG (2011) Perinatal pho-toperiod imprints the circadian clock. Nat Neurosci 14(1):25–27.
25. Kelso JAS (1995) Dynamic Patterns: The Self-Organization of Brain and Behavior (MITPress, Cambridge, MA).
26. Pittendrigh CS, Daan S (1976) A functional analysis of circadian pacemakers in nocturnalrodents. V. Pacemaker structure: A clock for all seasons. J Comp Physiol 106:333–355.
27. Aton SJ, Huettner JE, Straume M, Herzog ED (2006) GABA and Gi/o differentiallycontrol circadian rhythms and synchrony in clock neurons. Proc Natl Acad Sci USA103(50):19188–19193.
28. Freeman GM, Jr, Krock RM, Aton SJ, Thaben P, Herzog ED (2013) GABA networksdestabilize genetic oscillations in the circadian pacemaker. Neuron 78(5):799–806.
29. Yamada J, et al. (2004) Cl- uptake promoting depolarizing GABA actions in immaturerat neocortical neurones is mediated by NKCC1. J Physiol 557(Pt 3):829–841.
30. Belenky MA, et al. (2010) Cell-type specific distribution of chloride transporters in therat suprachiasmatic nucleus. Neuroscience 165(4):1519–1537.
31. Kanaka C, et al. (2001) The differential expression patterns of messenger RNAs en-coding K-Cl cotransporters (KCC1,2) and Na-K-2Cl cotransporter (NKCC1) in the ratnervous system. Neuroscience 104(4):933–946.
32. Kahle KT, et al. (2005) WNK3 modulates transport of Cl- in and out of cells: Impli-cations for control of cell volume and neuronal excitability. Proc Natl Acad Sci USA102(46):16783–16788.
33. Diekman CO, et al. (2013) Causes and consequences of hyperexcitation in central clockneurons. PLOS Comput Biol 9(8):e1003196.
34. Kim JK, Forger DB (2012) A mechanism for robust circadian timekeeping via stoi-chiometric balance. Mol Syst Biol 8:630.
35. DeWoskin D, Geng W, Stinchcombe AR, Forger DB (2014) It is not the parts, but howthey interact that determines the behaviour of circadian rhythms across scales andorganisms. Interface Focus 4(3):20130076.
36. Daan S, Berde C (1978) Two coupled oscillators: Simulations of the circadian pace-maker in mammalian activity rhythms. J Theor Biol 70(3):297–313.
37. Kronauer RE (1987) Temporal subdivision of the circadian cycle. Lect Math Life Sci19:63–120.
38. Oda GA, Friesen WO (2002) A model for “splitting” of running-wheel activity inhamsters. J Biol Rhythms 17(1):76–88.
39. Aton SJ, Colwell CS, Harmar AJ, Waschek J, Herzog ED (2005) Vasoactive intestinalpolypeptide mediates circadian rhythmicity and synchrony in mammalian clock neu-rons. Nat Neurosci 8(4):476–483.
40. Wagner S, Castel M, Gainer H, Yarom Y (1997) GABA in the mammalian supra-chiasmatic nucleus and its role in diurnal rhythmicity. Nature 387(6633):598–603.
41. Liou SY, Albers HE (1990) Single unit response of neurons within the hamster su-prachiasmatic nucleus to GABA and low chloride perfusate during the day and night.Brain Res Bull 25(1):93–98.
42. Sumová A, Jác M, Sládek M, Sauman I, Illnerová H (2003) Clock gene daily profiles andtheir phase relationship in the rat suprachiasmatic nucleus are affected by photo-period. J Biol Rhythms 18(2):134–144.
43. Azzi A, et al. (2014) Circadian behavior is light-reprogrammed by plastic DNAmethylation. Nat Neurosci 17(3):377–382.
44. Iyer R, Wang TA, Gillette MU (2014) Circadian gating of neuronal functionality: Abasis for iterative metaplasticity. Front Syst Neurosci 8:164.
45. Yan L, et al. (2007) Exploring spatiotemporal organization of SCN circuits. Cold SpringHarb Symp Quant Biol 72:527–541.
46. Noguchi T, et al. (2010) Dual-color luciferase mouse directly demonstrates coupledexpression of two clock genes. Biochemistry 49(37):8053–8061.
47. Vandesompele J, et al. (2002) Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol 3(7):research0034–research0034.11.
48. Hoppensteadt FC, Izhikevich EM (1997) Weakly Connected Neural Networks (Springer,New York).
49. Hong JH, Jeong B, Min CH, Lee KJ (2012) Circadian waves of cytosolic calcium con-centration and long-range network connections in rat suprachiasmatic nucleus. Eur JNeurosci 35(9):1417–1425.
50. Fan J, et al. (2015) Vasoactive intestinal polypeptide (VIP)-expressing neurons in thesuprachiasmatic nucleus provide sparse GABAergic outputs to local neurons with
E3928 | www.pnas.org/cgi/doi/10.1073/pnas.1421200112 Myung et al.

circadian regulation occurring distal to the opening of postsynaptic GABAA iono-tropic receptors. J Neurosci 35(5):1905–1920.
51. Hastie T, Tibshirani R, Friedman J (2009) The Elements of Statistical Learning(Springer, New York), 2nd Ed.
52. Grant M, Boyd S, Ye Y (2008) CVX: Matlab software for disciplined convex pro-gramming. Available at http://cvxr.com/cvx/.
53. Friedman J, Hastie T, Tibshirani R (2010) Regularization paths for generalized linearmodels via coordinate descent. J Stat Softw 33(1):1–22.
54. Ermentrout GB, Terman DH (2010) Mathematical Foundations of Neuroscience(Springer, New York).
55. Kononenko NI, Dudek FE (2004) Mechanism of irregular firing of suprachiasmaticnucleus neurons in rat hypothalamic slices. J Neurophysiol 91(1):267–273.
Myung et al. PNAS | Published online June 30, 2015 | E3929
NEU
ROSC
IENCE
APP
LIED
MATH
EMATICS
PNASPL
US