from the departamento de fisicoquimica-facultad de farmacia
TRANSCRIPT

J. Phy8iol. (1967), 190, pp. 481-497 481With 6 text-ftgure8P'rinted in Great Britain
TRANSPORT AND DISTRIBUTION OF SODIUMACROSS FROG SKIN
BY M. CEREIJIDO AND CATALINA A. ROTUNNOFrom the Departamento de Fisicoquimica-Facultad de Farmacia
y Bioquitmica, Juntn 956, Buenos Aires, Argentina
(Received 7 October 1966)
INTRODUCTION
1. The time course of 22Na influx across frog skin mounted as a flatsheet between two lucite chambers has been studied. The flux reachesits maximal steady value in about 30 min.
2. The time course of changes in 22Na specific activity in the cells hasbeen followed by two different methods: (1) periodical measurements of22Na activity in pieces of skin mounted in a special device in which theouter facing membrane was in contact with the tracer and (2) measure-ment of uptake of 24Na in individual pieces of skin suspended in Ringersolution. Under both circumstances the skin failed to exchange all its Na.
3. Considerations on the basis of the kinetics of 22Na influx and 22Naspecific activity indicate that there exist at least two different Na compart-ments in the epithelium, one of them being directly involved in Natransport.
4. The 22Na specific activity profile was studied in skins which had pre-viously been mounted as a flat sheet between two chambers with tracer inone of them. This was carried out by removing the skin from the chamber;freezing, slicing and analysing the slices for Na and 22Na. The resultsindicate that both Na compartments are distributed across the wholeepithelium.
5. The results of these studies are taken to indicate: (a) that the Napartition in the tissue is not a result of the Na being contained in differentcellular layers; (b) that Na transport across frog skin is carried out by allthe epithelial cells and is not restricted to those of a particular layer;(c) that Na entry from an outer solution containing from 1 to 10 mM-Naoccurs into the transporting cells down an electrochemical potentialgradient and therefore need not involve an active mechanism at the outerborder of the cells.

M. CEREIJIDO AND CATALINA A. ROTUNNO
INTRODUCTION
Current models of Na movement and distribution in frog skin have threemain characteristics: (1) Na is supposed to be homogeneously distributedin a unique cellular compartment (Hoshiko & Ussing, 1960; Curran,Herrera & Flanigan, 1963; Ussing, 1964; Cereijido, Herrera, Flanigan &Curran, 1964; Ussing & Windhager, 1964); (2) Na is assumed to enter fromthe outside bathing solution into the cell compartment by a passivemechanism; and (3) the active step in Na net movement across the frogskin is assumed to be located at the boundary between the cells and theinside bathing solution. Yet recent studies by Rotunno, Pouchan &Cereijido (1966) have clearly demonstrated that skins with 1-10 mM-Naon the outside bathing solution and 97 mm-Na in the cells were able totransport Na. Since, under the conditions they used, the cells were onlysome 18 mV negative with respect to the outside bathing solution, the Naconcentration in the cells was not consistent with a passive entry mechanism.Thus Na seems to be actively transported at the outside-solution/cellboundary. However, they also carried out a study of.the distribution ofNa-K dependent (or ouabain sensitive) adenosintriphosphatase (Na-K-ATPase) across the epithelium and, on the basis that three sodium ions areactively transported per molecule of ATP split, they could not findenough enzyme at the outer layer of cells to account for the active trans-port. Therefore, to explain the high Na content of the epithelium, theysuggested that Na in the cell might be compartmentalized. The presentwork explores that possibility. It will be shown that only 37 % of the Nain the epithelium is directly involved in the active transport.
METHODS
All studies were performed in vitro on the abdominal skin of specimens of either sex of theSouth American Frog Leptodactylua ocellatuw (L., 1758) during all seasons over a period of2 years. Animals were kept in a moist sink and periods in captivity never exceeded twomonths. Experiments were carried out at room temperature (20-22o C). Electrjeal potentialdifference was measured by connecting the chambers through agar-Ringer bridges to calomelhalf cells and these cells to a Keithley 200B DC electrometer (imput impedance, 10'4Q).Current was passed using Ag-AgCl electrodes and was measured with a Simpson micro-ammeter. Whenever possible the preparation was kept short-circuited.
Preparation of the 8olution8. Sulphate-Ringer solution contained 2-5-5-0 mM-Na2SO,2-4 mM-KHCO3, 1-2 mM-CaSO4 and 146 mm sucrose. Chloride-Ringer contained 10 mM-NaCl, 2-4 mM-KHCO3, 1 mM-CaCl2 and 204 mm sucrose. All reagents used were of Analargrade. After the solutions were gassed with air they gave a pH of 8-2. Low-Na Ringer wasused in order to reduce the amount of Na in the extracellular space and therefore minimizethe errors in the estimation of intracellular Na.Na analysis. The piece of skin was placed in polystyrene test tube with 3-0 ml. of 0-1
482

SODIUM IN FROG SKIN 483N-HNO3 stoppered with Parafilm (Marathon) paper and extracted for 48-96 hr. Aliquotswere then diluted and Na was measured by flame photometry (Crudo-Caamanio flame photo-meter, Buenos Aires). Calibration curves, obtained with 'standard solutions of Na alsocontainingK at a concentration 100 times higher than in the samples of tissue, were identicalwith those obtained in absence of K.
Inulin spaces. Pieces of skin were immersed in Ringer solution containing 0 04 /tc/ml. of[14C]inulin (New England, Nuclear) for at least 40 min. They were then removed, blotted andtreated as described in Na analy8si. Duplicate samples of bathing solution and extractionfluid were placed in planchets. In those experiments in which [14C]inulin was used 24Na wasused instead of 22Na. After allowing sufficient time for 24Na to decay, the planchets werecounted in a windowless gas flow counter and the 14C activity was compared with that inthe bathing solution in order to estimate the extracellular space. The average estimate ofextracellular space was 15 9 + 0 9 /ul. cm-2 (52) [mean + S.E. of mean (number of observa-tions)]. This figure agreed satisfactorily with the extracellular space 13- 6 + 0-9 1d. cm-2 (34)obtained with preparations in which the frog skin was mounted as a flat sheet between twolucite chambers and [14C]inulin was added to the inside bathing solution. When [14C]inulinwas added only to the outside solution virtually no radioactivity was found in the skin. Thefact that the inulin spaces did not differ significantly when both sides of the skin werebathed in a solution of inulin and when inulin was added only to the inside bathing solutionis also in agreement with electronmicroscope pictures of frog skin (R. pipiens) obtained byFarquhar & Palade (1964) indicating that extracellular spaces are confined to the corion anda narrow intercellular space of the epithelium.
22Na flux. The skin was rinsed in sulphate-Ringer solution and then mounted as a flat sheetbetween two lucite chambers of 3-14 cm2 area. The volume of each chamber was 5 0 ml.After allowing '40 min for equilibration, 1*5 ,uc of 22Na was added to the outside chamber.Duplicate samples of 100 ,d. were withdrawn periodically from the inside bathing solutionand the volume replaced with fresh Ringer. At the end of the experiment, samples weretaken from the outside bathing solution and analysed for Na and 22Na.Time course of the 22Na specific activity in the cells. The assumption was made that the
preparation was in steady state and therefore Na content of the tissue was constant. Thisseems justifiable because the Na content in the epithelial cells of this skin at the end ofexperiment was found to be 0 074 + 0 004 (13) ,umole/mg dry weight of membrane whichcompares with 0 071 + 0-004 (16) ,umole/mg, the value found in control skins. Hence, thecounts/min measured at any given moment divided by the Na content of the epitheliummeasured at the end of the experiment were taken as a measure of the 22Na specific activityof the tissue. The arrangement used to measure the time course of 22Na activity in the frogskin is shown in Fig. 1. It consists of a cylindrical lucite holder attached to a flat plate topermit reproducible positioning of the preparation both in the loading solution and in thecounter. The skin was tied in place with the outer side facing outward by thread fitted intoa groove machined in the cylinder at 0-2mm from the lower end. The edge of the skin down tothe border and part of the side of the cylinder were painted with collodion (Parlodion,Merck) dissolved in acetone. The exposed area was 4 79 cm2. The skin was placed in abeaker with 30 ml. sulphate-Ringer solution stirred by a magnetic stirrer. A constant flow ofRinger solution was passed through the inside chamber. The diameter of the cannulae, thespeed of the inflow and the suction were so adjusted as to produce a constant flow of 5-0 ml./min and keep the inside bathing solution volume at 1-5 ml. The preparation was alwaysequilibrated for at least 40 min before 10 'aC of 22Na was added to the outside bathing solu-tion. The skin remained in the loading solution for 10 min, after which it was removed andblotted carefully with soft paper and the whole assembly placed in a fixed position above thecrystal detector. It was counted for 1 min. The loading and counting periods were repeated9 times. At the end of the experiment the exposed part of the skin was cut out and its Nacontent determined. The holder with the edge of the skin covered by the collodion wascounted. This value was subtracted from those of all previous measurements. This proce-

484 M. CEREIJIDO AND CATALINA A. ROTUNNOdure insured that the counts/min used in the calculation of the 22Na specific activity werethose belonging to Na in the transporting tissue. In order to measure the radioactivity ofthe loading solution with the same geometry, the rest of the skin was then removed from theholder, replaced by a piece of Parafilm paper and samples of 100 1tl. of loading solution putinto the cylinder and counted. Samples of the loading solution were also analysed for Na. Theamount of Na in the cells was obtained by difference between the total Na in the tissue andin the corion. Because of the big volume of Ringer used on the inside no [14C]inulin wasadded and the Na content in the extracellular space was not measured but calculated fromthe measured Na concentration in the Ringer (10 mM) and the average value of the extra-
Compressed air id
-l_ ,.To suction pump
- t ~~~Skin
CrystalSti rrer detector
Fig. 1. Arrangement for measuring the time course of the radioactivity of 22Na inthe skin. On the left the skin is in contact with the loading solution; after 10 minit is removed, blotted and placed in a fixed position on a scintillation counter for1 min. The whole procedure is repeated 9 times. At the end of the experiment theNa content of the exposed skin is determined. The same assembly shown on theright but with a piece of Parafilm paper replacing the skin is used to measure theradioactivity of 22Na in samples of the loading solution.
cellular space (13-6 ,u. cm-2) mentioned before. This yields about 0-14 ,umoles of Na. Thisamount is considerably smaller than that in the tissue (0 4-0 6 jctmole; see also Rotunnoet al. 1966) and therefore any variation of the Na content of the extracellular space should notsignificantly alter the computation of Na in the cells. This method is similar in several re-spects to the one used by Curran & Cereijido (1965) for K flux measurements.
24Na uptake. The skin was placed on a filter paper moistened in sulphate-Ringer with theouter side in contact with the paper. Eight to twelve pieces of 1-16 cm2 were cut out with acork borer and put into a beaker with 100 ml. Ringer gassed with air and stirred with amagnetic stirrer. Fifty-two pieces of skin from five frogs were processed together in the same

SODIUM IN FROG SKIN 485
experiment. After all the pieces were obtained (40 min) they were removed to a new beakercontaining 100 ml. of fresh Ringer with [14C]inulin and allowed to equilibrate with the tracerfor 40 min. Then 50 gc of 24Na was added to the bathing medium. Pieces of skin were re-moved periodically, placed on filter paper, carefully blotted, and put into polystyrene testtubes, and the 24Na activity was counted. At the end of the experiment samples of bathingsolution were withdrawn for 24Na, Na and [14C]inulin measurements. Once the pieces of skinwere counted, 3-0 ml. of 0-1N-HNO3 was added, the tubes were stoppered with Parafilmpaper and their content of Na and [14C]inulin measured. With these data the 24Na specificactivity of the cells and that of the loading solution were calculated.The assumption is made that when a flux of tracer reaches the steady-state level the
specific activity in the compartment from which the flux is coming should be constant (seeSolomon, 1960). Figure 6a shows the model generally used in studying the kinetics of Natracer movements across epithelial membranes (Schoffeniels, 1957; Leaf, 1958; Hoshiko &Ussing, 1960; Curran et al. 1963; Solomon, 1964; Cereijido et al. 1964 and Herrera, 1965).Compartment 2 is taken to represent the compartment ofNa involved in Na transport acrossthe frog skin. When 22Na is added to the outside bathing solution, the variations in theamount of tracer in compartment 2 and the inside bathing solution are given by the followingequations:
dP2/dt =Jl2P*1-P*2 (J21+ J23) +J32P*3 (1)
dP3/dt= J23P*2 -.J32P*3, (2)Pi = total activity of 22Na in compartment i (counts/min);Jij = Na flux from compartment i to j (4amoles.hr-1. cm-2);P*i = 22Na specific activity in compartment i (counts/min.,umole).Since the preparation is assumed to be in a steady state, the Na fluxes are constant. Dur-
ing the length of the experimental period (90 min maximal), p*1 remains constant and p*3may be taken as zero. These assumptions make p*2 the only variable on the right-hand sideof equations (1) and (2). dP3/dt is the rate of tracer appearance in compartment 3; when itreaches a constant value (dP2/dt)q,, it implies that p*2 is also constant. The aim of the firstset of experiments was to determine the moment at which this level is reached. The value ofthe influx of Na may be obtained by the equation
(dP3fdt),n(3J13 = pg * (3)
Slicing. The skins were mounted as a flat sheet between lucite chambers as describedabove. After 40-50 min of equilibration in the desired Ringer, 5 ,uc of 22Na was added to oneof the chambers. Samples of Ringer in the hot side were taken for Na and 22Na measurements.After a loading period, the skin was removed from the chamber, placed on a filter paper withthe inner side facing the paper, and blotted with soft tissue on the outer side. A piece of skinof 1-16 cm2 of the area exposed in the chamber was cut out with a cork borer and placed onthe stage of a freezing microtome (Jung Quick-Freeze Microtome) and frozen. The time fromthe removal of the skin from the loading chamber until it was frozen was usually 40 sec andnever more than 60 sec. The stage of the microtome was covered beforehand with a layerof 4 g/100 ml. agar which was frozen and cut to a thickness of about 1 mm. The knife wascooled both with a jet of CO2 and with dry ice placed on a basket on top of the knife. Sliceswere removed from the knife with a sharp needle made of stainless steel and were placed ona piece of aluminium paper of about 1 cm2. With this procedure, the slices were continuouslyfrozen until they were on the aluminium paper. This procedure minimizes the risk of loss orcontamination of the different layers when in contact with the knife. The piece of aluminiumpaper in which the tissue was placed was washed with distilled water and tared in a Mettlerultramicrobalance with an error of 2 ,ug. The slices were dried to constant weight at 90° C.The dry weight of the slices was usually between 50 and 300 ,ug. Those closer to the outerside were made thinner, since more discrimination in this region was desired. It was possible
31I Physiol. I90

486 M. CEREIJIDO AND CATALINA A. ROTUNNOto obtain 8-15 slices before connective tissue was reached. This was easily noted because ofthe sudden change in colour. Fortunately the difference between the dark epithelium and thewhite connective tissue was maximal at the junction level. Once the connective tissueappeared, the slicing was stopped and the remaining skin was put on aluminium paper.The connective tissue usually showed up partially (i.e. it was clearly reached in onearea while the other was still dark). If in the next slice the connective tissue also showedup partially it was taken as indication that the slices were not parallel to the outer borderand the whole experiment was discarded. In preliminary experiments, it was noted thatthis situation arose when the lateral part of the abdominal skin was used. Accordinglyspecial care was taken to use the central part of the skin in which the thickness wasmore uniform. After drying and weighing, the slices were analysed for Na and 22Na. Inorder to compare results obtained with different skins a parameter (normalized depth) wasdefined: the weight of a given slice was added to all previous slices (that is, between the outerborder and the given slice) and this value was divided by the sum of all individual slicesplus the remaining connective tissue from the same piece of skin. Since the amount of wateris probably larger in the connective tissue than in the epithelium, the 'normalized depth'would not correspond exactly with the actual location of slices. A slice of the connectivetissue of a given dry weight will actually correspond to a larger volume of fresh tissue thanone in the epithelium and this artifact will tend to increase the apparent Na content andcounts/min content of the connective tissue. Therefore conclusions on the basis of the valuesobtained on the connective tissue side will be avoided. Nevertheless, this problem should notinvalidate comparisons, for instance between two 22Na activity profiles or between a 22Naactivity profile and a profile of Na content.
RESULTS
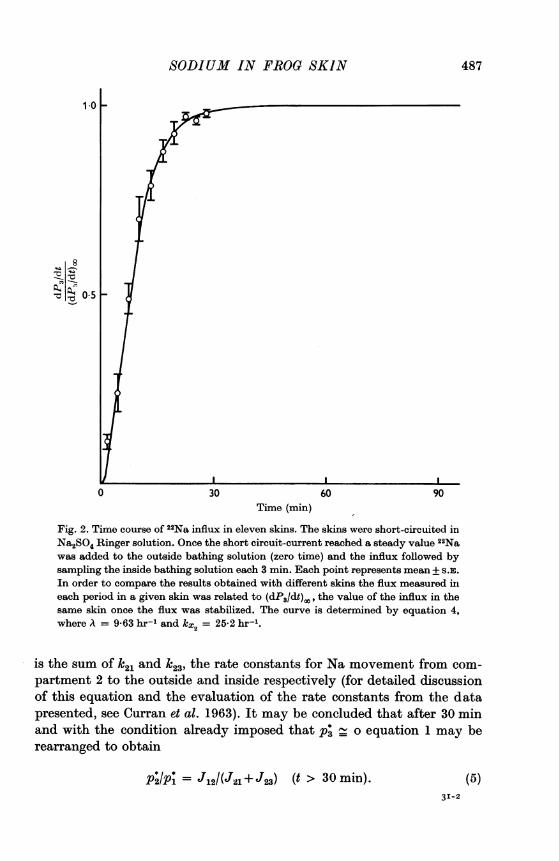
Time course of the 22Na influx. Studies of 22Na influxes carried out inR. pipiens (Curran et al. 1963; Cereijido et al. 1964) indicated that thesteady-state flux of Na tracer was reached in about 10-20 min. In pre-liminary studies with L. ocellatus, it was observed that the flux was alreadyin equilibrium by 30 min and continued stabilized during at least 90 min.In eleven experiments in which the skins were mounted in sulphate-Ringer solution, the time course of 22Na appearance in compartment 3was followed by sampling this compartment every 3 min. The results areshown in Fig. 2. The maximal influx of tracer is reached in about 30 min.The influx of Na was 0x24 + 0*04 #tmoles . hr-1 . cm-2 (11). It demonstratesthat the low concentration used to minimize errors in cell Na measure-ments does not alter the ion movement across the skin. This agrees withprevious findings of Curran & Cereijido (1965) indicating that the skinsustains active transport of Na and K when bathed in solutions with5 m-MNa on the inside. The line in Fig. 2 was calculated with an equationof the form
dt (dtP [-k-Axe-A+ A e-kx3t], (4)
where t is the time in hours after adding the tracer; kx3 is the rate constantfor Na movement from the connective tissue to the compartment 3 and A
SODIUM IN FROG SKIN
1.0
8* 05
2-_
0
487
30 60 90Time (min)
Fig. 2. Time course of 22Na influx in eleven skins. The skins were short-circuited inNa2SO4 Ringer solution. Once the short circuit-current reached a steady value 22Nawas added to the outside bathing solution (zero time) and the influx followed bysampling the inside bathing solution each 3 min. Each point represents mean + S.E.In order to compare the results obtained with different skins the flux measured ineach period in a given skin was related to (dP3/dt),,, the value of the influx in thesame skin once the flux was stabilized. The curve is determined by equation 4,where A = 9-63 hr-1 and kxc2 = 25-2 hr'1.
is the sum of k2l and k23, the rate constants for Na movement from com-partment 2 to the outside and inside respectively (for detailed discussionof this equation and the evaluation of the rate constants from the datapresented, see Curran et al. 1963). It may be concluded that after 30 minand with the condition already imposed that p* o equation 1 may berearranged to obtain
P2I/P = J12/(J21+ J23) (t > 30 min). (5)3I-2

488 M. CEREIJIDO AND CATALINA A. ROTUNNOSince the fluxes of Na are in steady state the following assumption shouldhold
J12 + J32 = J21+ J23- (6)
The inner membrane of the transporting cells has a very low Na permea-bility (Koefoed-Johnsen & Ussing, 1958; MacRobbie & Ussing, 1961);therefore J32 should have a very low value as compared with the otherthree fluxes in equation (6). Accordingly the relationship p2/p; given byequation (3) should have a value close to 1.Time course of the 22Na specific activity in the cells (p*). Compartment c
includes all the Na in the cells. Compartment 2 contains only the cell Nainvolved in Na transport. If all the Na in the cell were involved, com-partments c and 2 would be equal. Table 1 shows the values of pc/p;
TABLE 1. Time course of 22Na specific activity in the skin as measured with thetechnique depicted in Fig. 1 (mean+ S.E.; n = 13)
Time after adding22Na (min) (p*C/p*) 100
10 2-25+*04320 4-02 + 0*5830 5'34+ 0-8640 5-83+0.9150 6-71 + 0*9760 7*74+1*0670 8*41+1.1080 9-16+1.0090 9 69+ 1*20
obtained in 13 experiments (9 periods of 10 min in each experiment). Thefact that the value of pc/p; reached at 30 min is far below 1.0 may indicateeither that only a fraction of Na in the epithelium is involved in Na trans-port or Na fluxes across the inner barrier are higher than assumed. IfJ32were different from zero, Na fluxes across the inner barrier would washthe 22Na out of the cells and prevent its specific activity from reaching thevalue of this parameter in the outer solution. A decrease of these fluxeswith time (one of them J23, includes a component which depends onmetabolic energy) may conceivably explain the slow increase of pc/p;observed after 30 min. 24Na uptake was studied to test the possibility thatthe low p*/pl ratio was due to a high Na exchange across the inner barrier.24Na uptake. Although in this experiment the pieces of skin were not
mounted between two chambers but suspended in the loading solution thesame model of Fig. 6a and therefore equation (1) may be used. This timep3 is no longer negligible but is equal to p;. After the tracer distributionreaches steady state, the relationship p2/p; should be given by
P2VP; = (J12 + J32) / (J21 + J23) ((7)

SODIUM IN FROG SKIN
With steady-state condition (equation (6)) it follows that, once Na specificactivity in the epithelium achieves a constant value, the relationship
P = p* (t > 30 min) (8)
should hold. Figure 3 shows the time course of pc. It may be described bythe equation
P= 037p; (l-e-103). (9)
As might be expected on the basis of the studies of 22Na flux discussedabove, the tracer specific activity in the cell compartment equilibrates
15,000
_ PC10
._
A 10,000
0
o 0
c)
0 30 60 90Timne (min)
Fig. 3. Time course of 24Na specific activity in the cell compartment, p*, and in theloading solution, pl. The pieces of skin were suspended in Na2SO4 Ringer. After aperiod of equilibration 24Na was added to the bathing medium so that it waspresent on both sides of the skin. The curve is determined by equation (9). Eachpoint before 20 min represents a mean value of two pieces of skin. Those after 2 minare means of four determinations.
in less than 30 min. With the condition imposed by equation (8) it may beseen that
Pc = 037p* (t > 30 min). (10)
This means that only 37 % of the Na in the epithelium exchanges with thatin the loading solution. It also means that 63% is not directly involved in
489

M. CEREIJIDO AND CATALINA A. ROTUNNO
Na transport across the skin. However, on the basis of these results andthose of the previous section one cannot discriminate between the follow-ing possibilities: (a) Only 5-6% of the total Na is involved in Na transportand this fraction reaches the same specific activity as the outer loadingsolution; the remaining 31-32 % constitutes a third compartment thatcan be reached from the inside only but does not participate in Na trans-port across the skin. (b) Only 37 % of the total Na in the cells is involvedin Na transport but because it is washed toward the inner bathing solutionthe 22Na specific activity in the cells is only 5-6% of the specific activityin the outer solution.
TABLE 2. Profile of the Na content. The content was measured in slices of skin parallel to theouter surface. The skins had been previously mounted as a flat sheet between two chamberswith identical sulphate-Ringer solutions. The zero level of the depth corresponds to theouter anatomical border
Na contentDepth (%) (,moles/mg) dry weight
5 0-18710 0 15615 0-15220 0.15530 0-09240 0-099
Localization of the compartment responsible for Na transport. To obtainsome information on the characteristics of this compartment, the profileof the specific activity of the Na in the tissue was studied with a techniqueof the type described under Slicing (see Methods) in which sulphate-Ringer solution was used on both sides. 22Na tracer was added to the out-side bathing solution and the skin removed from the chamber at 40 min,that is, after a period longer than it takes for the influx of the tracer toreach the steady state value. Since the Na content/mg dry weight isaround 0-1 ,tmole and the dry weight of each slice was between 50 and300,g, determination of the Na content of each slice involved a consider-able error. To minimize this the following procedure was adopted: after22Na was counted in each individual slice, the slices belonging to a givenrange of normalized depth were pooled and the Na content was measured.Table 2 shows the profile of Na content of twenty-five skins. It clearlyshows a drop in Na content between the 20 and 30% of depth. On thebasis of the relationship of total water/extracellular water, Rotunno et al.(1966) estimated that the cells occupy the outer 28% of the total thick-ness. Therefore the drop in Na content may occur presumably near theepithelial/connective tissue boundary, which agrees with the high Nacontent of the cells and the low Na concentration of the Ringer. Thespecific activity of the 22Na of each slice (pc) was obtained by dividing thecounts/min of 22Na by the Na content corresponding to its normalized
490

SODIUM IN FROG SKIN
depth. This value was in turn divided by p;, the specific activity of 22Nain the loading solution, and plotted. The ordinate represented pc/p; andthe abscissa represented normalized depth. The area under the curve wasthen computed at intervals of 0-2 5; 2-5-5-0; 5 0-7 5; 7 5-10; 10-15;15-20; 20-25 and 25-30% of normalized depth. This distribution waschosen so as to include at least one individual measurement of each skinin each interval and, as said in Methods, it reflects the purpose of gettingmore and thinner slices (higher discrimination) near the outside-solution/cell boundary. Figure 4 summarizes the results obtained with twelve skins.Three facts may be pointed out: (1) The method does not involve gross
0.05
0 10 20 30 40 50
Normalized depth
Fig. 4. Profile of the 22Na specific activity. The skins were mounted as a flat sheetbetween two chambers with Na2SO4 Ringer solution and short-circuited. Afterallowing the skins to be 40 min in contact with 22Na on the outside they were re-moved from the chamber, frozen and sliced. Ordinate represents specific activityof the slices as fraction of this parameter in the outside solution. The zero inabscissa represents the outer anatomical border of the skin.
contaminations since the first set of slices which has from 0 to 2-5% of thetotal thickness has less than 4% of the specific activity of the loadingsolution which was in immediate contact with it. (2) The values of 22Naspecific activity are evenly distributed over a distance presumably coveredby the epithelium. (3) The 22Na specific activity in the cells reaches only5% of the value in the loading solution. This value compares remarkablywell with 5-8 %, the value obtained for the same loading period (40 min)
491

M. CEREIJIDO AND CATALINA R. ROTUNNO
in the experiments described in Table 1 which involved a completelydifferent technique.Another set of experiments was carried out in which two halves of the
same skin were mounted in chloride-Ringer on both sides. After the usualperiod of equilibration 5 4uc of 22Na was added to the outer bathing solution
0 30 _-
(a)
0 00
0 20 _-
010
0o0
8 9 0:-6.----o---- 0- 0 °0
8 08 81 1I-
(b)
0 30 1-
0
0 20
0 10
0o0
~~~~000 -.06-
=___0-- t
00
0
p 1 1
0 30 60 90Time (min)
Fig. 5. 22Na specific activity in the epithelium of paired skins mounted in NaClRinger solution, short-circuited and loaded from the outside (a) or the inside (b).Each point in each graph is an individual measurement. Straight lines were drawnwith equations (11) and (12), respectively.
of one half of the skin and to the inner bathing solution of the other half.Both skins were withdrawn from the chamber after the same period ofexposure to the tracer (5, 15, 30, 60 or 90 min). In this set of experimentsonly the outermost slices were taken and those belonging to the same
piece of skins put together in the same aluminum paper. This procedurewas adopted to avoid the risk of taking part of the connective tissue with
492

SODIUM IN FROG SKINhigh 22Na activity in the pieces of skin loaded from the inside. Accordinglythe slices reported in this section never exceeded 15% of normalized depthwhich is nearly a half of the epithelium. The reason for using chloride-Ringer solution instead of sulphate-Ringer was to check the possibilitythat the low 22Na exchange of the epithelial cells was at least in part dueto restrictions imposed by the sulphate impermeability of the membrane.Figure 5a shows the results obtained with skins loaded from the outsideand 5b those with inside loading. Each point represents an individualmeasurement. There are thirty points on each side of the figure. Thestraight lines were obtained by the least square method using only thepoints at 30, 60 and 90 min, i.e. those in which the Na transporting com-partment is expected to be in equilibrium with the influx of tracer when itis added to the outside bathing solution. The straight line of the loadingfrom the outside is described by the equation
y = O000t+0115 (11)
and the one for the inside loading by
y = 0'001t+0'087. (12)
The fact that in the skins loaded from the inside the points at 5 min are soclose to the straight line shows that the low value of 22Na specific activityat 30, 60 or 90 min may not be ascribed to restrictions to 22Na diffusionimposed by the connective tissue and the innermost epithelial cell layers.The values of the intercepts (1I l5 and 8&7 % of the total), indicate thatloading either from the outer or the inner side the same proportion of Nain the cells is interchanged. Since in the epithelium loaded from the insidethe specific activity of 22Na does not reach a 30 % of the value of thisparameter in the loading solution, the difference between the 4-10%reached at 30 min when loading from the outside (Table 1 and Figs. 4and 5a) and the 35% when loading from both sides (Fig. 3) may not beascribed to the existence of a subcompartment which may only be reachedfrom the inside and which contains 31-32% of the total Na. The compari-son of the values of pt/pl reached in sulphate- and chloride-Ringer solutionsuggests that, even though values obtained in chloride-Ringer are slightlyhigher, the phenomenon is not significantly affected by the anion.
DISCUSSION
The main purpose of the present investigation was to discover whetherthe Na in the epithelium ofthe frog skin is compartmentalized. The generalconclusion from these results is that Na is contained in at least two differentcompartments, one of them being directly involved in active transport.
493

494 21I. CEREIJIDO AND CATALINA A. ROTUNNO
For the purpose of discussion this compartment will be referred to as thetransporting compartment and the rest of the cellular Na as the non-transporting compartment.
Localization of the Na compartments. Figure 6b shows very schematicallythe model proposed by Ussing (1964) to account for membrane phenomenain frog skin. Cells are distributed in several layers and connected to eachother through low resistance paths (Ussing & Windhager, 1964). One maywonder where the two different Na compartments may le located. There
Outside Inside
(a) bathing 1 2 3 bathingsolution solution
(b) 12I 2U . 3
Shunts
2.:(c) 1 3
Fig. 6. Different models to account for Na distribution and movement across epi-thelial membranes. In (a) Na is assumed to be homogeneously distributed in aunique cellular compartment. In (b) Na is also assumed to be homogeneously dis-tributed in a unique cellular compartment but because of the high value of therelationship flux-to-the-interspace/flux-through-the-shunts, Na in the cells of theinner layers behaves as a non-transporting compartment. In model (c) it is assumedthat every cell has both, Na transporting and non-transporting compartments; thebold line of the models represents the inner-facing membrane (see text).
is strong evidence indicating that active mechanisms for Na and Ktransport are located along the cell-interspace borders (see Rotunno et al.1966). Since Na pumping mechanisms are oriented toward the insidebathing solution, Na leaking into the cells either from the outer bathingsolution or from the interspace will be actively extruded toward theinterspace. Na ions coming to the cells of the outer layer will follow mainly

SODIUM IN FROG SKIN
two routes: (1) some of them will be actively transported to the inter-space; (2) others will pass to inner layers to be, in turn, pumped towardthe interspace. Therefore the contribution of the inner layers to the netactive transport from compartment 1 to compartment 3 will be limited bythe diffusion of Na through the bridges. If the amount of Na that thefirst layer of cells pumps into the interspace were much greater than theamount of Na that leaves these cells via the bridges, most of the Na nettransport would be carried out by the first layer. If 22Na is added to theouter solution, by the time the influx reaches a quasi-steady-state level thefirst layer of cells will have its 22Na specific activity in equilibrium, but the22Na specific activity in the inner cells would be comparatively low. Thiswould suggest that the first layer would represent the transporting com-partment and the rest of the epithelium (compartment 2') the non-transporting one. But this possibility may be discarded since the profileof 22Na specific activity (Fig. 4) indicates that even the outermost celllayer has a masking compartment of the same magnitude as the rest of thecells. This fact, together with the observation that the exchange isessentially completed in 10 min regardless of whether the 22Na comes fromthe compartment 1 or 3 (see Fig. 5) leads to the conclusion that thetransporting compartment as well as the non-transporting compartmentis distributed uniformly throughout the whole epithelium as suggested inFig. 6c. This is a very well known fact in other tissues. Thus Harris &Steinbach (1956) reported that only a fraction of Na in the frog skeletalmuscle is readily exchangeable with 24Na in the loading medium. However,they ascribed this non-exchanging compartment to the connective tissuerather than to the muscle fibres. A clearer example of two different Nacompartments inside the cells was provided by the studies of Gold andSolomon (1955) and Harris (1964) working with red cells.The nature of the Na compartments. There is at present no indication to
suppose that Na within the cell is contained in two physical compartments,one of them being almost impermeable to Na. It is more likely that theyare chemical compartments, i.e. Na contained in the same regions but intwo different states. Thus one may envisage a situation in which only 37 %of the Na in the cell was in the free ionized form and the rest was bound.This view agrees with the evidence obtained by MacRobbie & Ussing (1961)that in sulphate-Ringer solution only 21 ,u of the 58 ,t epidermis is osmo-tically active (that is, 36 %). Cereijido & Curran (1965) demonstratedthat, under short-circuited conditions, the cells of the skin of R. pipiensare some 18 mV negative with respect to the outside bathing solution.Hoshiko (1961) reports -25 mV under the same conditions; Whittembury(1964) -25 mV in toad skin. Unpublished observations in L. ocellatusshowed values ranging from -12 to -40 mV. Even if Na enters by a
495

M. CEREIJIDO AND CATALINA A. ROTUNNO
passive mechanism from an outside solution with 5-10 mM-Na, the Naconcentration of the cells may be as high as 49 mm. Na content in theoutermost set of slices (Table 2) was 0-187 ,umole/mg dry weight. If only37% of Na were free, there would be 0-069 ,umole of Na+/mg dry weight.Since the amount of water in the cells is 2-589 ,ul/mg of dry weight (Rotun-no et al. 1966) the Na concentration in the transporting compartmentwould be around 26 7 mm. Thus Na enters from a solution with a con-centration 5-10 mm into cells with 26-7 mm. Na compartmentalizationwould provide a consistent explanation of Na penetration through a pas-sive mechanism across the outer border despite the high Na content of thecells. Compartmentalization also offers an explanation of the somewhatpuzzling finding of Rotunno et al. already mentioned in the Introduction,that, on the basis of three sodium ions being actively transported permolecule of ATP split, there is not Na-K-ATPase enough at the outerborder to account for the active transport at that level, and the enzymecontained in almost the whole epithelium would be needed. If only onethird of the Na in the cells is freely exchangeable the high Na content ofthe cells does not necessarily involve an active mechanism at the outerborder when the skin is bathed in solution with low Na concentrations.
The authors wish to express their sincere thanks to Dr D. S. Parsons and Dr P. F. Curranfor their valuable criticism of this work and helpful suggestions, and to Miss Marisa Battellifor her competent technical assistance. This work was supported in part by grants from theConsejo Nacional de Investigaciones Cientificas y T6cnicas de la Repuiblica Argentina and thePublic Health Service of U.S.A.
REFERENCES
CEREIJIDO, M. & CURRAN, P. F. (1965). Intracellular electrical potentials in frog skin.J. gen. Phy8iol. 48, 543-557.
CEREIJIDO, M., HERRERA, F. C., FLANIGAN, W. J. & CURRAN, P. F. (1964). The influence ofNa concentration on Na transport across frog skin. J. gen. Phy8iol. 47, 879-893.
CURRAN, P. F., HERRERA, F. C. & FLANIGAN, W. J. (1963). The effect of Ca and antidiuretichormone on Na transport across frog skin. II. Sites and mechanisms of action. J. gen.Phy8iol. 46, 1011-1027.
CURRAN, P. F. & CEREIJIDO, M. (1965). K fluxes in frog skin. J. gen. Physiol. 48, 1011-1033.
FARQUHAR, M. G. & PALADE, G. E. (1964). Functional organization of amphibian skin.Proc. nat. Acad. Sci. U.S.A. 51, 569-577.
GOLD, G. L. & SOLOMON, A. K. (1955), The transport of Na into human erythrocytes invivo. J. gen. Physiol. 38, 389-404.
HARRIS, E. J. (1964). External Na concentration and erythrocyte Na turnover. J. Physiol.172, 61-73.
HARRIS, E. J. & STEINBACH, H. B. (1956). The extraction of ions from muscle by water andsugar solution with a study of the degree of exchange with tracer of the Na and K inthe extracts. J. Physiol. 133, 385-401.
HERRERA, F. C. (1965). Effect of insulin on short-circuited current and sodium transportacross toad urinary bladder. Am. J. Physiol. 209, 819-824.
HOSHIKO, T. (1961). Electrogenesis in frog skin. In Biophysics of Physiological andPharma-cological Actions. Washington, D.C.: American Association for the Advancement ofScience.
496

SODIUM IN FROG SKIN 497HOSHIoO, T. & UssING, H. H. (1960). The kinetics of 24Na flux across amphibian skin and
bladder. Acta vhysiol. scand. 34. 74-81.KOEFOED-JOHNSEN, V. & USSING, H. H. (1958). The nature of the frog skin potential. Acta
physiol. scand. 42, 298-308.LEAF, A. (1958). Measurements of the permeability of the two surfaces of a living membrane.
Science, N.Y. 128, 144-145.MACROBBIE, ENID A. C. & USSING, H. H. (1961). Osmotic behaviour of the epithelial cells
of frog skin. Acta physiol. scand. 53, 348-365.ROTUNNO, CATALINA, A., POUCHAN, MARIA, I. & CEREIJIDO, M. (1966). Location of themechanism of active transport of sodium across the frog skin. Nature,'Lond. 210, 597-599.
SCHOFFENIELS, F. (1957). An isolated single electroplax preparation. II. Improved prepara-tion for studying ion flux. Biochim. biophys. Acta 26, 585-596.
SOLOMON, A. K. (1960). Compartmental methods of kinetic analysis. In Mineral Metabolism,ed. COMAR, C. L. & BRONNER, F. London: Academic Press.
SOLOMON, A. K. (1964). Validity of tracer measurements of fluxes in kidney tubules andother three compartment systems. In Transcellular Membrane Potentials and Ionic Fluxes,ed. SNELL, F. M. & NOELL, W. K. London: Gordon and Breach.
UssING, H. H. (1964). Transport of electrolytes and water across epithelia. Harvey Lect.UsSING, H. H. & WINDHAGEIL, E. E. (1964). Nature of shunt path and active sodium trans-
port path through frog skin epithelium. Acta physiol. scand. 61, 484-504.WHITTEMBURY, G. (1964). Electrical potential profile of the toad skin epithelium. J. gen.
Physiol. 47, 795-808.