fish lymphocytes differ in the expression of surface immunoglobulin
TRANSCRIPT

DEVELOPMENTAL AND COMPARATIVE IMMUNOLOGY. Vol. 61 pp. 473-479, 1982. 0145-305X/82/030473-07$03.00/0 Printed in the USA. Copyright (c) 1982 Pergamon Press Ltd. All rights reserved.
FISH LYMPHOCYTES DIFFER IN THE EXPRESSION OF SURFACE IMMUNOGLOBULIN!
Craig J, Lobb and L. William Clem Department of Microbiology
University of Uississippi Uedical Center Jackson, Mississippi 39216
ABSTRACT. Catfish peripheral blood and splenic lymphocytes were assayed
1.
for surface immunoglobulin using fifteen different mouse hybridoma antibodies to catfish immunoglobulin (Ig). These studies showed that this battery of monoclonal antibodies did not detect significant amounts of Ig on all lymphocytes. U~like polyclonal antisera which demonstrated nearly 100%+surface Ig cells, the monoclonal antibodi~s detected ~0% surface Ig cells. Furthermore, the percentage of Ig cells reactive with two of these monoclonals, tentatively shown to react with two different types of catfish light chains, was found to be nearly additive when the two antibodies were mixed. Thus it seems that fish lymphocytes, like their mammalian counterparts, have two different populations of lymphocytes; one which contains abundant surface Ig and one which does not. Whether these two types of cells represent the fish equivalents of B and T cells remains to be determined.
i~i;7~rk was supported by NSF grant PCM-79-04954 and NIH grant 1 R01 AI-
4.73

474 HYBRIDOMAS AGAINST FISH Ig Vol. 6, No. 3
INTRODUCTION
One of the milestones in modern immunobiology has been the development of a variety of surface marker and functional distinctions between T and B cells. In spite of these advances, however, considerable uncertainty and controversy still continues regarding the nature of the antigen receptor on T cells. This controversy has been particularly difficult to resolve when interpreting the results obtained with lower vertebrates such as fish. Reports with a variety of fish species have shown that most, if not all, fish lymphocytes possess abundant surface immunoglobulin determinants demonstrable by membrane immunofluorescence using heterologous polyclonal antisera to fish Ig (1-4). These findings suggesting the presence of surface Ig on all fish lymphocytes take on considerable importance since rather strong arguments by analogy indicate that fish have B and T cells (5, 6). One interpretation of these findings is that fish may represent phylogenetic proof that the primitive recognition unit for all lymphocytes may be membrane Ig. Although a recent study has suggested that the accounts of Ig on fish thymocytes (presumably representing T cells) might be artifactual, i.e. due to trivial carbohydrate crossreactivity (7), subsequent investigations by other workers (3) have refuted these findings. Since some of this controversy is likely attributable to differences in the polyclonal antisera employed in various laboratories, it seems reasonable to suggest that monoclonal antibodies to fish Ig may yield ~ore definitive answers. The investigation described here demonstrates that monoclonal antibodies to catfish Ig from a battery of fifteen different murine hybridoma lines do not reveal surface Ig determinants demonstrable by immunofluorescence on all catfish lymphocytes.
MATERIALS AND METHODS
Preparation of catfish immunoglobulin The predominant, if not exclusive, Ig in channel catfish (Ictalurus
punctatus) serum, representing a tetrameric Iglf-like molecule (8), was purified from pooled serum by anion exchange chromatography and gel filtration (9). This material exhibited a molecular weight of ~720,000 by gel filtration in neutral aqueous buffer, was composed of equimolar H (~fW ~70,000) and L (MW ~ 25,000) chains, and was immunoelectrophoretically pure when reacted with rabbit antisera to catfish serum. It was also antigenically identical to specifically purified catfish antibody to the DNP moiety when reacted with polyclonal rabbit and mouse antisera to catfish Ig (Lobb and Clem, in preparation).
Development of hybridoma cell lines BALB/c mice were hyperimmunized with the purified Ig. The spleen cells
from two mice were pooled and fused with the myeloma line P3-X63-Ag8 according to the techniques of KBhler and ltilstein (10). Following growth in selective media (HAT), the cultures were assayed for secreted antibodies to catfish Ig by solid-phase radioimmunoassay. Briefly polyvinyl 96 well microtiter plates were coated with 5~g of catfish Ig in phosphate buffered saline (PBS, 0.01 M phosphate, 0.15 M, NaCl, pH 7.4). After one hour the plates were washed 3 times in PBS containing 1% bovine serum albumin (BSA). Hybridoma culture supernatants were introduced and allowed to react with the bound Ig for 1 hour. 125he plates were washed 3 times with PBS containing 1% BSA and 250,000 cym of !-rabbit anti-mouse IgG placed into each well (specific activity~ 10 cpm/~g). Following another 1 hour incubation period the wells were washed, severed and counted in a gamma spectrometer (Auto-gamma 5110, Packard). Positive fusions were considered to be those wells with radioactivity greater than 3x the background level.

Vol. 6, No. 3 HYBRIDOMAS AGAINST FISH Ig 475
Positive cultures were then cloned in soft agar and positive representative clones passed as ascitic tumors in pristane-primed BALB/c mice. This approach resulted in the development of fifteen different hybridoma lines; each line originated from a different well in the original fusion,
Fluorescenc5 microscopy 2 x 10 washed blood or spleen leucocytes, composed of >75% lymphocytes
based on morphologic criteria, isolated from lymphoprep (Accurate Chemicals) interfaces, were stained with the primary antibody at a dilution of 1:10 in catfish RPtiT-1640 (243 mOs) containing 0.03M sodium azide for 20 min at 4°C, This level of azide was necessary to prevent "cold" patching and capping. Cells were washed three times and incubated with a 1:90 dilution of a fluorescein isothiocyanate conjugated IgG preparation of rabbit anti-mouse IgG. After 20 min at 4°C the cells were washed and the proportion of positive cells enumerated by fluorescence microscopy using a Zeiss microscope equipped with an epi-fluorescence condenser. Illumination was from a XBO 75W high pressure xenon lamp with an exciter filter for blue interference 455-490, a beam splitter FT 510, and a barrier filter for green interference 520-560. 200-300 lymphocyte appearing cells were counted for each value reported.
RESULTS
Monoclonal Anti-Catfish Ig Antibodies Can Distinguish Differences in Catfish Lymphocytes
Catfish peripheral blood and splenic lymphocytes were analyzed for the presence of surface membrane Ig with the fifteen independently-derived monoclonal antibodies as well as a polyclonal mouse anti-catfish Ig reagent (MAC). The results obtained with }~C indicated that virtually 100% of the lymphocytes from both blood and spleen exhibited surface fluorescence (Table 1). However when catfish lymphocytes were examined using the hybridoma-derived monoclonal reagents, the results indicated that less than 30% of the lymphocytes expressed surface Ig determinants reactive with any particular monoclonal reagent. These analyses also showed different levels of lymphocyte reactivity with the monoclonal battery. On the basis of these results the monoclonal reagents could be subdivided into three categories of reactivity. Six of the monoclonal reagents reacted demonstrably with at least 20% or more of the lymphocytes from at least one of the fish analysed. Four other monoclonals reacted visibly with 7-16% and the final group recognized none or very low (<5%) numbers of catfish lymphocytes.
Mixed Monoclonal Anti-Catfish Ig Antibodies Are Not Reactive lvith All Catfish 1YmJ>hocytes
Due to the varying degree of monoclonal reactivities with catfish lymphocytes, it was important to determine if the monoclonal reagents could be combined to obtain a higher percentage of lymphocytes expressing surface Ig. Five of the highest-reactive monoclonal reagents were, therefore, paired with one another in equal volumes and used to assay lymphocytes from catfish peripheral blood. The results (Table 2) indicated that although there may have been an increase in the total number of fluorescing lymphocytes with some combinations of monoclonal reagents, there was no combination which approached the 100% level seen with the polyclonal reagent, MAC. Similarly if all five of these particular reagents were pooled in equal proportions, 38% of the peripheral blood lymphocytes exhibited membrane fluorescence, In fact when all fifteen monoclonal reagents were equally combined into one pool, the percentage observed was ~0% Ig positive lymphocytes.

An
tib
od
z
MA~
3F
l2-l
3
Ell
-l
1H
6-l
4
Bll
-l
2Gl0
-5
1G7-
3 4D
l0-8
3
Dll
-18
3B
6-7
2F5-
9 2
Dll
-21
1
B6
-ll
2G4-
19
4A84
-16
4D
:S-l
N
AF
TABL
E 1
Per
cen
t S
urf
ace
Lm
mun
oflu
ores
cenc
e o
f C
atfi
sh P
erip
her
al B
lood
and
S
ple
nic
Lym
phoc
ytes
U
sing
Mou
se H
ybri
dom
a A
nti
bo
die
s to
Cat
fish
Im
mun
oglo
buli
n
Fis
h 1
F
ish
2
Fis
h 3
F
ish
4
Fis
h 5
Blo
od
S:e
leen
B
lood
B
loO
d S
:ele
en
Blo
od
S:e
leen
B
lood
>9
9 98
>
99
>9
9 >9
9 >9
9 >9
9 >
99
28
26
33
25
9
--
-21
20
25
20
8
--
-19
19
15
16
7
21
18
26
25
18
30
--
20
26
28
10
10
12
1
1
4 31
10
15
18
20
22
18
2
--
-14
9
7 13
3
16
8 14
15
1
0
14
10
2
--
-7
9 6
5 1
--
-<
1
<
1 2
--
8 1
1
8 3
2 <
1
--
<
1 5
3 2
2 6
2 1
--
-5
<
1 2
--
<
1 <
1
4 3
< 1
<
1
<
1 1
<
1 <
1
< 1
< 1
<
1
<
1 <
1
<
1 <
1
< 1
<
1 <
1
. <
1
~use a
nti
-catf
ish
Ig
was
a p
oly
clo
nal
asc
itic
flu
id
reag
ent
der
ived
fro
m
BA
LC/c
mic
e hy
perf
mm
uniz
ed w
ith
catf
ish
16S
se
rum
Ig
. T
he asc
itic
flu
id w
as
indu
ced
wit
h
the
ori
gin
al
P3 m
yelo
ma
lin
e.
b Nor
mal
asc
itic
flu
id d
eriv
ed f
rom
BA
LB/c
mic
e b
eari
ng
th
e o
rig
inal
P3 m
yelo
ma
lin
e.
..,..
""-~ "' ~ ::0
...... ~ til > ~ ...... z til
1-3
'zj
......
til =
......
OCI ~
..... . "' . z 0 . w
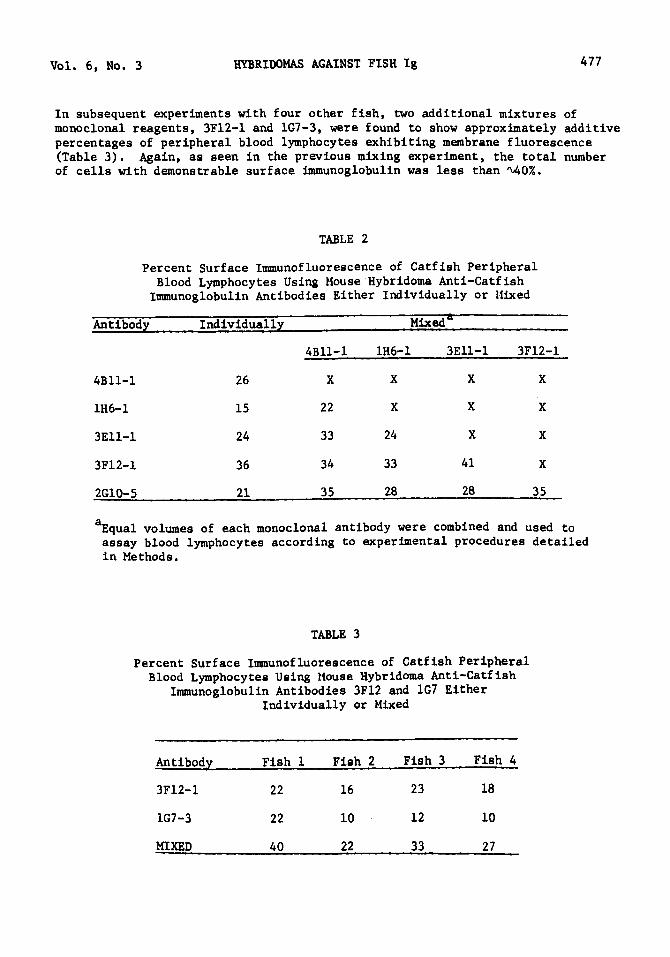
Vol. 6, No. 3 HYBRIDOMAS AGAINST FISH Ig 477
In subsequent experiments with four other fish, two additional mixtures of monoclonal reagents, 3Fl2-l and 1G7-3, were found to show approximately additive percentages of peripheral blood lymphocytes exhibiting membrane fluorescence (Table 3). Again, as seen in the previous mixing experiment, the total number of cells with demonstrable surface immunoglobulin was less than ~0%.
TABLE 2
Percent Surface Immunofluorescence of Catfish Peripheral Blood Lymphocytes Using Mouse Hybridoma Anti-Catfish
Immunoglobulin Antibodies Either Individually or Uixed
Antibodl Individualll Mixed
4Bll-l 1H6-l 3Ell-l 3Fl2-l
4Bll-l 26 X X X X
1H6-l 15 22 X X X
3Ell-l 24 33 24 X X
3Fl2-l 36 34 33 41 X
2Gl0-5 21 35 28 28 35
a Equal volumes of each monoclonal antibody were combined and used to assay blood lymphocytes according to experimental procedures detailed in Methods.
TABLE 3
Percent Surface Immunofluorescence of Catfish Peripheral Blood Lymphocytes Using tiouse Hybridoma Anti-Catfish
Immunoglobulin Antibodies 3F12 and 1G7 Either Individually or Mixed
Antibody Fish 1 Fiah 2 Fish 3 Fish 4
3F12-1 22 16 23 18
1G7-3 22 10 12 10
MIXED 40 22 33 27

478 HYBRIDOMAS AGAINST FISH Ig Vol. 6, No. 3
DISCUSSION
The purpose of the study reported here was to ascertain, by using a battery of murine hybridoma-derived antibodies to catfish immunoglobulin, if all catfish lymphocytes possess surface immunoglobulins demonstrable by immunofluorescence. Although each of the monoclonal reagents used have not yet been thoroughly characterized in terms of specificities (H vs L chains, isotypes, allotypes, etc.), the initial immunofluorescence results reported here when coupled with ongoing immunochemical studies strongly support the notion that all catfish peripheral lymphocytes do not contain significant amounts of surface immunoglobulin. Of particular importance in this regard are the results obtained with the monoclonal reagents 3Fl2 and 1G7. These reagents, based upon the results of radioimmunoassays and affinity chromatography experiments, have specificities for catfish L chains. Furthermore each of these monoclonal reagents appears to r~act with a different population of immunoglobulins present in the serum of all ( /4) fish examined. In addition when they are used together they will immunoprecipitate >90% of radioiodinated catfish Ig that will react with the polyclonal mouse anticatfish Ig reagent (Lobb and Clem, in preparation). Hence, it seems likely that the monoclonal reagents 3Fl2 and 1G7 have specificities for the catfish equivalents of kappa and lambda chains and thus may actually, when used as a mixture, be detecting all surface Ig positive catfish lymphocytes. This number of surface Ig positive cells would appear to represent ~407, of the total lymphocytes. In terms of the variability between different monoclonal reagents little can be said other than to suggest that some reagents showing no fluorescence may be directed to "buried" determinants on the cell surfaces whereas others may be detecting isotypic (or allotypic) differences that are not the same on all cells. Since monoclonal technology (and the resultant exquisite specificities) has not been previously applied to studies on lower vertebrate Igs, it would be premature to speculate further on this point. A final important question in the context of th~ study !eported here involves the functional significance of finding surface Ig and Ig lymphocytes in fish. Although definitive identification and separation of fish T and B cells is yet to be forthcoming, preliminary cell affinity column experiments using some of ~he monoclonal reagents described here have indicated the catfish surface Ig population contains those cells which undergo in vitro mitogenic responses to LPS. Similarly, the surface Ig- population contains those cells necessary for proliferative in vitro responses to ConA (Faulmann, Lobb, Clem, unpublished observations).--Thus, in light of these initial findings, it seems likely that catfish T-like and B-like cells may exhibit surface properties analogous to their mammalian counterparts.
1.
2.
3.
REFERENCES
Warr, G.W., DeLuca, D. and Marchalonis, J.J. Phylogenetic origins of immune recognition: Lymphocyte surface immunoglobulins in the goldfish, carassius auratus. R!£S· Natn. ~· Sci. U.S.A. 73, 2476, 1976.
Clem, L.W., McLean, W.E., Shankey, V.T. and Cuchens, 11.A. Phylogeny of lymphocyte hetererogenity. I. Membrane immunoglobulins of teleost lympho-cytes • .P!::!.· Comp. ~· 1, 105,1977.
Warr, G.W., DeLuca, D. and Griffin, B.R. Membrane immunoglobulin is present on thymic and splenic lymphocytes of the trout, ~ gairdneri. :!.· Imtn~· 123, 910, 1979.

Vol. 6, No. 3 HYBRIDOMAS AGAINST FISH Ig 479
4. Emmrich, F., Richter, R. and Ambrosius, H. Immunoglobulin determinants of the surface of lymphoid cells of carps. ~· I· Immun. 5, 76, 1975.
5. Etlinger, H.M., Hodgins, H.O. and Chiller, J.M. Evolution of the lymphoid system. I. Evidence for lymphoid heterogeneity in rainbow trout revealed by the organ distribution of mitogenic responses. I• ~· 116, 1547, 1976.
6. Cuchens, M.A. and Clem, L.W. Phylogeny of lymphocyte heterogeneity. Differential effects of temperature on Fish T-like and B-like cells. ~· 34, 219, 1977.
II. Cell.
7. Yamaga, K.M., Kubo, R. T. and Etlinger, U.H. Studies on the question of conventional immunoglobulin on thymocytes from primitive vertebrates. II. Delineation between Ig-specific and cross-reactive membrane components. J. Immun. 120, 2074, 1978.
8. Hall, S.J., Evans, E. E., Dupree, H.K., Acton, R.T., \o/einheimer, P.F., and Bennett, J.C. Characterization of a teleo~t tmmuhoglobulin: The immune macroglobulin from the channel catfish, Ictalurus punctatus. Comp. Biochem. Physiol. 46B, 187, 1973.
9. Lobb, C.J. and Clem, L.W. Phylogeny of immunoglobulin structure and function. X. Humoral immunoglobulins of the sheepshead, Archosargus probatocephalus. ~ Comp. Immun. 5, 271, 1981.
10. KBhler, G. and Milstein, C. Continuous cultures of fused cells secreting antibody of pred~fined specificity. Nature 256, 495, 1975.
Received Accepted
March 1982 April 1982