
original article
658 www.moleculartherapy.org vol. 19 no. 4, 658–666 apr. 2011
© The American Society of Gene & Cell Therapy
We have previously shown that the signaling pathway of the embryonic morphogen Sonic hedgehog (Shh) is recapitulated in the postnatal skeletal muscle in response to ischemia. We have also demonstrated that Shh is an indirect angiogenic agent upregulating various families of angiogenic growth factors and that Shh gene therapy improves angiogenesis and heart function in experimen-tal models of myocardial ischemia. Based on these find-ings, we hypothesized that Shh gene therapy is beneficial in an experimental model of peripheral ischemia. We found that intramuscular (i.m.) treatment with a plasmid encoding the Shh human gene (phShh) increased blood flow, capillary density, and arteriole density in mice in which peripheral circulation of the hindlimb was dis-rupted by removal of the common femoral artery. Shh gene therapy also enhanced vasculogenesis, by increas-ing the number of circulating bone marrow (BM)-derived endothelial precursors and improving the contribution of these cells to the process of neovascularization. Finally, phShh treatment induced upregulation of prototypical angiogenic, arteriogenic, and vasculogenic factors, such as vascular endothelial growth factor (VEGF), angiopoie-tin 1 (Ang-1), and stromal cell-derived factor-1 (SDF-1α). These data suggest that Shh gene therapy merits further investigation for its ability to trigger the expression of potent trophic factors and stimulate pleiotropic aspects of neovascularization in the setting of ischemia.
Received 21 July 2010; accepted 8 December 2010; published online 11 January 2011. doi:10.1038/mt.2010.292
IntroductIonSonic hedgehog (Shh) is a morphogen-regulating crucial epithe-lial-mesenchymal interactions during embryogenesis.1–5 We have previously demonstrated that Shh is an indirect angiogenic agent able to upregulate different families of angiogenic growth fac-tors.6 We have also shown that the Shh developmental pathway
is recapitulated in the adult skeletal muscle and myocardium in response to ischemia.7,8 Finally, we have found that intramyocardial gene transfer of naked DNA encoding human Shh has favorable effect on recovery from acute and chronic myocardial ischemia in adults, in terms of neovascularization, preservation of left ven-tricular function, and reduction of fibrosis and cardiac apoptosis.8 Importantly, Shh gene transfer also enhanced the contribution of bone marrow (BM)-derived endothelial progenitor cells (EPCs) to myocardial neovascularization.8 More recently, other investiga-tors have shown that genetic modification of mesenchymal stem cells (MSCs) with a Shh transgene improves MSC survival and angiogenic potential in the ischemic heart,9 while Shh favor neo-vascularization during ischemia.10 These data suggest that Shh therapy merits further investigation for its therapeutic potential in individuals with ischemic cardiovascular diseases.
In this study, we used a plasmid containing the amino-terminal domain coding sequence of the human Shh gene (phShh) to test the beneficial potentials of Shh gene therapy in an experimental model of peripheral limb ischemia. This is a pathologic condition characterized by occlusion of vessels of the arterial circulation of the legs, most often secondary to atherosclerosis, thrombosis, and/or inflammatory processes.11 Peripheral limb ischemia may lead to claudication, rest pain, skin ulceration, and critical limb ischemia.12 The prevalence of peripheral artery disease in people aged over 55 years is 10–25% and increases with age, with important clinical and economical implications.12 We found that, in 1-year-old-male mice (which represent middle-aged men), there is impaired activation of the Shh pathway in response to peripheral limb ischemia. In these animals, direct intramuscular (i.m.) injection of phShh induces functional activation of the Shh signaling pathway in the skeletal muscle and increases blood flow, capillary density, and arteriole density. We also observed that treatment with phShh increases the number of BM-derived circulating endothelial progenitors and enhances the contribution of these cells to the process of neovas-cularization of the ischemic muscle. Importantly, Shh gene therapy induces significant local upregulation of prototypical angiogenic, arteriogenic, and vasculogenic factors, thus demonstrating the
Correspondence: Roberto Pola, Department of Medicine, Division of Cardiovascular Research, St Elizabeth’s Medical Center, Tufts University School of Medicine, Boston, Massachusetts 02135, USA. E-mail: [email protected]
Pleiotropic Beneficial Effects of Sonic Hedgehog Gene Therapy in an Experimental Model of Peripheral Limb IschemiaMariangela Palladino1,2, Ilaria Gatto1, Valentina Neri1,2, Stefania Straino3, Marcy Silver2, Alessandra Tritarelli4, Andrea Piccioni1,2, Roy C Smith2, Eleonora Gaetani1, Douglas W Losordo5, Filippo Crea4, Maurizio Capogrossi3 and Roberto Pola1,2
1Laboratory of Vascular Biology and Genetics, Department of Medicine, A. Gemelli University Hospital, Catholic University School of Medicine, Rome, Italy; 2Department of Medicine, Division of Cardiovascular Research, St Elizabeth’s Medical Center, Tufts University School of Medicine, Boston, Massachusetts, USA; 3Laboratory of Vascular Pathology, IDIIRCCS Institute, Rome, Italy; 4Department of Cardiovascular Medicine, A. Gemelli University Hospital, Catholic University School of Medicine, Rome, Italy; 5Feinberg Cardiovascular Institute, Northwestern University, Chicago, Illinois, USA

Molecular Therapy vol. 19 no. 4 apr. 2011 659
© The American Society of Gene & Cell TherapyShh Gene Therapy for Peripheral Ischemia
ability of this treatment to regulate the expression of different fami-lies of proteins that are beneficial in the setting of ischemia.
resultsthe activation of the shh pathway is impaired in ischemic muscles of middle-aged miceWe induced unilateral hindlimb ischemia in young (8-weeks-old) and middle-aged (1-year-old) male mice and evaluated the expression levels of Gli1, the principal transcription factor of the Shh signaling pathway and a Shh-target gene. Mice were sacrificed at various time-points, hindlimb muscles were harvested, and Gli1 mRNA levels were evaluated by real-time PCR (Figure 1). Gli1 levels increased significantly upon induction of ischemia in young mice. Gli1 upregulation was already detectable 3 days after ischemia, peaked 7 days after ischemia, and then progres-sively decreased, returning to levels similar to baseline at day 21. Compared to young animals, Gli1 upregulation in response to ischemia was significantly impaired in middle-aged mice. As mentioned above, Gli1 is the principal transcription factor of the Shh signaling pathway and a Shh-target gene. Thus, its reduced upregulation constitutes evidence of impaired activation of the Shh pathway in response to ischemia in middle-aged mice.
treatment with phshh induces robust and diffuse activation of the shh pathway in the skeletal muscle of middle-aged miceA basic tenet of our study is that we can overexpress functional Shh in a controllable manner in mouse muscle tissues. To ensure that we have this capability, we tested different doses of phShh to artificially activate the Shh pathway in adult skeletal muscle. First, we evalu-ated the effects of these treatments on the expression levels of Gli1, which is, as mentioned above, the principal transcription factor of the Shh signaling pathway and a Shh-target gene. One-year-old mice were treated with either 100- or 200-μg phShh, as described in the Materials and Methods section. Some mice were treated with the empty plasmid and used as controls. Animals were sacrificed 7 days after treatment and hindlimb muscles were harvested and used to measure Gli1 expression by real-time PCR (Figure 2a). Gli1
expression was significantly increased in muscles of mice injected with phShh, compared to muscles treated with the empty plasmid. Gli1 expression was higher in muscles treated with 200-μg phShh than in those injected with 100-μg phShh. To further investigate the ability of phShh therapy to successfully activate the hedgehog path-way, the experiments were repeated in nls-Ptc1-LacZ mice, which carry a nondisruptive insertion of the LacZ gene under the control of the Ptc1 promoter. In these mice, LacZ expression corresponds to Ptc1 expression.6 Because Ptc1 is a classical downstream transcrip-tional target of the Shh pathway, LacZ upregulation constitutes evi-dence of successful activation of the Shh pathway.6 Nls-Ptc1-LacZ mice were treated with either 100- or 200-μg phShh, as described in the Materials and Methods section. Some nls-Ptc1-LacZ mice were treated with the empty plasmid and used as controls. Nls-Ptc1-LacZ mice were 1-year-old at the time of the experiments. Animals were sacrificed 7 days after treatment and hindlimb muscles were harvested and used to measure LacZ expression by assessment of β-gal activity (Figure 2b). Significant increment of β-gal activity was observed in muscles treated with phShh, compared to muscles treated with the empty plasmid. Also in the case of β-gal activity, the effect was stronger in mice treated with 200-μg phShh than in those injected with 100 μg phShh. In addition, we detected very robust expression of the human form of the Shh gene in the muscle of mice treated with 200-μg phShh (Figure 2c). To evaluate the range of expression of Ptc1 in response to phShh therapy, muscle of 1-year-old nls-Ptc1-LacZ mice treated with 200-μg phShh or the empty plasmid were also used for whole-mount X-gal staining (Figure 2d). This analysis showed that Ptc1 is robustly expressed in large areas of phShh-treated muscles, whereas muscles treated with the empty plasmid only showed negligible X-gal staining. These data demon-strate that treatment with 200-μg phShh is able to induce strong activation of the Shh pathway in large areas of the muscle, a con-dition that is important for the efficiency of treatment. Finally, to determine the identity of the cells responding to Shh treatment, we performed immunohistochemical analyses on X-gal-stained sections from skeletal muscles of 1-year-old nls-Ptc1-LacZ mice treated with 200 μg phShh. We found that X-gal was expressed in several interstitial cells (Figure 2e). These cells were not positive for the endothelial cell marker CD31 or the smooth muscle cell marker α-smooth muscle-actin (data not shown). In contrast, they stained positive for vimentin, a marker of mesenchyme-derived fibroblast cells (Figure 2f). These findings are consistent with previous data from our group and confirm that interstitial fibroblasts are crucial mediators of Shh activity in adult muscle tissues.6–8 Taken together, these data demonstrate the feasibility of Shh gene therapy in the adult skeletal muscle and show that treatment with 200-μg phShh is able to induce functional and robust activation of the Shh pathway in muscles of middle-aged animals.
shh gene therapy promotes blood perfusion and increases vascular density in ischemic hindlimb of middle-aged miceFor these experiments, we used 1-year-old C57BL/6J male mice. The rationale for treating these mice with phShh is that they display impaired activation of the Shh pathway in response to ischemia, as shown in Figure 1. Unilateral hindlimb ischemia was induced as described in the Materials and Methods section and i.m. treatment
16
14
12
10
8
6
4
2
0
gli1
mR
NA
exp
ress
ion
(isch
emic
/con
tral
ater
al h
indl
imb
ratio
)
Day 0 Day 3 Day 7 Day 14 Day 21
*
*
* Youngmice
Adultmice
Figure 1 Impaired activation of the sonic hedgehog (shh) signaling pathway is in ischemic muscles of middle-aged mice. Gli1 real-time PCR was performed at day 0, 3, 7, 14, and 21 after hindlimb ischemia. The ratio between Gli1 mRNA levels in the ischemic limb and the con-tralateral limb increases significantly in young (8-week-old) mice. In middle-aged (1-year-old) mice, Gli1 upregulation after ischemia is sig-nificantly impaired (P < 0.01).

660 www.moleculartherapy.org vol. 19 no. 4 apr. 2011
© The American Society of Gene & Cell TherapyShh Gene Therapy for Peripheral Ischemia
with 200 μg phShh (n = 10 mice) or an equal dose of empty plas-mid (n = 10) was performed immediately after surgery. Hindlimb blood flow was examined by laser Doppler perfusion imaging (Figure 3a,b). These studies revealed significant differences in limb perfusion between phShh-treated mice and controls. Indeed, although limb perfusion was equally severely reduced in the two groups of treatment immediately after surgery and at day 7 after ischemia, significant improvement of blood flow was detected in the hindlimb of mice treated with phShh at later time-points. At the end of the follow-up period (day 28 after induction of ischemia), blood perfusion ratio between the ischemic and the contralateral leg was 1.01 ± 0.05 in Shh-treated mice and 0.60 ± 0.02 in control animals (P < 0.01), indicating that phShh treatment induced complete recov-ery of blood flow in ischemic hindlimbs of middle-aged mice. We also determined and quantified capillary density by CD31 immunostain-ing in the ischemic adductor muscle of mice treated with phShh and
control (Figure 3c,d). These analyses were performed 28 days after induction of ischemia. Capillary density was significantly higher in the phShh-treated muscles compared to controls (P = 0.01). We also quantified arteriole density by α-smooth muscle-actin immunos-taining and determined that it was significantly increased in muscles treated with phShh compared to controls (P < 0.01) (Figure 3e,f). We also observed the presence of several large-diameter vessels in the phShh-treated group, a feature that was not present in mice treated with the control plasmid (Figure 3g,h).
shh gene therapy increases the number of BM-derived circulating endothelial progenitors after ischemia and increases their contribution to the process of neovascularizationWe investigated whether Shh gene therapy affects mobilization of BM-derived EPCs after hindlimb ischemia. Accordingly, we
10
8
6
4
2
0
gli1
mR
NA
exp
ress
ion
(phS
hh-t
reat
ed/-
untr
eate
d hi
ndlim
b ra
tio)
phShh 100 µg Empty plasmid Empty plasmidphShh 200 µg
a
8
7
5
6
3
4
2
1
0
β-G
alac
tosi
dase
act
ivity
(phS
hh-t
reat
ed/-
untr
eate
d hi
ndlim
b ra
tio)
P < 0.01P < 0.001
P < 0.001
P < 0.001
Empty plasmidphShh 200 µgEmpty plasmidphShh 100 µg
b
Empty plasmid phShh 200 µgH2O
β-Actin
Human Shh
540 bp
52 bp
c
e
d
Empty plasmid phShh 200 µg
f
Figure 2 treatment with plasmid encoding the sonic hedgehog human gene (phshh) induces activation of the sonic hedgehog (shh) path-way in the skeletal muscle of middle-aged mice. (a) phShh (100–200 μg) or empty plasmid was administered to 1-year-old mice. Gli1 expression was measured in hindlimb muscles 7 days after treatment. Significant increment of Gli1 expression was detected in phShh-treated compared to empty plasmid-treated muscles (P < 0.001). Increment was stronger in muscles treated with 200-μg phShh. (b) phShh (100–200 μg) or empty plasmid was administered to 1-year-old nls-Ptc1-LacZ mice. β-Gal activity was measured 7 days after treatment. It was significantly increased in muscles treated with 100 (P < 0.01) or 200 μg phShh (P < 0.001), compared to muscles treated with empty plasmid. (c) Real-time PCR (RT-PCR) showing strong expression of human Shh mRNA in the adductor muscle of mice treated with 200 μg of phShh. No signal is detectable in the muscles of mice treated with empty plasmid. (d) Whole mount X-gal staining showing strong and diffuse positive signal (blue staining) in large areas of nls-Ptc1-LacZ muscle treated with 200-μg phShh, whereas muscle treated with empty plasmid only shows negligible staining. (e) Presence of X-gal-positive cells (blue nuclei) in muscles of nls-Ptc1-LacZ mice treated with 200 μg phShh. (f) X-gal-expressing cells (blue nuclei) are positive for vimentin (brown cytoplasm).

Molecular Therapy vol. 19 no. 4 apr. 2011 661
© The American Society of Gene & Cell TherapyShh Gene Therapy for Peripheral Ischemia
performed experiments to quantify the number of circulating cells after hindlimb ischemia in 1-year-old mice treated with phShh or control plasmid. Treatment with 200 μg phShh (or control plas-mid) was performed by i.m. injection immediately after induction
of hindlimb ischemia. The effect of Shh treatment on the number of circulating progenitor cells was determined by fluorescence-activated cell sorting analysis, at baseline and 4 and 7 days after induction of ischemia. Five mice were studied in each experimental
Day 0
Day 7
Day 7
Day 14
Day 14
Day 21
Day 21
Day 28
Day 28
Control
Empty plasmid
phShh
phShh
1.2
1
0.8
0.6
0.4
0.2
0Before
ischemiaAfter
ischemia
P < 0.01
Isch
emic
/nor
mal
blo
od fl
ow r
atio
a
c
b
P = 0.01600
500
400
300
200
100
0Empty plasmid phShh
Cap
illar
ies/
mm
2
d
e
g
0
1
2
3
4
5
6 P < 0.01
Empty plasmid phShh
Art
erio
les/
mm
2
f
h
Figure 3 treatment with plasmid encoding the sonic hedgehog human gene (phshh) increases blood flow and vascular density in middle-aged mice after hindlimb ischemia. Laser Doppler perfusion imaging was performed in 1-year-old mice immediately after induction of hindlimb ischemia (day 0), and at day 7, 14, 21, and 28 after surgery. In color-coded images (a), normal perfusion is depicted in red. Immediately after surgical excision of the left femoral artery (day 0), marked reduction of blood flow is depicted in blue, both in control- and phShh-treated mice. Perfusion remained severely impaired for 14 days in animals treated with control plasmid, while only for 7 days in phShh-treated mice. At day 21, perfusion is depicted in yellow and red in phShh-treated mice and in light blue/green in control mice, indicating better blood flow in the group of animals treated with Shh. Finally, perfusion returned to normal levels by day 28 in phShh-treated mice (depicted in red), whereas it was still partially impaired in control animals (depicted in yellow). (b) Quantification of laser Doppler perfusion studies demonstrated that, at day 28 after surgery, blood flow ratio between the ischemic leg and the contralateral leg was about 1 in mice treated with 200 μg phShh (black line), whereas it was still significantly compromised in mice treated with empty plasmid (gray line) (P < 0.01). (c) Representative images of CD31 immunostaining in the adductor muscle of 1-year-old mice treated with 200 μg phShh or empty plasmid, 28 days after induction of ischemia. (d) At the end of follow-up, capillary density is significantly higher in phShh-treated mice compared to control mice (P = 0.01). (e) Representative images of immunostaining for α-smooth muscle (SM)-actin in the adductor muscle of 1-year-old mice treated with 200 μg phShh or empty plasmid, 28 days after ischemia. (f) At the end of follow-up, arteriole density is significantly higher in ischemic limbs of mice treated with phShh than in those of mice treated with empty plasmid (P < 0.01). (g) Representative image of a CD31+ large-diameter vessel in muscles of phShh-treated mice. (h) Representative image of α-SM-actin-positive large-diameter vessels in muscles of phShh-treated mice.
662 www.moleculartherapy.org vol. 19 no. 4 apr. 2011
© The American Society of Gene & Cell TherapyShh Gene Therapy for Peripheral Ischemia
group at each time-point. Treatment with phShh resulted in signifi-cant increase of the number of circulating CD45−/Sca-1+/Flk-1+ cells (P < 0.01) (Figure 4a), which are endothelial-committed pro-genitor cells with proangiogenic potential.13 To further investigate the role of Shh in vasculogenesis, we used a BM transplantation model, in which wild-type mice were transplanted with BM cells derived from nls-Ptc1-LacZ mice. After induction of ischemia, sev-eral X-gal positive cells were detected in the ischemic hindlimb of recipient mice (Figure 4b), thus demonstrating that BM-derived Ptc1-expressing cells are present in the ischemic site. Since Ptc1 expression constitutes evidence of functional activation of the Shh pathway,6 these findings indicate that Shh plays a direct role on these cellular elements. Based on this observation, we treated recip-ient mice with 200-μg phShh or control plasmid and found that, after induction of ischemia, the number of X-gal positive cells in the ischemic site was significantly higher in phShh-treated animals (P < 0.01) (Figure 4c). Double fluorescent staining for β-gal and CD31 demonstrated that several of these cells had incorporated into vas-cular structures (Figure 4d). These findings demonstrate that Shh gene therapy increases the number of cells that migrate from the BM to the site of ischemia and contribute to the process of neovas-cularization. These cells are β-gal positive, thus express Ptc1, which indicates that they are directly responding to Shh.
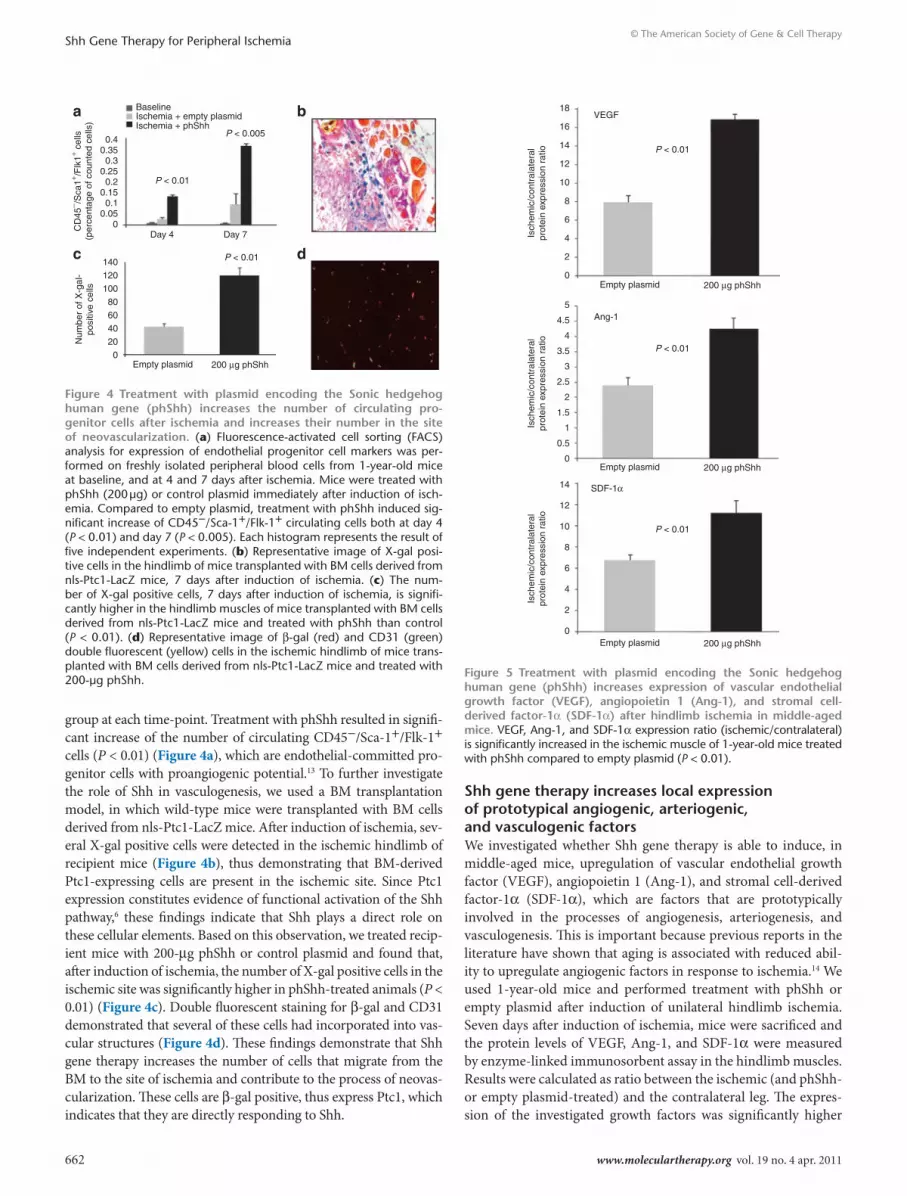
shh gene therapy increases local expression of prototypical angiogenic, arteriogenic, and vasculogenic factorsWe investigated whether Shh gene therapy is able to induce, in middle-aged mice, upregulation of vascular endothelial growth factor (VEGF), angiopoietin 1 (Ang-1), and stromal cell-derived factor-1α (SDF-1α), which are factors that are prototypically involved in the processes of angiogenesis, arteriogenesis, and vasculogenesis. This is important because previous reports in the literature have shown that aging is associated with reduced abil-ity to upregulate angiogenic factors in response to ischemia.14 We used 1-year-old mice and performed treatment with phShh or empty plasmid after induction of unilateral hindlimb ischemia. Seven days after induction of ischemia, mice were sacrificed and the protein levels of VEGF, Ang-1, and SDF-1α were measured by enzyme-linked immunosorbent assay in the hindlimb muscles. Results were calculated as ratio between the ischemic (and phShh- or empty plasmid-treated) and the contralateral leg. The expres-sion of the investigated growth factors was significantly higher
0.40.350.3
0.250.2
0.150.1
0.050C
D45
− /Sca
1+ /Flk
1+ cel
ls(p
erce
ntag
e of
cou
nted
cel
ls)
Day 4 Day 7
P < 0.01
P < 0.01
P < 0.005
BaselineIschemia + empty plasmidIschemia + phShh
140
120
100
80
60
40
20
0
Num
ber
of X
-gal
-po
sitiv
e ce
lls
Empty plasmid 200 µg phShh
a b
c d
Figure 4 treatment with plasmid encoding the sonic hedgehog human gene (phshh) increases the number of circulating pro-genitor cells after ischemia and increases their number in the site of neovascularization. (a) Fluorescence-activated cell sorting (FACS) analysis for expression of endothelial progenitor cell markers was per-formed on freshly isolated peripheral blood cells from 1-year-old mice at baseline, and at 4 and 7 days after ischemia. Mice were treated with phShh (200 μg) or control plasmid immediately after induction of isch-emia. Compared to empty plasmid, treatment with phShh induced sig-nificant increase of CD45−/Sca-1+/Flk-1+ circulating cells both at day 4 (P < 0.01) and day 7 (P < 0.005). Each histogram represents the result of five independent experiments. (b) Representative image of X-gal posi-tive cells in the hindlimb of mice transplanted with BM cells derived from nls-Ptc1-LacZ mice, 7 days after induction of ischemia. (c) The num-ber of X-gal positive cells, 7 days after induction of ischemia, is signifi-cantly higher in the hindlimb muscles of mice transplanted with BM cells derived from nls-Ptc1-LacZ mice and treated with phShh than control (P < 0.01). (d) Representative image of β-gal (red) and CD31 (green) double fluorescent (yellow) cells in the ischemic hindlimb of mice trans-planted with BM cells derived from nls-Ptc1-LacZ mice and treated with 200-μg phShh.
18
16
14
12
10
8
6
4
2
0
14
12
10
8
6
4
2
0
VEGF
P < 0.01
Empty plasmid 200 µg phShh
Isch
emic
/con
tral
ater
alpr
otei
n ex
pres
sion
rat
io
5
4
4.5
3.5
3
2.5
2
1.5
1
0.5
0
Ang-1
SDF-1α
P < 0.01
P < 0.01
Empty plasmid 200 µg phShh
Empty plasmid 200 µg phShh
Isch
emic
/con
tral
ater
alpr
otei
n ex
pres
sion
rat
ioIs
chem
ic/c
ontr
alat
eral
prot
ein
expr
essi
on r
atio
Figure 5 treatment with plasmid encoding the sonic hedgehog human gene (phshh) increases expression of vascular endothelial growth factor (VeGF), angiopoietin 1 (Ang-1), and stromal cell-derived factor-1α (sdF-1α) after hindlimb ischemia in middle-aged mice. VEGF, Ang-1, and SDF-1α expression ratio (ischemic/contralateral) is significantly increased in the ischemic muscle of 1-year-old mice treated with phShh compared to empty plasmid (P < 0.01).

Molecular Therapy vol. 19 no. 4 apr. 2011 663
© The American Society of Gene & Cell TherapyShh Gene Therapy for Peripheral Ischemia
in animals treated with phShh than in mice treated with control (Figure 5).
dIscussIonThis study presents several new findings. First, it shows that the upregulation of the Shh pathway, which physiologically occurs in young mice after induction of ischemia, is significantly impaired in middle-aged animals. Second, it demonstrates the feasibility of Shh gene therapy in the adult skeletal muscle and shows that treat-ment with phShh is able to induce significant and functional acti-vation of the Shh pathway in muscles of middle-aged mice. Third, it provides evidence that Shh gene therapy has beneficial effects in an experimental model of peripheral limb ischemia, in terms of increased angiogenesis, arteriogenesis, and vasculogenesis. Finally, it shows that Shh gene therapy increases the expression of multiple growth factors that are important for neovascularization in the ischemic skeletal muscle.
The demonstration of impaired activation of the Shh path-way in response to ischemia in middle-aged mice is interesting because it is consistent with the concept that important mecha-nisms of angiogenesis are altered by aging. In particular, it has been shown that aging is associated with impaired angiogenesis, due to reduced activity of hypoxia-inducible factor-1,15 defects in activation of nitric oxide synthase,16 and alterations in matrix com-position, inflammatory response, and expression of growth fac-tors.17,18 Deficient angiogenesis has been observed in several aging tissues, including myocardium, skeletal muscle, brain, kidney, retina, and peripheral nerve.19–24 Our data show that 1-year-old mice are unable to efficiently activate the Shh pathway in the isch-emic skeletal muscle, compared to young (8-weeks-old) mice. The molecular and cellular mechanisms underlying this phenomenon remain to be elucidated, but this is beyond the scope of our study. Further investigation is needed to understand whether impaired activation of the Shh pathway depends on reduced production of the Shh ligand, altered function, or expression of the Shh recep-tors or transcription factors, or increased expression of negative modulators of the Shh pathway. Additional studies using older animals are also needed to better clarify the relationship between aging and impairment of the Shh pathway. However, our findings suggest that dysregulation of the Shh pathway might be a novel contributor to deficient angiogenesis in aging tissues.
Impaired activation of the Shh pathway in the skeletal muscle of 1-year-old mice undergoing ischemia constitutes the rationale for treating these mice with Shh therapy. In the present study, we show that phShh is able to induce functional activation of the Shh pathway in the skeletal muscle of these mice, resulting in Gli1—the principal transcription factor of the Shh pathway—expression levels that are similar to those observed in young mice upon induction of ischemia. Importantly, in middle-aged mice, phShh-induced upregulation of the Shh pathway is mirrored by the activation of an efficient angiogenic process in response to hindlimb ischemia. Recently, it has been suggested that therapies that incorporate multiple growth factors, approaches that com-bine cell and gene therapy, and/or the administration of “mas-ter switch” agents that activate numerous downstream pathways are among the most credible and plausible steps forward in the field of therapeutic angiogenesis.25 This concept is based on the
evidence that the genesis, growth, and maintenance of an effi-cient and durable neovasculature occur through complex inter-actions and crosstalk between mechanisms that concomitantly involve angiogenesis, arteriogenesis, and vasculogenesis.26,27 In this study, we demonstrate that i.m. administration of a plasmid containing the human Shh gene induces simultaneous activation of angiogenic, arteriogenic, and vasculogenic mechanisms, with significant beneficial effects in a model of peripheral artery dis-ease. Previous studies from our group have demonstrated that Shh is an indirect angiogenic agent upregulating different families of angiogenic growth factors.6,7 We have also previously shown that treatment with phShh improves vascularization and function in experimental models of myocardial ischemia.8 Here, we demon-strate that phShh therapy can also be used for ischemic diseases of the skeletal muscle and confirm the ability of this treatment to induce upregulation of angiogenic factors and promote therapeu-tic angiogenesis. Not only phShh upregulates VEGF expression in treated muscles, but also induces increased expression of Ang-1, consistent with our previous demonstration that Shh is able to modulate the expression of angiopoietins in the adult.6 Ang-1 is a crucial regulator of smooth muscle cell recruitment and, not sur-prisingly, its upregulation leads to the formation of large vascular structures endowed of a well-structured tunica media.6 Here, we show that phShh treatment has the ability to upregulate Ang-1 expression in the skeletal muscle and increase the number of arte-rioles and large-diameter vessels.
Data from our group and other investigators have previously suggested that Shh might also be involved in the regulation of EPC biology and function.8,28,29 Here, we demonstrate that phShh gene therapy has direct effects on BM-derived progenitor cells. This is demonstrated by the fact that these cells express the Shh-target gene Ptc1 in response to Shh treatment, thus indicating that they are directly responding to Shh. We have previously shown that phShh therapy enhances the contribution of EPCs to the pro-cess of neovascularization,8 but the demonstration of a direct effect of phShh on endothelial progenitors in vivo was lacking. Indeed, although it is likely that phShh-induced vasculogenesis also occurs indirectly, through the upregulation of VEGF and SDF-1α—which are potent modulators of the mobilization and differentiation of EPCs30—our findings demonstrate that stimula-tion of endothelial progenitors also occurs directly in response to treatment with phShh. The fact that, at the level of the ischemic site, BM-derived cells express Ptc1 provides evidence that phShh acts on these cells in a direct manner. This is consistent with the previous demonstration that Shh protein promotes BM-derived EPC migration and proliferation in vitro.31 Our findings also strengthen the concept that Shh is a stem cell regulator and gov-erns stem cell fate, as indicated by its role during embryogenesis and a large body of evidence in the literature.1–5,32,33
In this scenario, it is reasonable to hypothesize that, upon induction of skeletal muscle ischemia, treatment with phShh increases local upregulation of prototypical growth factors, such as VEGF, Ang-1, and SDF-1α in interstitial fibroblasts. These growth factors are potent angiogenic, arteriogenic, and vasculo-genic agents that enhance the process of neovascularization of the ischemic site. Concomitantly, phShh therapy seems to have direct effects on BM-derived progenitor cells, which upregulate Ptc1 in

664 www.moleculartherapy.org vol. 19 no. 4 apr. 2011
© The American Society of Gene & Cell TherapyShh Gene Therapy for Peripheral Ischemia
the course of ischemia and increase their contribution to the pro-cess of neovascularization upon treatment with phShh. Therefore, our data indicate that Shh gene therapy displays its beneficial effects both directly and indirectly, through the stimulation of the Shh pathway in EPCs and the orchestrated production of fibro-blast-derived multiple growth factors that are eventually respon-sible for activating and/or enhancing angiogenesis, arteriogenesis, and vasculogenesis.
This study has some potential limitations. We used an acute model of hindlimb ischemia, with the main trauma being pro-duced in the thigh area of relatively healthy animals, whereas in humans peripheral limb ischemia most often develops chroni-cally, affects the distal leg area, and is usually associated with systemic pathological conditions, such as hypertension, diabetes, and hypercholesterolemia. However, no animal model accurately reflects the human condition and, to our knowledge, no experi-mental model of chronic, severe lower extremity ischemia cur-rently exists. Similarly, there is no animal with a lower extremity anatomy similar to the human anatomy. Accordingly, our results must be interpreted with caution when considering their potential clinical implications in humans. Another potential limitation is that, although we have demonstrated that phShh therapy results in robust activation of the Shh pathway in large areas of the mus-cle, it is possible that the expression profile of the Shh plasmid is less efficient in humans, especially considering the different size between murine and human skeletal muscles. Finally, it cannot be excluded that the time required by Shh gene therapy to exert its beneficial effects, which in our experimental model is about 3–4 weeks, might be too long to allow for clinically efficient revas-cularization in human subjects affected by acute peripheral limb ischemia.
In summary, our findings demonstrate that Shh gene therapy is feasible in the adult skeletal muscle and merits additional inves-tigation for its pleiotropic beneficial effects on neovascularization in an experimental model of limb ischemia.
MAterIAls And MethodsAnimals. We used young (8-weeks-old) and middle-aged (1-year-old) C57BL/6J male mice (Charles River Laboratories, Wilmington, MA). For some experiments, we also used 1-year-old nls-Ptc1-lacZ mice. For the BM transplantation model, we used 10-month-old nls-Ptc1-lacZ and FVB/NJ male mice (Charles River Laboratories). At the time of induction of isch-emia, these mice were 1-year-old. R.P. maintains a colony of nls-Ptc1-lacZ mice at his institution (mice were originally kindly provided by Matthew Scott, Stanford University). All the experiments were approved by the authors’ institutional review boards and ethics committees.
Ischemic hindlimb model. Unilateral hindlimb ischemia was induced as previously established34 and described by our group.6,7 Briefly, all animals were anesthetized and the left femoral artery was excised from its proxi-mal origin as a branch of the external iliac artery till the bifurcation into saphenous and popliteal arteries. Induction of ischemia was confirmed by laser Doppler perfusion imaging (Lisca, Peri-Med AB, Jarfalla, Sweden), as described below.
Analysis of the activation of the Shh pathway in response to ischemia. Unilateral hindlimb ischemia was induced in young (8-weeks-old) and middle-aged (1-year-old) C57BL/6J mice, as described above. Mice were sacrificed before ischemia (day 0) and 3, 7, 14, and 21 days after ischemia. These time-points were chosen because they encompass both early and late
changes in gene expression in response to ischemia and have been estab-lished previously.7,34 After sacrifice, muscles were harvested and the acti-vation of the Shh pathway was evaluated by quantification of gli1 mRNA levels by real-time PCR, as previously described.8
Description of the human Shh plasmid. The native full-length Shh gene product undergoes an autoprocessing reaction during its biogenesis, result-ing in the generation of amino- and carboxy-terminal domain products.35 Biologic activity is contained in the aminoterminal cleavage product. However, during autoprocessing, the amino-terminal domain products are cholesterol modified and this modification causes the amino-terminal protein to be tightly associated and tethered to the cell.36 As this could be a potentially disadvantageous phenomenon for a local gene therapy approach, the amino-terminal domain of human Shh coding sequence was selected to make a Shh plasmid, using a pCMV-ScriptPCR mam-malian expression vector (Stratagene, Agilent Technologies, Santa Clara, CA). The human Shh plasmid (phShh) that we used is a 4,878-bp plasmid that contains the 600 bp amino-terminal domain coding sequence of the human Shh gene. Expression of Shh gene is modulated by the presence of promoter sequences from cytomegalovirus to drive Shh expression. Downstream from the Shh complementary DNA is an SV40 polyadenyla-tion sequence. The plasmid also contains a gene that confers neomycin/kanamycin resistance to the host cells. Transgene expression in vitro has been assessed previously.8
Assessment of transgene expression in vivo. We have previously shown that, in experimental models of myocardial ischemia in rats, intramyocar-dial treatment with phShh induces significant improvement of capillary density, reduces fibrosis, and increases multiple wall motion scores.8 Based on the relative muscle masses of rat myocardium and mouse hindlimb mus-cles, we calculated a dose equivalent of phShh to use in mouse hindlimb. Based on this calculation, we tested the ability of two different doses of phShh (100 and 200 μg) to induce functional activation of the Shh path-way in murine hindlimb muscles. These two doses were tested in 1-year-old nls-Ptc1-lacZ mice, which carry a nondisruptive insertion of the lacZ gene under the control of the Ptc1 promoter.6 Treatment was performed by respectively injecting 20 and 40 μg phShh in five separate sites of the hindlimb. Mice treated with empty plasmid were used as controls. One week after treatment, mice were sacrificed and hindlimb muscles were har-vested. Quantification of β-gal activity was used to assess Ptc1 expression. Results are expressed as the ratio between β-gal activity in the treated leg versus the untreated leg. Additional experiments were performed to detect Gli1 expression in the mouse hindlimb in response to 100 and 200 μg phShh treatments. These experiments were conducted in 1-year-old C57BL/6J male mice. Treatment was performed as outlined above (five unilateral i.m. injections of 20 or 40 μg phShh in five separate sites of the hindlimb—total dose 100 or 200 μg). Mice were sacrificed 7 days after treatment and muscles were harvested. Expression of Gli1 was detected by real-time PCR. Total RNA was extracted, reverse transcribed, and amplified by PCR, as previ-ously described.8 Results are expressed as the ratio between Gli1 expres-sion in the treated leg versus the untreated leg. We also performed PCR to detect expression of human Shh in muscles treated with 200-μg phShh. PCR was performed using a Taqman 7300 apparatus (Applied Biosystems, Carlsbad, CA), with the following experimental conditions: hold for 2 minutes at 50 °C, and 10 minutes at 95 °C followed by 2-step PCR for 40 cycles of 95 °C for 15 seconds and 60 °C for 60 seconds. Primer sequences were as follows: Shh: forward 5′-GAGCAGACCGGCTGATGACT-3′, reverse 5′-AGAGATGGCCAAGGCATTTAAC-3′and FAM-AGAGGTG CAAAGACA-MGB. Expression of mouse β-actin was used as internal control. PCR products were analyzed by 1.5% agarose gel electrophoresis. For X-gal staining, tissues from nls-Ptc1-LacZ mice were fixed in 0.2% glu-taraldehyde, washed, stained overnight at 37 °C in 1 mg/ml X-gal, 5 mmol/l potassium ferricyanide, 5 mmol/l potassium ferrocyanide, 2 mmol/l MgCl2, 0.01% sodium deoxycholate, 0.02% Nonidet P-40, 50 mmol/l Na2HPO4

Molecular Therapy vol. 19 no. 4 apr. 2011 665
© The American Society of Gene & Cell TherapyShh Gene Therapy for Peripheral Ischemia
pH 8, and visualized as whole mounts or paraffin sections. For immuno-histochemistry on X-gal-stained paraffin sections, endothelial cells were identified using a rat monoclonal antibody against mouse CD31 (BD, Franklin Lakes, NJ) and a biotinylated goat immunoglobulin against rat. For periendothelial cells, a mouse monoclonal antibody against smooth muscle α-actin conjugated with alkaline phosphatase (Sigma, St Louis, MO) was used. Staining for vimentin was done with goat serum against vimentin (Sigma) compared with normal goat serum (Sigma) using horse-radish peroxidase-conjugated donkey secondary antibody against goat (Jackson Immunoresearch, West Grove, PA).
Blood flow measurements. A total of 20 (1-year-old) C57BL/6J male mice were used for these experiments. Ischemia of the left hindlimb was induced as described above. Ten mice were treated with 200-μg phShh, directly injected in the skeletal muscles of the left hindlimb immediately after induction of ischemia. Ten mice received injection of the empty plasmid and were used as controls. Blood flow was measured in ischemic and con-tralateral hindlimbs by laser Doppler perfusion imaging (Lisca) at baseline, immediately after induction of ischemia (day 0, to confirm efficient induc-tion of ischemia), and 7, 14, and 28 days after ischemia, as established.34 Before imaging, excess hairs were removed from the limbs using depilatory cream and mice were placed on a heating plate at 37 °C. Low or no perfu-sion was displayed in dark blue, whereas high perfusion was displayed in red. To avoid the influence of ambient light and temperature, results were expressed as the ratio between perfusion in the left (ischemic) versus right (nonischemic) limb.
Quantification of vascular density. After completion of blood flow mea-surements, 28 days after induction of ischemia, mice were sacrificed and both right and left adductor muscles were excised and fixed in 4% buffered formalin overnight. Muscles were then embedded in paraffin and sectioned at 3 μm. CD31 immunofluorescent staining was used to label endothelial cells, as previously described.7,34 Capillaries and large-diameter vessels were identified by CD31-immunopositive staining and appropriate morphology and counted by two independent operators blinded to the treatment regi-men under a ×20 objective and a ×5 lens. The number of capillaries from 30 randomly selected different fields was counted for each muscle. Capillary density was defined as mean number of capillaries per square millimeter, as established.34 For arteriole counting, sections were incubated with 1–10 μg/ml mouse monoclonal anti-smooth muscle α-actin antibody (clone 1A4; Sigma Immunochemicals, St Louis, MO), stained with rhodamine-con-jugated goat anti-mouse polyclonal antibody (Sigma Immunochemicals), and observed under a Zeiss Axiovert fluorescence microscope (Zeiss, Thornwood, NY). Arterioles were identified using established criteria:37 circular structure, α-smooth muscle-actin positivity around the entire cir-cumference of the vessel, diameter between 4 and 41 μm. Arteriole density was expressed as number of arterioles per square millimeter per section. Images were acquired with image analyzer KS300 software.
Flow cytometry. A total of 30 middle-aged (1-year-old) C57BL/6J male mice were used for these experiments. An equal number of 8-weeks-old C57BL/6J mice were also used. Hindlimb ischemia was induced as described above. Immediately after induction of ischemia, mice were treated with 200 μg phShh or empty plasmid, as described above. Blood samples (100 μl) were obtained through intracardiac puncture under anesthesia at day 4 and 7 after induction of ischemia. In some mice, blood samples were obtained in baseline conditions (without induction of isch-emia) and used as internal controls. The number of circulating progeni-tor cells was determined by fluorescence-activated cell sorting. Blood was incubated at room temperature for 30 minutes in the dark with mono-clonal antibodies for phycoerythrin-Cy5-conjugated anti-mouse-CD45 (BD), fluorescein isothiocyanate-conjugated anti-mouse-Sca-1 (BD), and phycoerythrin-conjugated anti-mouse-Flk-1 (BD). Nonviable cells were excluded using Immunoprep reagent system (Coulter Q-Prep, Beckman Coulter, Brea, CA). Cells were gated for low expression of CD45, then cells
were analyzed for events double-positive for Sca-1 and Flk-1 and presented as a percent of total viable mononuclear cells. All samples were analyzed using a FACScan flow cytometer (BD), and the data analyzed by Cell Quest software (BD). Relevant isotype controls were also included to con-firm specificity and for compensation setting. At least 100,000 events were acquired. Fluorescence-activated cell sorting analyses were performed on at least five different experiments in each experimental set.
BM transplantation model and detection of lacZ-expressing cells. BM transplantation was performed as described.38 Briefly, the BM of recipi-ent mice was ablated with lethal irradiation (9.0 Gy), then 5 × 106 donor BM cells were injected intravenously. FVB/NJ mice were used as recipient animals, whereas nls-Ptc1-LacZ mice were used as donors. Recipient and donor mice were 10-month-old at the time of transplantation. Ten FVB/NJ mice were used and the transplanted BM was allowed to regenerate for 6–8 weeks before induction of ischemia. Immediately after induction of ischemia, mice were treated with i.m. injections of 200 μg phShh (n = 5) or control plasmid (n = 5) as described above. Mice were sacrificed 10 days later. After euthanasia, hindlimb muscles were harvested and processed for β-gal (or X-gal) and CD31 immunostaining, as previously described.8 The number of X-gal positive cells was counted by two independent operators blinded to the treatment regimen. The number of cells from 30 randomly selected different fields was counted for each muscle. Results are expressed as number/field.
Analysis of the expression levels of growth factors in mice treated with phShh and controls. Twelve middle-aged (1-year-old) C57BL/6J male mice underwent surgical induction of left hindlimb ischemia as described above. Immediately after ischemia, mice were treated with i.m. injection of 200 μg phShh (n = 6) or control plasmid (n = 6) in the left hindlimb. Mice were sacrificed 7 days later and muscles were harvested to measure local expression levels of VEGF165, Ang-1, and SDF-1α proteins, by enzyme-linked immunosorbent assay (R&D Systems, Minneapolis, MN and USCN Life Science, Wuhan, China), as previously described.39 Results are pre-sented as ratio between the injured muscle and the contralateral side.
Statistical analyses. All data are expressed as mean value ± SD. Statistical comparisons between groups were performed by Student’s t-test. A P value <0.05 was considered statistically significant.
AcKnoWledGMentsThis study was supported by grant RCSNE3 from the “Fondazione Roma,” NIH grant 1R21HL089684, and grant RBID08MAFS (FIRB-IDEAS) from the Italian Department of University and Research (MIUR). We are grateful to Matthew Scott (Stanford University) for providing us with the nls-Ptc1-LacZ mice. The authors declared no conflict of interest.
reFerences1. Johnson, RL and Tabin, CJ (1997). Molecular models for vertebrate limb development.
Cell 90: 979–990.2. Pepicelli, CV, Lewis, PM and McMahon, AP (1998). Sonic hedgehog regulates
branching morphogenesis in the mammalian lung. Curr Biol 8: 1083–1086.3. Ramalho-Santos, M, Melton, DA and McMahon, AP (2000). Hedgehog signals
regulate multiple aspects of gastrointestinal development. Development 127: 2763–2772.
4. Porter, JA, Young, KE and Beachy, PA (1996). Cholesterol modification of hedgehog signaling proteins in animal development. Science 274: 255–259.
5. Dassule, HR, Lewis, P, Bei, M, Maas, R and McMahon, AP (2000). Sonic hedgehog regulates growth and morphogenesis of the tooth. Development 127: 4775–4785.
6. Pola, R, Ling, LE, Silver, M, Corbley, MJ, Kearney, M, Blake Pepinsky, R et al. (2001). The morphogen Sonic hedgehog is an indirect angiogenic agent upregulating two families of angiogenic growth factors. Nat Med 7: 706–711.
7. Pola, R, Ling, LE, Aprahamian, TR, Barban, E, Bosch-Marce, M, Curry, C et al. (2003). Postnatal recapitulation of embryonic hedgehog pathway in response to skeletal muscle ischemia. Circulation 108: 479–485.
8. Kusano, KF, Pola, R, Murayama, T, Curry, C, Kawamoto, A, Iwakura, A et al. (2005). Sonic hedgehog myocardial gene therapy: tissue repair through transient reconstitution of embryonic signaling. Nat Med 11: 1197–1204.
9. Ahmed, RP, Haider, KH, Shujia, J, Afzal, MR and Ashraf, M (2010). Sonic Hedgehog gene delivery to the rodent heart promotes angiogenesis via iNOS/netrin-1/PKC pathway. PLoS ONE 5: e8576.

666 www.moleculartherapy.org vol. 19 no. 4 apr. 2011
© The American Society of Gene & Cell TherapyShh Gene Therapy for Peripheral Ischemia
10. Benameur, T, Soleti, R, Porro, C, Andriantsitohaina, R and Martínez, MC (2010). Microparticles carrying Sonic hedgehog favor neovascularization through the activation of nitric oxide pathway in mice. PLoS ONE 5: e12688.
11. Schainfeld, RM and Isner, JM (1999). Critical limb ischemia: nothing to give at the office? Ann Intern Med 130: 442–444.
12. Ouriel, K (2001). Peripheral arterial disease. Lancet 358: 1257–1264.13. Iwakura, A, Luedemann, C, Shastry, S, Hanley, A, Kearney, M, Aikawa, R et al. (2003).
Estrogen-mediated, endothelial nitric oxide synthase-dependent mobilization of bone marrow-derived endothelial progenitor cells contributes to reendothelialization after arterial injury. Circulation 108: 3115–3121.
14. Rivard, A, Fabre, JE, Silver, M, Chen, D, Murohara, T, Kearney, M et al. (1999). Age-dependent impairment of angiogenesis. Circulation 99: 111–120.
15. Rivard, A, Berthou-Soulie, L, Principe, N, Kearney, M, Curry, C, Branellec, D et al. (2000). Age-dependent defect in vascular endothelial growth factor expression is associated with reduced hypoxia-inducible factor 1 activity. J Biol Chem 275: 29643–29647.
16. Bach, MH, Sadoun, E and Reed, MJ (2005). Defects in activation of nitric oxide synthases occur during delayed angiogenesis in aging. Mech Ageing Dev 126: 467–473.
17. Rivard, A and Isner, JM (1998). Angiogenesis and vasculogenesis in treatment of cardiovascular disease. Mol Med 4: 429–440.
18. Sadoun, E and Reed, MJ (2003). Impaired angiogenesis in aging is associated with alterations in vessel density, matrix composition, inflammatory response, and growth factor expression. J Histochem Cytochem 51: 1119–1130.
19. Edelberg, JM, Tang, L, Hattori, K, Lyden, D and Rafii, S (2002). Young adult bone marrow-derived endothelial precursor cells restore aging-impaired cardiac angiogenic function. Circ Res 31: 90.
20. Smythe, GM, Shavlakadze, T, Roberts, P, Davies, MJ, McGeachie, JK and Grounds, MD (2008). Age influences the early events of skeletal muscle regeneration: studies of whole muscle grafts transplanted between young (8 weeks) and old (13-21 months) mice. Exp Gerontol 43: 550–562.
21. Gao, P, Shen, F, Gabriel, RA, Law, D, Yang, E, Yang, GY et al. (2009). Attenuation of brain response to vascular endothelial growth factor-mediated angiogenesis and neurogenesis in aged mice. Stroke 40: 3596–3600.
22. Khang, G, Kim, SW, Cho, JC, Rhee, JM, Yoon, SC and Lee, HB (2001). Preparation and characterization of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) microspheres for the sustained release of 5-fluorouracil. Biomed Mater Eng 11: 89–103.
23. Hughes, S, Gardiner, T, Hu, P, Baxter, L, Rosinova, E and Chan-Ling, T (2006). Altered pericyte-endothelial relations in the rat retina during aging: implications for vessel stability. Neurobiol Aging 27: 1838–1847.
24. Pola, R, Aprahamian, TR, Bosch-Marcé, M, Curry, C, Gaetani, E, Flex, A et al. (2004). Age-dependent VEGF expression and intraneural neovascularization during regeneration of peripheral nerves. Neurobiol Aging 25: 1361–1368.
25. Gupta, R, Tongers, J and Losordo, DW (2009). Human studies of angiogenic gene therapy. Circ Res 105: 724–736.
26. Carmeliet, P (2000). Mechanisms of angiogenesis and arteriogenesis. Nat Med 6: 389–395.
27. Semenza, GL (2007). Vasculogenesis, angiogenesis, and arteriogenesis: mechanisms of blood vessel formation and remodeling. J Cell Biochem 102: 840–847.
28. Yamazaki, M, Nakamura, K, Mizukami, Y, Ii, M, Sasajima, J, Sugiyama, Y et al. (2008). Sonic hedgehog derived from human pancreatic cancer cells augments angiogenic function of endothelial progenitor cells. Cancer Sci 99: 1131–1138.
29. Nakamura, K, Sasajima, J, Mizukami, Y, Sugiyama, Y, Yamazaki, M, Fujii, R et al. (2010). Hedgehog promotes neovascularization in pancreatic cancers by regulating Ang-1 and IGF-1 expression in bone-marrow derived pro-angiogenic cells. PLoS ONE 5: e8824.
30. Yamaguchi, J, Kusano, KF, Masuo, O, Kawamoto, A, Silver, M, Murasawa, S et al. (2003). Stromal cell-derived factor-1 effects on ex vivo expanded endothelial progenitor cell recruitment for ischemic neovascularization. Circulation 107: 1322–1328.
31. Fu, JR, Liu, WL, Zhou, JF, Sun, HY, Xu, HZ, Luo, L et al. (2006). Sonic hedgehog protein promotes bone marrow-derived endothelial progenitor cell proliferation, migration and VEGF production via PI 3-kinase/Akt signaling pathways. Acta Pharmacol Sin 27: 685–693.
32. Blank, U, Karlsson, G and Karlsson, S (2008). Signaling pathways governing stem-cell fate. Blood 111: 492–503.
33. Sims, JR, Lee, SW, Topalkara, K, Qiu, J, Xu, J, Zhou, Z et al. (2009). Sonic hedgehog regulates ischemia/hypoxia-induced neural progenitor proliferation. Stroke 40: 3618–3626.
34. Couffinhal, T, Silver, M, Zheng, LP, Kearney, M, Witzenbichler, B and Isner, JM (1998). Mouse model of angiogenesis. Am J Pathol 152: 1667–1679.
35. Roelink, H, Porter, JA, Chiang, C, Tanabe, Y, Chang, DT, Beachy, PA et al. (1995). Floor plate and motor neuron induction by different concentrations of the amino-terminal cleavage product of sonic hedgehog autoproteolysis. Cell 81: 445–455.
36. Porter, JA, Ekker, SC, Park, WJ, von Kessler, DP, Young, KE, Chen, CH et al. (1996). Hedgehog patterning activity: role of a lipophilic modification mediated by the carboxy-terminal autoprocessing domain. Cell 86: 21–34.
37. Schgoer, W, Theurl, M, Jeschke, J, Beer, AG, Albrecht, K, Gander, R et al. (2009). Gene therapy with the angiogenic cytokine secretoneurin induces therapeutic angiogenesis by a nitric oxide-dependent mechanism. Circ Res 105: 994–1002.
38. Takahashi, T, Kalka, C, Masuda, H, Chen, D, Silver, M, Kearney, M et al. (1999). Ischemia- and cytokine-induced mobilization of bone marrow-derived endothelial progenitor cells for neovascularization. Nat Med 5: 434–438.
39. Straface, G, Aprahamian, T, Flex, A, Gaetani, E, Biscetti, F, Smith, RC et al. (2009). Sonic hedgehog regulates angiogenesis and myogenesis during post-natal skeletal muscle regeneration. J Cell Mol Med 13(8B): 2424–2435.







