fen bİlİmlerİ enstİtÜsÜ doktora tezİ apis mellifera...
TRANSCRIPT

ANKARA ÜNİVERSİTESİ
FEN BİLİMLERİ ENSTİTÜSÜ
DOKTORA TEZİ
İRAN BAL ARILARI (Apis mellifera meda)’NIN VARROA PARAZİTİ
(Varroa destructor)’NE KARŞI KORUNMA YETENEĞİNİN
DEĞERLENDİRİLMESİ
Amir NAJI KHOEI
ZOOTEKNİ ANABİLİM DALI
ANKARA
2012
Her hakkı saklıdır

i
ÖZET
Doktora Tezi
İRAN BAL ARILARI (Apis mellifera meda)’NIN VARROA PARAZİTİ
(Varroa destructor)’NE KARŞI KORUNMA YETENEĞİNİN DEĞERLENDİRİLMESİ
Amir NAJI KHOEI Ankara Üniversitesi
Fen Bilimleri Enstitüsü Zootekni Anabilim Dalı
Danışman: Prof. Dr. A. Çetin FIRATLI
Varroa destructor, bal arılarının (Apis mellifera L.) en önemli parazitlerinden biridir. Bu araştırmada İran bal arılarının (Apis mellifera meda) Varroa destructor'e karşı direnç özelliklerini araştırmak amacıyla, İran'ın Doğu Azerbaycan ilinin 5 farklı ilçesinden materyal olarak 60 bal arısı kolonisi ile Tebriz Üniversitesi Ziraat Fakültesi’nde deneme kurulmuştur. Çalışmada; tımarlama davranışı (TD), sır sökme ve petek gözü temizleme davranışı (SSPT) ve varroaya karşı duyarlı hijyen davranışın (VDH) incelenmiştir. Sonuçlar kovanın dip tahtasına düşen varroa sayısının, arı yoğunluğu ve sırlanmış yavrulu petek gözlerinin sayısından etkilenmemektedir. Kovanın dip tahtasına düşen varroa sayısı ile yavruların varroa bulaşıklığı ve foretik varroa bulaşıklığı arasında önemli korelasyon bulunmuştur. Dondurularak öldürülen yavrulara karşı arıların SSPT davranışlarıyla, yavru ve ergin arıların varroa bulaşıklığı arasında önemli negatif korelasyon bulunmuştur. Yani SSPT özelliğinin yüksek düzeyde olması kolonideki varroa bulaşığını azaltmaktadır. Araştırmada VDH özelliği, petek gözlerindeki bulaşıklığın azalmasına dayanarak hesaplanmıştır. Varroaya karşı duyarlı hijyen davranışıyla yavruların ve ergin arıların bulaşıklığı arasında negatif korelasyon bulunması, bulaşık gözlerin arılar tarafından temizlendiğini göstermektedir. Araştırma gerçekleşen kalıtım derecesi hesaplama tekniğine uygun deneme desenine göre yürütülmüştür. İncelenen özelliklere ait kalıtım dereceleri, diğer araştırmacıların buldukları ile uyum içerisindedir. Araştırma sonuçları varroaya karşı korunma özelliği taşıyan bal arısı hatlarının yetiştirilebileceğini göstermektedir. Mart 2012, 81 sayaf Anahtar kelimeler: İran bal arısı, Varroa destructor, tımarlama, sır sökme ve petek gözü temizleme davranışı, varroaya karşı duyarlı hijyen davranışı.

ii
ABSTRACT
Ph.D. Thesis
THE EVALUATION OF RESISTANCE ABILITY OF IRANIAN HONEY BEES (Apis mellifera meda) AGAINST VARROA (Varroa destructor) MITE
Amir NAJI KHOEI
Ankara University Graduate School of Natural and Applied Sciences
Department of Animal Science
Supervisor: Prof. Dr. A. Çetin FIRATLI
The parasitic mite, Varroa destructor, is a serious pest of honey bees, Apis mellifera L. To investigate the resistance ability of Iranian honeybees (Apis mellifera meda) against varroa, 60 honey bee colonies from five different regions of East Azarbayjan province were purchased and brought to the Agricultural Faculty of Tabriz University. The main objective of this study is to evaluate the resistance behavior of Iranian honey bees against varroa and the possibility of breeding the resistant bees. In the present study, grooming (GRO), hygienic (HYG) and varroa sensitive hygienic behaviors (VSH) individually and the combined effects have been studied. The results show that the number of adult bees and number of sealed brood will not affect the number of varroa mites falling on the hive bottom board. The correlation between the number of varroa mites falling on the hive bottom board with the number of mites in phoretic phase and the number of varroa infested brood was found significant. Results of correlation analysis indicate that hygienic behavior of bees against frozen brood is negatively and significantly correlated with the number of varroa in sealed brood and on adult bees. Thus the high frequencies of hygienic behavior reduce varroa infestation in the colony. In this study, VSH is investigated based on the reduction of brood infestation. A negative correlation between VSH behavior and the number of varroa in sealed brood and on adult bees indicated the cleaning of infested cells by bees and reducing varroa infestation. The experiment was designed to calculate the Realized Heritability of traits under study. The estimated heritabilties for traits under study are consistent with the findings of other researches. The results show that these heritable traits can possibly be used in order to breed honey bees lines with resistance ability against varroasis. March 2012, 81 pages Key Words: Iranian honey bee, Varroa destructor, grooming, uncapping and removing, varroa sensitive hygiene

iii
TEŞEKKÜR
Bu çalışmanın yürütülmesinde bana yol gösteren ve desteğini esirgemeyen, değerli
danışman hocam Prof. Dr. A. Çetin FIRATLI’ya (Ankara Üniversitesi Ziraat Fakültesi
Zootekni Bölümü), bilgi ve deneyimlerini her fırsatta bana aktaran değerli hocalarım
Prof. Dr. H. Vasfi GENÇER (Ankara Üniversitesi Ziraat Fakültesi Zootekni Bölümü)
ve Prof. Dr. Mete KARACAOĞLU’na (Adnan Menderes Üniversitesi Ziraat Fakültesi
Zootekni Bölümü), Genetik ve Islah açısından çalışmanın değerlendirilmesinde çok
değerli yardımlarını gördüğüm hocalarım Prof. Dr. Numan AKMAN (Ankara
Üniversitesi Ziraat Fakültesi Zootekni Bölümü) ve Prof. Dr. Mehmet Ali YILDIZ’a
(Ankara Üniversitesi Ziraat Fakültesi Zootekni Bölümü), istatistik analizlere yön veren
sayın Prof. Dr. Ensar BAŞPINAR’a (Ankara Üniversitesi Ziraat Fakültesi Zootekni
Bölümü), özveriyle zamanlarını ayırarak istatistik analizleri yapan değerli arkadaşlarım
Doç. Dr. Sadeg ALİJANİ’ye (Tebriz Üniversitesi Ziraat Fakültesi Zootekni Bölümü),
Araş. Gör. Yeliz KAŞKO ARICI’ya, Araş. Gör. Rabia ALBAYRAK’a, tezin yazımında
yardımlarından dolayı değerli arkadaşlarım Araş. Gör. Yasin KAHYA’ya ve Araş. Gör.
Gonca ÖZMEN ÖZBAKIR’a, saha çalısmalarımda bana eşlik ederek yardım eden
değerli arkadaşlarım Peyman GOLAMZADE’ye ve Amin YUSEFİ’ye teşekkürlerimi
sunarım. Tebriz Üniversitesi Ziraat Fakültesi Dekanı Sayın Prof. Dr. Safar
NASROLLAHZADE’ye araştırma merkezinde çalışmamın yürütülmesini desteklediği
için teşekür ederim .
Çalışmalarım süresince bana her zaman destek olan sevgili eşim ile oğluma ve
İRAN’daki ailelerim NAJİ KHOEİ’lere ve PASHMİNE’lere en derin duygularla
teşekkür ederim.
Amir NAJİ KHOEİ
Ankara, Mart 2012

iv
İÇİNDEKİLER
ÖZET ................................................................................................................................. i ABSTRACT ..................................................................................................................... ii TEŞEKKÜR ................................................................................................................... iii İÇİNDEKİLER .............................................................................................................. iv KISALTMALAR ........................................................................................................... vi ŞEKİLLER DİZİNİ ...................................................................................................... vii ÇİZELGELER DİZİNİ ............................................................................................... viii 1.GİRİŞ ............................................................................................................................ 1 2. KAYNAK ÖZETLERİ ............................................................................................... 5 2.1 Bal Arısı (Apis mellifera L.) Kolonilerinde Varroa Parazitinin Geçmişi ve Yayılması .......................................................................................................................... 5 2.1.1 Varroa jacobsoni ve Varroa destructor’un farklı haplotipleri ve konukçuları .. 6 2.1.2 Varroanın yaşam döngüsü .................................................................................... 6 2.1.3 Varroanın ergin arılarda, yavrularda ve tüm kolonideki etkileri ..................... 9 2.1.4 Varroanın neden olduğu viral hastalıkları ........................................................ 12 2.2 Tımarlama Davranışı .............................................................................................. 13 2.3 Sır Sökme ve Petek Gözü Temizleme Davranışı .................................................. 16 2.3.1 Sır sökme ve petek gözü temizleme davranışın testi ......................................... 19 2.4 Bal Arılarında Varroaya Karşı Duyarlı Hijyen Davranışı (VDH) ..................... 19 2.5 Bal Arılarında Kalıtım Derecesinin Tahmini ....................................................... 23 2.6 İz-katsayısı ............................................................................................................... 25 3. MATERYAL VE YÖNTEM .................................................................................... 27 3.1 Materyal ................................................................................................................... 27 3.2 Yöntem ..................................................................................................................... 29 3.2.1 Tımarlama davranışının (TD) ölçülmesi ............................................................ 29 3.2.2 Sır sökme ve petek gözü temizleme (SSPT) davranışının ölçülmesi ............... 31 3.2.3 Varroaya karşı duyarlı hijyen (VDH) davranışın ölçülmesi ............................ 33 3.2.4 Kalıtım derecesi .................................................................................................... 34 3.2.5 İstatistik analizler ................................................................................................. 35 4. ARAŞTIRMA BULGULARI ................................................................................... 37 4.1 Başlangıç Generasyonda Tımarlama Davranışı (TD) ile İlgili Özellikler.......... 37 4.2 Başlangıç Generasyonda Sır Sökme ve Petek Gözü Temizleme Davranışı (SSPT) ............................................................................................................................ 41 4.3 Başlangıç Generasyonda Varroaya Karşı Duyarlı Hijyen (VDH) Davranışı .... 47 4.4 Döl Generasyonunda Araştırılan Özellikler ......................................................... 50 4.4.1 Döl generasyonunda tımarlama davranışı (TD)................................................ 50 4.4.2 Döl generasyonunda sır sökme ve petek gözü temizleme (SSPT) davranışı ... 52 4.4.3 Döl generasyonunda varroaya karşı duyarlı hijyen (VDH) davranışı ............ 54 4.5 Gerçekleşen Kalıtım Derecesi ................................................................................ 55 4.5.1 Tımarlama davranışının (TD) kalıtım derecesi ................................................. 56 4.5.2 Sır sökme ve petek gözü temizleme (SSPT) davranışın kalıtım derecesi ........ 57 4.5.3 Varroaya karşı duyarlı hijyen (VDH) davranışına ait kalıtım derecesi ......... 58

v
5. TARTIŞMA VE SONUÇ .......................................................................................... 60 5.1 Tımarlama Davranışı (TD) .................................................................................... 60 5.2 Sır Sökme ve Petek Gözü Temizleme (SSPT) Davranışı ..................................... 62 5.3 Varroaya karşı Duyarlı Hijyen Davranışı (VDH) ................................................ 64 5.4 Kalıtım Derecesi ...................................................................................................... 66 KAYNAKLAR .............................................................................................................. 69 ÖZGEÇMİŞ ................................................................................................................... 80

vi
KISALTMALAR
ADV 1000 arı başına kovanın dip tahtasına düşen varroa
FAVO Foretik aşamada olan varroa oranı
FAVS Foretik aşamada olan toplam varroa sayısı
KAY Kolonideki arı yoğunluğu
KDV Kovan dip tahtasına düşen varroa sayısı
SSPT Sır sökme ve petek gözü temizleme davranışı
SYPS Sırlanmış yavrulu petek gözü sayısı
TD Tımarlama davranışı
VBAZ Varroa bulaşıklığının oranındaki azalma
VDH Varroaya karşı duyarlı hijyen davranışı
VBYO Varroa bulaşık yavru oranı
VBYS Varroa bulaşık toplam yavru sayısı

vii
ŞEKİLLER DİZİNİ
Şekil 2.1 Varroa destructor .............................................................................................. 7 Şekil 2.2 Varroanın yaşam döngüsü.................................................................................. 8 Şekil 2.3 Varroa bulaşmış pupa ve larvalar ...................................................................... 9 Şekil 2.4 Arının çenesiyle bedeninin bir kısmı kopartılmış bir Varroa ......................... 14 Şekil 3.1 Deneme materyalinin alındığı yöreleri gösteren İran haritası .......................... 27 Şekil 3.2 Kovanın alt kısmına uygulanmış tel ızgara ve onun altına konulmuş
yapışkan kağıt ile üzerindeki varroalar. ........................................................... 31 Şekil 3.3 Sırlanmış petek gözlerindeki yavruların sıvı azot ile dondurarak öldürülmesi 32 Şekil 3.4 Sırlanmış petek gözlerinde varroa bulaşıklığının tespit edilmesi .................... 33 Şekil 3.5 FAVO ve VBYO faktörleriyle ADV karakteri arasındaki ilişkiler ................. 35

viii
ÇİZELGELER DİZİNİ
Çizelge 2.1 Bal arılarında (Apis mellifera) Varroa destructor ile taşınan ve bulaştırılan viral hastalıkları .......................................................................................... 12
Çizelge 3.1 Deneme planı ve süresi ................................................................................ 29 Çizelge 4.1 Başlangıç generasyonunda tımarlama davranışı ile ilgili olan özelliklere
ilişkin tanımlayıcı istatistikler ..................................................................... 37 Çizelge 4.2 Başlangıç generasyonunda 1000 arı başına kovanın dip tahtasına düşen
varroa sayısının yörelere göre tanımlayıcı istatistikleri ve Kruskal-Wallis testi sonuçları .............................................................................................. 38
Çizelge 4.3 Başlangıç generasyonunda tımarlama ile ilgili özelliklerin arasındaki Spearman’ın sıralama korelasyon katsayıları ............................................. 39
Çizelge 4.4 Başlangıç generasyonunda foretik aşamada olan varroa oranının yörelere göre tanımlayıcı istatistikleri ve Kruskal-Wallis testi sonuçları ................ 39
Çizelge 4.5 Başlangıç generasyonunda varroa bulaşık yavru oranının yörelere göre tanımlayıcı istatistikleri ve Kruskal-Wallis testi sonuçları ........................ 40
Çizelge 4.6 Başlangıç generasyonunda varroa bulaşık yavru oranı bakımından yöreler arasında ikili karşılaştırma analizi (p değerleri) .......................................... 40
Çizelge 4.7 Başlangıç generasyonunda tımarlama davranışı ile ilgili olan özelliklerin arasında her yöre için Spearman’ın sıralama korelasyon katsayıları .......... 41
Çizelge 4.8 Başlangıç generasyonun SSPT grubuna ait tanımlayıcı istatistikler ........... 42 Çizelge 4.9 Başlangıç generasyonun SSPT grubunda temizleme süreleri bakımından
yöreler içerisinde tekrarlanan ölçümlü varyans analizinin sonuçları ......... 43 Çizelge 4.10 Başlangıç generasyonun SSPT grubundaki kolonilerin sır sökme ve petek
gözü temizleme süresi bakımından yörelere göre karşılaştırılması ............ 44 Çizelge 4.11 Başlangıç generasyonun SSPT grubunda foretik aşamada olan varroa
oranlarına ait tanımlayıcı istatistikler ve Kruskal-Wallis testi sonuçları ... 44 Çizelge 4.12 Başlangıç generasyonun SSPT grubunda varroa bulaşık yavru oranlarına
ait tanımlayıcı istatistikler ve Kruskal-Wallis test sonuçları ...................... 45 Çizelge 4.13 Başlangıç generasyonun SSPT grubunda, varroa bulaşık yavru oranlarının
p değerlerine bakarak yörelerin ikili karşılaştırılması ................................. 45 Çizelge 4.14 Başlangıç generasyonun SSPT grubunda her yöre için ölçülen değişkenler
arasındaki Spearman’ın sıralama korelasyon katsayıları ............................ 46 Çizelge 4.15 Başlangıç generasyonun SSPT grubunda ölçülen değişkenlerin arasındaki
Spearman’ın sıralama korelasyon katsayıları ............................................. 46 Çizelge 4.16 Başlangıç generasyonun SSPT grubunda 3 alt grubun tanımlayıcı
istatistikleri ve Kruskal-Wallis testinin sonuçları ...................................... 47 Çizelge 4.17 Başlangıç generasyonun VDH grubunda ölçülen değişkenlere ait
tanımlayıcı istatistikler ............................................................................... 48 Çizelge 4.18 Başlangıç generasyonun VDH grubunda 3 alt grubun tanımlayıcı
istatistikleri ve Kruskal-Wallis testinin sonuçları ....................................... 49 Çizelge 4.19 Başlangıç generasyonun VDH grubunda varroa bulaşıklığıyla ilgili olan
özellikler arasında Spearman’ın sıralama korelasyon katsayıları ............... 49

ix
Çizelge 4.20 Döl generasyonunun tüm kolonilerinde tımarlama davranışı ile ilgili özelliklere ilişkin tanımlayıcı istatistikler .................................................. 50
Çizelge 4.21 Damızlığa seçilen kolonilerin döllerinde tımarlama davranışı ile ilgili olan özelliklere ilişkin tanımlayıcı istatistikler .................................................. 51
Çizelge 4.22 Kontrol grubu olarak rasgele seçilen kolonilerin döllerinde tımarlama davranışı ile ilgili olan özelliklere ilişkin tanımlayıcı istatistikler ............. 51
Çizelge 4.23 Damızlığa seçilen kolonilerin döllerinde SSPT davranışı ile ilgili olan özelliklere ilişkin tanımlayıcı istatistikler .................................................. 52
Çizelge 4.24 Kontrol grubu olarak rasgele seçilen kolonilerin döllerinde SSPT davranışı ile ilgili olan özelliklere ilişkin tanımlayıcı istatistikler ............................. 53
Çizelge 4.25 Damızlığa seçilen kolonilerin döllerinde VDH davranışı ile ilgili olan özelliklere ilişkin tanımlayıcı istatistikler .................................................. 54
Çizelge 4.26 Kontrol grubu olarak rasgele seçilen kolonilerin döllerinde VDH davranışı ile ilgili olan özelliklere ilişkin tanımlayıcı istatistikler ............................. 55
Çizelge 4.27 Tımarlama özelliğine göre damızlığa seçilen ve rasgele oluşturulan kontrol grubu kolonileri ile bunların döllerinde saptanan yavrulu gözlerde varroa bulaşıklığına ait tanımlayıcı istatistikler ..................................................... 57
Çizelge 4.28 Sır sökme ve petek gözü temizleme özelliğine göre damızlığa seçilen ve rasgele oluşturulan kontrol grubu kolonileri ile bunların döllerinde saptanan 24 ve 48 saat dilimlerde SSPT davranışına ait tanımlayıcı istatistikler ...... 58
Çizelge 4.29 VDH özelliğine göre damızlığa seçilen ve rasgele oluşturulan kontrol grubu kolonileri ile bunların döllerinde saptanan bulaşıklık oranındaki azalmaya ait tanımlayıcı istatistikler ........................................................... 59

1
1.GİRİŞ
Varroa parazitinin İran’a giriş tarihi kesin olarak bilinmemektedir. İlk kez 1981 yılında
Poorasgar tarafından İran’ın Azerbaycan bölgesinden gönderilen parazit örneklerine
dayanarak Ruttner varroa parazitinin İran’da bulunduğunu rapor etmiştir. Ruttner
(1981)’in raporuna ve 1984 yılında İran’da parazitin dağılımı ve bal arısı kolonilerinde
neden olduğu kayıplar dikkate alınarak varroa zararlısının İran’a 1979-1980 yıllarında
girdiği ileri sürülmektedir (Mossadegh ve Komeili 1987).
Sosyal yaşamlarda davranış mekanizması oldukça karışık ve hassastır. Özellikle bir ana
arının çok sayıda erkek arı ile çiftleşmesi sonucu kolonilerde oluşan farklı akrabalık
kombinasyonları, genetik yapı, besin durumu, yavru miktarı, iklim, mevsim, stres
faktörleri ve daha genel olarak çevre koşulları gibi birçok faktör arılarda davranış
değişikliklerine yol açabilmektedir. Bu değişkenler hızla dalgalanabildiği için bu
konularda yapılacak çalışmalarda çok hassas davranmak ve ince ayrıntıları bile ihmal
etmemek gerekir.
Arıcılık endüstrisi sadece kovan ve arı ürünlerinden ibaret değildir. Arıların ekolojide
oynadıkları en önemli rollerden biri bitkilerin tozlaşmasında oynadıkları roldür.
Özellikle doğada diğer biyolojik unsurların azalması tozlaşmada bal arılarının önemini
daha da artırmaktadır. Tarım alanlarına bal arısı kolonilerinin taşınabilme özelliği bu
böceği bitkisel üretimde önemli kılmaktadır.
Dışarıdan ana arı getirme yasağı nedeniyle İran bal arısı kaynaklarının sadece İran için
değil, bütün dünya için gen kaynağı olarak önemi fark edilmeli ve korunmalıdır.
Bienefeld vd. (1996) İran bal arısı Apis mellifera meda’nın A. m. carnica ve A. m.
ligustica’dan etkilenmediğini savunmuşlardır. Benzer şekilde Tahmasebi vd. (1998)
İran bal arısı populasyonlarına yönelik çalışmalarda, İran bal arısı populasyonunun A.
m. carnica, A. m. ligustica, A. m. caucasica, A. m. anatoliaca, A. m. syriaca, A. m.

2
yemenitica ve A. m. lamarkii’den tamamen ayrıldığını ve bu arılardan etkilenmediğini
belirlemişlerdir.
Belirli kurallara göre yapılan taksonomik sınıflandırmada İran bal arısı bir alt tür
(coğrafik ırk) olarak ilk kez Goetze tarafından Apis mellifera meda sistematik adıyla
tanımlanmıştır. Bu araştırmacının tanımlamasına göre işçi arıda skutellum ve karın
tergitleri sarı-kahverenginde ve ilk üç tergit siyah rengindedir (Shahrestani 1979).
Genetik ıslah çalışmaları arıcılığın önemli konularındandır. ABD ve AB ülkelerinde;
varroa zararlısına, Avrupa ve Amerikan yavru çürüklüğüne, kireç ve nosema
hastalıklarına dirençli, bal verimi yüksek ve oğul verme eğilimi düşük olan ırklar ve
hatlar oluşturulmuştur.
Islah yöntemleri kullanılarak bal arılarının ekonomik açıdan önemli olan genetik
özellikleri geliştirilebilir. Bu ıslah kriterlerinin oluşturulmasında koloni yönetimi ve ana
arı yetiştirme ve yapay tohumlama tekniklerinin kullanımı arıcılık sektöründe ıslah
programlarının başarıya ulaşmasında kullanılması gereken yollar olarak bilinmektedir.
İran'da bu çalışılmalar öncelikli olarak ele alınmalı, yerli populasyonların genetik
zenginlikleri ortaya çıkarılmalıdır.
Varroa zararlısının yavru ve ergin arılara doğrudan zararının yanında tulumsu
çürüklüğü, felç gibi viral hastalıkların yayılmasına da neden olmaktadır. Birçok bilimsel
çalışmaya göre son yıllarda yaygın görülen Koloni Çökme Bozukluğu (CCD) ile
ilişkilendirilen etmenlerden biri ve en önemlisi Varroa destructor parazitidir (Haddad
2011).
Bu çalışmada İran bal arılarının (Apis mellifera meda) Varroa destructor parazitine
karşı direnç özelliklerinin değerlendirilmesi hedeflenmiştir. Bu yüzden deneme
kolonilerinde varroaya dayanıklılık davranışları; örneğin tımarlama davranışı, sır sökme
ve petek gözü temizleme ve varroaya karşı duyarlı hijyen davranışı uygun yöntemlerle
incelenmiştir. Araştırmada tımarlama davranışını incelemek üzere, kovan dip tahtasına

3
düşen varroa sayısı yanında kolonilerde ergin arı yoğunluğu, sırlanmış petek gözü sayısı
ve bunlara bulaşan varroa sayıları da belirlenmiştir. Sır sökme ve petek gözü temizleme
davranışını incelemek için yavrulu bir çerçevede belli sayıda (210) yavru, sıvı azot
kullanarak öldürülmüş ve belli bir süre (24, 48 ve 72 saat) içerisinde arılar tarafından
temizlenmesi izlenmiştir. Varroaya karşı duyarlı hijyen (VDH) davranışının
incelenmesinde, her deneme kovanına varroa bulaşıklığının oranı belirlenmiş yavrulu
bir çerçeve verilerek bir hafta sonra varroa bulaşıklığının oranı tekrar ölçülmüştür. Elde
edilen verilere dayanarak araştırılan özellikler arasındaki ilişkiler ve etkileşimlerinin
populasyondaki düzeyleri belirlenerek, gerçekleşen kalıtım derecesi (h2) tahmin
edilmiştir. Gerçekleşen kalıtım derecesi ebeveyn olarak seçilen grup ile rasgele seçilen
grubun dölleri arasındaki farkın, ebeveynler arasındaki farka oranıdır.
Varroa zararlısı nedeniyle meydana gelen koloni ölümleri ve ürün kayıpları ile bu akara
karşı üretilen ve ithal edilen ilaçların giderleri, arıcılık maliyetlerini artıran önemli
faktörlerdir. İran arıcılarının varroa zararlısını kontrol etmek için kullandıkları
kimyasallar bal ve bal mumunda kalıntı bırakmasının yanısıra akarların bu ilaçlara
direnç geliştirmesi varroa zararlarını daha da büyütmekte ve kalıcı kılmaktadır.
Bal arısı ürünlerinde kimyasalların kalıntı sorununu gidermek ve akarların bu
kimyasallara direnç kazanmasını geciktirmek için kimi biyolojik mücadele yöntemleri
geliştirilmiştir. Paraziti erkek arı gözlerinde tuzağa düşürerek gözler sırlandıktan sonra
peteği kovandan çıkararak imha etmek ve kovanın dip tahtasına tel ızgara yerleştirip
altına yapışkan beyaz bir kâğıt koyarak kovanın dip tahtasına düşen akarların ölmesini
sağlamak, biyolojik mücadele yöntemlerden bazıları olarak kullanılmaktadır.
Biyolojik yöntemlerden biri de varroaya karşı dirençli arı hatlarının geliştirilmesidir.
Islah yöntemleri ile elde edilebilecek böyle hatlarda diğer mücadele yöntemlerine gerek
kalmadan varroa paraziti denetim altına alınabilmektedir.
Yukarıda kısaca önemi vurgulanan hijyenik davranış özellikleri, koloni performansı,
bazı hastalık ve parazitlere direnç gibi güncel sorunların çözümünde kullanılabilmesi

4
için iyice irdelenmesine ihtiyaç duyulmaktadır. İran genelinde yapılacak seleksiyon
çalışmalarında da kullanılabilecek yöntem ve koşulların belirlenmesi amacıyla bu
çalışmada Azerbaycan bölgesi bal arısı populasyonlarında hijyenik davranış yönünde
bir seleksiyon çalışması ile katkı yapılması hedeflenmiştir.
Araştırılan özelliklere uygulanıp başarı sağlayacak genetik ıslah çalışmaları varroaya
karşı dirençli arıların yetiştirilmesi ve arıcılığın ekonomik değerinin arttırılmasına katkı
sağlayacaktır.

5
2. KAYNAK ÖZETLERİ
ABD, AB ülkeleri, Avustralya gibi birçok ülke ve bölgede parazitlere ve bulaşıcı
hastalıklara karşı dirençli bal arısı hatlarının geliştirilmesi çalışmalarına hız verilmiştir.
Arı genetiği uzmanları, arı ırklarının, ekotiplerin hatta kolonilerin varroa parazitine karşı
farklı düzeylerde duyarlılık gösterdiklerini ortaya koymuş, parazite dayanıklı
kolonilerden yetiştirilecek ana arılar ile oluşturulan kolonilerin parazite daha dayanıklı
olduğunu kanıtlayan bulgular elde etmişlerdir. Bu çalışmalara devam edilip uygulamaya
aktarılması halinde varroa ile mücadelede büyük bir başarı sağlanabileceği ifade
edilmektedir ( Morse 1990, Nasr 1998, Palacio vd. 2000).
Bal arılarında son yıllardaki koloni kayıpları tüm dünyada rapor edilmektedir. Bu
kayıpların nedeni olarak birçok etken ileri sürülse de çoğu araştırmacılar birkaç etkenin
sorumlu olduğu konusunda fikir birliği içindedir. Koloni kayıplarının son yıllardaki en
önemli etkenleri arasında varroa ve taşıdığı virüsler gösterilmektedir. Varroa paraziti arı
kolonilerini zayıflatarak, diğer hastalıklar için ortam hazırlamaktadır. Varroa akarının
bal arıları ile iyi bir uyum içerisinde olduğundan kullanılan ilaçlar yeterince etkili
olmamakta veya varroa paraziti kısa zamanda ilaçlara direnç kazanmaktadır. Etkili
ilaçların aynı zamanda bal ve diğer arı ürünlerinde hem insan, hem de arı sağlığını
tehdit eden kalıntı sorunu çıkarması, varroa sorununun göründüğünden çok daha fazla
olumsuz etkilere yol açmaktadır. Bu nedenle varroa parazitine karşı dirençli arı
hatlarının geliştirilmesi çalışmalarına ağırlık verilmesi önem taşımaktadır.
2.1 Bal Arısı (Apis mellifera L.) Kolonilerinde Varroa Parazitinin Geçmişi ve
Yayılması
Varroa jacobsoni Oud olarak adlandırılan akar bal arılarının ciddi bir ektoparazitidir ve
ilk kez Oudemans (1904) tarafından Java’daki Apis cerana kolonilerinde, 1951’de de
Singapur’da bulunmuştur (Morse 1969).

6
Bu parazit Apis mellifera kolonilerinde ilk kez Hong Kong ve Filipinlerde (1962-63)
bulunmuştur. Sonraki yıllarda çok hızlı şekilde dünyaya yayılmış, bugün mellifera
türünün bulunduğu her yerde görülmektedir (Denmark vd. 1991).
2.1.1 Varroa jacobsoni ve Varroa destructor’un farklı haplotipleri ve konukçuları
Asya’nın her tarafında V. jacobsoni ile bulaşık A. cerana kolonileri arasında gözlenen
genotipik, fenotipik ve üreme özellikleri ile ilgili farklılıklar, V. jacobsoni’nin en az iki
türü olabileceğini düşündürmüştür. Bu noktadan hareket eden Anderson ve Trueman
(2000)’nın yeni sınıflandırmasına göre Malezya-Endonezya bölgesinde A. cerana’ya
bulaşan V. jacobsoni’nin 9 haplotipi (mtDNA Co-I gen dizisine göre) vardır. V.
jacobsoni’nin tanımlanmasında Java haplotipi ilk örnek olarak içinde olduğumuz
yüzyılın başında bulunmuştur. Asya anakarasında A. cerana’ya bulaşan varroanın 6
haplotipi V.destructor olarak adlandırılmıştır. A. cerana’dan alınan dişi varroaların, A.
mellifera’dan alınan dişilerden daha küçük olduğu, virülenslerinin farklı olduğu,
Avrupa’daki A. mellifera’larda üreyen fenotipik olarak benzer akarların mitokondriyal
DNA (mt DNA) sitokrom oksidaz I (CO-I) gen sekanslarının % 6.7 oranında farklılık
gösterdiği görülmüştür. Varroanın birden fazla türü olabileceği düşüncesine kaynaklık
eden temel farklılıklar bunlardır (Anderson ve Trueman 2000).
Son araştırmalar varroanın birçok farklı haplotipinin olduğunu göstermektedir. Bunların
her biri Asya’daki bir A. cerana ekotipinin doğal parazitidir. V.destructor’un Kore ve
Japon olarak adlandırılmış sadece 2 haplotipi Apis mellifera’nın paraziti olmuştur.
Bunun nedeni ise Apis mellifera larvaları tarafından salgılanan bir uyarıcının bu
haplotiplerin üremesine yol açmasıdır. Haplotiplerin %90’ı ise bu uyarıcıyı
algılayamadıkları için Apis mellifera’da parazitik olamamışlardır (Anderson 2006).
2.1.2 Varroanın yaşam döngüsü
Dişi varroa kırmızımsı kahverengi, vücut uzunluğu 1.00-1.77 mm, sırtüstü genişliği
1.50-1.99 mm, düğme şeklinde ve sekiz bacaklıdır (Şekil 2.1). Erkek varroa ise sarımsı,

7
bacaklar hafifçe bronzlaşmış, vücut uzunluğu 0.75-0.98 mm, sırtüstü genişliği 0.70-0.88
mm’dir (Denmark vd. 1991).
Şekil 2.1 Varroa destructor (Anonymous 2009)
V.destructor sadece erkek ve işçi arı pupası olan sırlanmış petek gözlerinde
üreyebilmektedir. Bu nedenle yavrulu petek gözünün kapatılmasından 15-20 saat önce,
dişi varroa üzerinde bulunduğu genç ergin arıdan ayrılarak 5. deri değiştirme evresini
geçiren bir larvaya geçmektedir (Şekil 2.2). Dişi varroa, petek gözü sırlandıktan 60-70
saat sonra ilk yumurtasını erkek varroa çıkacak nitelikte (n=7 kromozom) yumurtlar ve
bundan sonra 30’ar saatlik aralıklarla yumurtlamaya devam ederek 4-5 adet de dişi
varroa çıkacak nitelikte (n=14 kromozom) yumurta bırakır (Ibrahim 2005). Genel
olarak isçi arı gözlerinde 3, erkek arı gözlerinde 5 dişi varroa ergin hale gelebilmektedir
(Şekil 2.3). Ana arı yüksüklerindeki dişi varroalar erginleşemeden ana arı gelişimini
tamamlayarak gözü terk ettiğinden, varroanın ana arı yüksüklerinde çoğalma şansı
yoktur.
Varroanın yumurtadan çıkma ve erginleşme dönemi proto-nimf (proto-nymph) ve
deuto-nimf (deuto-nymph) evrelerinden oluşmaktadır (Ifantidis 1997).

8
Şekil 2.2 Varroanın yaşam döngüsü
Varroa jacobsoni nimfinin oluşumu yumurta kabuğunun içerisinde gerçekleşerek
yumurtanın bırakılmasından birkaç saat sonra görülebilir. Yumurta-nimf aşamasıyla
olgunluk dönemi arasındaki sürenin ortalaması dişilerde ve erkeklerde sırayla 5.8 ve 6.6
gündür (Ifantidis 1997). Colin vd. (1999) ise bu süreyi dişilerde 5-6 gün, erkeklerde 7-8
gün hesaplamışlardır. Erkeklerde yumurtlanmadan çıkışa kadar geçen süre 30 saat,
protonimf 52 ve deutonimf aşaması 72 saat sürmektedir. Dişilerde ise 23±1 saat
yumurta aşaması, 30±3 saat protonimf ve 81±4 saat deutonimf aşaması sürmektedir
(Martin 1994). Sırlanmış petek gözlerindeki olgun dişi varroalar öz erkek kardeşleriyle
birkaç kez çiftleşmektedirler. İlk kız kardeş en az 8 kez, ikincisi ise en az 4 kez erkek
kardeşiyle çiftleşir. Bu iki kız kardeş üçüncü kardeşlerine göre daha iyi döllenme
şansına sahiptirler. Çünkü üçüncü kız kardeş erkek kardeşiyle iki kere çiftleşebilmekte
ve çok az sperm alabilmektedir.

9
Şekil 2.3 Varroa bulaşmış pupa ve larvalar (Anonymous 2009)
Başkalaşım dönemini tamamlayan genç arı, petek gözünden çıkarken, ana varroa ve
olgunlaşmış kızları da petek gözünü terk ederler. Dişi varroalar kolayca diğer genç
arılara aktarılarak beşinci deri değiştirme evresini geçiren bir larvaya parazitlik
yapmadan önce, 4-13 gün foretik aşamada kalabilmektedirler. Genelde genç ergin arının
petek gözünden çıkmasından önce erkek ve olgunlaşmamış dişi varroalar sırlanmış
petek gözü dışında yaşayamadıklarından hücrenin içinde ölür (Ibrahim 2005). Martin ve
Kemp (1997) bir varroanın yaşam boyunca 2-3 generasyon ürettiğini hesaplamışlardır.
2.1.3 Varroanın ergin arılarda, yavrularda ve tüm kolonideki etkileri
Varroa zararlısı Apis mellifera L. kolonilerinde isçi ve erkek arıların sırlanmış petek
gözlerinde üreyerek 1 yıl içerisinde sayısını 2000 misline çıkarabilmektedir (Vandame
vd. 2000). Bu da koloninin çökmesine neden olur (Martin vd. 1998, Martin 2001).
Varroa zararlısı işçi ve erkek arı yavrularının kan sıvısından beslenerek ergin arılarda;
vücut ağırlığının düşmesi, karın ve kanatlarda deformasyon, erkek arıların uçuş zamanı
ve çiftleşme kabiliyetinde değişiklik, işçi arılarda yaşam süresinin azalması,
hipofaringel bezlerin gelişmemesi, parazit mücadelesinde kullanılan ilaçlara karşı
dayanıksızlık ve viral hastalıkların çoğalmasına neden olmaktadır. Bal arısı
kolonilerinin çökmesine neden olan diğer önemli faktörler Deforme Kanat Virüsün
(DWV-deformed wing virus) ve Akut Arı Paraliz Virüsünün (APBV-Acut Bee

10
Paralysis Virus) varroa tarafından taşınması ve bulaştırılmasıdır. (Ball 1994, Shimanuki
vd. 1994, Rinderer vd. 1999).
Üzerlerinde 4-6 varroa bulunan larvalar gelişmelerini sürdürebilirler. Fakat üzerlerinde
daha fazla varroa bulunan larvalar gelişemeyip ölürler veya kanatsız, tek kanatlı,
gelişememiş kanat, eksik bacaklı veya kısa karınlı bireyler olurlar. Varroa ile bulaşık
arılar huzursuzdur. Ana arının yumurtlama gücü azalır. Üzerlerinde varroa bulunan isçi
arılar yavruların bakım ve besleme işini ihmal ettiklerinden koloni zayıflar. Ayrıca
‘Parazitik Akar Sendromu’ adı verilen belirtiler de gözlenebilmektedir. Bu hastalığın
etkiki olduğu kovanlarda V.destructor mevcuttur, ergin arı populasyonu azalmaya
başlamış, ana arının yumurtlama performansı düşmüş, yavru gözlerinde noktalı delikler,
yavru çürüklüğüne benzer hastalık belirtileri ve ölen yavruların gözlerde ‘C’ seklinde
kalması gibi belirtiler gözlenmektedir (Hung vd. 1995).
Varroaların gelişmesi için en uygun sıcaklık 34°C’dir. Varroaların gelişme ve
çoğalmasına; genetik faktörler, koloni koşullarının uygunluğu, yavru alanının miktarı,
koloninin varroa bulaşıklık oranı etki etmektedir. Varroanın çoğalmasında üzerinde
geliştiği larvanın cinsiyeti ve ırkı da etkilidir. Kolonilerde yavru üretimi ne kadar erken
başlar, ne kadar geç biterse varroaların üreme hızı ve gelişmesi de o oranda artmaktadır.
Arıların uçuş alanı içerisinde fazla sayıda parazitle bulaşık koloninin bulunması, arıların
kovanlarını şaşırmaları, petek takviyesi ve etkisiz mücadele arılıktaki diğer kolonilerin
de bu parazitle bulaşmasına neden olmaktadır (Kumova 2004).
Varroa ile mücadelede kimyasal ve biyolojik yöntemler uygulanmaktadır. Varroa
mücadelesinde sentetik kimyasal olarak; Amitraz, Fluvalinate, Flumethrin ve
Bromopropylate gibi etken maddelere sahip ruhsatlı ilaçlar kullanılmaktadır. Bu
ilaçların etkili olabilmesi için en uygun zaman kovanda kapalı yavrunun hiç olmadığı
veya çok az olduğu erken ilkbahar ile kapalı yavrunun sona erdiği bal hasadından
sonraki geç sonbahardır. Çünkü ilaçlar kapalı yavru gözü içindeki varroaları
öldürememektedir. Varroa mücadelesinde başarılı olmak mücadelenin uygun zamanda,
uygun ilaçla ve uygun dozda yapılmasına bağlıdır. Sentetik kimyasalların bal ve bal

11
mumunda kalıntı bırakarak insan sağlığını olumsuz yönde etkileyebilmesi ve akarların
bu ilaçlara direnç geliştirmesinden dolayı son zamanlarda varroa mücadelesinde formik
asit, laktik asit ve oksalik asit gibi organik asitler ile thymol gibi uçucu yağlardan oluşan
doğal ilaçların kullanımına başlanmıştır. Biyolojik mücadelede ise en çok kullanılan
yöntem; kovana erkek arı gözü bulunan peteklerin verilmesidir. Varroa erkek yavru
gözlerini daha fazla tercih ettiği için bu gözlere yumurtalarını bırakır. Gözler
kapandıktan sonra bu çerçeveler kovandan alınarak varroa nüfusu azaltılmış olur.
Ayrıca tel tabanlı ve polen çekmeceli kovanlar da kullanılarak düşen akarların tekrar
kovana dönmesi engellenebilir (Kumova 2004).

12
2.1.4 Varroanın neden olduğu viral hastalıkları
Çizelge 2.1 Bal arılarında (Apis mellifera) Varroa destructor ile taşınan ve bulaştırılan viral hastalıkları (Kevan vd. 2006)
Virüs Semptomlar Kaynaklar ABPV = Acute Bee Paralysis Virus (Akut Arı Felci Virüsü)
Ancak varroa ile birleşerek larva, pupa ve ergin arıları öldürmektedir. Diğer arılar sağlıklı görünür.
Anderson (1995) Bailey (vd. 1979)
BQCV = Black Queen Cell Virus (Siyah Ana Arı Hücre Virüsü)
Ana arıyı pupadan önce ve pupa aşamasında etkilemektedir. Çoğunlukla Nosema apis’ile birliktedir ve ana arı yüksüklerinde yavrular cansız ve kararmış bulunur.
Anderson (1995)
BVX = Bee Virus X (Arı X Virüsü)
Ergin arıların yaşam süresin azaltmaktadır. Bailey (1981)
BVY = Bee Virus Y (Arı Y Virüsü)
Nosema apis ile birliktedir. Anderson (1995)
CBPV = Chronic Bee Paralysis Virus (Kronik Arı Felci Virüsü)
Virüs bulaşmış ergin arı, yerinden çıkmış kanatlar, tüysüz ve yağlı bedenle titreyerek yorgun sürünmektedir.
Anderson (1995)
CWV = Cloudy Wing Virus (Bulanık Kanat Virüsü)
Kanatlar biraz bulanık görünür. Hornitzky (1987)
DWV = Deformed Wing Virus (Deforme olmuş Kanat Virüsü)
Bulaşık şiddetli olduğunda deforme olmuş kanatlar ve kısa yaşam süresi (<48 saat) görünür.
Yang ve Cox-Foster (2005)
KBV = Kashmir Bee Virus (Kaşmir Arı Virüsü)
Diğer patojenler örneğin Nosema apis ve Melissococcus pluton ile birleşmedikçe zararsızdır. Hem A.cerana hem de A.mellifera’ya bulaşmaktadır.
Anderson (1995)
KV = Kakugo Virüsü Virüs arının beyninde yerleşerek arı saldırgan olur. Fujiyuki (2004)
SBV = Sac Brood Virus (Tulumsu Yavru Virüsü)
Tulumsu çürüklük -
SBPV = Slow Bee Paralysis Virus (Yavaşça Arı Felci Virüsü)
Arıların ön bacakları felç olarak 12 saat sonra ölürler. Anderson (1995)

13
2.2 Tımarlama Davranışı
Bir işçi arı bacakları ve ağız parçaları yardımıyla üzerindeki parazitleri
temizleyebilmektedir. Bir işçi arı kendini tımarlayamazsa özel hareketler ve danslar ile
diğer arıların ilgisini çekerek paraziti üzerinden temizlettirir (Peng vd. 1987).
Tımarlama davranışı sıklığı Apis mellifera’da Apis cerana’ya göre daha azdır. Arıların
alt çenesiyle parazitleri yaradıkları birçok araştırmanın sonucunda belirlenmiştir (Şekil
2.4) (Ruttner ve Hanel 1992, Rosenkranz vd. 1997).
Moosbeckhofer (1992), yaralanmış parazit sayısı ile kolonideki bulaşıklık seviyesi
arasında negatif korelasyon bulmuştur. Böylece tımarlama davranışı, yaralanan zararlı
oranıyla ifade edilerek varroaya karşı dirençli kolonilerin seleksiyonunda yararlı bir
parametre olarak kullanılmaktadır (Fries vd. 1996). Tımarlama davranışı bal arısının
varroaya karşı önemli bir korunma mekanizmasıdır. Ancak tımarlama özelliği tek
başına, arıların varroaya karşı dirençli olduklarını göstermemektedir. Bu nedenle ıslah
programlarında her iki özellik, sır sökme ve tımarlama bakımından üstün olan koloniler
seçilerek dirençli arılar yetiştirmek mümkün olmaktadır (Spivak 1996).
Apis mellifera kolonilerinde foretik aşamada olan varroa akarları arıların tımarlama
davranışı ile uzaklaştırılır. Ancak bu davranış A. mellifera kolonilerinde A. cerana’ya
kıyasla daha az etkilidir (Fries vd. 1996). Varroa akarının sırtındaki düzenli ve küçük
çukurlar parazitin gelişme döneminde oluşan bozukluklardır ve bal arısı ya da predatör
eklembacaklılar tarafından yapılan hasarlardan ayrı olarak dikkate alınmalıdır (Davis
2009). Peng vd. (1987) tarafından yapılan bir çalışmada A. cerana’nın geniş ölçüde
tımarlama davranışı sonucunda deneme kolonilerine (n = 270) verilen varroaların
%99’undan daha fazlası uzaklaştırılmıştır. A.mellifera kolonilerinde (n = 270) ise
sadece zararlıların %3’ü tımarlama etkisiyle temizlenmiştir.
Thakur vd. (1997) kovan içerisine kızılötesi kamera yerleştirerek arıların varroaya karşı
aktif tımarlama yapmasını ve parazitlerin arılar tarafından ısırılmasını kaydetmişlerdir.

14
Bu araştırmacıların kayıtlarına göre; bazı arılar tımarlama aktivitesini defalarca
yapmaktadırlar. Moore vd. (1995)’e göre 4-31 günlük yaştaki bazı Apis mellifera arıları
tımarlama davranışı yapmak için özelleşmişlerdir. Bu arılar zamanlarının %84'ünü diğer
arıları tımarlayarak geçirmektedirler ve asla tarlacılık yapmamaktadırlar.
Bir koloninin tımarlama potansiyeli, arıların üzerinden kovanın dip tahtasına düşen
canlı parazit sayımlarıyla belirlenmektedir.
Şekil 2.4 Arının çenesiyle bedeninin bir kısmı kopartılmış bir Varroa (Anonymous
2008)
A. cerana’nın varroaya karşı geliştirdiği en az 4 davranış mekanizması California
Üniversitesi (Davis)’nden Peng vd. (1987) tarafından açıklanmıştır. Varroa bulaştırılan
arıların tımarlama hareketi ile varroaları üzerlerinden attıkları, eğer atamazlarsa
tımarlama dansı yaparak yakınlarında bulunan işçi arıların yardımıyla varroanın
üzerlerinden atılmasını sağladıkları gözlenmiştir. Daha sonra tımarlama yapan işçi
arının varroaya sürekli vurarak kovan dışına attığı görülmüştür. Varroanın atılması
zorlaştığında, diğer arılar varroalı arıyı hırpalayarak akarın arının üzerinden atılmasını
sağlamışlardır. Bu tip davranışlar A.mellifera’da gözlenmesine karşın A.cerana’da
görülen sıklıkta olmamıştır (Peng vd. 1987).

15
Rivera (2006), bal arısı kolonilerinin davranışlarından yararlanarak kolonilerin varroaya
karşı hangi düzeyde direnç gösterdiğini belirleyen yöntemi geliştirmiştir. Bu yönteme
göre her koloniden toplanan 10 arı şeffaf plastik kaplara konulmuştur. Yine
kolonilerden toplanan canlı varroadan birer adet arıların bulunduğu ortama bırakılarak,
arıların bunlara karşı davranışları gözlenmiştir. Varroayı ısırma, tımarlama ve vurma
davranış sayıları not edilerek puanlama yapılmış ve direnç gösteren koloniler
belirlenmiştir.
Bir koloniden alınmış arı örneklerinde, varroa bulaştırılmış ergin arıların davranışıyla
onları tımarlayan arıların davranışını izleyerek tımarlama özelliği ölçülebilir, ancak
koloniye ait tımarlama davranışını belirlemek için yeterli bir yöntem değildir.
Szabo vd. (1996)’ya göre laboratuvar koşullarında kafes içerisinde hapsedilmiş arılar
üzerinde yapılan denemelerden elde edilmiş gözlemlere dayanarak parazitlere karşı
tepki gösteren arı hatları belirlemek mümkün olmaktadır. Ancak bu gözlemlerle tüm
koloniye ait tımarlama davranışı arasında ilişki kurmak zordur (Boecking ve Spivak
1999).
Mangum’un (1997) yaptığı simülasyon çalışmasının sonuçlarına göre kovan içerisinde
varroanın kontrol altına alınması arıların tımarlama davranışı etkinliğine ve sıklığına
bağlıdır.
Doğal koşullarda dip tahtasına düşmüş olan varroalar üzerindeki hasarlar incelendiğinde
parazitlerin bir kısmının arılar tarafından yapılan belli bir savunma davranışı nedeniyle
öldüğü belirlenmiştir (Bienefeld vd. 1999).
Pettis ve Pankiw’e (1998) göre işçi arılar diğer arıların üzerindeki Acarapis woodi
parazitini bularak temizlemektedirler. Tımarlama dansı Apis mellifera’da ilk kez Milum
(1947) tarafından açıklanmıştır. Bir arının tımarlama dansı diğer arıların da tımarlama
yapmasını teşvik etmektedir (Ledezma 2000).

16
2.3 Sır Sökme ve Petek Gözü Temizleme Davranışı
Bal arıları da tüm canlılar gibi hastalık ve zararlıların tahdidi altındadır. Bunlarla
mücadele amaçlı birçok kimyasal kullanıma sokulmuş olmasına karşın ürünlerde kalıntı
sorunu, hedef organizmanın zaman içerisinde direnç kazanması gibi durumlar bu
uygulamalar sakıncalı olarak görülmektedir. Son yıllarda sorunlara karşı genetik ıslah
amaçlı çalışmalar olduğu görülmektedir.
Bal arılarının hastalık ve zararlılara karşı kullandığı genetik temelli direnç özelliği 2
farklı grupta toplanmaktadır.
a. Fizyolojik mekanizma
Larva ya da ergin bal arısı, salgıladığı kimi maddelerle hastalık etmeninin gelişimini ya
da üremesini baskı altına alabilmektedir. ABD’de yapılan araştırmalarda Amerikan
yavru çürüklüğüne direnç gösteren koloniler Rothenbuhler ve arkadaşları tarafından
bulunarak, ‘Brown hat’ adı verilen arı hattı oluşturulmuştur (Laidlaw ve Page 1997).
b. Davranış mekanizması
Bal arılarında en iyi bilinen direnç mekanizması hijyenik davranıştır. Bu davranış ilk
olarak 1937 yılında Park tarafından; hastalıklı sırlanmış kuluçka gözlerinin açılması ve
hastalıklı larvaların bu gözlerden çıkarılarak kovan dışına atılması olarak
tanımlanmıştır. Hijyenik davranış; Amerikan ve Avrupa yavru çürüklüklerine, kireç
hastalığına ve varroa parazitine karşı etkili bir davranış mekanizması olarak
gösterilmiştir (Laidlaw ve Page 1997).
İşçi arıların kapalı petek gözlerindeki Varroa destructor ya da yavru çürüklüğü
hastalığına bulaşmış yavruları bulma ve çıkarma yeteneğine hijyenik davranış denir
(Peng vd. 1987, Rath ve Drescher 1990, Boecking 1992a). Rothenbuhler (1964b)

17
hastalığa dayanıklı (hijyenik) ve yatkın (hijyenik olmayan) bal arısı hatlarının arasında
deneysel melezlemeler yapmış ve hijyenik davranışın 2 lokuslu resesif bir özellik
olduğunu ileri sürmüştür. Bu araştırmacıya göre; bir lokus sır sökme davranışını (u),
diğeri ise petek gözlerini temizleme davranışını (r) kontrol etmektedir. Ancak daha
soraki yıllarda QTL (Quantitative trait loci) analizi çalışmalarında hijyenik davranış ile
ilişkili 7 genetik marker bulunarak bu özelliğin poligenik olduğu ileri sürülmüştür
(Wilkes ve Oldroyd 2002).
Sır sökme ve petek gözü temizleme davranışın başlaması arıların koku duyarlılığına ve
bulaşık yavrunun salgıladığı uyarıcının şiddetine bağlıdır (Gramacho ve Spivak 2003).
Sır sökme ve petek gözü temizleme özellikli koloniler Amerikan yavru çürüklüğüne
(Rothenbuhler 1964a, 1964b, Spivak 1996) ve Kireç hastalığına da (Gilliam vd. 1983)
dayanıklı olmaktadır. Ayrıca sır sökme ve petek gözü temizleme davranışı Varroa
destructor parazitine karşı da bir direnç mekanizması olarak kabul edilmektedir
(Anderson ve Trueman 2000). Çünkü arılar sırlanmış petek gözlerinden paraziti bularak
temizleme yeteneğine sahiptirler (Peng vd. 1987, Boecking ve Drescher 1991, Spivak
1996). Hijyenik özellikli arılar tarafından varroa bulaşmış yavruları çıkarmada
Gramacho (1999) hijyenik sürecin ilk aşaması olarak bulaşık petek gözü sırında delik
açmanın öncelikli olduğunu savunmuştur (Gramacho ve Spivak 2003).
Bal arılarında hijyenik davranışı incelemek üzere birçok deneme yöntemi
geliştirilmiştir. Hijyenik davranış üzerine yapılan ilk araştırmalar, kolonilerin Amerikan
yavru çürüklüğüne neden olan patojen Bacillus larvae bakterisiyle bulaştırılmış
(Rothenbuhler 1964b) ya da siyanitle öldürülmüş yavrulara karşı tepki seviyelerini
belirlemişlerdir (Jones ve Rothenbuhler 1964). Kimi araştırmacılar ise iğneyle
öldürülmüş (Newton ve Ostasiewski 1986), dondurularak öldürülmüş (Taber 1982) ve
kireç hastalığı bulaştırılmış (Taber ve Gilliam 1987) yavrulara karşı kolonilerin tepki
seviyesini belirleyen denemeler yürütmüşlerdir. Daha sonraki yıllarda hijyenik davranışı
incelemek için en uygun ve güvenilir yöntemin sırlanmış petek gözlerindeki yavruları
sıvı nitrojen ile dondurarak öldürmek olduğu ileri sürülmüştür (Spivak ve Downey
1998).

18
Harris (2008) varroaya karşı duyarlı (VDH) (Varroa sensitive hygiene: VSH)
kolonilerde varroa ile bulaştırılmış işçi ve erkek arı yavrularına karşı arıların tepkilerini
laboratuvar ve saha testleri kullanarak kıyaslamış, VDH özelliği olan arıların 1 haftalık
sürenin sonunda daha az sayıda varroa ile bulaşık erkek arı pupası temizlediklerini
saptamıştır.
Hijyenik davranış varroa populasyonunun gelişmesini 3 şekilde engellemektedir: 1)
pupalar çıkarılırken olgunlaşmamış varroalar öldürülür, böylece üreme yapabilen zararlı
başına döl sayısı azalır, 2) petek gözünden pupalar çıkarılırken kaçmayı başaran olgun
dişi zararlıların foretik aşaması uzamaktadır, 3) foretik aşamada tımarlamaya maruz
kalan ergin zararlıların yaralanması ve ölmesi ihtimali artmaktadır (Spivak 1996).
Spivak ve Reuter (2001) tarafından yapılan bir araştırmada hijyen özellikli koloniler,
aynı populasyon yoğunluğu ve yavru alanına sahip Starline1 kolonilerinden daha çok bal
üreterek daha az kireç hastalığına yakalanmıştır. Hijyenik davranış karmaşık bir özellik
olarak genetik faktörlerden daha çok çevre koşullarından etkilenmektedir. Bazı çevre
faktörleri, örneğin işçi arıların yaşı (Thompson 1964) ve nektar akımı (Momot ve
Rothenbuhler 1971), hijyenik davranışı etkilemektedir. Milne (1985) bulaşık petek
gözlerini açma ve temizleme davranışının kalıtım derecesini sırasıyla 0.144, 0.022
olarak düşük bulmuş ve bu özellikler tarafından görülen farklılıklarda çevre koşullarının
daha etkili olduğunu savunmuştur.
______________________________________________________
1 Starline, A. m. ligustica arılarından geliştirilmiş hibrit arılardır.

19
2.3.1 Sır sökme ve petek gözü temizleme davranışın testi
Kolonilerden farklı zamanlarda farklı sonuçlar alınabileceğinden kolonilerin sezonda iki
kez test edilmesi gerekmektedir. Kolonilerin hijyenik yeteneğe sahip olup olmadığını
anlamak için iki yöntem kullanılmaktadır.
a) Petek kesme yöntemi
Sırlanmış kuluçka peteğinden 100 kuluçka gözü içeren bir parça kesilerek 24 saat -22
°C’de bekletildikten sonra kuluçka peteğindeki yerine yerleştirilerek koloniye verilir.
b) Sıvı nitrojen yöntemi
Sıvı nitrojen yöntemi uygulandığında işçilikten ve zamandan kazanılır. Yüz petek
gözünü içine alabilecek büyüklükteki, alt kısmı ve üst kısmı açık olan metal küp ya da
silindirin içine 250 ml sıvı nitrojen dökülerek 5 dakika beklemek suretiyle kapalı gözlü
kuluçka gözlerinin içinde bulunan yavruların ölmesi sağlanır. Her iki yöntemde kapalı
petek gözleri içerisindeki ölü arılar çerçeve ile koloniye geri verilerek 24 saat tutulur.
Bu sürenin sonunda petekler kolonilerden alınarak, arıların kaç adet gözü temizlediği
sayılır (Spivak vd. 1998b).
2.4 Bal Arılarında Varroaya Karşı Duyarlı Hijyen Davranışı (VDH)
Bu bölümde yer alan alıntılarda geçen özellik adı kısaltmalarından SMR (suppressed
mite reproduction) ve VSH (varroa sensitive hygiene) aynı özelliği tanımladıkları için
izlenebilirlik açısından Türkçeleştirilerek VDH (varroaya karşı duyarlı hijyen davranışı)
olarak kullanılması uygun görülmüştür. Varroanın doğal konukçusu A. cerana’dır. Bu
tür evrim sonucu bu akara karşı tımarlama ve ısırma davranışı ile kapalı kuluçka
peteklerindeki varroaları belirleyip kovan dışına atma davranışını geliştirmiştir.
Varroa, Apis mellifera türüne geçtiği ilk yıllarda çok büyük koloni kayıplarına neden
olmuştur. İlk zamanlar bu zararlıya karşı yoğun bir şekilde kimyasal mücadele

20
yöntemleri geliştirilerek zararı engellenmeye çalışılmıştır. Günümüzde, varroanın ona
karşı kullanılan birçok kimyasala direnç kazanmış olması nedeniyle genetik dirençli bal
arısı hatları oluşturulma yolu ağırlık kazanmıştır. Bal arılarında varroaya karşı ilk
dirençli hat oluşturma çalışmalarında her koloniden kapalı kuluçka gözlerinden 100 adet
açılarak erkek, dişi ve ergin olmayan akarların sayılması yolu izlenmiştir. Bu peteklerde
en az varroa bulunan kolonilerden ana arı yetiştirilerek bu parazite karşı dirençli hatlar
oluşturulmuştur (Laidlaw ve Page 1997).
Bal arısı kolonilerinde varroa parazitinin sayısal artışını sınırlayan diğer mekanizma,
konukçuya özgü faktör ve özelliklerdir. Yavruların üzerinde yumurta bırakan dişi varroa
sayısı konukçunun cinsine, türüne ve ırkına göre değişmektedir. VDH özelliği bal
arılarında Varroa destructor parazitinin sayısal varlığını kontrol edebilmektedir (Harbo
ve Harris 2005). Bazı bal arısı ırklarına ait kolonilerde ergin işçi arılar, üreyebilen
parazitlerle bulaşık pupaları belirleyerek çıkarmakta, kısır parazitlerle bulaşık pupaları
ise çıkarmamaktadırlar. Harris (2007) bu özelliği ‘varroaya karşı duyarlı hijyen’
davranışı olarak isimlendirmiştir.
Harris ve Harbo (2000) tarafından yapılan bir araştırmada; VDH davranışını incelemek
üzere, dirençli ve dirençsiz koloni ana arılarını birbirleri ile değiştirerek bunların
döllerinin varroa yoğunluğu üzerindeki etkisi araştırılmıştır. Buna göre, dirençli ve
duyarlı olmalarına bağlı olarak varroa populasyonunun değiştiği saptanmıştır.
Spivak’ın ve Harbo’nun varroaya karşı duyarlı hijyen davranışı önceleri SMR
adlandırdıkları sonra ise Harris’in (2007) tanımına uydukları görülmektedir.
Arılar petek gözlerinde yumurtlamaya başlayan varroa parazitlerini belirleyerek ortadan
kaldırmaktadırlar. Varroanın üremesi, işçi arıların parazitleri çıkarma davranışını
tetiklemektedir. Üreme yapamayan veya kısır varroalar belirlenemedikleri için petek
gözlerinde bırakılmaktadırlar. Böylece işçi arılar tarafından varroa parazitlerinin
yavrulu petek gözlerinden uzaklaştırılmasının seçici bir özellik olduğu ortaya
konulmuştur (Ibrahim ve Spivak 2006). Varroa ile bulaşmış petek gözleri sırlandıktan

21
sonraki 4-7 gün içerisinde yavru varroalar henuz gelişme döneminde iken arılar
tarafından sökülerek temizlenmektedir (Thakur vd. 1997).
Harbo ve Harris (2001) tarafından yapılan bir çalışmanın sonuçlarına göre; (1) bal
arıları tek bir direnç özelliği (varroaya karşı duyarlı hijyen) bakımından seleksiyona tabi
tutulduklarında, Varroa destructor’a karşı direnç göstermektedirler, (2) varroa dirençli
kolonilerden yetiştirilen ana arılar doğal çiftleşme yaptıklarında bile döllerinde bu
özellik önemli düzeyde korunmaktadır.
VDH özelliği olan A. mellifera kolonileri, yüksek düzeyde varroa ile bulaşmış yavrulu
petek gözlerini temizlemektedirler (Harbo ve Harris 2005).
İşçi arı yavru gözlerindeki varroa bulaşıklığı erkek arı yavru gözlerine kıyasla VDH
özelliğini daha çok tetiklemektedir. Bu nedenle VDH özelliğine sahip kolonilerde erkek
arı yavrusu fazla olduğunda varroa bulaşıklığının da yüksek olduğu görülmektedir
(Harris 2008). VDH kolonilerinde ergin arılar petek gözlerindeki varroaları temizlerken
kaçan dişi varroalar bu arıların üzerine geçmektedirler ya da peteklerin üstünde
dolaşırken arıların saldırısına uğrarlar (Aumeier ve Rosenkranz, 2001).
Thakur’a (1997) göre bal arıları açıkta dolaşan varroaları yakalayarak ısırmaktadırlar.
Ayrıca serbest kalmış dişi varroalar ergin arıların üstünde foretik aşamaya başlayarak
tımarlama davranışına maruz kalmaktadırlar. Bunun için VDH ana mekanizma olarak
diğer özellikleri ortaya çıkarmakta, örneğin parazitin foretik aşamasını uzatarak arıların
tımarlama davranışını artırmaktadır (Ibrahim vd. 2007).
Parazitlerin kısırlığı ile varroaya karşı duyarlı hijyen davranışının ilişkili olduğu
düşüncesiyle 2005 yılına kadar birçok araştırmacı tarafından kısır parazit (NR) oranına
dayanarak seleksiyon yapılmıştır (Harbo ve Harris 2005, Ibrahim ve Spivak 2006).
Ibrahim ve Spivak (2006) tarafından yapılan bir araştırmada, VDH özelliğine sahip
arıların Minesota hijyen stoku arıları ile kıyaslandığında, dondurularak öldürülmüş
yavruları daha hızlı temizleyerek daha hijyen davrandıkları belirlenmiştir. Minesota

22
hijyen stoku arıları, yavru çürüklüğüne ve kireç hastalığına dayanıklı özelliği olan
arılardır. Bu arılar ABD’nın Minesota Üniversitesinde çalışan Spivak tarafından A. m.
ligustica arılarından geliştirilmiştir.Diğer bir araştırmada, başka kolonilerden alınan
varroa ile bulaşık yavrulu petekler 1 hafta boyunca VDH özellikli arılara verildiğinde
parazit kısırlığı artış göstermiştir. Parazit kısırlığının artışı varroa bulaşıklığının azalma
oranına bağlı olarak, VDH özelliğinin sonucunda meydana gelmektedir (Harbo ve
Harris 2005, 2009).
Son araştırmalar VDH özellikli arıların yavrulu petek gözündeki parazitleri, kısır olup
olmadıklarına bakmaksızın temizlediğini ortaya koymaktadır (Harris vd. 2009, 2010).
Bu nedenle parazitlerde kısırlığın artış nedeni hijyenik özelliğinin başka halleri
olmalıdır. Örneğin, bir VDH özellikli kolonide bazen ağzı açılmış yavrulu petek gözü,
hijyen olmayan arılar tarafından tekrar kapatılır (Arathi vd. 2006), ki bu işlem VDH
kolonilerinde sık sık yapılmaktadır (Harris 2008). Ancak yavrulu petek gözlerinin
açılması nedeniyle varroa’nın üreme döngüsü bozulur ve içerisinde kısır varroa kalarak
kapatılır. Parazitin üremesi veya yavrularının salgıladığı koku VDH davranışının
başlatılmasında etkili değildir. Ne parazitten gelen kokular ne de yetişkin varroaların
hareketleri varroa bulaşmış yavruların çıkarılmasını açıklamaktadır (Aumeier ve
Rosenkranz 2001). Bu nedenle VDH davranışının başlatılması olasılıkla bulaşık
konukçu arı yavrusunun kokusundan kaynaklanmaktadır (Martin vd. 2002).
VDH özelliği olan arıların yetiştirilmesinde daha etkili bir yöntemin geliştirilmesi,
VDH’ya dayanarak seleksiyon yapılacaksa, 2-6 ay saha denemelerini gerektirir. Ergin
arıların VDH davranışının doğrudan ölçülebilmesi nedeniyle seleksiyonla hızlı bir
ilerleme sağlanması beklenebilir. Günümüzdeki ıslah çalışmaları bu davranışın dolaylı
ölçümlerine, örneğin yavrulardaki varroa bulaşıklığının azalma hızına
dayandırılmaktadır. VDH davranışının belirlenmesindeki hızlı yöntem, varroa bulaşmış
başka koloni yavrularının VDH kolonilerinde 40 saat ya da 1 hafta süresince izlenerek
bulaşıklığın azalma hızının saptanmasıdır. Kolonilerdeki parazit populasyonu büyümesi
ile 40 saat sonraki azalan bulaşıklık ve 1 hafta sonraki azalan parazit doğurganlığı
arasında negatif korelasyon bulunmuştur (Harris 2007, Villa vd. 2009).

23
2.5 Bal Arılarında Kalıtım Derecesinin Tahmini
Kalıtım derecesi, üzerinde durulan karaktere (verime) ait fenotipik varyasyonda
genotipik değerler arası farklılığın payıdır ve h2 ile gösterilir:
h2 = VA / VP
Kalıtım derecesinin alacağı en yüksek değer 1’dir. Bu da toplam varyasyonda genotip
dışındaki varyasyon kaynaklarının rol oynamadıkları hallerde söz konusu olabilir.
Kalıtım derecesinin alacağı en küçük değer ise sıfırdır ki bu da ancak populasyondaki
bütün fertler aynı genotipte oldukları zaman gerçekleşebilir. Birinci hale kalitatif
karakterlerde, ikinci hale ise akraba hatlarda veya uzun süre kapalı tutulan, başka
sürülerden gen aktarması yapılmayan sürülerde yaklaşılabilir. Gerçekte durum 0 < h2 <
1 eşitliği ile ifade edilir (Düzgüneş vd. 2003).
Bal arılarında kalıtım derecesinin hesaplanması için çeşitli teknikler Rinderer (1977)
tarafından tanımlanarak tartışılmıştır. Bu yöntemler cinsiyetin belirlenmesinde haplo-
diploid metodu ve kast sistemini dikkate almaktadır. Kalıtım derecesinin
belirlenmesinde bilinen metotlar ( özellikle ebeveyn-döl regresyonu ) bal arısı için
uygun değildir; çünkü ana arı ve erkek arılar farklı bir gelişme modeline sahip olarak
anatomik, genetik ve fizyolojik tüm özellikleri bakımından farklıdırlar. Bir kolonide
aynı zamanda üvey kardeşler (ana bir baba farklı) ve öz kardeşler (ana ve baba bir)
birlikte bulundukları için ebeveyn-döl regresyonundan faydalanarak kalıtım derecesi
tahmin edilemez (Falconer 1989). Rinderer’e (1977) göre kardeş analizlerindeki
problemler suni tohumlama yapılarak giderilebilir. Ana arı 20 erkek arının semeninin
karışımı ile tohumlandığında h2 tahmininde hem baba hem de ana unsurları geçerlidir.
İncelenen davranış koloniye ait bir davranışsa her çiftleşmede erkeklerin sayısını
dikkate almayarak sadece baba unsuru kullanılabilir (Rinderer 1986).
Kanca sayısı gibi işçi arılara özgü özelliklerin kalıtım derecesi, kontrollü çiftleştirmelere
ihtiyaç duyulmadan, bir populasyondan rasgele seçilen kolonilere ait veriler kullanılarak

24
hesaplanabilir. Koloniler içi işçi arıların akrabalık derecesini hesaplamak için
geliştirilmiş olan formül, ana arıyla çiftleşen erkek arıların sayısı ve bu erkek arılar arası
akrabalık derecesinin bir fonksiyonudur (Oldroyd ve Moran 1983). Böylece her bir
özelliğe ait kalıtım derecesi, grup içi korelasyon katsayısı akrabalık derecesine
bölünerek hesaplanabilir (Düzgüneş vd. 2003):
h2 = r P / r G
Bu eşitlikte;
h2 = kalıtım derecesi
r P = gruplar içerisindeki fertlerin birbirlerine benzerlik derecesi
r G = akrabalık derecesi
Damızlık olarak seçilenlerin üstünlüğünde her türlü gen etkisinin payı vardır.
Seleksiyon devam ettikçe eklemeli olmayan genlerin ayrı ayrı döllere dağılımları
(açılma) sonucunda, genotipte etkileri eklemeli olmayan genlerin payı azalır ve
hesaplanacak kalıtım derecesi dar anlamlı olmaya yönelir. 3-4 generasyon sonunda elde
edilen toplam ilerlemenin toplam seleksiyon üstünlüğüne bölünmesiyle bulunacak
kalıtım derecesi dar anlamlıdır ve buna gerçekleşen kalıtım derecesi denir (Düzgüneş
vd. 2003).
Boecking vd. (2000) Apis mellifera L. arılarında hijyenik davranışın kalıtım derecesini
ana-kız regresyon yöntemiyle hesaplamışlardır. Arıların, V.destructor’a bulaştırılmış
pupalara ve sırlanmış petek gözlerindeki iğneyle öldürülmüş yavrulara karşı hijyenik
davranışlarının ölçülmesiyle, veriler elde edilmiştir. İğneyle öldürülmüş yavrulara karşı
hijyenik davranışın kalıtım derecesi (h2 = 0.36 ± 0.30) ve tekrarlanma derecesi (r =
0.46) varroayla bulaştırılmış pupalara (h2 = 0.18 ± 0.27, r = 0.24) göre daha yüksek
tahmin edilmiştir. Arıların, V.destructor’a bulaştırılmış ve iğneyle öldürülmüş yavrulara
karşı hijyenik davranışlar arasında istatistik olarak yüksek derecede genotipik (rg =
0.61±0.51) ve fenotipik korelasyon (rp = 0.11) bulunmuştur (Boecking vd. 2000).

25
Moran (1984), gerçekleşen kalıtım derecesi hesapladığında arılarda doğal çiftleşme
nedeniyle kalıtım derecesinin h2=∆G/i değil, h2=(∆G/i)×(3/2) olarak hesaplanmasını
önermiştir. Bu yaklaşımdan yararlanarak ilk generasyonda dirençliliğin gerçekleşen
kalıtım derecesi 0.3 olarak elde edilmiştir (Kulincevic vd. 1992).
Harbo ve Harris (1999) araştırmalarında öz kardeşler arası benzerlik metoduna göre sır
sökme ve petek gözü temizleme davranışının, petek gözlerinin kapalı olma süresinin,
VDH özelliğinin ve fiziksel hasara uğratılmış ergin varroa sayısının kalıtım derecesini
sırasıyla 0.65, 0.89, 0.46 ve 0.00 hesaplamışlardır.
2.6 İz-katsayısı
Kantitatif bir karakterin etkenleri ile ilişkileri Path-katsayısı (İz-katsayısı) ve bunun
karesi olan belirleme katsayısı ile açıklanabilir. Bu katsayılar söz konusu karakterin
bunu etkileyen faktörlerden her birine ayrı ayrı ve bir arada ne ölçüde bağlı olduğu,
başka bir deyişle, sebep-sonuç ilişkisini ortaya konmuş olur. İncelenen kantitatif
karakterin gösterdiği değişimin (standart sapmanın) belli bir etkenden ileri gelen kısmı,
o etkene ait İz-(Path)-katsayısı olarak bilinir (Düzgüneş vd. 2003).
Path analizi metodu 1921 yılında Prof. Sewall Wright tarafından ‘Correlation and
causation’ adlı çalışma ile ilk kez ve tam olarak açıklanmıştır. Bir sebep-sonuç
sisteminde sebep değişkenlerinin sonuç değişkenine etkilerinin belirlenmesinde path
katsayılarından yararlanılır (Akçalı 1979).
Yukarıda yapılan tanımlama, istatistik olarak
P XA = S(X.A) / S(X)
şeklinde ifade edilir. Burada S(X) = X karakterine ait müşahede edilen (bütün
faktörlerin etkisi ile meydana gelen) standart sapma; S(X.A) = X karakterinin yalnız A

26
sebebiyle (diğer faktörler sabit tutulduğu zaman) göstereceği standart sapmadır
(Düzgüneş vd. 2003).
Path katsayıları aynı zamanda standardize edilmiş kısmi regresyon katsayıları olarak da
tarif edilir. Eğer sebep değişkenleri birbirlerinden bağımsız iseler, sebep
değişkenlerinden sonuç değişkenine giden path katsayıları bu değişkenlerle sonuç
değişkeni arasındaki korelasyon katsayısına eşittir (Akçalı 1979).

27
3. MATERYAL VE YÖNTEM
3.1 Materyal
Araştırma materyali olarak Doğu Azerbaycan ilinde Azerşehir, Bostan Abad, Şebster,
Aher ve Tebriz yörelerinde 5 farklı arılıktan 60 İran bal arısı (A. mellifera meda)
kolonisi alınmıştır (Şekil 3.1). Bu kolonilerden 50’si çalışmanın yürütüleceği Tebriz
Üniversitesi Araştırma Merkezi arılığına getirilmiş, 10 koloni varroa yetiştirmek üzere
uzak bir bölgeye taşınmıştır. Elli deneme kolonisi 25’er koloniden oluşan 2 gruba
ayrılarak, alındıkları yöreleri belirleyen harfler (A, G, vb. ) ile işaretlenmiştir. Birinci
grup 101’den 125’e, ikinci grup ise 201’den 225’e kadar numaralandırılmıştır. Birinci
grupta varroaya karşı duyarlı hijyen davranışı (VDH), ikinci grupta ise sır sökme ve
petek gözü temizleme davranışları (SSPT) çalışılmıştır. Ayrıca her iki grupta da
tımarlama davranışı (TD) ölçülmüştür.
Şekil 3.1 Deneme materyalinin alındığı yöreleri gösteren İran haritası
Araştırma iki aşamada gerçekleştirilmiştir. İlk aşamada 2009 yılında yaklaşık 6 aylık bir
sürede deneme kolonilerinde ölçümler yapılmış, ikinci aşamada ise 2010 yılında
başlangıç kolonilerinin arasından seçilen direnç ve kontrol gruplarının rasgele
çiftleştirmeler ile döl generasyonunun oluşturulması şeklinde yürütülmüştür.

28
Denemelere başlamadan önce, hem başlangıç (2009 yılı) hem de döl generasyonu
kolonilerinin (2010 yılı); arılı petek ile sırlanmış yavrulu petek gözü sayıları
bakımından eşitlenmesine çalışılmıştır. Koloniler arasındaki arı yoğunluğu farklılığının
KDV değişkeni üzerine olan etkisini gidermek üzere 1000 arı başına kovanın dip
tahtasına düşen varroa sayımlarına (ADV) bakılarak analizler yapılmıştır.
Bir generasyona mensup kolonilerin %20'sinin damızlığa ayrılacağı ve her 2 özelliğin
aynı önemde olduğu kabul edilmiştir. Elde edilen ölçüm değerleri kullanılarak her 25
kolonilik ebeveyn grubunda önce 5 koloni kontrol grubu olarak rasgele seçilmiştir.
Geriye kalan 20 koloni arasından en yüksek fenotipik değerlere sahip 4 koloni (%20)
direnç grubu olarak seçilmiştir. Kontrol grubundaki her koloniden 2’şer ana arı ve
direnç grubundaki her koloniden 5’er ana arı yetiştirilerek doğal çiftleştirilmişler ve bu
ana arılar ile 60 kolonilik döl generasyonu oluşturulmuştur. Bu kolonilerde, araştırılan
özelliklerinin fenotipik değerleri ebeveyn generasyonuna uygulanan yöntemlerle
belirlenmiştir. Ebevey-döl generasyonu verilerine dayanarak araştırılan özelliklerin
kalıtım dereceleri hesaplanmıştır. Araştırma yukarıda yazılanlardan da anlaşılacağı
üzere gerçekleşen kalıtım derecesi hesaplama tekniğine uygun deneme desenine göre
yürütülmüştür.

29
Çizelge 3.1 Deneme planı ve süresi Tarih Uygulama 01.10.2008-15.10.2008 İran arısı tarama gezisi 15.05.2009-30.05.2009 İran arısı kolonilerinin satın alınarak Tebriz
Üniversitesi Araştırma Merkezine getirilmesi 04.06.2009 Kolonilerin alındığı yörelere göre
numaralandırılması 10.06.2009-25.06.2009 Kovanlarda dip tahtasına düşen varroa sayısını
belirlemek için kovanların altına tel ızgara uygulanması
30.06.2009 Kolonilerin eşitlenmesi 05.07.2009-22.09.2009 Araştırılan özelliklerin ölçülmesi 06.11.2009-19.03.2010 Kolonilerin kışlatılması 03.05.2010 Ebeveyn grubunda damızlık olarak seçilen
kolonilerden ana arıların yetiştirilmesi 15.05.2010 Yetiştirilen ana arılardan döl generasyonunun
oluşturulması 11.07.2010-28.09.2010 Döl generasyonunda araştırılan özelliklerin
ölçümü Ekim 2010 Denemenin tamamlanması
3.2 Yöntem
3.2.1 Tımarlama davranışının (TD) ölçülmesi
Kovanın dip tahtasına düşen varroaların sayımlarına başlamadan önce deneme
kolonilerinde varroa populasyonundaki değişmenin izlenmesi amacıyla başlangıç
yoğunluğunun tahmin edilmesi gerekmektedir. Bir bal arısı kolonisinde, varroanın ergin
yaşamı 2 ana evreden oluşmaktadır; foretik aşama ve sırlanmış yavrulu petek gözündeki
üreme aşaması. Bu nedenle kolonideki varroa yoğunluğunun tahmin edilmesi için her 2
aşamadaki varroa sayısının belirlenmesi gerekir. Ergin arılar üzerindeki parazit
bulaşıklığını belirlemek için deneme kolonilerinin kuluçkalığının her birinden alınan
yavrulu petek üzerindeki işçi arılardan 150-200 adedi koloroformlu bir kavanozda
toplanmıştır. İşçi arılar ve üzerlerindeki parazitlerin koloroform etkisiyle birbirlerinden
ayrılması sonucunda, işçi arı ve parazit sayımları yapılmış, buradan da foretik
aşamadaki varroa oranı belirlenmiştir.

30
Kolonideki tüm foretik aşamada olan varroa bulaşıklığının belirlenmesi için her
kolonide işçi arı sayısının tahmin edilmiştir. Bir peteğin bir yüzü arıyla kaplandığında
yaklaşık 1500 ergin işçi arı olacağı hesabıyla kolonilerin arı sayıları hesaplanmıştır
(Burgett ve Burikam 1985, Ibrahim vd. 2007). Sonuçta ergin işçi arıların bulaşıklık
oranı ergin işçi arı sayısı ile çarpılarak kolonideki tüm foretik aşamada olan varroa
sayısı tahmin edilmiştir.
Kolonideki varroa yoğunluğunun tahmin edilmesi için yavrulu gözlerden parazitle
bulaşık olanların oranı da bilinmelidir. Bu amaçla her koloniden sırlanmış yavrulu bir
çerçeve alınarak her yüzünde 50 petek gözünün sırı sökülerek varroa bulaşmış yavru
oranı saptanmıştır (Harris 2007).
Ayrıca her kolonide toplam sırlanmış yavrulu petek gözü sayısın belirlemek için içi tel
ile 10 eşit bölmeye bölünmüş boş bir çerçeve kullanılmıştır. Her kuluçka peteği önlü
arkalı yaklaşık 7000 petek gözü (her yüzünde 3500 petek gözü) içermektedir. Bölünmüş
boş çerçeve sırlanmış petek üzerine yerleştirilerek kapalı yavru sayısı tahmin edilmiştir.
Sonuçta yavruların bulaşıklık oranı ile toplam sırlanmış petek gözü sayısı çarpılarak
kolonideki yavrulara ait toplam varroa bulaşıklığı tahmin edilmiştir. Tüm foretik
aşamadaki ve sırlanmış petek gözlerindeki varroa sayısı toplamı ile her koloniye ait
varroa yoğunluğu hesaplanmıştır (Mondragon vd. 2005).
Kovanın dip tahtasına düşen varroa sayısını belirlemek için deneme kolonilerinde
kovanın dip tahtasına göz çapı 3 milimetre olan tel ızgara yerleştirilmiş, ızgaranın altına
yapışkan beyaz bir kâğıt konulmuştur. Her hafta sonunda parazitler sayıldıktan sonra
beyaz kâğıtlar yenilenmiş ve bu işlem 5 hafta boyunca tekrarlanmıştır (Arechavaleta-
Velasco ve Guzman-Novoa 2001).

31
Şekil 3.2 Kovanın alt kısmına uygulanmış tel ızgara (sağ) ve onun altına konulmuş
yapışkan kağıt ile üzerindeki varroalar (sol).
Varroa zararlısı üreme yapmak için erkek arı petek gözlerini tercih etmektedir. Bunun
nedeni ise erkek arı gözlerinin hacim olarak daha büyük olmasıdır. Varroa zararlısı
sırlanmış işçi arı gözünde ortalama 1.3-1.4 döl yetiştirir iken sırlanmış yavrulu erkek
gözünde 2.2-2.6 döl yetiştirebilmektedir (Calderone 2001, 2006).
Erkek arı oluşumu sürecinde süre uzun, varroa yoğunluğu fazla ve yavru gözü
temizlemede işçi arıların tercihi dişi yavrulu petek gözleri olması nedeniyle denemede
erkek arıların varroa yoğunluğu üzerine olan etkisini gidermek amacıyla kolonilerdeki
sırlanmış erkek arı petek gözleri iki haftada bir temizlenmiştir.
3.2.2 Sır sökme ve petek gözü temizleme (SSPT) davranışının ölçülmesi
Araştırmada bu özellik sıvı azot ile dondurulmuş belirli sayıda yavrulu petek gözünü
kovan içerisinde izleyerek, 24, 48 ve 72 saatlik sürelerde sırı sökülen ve içi boşaltılan
gözlerin sayıları ile ölçülmüştür. Bunun için 9 santimetre çapında ve 10 santimetre
yüksekliğinde (hacmi 636 cm3) olan bir PVC boru kullanılmıştır. Silindirin iç alanında
yerleşen tüm yavruların (210 petek gözü) dondurularak öldürülmesi için en az silindir
hacminin 1/3’ü kadar sıvı azot yeterli olmaktadır. Testleri yapmak için her deneme
kolonisinde petek gözü yeni sırlanmış bir petek seçildikten sonra PVC boru sırlanmış

32
petek gözleri üstüne bastırılmış ve sıvı nitrojen yavaş yavaş silindirin içine dökülerek
sırlanmış petek gözlerindeki yavrular dondurularak öldürülmüştür.
Şekil 3.3 Sırlanmış petek gözlerindeki yavruların sıvı azot ile dondurarak öldürülmesi
Sıvı nitrojen buharlaştıktan sonra PVC silindir kaldırılıp yavruları dondurularak
öldürülmüş petek deneme kolonilerinde yavru alanının ortasına verilmiş; 24, 48 ve 72
saat sonra çerçeve dışarı alınarak sırı sökülmüş ve temizlenmiş petek gözleri sayılmıştır.
(Spivak ve Reuter 2005).
Bu sayımları sayısal olarak analiz etmek için aşağıdaki eşitlik kullanılmıştır:
X(T=24,48,72) = [C (T=0) - C (T=24,48,72)] /C (T=0)
X(T=24,48,72) = 24, 48 ve 72 saat içerisinde sırı sökülmüş ve temizlenmiş petek gözü oranı
C = sırlanmış petek gözü sayısı
U = sırı sökülmüş ve temizlenmiş petek gözü sayısı
T=0: sıvı azot kullanıldığı zaman
T=24, 48 ve 72: sıvı azot kullanıldıktan 24, 48 ve 72 saat sonrası
(Wilkes ve Oldroyd 2002).

33
Bu işlemler hem ebeveyn grubu hem de döl generasyonuna ait kolonilerde
uygulanmıştır.
3.2.3 Varroaya karşı duyarlı hijyen (VDH) davranışın ölçülmesi
Bu davranışın testine başlamadan 6 hafta önce deneme kolonilerinde varroa mücadelesi
yapmak üzere her koloniye etkili maddesi fluvalinate olan birer şerit verilmiştir. Ayrıca
yavru çürüklüğü hastalığını önlemek için kolonilere 1:1 oranında hazırlanmış su ve toz
şeker şurubuyla her 15 günde bir antibiyotik uygulanmıştır. Fluvalinate içeren şeritler
çıkarıldıktan sonra VDH davranışını incelemek üzere yüksek derecede varroa bulaşık
kolonilerden petek gözleri yeni kapatılmış petekler alınarak deneme kolonilerine
verilmiştir. Ayrıca bu peteklerin bulaşıklık oranı (1 ya da 1’den fazla parazite bulaşmış
petek gözlerinin toplamı) her çerçevenin boyuna bir hat üzerinde (çerçevenin her
yüzünde 50 petek gözü) olmak üzere toplam 100 adet yavrulu petek gözü incelenerek
belirlenmiştir.
Şekil 3.4 Sırlanmış petek gözlerinde varroa bulaşıklığının tespit edilmesi
Deneme kolonilerinde yavru oranının değişmesini engellemek için önce her koloniden
birer yavrulu çerçeve çıkartılmıştır. Bir hafta sonra varroa bulaşık çerçevelerde bulaşık

34
petek gözleri aynı yöntemle sayılmıştır. Varroa bulaşmış petek gözü sayısı varroa
bulaşmamış petek gözü sayısına oranı denemenin başlangıcında ve deneme
başlangıcından bir hafta sonra hesaplanmıştır. Böylece bu oranların karşılaştırılması ile
petek gözlerindeki varroa bulaşıklığı arasındaki değişim miktarı elde edilmiştir. Varroa
bulaşıklık düzeyindeki değişim miktarı koloniye ait VDH özelliğinin göstergesidir.
3.2.4 Kalıtım derecesi
Bilindiği üzere; seleksiyonla bir generasyonda sağlanan ilerleme, kalıtım derecesi ve
seleksiyon üstünlüğünün bir fonksiyonudur:
∆G = i . h2
Bu eşitlikten : h2 = ∆G/ i
bulunur. Görülüyor ki seleksiyonla bir generasyonda elde edilen değişme (∆G) ile
seleksiyon üstünlüğü (i) bilindiğinde kalıtım derecesi hesaplanabilir. Ancak kalıtım
derecesini bu yöntemle hesaplayabilmek için generasyonlar boyu çevre etkisinin
değişimini etkisiz hale getirmek gerekir. Bu yüzden bu hata kaynağını etkisiz bırakmak
için aynı populasyondan seleksiyona başlamadan önce bir grup hayvan rasgele ayrılıp
rasgele çiftleştirme ile kontrol grubu oluşturulmaktadır ve devam ettirilmelidir. Kalıtım
derecesi ebeveynler arasındaki farktan döllere geçen kısım olarak da tanımlanmaktadır.
Bu tanım matematik olarak:
2 ( ) / ( )S K S Kh Y Y P P= − −
şeklinde ifade edilebilir. SY = seçilen ebeveyn grubunun döllerine ait ortalama, KY =
seçilmeyen, kontrol grubunun döllerine ait ortalama, SP ve KP de sırasıyla sözü edilen
ebeveyn gruplarına ait ortalamalardır (Düzgüneş vd. 2003).

35
Üzerinde durulan özellikler yalnız dişilerde (işçi arılarda) görüldüğünden erkekler
tarafında seleksiyon üstünlüğü tespit edilemez. Bu engeli aşmak, yani baba olarak ana
arılarla çiftleşen erkek arıların döllerdeki etkilerini bütün döl gruplarında eşitlemek için
bütün ana arıların aynı çiftleşme bölgesinde çiftleşmesi sağlanmış ve kalıtım
derecelerinin tahmininde de aşağıdaki eşitlik kullanılmıştır:
2 2( ) / ( )S K S Kh Y Y P P= − −
3.2.5 İstatistik analizler
Kovanın dip tahtasına düşen varroa sayısına (KDV), kolonideki arı yoğunluğu (KAY),
foretik aşamada olan varroa sayısı (FAVS), varroa bulaşık toplam yavru sayısı (VBYS)
ve sırlanmış petek gözü sayısı (SYPS) etkilidir. Bu nedenle araştırmada KDV değişkeni
ile KAY, FAVS, VBYS ve SYPS değişkenlerinin arasındaki ilişkilerin incelenmesi için
korelasyon katsayıları, İz-(Path)-katsayısı ve belirleme katsayısı hesaplanmıştır.
Şekil 3.5 FAVO ve VBYO faktörleriyle ADV karakteri arasındaki ilişkiler

36
İz-katsayılarını elde etmek için kullanılan eşitliklerin daha kolay izlenmesi için şekil
3.5’te sonuç X, sebepler de A ve B harfleriyle gösterilmiştir. Bu durumda:
PXA = ( rxA – rAB rxB ) / (1 – r2AB)
PXB = ( rxB – rAB rxA ) / (1 – r2AB)
eşitlikleri kullanılarak İz-katsayıları hesaplanmıştır. Bu eşitliklerde:
PXA = ADV’ye (X’e) foretik aşamadaki varroa oranından (FAVO) gelen İz-katsayısı ve
PXB = ADV’ye (X’e) varroa bulaşık yavru oranından (VBYO) gelen İz-katsayısıdır.
ADV’nin FAVO ve VBYO tarafından toptan belirleme katsayısı ise aşağıdaki eşitlikten
hesaplanmıştır:
R2X.AB = P2
XA + P2XB + 2rABPXAPXB
Bu eşitlikte:
R2X.AB = ADV’nin FAVO ve VBYO tarafından toptan belirleme katsayısı, P2
XA=
ADV’nin FAVO tarafından belirleme katsayısı, P2XB= ADV’nin VBYO tarafından
belirleme katsayısı, rAB= FAVO ile VBYO arasındaki korelasyon katsayısıdır.
Üzerinde durulan özellikler bakımından elde edilen gözlemler varyans analizi tekniğinin
ön şartlarını sağlamadığı için Kruskal-Wallis testi ile değerlendirilmiştir. Denemede
yöre faktörünün A, G, K, N, S olmak üzere 5 hali mevcuttur. İncelenen özellikler
bakımından yöreler arasındaki farklılıkların saptanmasında parametrik olmayan
Kruskal-Wallis testinin ikili karşılaştırma (All Pairwise Comparisons) yöntemi
kullanılmıştır. Ayrıca, çoklu karşılaştırma analizlerinin sonuçlarına dayanarak yöreler
arasında ve içerisinde incelenen özellikler ve değişkenler bakımından fark görüldüğü
yerlerde Spearman’ın sıralama korelasyon katsayıları hesaplanmıştır.

37
4. ARAŞTIRMA BULGULARI
4.1 Başlangıç Generasyonda Tımarlama Davranışı (TD) ile İlgili Özellikler
Önceden belirtildiği gibi başlangıç generasyonda 50, 1. generasyonda ise 60 kolonide
arı yoğunluğu (KAY), foretik aşamada olan varroa oranı (FAVO), varroa bulaşık yavru
oranı (VBYO), sırlanmış yavrulu petek gözü sayısı (SYPS) ve 5 hafta boyunca kovanın
dip tahtasına düşen varroa sayısı (KDV) incelenmiştir. Analizlerde KDV değişkeni
kovandaki 1000 arı başına dip tahtasına düşen varroa (ADV) sayısı olarak
değerlendirilmiş, arı yoğunluğu bakımından standardizasyon yapıldığı varsayılmıştır.
Tımarlama davranışı ile ilgili olan özelliklere ait tanımlayıcı değerler çizelge 4.1’de
verilmiştir.
Çizelge 4.1 Başlangıç generasyonunda tımarlama davranışı ile ilgili olan özelliklere ilişkin tanımlayıcı istatistikler (n= 50 Koloni)
Değişken Ortalama ± Standart Hata Varyasyon Katsayısı,% KAY 16170 ± 599 26.19
Arılı petek sayısı 5.4 ± 0.20 26.19 SYPS 3840 ± 212 39.01
Örneklenen arı sayısı 198.6 ± 9.47 33.73 Örneklerde foretik aşamadaki varroa
sayısı
9.6 ± 1.42 103.86
FAVO,% 5 ± 0.7 105.09 FAVS 802 ± 127 112.43
VBYO,% 7 ± 1.0 106.88 VBYS 248.2 ± 39.00 111.21 KDV 371.5 ± 59.90 114.02 ADV 24.6 ± 3.95 113.62
Kovan dip tahtasına düşen varroa sayısı ile ergin arı sayısı ve sırlanmış yavrulu petek
gözü sayısı arasında korelasyonun önemli bulunmaması nedeniyle bu iki değişken
sonuca etkili faktörler olarak değerlendirilmemiştir.

38
Varroa zararlısının istilasına uğramış bir bal arısı kolonisinde akarın kovan dibine
düşmesinin nedenlerinden önde geleni arıların hijyenik davranışlarıdır. Bunun
göstergelerinden biri olduğu yaklaşımıyla araştırmada tabana düşen varroalar
sayılmıştır. Ancak tabana düşen varroa sayısı bakımından bölgeler arası farkın önemli
bulunmaması (Çizelge 4.2) bu özellik ile ilişkili başka faktörlerin dikkate alınmasını
gerektirmiştir. Bu nedenle ergin arılarda FAVO ve Varroa bulaşık yavru oranlarının
(VBYO) analizi yapılmıştır.
Çizelge 4.2 Başlangıç generasyonunda 1000 arı başına kovanın dip tahtasına düşen varroa sayısının yörelere göre tanımlayıcı istatistikleri ve Kruskal-Wallis testi sonuçları (n=10 koloni)
Yöre Ortalama ± Standart Hata Minimum Maksimum Rank ortlamaları
A 25.6 ± 8.43 2.67 90.33 27.3 G 10.0 ± 2.60 2.31 27.53 15.4 K 32.0 ± 14 5.00 129.6 23.8 N 25.1 ± 6.91 6.13 77.4 30.0 S 30.3 ± 8.44 5.33 91.56 31.1
SD = 4, P > 0.05
Yörelere göre yapılan analizlerde kovanın dip tahtasına düşen varroa sayısıyla
kolonideki arı yoğunluğu ve sırlanmış yavrulu petek gözü sayısı arasında istatistik
olarak önemli korelasyon bulunmamıştır. Kruskal-Wallis testinin sonuçları (Çizelge
4.2) ADV bakımından yöreler arası farkın önemli olmadığını göstermiştir. ADV, FAVO
ve VBYO değişkenleri arasında korelasyonlar önemli bulunmuştur (Çizelge 4.3). Bu
yüzden tımarlama davranışının tespitinde, bu davranışı daha iyi ifade eden değişkeni
belirlemek için söz konusu değişkenlerin arasındaki İz-katsayıları ve belirleme
katsayıları hesaplanmıştır. Şekil 4.1’de incelenen özellik olan 1000 arı başına kovanın
dip tahtasına düşen varroa sayısı (ADV) ve bunu etkileyen nedenler (FAVO ve VBYO)
ile aralarındaki ilişki gösterilmiştir.

39
Çizelge 4.3 Başlangıç generasyonunda tımarlama ile ilgili özelliklerin arasındaki Spearman’ın sıralama korelasyon katsayıları (n=50 koloni)
Değişken KAY SYPS FAVO VBYO
SYPS 0.450** - - -
FAVO -0.083 -0.236 - -
VBYO 0.007 -0.270 0.703** -
ADV -0.235 -0.186 0.314* 0.442** ** P< 0.01, * P< 0.05
İz-katsayılarının hesaplanmasında 3.2.5 bölümünde açıklanan eşitlikler kullanılmıştır.
ADV’ye foretik aşamadaki varroa oranından (FAVO) gelen İz-katsayısı 0.14, varroa
bulaşık yavru oranından (VBYO) gelen ise 0.53 olarak bulunmuştur. Belirleme
katsayıları da İz-katsayılarının karesi olarak sırasıyla 0.019 ve 0.281’dir. ADV’nin
FAVO ve VBYO tarafından toptan belirleme katsayısı ise 0.425 olarak hesaplanmıştır:
Kruskal-Wallis testi sonuçlarına göre başlangıç generasyonun kolonilerinde foretik
aşamadaki varroa oranı bakımından yöreler arasındaki fark önemli bulunmuştur (p <
0.01) (Çizelge 4.4).
Çizelge 4.4 Başlangıç generasyonunda foretik aşamada olan varroa oranının yörelere göre tanımlayıcı istatistikleri ve Kruskal-Wallis testi sonuçları (n=10 koloni)
Yöre Ortalama ± Standart Hata
Minimum Maksimum Rank ortalamaları
A 5 ± 1.1 2.3 14.7 31.1 A G 1 ± 0.4 0.0 4.3 12.3 B K 10 ± 2.5 0.0 23.1 34.6 A N 5 ± 1.5 0.6 16.9 27.2 A S 3 ± 0.8 0.6 7.0 22.3 AB
SD = 4, P < 0.01
Ergin arılarda varroa bulaşıklığı (foretik aşaması) bakımından G ile A, K ve N yöreleri
arasında farklar istatistik olarak önemli (p < 0.01) bulunmuştur (Çizelge 4.4).

40
Çeşitli yörelerden alınan kolonilerde, yavruların varroa bulaşıklığına ilişkin veriler
çizelge 4.5’da verilmiştir. Kruskal-Wallis testi ve yörelerin ikili karşılaştırma
sonuçlarına göre yavruların varroa bulaşıklığı bakımından G yöresi ile diğer yöreler
arasında farklar (p < 0.05, 0.01) önemlidir (Çizelge 4.6).
Çizelge 4.5 Başlangıç generasyonunda varroa bulaşık yavru oranının yörelere göre tanımlayıcı istatistikleri ve Kruskal-Wallis testi sonuçları (n=10 koloni)
Yöre Ortalama ± Standart Hata Minimum Maksimum Rank
Ortlamaları A 6 ± 1.0 0 11 29.3 A G 2 ± 0.5 0 6 10.9 B K 12 ± 4.0 0 40 31.6 A N 9 ± 2.3 0 22 30 A S 6 ± 1.2 0 12 25.7 A
SD = 4, P < 0.01
Çizelge 4.6 Başlangıç generasyonunda varroa bulaşık yavru oranı bakımından yöreler arasında ikili karşılaştırma analizi (p değerleri)
Yöre A G K N
G 0.005** - - -
K 0.717 0.001** - -
N 0.908 0.003** 0.805 -
S 0.585 0.023* 0.364 0.508
Farklı yörelerden alınan kolonilerde incelenen özellikler arasındaki korelasyon
katsayıları çizelge 4.7’de verilmiştir. Elde edilen değerler, tüm deneme kolonilerinde
hesaplanan korelasyon katsayılarıyla uyum sağlamaktadır. Yöreler içerisinde tahmin
edilen korelasyon katsayılarına göre yavrular ile ergin arılardaki varroa bulaşıklığı
yüksek derecede ilişkili bulunmuştur.
Denemenin başında tüm kolonilerde foretik aşamadaki varroa sayımları yapılmıştır.
Başlangıçta ölçülen foretik aşamadaki varroa sayısı ile 5 hafta boyunca haftada bir
kovanın dip tahtasına düşen varroa sayısı arasındaki ilişki ölçülmüştür.

41
Buna göre beşinci hafta sayımı ile foretik bulaşıklık arasında korelasyon r =0.295
(p<0.05; n=50) iken başlangıçta ölçülen foretik aşamadaki varroa sayısı ile birinci hafta
sonunda kovanın dip tahtasına düşen varroa sayısı arasındaki korelasyon r =0.347
(p<0.05; n=50) bulunmuştur. Korelasyonun birinci hafta için yüksek olması nedeniyle
döl generasyonunda 2 haftalık sayım yeterli görülmüştür.
Çizelge 4.7 Başlangıç generasyonunda tımarlama davranışı ile ilgili olan özelliklerin arasında her yöre için Spearman’ın sıralama korelasyon katsayıları
İlişkili Özellikler Yöreler
A G K N S
FAVO VBYO 0.745* -0.372 0.952** 0.954** 0.043
FAVO ADV 0.427 0.176 0.539 0.491 -0.061
ADV VBYO 0.279 -0.169 0.539 0.593* 0.202
A=Tabriz, G=Azarshahr, K=Ahar, N=Bostanabad, S=Shabestar; * P< 0.05 ve ** P< 0.01
4.2 Başlangıç Generasyonda Sır Sökme ve Petek Gözü Temizleme Davranışı
(SSPT)
Bal arılarının hastalıklara, parazitlere ve her türlü bulaşıklığa karşı gösterdikleri tepki
hijyenik davranış adı altında toplanır. Sırlanmış petek gözlerindeki hasta ve ölü
yavruları temizleme özelliği de hijyenik davranışın önemli unsurlarından biridir. Bu
özelliğin ölçümü ile elde edilen verilere ilişkin tanımlayıcı değerler çizelge 4.8’de
verilmiştir. Ölçümler, sıvı azot ile öldürülmüş yavrulu petek kovanlara verildikten 24,
48 ve 72 saat sonra yapılmış, 24 saat içerisinde temizlenen petek gözü sayısı diğer
zamanlara göre daha yüksek değişim (CV) göstermiştir.

42
Çizelge 4.8 Başlangıç generasyonun SSPT grubuna ait tanımlayıcı istatistikler (n=25 koloni)
Değişken Ortalama ± Standart Hata Varyasyon Katsayısı,%
KAY 16680 ± 876 26.26 SYPS 3584 ± 308 43.01 FAVS 1164 ± 219 94.19
FAVO,% 7 ± 1.2 89.19 VBYS 378.8 ± 66.3 87.56
VBYO,% 10 ± 1.7 78.99 24 saat içerisinde
temizlenmiş yavrulu petek gözü oranı,%
63 ± 4.7
37.01
48 saat içerisinde temizlenmiş yavrulu petek
gözü oranı,%
90 ± 2.0
11.41
72 saat içerisinde temizlenmiş yavrulu petek
gözü oranı,%
97 ± 0.9
5.05
KDV 453 ± 110 121.11 ADV 30.5 ± 7.23 118.56
Dondurularak öldürülmüş yavruların petek gözlerinden temizlenmesi bakımından elde
edilen gözlemler faktöriyel düzende tekrarlanan ölçümlü varyans analizi tekniği
(Repeated measurement ANOVA) ile değerlendirilmiştir. Denemede yöre faktörünün A,
G, K, N, S olmak üzere 5 seviyesi ve süre faktörünün 24 saat, 48 saat ve 72 saat olmak
üzere 3 seviyesi ile ölçüm yapılmıştır. Tekrarlanan ölçümler zaman faktörünün
seviyelerinde gerçekleştirilmiştir (Çizelge 4.9). Analizleri yapmak üzere aşağıdaki
model oluşturulmuştur:
yijk = µ + bi + dk(i) + cj + bcij + cdjk(i) + eijk
(i = 1, 2, …, l; j = 1, 2, …, m ve k = 1, 2, …, ni)
Buradan :
yijk : i. yörede k. koloninin j. sürede ölçüm değeri,
µ : populasyon ortalaması
bi : yöre faktörünün i. halinde etki miktarı,

43
dk(i) = yöre faktörünün i. halinde yer alan k. koloninin rasgele etkisi,
cj = süre faktörünün j. seviyesinin etki miktarı,
bcij = yöre × süre interaksiyonun etkisi,
cdjk(i) = yöre faktörünün i. hali için süre × koloni interaksiyonu,
eijk = tesadüfi çevre faktörlerinin etkisi (N, 0, б2e).
Yapılan varyans analizi sonucunda Süre*Yöre interaksiyonu (P<0.01) ve yöreler
arasında yavrulu petek gözü temizleme süresi bakımından istatistik olarak önemli fark
bulunmuştur (P<0.01) (Çizelge 4.9).
Çizelge 4.9 Başlangıç generasyonun SSPT grubunda temizleme süreleri bakımından yöreler içerisinde tekrarlanan ölçümlü varyans analizinin (Repeated measurment ANOVA) sonuçları
Varyasyon
kaynakları
Serbestlik
derecesi
Kareler
toplamı
Kareler
ortalaması
F P
Yöre 4 0.435 0.109 4.40 0.01
Hata1 20 0.494 0.025 - -
Süre 2 1.573 0.787 92.35 0.000
Süre*Yöre 8 0.376 0.047 5.51 0.000
Hata2 40 0.341 0.009 - -
Arılıklar ortalamaları arasındaki farklar Duncan testi ile irdelenmiştir. Yapılan varyans
analizi sonuçlarına göre S yöresinin kolonilerinde 24 saat içerisinde temizlenmiş petek
gözü oranı ortalaması diğer bölgelere kıyasla önemli fark göstermiştir (p < 0.05).
Duncan testi sonuçları ortalamaların harfli gösterimi ile çizelge 4.10’da verilmiştir.
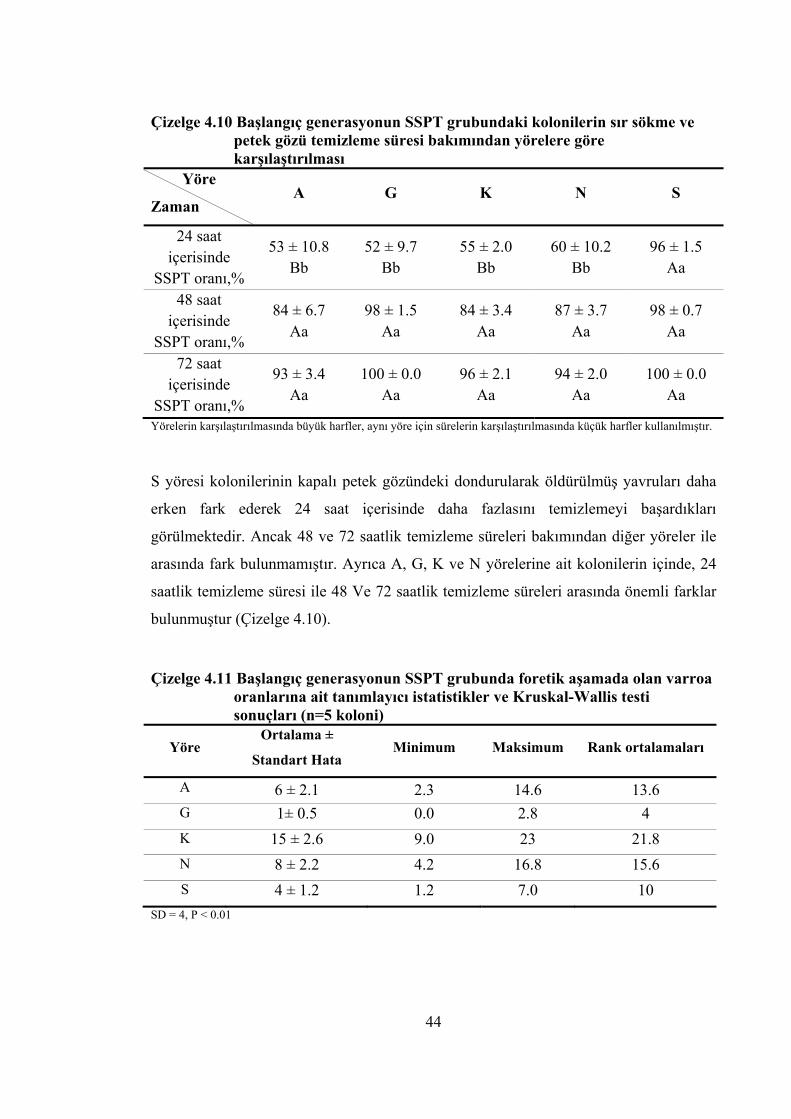
44
Çizelge 4.10 Başlangıç generasyonun SSPT grubundaki kolonilerin sır sökme ve petek gözü temizleme süresi bakımından yörelere göre karşılaştırılması
Yöre
Zaman A G K N S
24 saat içerisinde
SSPT oranı,%
53 ± 10.8 Bb
52 ± 9.7 Bb
55 ± 2.0 Bb
60 ± 10.2 Bb
96 ± 1.5 Aa
48 saat içerisinde
SSPT oranı,%
84 ± 6.7 Aa
98 ± 1.5 Aa
84 ± 3.4 Aa
87 ± 3.7 Aa
98 ± 0.7 Aa
72 saat içerisinde
SSPT oranı,%
93 ± 3.4 Aa
100 ± 0.0 Aa
96 ± 2.1 Aa
94 ± 2.0 Aa
100 ± 0.0 Aa
Yörelerin karşılaştırılmasında büyük harfler, aynı yöre için sürelerin karşılaştırılmasında küçük harfler kullanılmıştır.
S yöresi kolonilerinin kapalı petek gözündeki dondurularak öldürülmüş yavruları daha
erken fark ederek 24 saat içerisinde daha fazlasını temizlemeyi başardıkları
görülmektedir. Ancak 48 ve 72 saatlik temizleme süreleri bakımından diğer yöreler ile
arasında fark bulunmamıştır. Ayrıca A, G, K ve N yörelerine ait kolonilerin içinde, 24
saatlik temizleme süresi ile 48 Ve 72 saatlik temizleme süreleri arasında önemli farklar
bulunmuştur (Çizelge 4.10).
Çizelge 4.11 Başlangıç generasyonun SSPT grubunda foretik aşamada olan varroa oranlarına ait tanımlayıcı istatistikler ve Kruskal-Wallis testi sonuçları (n=5 koloni)
Yöre Ortalama ±
Standart Hata Minimum Maksimum Rank ortalamaları
A 6 ± 2.1 2.3 14.6 13.6 G 1± 0.5 0.0 2.8 4 K 15 ± 2.6 9.0 23 21.8 N 8 ± 2.2 4.2 16.8 15.6 S 4 ± 1.2 1.2 7.0 10
SD = 4, P < 0.01
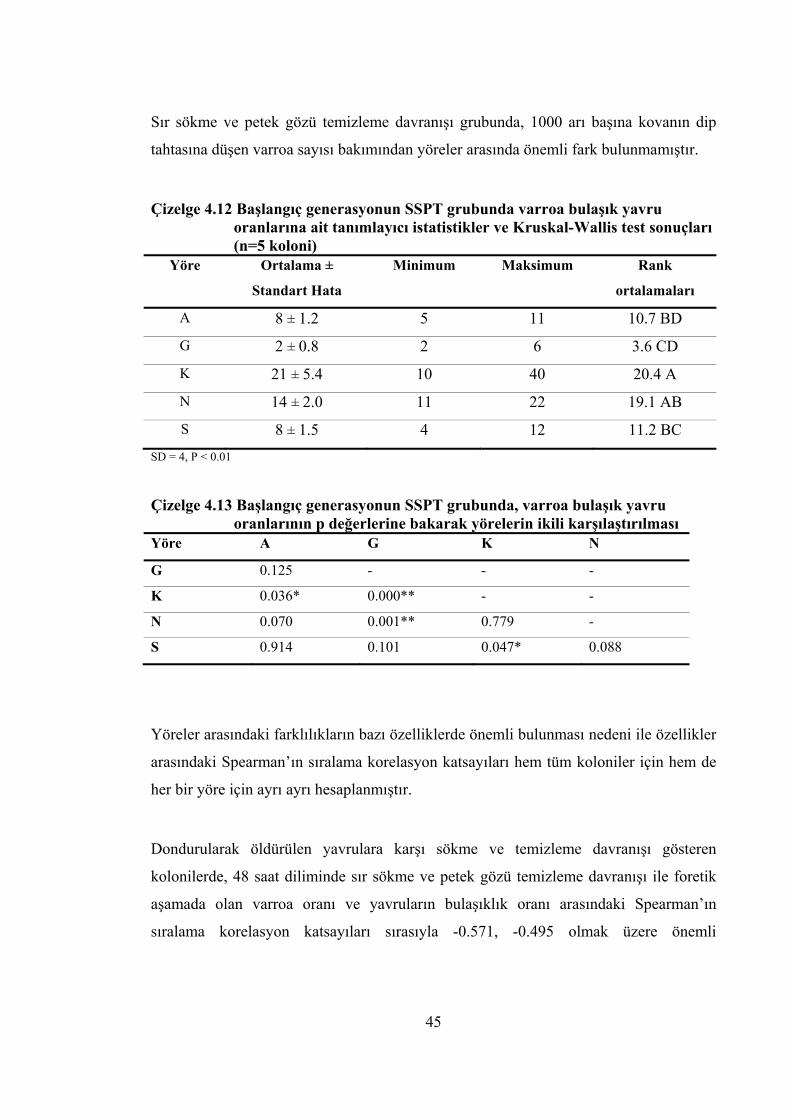
45
Sır sökme ve petek gözü temizleme davranışı grubunda, 1000 arı başına kovanın dip
tahtasına düşen varroa sayısı bakımından yöreler arasında önemli fark bulunmamıştır.
Çizelge 4.12 Başlangıç generasyonun SSPT grubunda varroa bulaşık yavru oranlarına ait tanımlayıcı istatistikler ve Kruskal-Wallis test sonuçları (n=5 koloni)
Yöre Ortalama ±
Standart Hata
Minimum Maksimum Rank
ortalamaları
A 8 ± 1.2 5 11 10.7 BD
G 2 ± 0.8 2 6 3.6 CD
K 21 ± 5.4 10 40 20.4 A
N 14 ± 2.0 11 22 19.1 AB
S 8 ± 1.5 4 12 11.2 BC SD = 4, P < 0.01
Çizelge 4.13 Başlangıç generasyonun SSPT grubunda, varroa bulaşık yavru oranlarının p değerlerine bakarak yörelerin ikili karşılaştırılması
Yöre A G K N
G 0.125 - - -
K 0.036* 0.000** - -
N 0.070 0.001** 0.779 -
S 0.914 0.101 0.047* 0.088
Yöreler arasındaki farklılıkların bazı özelliklerde önemli bulunması nedeni ile özellikler
arasındaki Spearman’ın sıralama korelasyon katsayıları hem tüm koloniler için hem de
her bir yöre için ayrı ayrı hesaplanmıştır.
Dondurularak öldürülen yavrulara karşı sökme ve temizleme davranışı gösteren
kolonilerde, 48 saat diliminde sır sökme ve petek gözü temizleme davranışı ile foretik
aşamada olan varroa oranı ve yavruların bulaşıklık oranı arasındaki Spearman’ın
sıralama korelasyon katsayıları sırasıyla -0.571, -0.495 olmak üzere önemli
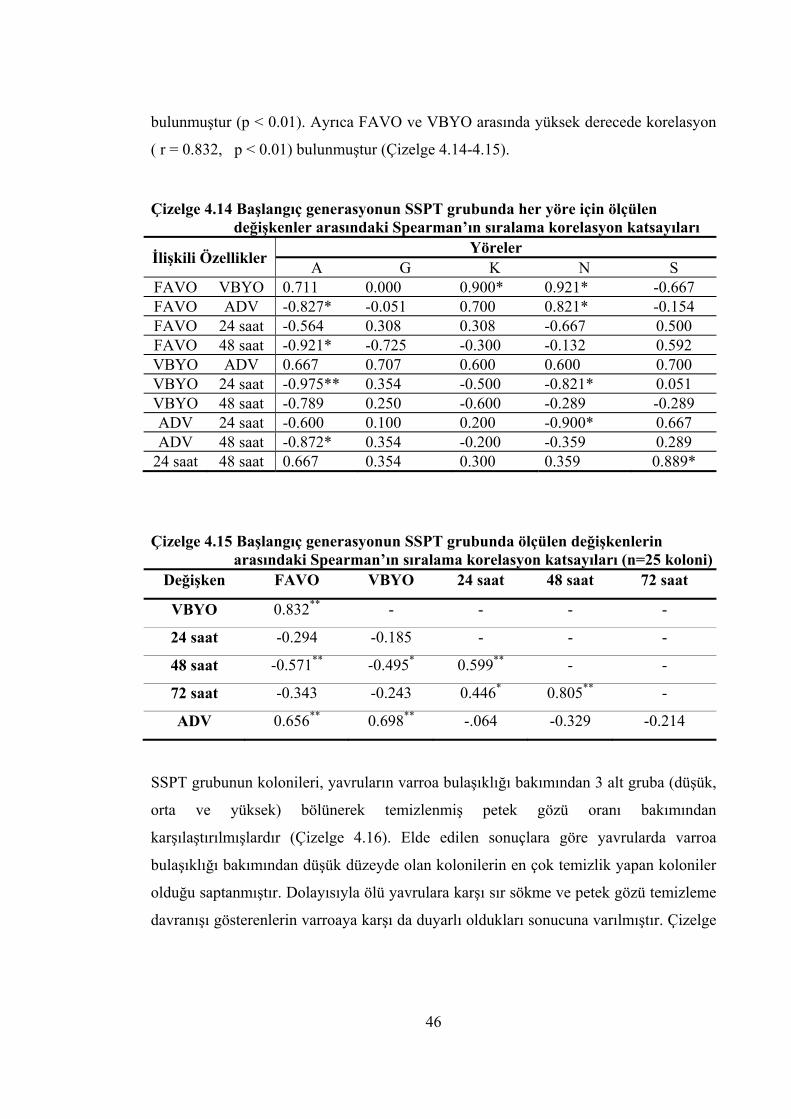
46
bulunmuştur (p < 0.01). Ayrıca FAVO ve VBYO arasında yüksek derecede korelasyon
( r = 0.832, p < 0.01) bulunmuştur (Çizelge 4.14-4.15).
Çizelge 4.14 Başlangıç generasyonun SSPT grubunda her yöre için ölçülen değişkenler arasındaki Spearman’ın sıralama korelasyon katsayıları
İlişkili Özellikler Yöreler A G K N S
FAVO VBYO 0.711 0.000 0.900* 0.921* -0.667 FAVO ADV -0.827* -0.051 0.700 0.821* -0.154 FAVO 24 saat -0.564 0.308 0.308 -0.667 0.500 FAVO 48 saat -0.921* -0.725 -0.300 -0.132 0.592 VBYO ADV 0.667 0.707 0.600 0.600 0.700 VBYO 24 saat -0.975** 0.354 -0.500 -0.821* 0.051 VBYO 48 saat -0.789 0.250 -0.600 -0.289 -0.289 ADV 24 saat -0.600 0.100 0.200 -0.900* 0.667 ADV 48 saat -0.872* 0.354 -0.200 -0.359 0.289
24 saat 48 saat 0.667 0.354 0.300 0.359 0.889*
Çizelge 4.15 Başlangıç generasyonun SSPT grubunda ölçülen değişkenlerin arasındaki Spearman’ın sıralama korelasyon katsayıları (n=25 koloni)
Değişken FAVO VBYO 24 saat 48 saat 72 saat
VBYO 0.832** - - - -
24 saat -0.294 -0.185 - - -
48 saat -0.571** -0.495* 0.599** - -
72 saat -0.343 -0.243 0.446* 0.805** -
ADV 0.656** 0.698** -.064 -0.329 -0.214
SSPT grubunun kolonileri, yavruların varroa bulaşıklığı bakımından 3 alt gruba (düşük,
orta ve yüksek) bölünerek temizlenmiş petek gözü oranı bakımından
karşılaştırılmışlardır (Çizelge 4.16). Elde edilen sonuçlara göre yavrularda varroa
bulaşıklığı bakımından düşük düzeyde olan kolonilerin en çok temizlik yapan koloniler
olduğu saptanmıştır. Dolayısıyla ölü yavrulara karşı sır sökme ve petek gözü temizleme
davranışı gösterenlerin varroaya karşı da duyarlı oldukları sonucuna varılmıştır. Çizelge

47
4.16’da düşük, orta ve yüksek gruplara ayrılan koloniler sırasıyla A, B ve C harfleri ile
gösterilmiştir.
Çizelge 4.16 Başlangıç generasyonun SSPT grubunda 3 alt grubun tanımlayıcı istatistikleri ve Kruskal-Wallis testinin sonuçları
Varroa Bulaşık Yavru Oranı,% Grup Koloni
Sayısı Ortalama ±
Standart Hata Minimum Maksimum Rank
ortalamaları A 9 3 ± 0.6 2 6 5.0 C B 8 10 ± 0.6 7 12 13.6 B C 8 19 ± 3.4 12 40 21.4 A
48 Saat Süre İçerisinde Temizlenmiş Petek Gözü Oranı,% A 9 99 ± 0.8 92 100 19.6 A B 8 88 ± 4.5 66 100 11.6 B C 8 83 ± 1.9 75 90 7.1 C
SD = 2 P < 0.01, SD = 2 P< 0.01
4.3 Başlangıç Generasyonda Varroaya Karşı Duyarlı Hijyen (VDH) Davranışı
VDH özelliği araştırılan deneme gurubunda, elde edilen verilere ilişkin tanımlayıcı
değerler çizelge 4.17’de verilmiştir. Bu özelliği belirlemek için ölçülen değişkenler
arasında değişim katsayısı büyük olan, sırlanmış petek gözlerindeki varroa
bulaşıklığının azalma miktarı olduğu görülmüştür (Çizelge 4.17). Bu nedenle
bulaşıklığın azalma miktarı bu özelliğin en önemli belirteci olarak ölçülmesi gereken
parametredir.
Kruskal-Wallis testi sonuçlarına göre VDH özelliği bakımından yöreler arasında önemli
fark bulunmamıştır. Değişik bir karşılaştırma yapmak üzere VDH grubunda yer alan
koloniler yavrulardaki varroa bulaşıklığı bakımından 3 alt gruba bölünmüştür. Bu alt
gruplarda yavruların varroa bulaşıklığına ve parazitin petek gözlerinden temizlenmesine
ait tanımlayıcı değerler ile Kruskal-Wallis testinden elde edilen rank ortalamaları
Çizelge 4.18’de verilmiştir. Bu çizelgede düşük, orta ve yüksek gruplara ayrılan
koloniler sırasıyla A, B ve C harfleri ile gösterilmiştir.

48
Çizelge 4.17 Başlangıç generasyonun VDH grubunda ölçülen değişkenlere ait tanımlayıcı istatistikler (n=25 koloni)
Değişken Ortalama ±
Standart Hata
Minimum Maksimum Varyasyon
Katsayısı,%
Varroa bulaşık petek gözü sayısı
24.76 ± 2.610 3.00 49.00 52.66
Varroa bulaşıklığı olmayan petek gözü sayısı
75.24 ± 2.610 51.00 97.00 17.33
Başlangıçta varroa bulaşık petek gözlerinin varroa bulaşık olmayan petek
gözlerine oranı
0.37 ± 0.050 0.03 0.96 68.36
Bir hafta sonra varroa bulaşmış petek gözü sayısı
17.64 ± 1.970 3.00 38.00 55.73
Bir hafta sonra varroa bulaşıklığı olmayan petek
gözü sayısı 82.36 ± 1.970 62.00 97.00 11.94
Bir hafta sonra varroa bulaşık petek gözlerinin varroa bulaşık olmayan petek gözlerine oranı
0.23 ± 0.031 0.03 0.61 67.13
Bulaşıklık oranındaki azalma,%
13 ± 3.1 0.00 47 113.90
VBYO,% 3 ± 0.4 0.00 8 79.35 FAVO,% 2 ± 0.4 0.00 11.3 86.25
ADV 18.68 ± 2.920 2.87 56.89 78.17
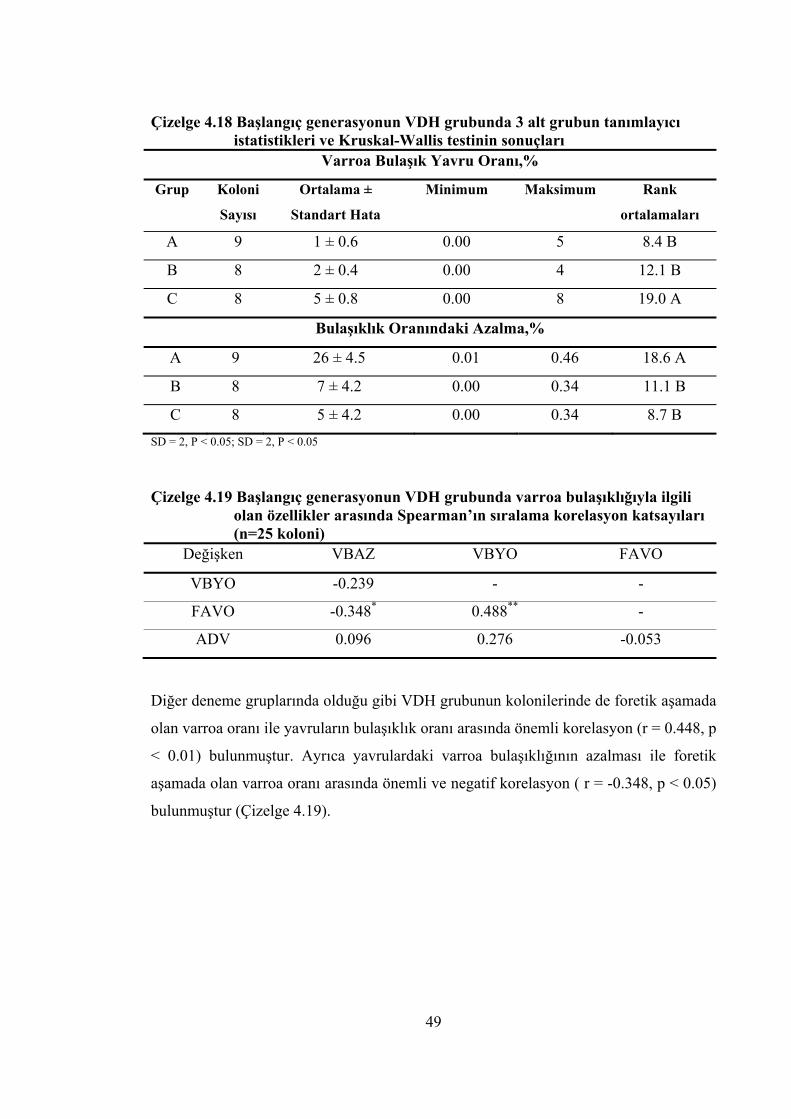
49
Çizelge 4.18 Başlangıç generasyonun VDH grubunda 3 alt grubun tanımlayıcı istatistikleri ve Kruskal-Wallis testinin sonuçları
Varroa Bulaşık Yavru Oranı,%
Grup Koloni
Sayısı
Ortalama ±
Standart Hata
Minimum Maksimum Rank
ortalamaları
A 9 1 ± 0.6 0.00 5 8.4 B
B 8 2 ± 0.4 0.00 4 12.1 B
C 8 5 ± 0.8 0.00 8 19.0 A
Bulaşıklık Oranındaki Azalma,%
A 9 26 ± 4.5 0.01 0.46 18.6 A
B 8 7 ± 4.2 0.00 0.34 11.1 B
C 8 5 ± 4.2 0.00 0.34 8.7 B SD = 2, P < 0.05; SD = 2, P < 0.05
Çizelge 4.19 Başlangıç generasyonun VDH grubunda varroa bulaşıklığıyla ilgili olan özellikler arasında Spearman’ın sıralama korelasyon katsayıları (n=25 koloni)
Değişken VBAZ VBYO FAVO
VBYO -0.239 - -
FAVO -0.348* 0.488** -
ADV 0.096 0.276 -0.053
Diğer deneme gruplarında olduğu gibi VDH grubunun kolonilerinde de foretik aşamada
olan varroa oranı ile yavruların bulaşıklık oranı arasında önemli korelasyon (r = 0.448, p
< 0.01) bulunmuştur. Ayrıca yavrulardaki varroa bulaşıklığının azalması ile foretik
aşamada olan varroa oranı arasında önemli ve negatif korelasyon ( r = -0.348, p < 0.05)
bulunmuştur (Çizelge 4.19).
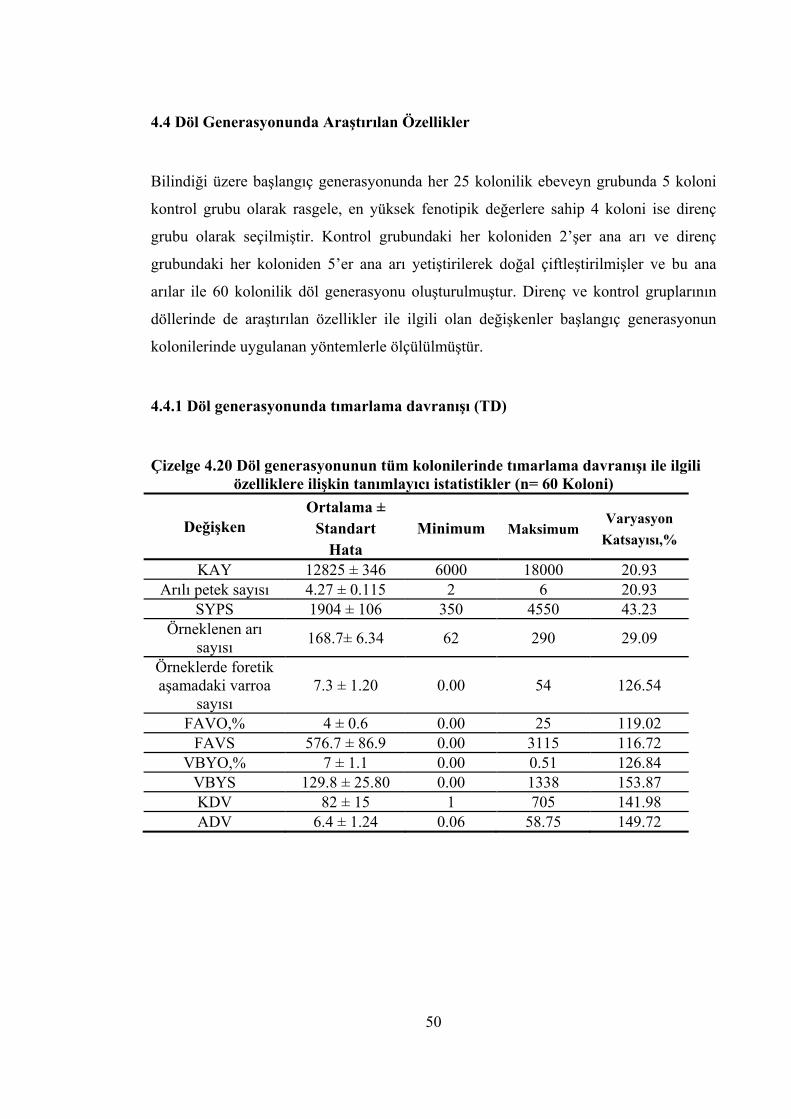
50
4.4 Döl Generasyonunda Araştırılan Özellikler
Bilindiği üzere başlangıç generasyonunda her 25 kolonilik ebeveyn grubunda 5 koloni
kontrol grubu olarak rasgele, en yüksek fenotipik değerlere sahip 4 koloni ise direnç
grubu olarak seçilmiştir. Kontrol grubundaki her koloniden 2’şer ana arı ve direnç
grubundaki her koloniden 5’er ana arı yetiştirilerek doğal çiftleştirilmişler ve bu ana
arılar ile 60 kolonilik döl generasyonu oluşturulmuştur. Direnç ve kontrol gruplarının
döllerinde de araştırılan özellikler ile ilgili olan değişkenler başlangıç generasyonun
kolonilerinde uygulanan yöntemlerle ölçülülmüştür.
4.4.1 Döl generasyonunda tımarlama davranışı (TD)
Çizelge 4.20 Döl generasyonunun tüm kolonilerinde tımarlama davranışı ile ilgili özelliklere ilişkin tanımlayıcı istatistikler (n= 60 Koloni)
Değişken Ortalama ±
Standart Hata
Minimum Maksimum Varyasyon
Katsayısı,%
KAY 12825 ± 346 6000 18000 20.93 Arılı petek sayısı 4.27 ± 0.115 2 6 20.93
SYPS 1904 ± 106 350 4550 43.23 Örneklenen arı
sayısı 168.7± 6.34 62 290 29.09
Örneklerde foretik aşamadaki varroa
sayısı 7.3 ± 1.20 0.00 54 126.54
FAVO,% 4 ± 0.6 0.00 25 119.02 FAVS 576.7 ± 86.9 0.00 3115 116.72
VBYO,% 7 ± 1.1 0.00 0.51 126.84 VBYS 129.8 ± 25.80 0.00 1338 153.87 KDV 82 ± 15 1 705 141.98 ADV 6.4 ± 1.24 0.06 58.75 149.72
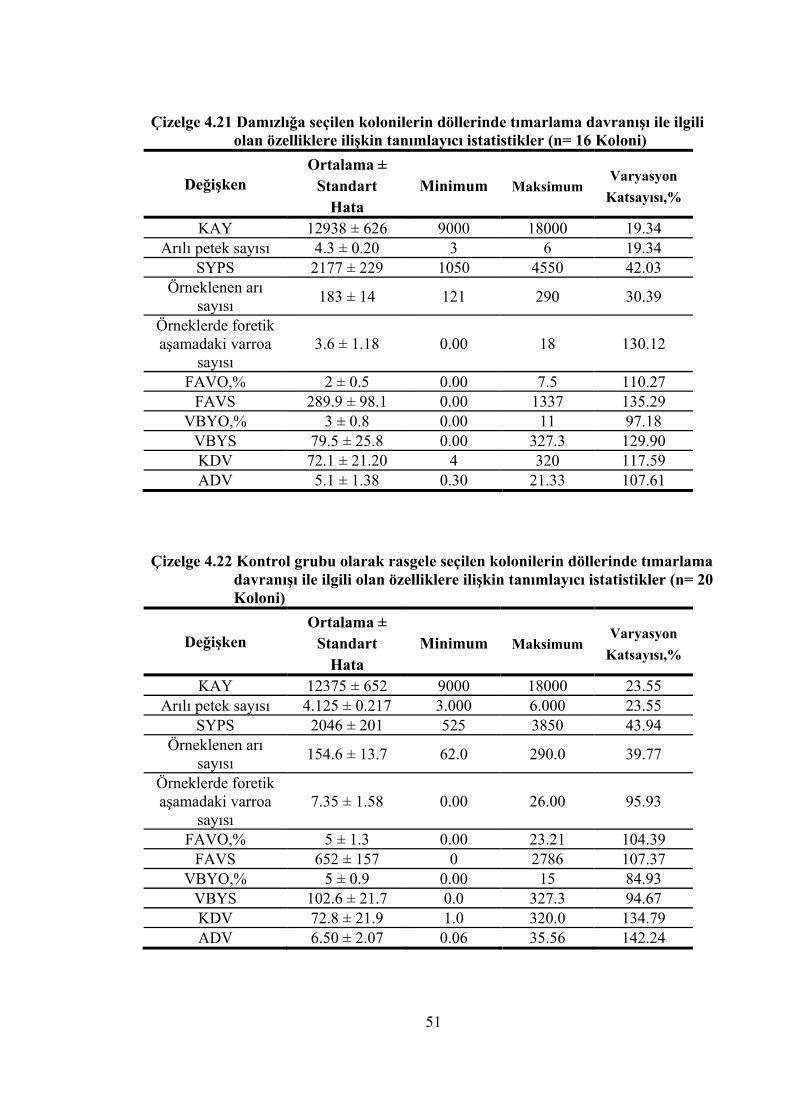
51
Çizelge 4.21 Damızlığa seçilen kolonilerin döllerinde tımarlama davranışı ile ilgili olan özelliklere ilişkin tanımlayıcı istatistikler (n= 16 Koloni)
Değişken Ortalama ±
Standart Hata
Minimum Maksimum Varyasyon
Katsayısı,%
KAY 12938 ± 626 9000 18000 19.34 Arılı petek sayısı 4.3 ± 0.20 3 6 19.34
SYPS 2177 ± 229 1050 4550 42.03 Örneklenen arı
sayısı 183 ± 14 121 290 30.39
Örneklerde foretik aşamadaki varroa
sayısı 3.6 ± 1.18 0.00 18 130.12
FAVO,% 2 ± 0.5 0.00 7.5 110.27 FAVS 289.9 ± 98.1 0.00 1337 135.29
VBYO,% 3 ± 0.8 0.00 11 97.18 VBYS 79.5 ± 25.8 0.00 327.3 129.90 KDV 72.1 ± 21.20 4 320 117.59 ADV 5.1 ± 1.38 0.30 21.33 107.61
Çizelge 4.22 Kontrol grubu olarak rasgele seçilen kolonilerin döllerinde tımarlama davranışı ile ilgili olan özelliklere ilişkin tanımlayıcı istatistikler (n= 20 Koloni)
Değişken Ortalama ±
Standart Hata
Minimum Maksimum Varyasyon
Katsayısı,%
KAY 12375 ± 652 9000 18000 23.55 Arılı petek sayısı 4.125 ± 0.217 3.000 6.000 23.55
SYPS 2046 ± 201 525 3850 43.94 Örneklenen arı
sayısı 154.6 ± 13.7 62.0 290.0 39.77
Örneklerde foretik aşamadaki varroa
sayısı 7.35 ± 1.58 0.00 26.00 95.93
FAVO,% 5 ± 1.3 0.00 23.21 104.39 FAVS 652 ± 157 0 2786 107.37
VBYO,% 5 ± 0.9 0.00 15 84.93 VBYS 102.6 ± 21.7 0.0 327.3 94.67 KDV 72.8 ± 21.9 1.0 320.0 134.79 ADV 6.50 ± 2.07 0.06 35.56 142.24

52
4.4.2 Döl generasyonunda sır sökme ve petek gözü temizleme (SSPT) davranışı
Araştırmada başlangıç generasyonunun SSPT grubundaki kolonilerde ve döl
generasyonunun SSPT bakımında damızlığa seçilen kolonilerinde sır sökme ve petek
gözü temizleme hızını incelemek üzere 24, 48 ve 72 saat dilimlerde kapalı ve
temizlenmemiş petek gözlerinin oranına bakılmıştır. Başlangıç generasyonunun SSPT
grubunun kolonilerinde 24, 48 ve 72 saat dilimlerde sır sökme ve petek gözü temizleme
hızına ait ortalamalar sırasıyla 0.63±0.047, 0.78±0.042 ve 0.85±0.044 iken damızlığa
seçilen kolonilerin döllerinde 24 ve 48 saat dilimlerde sırasıyla 0.66±0.049 ve
0.86±0.040 bulunmuştur.
Çizelge 4.23 Damızlığa seçilen kolonilerin döllerinde SSPT davranışı ile ilgili olan özelliklere ilişkin tanımlayıcı istatistikler (n= 20 Koloni)
Değişken Ortalama ±
Standart Hata
Minimum Maksimum Varyasyon
Katsayısı,%
KAY 13725 ± 568 9000 18000 18.52 SYPS 2011 ± 126 1050 2975 27.96 FAVS 826 ± 178 0.00 3115 96.17
FAVO,% 6 ± 1.4 0.00 2.6 102.78 VBYS 237 ± 67 0.00 1338.8 126.22
VBYO,% 11 ± 2.5 0.00 5.1 100.52 24 saat içerisinde
temizlenmiş yavrulu petek gözü
oranı;%
66.2 ± 4.9 30 100 33.73
48 saat içerisinde temizlenmiş
yavrulu petek gözü oranı,%
93.9 ± 2.0 61.4 100 9.96
KDV 119.7 ± 35.3 3.0 705.0 131.85 ADV 8.84 ± 2.87 0.25 58.75 145.23

53
Çizelge 4.24 Kontrol grubu olarak rasgele seçilen kolonilerin döllerinde SSPT davranışı ile ilgili olan özelliklere ilişkin tanımlayıcı istatistikler (n=10 koloni)
Değişken Ortalama ± Standart
Hata
Minimum Maksimum Varyasyon Katsayısı,%
KAY 11400 ± 900 9000 18000 24.97 SYPS 2485 ± 290 1050 3850 36.94 FAVS 522 ± 156 0.00 1488 94.80
FAVO,% 5 ± 1.2 0.00 14.17 75.73 VBYS 89.4 ± 30.2 0.00 327.30 106.86
VBYO,% 3 ± 1.0 0.00 11 95.51 24 saat içerisinde
temizlenmiş yavrulu petek gözü oranı,%
64.1 ± 5.2 37 93 25.81
48 saat içerisinde temizlenmiş
yavrulu petek gözü oranı,%
91.6 ± 2.1 81 100 7.41
KDV 78.2 ± 33 1.00 320.0 134.85 ADV 7.98 ± 3.6 0.06 35.56 142.58

54
4.4.3 Döl generasyonunda varroaya karşı duyarlı hijyen (VDH) davranışı
Çizelge 4.25 Damızlığa seçilen kolonilerin döllerinde VDH davranışı ile ilgili olan özelliklere ilişkin tanımlayıcı istatistikler (n=20 koloni)
Değişken Ortalama ± Standart
Hata
Minimum
Maksimum
Varyasyon Katsayısı,
% Varroa bulaşık petek gözü sayısı
31.2 ± 1.50 18 44 21.53
Varroa bulaşıklığı olmayan petek gözü sayısı
68.8 ± 1.50
56
82
9.76
Başlangıçta varroa bulaşık petek gözlerinin varroa bulaşık olmayan petek gözlerine oranı
0.47 ± 0.032
0.21
0.78
31.37
Bir hafta sonra varroa bulaşmış petek gözü sayısı
24.2 ± 1.74
10
37
32.11
Bir hafta sonra varroa bulaşıklığı olmayan petek gözü sayısı
75.6 ± 1.66 63 88 9.83
Bir hafta sonra varroa bulaşık petek gözlerinin varroa bulaşık olmayan petek gözlerine oranı
0.33 ± 0.030 0.11 0.58 40.74
Bulaşıklık oranındaki azalma,%
13 ± 15 4 25 50.08
VBYO,% 5 ± 1.8 0.00 37 146.09 FAVO,% 2 ± 0.6 0.00 11 103.25 ADV 3.36 ± 0.837 0.29 14.20 111.21
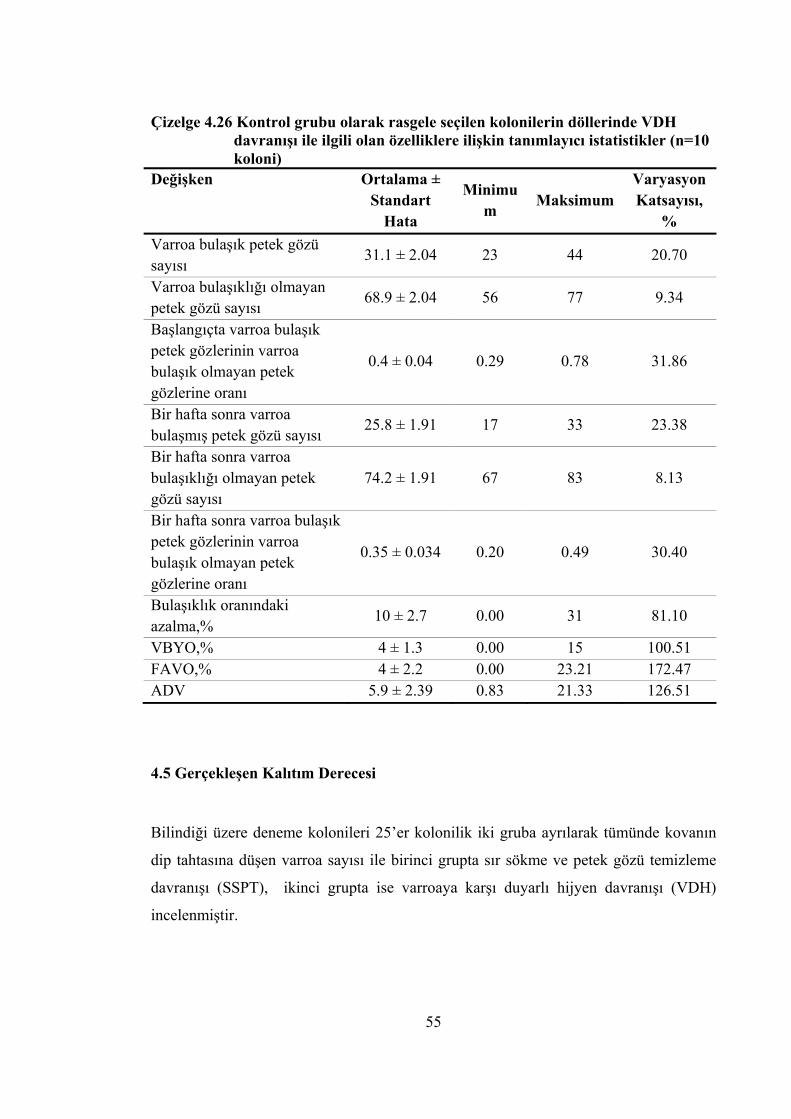
55
Çizelge 4.26 Kontrol grubu olarak rasgele seçilen kolonilerin döllerinde VDH davranışı ile ilgili olan özelliklere ilişkin tanımlayıcı istatistikler (n=10 koloni)
Değişken Ortalama ± Standart
Hata
Minimum
Maksimum Varyasyon Katsayısı,
% Varroa bulaşık petek gözü sayısı
31.1 ± 2.04 23 44 20.70
Varroa bulaşıklığı olmayan petek gözü sayısı
68.9 ± 2.04 56 77 9.34
Başlangıçta varroa bulaşık petek gözlerinin varroa bulaşık olmayan petek gözlerine oranı
0.4 ± 0.04 0.29 0.78 31.86
Bir hafta sonra varroa bulaşmış petek gözü sayısı
25.8 ± 1.91 17 33 23.38
Bir hafta sonra varroa bulaşıklığı olmayan petek gözü sayısı
74.2 ± 1.91 67 83 8.13
Bir hafta sonra varroa bulaşık petek gözlerinin varroa bulaşık olmayan petek gözlerine oranı
0.35 ± 0.034 0.20 0.49 30.40
Bulaşıklık oranındaki azalma,%
10 ± 2.7 0.00 31 81.10
VBYO,% 4 ± 1.3 0.00 15 100.51 FAVO,% 4 ± 2.2 0.00 23.21 172.47 ADV 5.9 ± 2.39 0.83 21.33 126.51
4.5 Gerçekleşen Kalıtım Derecesi
Bilindiği üzere deneme kolonileri 25’er kolonilik iki gruba ayrılarak tümünde kovanın
dip tahtasına düşen varroa sayısı ile birinci grupta sır sökme ve petek gözü temizleme
davranışı (SSPT), ikinci grupta ise varroaya karşı duyarlı hijyen davranışı (VDH)
incelenmiştir.

56
Araştırmada hijyenik davranış özellikleri yalnız işçi arılarda görüldüklerinden kalıtım
derecelerinin tahmininde aşağıdaki eşitlik kullanılmıştır:
2 2( ) / ( )S K S Kh Y Y P P= − −
SY = seçilen ebeveyn grubunun döllerine ait ortalama, KY = kontrol grubunun döllerine
ait ortalama, SP = seçilen ebeveyn grubuna ait ortalama ve KP = kontrol grubunun
ortalamasıdır.
Ayrıca araştırılan özelliklerin kalıtım dereceleri Moran (1984)’ın önerdiği
h2=(∆G/i)×(3/2) gerçekleşen kalıtım derecesi eşitliğine dayanarak da hesaplanmıştır.
4.5.1 Tımarlama davranışının (TD) kalıtım derecesi
Bal arısının varroaya karşı çeşitli davranışları, örneğin tımarlama davranışı, varroaya
karşı duyarlı hijyenik davranışı ve sır sökme ile petek gözlerini temizleme davranışı,
kovanın dip tahtasına düşen varroa sayımlarıyla değerlendirilebilir. Ancak kovan
içerisinde tabana düşen parazit sayısı iki faktörden, yavruların ve ergin arıların varroa
bulaşıklığından etkilenmektedir. Foretik aşamada olan varroa sayısı, yavrulara bulaşan
varroa sayısı ve kovanın dip tahtasına düşen varroa sayısı değişkenlerinin arasında
pozitif ve önemli korelasyon (Çizelge 4.3) bulunduğundan, koloniye ait hijyenik
davranışta bu değişkenlerin önemli etken olduğu ortaya çıkmıştır. Hesaplanan İz-
katsayısı ve belirleme katsayısı yavrulu petek gözlerinin varroa bulaşıklığının, kovanın
dip tahtasına düşen parazit sayısı üzerinde çok etkili olduğunu göstermiştir. Bu nedenle
tımarlama davranışı incelenen başlangıç kolonileri arasından döl generasyonunu
oluşturmak üzere damızlık kolonilerin seçiminde yavrularda varroa bulaşıklığı
değişkeni kullanılmıştır.
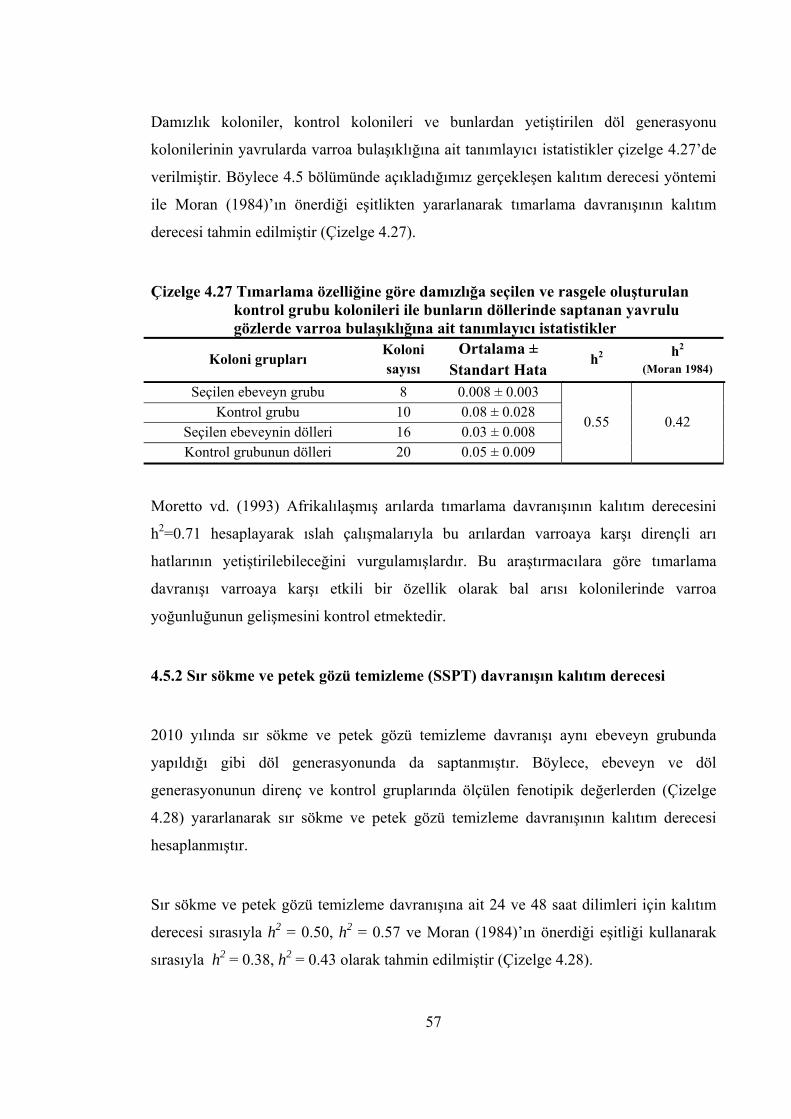
57
Damızlık koloniler, kontrol kolonileri ve bunlardan yetiştirilen döl generasyonu
kolonilerinin yavrularda varroa bulaşıklığına ait tanımlayıcı istatistikler çizelge 4.27’de
verilmiştir. Böylece 4.5 bölümünde açıkladığımız gerçekleşen kalıtım derecesi yöntemi
ile Moran (1984)’ın önerdiği eşitlikten yararlanarak tımarlama davranışının kalıtım
derecesi tahmin edilmiştir (Çizelge 4.27).
Çizelge 4.27 Tımarlama özelliğine göre damızlığa seçilen ve rasgele oluşturulan kontrol grubu kolonileri ile bunların döllerinde saptanan yavrulu gözlerde varroa bulaşıklığına ait tanımlayıcı istatistikler
Koloni grupları Koloni sayısı
Ortalama ± Standart Hata
h2 h2 (Moran 1984)
Seçilen ebeveyn grubu 8 0.008 ± 0.003
0.55 0.42 Kontrol grubu 10 0.08 ± 0.028
Seçilen ebeveynin dölleri 16 0.03 ± 0.008 Kontrol grubunun dölleri 20 0.05 ± 0.009
Moretto vd. (1993) Afrikalılaşmış arılarda tımarlama davranışının kalıtım derecesini
h2=0.71 hesaplayarak ıslah çalışmalarıyla bu arılardan varroaya karşı dirençli arı
hatlarının yetiştirilebileceğini vurgulamışlardır. Bu araştırmacılara göre tımarlama
davranışı varroaya karşı etkili bir özellik olarak bal arısı kolonilerinde varroa
yoğunluğunun gelişmesini kontrol etmektedir.
4.5.2 Sır sökme ve petek gözü temizleme (SSPT) davranışın kalıtım derecesi
2010 yılında sır sökme ve petek gözü temizleme davranışı aynı ebeveyn grubunda
yapıldığı gibi döl generasyonunda da saptanmıştır. Böylece, ebeveyn ve döl
generasyonunun direnç ve kontrol gruplarında ölçülen fenotipik değerlerden (Çizelge
4.28) yararlanarak sır sökme ve petek gözü temizleme davranışının kalıtım derecesi
hesaplanmıştır.
Sır sökme ve petek gözü temizleme davranışına ait 24 ve 48 saat dilimleri için kalıtım
derecesi sırasıyla h2 = 0.50, h2 = 0.57 ve Moran (1984)’ın önerdiği eşitliği kullanarak
sırasıyla h2 = 0.38, h2 = 0.43 olarak tahmin edilmiştir (Çizelge 4.28).

58
Çizelge 4.28 Sır sökme ve petek gözü temizleme özelliğine göre damızlığa seçilen ve rasgele oluşturulan kontrol grubu kolonileri ile bunların döllerinde saptanan 24 ve 48 saat dilimlerde SSPT davranışına ait tanımlayıcı istatistikler
SSPT 24 saat Koloni grupları Koloni
sayısı Ortalama ±
Standart Hata h2 h2
(Moran 1984)
Seçilen ebeveyn grubu 4 0.734 ± 0.052
0.50 0.38 Kontrol grubu 5 0.651 ± 0.083 Seçilen ebeveynin dölleri 20 0.662 ± 0.049 Kontrol grubunun dölleri 10 0.641 ± 0.052
SSPT 48 saat Seçilen ebeveyn grubu 4 1.00 ± 0.000
0.57 0.43 Kontrol grubu 5 0.92 ± 0.015 Seçilen ebeveynin dölleri 20 0.939 ± 0.020 Kontrol grubunun dölleri 10 0.916 ± 0.021
4.5.3 Varroaya karşı duyarlı hijyen (VDH) davranışına ait kalıtım derecesi
Harbo ve Harris’e (2005) göre VDH özelliği, bal arılarında kalıtıma bağlı bir özellik
olarak varroa parazitinin yoğunluğunu kontrol etmektedir. Bu araştırmacılar öz
kardeşler arası benzerlik metoduyla VDH özelliğinin kalıtım derecesini h2= 0.46
hesaplamışlardır (Harbo ve Harris 1999).
Araştırmada VDH özelliğinin fenotipik değerlerinin elde edilmesi için ebeveyn
grubunda yapılan uygulamalar döl generasyonunda da tekrarlanmıştır (Çizelge 4.25-
4.26). Böylece gerçekleşen kalıtım derecesi yönteminden faydalanarak sözü edilen
özelliğin kalıtım derecesi h2 = 0.23, Moran (1984)’ın önerdiği eşitlik kullanılarak da h2
= 0.17 tahmin edilmiştir (Çizelge 4.29).
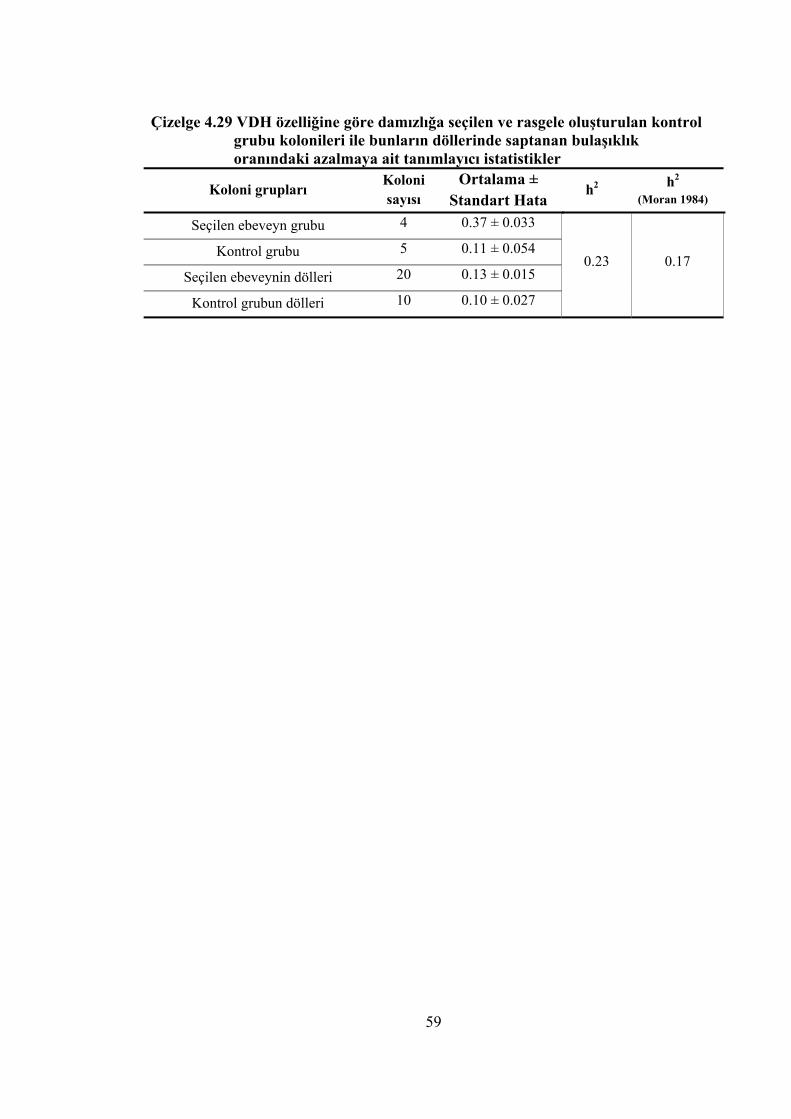
59
Çizelge 4.29 VDH özelliğine göre damızlığa seçilen ve rasgele oluşturulan kontrol grubu kolonileri ile bunların döllerinde saptanan bulaşıklık oranındaki azalmaya ait tanımlayıcı istatistikler
Koloni grupları Koloni sayısı
Ortalama ± Standart Hata
h2 h2 (Moran 1984)
Seçilen ebeveyn grubu 4 0.37 ± 0.033
0.23 0.17 Kontrol grubu 5 0.11 ± 0.054
Seçilen ebeveynin dölleri 20 0.13 ± 0.015
Kontrol grubun dölleri 10 0.10 ± 0.027

60
5. TARTIŞMA VE SONUÇ
5.1 Tımarlama Davranışı (TD)
Arıların alt çeneleriyle parazitleri yaraladıkları birçok araştırmayla belirlenmiştir
(Ruttner ve Hanel 1992, Rosenkranz vd. 1997). Bu nedenle tımarlama davranışı kovan
dip tahtasına düşen akarların yaralı olanlarının oranıyla ifade edilerek varroaya karşı
dirençli kolonilerin seleksiyonunda yararlı bir parametre olarak kullanılmaktadır (Fries
vd. 1996). Tımarlama davranışı bakımından bal arısı kolonileri arasındaki farkların
belirlenmesinde yaralı parazit sayısının kullanılması Boecking ve Spivak 1999, Fries
vd. 1996, Rinderer vd. 2001, Arechavaleta-Velasco ve Guzman-Novoa 2001, tarafından
da benimsenmiştir. Birçok araştırmacı da (Peng vd. 1987, Ruttner ve Hanel 1992, Fries
vd. 1996, Rosenkranz vd. 1997) Apis cerana ve Apis mellifera kolonilerinde tımarlama
davranışını incelemek üzere, işçi arılar tarafından yaralanarak öldürülmüş varroa
sayısını kullanmışlardır.
Mondragon’a (2005) göre kolonideki varroa yoğunluğunun değişimini incelemek için
en uygun yöntem, döllenmiş dişi varroa ile yaralanmış varroa sayısını belirlemektir.
Ancak bu yolla tımarlama davranışının tespitinde birçok zorluk vardır. Genelde dip
tahtasına düşenler arasından binoküler altında yaralanmış parazitlerin sayımı zor bir iş
olarak çok zaman harcamayı gerektirmektedir. Kaldı ki, kovan tabanında ölmüş
olanların dışında canlı, sağlam, yaralı ve zayıf varroalar da tımarlama davranışının
sonucu olabilir (Fries vd. 1996, Thakur vd. 1997).
Kolonilerde foretik aşamada olan varroalar arıların tımarlama davranışı ile uzaklaştırılır
(Fries vd. 1996), ancak bazı akarların sırtında görülen çukurların parazitin gelişme
döneminden kalma olduğu ve bal arısı tarafından yapılan hasarlardan ayrı olduğu
bilinmelidir (Davis 2009). Mooretto’ya (1993) göre, koloniye ait tımarlama davranışının
belirlenmesinde arıların üzerinden kovanın dibine düşen canlı ve yaralı parazit sayısı
kullanılmalıdır.

61
Bu çalışmada kovan dip tahtasına serilen yapışkan kağıt üzerinde toplanan parazitlerin
sayımının tımarlama davranışının sonucu olduğu yaklaşımı uygun görülmüştür.
Ayrıca; kovan dip tahtasına düşen varroa sayısı değişkeninin kolonideki arı yoğunluğu,
sırlanmış yavrulu petek gözü sayısı, varroa bulaşık yavru sayısı ve foretik aşamada olan
varroa sayısı değişkenlerinden etkilenip etkilenmediğini belirlemek için bu değişkenler
de ölçülmüştür.
Sır sökme ve petek gözü temizleme özelliği gösteren kolonilerde sırlanmış petek
gözlerindeki yavruları temizlemenin düzeyi temizlik yapan arı sayısına bağlıdır (Arathi
vd. 2000). Palacio vd. (2010) tarafından yapılan bir araştırmada sır sökme ve petek gözü
temizleme (SSPT) özelliği olan kolonilerde kireç hastalığına bulaştırılmış yavrulu
gözleri inceleyen, göz sırını söken ve gözleri temizleyen arıların oranları sırasıyla 16.57,
4.29 ve 3.05 iken SSPT özelliği olmayan kolonilerde sözü edilen davranışları sergileyen
arı oranı sırasıyla 5.43, 1.10 ve 1.19 olarak saptanmıştır.
Bu araştırmada da kolonilerde arı yoğunluğu ile 1000 arı başına kovanın dip tahtasına
düşen varroa sayısı arasında korelasyon önemli bulunmamıştır (Çizelge 4.3). Başka bir
deyişle tımarlama davranışının etkisiyle kovanın dip tahtasına düşen akarların oranı
kolonideki arı yoğunluğuna değil tımarlama yapan arıların miktarına bağlı olduğu
sonucuna varılmıştır.
Yukarıdaki araştırmaların sonuçlarına dayanarak bu çalışmada tımarlama davranışının
daha doğru tespit edilmesi için sadece yaralanmış parazitlerin sayımları değil, kovanın
tabanına düşen tüm varroa sayımları ele alınmıştır. Elde edilen sonuçlara göre kovanın
dip tahtasına düşen varroa sayısı ile foretik aşmadaki varroa bulaşıklığı ve yavrulardaki
varroa bulaşıklığı arasında önemli korelasyon bulunmuştur. Kovanın dip tahtasına
düşen varroa sayısını daha çok etkileyen faktörü belirlemek için bu değişkenler arasında
İz-katsayısı ve belirleme katsayısı hesaplanmıştır. Buna göre kolonideki varroa
yoğunluğunu ve kovanın dip tahtasına düşen parazit sayısını etkileyen en önemli
değişkenin yavrulu gözlerdeki varroa bulaşıklığı olduğu saptanmıştır. Bu sonuçlar

62
varroa zararlısının 12 gün sırlanmış yavrulu petek gözünde ve yaklaşık 5 gün foretik
aşama olan yaşam döngüsü (Fries vd. 1994) nedeniyle beklenen durumdur. Varroa
bulaşıklığı olan bal arısı kolonisinde parazitin üremesini ve yavrularının yetiştirilmesini
sağlayan en iyi ortam yavrulu petek gözleridir. Bu nedenle varroa yoğunluğu, sırlanmış
petek gözünde ergin arıların üzerindekinden her zaman daha fazladır.
5.2 Sır Sökme ve Petek Gözü Temizleme (SSPT) Davranışı
Petek gözlerindeki ölü ve hasta yavruları temizleme davranışı yönünde seçilmiş
koloniler, seçilmeyenlere kıyasla daha düşük varroa bulaşıklığı göstermişlerdir (Spivak
ve Reuter 1998a, Spivak ve Reuter 1998b, İbrahim vd. 2007).
Hastalıklara karşı farklı seviyelerde dayanıklılık gösteren arı hatları karşılaştırıldığında,
hastalığa karşı dayanıklılığın 4 farklı özellik olduğu ileri sürülmektedir. Bu
özelliklerden biri olan sır sökme ve petek gözü temizleme davranışı 2 aşamalı olarak
gerçekleşmektedir. Birinci aşamada ölü larvaların bulunduğu gözlerin sırlarının bir
kısım işçi arılar tarafından delindiği, başka işçi arılar tarafından delinen gözlerdeki
yavruların dışarı atıldığı belirtilmektedir (Milne 1983).
Araştırmada 48 saat süre içerisinde temizlenmiş yavrulu petek gözü oranı ile yavrularda
varroa bulaşıklığı oranı (r = -0.495 ) ve foretik aşamadaki varroa oranı (r = -0.571)
arasında önemli korelasyonlar bulunmuştur. Demek ki dondurularak öldürülmüş
yavruları temizleyen koloniler varroa bulaşıklığına da duyarlı olarak foretik aşamadaki
ve sırlanmış petek gözlerindeki parazitleri temizlemektedirler.
Araştırmada SSPT grubunun kolonilerinde sır sökme ve petek gözü temizleme
davranışının yanısıra yavrularda varroa bulaşıklığı, foretik aşamada olan varroa oranı ve
kovan dip tahtasına düşen varroa sayısı değişkenleri de ölçülmüştür. Yavrulardaki
varroa bulaşıklığıyla SSPT davranışının arasındaki ilişkiyi incelemek için başlangıç
generasyonun SSPT özelliği gösteren kolonilerden elde edilen verilere dayanarak, bu
koloniler yavrulardaki varroa bulaşıklığı düzeyi bakımından düşük, orta ve yüksek

63
olarak 3 alt gruba ayrılmıştır. Bu alt grupları SSPT davranışı bakımından
karşılaştırdığımızda, daha çok temizlik yapan kolonilerde yavrulardaki varroa
bulaşıklığının daha düşük olduğu görülmüştür. Bu sonuçlara göre ölü yavrulara karşı
SSPT davranışı gösteren koloniler varroa parazitine karşı da dirençli olmaktadırlar.
Birçok çalışmada, SSPT davranışı yönünde yapılan seleksiyonun, kolonilerin bal
üretimini ve hırçınlık davranışını etkilemediği gösterilmiştir (Spivak 1996, Spivak ve
Reuter 1998a). Araştırmacılar, ticari olarak çoğaltılabilecek bu özellikteki arı hatlarının
arıcılık endüstrisi için yararlı olacağı görüşünü taşımaktadırlar (Spivak 1998).
Jevtic vd. (2009) Sırbistan’da Karniyol ( Apis mellifera carnica Poll.) arısının 3’ü
dağlık ve 3’ü daha düşük rakımlı vadi olmak üzere 6 farklı yöresel populasyonun kendi
doğal koşullarında iğne ile öldürülmüş yavrulu gözlerde sır sökme ve petek gözü
temizleme davranışlarını incelemişlerdir. Çalışmada 2 generasyonda ve 2 yıl süre ile 24
ve 48 saatlik sürelerde sır sökme ve petek gözü temizleme davranışlarının test
sonuçlarına bakılmıştır. 48 saat süre içerisinde sır sökme ve petek gözü temizleme
oranları bakımından populasyonlar arasındaki farklılık istatistik olarak çok yüksek
çıkmıştır.
Bu araştırmada da farklı yörelerden alınan koloni gruplarında hem sır sökme ve petek
gözü temizleme hem de VDH ile tımarlama özellikleri bakımından önemli farklılıklar
bulunmuştur.
İbrahim vd. (2007) tarafından yavrularda varroa bulaşıklık oranı ve ergin arılarda
bulaşıklık oranı (bağımlı değişkenler), sır sökme ve petek gözü temizleme davranışının
(bağımsız değişken) fonksiyonu olarak incelenmiştir. Sır sökme ve petek gözü
temizleme davranışının sözü edilen bağımlı değişkenler tarafından belirleme katsayıları
sırasıyla 0.376 ve 0.193 olarak hesaplanmıştır.
Yapılan bir çalışmada, gözlem kovanlarına alınan 2 hijyenik 2 de hijyenik olmayan
kolonide iğneleme yöntemi ile öldürülmüş yavrulara karşı hijyenik davranış testi

64
yapılmıştır. Kolonilerin ölü yavruları 1, 2, 3, 4, 5, 6 ve 24 saat sonra temizleme kayıtları
alınmış ve SSPT özelliği olan koloniler ile olmayanlar arasında temizleme süresi
bakımından önemli farklılıklar gözlenmiştir. Bu gözlemlere dayanarak SSPT özelliğine
sahip hatların kısa süre içerisinde saptanabileceği vurgulanmıştır (Palacio vd. 2005).
Bu çalışmada da sır sökme ve petek gözü temizleme süresi bakımından yöreler arasında
istatistik olarak önemli farklar bulunmuştur. Ayrıca varyans analizi sonuçlarına göre
süre faktörü ile yöre faktörü arasındaki karşılıklı etkileşim önemli (P < 0.01)
bulunmuştur. Bu nedenle İran bal arısı populasyonlarında da SSPT özelliğinin farklı
frekanslarda bulunduğu kanıtlanarak geniş kapsamlı araştırmalar yapıldığı takdire
varroa parazitine karşı dayanaklı arı hatlarının yetiştirilebileceği sonucuna varılmıştır.
Spivak’a (1996) göre ıslah programlarında sır sökme ile petek gözü temizleme ve
tımarlama davranışlarına dayanarak varroaya karşı dirençli arıların yetiştirilmesi
mümkün olmaktadır.
İran balarısı ile yürütülen bu araştırmada, arıların sır sökme ve petek gözü temizleme
davranışına bakılarak elde edilen kalıtım dereceleri, incelenen özellik yönünde
yapılacak bir seleksiyon ile iyileştirilme sağlanabileceğini göstermektedir.
5.3 Varroaya karşı Duyarlı Hijyen Davranışı (VDH)
Hijyenik davranış gösteren arılar varroa populasyonunun gelişmesini 3 şekilde
engellemektedir: 1) pupalar gözlerden sökülürken olgunlaşmamış varroalar öldürülür,
böylece üreyebilen zararlı başına döl sayısı azalır, 2) petek gözünden pupalar
çıkarılırken kaçmayı başaran olgun dişi zararlıların foretik aşaması uzar, 3) arıların
tımarlama etkisiyle, sırı sökülmüş petek gözlerinden kaçmaya çalışan ergin zararlıların
yaralanması ile parazitlerin ölüm oranı artar (Spivak 1996).

65
Sır sökme ve petek gözü temizleme davranışını incelemek üzere uygulanan sıvı azot ile
dondurularak öldürülen yavruların çıkarılmasına dayanan yöntem (Spivak ve Reuter,
1998), VDH davranışının da belirlenmesini sağlar (Boecking ve Drescher 1992, Spivak
1996).
Bazı kolonilerde foretik aşamanın uzaması parazitlerin üreme şansını azaltarak arıların
tımarlama davranışına maruz kalmalarını sağlamaktadır (Ruttner ve Hanel 1992). Bu
yüzden foretik aşamanın uzunluğuna dayanarak kolonilerin seleksiyonu, varroaya karşı
dirençli olma özelliğini geliştirmektedir. Nasıl olursa arıların VDH ve tımarlama
davranışları yavrulara ve ergin arılara parazitlik yapan varroaların sayısını
azaltmaktadır. VDH nedeniyle petek gözünden çıkartılan varroalar ya ölür ya da foretik
aşamasında olan parazit populasyonuna katılarak yoğunluğu artırır. Rus bal arılarında
foretik aşamanın süresi, ya yavruların çekici olmamasından (Guzman vd. 2007), ya kış
sezonunda kolonide yavru bulunmamasından (Tubbs vd. 2003) ya da oluşum dönemi
uzun olan erkek arı gözlerine girmesinden (Rinderer vd. 2001, Guzman vd. 2007)
etkilenmektedir.
Çalışmada, yavrularda varroa bulaşıklığı bakımından düşük grupta VDH özelliğinin
daha yüksek seviyede olduğu saptanmıştır. Ayrıca düşük, orta ve yüksek olmak üzere
üç alt gruba bölünen VDH kolonileri arasında varroaya karşı duyarlı hijyen davranışı
bakımından önemli fark (P < 0.05) bulunmuştur.
Vandame vd. (2002) Meksika’da yaptıkları bir çalışmada, Varroa destructor’un Avrupa
(EHB) bal arılarında Afrika (AHB) bal arılarından daha fazla yaygın bulduğunu ve bu
parazite karşı iki direnç mekanizmasını inceleyerek Afrika (AHB) bal arılarının Avrupa
(EHB) bal arılarına göre daha fazla tımarlama davranışı gösterdiklerini saptamışlardır.
Ayrıca yavrularda varroa bulaşıklığının temizlenmesi bakımından da özellikle yavrulu
petek gözlerinin kapandığı 5 ile 7. günler arasında Avrupa (EHB) bal arıları %8.03
oranında temizlerken Afrika (AHB) bal arıları %32.46 oranında temizleme davranışı
göstermişlerdir.

66
Mısırda yapılan benzer bir çalışmada, Karniyol (A. m. carnica) arıları ile Lamarkii (A.
m. lamarckii) arıları üzerinde incelenen VDH davranışı testlerinde Lamarkii arıları
Karniyol arılarından önemli düzeyde daha yüksek hijyenik davranış göstermişlerdir
(Kamel vd. 2003).
İran bal arıları ile yürütülmüş olan bu araştırmada, VDH özelliği, petek gözlerindeki
bulaşıklığın azalmasına dayanarak hesaplanmıştır. Elde edilen sonuçlara göre
yavrulardaki varroa bulaşıklığı düşük olan kolonilerde daha yüksek düzeyde VDH
davranışı olduğu görülmüştür (Çizelge 4.19). Başka bir deyişle arıların parazite karşı
duyarlı olduğu kolonilerde yavrularda varroa bulaşıklığı daha az bulunmaktadır. Birçok
araştırma (Spivak 1996, Harbo ve Harris 2005, İbrahim 2005, İbrahim vd. 2007, Harris
2007, Harris 2008, Harbo ve Harris 2009, Harris vd. 2010) bu sonuçları
desteklemektedir. Ayrıca foretik aşamada olan varroa bulaşıklığı bakımından yöreler
arasındaki farklara bakarak birçok dayanıklılık özellikleri ortaya çıkmaktadır. Arılar
tarafından sırlanmış petek gözlerindeki parazitlerin belirlenip çıkartılması nedeniyle
yüksek derecede VDH davranışı gösteren kolonilerde yavrularda varroa bulaşıklığı
azalırken foretik bulaşıklık (r = -0.348, P < 0.05) artmaktadır.
5.4 Kalıtım Derecesi
Moretto vd. (1993) araştırmalarında, varroaya karşı tımarlama davranışının kalıtım
derecesini 0.71 hesaplamışlardır. Milne (1985) ise sır sökme ve petek gözü temizleme
davranışlarının kalıtım derecelerini sırasıyla 0.144 ve 0.22 bulmuştur.
Harbo (1995) bal arısı kolonilerinde kireç hastalığına yakalanan yavrulara karşı arıların
hijyenik davranışlarının kalıtım derecesini 0.65 olarak hesaplamıştır.
Boecking vd. (2000) yaptıkları bir araştırmada, petek gözlerindeki varroalara karşı
arıların hijyenik davranışının kalıtım derecesini h2=0.18 ± 0.27 ve ölü yavrular bulunan
petek gözlerinin temizlenmesinin kalıtım derecesini h2=0.36 ± 0.30 hesaplamışlardır.
Harbo ve Harris (1999) tarafından yapılan araştırmada öz kardeşler arası benzerlik

67
metodu kullanılarak varroaya karşı duyarlı hijyen davranışının kalıtım derecesi h2= 0.46
hesaplanmıştır.
Yukarıda bildirilen araştırmaların tımarlama, 24 ve 48 saat içerisinde sır sökme ve petek
gözü temizleme ve varroaya karşı duyarlı hijyen davranışı için, hesaplanan h2 değerleri
ile bu araştırmada sırasıyla 0.28, 0.50, 0.57 ve 0.25 olarak hesaplanmış olan kalıtım
dereceleri ile benzerdir (Milne 1985, Moretto vd. 1993, Harbo 1995, Harbo ve Harris
1999, Boecking vd. 2000).
Bu araştırmadan elde edilen sonuçlara göre, foretik aşamadaki parazit oranını ve
kovanın dip tahtasına düşen varroa sayısını etkileyen yavrularda varroa bulaşıklığı,
tımarlama davranışının belirlenmesinde incelenmesi gereken önemli bir faktördür.
Ayrıca bu sonuçlar, yavrulardaki varroa bulaşıklığının doğrudan VDH özelliğinden
etkilenmesini açıklamaktadır. Bu yüzden varroaya karşı direnç özelliklerinin
değerlendirilmesinde ve bu özelliği gösteren kolonilerin seçilmesinde yavru
bulaşıklığının tespit edilip kullanılması, hem araştırmacılara hem de arıcılara
önerilebilecek bir sonuçtur. Sonuçlar varroa ile mücadelede başarılı hatların
yetiştirilmesinin mümkün olduğunu göstermektedir.
Tek üreme ve çoğalma yeri bal arısı kolonisi olan varroa gerekli önlem alınmadığında
hızla çoğalarak ya kolonilerin verimlerinin düşmesine ve ileri aşamalarda sönmesine ya
da koloni bireylerini zayıf düşürerek diğer hastalıklara karşı daha duyarlı hale gelmesine
neden olmaktadır. Bu durumlar bilindiğinden parazite karşı çok fazla sayıda mücadele
yöntemleri geliştirilmiş olup bunların başında da kimyasal mücadele yöntemleri
gelmektedir. Kimyasallar varroa mücadelesinde etkili olmakta ancak bu maddelerin
bilinçsiz kullanımı arıların sağlıklarını tehdit ettiği gibi arı ürünlerinde kalıntı bırakarak
insan sağlığını da tehdit etmektedir. Bunun yanında hedef organizmalarda kimyasallara
karşı direnç oluşum riski yüksektir. Günümüzde, ergin arıların üzerinde veya kapalı
yavru gözlerinin içinde yaşayan varroanın tamamen yok edilmesi mevcut metotlarla
mümkün değildir. Kaldı ki birçok ülkede hastalık ve zararlılara karsı kimyasal
kullanılması yasaklanmıştır. Verimli bir arıcılık ve sağlıklı arı ürünleri üretebilmek için

68
kovandaki varroa yoğunluğunu azaltmak gerekmektedir. Bu yüzden ileriki yıllarda arı
kolonilerini ıslah yöntemleri ile hastalıklara karsı dirençli hale getirmek büyük önem
kazanmıştır.
Bu araştırma, İran’ın Doğu Azerbaycanı bal arısı populasyonlarında varroaya karşı
direnç özelliklerinin bulunduğunu kanıtlamış, İran’ın diğer bölgelerinde de çalışılan
özellikleri taşıyan populasyonların bulunabileceği görüşünü güçlendirmiştir.
İran’da Varroa destructor’e karşı dirençli arıların yetiştirilmesi, arıcılar için bu parazitin
neden olduğu birçok ekonomik sorunu giderebilecektir. İlk adım olarak İran bal arısı
(Apis mellifera meda) populasyonları içerisinde varroaya karşı dirençli olan kolonilerin
belirlenmesi ve çoğaltılması öncelikli hedef olmalıdır.

69
KAYNAKLAR
Akçalı, Y. 1979. Bazı tarla bitkilerinde çeşitli morfolojik özelliklerin verim üzerindeki
etkilerinin saptanmasında path analizi yönteminin kullanılması üzerinde
araştırmalar. Doktora tezi, Fenbilimleri Enstitüsü, Ankara Üniversitesi.
Anderson, D. 2006. Clarification of aspects of Varroa reproduction-first stage of a
possible new control method. RIRDC Publication No 06/007. RIRDC Project No
CSE-87A.
Anderson, D. L. 1995. Viruses of Apis cerana and Apis mellifera. In: P. G. Kevan (ed).
The Asiatic Bee Hive: Apiculture, Biology, and Role in Sustainable Development
in Tropical and Subtropical Asia. Enviroquest, Ltd., Cambridge, ON, Canada. pp.
161-170.
Anderson, D. L. and Trueman, J. W. H. 2000. Varroa jacobsoni (Acari: Varroidae) is
more than one species. Experimental and Applied Acarology, vol. 24, pp.165–
189.
Anonymous. 2009. Varroa destructor. Web sitesi: http://en.wikipedia.org/wiki/Varroa_
destructor.
Anonymous. 2008. Bee enemies. Web Sitesi: http://www.beebehavior.com/bee_ enemies.php.
Erişim Tarihi (18/01/2009).
Arathi, H. S., Burns, I. and Spivak, M. 2000. Ethology of hygienic behavior in the
honeybee Apis mellifera L. (Hymenoptera: Apidae) behavioural repertoire of
hygienic bees. Ethology, vol. 106, pp: 365–379.
Arathi, H.S., Ho, G. and Spivak, M. 2006. Inefficient task partitioning among
nonhygienic honeybees, Apis mellifera L., and implications for disease
transmission. Anim. Behav., vol. 72, pp. 431–438.
Arechavaleta-Velasco, M. E. and Guzman-Novoa, E. 2001. Relative effect of four
characteristics that restrain the population growth of the mite Varroa destructor in
honey bee (Apis mellifera) colonies. Apidologie, vol. 32 (157), pp. 157-174.

70
Aumeier, P. and Rosenkranz, P. 2001. Scent or movement of Varroa destructor mites
does not elicit hygienic behaviour by Africanized and Carniolan honey bees.
Apidologie, vol. 32, pp. 253–263.
Bailey, L. 1981. Honey Bee Pathology. Academic Press, New York & London. 124 pp.
Bailey, L., Carpenter, J. M. and Woods. R. D. 1979. Egypt bee virus and Australian
isolates of Kashmir bee virus. Journal of General Virology; vol. 43, pp. 641-647.
Ball, B. 1994. Host-parasite-pathogen interactions. In Matheson, A (ed) New
perspectives on Varroa . IBRA, Cardiff, Wales, pp 5-11.
Bienefeld, K., Zaautke, F., Pbronin, D. and Mazeed, A. 1999. Recording the proportion
of damaged Varroa jacobsoni Oud. in the debris of honey bee colonies (Apis
mellifera). Apidologie, vol. 30, pp. 249-256.
Bienefeld, K., Tahmasebi, G. H., Keller, R., Kauhausen-Keller, D. and Ruttner, F. 1996.
Report on the present situation of Apis mellifera meda in Iran. Apidologie, vol.
27(4), pp. 307-308.
Boecking O., and Drescher W. 1992. The removal response of Apis mellifera L.
colonies to brood in wax and plastic cells after artificial and natural infestation
with Varroa jacobsoni Oud. and to freeze-killed brood, Exp. Appl. Acarol. vol.
16, 321 329.
Boecking, O. 1992a. Removal behavior of Apis mellifera towards sealed brood cells
infested with Varroa jacobsoni: Techniques, extent and efficiency. Apidologie,
vol. 23, 371–373.
Boecking, O., Bienefeld, K. and Drescher, W. 2000. Heritability of Varroa -specific
hygienic behaviour in honey bees (Hymenoptera: Apidae). Journal of Animal
Breeding and Genetics, vol. 117, 417-424.
Boecking, O. and Drescher, W. 1991. Response of Apis mellifera L. colonies to brood
infested with Varroa jacobsoni Oud. Apidologie, vol. 22, pp.237–241.
Boecking, O. and Spivak, M. 1999. Behavioral defenses of honey bees against Varroa
jacobsoni Oud. Apidologie, vol. 30, pp. 141–158.

71
Burgett, M. and Burikam, I. 1985. Number of adult honey bees (Hymenoptera: Apidae)
occupying a comb: a standard for estimating colony populations. J. Econ.
Entomol. vol. 78, pp. 1154-1156.
Calderone, N. W. and Lin, S. 2001. Behavioural responses of Varroa destructor (Acari:
Varroidae) to extracts of larvae, cocoons and brood food of worker and drone
honey bees, Apis mellifera (Hymenoptera: Apidae). Physiological Entomology,
vol. 26, pp. 341-350.
Calderone, N. W. 2006. Drone Brood Removal for the Management of Varroa
destructor. Web sitesi: http://www.masterbeekeeper.org/pdf/dronecomb_exchange.pdf.
Erişim Tarihi: 30.01.2011
Colin, M.E., Fernandez, P.G. and Hamida, T.B. 1999. Varroosis. In Colin ME (Ed),
Ball BV (Ed), Kilani M (Ed): Bee Disease Diagnosis, Zaragoza. CIHEAM-
IAMZ, 121-142.
Davis, A. R. 2009. Regular dorsal dimples on Varroa destructor– Damage symptoms or
developmental origin?. Apidologie, vol. 40, 151-162.
Denmark, H. A., Cromroy, H. L., Cutts, L. and Sanford, M. T. 1991. Varroa Mite,
Varroa jacobsoni Oudemans. Entomology Circular No. 347.
Düzgüneş, O., Eliçin, A. ve Akman, N. 2003. Hayvan ıslahı. Ankara Üniversitesi, Ziraat
Fakültesi, Zootekni Bölümü, pp. 298, Ankara.
Falconer, D. S. 1989. Introduction to Quantitative Genetics. 3nd Edn. Longmans:
London.
Fries, I., Camazine, S. and Sneyd, J. 1994. Population dynamics of Varroa jacobsoni: a
model and a review. Bee World, vol. 75, pp. 5-28.
Fries, I., Huazhen, W., Wei, S. and Jin, C. S. 1996. Grooming behavior and damaged
mites (Varroa jacobsoni) in Apis cerana cerana and Apis mellifera ligostica.
Apidologie, vol. 27, pp. 3-11.
Fujiyuki, T. 2005. Kakugo virus from brains of aggressive worker honeybees. Adv
Virus Res., vol. 65, pp.1-27.

72
Gilliam, M. and Taber, S. III. Richardson, G. V. 1983. Hygienic behavior of honey bees
in relation to chalkbrood disease. Apidologie, vol. 14, pp.29-39.
Gramacho, K. P. and Spivak. M. 2003. Differences in olfactory sensitivity and
behavioral responses among honey bees bred for hygienic behavior. Behav Ecol
Sociobiol, vol. 54, pp.472–479.
Guzman, L. I., Rinderer, T. E. and Frake, A. M. 2007. Growth of Varroa destructor
(Acari: Varroidae) populations in Russian honey bee (Hymenoptera: Apidae)
colonies. Ann. Entomol. Soc. Am, vol. 100, pp. 187195.
Haddad, N. 2011. Honey bee viruses, diseases and hive management in the middle east
and their relation to the colony collapse disorder and bee losses. U. Bee J. vol.
11(1), pp. 17-24.
Harbo, J. R. 1995. Observation hygienic behavior and resistance to chalkbrood. Am.
Bee. J. vol. 135, pp. 828.
Harbo, J. R. and Harris, J. W. 2000. Changes in reproduction of Varroa destructor after
honey bee queens were exchanged between resistant and susceptible colonies.
Apidologie vol. 31, pp. 689-699.
Harbo, J. R. and Harris, J. W. 2001. Resistance to Varroa destructor (Mesostigmata:
Varroidae) When Mite-Resistant Queen Honey Bees (Hymenoptera: Apidae)
Were Free-Mated with Unselected Drones. Journal of Economic Entomology, vol.
94(6), pp. 1319-1323(5).
Harbo, J.R. and Harris, J.W. 2005. Suppressed mite reproduction explained by the
behaviour of adult bees. Journal of Apicultural Research, vol. 44, pp. 21-23.
Harbo, J.R. and Harris, J.W. 2009. Responses to Varroa by honey bees with different
levels of varroa sensitive hygiene. J. Apic. Res., vol. 48, pp. 156–161.
Harris, J. W. 2007. Bees with varroa sensitive hygiene preferentially remove mite
infested pupae aged ≤ five days post capping. Journal of Apicultural Research,
vol. 46(3), pp. 134-139.

73
Harris, J. W. 2008. Effect of brood type on varroa -sensitive hygiene by worker honey
bees (Hymenoptera: Apidae). Ann. Entomol. Soc. Am., vol. 101(6), pp. 1137-
1144.
Harris, J.W., Danka, R. G. and Villa, J. D. 2009. Hygienic activity toward Varroa mites
in capped brood is not dependent on mite reproductive status. Am. Bee J., vol.
149, pp. 587–588.
Harris, J.W., Danka, R. G. and Villa, J. D. 2010. Honey bees (Hymenoptera: Apidae)
with the trait of Varroa Sensitive Hygiene remove brood with all reproductive
stages of Varroa mites (Mesostigmata: Varroidae). Ann. Entomol. Soc. Am.,
vol. 103, pp. 146–152.
Hung, A. C. F., Adams, J. R. and Shimanuki, H. 1995. Bee parasitic mite syndrome (II):
the role of Varroa mite and viruses. American Bee Journal, vol. 135(10),
pp.702-704.
Ibrahim, A. M. 2005. The effect of two honey bee mechanisms of resistance against the
parasitic mite, Varroa destructor: Hygienic behavior and suppression of mite
reproduction. Doktora tezi.
Ibrahim, A. and Spivak, M. 2006. The relationship between hygienic behavior and
suppression of mite reproduction as honey bee (Apis mellifera) mechanisms of
resistance to Varroa destructor. Apidologie, vol. 37, pp. 31–40.
Ibrahim, A., Reuter, G.S. and Spivak, M. 2007. Field trial of honey bee colonies bred
for mechanisms of resistance against Varroa destructor, Apidologie, vol. 38, pp.
67–76.
Ifantidis. M. D. 1997. Ontogenesis of Varroa jacobsoni Oud. CIHEAM-Options
Mediterraneennes.
Jevtic, G., Mladenovic, M., Nedic, N. and Andelkovic, B. 2009. The temperament traits
ant the hygienic behavior of honey bee (Apis mellifera carnica Poll.) from serbia.
Apimondia. 2009. France.

74
Jones, R. L. and Rothenbuhler, W. C. 1964. Behavior genetics of nest cleaning in honey
bees II. Responses of two inbred lines to various amounts of cyanide-killed brood.
Animal Behaviour. vol. 12, pp. 584-588.
Kamel, S. M., Strange, J. P. and Sheppard, W. S. 2003. A scientific note on hygienic
behavior in Apis mellifera lamarckii and A. m. carnica in Egypt. Apidologie. vol.
34 189–190.
Kevan, P. G., Hannan, M. A., Ostiguy, N. and Guzman-Novoa. E. 2006. A summary of
the Varroa -Virus disease complex in honey bees. American Bee Journal, August
2006, pp. 694-697.
Kulincevic, J. M., Rinderer, T. E., Mladjan, V. J. and Buco, S. M. 1992. Five years of
bi-directional genetic selection for honey bees resistant and susceptible to Varroa
jacobsoni. Apidologie, vol. 23, 443-452.
Kumova, U. 2004. Varroa ile Mücadele Yöntemleri. II.Marmara Arıcılık Kongresi
Bildiri Kitabı. Uludag Arıcılık Derneği Yayın No: 2 83–131 Uludag Üniv.
Basımevi, Bursa.
Laidlaw, H. H. and Page, E. R. 1997. Queen Rearing and Bee Breeding. Wicwas Press.
Cheshire, Connecticticut,USA. 1-224.
Ledezma, J. 2000. Differential grooming responses of European and Africanized
honeybees Apis mellifera to infestation by the parasitic mite Varroa jacobsoni. In
partial fulfillment of the requirements for the degree of master of science. Major
subject: Biology. University of Texas-Pan American.
Mangum, W. A. 1997. Modeling the population biology and the population genetic
dynamics of the honey bee, Apis mellifera L., when parasitized by the mite,
Varroa jacobsoni Oudemanns. Am. Bee J. vol. 137, pp. 226.
Martin, C., Provost, E., Bagnères, A.G., Roux, M., Clément, J.L. and Le Conte, Y.
2002. Potential mechanism for detection by Apis mellifera of the parasitic mite
Varroa destructor inside sealed brood cells. Physiol. Entomol., vol. 27, pp. 175–
188.
Martin, S. 1994. Ontogenesis of the mite Varroa jacobsoni Oud. In worker brood of the
honeybee Apis mellifera L. under natural conditions. Exp. Appl. Acarol. 18, (2),
pp. 85-99.

75
Martin, S. 2001. The role of Varroa and viral pathogens in the collapse of honeybee
colonies: a modeling approach. J. Appl. Ecol., vol. 38, pp. 1082-1093.
Martin, S., Hogarth, A., Van Breda, A. and Perrett, J. 1998. A scientific note on Varroa
jacobsoni Oudemans and the collaps of Apis mellifera colonies in the United
Kingdom. Apidologie. vol. 29, pp. 369-370.
Martin, S. J. and Kemp, D. 1997. Average number of reproductive cycles performed by
Varroa jacobsoni in honey bee (Apis mellifera) colonies. Journal of Apicultural
Research vol. 36(3/4), pp. 113–123.
Milne, C. P. 1985. Estimates of the heritabilities and genetic correlation between two
components of honey bee (Hymenoptera: apidae) hygienic behavior: uncapping
and removing. Ann. Entomol. Soc. Am. vol. 78, pp. 841-844.
Milne, C. 1982. Laboratory measurement of brood disease resistance in the honeybee. 1.
Uncapping and removal of freeze-killed brood by newly emerged workers in
laboratory test cages. J. of Apic. Res. vol. 21(2), pp. 111-114.
Milne, C. 1983. Honeybee (Hymenoptera: Apidae) hygienic behavior and resistance to
chalkbrood. Ann. Entomol. Soc. Of Amer. vol. 76(3), pp. 384-387.
Milum, G. V. 1947 Grooming dance and associated activities of the honey bee colony.
Gleanings in Bee Culture. pp: 676-685.
Momot, J. P. and Rothenbuhler, W. C. 1971. Behaviour genetics of nest cleaning in
honeybees. VI. Interactions of age and genotype of bees, and nectar . J. Apic.
Res., vol. 10, pp.11–21.
Mondragon, L., Spivak, M. and Vandame, R. 2005. A multifactorial study of the
resistance of honeybees Apis mellifera to the mite Varroa destructor over one
year in Mexico1. Apidologie, vol. 36, pp. 345–358
Moore, D., Angel, J. E., Cheeseman, I. M., Robinson, G. E. and Fahrbach, S. E. 1995. A
highly specialized social grooming honey bee (Hymenoptera: Apidae). J. Insect.
Behav. vol. 8, pp. 855-861.
Moosbeckhofer, R. 1992. Beobachtungen zum auftreten beschädigter varroamilben im
natürlichen totenfall bei völkern von Apis mellifera carnica. Apidologie, vol. 23,
pp. 523–531.

76
Moran, C. 1984. Sex linked effective population size in control populations, with
particular reference to honeybees (Apis mellifera L). Theor. Appl. Genet. Vol. 67,
pp. 317-322.
Moretto, G., Gonçalves, L. S. and de Jong, D. 1993. Heritability of Africanized and
European honey bee defense behavior against the mite Varroa jacobsoni. Braz. G.
Genet. vol. 16, pp. 71–77.
Morse, R. A. 1969. Asian bee mites. Glean Bee Cult. vol. 97, 287-315.
Morse, R. A. 1990. The Cornell project to breed mite resistant bees from queen
honeybees imported from Great Britain. Amer. Bee J. vol. 130, pp. 186.
Mossadegh, S. and Komeili, B. A. 1987. Mites of honey bee. Academic press. Ahvaz.
pp. 4-10.
Nasr, M. E. 1998. Technology transfer of improved honey bee stocks and management
practices to beekeepers in Ontario. Annual Report for 1997–1998. ABA, Bayfield,
Ontario, Canada. ,
Newton, D. C. and Ostasiewski, N. J. 1986. A Simplified Bioassay for Behavioral
Resistance to American Foulbrood in Honey-Bees (Apis mellifera L). American
Bee Journal, vol. 126(4), 278-281.
Oldroyd, B. and Moran, C. 1983. Heritability of worker characters in the honeybee
(Apis mellifera). Australian Journal of Biological Science, vol. 36, 323-32.
Palacio, M. A., Flores, J. M., Figini, E., Ruffinengo, S., Escande, A., Bedascarrasbure,
E., Rodriguez, E. and Gonçalves, L. S. 2005. Evaluation of the time of uncapping
and removing dead brood from cells by hygienic. Genet. Mol. Res. vol. 4 (1), pp.
105-114.
Palacio, M. A., Figini, E. E., Ruffinengo, S. R., Rodriguez, E. M., Del Hoyo, M. L. and
Bedascarrasbure, E. L. 2000. Changes in a population of Apis mellifera L.
selected for hygienic behaviour and its relation to brood disease tolerance.
Apidologie. vol. 31, pp. 471–478.
Palacio, M., Rodriguez, E., Goncalves, L., Bedascarrasbure, E. and Spivak, M. 2010.
Hygienic behaviors of honey bees in response to brood experimentally pin-killed
or infected with Ascosphaera apis. Apidologie vol. 41, pp. 602–612.

77
Peng, Y. S., Fang, Y., Xu, S. and Ge, L. 1987. The resistance mechanisms of the Asian
honey bee, Apis cerana Fabr., to an ectoparasitic mite, Varroa jacobsoni
Oudemans. J. Invert. Pathol. vol. 49, pp. 54–60.
Pettis, J. S. and Pankiw, T. 1998. Grooming behavior by Apis mellifera L. in the
presence of Acarapis woodi (Rennie) (Acari: Tarsonemidae).
Rath, W. and Drescher, W. 1990. Response of Apis cerana Fabr. colonies towards
brood infested with Varroa jacobsoni Oud. and infestation rate of colonies in
Thailand, Apidologie vol. 21, pp. 311–321.
Rinderer, T., De Guzman, L., Lancaster, V., Delatte, G. and Stelzer, J. 1999. Varroa in
the mating yard: 1. The effects of Varroa jacobsoni and Apistan R on drone
honey bees. 2. The effects of Varroa and fluvalinate on drone mating
competitiveness. 3. The effects of formic acid gel formulation on drone
reproduction. American Bee Journal, vol. 139 (2), pp. 34-139; (3), pp. 225-227;
(4), pp. 304-307.
Rinderer, T. E. 1977. Measuring the heritability of characters of the honeybee. Journal
of Apicultural Research, vol. 16, pp. 95-8.
Rinderer, T. E. 1986. Bee Genetics and Breeding. Harcourt Brace Jovanovich, 426,
United States.
Rinderer, T. E., de Guzman, L. I., Delatte, G. T., Stelzer, J. A., Lancaster, V. A.,
Kuznetsov, V., Beaman, L., Watts, R. and Harris, J. W. 2001. Resistance to the
parasitic mite Varroa destructor in honey bees from far-eastern Russia,
Apidologie, vol. 32, pp. 381–394.
Rivera-Marchand, B. 2006. Africanized honey bees in Puerto Rico. Ph.D. Thesis,
University of Puerto Rico. pp. 1-100.
Rosenkranz, P., Fries I., Boecking O. and Stürmer, M. 1997. Damaged Varroa mites in
the debris of honey bee (Apis mellifera L.) colonies with and without hatching
brood, Apidologie vol. 28, 427–437.
Rothenbuhler, W. C. 1964a. Behavior genetics of nest cleaning in honey bees I.
Responses of four inbred lines to disease-killed brood. Animal Behaviour. vol. 12,
pp. 578-583.

78
Rothenbuhler, W. C. 1964b. Behavior genetics of nest cleaning in honey bees IV.
Response of F1 and backcross generations to disease-killed brood. American
Zoologist. vol. 4, pp. 111-123.
Ruttner F., Hänel H. 1992. Active defense against Varroa mites in a carniolan strain of
honey bee (Apis mellifera carnica Pollmann), Apidologie vol. 23, 173–187.
Shahrestani, N. 1979. Honey bee rearing. Academic Press. Tehran. 380 pp.
Shimanuki, H., Calderone, N. and Knox, D. 1994. Parasitic mite syndrome: the
symptoms. American Bee Journal. vol. 134(12), pp. 827-828.
Spivak M. and Reuter, G. S. 1998a. Performance of hygienic honey bee colonies in a
commercial apiary. Apidologie, vol. 29, pp. 291-302
Spivak, M. and Reuter, G. S. 1998b. Honey bee hygienic behavior. Am. Bee J. vol. 138,
pp. 283-286
Spivak, M. 1996. Honey bee hygienic behavior and defense against Varroa jacobsoni.
Apidologie, vol. 27, pp.245–260.
Spivak, M. and Downey, D. L. 1998. Field assays for hygienic behavior in honey bees
(Hymenoptera : Apidae). Journal of Economic Entomology, vol. 91(1), pp. 64-70.
Spivak, M. and Reuter, G. S. 2001. Varroa destructor Infestation in Untreated Honey
Bee (Hymenoptera: Apidae) Colonies Selected for Hygienic Behavior. J. Econ.
Entomol. vol. 94(2), pp. 326-331.
Spivak, M. and Reuter, G. S. 2005. A Sustainable Approach to Controlling Honey Bee
Diseases and Varroa Mites. Web sitesi: http://www.extension.org/mediawiki
/files/4/40/AsustainableApproachtoControllingHoneyBeeDiseaseandVarroaMites.
pdf. Erişi Tarihi: 25/02/2009.
Szabo, T. I., Walker, C. R. T. and Mueller, A. E. 1996. Grooming behavior as a Varroa
resistance characteristic in honey bee colonies. Am. Bee J. vol. 136, pp. 515-517.
Taber, S. 1982. Determining resistance to brood diseases. American Bee Journal, vol.
122(6), 422-425.
Taber, S., and Gilliam, M. 1987. Test for resistance to chalkbrood disease, Ascosphaera
apis, of honeybees. The XXXIst International Congress of Apiculture, Warsaw,
Poland, pp. 145-148.

79
Tahmasebi, G. H., Ebadi, R., Esmaili, M. and Kambusia, J. 1998. Morphological study
of honeybee (Apis mellifera L.) in Iran. J. Agric. Sci. And Nat. Res. vol. 2(1), pp.
89-101.
Thakur, R. K., Bienenfeld, K. and Keller, R. 1997. Varroa defense behavior in Apis
mellifera carnica. Am. Bee J. vol. 137, pp. 143-148.
Thompson, V. C. 1964. Behaviour genetics of nest cleaning in honey bees. III. Effect of
age of bees of a resistant line on their response to disease-killed brood. J. Apic.
Res. vol. 3, pp. 25–30.
Tubbs, H., Harper, C., Bigalk, M., Bernard, S. J., Delatte, G. T., Sylvester, H. A. and
Rinderer, T. E. 2003. Commercial management of ARS Russian honey bees. Am.
Bee J. vol. 144, pp. 819-820.
Vandame, R., Colin, M., Morand, S. and Otero-Colina, G. 2000. Levels of compatibility
in a new host parasite association: Apis mellifera / Varroa jacobsoni. Can. J. Zool.
vol. 78, pp. 2037-2044.
Vandame, R., Morand, S., Colin, M.E. and Belzunces, L.P. 2002. Parasitism in the
social bee Apis mellifera: quantifying costs and benefits of behavioral resistance
to Varroa destructor mites. Apidologie, vol. 33, pp. 433–445.
Villa, J.D., Danka, R.G. and Harris, J.W. 2009. Simplified methods of evaluating
colonies for levels of Varroa sensitive hygiene (VSH). J. Apic. Res., vol. 48, pp.
162–167.
Wilkes, K. and Oldroyd, B. 2002. Breeding hygienic disease resistant bees. A report for
the Rural Industries Research and Development Corporation. RIRDC Publication
No 02/048, RIRDC Project No. US-39A.
Yang, X. and D. L. Cox-Foster. 2005. Impact of an ectoparasite on the immunity and
pathology of an invertebrate: Evidence for host immunosuppression and viral
amplification. Proceeding of the National Academy of Sciences of the United
States of America, 102, pp. 7470-7475.

80
ÖZGEÇMİŞ
Adı Soyadı : Amir NAJI KHOEI
Doğum Yeri : Iran - Khoy
Doğum Tarihi : 23.08.1967
Medeni Hali : Evli
Yabancı Dili : İngilizce
Eğitim Durumu (Kurum ve Yıl)
Lise : Ayatollah Talegani Lisesi, 1985
Lisans : Tebriz Üniversitesi Ziraat Fakültesi Zootekni Bölümü (1990)
Yüksek Lisans : Mazandaran Üniversitesi Ziraat Fakültesi Zootekni Bölümü (2001)
Çalıştığı Kurum/Kurumlar ve Yıl
Tebriz Üniversitesi Ziraat Fakültesi Zootekni Bölümü 1993-2012
Yayınları
Naji-Khoei, A. Fıratlı, Ç. Alijani, S. Kaşko-Arıcı. Khodie, S. Behavioral defenses
of Iranian honey bees (Apis mellifera meda) against Varroa destructor. Annals of
Biological Research, 2011, 2 (6):510-516.
Naji-Khoei, A. Nejati-Javaremi, A. Tahmasebi, G. H. 2007. Using simulation to
predict the impact of using economic weights in genetic improvement of honey
bees. Pajouhesh and Sazandegi, 76, pp. 29-34.
Naji-Khoei, A. Nejati-Javaremi, A. Tahmasebi, G. H. 2003. Using simulation to predict the impact of using economic weights in genetic improvement of honey bees. Fifth Iranian seminar on honey bee research.

81
Naji-Khoei, A. Nejati-Javaremi, A. Tahmasebi, G. H. 2000. Economic weight estimation of some important characteristics in the honeybee. Fourth Iranian seminar on honey bee research.
4- Özdil, F. Fakhri, B. Naji khoei, A. Erkan, C. Yildiz, M. A. 2008. HinfI and DraI digestion of the COI-COII intergenic region in honey bee (Apis mellifera L.) Population from Turkey and western part of Iran. 3rd European conference of Apidology . Belfast 8-11 September 2008.
5- Gedik Y. Fakhri, B. Naji Khoei, A. Meydan H. Özdil, F. Yildiz, M. A. 2007. Molecular characterization of A. m. meda populations from the west part of Iran. IBRA International conference on Recent Trends in Apicultural Science. 10-14 June 2007.