exploring spatiotemporal patterns in early stages of primary succession on former lignite mining...
TRANSCRIPT

- Exploring spatiotemporal patterns in early stages of primary succession - 267Journal of Vegetation Science 19: 267-276, 2008doi: 10.3170/2008-8-18369, published online 1 February 2008© IAVS; Opulus Press Uppsala.
Exploring spatiotemporal patterns in early stages of primary succession on former lignite mining sites
Felinks, Birgit1* & Wiegand, Thorsten2
1UFZ Helmholtz Centre for Environmental Research - UFZ Department of Conservation Biology, DE-04318 Leipzig, Germany; 2UFZ Helmholtz Centre for Environmental Research - UFZ, Department of Ecological Modelling, DE-04318 Leipzig, Germany;
E-mail [email protected]; *Corresponding author; Present address Anhalt University of Applied Sciences, Strenzfelder Allee 28, DE-06406 Bernburg, Germany; Fax +49 347135591131; E-mail [email protected]
AbstractQuestions: 1. Does random colonization predominate in early stages of primary succession? 2. Do pioneer species facilitate the establishment of later arriving species? 3. Does an initially random distribution change to an aggregated pattern with ongoing succession?Location: Lignite mining region of Lower Lusatia, eastern Germany.Methods: Individual plants were mapped along a 2 m × 28 m transect during three successive years and classified into two groups (1) the pioneer Corynephorus canescens and (2) ‘all other species’. Using the pair-correlation function, univariate point pattern analysis was carried out by applying a hetero-geneous Poisson process as null model. Bivariate analysis and a toroidal shift null model were applied to test for independence between the spatial patterns of the two groups separately for each year, as well by exploring spatiotemporal patterns from different years. Results: In the first year Corynephorus and ‘all other species’ showed an aggregated pattern on a spatial scale > 40 cm and in the second and third years a significant attraction for distances between 4 and 12 cm, with an increasing radius in the third year. The analyses of interspecific spatiotemporal dynamics revealed a change from independence to attraction between distances of 4 cm and 16 cm when using Corynephorus as focal species. However, applying ‘all other species’ as focal points results in a significant attraction at distances up to 60 cm in the first year and a diminishing attraction in the second and third years with distances ≤ 6 cm. Conclusions: Facilitative species-species interactions are present in early stages of primary succession, resulting mainly from pioneer species acting as physical barriers and their ability to capture diaspores being drifted by secondary dispersal along the substrate surface. However, due to gradual establishment of perennial species and their ability of lateral extension by vegetative dispersal, facilitation may influence spatial pattern formation predominantly on short temporal and fine spatial scales.
Keywords: Corynephorus; Facilitation; Pair-correlation func-tion; Point pattern analysis; Species interaction.
Nomenclature: Wisskirchen & Haeupler (1998).
Introduction
The study of vegetation patterns on different spatial scales has long been of great importance in plant ecol-ogy (e.g. Watt 1947; Lepš 1990). When studying the interactions between different species or life stages, the aim is to detect attraction, suggesting positive associa-tion between two species or two life stages, repulsion, implying (for example) interspecific competition (Kenkel 1988; Callaway & Walker 1997; Bossdorf et al. 2000; Pélissier & Goreaud 2001; Couteron et al. 2003; Wiegand et al. 2007a) or segregation at larger scales implying (for example) use of different habitats in case of envi-ronmental heterogeneity. As the spatial pattern of one or few dominant species may affect the spatial pattern of associated species, an interpretation of spatial pattern may also help to refine our understanding of ecological processes relevant in vegetation succession (Dale 1999). In general, competition is assumed to cause a systematic change in spatial pattern from random or aggregated to (more) regular pattern as a result of density dependent mortality (e.g. Sterner et al. 1986; Moeur 1997; Batista & Maguire 1998; Barot et al. 1999; Freeman & Ford 2002; Woodall & Graham 2004). The negative effects of interspecific competition can be offset by facilita-tion if physical conditions are sufficiently improved by the central species ‘micro-ecosystems’ (Garner & Steinberger 1989; Bertness & Callaway 1994). Thus, particularly in dryland ecosystems a large proportion of vegetation reveals an aggregated pattern (Prentice & Werger 1985; Aguiar & Sala 1994; Pugnaire et al. 1996; Tirado & Pugnaire 2003; but see Wiegand et al. 2006). The interest in studying spatial pattern has also been driven by the need to justify the relevant scale of the study and to develop hypotheses on the underlying processes that can be tested in experimental studies (Cale et al. 1989; Perry et al. 2002; Wiegand & Moloney 2004; Wiegand et al. 2007a).
However, studying the interplay between processes and emerging patterns is often complicated in established

268 Felinks, B. & Wiegand, T.
an initially random distribution change to an aggregated pattern with ongoing succession?
Methods
Study area, species, and data collection
The study was carried out in a lignite mining region of Lower Lusatia, eastern Germany (51°47’ N, 13°50’ E). Mean annual temperature is 8.9 °C and mean an-nual precipitation ca. 590 mm (Anon. 1961-1990). The heaps consist of predominantly nutrient-poor, sandy substrates (Wiegleb & Felinks 2001). On major parts of the heaps restoration measures such as levelling or amelioration have been carried out and large areas have been afforested or are currently used for agriculture. Nevertheless, some sites were left to spontaneous suc-cession. According to the prevailing substrate conditions the present vegetation on such sites is characterized by sparse vegetation, Corynephorus canescens dominated grasslands, species-rich psammophytic grasslands with varying cover of mosses and lichen- or Calamagrostis epigejos stands (Wiegleb & Felinks 2001).
Field sampling for the present study was carried out on a site where mining activities were closed down ca. 20 years ago. Due to ongoing restoration measures, vegeta-tion succession ‘started’ ca. 5 years before the study was initiated. A linear 2 m × 28 m transect was set up in a representative area within the study site, revealing a slight 5% slope from the right to the left (Fig. 1). The point locations of all individual plants were mapped during three successive years (1995-1997) using a 1-m² frame divided in 10-cm² grid fields, which allows a recording of the x and y co-ordinates of the individuals with a precision of ca. 2 cm. The point-data were digitized with AutoCAD 13TM and the data were stored as a list of x and y co-ordinates. Because it is not easy to map individual sprouts of grasses, tufts with a diameter > 5 cm were approximated by several points representing the integer co-ordinates within a polygon of the respective diameter (see App. 1, Fig. A1). Note that the approximation of larger individuals as points may introduce clustering at small scales which correspond to the typical tuft size (Wiegand et al. 2006; see below).
The species were classified into two groups: (1) the pioneer species Corynephorus canescens and (2) ‘all other species’. Corynephorus is considered as a typical pioneer on sandy substrates (Marshall 1967; Symonides 1985). This perennial is capable of renewing its tufts layer by layer and establishing a rich secondary root system. Furthermore, it is characterized by a short pre-reproductive period and a high fecundity of individuals. The diaspores are dispersed by wind. A further 14 annual
vegetation where many processes and factors operate simultaneously. One approach to unravel this question is to study simpler systems, such as former mining sites where primary succession occurs (Wiegleb & Felinks 2001). The general patterns described above can then be translated into a simple conceptual model. The ini-tial successional stage will be characterized by random colonization with pioneer species. Once established, the pioneer species will facilitate the establishment and growth of later arriving species, hence, the random dis-tribution should change to an aggregated univariate and attracted bivariate pattern. In later successional stages, when vegetation cover has become dense, competition processes should prevail causing more regular univariate patterns and repulsive bivariate patterns.
One approach to explore spatial pattern formation during succession is to use point pattern analysis of fully mapped point locations of plants (Ripley 1981; Upton & Fingleton 1985; Cressie 1991; Dale et al. 2002; Diggle 2003). Spatial statistics such as Ripley’s K function (Rip-ley 1977, 1981) and the pair-correlation function (Stoyan & Stoyan 1994; Wiegand & Moloney 2004) can be used to describe the small-scale spatial correlation structure of a point pattern which contains information on the type of interactions among plants. However, spatial pattern analyses are usually based on a single point in time and few studies have investigated point pattern in the context of ongoing primary succession (but see Zhang & Skarpe 1995; Peterson & Squiers 1995; Wiegand et al. 1998; Malkinson et al. 2003; McDonald et al. 2003). Because the relative importance of ecological processes and the nature of ecological interactions between individuals may change with time (Connell & Slatyer 1977; Tilman 1985; Bertness & Callaway 1994) it is also expected that spatial pattern will change during the course of succession (Juhász-Nagy & Podani 1983; Bartha et al. 1995).
The present study explores the spatiotemporal pat-terns observed during three years in early stages of primary succession on former lignite mining sites in eastern Germany. As a result of the long lasting, open-cast mining activities neither plants nor diaspores remained on the sites, thus, they provide a unique opportunity for the study of primary succession in central Europe. Our data are also of general interest since little attention has been dedicated to spatial pattern of plant communities dominated by grasses and herbs in temperate regions.
Based on a classification of the mapped plants into the pioneer Corynephorus canescens and a second group which encompasses all other species (‘all other species’) we asked the following questions guided by the conceptual model of primary succession. First, does random colonization predominate in early stages of pri-mary succession? Second, do pioneer species facilitate the establishment of later arriving species? Third, does

- Exploring spatiotemporal patterns in early stages of primary succession - 269
and 22 perennial species were encompassed in the group of ‘all other species’. Throughout the three sampling years the mean proportion of annual individuals is 65% and the perennials 35%, respectively. Conyza canadensis, Arenaria serpyllifolia and Corispermum leptopterum represent almost 95% of the annual individuals and Achillea millefolium, Tussilago farfara, Artemisia camp-estris, Echium vulgare, Ononis repens and Calamagrostis epigejos representing ca. 90% of the perennials. Conyza and Corispermum are wind dispersed, the diaspores of A. serpyllifolia are mainly dispersed by gravity. Except the anemochorous A. millefolium and Calamagrostis all perennial species are mainly dispersed by gravity. All perennials, except E. vulgare, are capable of vegetative dispersal by elongated rhizomes or stolons.
Statistical analyses
We used the pair-correlation function (Stoyan & Stoyan 1994; Wiegand & Moloney 2004) to explore the observed spatiotemporal patterns. The pair correlation function is closely related to Ripley’s K-function (Ripley 1976); both are based on the distribution of distances of all pairs of points of the patterns. The univariate K-function K(r) can be defined as the expected number of points within distance r of an arbitrary point (without counting the focal points), divided by the intensity λ of pattern (Ripley 1976). The univariate pair-correlation function g(r) is related to the derivative of the K-function i.e. g(r) = K’(r)/(2πr) (Ripley 1977; Stoyan & Stoyan 1994). Analogously, the bivariate K-function K12(r) is the expected number of type 2 points within distance r of an arbitrary type 1 point, divided by the intensity of type 2 points. Note that we do not combine the two bivariate functions K12 and K21 into a single estimator as proposed by Lotwick & Silverman (1982). Further details can be found in standard textbooks (e.g. Stoyan & Stoyan
1994; Diggle 2003). The pair-correlation function is a non-cumulative statistic which isolates different distance classes and does not integrate the ‘memory’ of small scale second-order effects to larger scales, as does Ripley’s K (Wiegand & Moloney 2004) and is recommended for exploratory data analysis (Stoyan & Stoyan 1994; Stoyan & Penttinen 2000). We followed the approach of Wiegand & Moloney (2004) for considering edge effects and for estimating the pair-correlation function.
In order to describe the spatiotemporal structure of the observed patterns and to relate them to expectations from the conceptual model we compared the observed patterns to simple null models. Significant departure from a given null model was tested by comparing the pair-correlation function of the data to simulation envelopes generated from multiple Monte Carlo simulations of the null model. We used ca. 5% simulation envelopes which were the 25th lowest and highest values from 999 Monte Carlo simulations of the null-model. The 999 simulations of the null model generated sufficiently smooth envelopes for our explorative data analysis.
Univariate analysis Visualization of the univariate pattern of the 1995
snapshot (Fig. 1A) suggested a gradient with low point densities at the left end of the gradient and high densi-ties at the right end. Since the highest point is at the right end of the transect, a clumping of individuals due to fine scale erosion or washing away of diaspores has to be considered as irrelevant. This pattern may also result from an initial establishment on the right end of the transect previous to our study. However, the gradient in point density observed in the first year almost disap-peared during the next two years (Fig. 1B, C). To be able to describe the small-scale structure of the univariate patterns, which might be superposed to the larger scale gradient, we used a heterogeneous Poisson process as
Fig. 1. The observed spatial patterns of three years of primary succession on a former lignite mining site along a 2 m × 28 m transect. A = (1995), B = (1996) and C = (1997). Grey dots = Corynephorus canescens, = ‘all other species’.

270 Felinks, B. & Wiegand, T.
null model which retained the large-scale structure of the pattern but removed its small-scale correlation structure. In turn, if the pattern was not heterogeneous, the heterogeneous Poisson process will collapse to a homogeneous Poisson process. Note that approximating larger individuals as points will introduce (when using the heterogeneous Poisson null model) a clustering up to scales which correspond to the typical tuft sizes. An approach to deal with plants of finite size was recently proposed by Wiegand et al. (2006), but because of dif-ficulties in unambiguously defining individuals in our data set (see detailed data in App. 1) we decided to ap-proximate larger tufts by various points.
In heterogeneous Poisson point processes the occur-rence of any point is independent of that of any other, but the points are distributed in accordance with an intensity function λ (x, y) that varies with location (x, y) (Stoyan & Stoyan 1994; Wiegand & Moloney 2004; Wiegand et al. 2007b). We used an Epanečnikov kernel, a non-parametric method recommended by Stoyan & Stoyan (1994), to estimate the intensity function of a given point pattern. In short, for a given location (x, y) the intensity λ (x, y) is constructed by using a moving window with circular shape and radius h around location (x, y) and summing up all points in the circle, but weighting them with the Epanečnikov kernel eh(d), according to their distance d from the focal location (x, y). For distance d smaller than the bandwidth h the Epanečnikov kernel is defined as eh(d) = 0.75(1 - d2/h2)/h and otherwise it is eh(d) = 0. Thus, spatial structures smaller than the bandwidth h are removed from the pattern. To avoid edge effects if the moving window was partly outside the study region we divided by the area of the moving window inside the study region. To capture the broad gradient in point density along the gradient we selected a bandwidth h = 0.8 m which is almost half the width of the transect. If the observed pair-correlation function was above or below the simulation envelopes, the pattern showed additional small-scale aggregation or regularity, respectively.
Bivariate spatial analysesIn our bivariate analyses we were interested to find
indications of positive or negative association between two patterns. This question suggests the use of a toroidal shift null model (Goreaud & Pélissier 2003) which pro-vides a non-parametric test for independence between two patterns. This null model preserves the separate, and sometimes complicated, second order structures of both pattern but breaks the dependence between them. One way of achieving this is by simulations that involve random shifts of the whole pattern two relative to pattern one, which is kept fixed (Dixon 2002). In practice, a rectangular study region is treated as a torus where the upper and lower edges are connected and the
right and left edges are connected. If the observed pair-correlation function was above or below the simulation envelopes the two patterns showed attraction or repul-sion, respectively. Note that the toroidal shift does not change the spatial structure of the component patterns; approximating larger individuals as points does therefore not introduce artefacts.
Bivariate spatiotemporal analysesWe also used the independence null model for study-
ing the inter- and intraspecific spatiotemporal patterns in different years. We used the toroidal shift method to randomize the later pattern and thus test for independ-ence of the pattern at year t+1 (or t+2) from a pattern at year t. When we analysed two patterns of the same species group from different years and the observed pair-correlation function was at small scales above the simulation envelopes the pattern showed an expansion from the focal points of the earlier pattern. If we analysed two patterns of different species groups and the observed pair-correlation function was at small scales above or below the simulation envelopes, pattern 2 was facilitated or repulsed by the earlier pattern, respectively.
Point pattern analysis was carried out in three steps including univariate analyses to answer questions one and three, bivariate analyses of the patterns observed at each of the three years to answer question two, and finally bivariate analyses of all pattern combinations taken from different years to look at the dynamic aspect of question two. For all analyses we used the grid-based software Programita. Details on the implementation of the second-order statistics are provided in Wiegand & Moloney (2004). We used a spatial resolution of 2 cm for all analyses, which approximates the mapping accuracy of the point patterns
Results
Colonization and establishment
During the three years of field sampling a continuous increase in the number of individuals of Corynephorus as well as of ‘all other species’ was observed (Fig. 1). In the first year Corynephorus could be mapped by single sprouts, by the second year many of the sprouts had increased to tufts with a mean diameter of 5 cm. In the third year the mean diameter of the tufts had extended to 10 cm. The increasing tuft size indicates that Corynephorus has to be considered as a successful pioneer colonizer. The space between the grass tufts was colonized by Corynephorus and ‘all other species’. Other species growing directly in the Corynephorus tufts were merely observed.
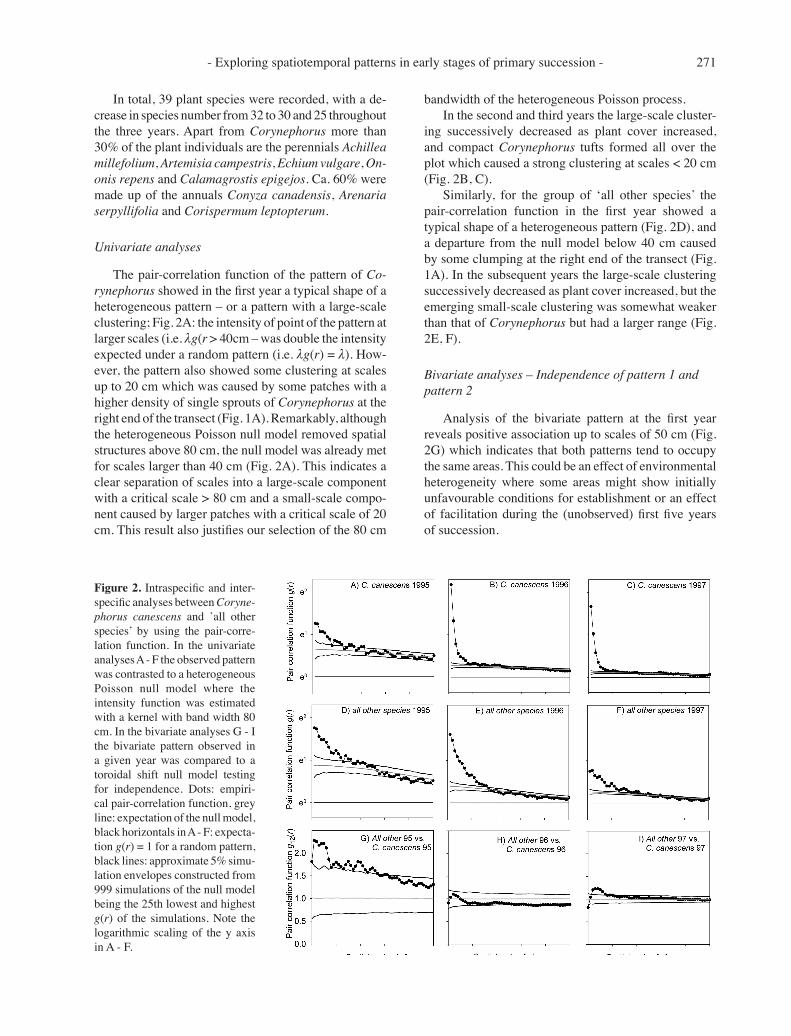
- Exploring spatiotemporal patterns in early stages of primary succession - 271
In total, 39 plant species were recorded, with a de-crease in species number from 32 to 30 and 25 throughout the three years. Apart from Corynephorus more than 30% of the plant individuals are the perennials Achillea millefolium, Artemisia campestris, Echium vulgare, On-onis repens and Calamagrostis epigejos. Ca. 60% were made up of the annuals Conyza canadensis, Arenaria serpyllifolia and Corispermum leptopterum.
Univariate analyses
The pair-correlation function of the pattern of Co-rynephorus showed in the first year a typical shape of a heterogeneous pattern – or a pattern with a large-scale clustering; Fig. 2A: the intensity of point of the pattern at larger scales (i.e. λg(r > 40cm – was double the intensity expected under a random pattern (i.e. λg(r) = λ). How-ever, the pattern also showed some clustering at scales up to 20 cm which was caused by some patches with a higher density of single sprouts of Corynephorus at the right end of the transect (Fig. 1A). Remarkably, although the heterogeneous Poisson null model removed spatial structures above 80 cm, the null model was already met for scales larger than 40 cm (Fig. 2A). This indicates a clear separation of scales into a large-scale component with a critical scale > 80 cm and a small-scale compo-nent caused by larger patches with a critical scale of 20 cm. This result also justifies our selection of the 80 cm
bandwidth of the heterogeneous Poisson process.In the second and third years the large-scale cluster-
ing successively decreased as plant cover increased, and compact Corynephorus tufts formed all over the plot which caused a strong clustering at scales < 20 cm (Fig. 2B, C).
Similarly, for the group of ‘all other species’ the pair-correlation function in the first year showed a typical shape of a heterogeneous pattern (Fig. 2D), and a departure from the null model below 40 cm caused by some clumping at the right end of the transect (Fig. 1A). In the subsequent years the large-scale clustering successively decreased as plant cover increased, but the emerging small-scale clustering was somewhat weaker than that of Corynephorus but had a larger range (Fig. 2E, F).
Bivariate analyses – Independence of pattern 1 and pattern 2
Analysis of the bivariate pattern at the first year reveals positive association up to scales of 50 cm (Fig. 2G) which indicates that both patterns tend to occupy the same areas. This could be an effect of environmental heterogeneity where some areas might show initially unfavourable conditions for establishment or an effect of facilitation during the (unobserved) first five years of succession.
Figure 2. Intraspecific and inter-specific analyses between Coryne-phorus canescens and ’all other species’ by using the pair-corre-lation function. In the univariate analyses A - F the observed pattern was contrasted to a heterogeneous Poisson null model where the intensity function was estimated with a kernel with band width 80 cm. In the bivariate analyses G - I the bivariate pattern observed in a given year was compared to a toroidal shift null model testing for independence. Dots: empiri-cal pair-correlation function, grey line: expectation of the null model, black horizontals in A - F: expecta-tion g(r) = 1 for a random pattern, black lines: approximate 5% simu-lation envelopes constructed from 999 simulations of the null model being the 25th lowest and highest g(r) of the simulations. Note the logarithmic scaling of the y axis in A - F.

272 Felinks, B. & Wiegand, T.
In the second year we observed a tendency to repul-sion which becomes marginally significant at scales > 50 cm and in the third year a significant attraction at scales between 4 cm and 12 cm (Fig. 2H, I). The small-scale repulsion found in the third year could be an artefact of idealizing plants of finite size as points (Wiegand et al. 2006) but may also indicate competition between the two patterns. The attraction at scales up to 12 cm indi-cates a facilitation effect where individuals of ‘all other species’ grow in the neighbourhood of the colonizer Corynephorus.
Bivariate analyses – Spatiotemporal pattern
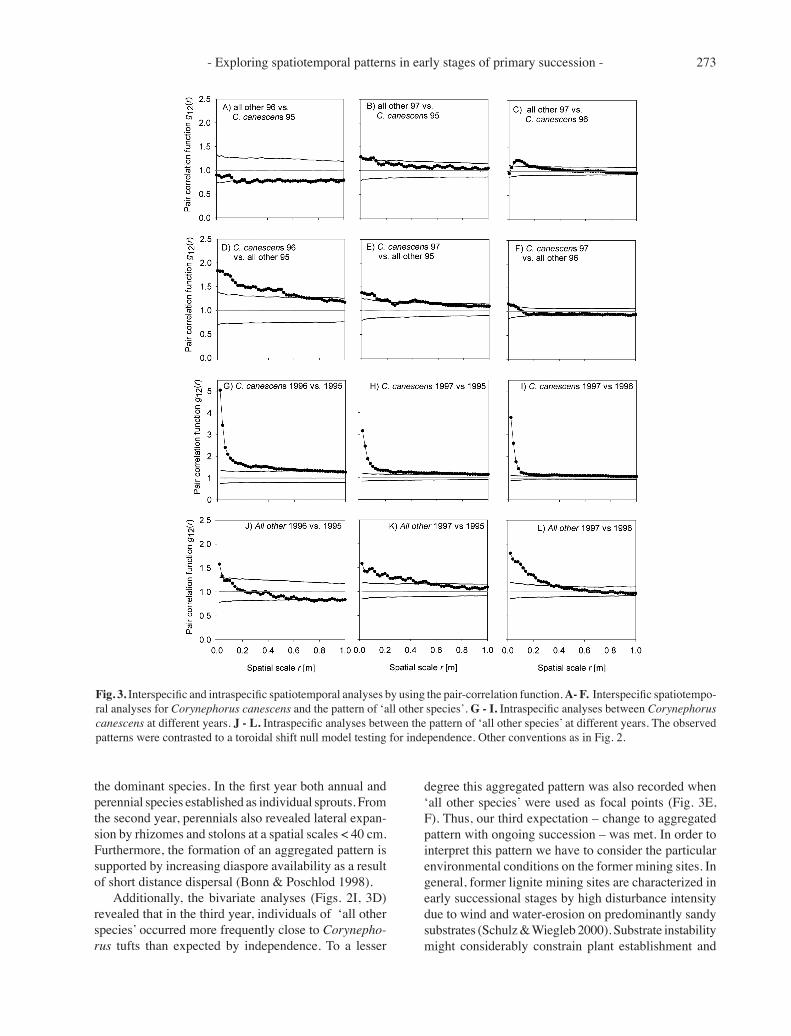
The analysis of point pattern of Corynephorus from the first year in relation to pattern of ‘all other species’ from the second year revealed no significant attraction or repulsion between the two species groups (Fig. 3A) but showed a tendency to repulsion. This is contrary to our expectation, but it has to be considered that the individual Corynephorus sprouts do not act as physical barriers for diaspores being drifted over the bare substrate surface. However, point pattern of Corynephorus taken from the second year and ‘all other species’ from the third year clearly showed an attraction between distances of 4 cm and 16 cm (Fig. 3C). In contrast, the analysis of Corynephorus in 1995 and ‘all other species’ in 1997 revealed only a weak significant attraction at spatial scales between 0 and 12 cm (Fig. 3B). This short memory indicates that colonization processes at the initial suc-cession phase are very dynamic and facilitation effects may interfere with effects resulting from biological traits of the colonizing species as, for example, the ability of lateral expansion by rhizomes.
Analysis of the spatiotemporal pattern of ‘all other species’ vs Corynephorus showed a significant attraction for spatial scales up to r = 60 cm (Fig. 3D) for the 1996 Corynephorus pattern in relation to the 1995 pattern of ‘all other species’. This attraction is probably due to a few large individuals of Ononis repens or Artemisia campestris which may efficiently capture diaspores by their outspread branches. However, when relating pat-tern of Corynephorus of the third year with pattern of ‘all other species’ from the second year, we found only weak attraction which was restricted to spatial scales ≤ 6 cm (Fig. 3F). Considering the influence of ‘all other species’ of the first year on Corynephorus of the third year, low but significant attraction is reported up to 20 cm (Fig. 3E). Thus, it has to be assumed that the colonizer species Corynephorus was facilitated by the previous spatial pattern of all other species, especially in the second year.
Strong attraction can also be inferred from an analysis of intraspecific spatiotemporal analyses (Fig. 3G-L).
Interaction of point pattern for Corynephorus from the first with the second year results, in all cases, in an at-traction up to 60 cm (Fig. 3G-I) and simply indicates considerable growth of perennial tufts. The patterns of all other species were only weakly associated between the first two years and even showed a tendency to repul-sion (Fig. 3J, K). The pattern of the third year showed strong positive association to the patterns of the two previous years indicating that this species group was able to form larger individuals due to the development of rhizomes and stolons during the second and third years of observation (Fig. 3L).
Discussion
The detailed exploratory analyses of the spatiotempo-ral patterns of primary succession observed at a former lignite mining site partly confirmed our conceptual model in that strong and facilitative species-species interac-tions were found. However, the expectation of random colonization (e.g. Bartha 1992; Geißelbrecht-Taferner et al. 1997; Tirado & Pugnaire 2003) was not met for the first year of observation; the two species groups already showed aggregated patterns at a larger scale. This is not necessarily evidence against our conceptual model; the patterns at the first year of observation could already be the result of expansion after random colonization prior to our observation period. A pattern formation as a consequence of microrelief characteristics is less likely, because the entire transect was colonized at the final year of observation.
A comparison of the results of the intra- and inter-specific analyses of the same year, and of subsequent years, clearly showed that the colonization of bare soils at former mining sites is a non-random process where strong positive and negative plant-plant interactions oc-cur. The strongest positive effect was demonstrated by intraspecific analyses. For Corynephorus this was simply an increase in tuft size (Fig. 3G-I; see also Symonides 1985). For ‘all other species’ an interesting pattern was observed: in the second year a positive association to the pattern of the first year occurred only in the immediate 4 cm neighbourhood and showed a tendency to repulsion at larger scales (Fig. 3J). This indicates that few of the plants of 1995 served as ‘expansion nuclei’ for the pat-tern of the second year and that a substantial proportion of new recruitment occurred in the second year at some distance from the 1995 plants. Comparing the pattern showing Fig. 1A, B confirms this. However, in the first or second years to the third year of observed succession an aggregated pattern was observed at a spatial scale < 40 cm (Fig. 3K-L), which corresponds to biological characteristics, such as dispersal and growth form, of
- Exploring spatiotemporal patterns in early stages of primary succession - 273
Fig. 3. Interspecific and intraspecific spatiotemporal analyses by using the pair-correlation function. A- F. Interspecific spatiotempo-ral analyses for Corynephorus canescens and the pattern of ‘all other species’. G - I. Intraspecific analyses between Corynephorus canescens at different years. J - L. Intraspecific analyses between the pattern of ‘all other species’ at different years. The observed patterns were contrasted to a toroidal shift null model testing for independence. Other conventions as in Fig. 2.
the dominant species. In the first year both annual and perennial species established as individual sprouts. From the second year, perennials also revealed lateral expan-sion by rhizomes and stolons at a spatial scales < 40 cm. Furthermore, the formation of an aggregated pattern is supported by increasing diaspore availability as a result of short distance dispersal (Bonn & Poschlod 1998).
Additionally, the bivariate analyses (Figs. 2I, 3D) revealed that in the third year, individuals of ‘all other species’ occurred more frequently close to Corynepho-rus tufts than expected by independence. To a lesser
degree this aggregated pattern was also recorded when ‘all other species’ were used as focal points (Fig. 3E, F). Thus, our third expectation – change to aggregated pattern with ongoing succession – was met. In order to interpret this pattern we have to consider the particular environmental conditions on the former mining sites. In general, former lignite mining sites are characterized in early successional stages by high disturbance intensity due to wind and water-erosion on predominantly sandy substrates (Schulz & Wiegleb 2000). Substrate instability might considerably constrain plant establishment and

274 Felinks, B. & Wiegand, T.
growth (Roughgarden & Diamond 1986; Prach 1987; van der Valk 1992). Thus, the first colonizers which established on bare ground may act as ‘nurse plants’ and facilitate the subsequent establishment of later ar-riving individuals via different mechanisms (Malkinson et al. 2003). Furthermore, both the dominant annual and perennial species are mainly dispersed by gravity. Thus, the physical barriers formed by the dense and compact tufts of Corynephorus in the second and third years of observation, but also the larger – though less dense – tufts of perennial species efficiently capture diaspores which are drifted by secondary dispersal above the substrate surface (van der Valk 1992; Chambers & MacMahon 1994). Consequently, establishment of new species or individuals occurs predominantly at the edges of already existing vegetation structures (Grubb 1985; Armesto et al. 1991; Salonen 1992; Salonen et al. 1992; del Moral 1993; Aguiar & Sala 1994; Holmgren et al. 1997; Schütz 2000; Fuller & del Moral 2003).
Furthermore, sand movement may be considerably reduced at the edges of established plants because of reduced wind speed or water erosion. As a result, a more benign environment is provided on a fine spatial scale where survival probabilities of newly arriving species are higher. Consequently, founder effects (Paine 1980) may trigger vegetation development in early successional stages via positive feedback mechanisms; pioneer species establish and they contribute to an increase of substrate stability. These conditions allow the establishment of later arriving species which contribute to increasing substrate stability. Such a phenomenon that randomly established initial colonizers may provide ‘nuclei’ for the following vegetation development was referred to as nucleation when it occurs during primary succession (Yarranton & Morrison 1974; Blundon et al. 1993).
An interesting outcome of our analyses is the un-expected dynamic relationship between the two types of point pattern (Fig. 2G-I). The two patterns occurred in the first year of colonization in the same areas of the transect, they were independent with tendency to repul-sion in the second year and in the third year, individuals occurred more frequent close to Corynephorus tufts than expected at random.
Results of the interspecific spatiotemporal analyses revealed that points of year (t) were, in most cases, signifi-cantly attracted by points of the year (t – 1), irrespective of the type of points that were set as pattern 1 (t - 1) or pattern 2 (t) (Fig. 3A-F). The bivariate analyses of the relationship of points of year (t – 2) with point pattern from year (t) showed that although attraction of the two types of point is still present and significant, it is less intensive and restricted to a narrower spatial scale. This indicates that the initial three year phase of the succession proceeded quite dynamic with a relatively short ‘memory’ of two years. Such a dynamic
pattern development might be partly influenced by positive interaction of the colonizing plants themselves (Zobel et al. 1993, 1996). Whereas the compact tufts of Corynephorus may pronounce the spatial pattern established in initial succession stages, the observed facilitation effect of ‘all other species’ diminishes within a shorter period of time, probably as a consequence of the vegetative dispersal of the dominant perennial species.
Our analyses, which investigate spatiotemporal pattern of succession, are of special interest since such analyses are not common (but see e.g. Peterson & Squiers 1995; Zhang & Skarpe 1995; Wiegand et al. 1998; Mal-kinson et al. 2003; McDonald et al. 2003) and at present few detailed analyses of founder effects in former mining sites are available (Bartha 1992; Tischew 2004). Ac-cording to our spatiotemporal point-pattern analyses it might be concluded that the establishment of new plant individuals benefits from already existing vegetation either because of the presence of physical barriers or from enhanced diaspore availability. However, the resulting spatial patterns could be detected only during a short time period. Consequently, facilitation may influence spatial pattern formation predominantly at the early stages of a primary succession on rather short temporal and fine spatial scales. Since establishment from the diaspore bank will increase and physical barriers will become less important, and additionally environmental condi-tions might be less extreme, facilitation may be quickly substituted by competition with ongoing succession (Zobel et al. 1993; Bertness & Callaway 1994; Callaway & Walker 1997) – at least as long as bare sites are not recreated by disturbance events. However, evidence for increasing importance of competition cannot be inferred from the results of the present study, because we only monitored three years.
Acknowledgements. Data collection was funded by the German Federal Ministry of Education and Science (BMBF, Grant-No.0334896) and the Lausitz and Central-German Min-ing Administration Company (LMBV). We thank Philip Dixon and three anonymous reviewers for their helpful comments on an earlier version of this manuscript.
References
Anon. 1961-1990. DWD Deutscher Wetterdienst http://www.dwd.de/de/FundE/Klima/KLIS/daten/online/nat/index_mittelwerte.htm.
Aguiar, M.R. & Sala, O.E. 1994. Competition, facilitation, seed distribution and the origin of patches in a Patagonian steppe. Oikos 70: 26-34.
Armesto, J.J., Pickett, S.T.A. & McDonnell, M.J. 1991. Spatial heterogeneity during succession: A cyclic model of invasion and exclusion. Springer, New York, NY, US.

- Exploring spatiotemporal patterns in early stages of primary succession - 275
Barot, S., Gignoux, J. & Menaut, J.C. 1999. Demography of a savanna palm tree: Prediction from comprehensive spatial pattern analyses. Ecology 80: 1987-2005.
Bartha, S. 1992. Preliminary scaling for multi-species coalitions in primary succession. Abstracta Botanica 16: 31-41.
Bartha, S., Czárán, T. & Oborny, B. 1995. Spatial constraints masking community assembly rules: a simulation study. Folia Geobotanica and Phytotaxonomica 30: 471-482.
Batista, J.L.F. & Maguire, D.A. 1998. Modelling the spatial structure of tropical forest. Forest Ecology and Manage-ment 110: 293-314.
Bertness, M.D. & Callaway, R.M. 1994. Positive interactions in communities. Trends in Ecology and Evolution 9: 187-191.
Blundon, D.J., MacIsaac, D.A. & Dale, M.R.T. 1993. Nuclea-tion during primary succession in the Canadian Rockies. Canadian Journal of Botany 71: 1093-1096.
Bonn, S. & Poschlod, P. 1998. Ausbreitungsbiologie der Pflan-zen Mitteleuropas. Quelle & Meyer, Wiesbaden, DE.
Bossdorf, O., Schurr, F. & Schumacher, J. 2000. Spatial patterns of plant association in grazed and ungrazed shrublands in the semi-arid Karoo, South Africa. Journal of Vegetation Science 11: 253-258.
Cale, W.G., Henebry, G.M. & Yeakley, J.A. 1989. Inferring process from pattern in natural communities – can we understand what we see? BioScience 39: 600-605.
Callaway, R.M. & Walker, L.R. 1997. Competition and fa-cilitation: a synthetic approach to interactions in plant communities. Ecology 78: 1958-1965.
Chambers, J.C. & MacMahon, J.A. 1994. A day in the life of a seed: movements and fates of seeds and their implica-tions for natural and managed systems. Annual Review of Ecology and Systematics 25: 263-292.
Connell, J.H. & Slatyer, R.O. 1977. Mechanisms of succession in natural communities and their role in community stability and organization. American Naturalist 111: 1119-1144.
Couteron, P., Seghieri, J. & Chadoeuf, J. 2003. A test for spa-tial relationships between neighbouring plants in plots of heterogeneous plant density. Journal of Vegetation Science 14: 163-177.
Cressie, N.A.C. 1991. Statistics for spatial data. Wiley, New York, NY, US.
Dale, M.R.T. 1999. Spatial pattern analysis in plant ecology. Cambridge University Press, Cambridge, UK.
Dale, M.R.T., Dixon, P., Fortin, M.-J., Legendre, P., Myers, D.E. & Rosenberg, M.S. 2002. Conceptual and mathe-matical relationships among methods for spatial analysis. Ecography 25: 558-577.
del Moral, R. 1993. Mechanisms of primary succession on volcanoes: A view from Mount St Helens. In: Miles, J. & Walton, D. (eds.) Primary succession on land, pp. 79-100.Blackwell, Oxford. UK.
Diggle, P.J. 2003. Statistical analysis of spatial point pattern. Academic Press. London, UK.
Dixon, P.M. 2002. Ripley’s K function. In: El-Shaaraui, A.H. & Piergorsch, W.W. (eds.) The encyclopedia of environme-trics, pp. 1796-1803. Wiley, New York, NY, US.
Fowler, N.L. 1988. What is a safe site? Neighbor, litter, germi-nation date, and patch effects. Ecology 69: 947–961.
Freeman, E.A. & Ford, E.D. 2002. Effects of data quality on analysis of ecological pattern using the statistical function. Ecology 83: 35-46.
Fuller, R.N. & del Moral, R. 2003. The role of refugia and dispersal in primary succession on Mount St. Helens, Washington. Journal of Vegetation Science 14: 637-644.
Garner, W. & Steinberger, Y. 1989. A proposed mechanism for the formation of ‘Fertile Islands’ in the desert ecosystem. Journal of Arid Environments 16: 257-262.
Geißelbrecht-Taferner, L., Geißelbrecht, J. & Mucina, L. 1997. Fine-scale spatial population patterns and mobility of winter-annual herbs in a dry grassland. Journal of Vege-tation Science 8: 209-216.
Goreaud, F. & Pélissier, R. 2003. Avoiding misinterpretation of biotic interactions with the intertype K12-funktion: population independence vs. random labelling hypotheses. Journal of Vegetation Science 14: 681-692.
Grubb, P.J. 1985. Plant populations and vegetation in relation to habitat, disturbance and competition. In: White, J. (ed.) The population structure of vegetation. Handbook of Vegetation Science, Vol. 3, pp. 595-621. Junk, Dordrecht, NL.
Holmgren, M., Scheffer, M. & Huston, M.A. 1997. The inter-play of facilitation and competition in plant communities. Ecology 78: 1966-1975.
Juhász-Nagy, P. & Podani, J. 1983. Information theory methods for the study of spatial processes and succession. Vegetatio 51: 129-140.
Kenkel, N.C. 1988. Pattern of self-thinning in jack pine: testing the random mortality hypothesis. Ecology 69: 1017-1024.
Lepš, J. 1990. Can underlying mechanisms be deduced from observed patterns. In: Krahulec, F., Agnew, A.D.Q., Ag-new, S. & Willems, J.H (eds.) Spatial processes in plant communities, pp. 1-11. Academia, Praha and SPB, The Hague, NL.
Lotwick, H.W. & Silverman, B.W. 1982. Methods for analysing spatial processes of several types of points. Journal of the Royal Statistical Society Series B 44: 406-413.
Malkinson, D., Kadmon, R. & Cohen, D. 2003. Pattern analysis in successional communities – an approach for studying shifts in ecological interactions. Journal of Vegetation Science 14: 213-222.
Marshall, J.K. 1967. Corynephorus canescens (L.) P.Beauv. Journal of Ecology 55: 207-220.
McDonald, R.I., Peet, R.K. & Urban, D.L. 2003. Spatial pat-tern of Quercus regeneration limitation and Acer rubrum invasion in a Piedmont forest. Journal of Vegetation Science 14: 441-450.
Muller, C.H. 1953. The association of desert annuals with shrubs. American Journal of Botany 40: 53–60.
Moeur, M. 1997. Spatial models of competition and gap dynamics in old-growth Tsuga heterophylla/Thuja plicata forests. Forest Ecology and Management 94: 175-186.
Paine, R.T. 1980. Food-webs: linkage, interaction, strength and community infrastructure. Journal of Animal Ecology 49: 667-685.
Pélissier, R. & Goreaud, F. 2001. A practical approach to the study of spatial structure in simple cases of heterogeneous vegetation. Journal of Vegetation Science 12: 99-108.

276 Felinks, B. & Wiegand, T.
Perry, J.N., Liebhold, A.M., Rosenberg, M.S., Dungan, J., Miriti, M., Jakomulska, A. & Citron-Pousty, S. 2002. Il-lustrations and guidelines for selecting statistical methods for quantifying spatial pattern in ecological data. Ecography 25: 578-600.
Peterson, C.J. & Squiers, E.R. 1995. An unexpected change in spatial pattern across 10 years in an aspen-white-pine forest. Journal of Ecology 83: 847-855.
Prach, K. 1987. Succession of vegetation on dumps from strip coal mining, N.W. Bohemia, Czechoslovakia. Folia Geobotanica and Phytotaxonomia 22: 339-354.
Prentice, I.C. & Werger, M.J.A. 1985. Clump spacing in a desert dwarf shrub community. Vegetatio 63: 133-139.
Pugnaire, F.I., Haase, P., Puigdefábregas, J., Cueto, M., Incoll, L.D. & Clark, S.C. 1996. Facilitation and succession under the canopy of a leguminous shrub in a semi-arid environ-ment in south-east Spain. Oikos 76: 455-464.
Ripley, B.D. 1976. The second-order analysis of stationary point processes. Journal of Applied Probability 13: 255-266.
Ripley, B.D. 1977. Modelling spatial patterns. Journal of the Royal Statistical Society. Series B (Methodological) 39: 172-212.
Ripley, B.D. 1981. Spatial statistics. Wiley, New York, NY, US.Roughgarden, J. & Diamond, J. 1986. Overview: The role of
species interaction in community ecology. In: Diamond, J. & Case, T.J. (eds.) Community ecology, pp. 333-343. Harper & Row, New York, NY, US.
Salonen, V. 1992. Effects of artificial plant cover on plant colonization of a bare peat surface. Journal of Vegetation Science 3: 109-112.
Salonen, V., Penttinen, A. & Särkkä, A. 1992. Plant colonization of a bare peat surface: population changes and spatial pat-terns. Journal of Vegetation Science 3: 113-118.
Schulz, F. & Wiegleb, G. 2000. Development options of natural habitats in a post-mining landscape. Land degradation and development 11: 99-110.
Schütz, W. 2000. The importance of seed regeneration strategies for the persistence of species in the changing landscape of Central Europe. Zeitschrift für Ökologie und Naturschutz 9: 73-83.
Sterner, R.W., Ribic, C.A. & Schatz, G.E. 1986. Testing for life historical changes in spatial patterns of four tropical tree species. Journal of Ecology 74: 621-633.
Stoyan, D. & Penttinen, A. 2000. Recent applications of point process methods in forestry statistics. Statistical Science 15: 61-78
Stoyan, D. & Stoyan, H. 1994. Fractals, random shapes and point fields. Methods of geometrical statistics. Wiley, London, UK.
Symonides, E. 1985. Population structure of psammophyte vegetation. Tüxenia 5: 259-271.
Tilman, D. 1985. The resource-ratio hypothesis of plant suc-cession. American Naturalist 125: 827-852.
Tirado, R. & Pugnaire, F.I. 2003. Shrub spatial aggregation and consequences for reproductive success. Oecologia 136: 296-301.
Tischew, S. (ed.) 2004. Renaturierung nach dem Braunkohle-abbau. Teubner, Stuttgart, Leipzig, Wiesbaden, DE.
Upton, G. & Fingleton, B. 1985. Spatial data analysis by
example. Volume 1: Point pattern and quantitative data. Wiley, London, UK.
van der Valk, A.G. 1992. Establishment, colonization and per-sistence. In: Glenn-Lewin, D.C., Peet, R.K. & Veblen, T.T. (eds.) Plant succession. Theory and prediction, Population and community biology, Series 11, pp. 60-102. Chapman & Hall, London, UK.
Watt, A.S. 1947. Pattern and process in the plant community. Journal of Ecology 35: 1-22.
Weiher, E. & Keddy, P.A. 1995. Assembly rules, null models, and trait dispersion: new questions from old patterns. Oikos 74: 159-164.
Wiegand, T. & Moloney, K.A. 2004. Rings, circles, and null-models for point pattern analysis in ecology. Oikos 104: 209-229.
Wiegand, T., Moloney, K.A. & Milton, S.J. 1998. Population dynamics, disturbance, and pattern evolution: Identifying the fundamental scales of organization in a model ecosys-tem. American Naturalist 152: 321-337.
Wiegand, T. Kissling, W.D., Cipriotti, P.A. & Aguiar, M.R. 2006. Extending point pattern analysis to objects of finite size and irregular shape. Journal of Ecology 94: 825-837.
Wiegand, T., Gunatilleke, C.V.S., Gunatilleke, I.A.U.N. & Okuda, T. 2007a. Analyzing the spatial structure of a Sri Lankan tree species with multiple scales of clustering. Ecology 88: 3088-3102.
Wiegand, T., Gunatilleke, C.V.S. & Gunatilleke I.A.U.N. 2007b. Species associations in a heterogeneous Sri Lankan Dip-terocarp forest. The American Naturalist 170: E77–E95.
Wiegleb, G. & Felinks, B. 2001. Primary succession in post-mining landscapes of Lower Lusatia – chance or necessity. Ecological Engineering 17: 199-217.
Wisskirchen, H. & Haeupler, H. 1998. Standardliste der Farn- und Blütenpflanzen Deutschlands. Ulmer, Stuttgart, DE.
Woodall, C.W. & Graham, J.M. 2004. A technique for con-ducting point pattern analysis of cluster plot stem-maps. Forest Ecology and Management 198: 31-37.
Yarranton, G.A. & Morrison, R.G. 1974. Spatial dynamics of a primary succession: nucleation. Journal of Ecology 62: 417-428.
Zhang, W. & Skarpe, C. 1995. Small scale species dynamics in semi-arid steppe vegetation in Inner Mongolia. Journal of Vegetation Science 6: 583-592.
Zobel, K., Zobel, M. & Peet, R.K. 1993. Change in pattern diversity during secondary succession in Estonian forests. Journal of Vegetation Science 4: 489-498.
Zobel, M., Suurkask, M., Rosén, E. & Pärtel, M. 1996. The dynamics of species richness in an experimentally resto-red calcareous grassland. Journal of Vegetation Science 7: 203-210.
Received 14 July 2006;Accepted 24 July 2007;
Co-ordinating Editor: P.M. Dixon.
For App. 1, see below (online version)also available at JVS/AVS Electronic Archives;
www.opuluspress.se/