evolucija arhitekture genoma - bio.bg.ac.rs · veliki deo genoma čine mobilni genetički elementi...
TRANSCRIPT

Evolucija arhitekture genoma

Brzina neutralne supstitucije
Stopa neutralne supstitucije jednaka je stopi mutacije (v).
Populacija veličine Ne → 2Ne alela (kopija gena)
Ne2
1- ista verovatnoća fiksacije za bilo koji alel
Nev2 - broj novih alela u populaciji
- verovatnoća fiksacije mutiranog alela po generacijivNe
Nev 2
12
Stopa supstitucije ne zavisi od N (Ne): -U velikoj populaciji više realizovanih mutacija, ali manja verovatnoća fiksacije driftom-U maloj populaciji manje realizovanih mutacija, ali veća verovatnoća fiksacije driftom

Skoro neutralne mutacije
Pokazalo se i da neke nesinonimne zamene nukleotida (očekivano selektivno prepoznatljive) imaju sličnu stopu supstitucije kao neutralne.
Ohta (1972, 1977) – matematički model:
mutacije se ponašaju kao selektivno neutralne ako
Nes
2
1
U velikim populacijama efekat drifta je mali, mali broj mutacija je efektivno neutralan.U malim populacijama efekat drifta je veliki, veći broj mutacija je efektivno neutralan.

Pregled veličine genoma
virusi prokarioti eukarioti
Virusi:-od 1- 4 Kb sa 2 gena (cirkovirusi) -do 1.2 Mb sa 1018 gena (mimivirus) i 1200 gena (megavirus)
Najmanji bakterijski genomi:-Mycoplasma genitalium (slobodno živeća) – 482 gena-Hodgkinia cicadicola (obligatni parazit) – 169 gena

Prokarioti: 0.5 – 9Mb do oko 8000 gena
Eukarioti: Više od 100Mb Obično više od 13000 gena (višećelijski)
Količina energije po genu – Ključna za veličinu genoma Mitohondrije neophodne za “finansiranje” eukariotskih genoma
Mitohondrije snabdevaju ćelije 105 – 106 puta većom energijom po genu u odnosu na prokariote
“Osnovna razlika između prokariota i eukariota nije samo u jedru već i u ekstremnoj polarizaciji eukariotskih ćelija – sa džinovskim jedarnim genomima koje energetski podržavaju mnogobrojni mali mitohondrijski genomi“ (Lane, 2011).
Pregled veličine genoma
prokarioti eukarioti

Razlike u organizaciji genoma Prokarioti (i virusi):
80% do 95% genoma kodirajuće
Eukarioti: Genomi veći i 100 000 puta, a samo 100 puta više kodirajućih nizova
Veliki deo genoma čine mobilni genetički elementi (3 – 53% genoma životinja, kod biljaka i do 80%)
Introni:
Jednako učešće introna i egzona u genima u genomima oko 100Mb
Genomi preko 25000Mb – 95% sekvence u genima čine introni.
Intergenska DNK: Genomi manji od 1 Mb – 20% intergenske DNK
Genomi veći od 10 Mb – i preko 80% intergenske DNK
Broj gena: Kod prokariota – 1000 gena po Mb
Kod eukariota – oko 12 gena po Mb
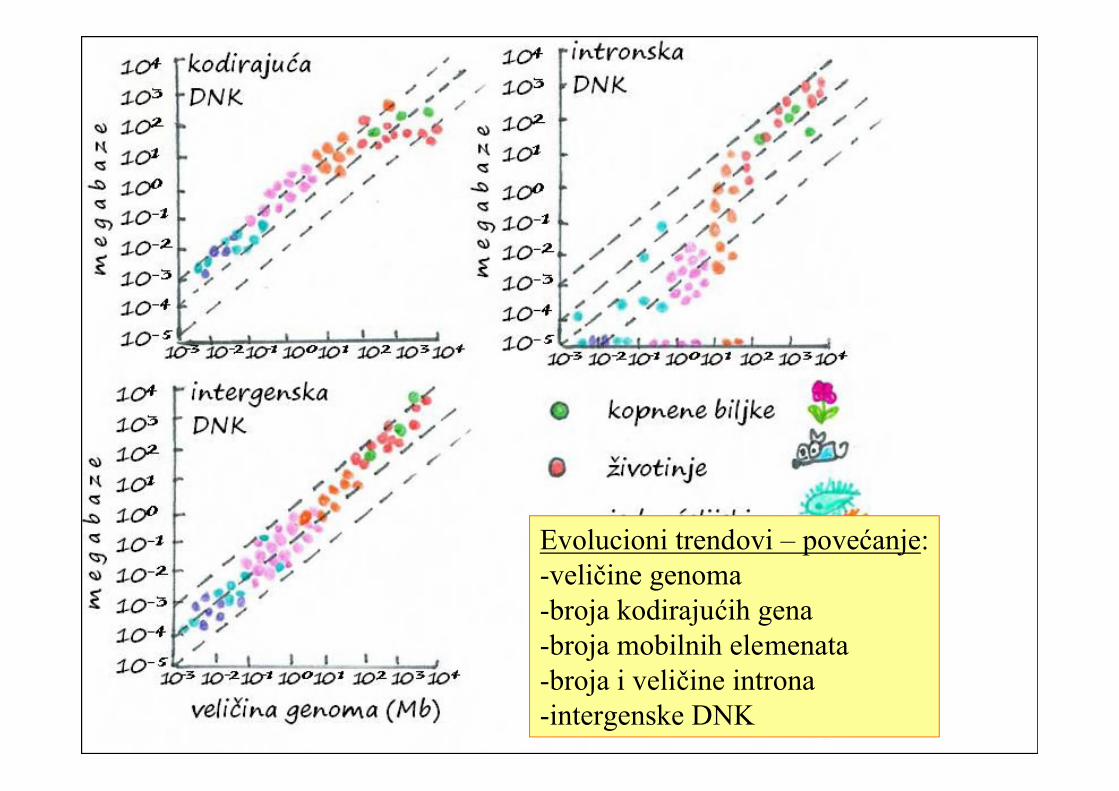
Evolucioni trendovi – povećanje:-veličine genoma-broja kodirajućih gena-broja mobilnih elemenata-broja i veličine introna-intergenske DNK

Koji deo genoma nam govori o fenotipskoj složenosti?
“G paradoks” – paradoks broja gena (koji kodiraju proteine)
Korelacija između biološke složenosti i količine ncDNK!

Evolucioni mehanizmi u oblikovanju genoma
Veliki genomi su evoluirali u malim populacijama!
Smanjenje populacije → povećanje verovatnoće akumulacije mutacija sa štetnim efektima (genetički drift)
Veći organizmi → manje populacije Prosečna veličina prokariotske populacije 1023 → mali genomi (efikasna
purifikujuća selekcija), mala varijansa u veličini genoma.
Nes
2
1
dobitak mt veće ćelije (sporija deoba, manje pop) višećeličnost
male pop.
neutralne mutacije – dodaci genomu (drift)
mala efikasnost purifikujuće selekcijeakumulacija
mutacija(viškova u genomu)

Uloga selekcije u oblikovanju genoma
Genska amplifikacija i adaptivna mutageneza Kod bakterija (LacZ- u LacZ+) – pojava adaptivne mutacije
češće nego što se očekuje po stopi mutacija - usled amplifikacije gena
Kod eukariota – oblici adaptivne mutageneze inicirani stresom i mobilnim elementima “programi” stresom indukovane mutageneze
uloga mobilnih elemenata u evolvabilnosti

Mobilni genetički elementi kod eukariota

Efekti ME
Direktni – ugrađivanjem u egzone, introne, promotore, pojačavače → menjaju okvir čitanja, nivo ekspresije gena, alternativno iskrajanje, itd. Npr. kod ljudi – više od 1000 gena ima ME u promotorima Npr. kod nekih primata u promotor gena za alfa-amilazu ubačen
ERV → promena ekspresije – iz pankreasa u pljuvačne žlezde (hrana bogata skrobom)
ME sa sopstvenim regulatornim elementima → promena obrasca ekspresije gena domaćina
Efekat ME na drugim nivoima regulacije: Kod ljudi 12% miRNK potiče od ME → kontrola ekspresije na
hiljade gena SINE nizovi – vezuju se za kompleks RNK polimeraze II ili sa
proteinskom kinazom R (PKR), itd.

Pasivni – nehomologne rekombinacije → hromozomski rearanžmani i duplikacije/delecije gena Npr. kod Anthropoidea Alu-Alu rekombinacija → duplikacija gena za
hormon rasta (između 5 i 8 kopija GH)
Npr. kod Cercopithecoidea Alu-Alu rekombinacija (pre 40 mil. god)→duplikacija opsinskih (OPN) gena → trihromatsko viđenje
ME česti oko gena koji brzo evoluiraju i čija uloga u metabolizmu zahteva fleksibilnost (odgovori na životnu sredinu – imunski odgovori ili signalni putevi)
“genske fabrike” – regioni bogati ME → nejednaki krosingover → veći broj multigenskih familija
Konzervisani regioni – malo ME Npr. Hox geni kod kičmenjaka (suprotno od Hox jata beskičmenjaka)
Efekti ME

Evoluciona uloga - evolvabilnost
“Živi fosili” imaju malo ME Latimeria (400 mil.god) i Sphenodon (220 mil.god. – 0.11% SINE i 2.59%
LINE elemenata u genomu)
Neurospora crassa – RIP mehanizam zaštite od ME i svake nove genske kopije
Većinska adaptivna strategija je, ipak, ne eliminacija ME, većnjihova regulacija → povećavaju evolucioni potencijal!!!

Mehanizmi kontrole
Hipoteza “odbrane domaćina” – primarna funkcija mehanizama epigenetičke regulacije ekspresije gena bila je odbrana domaćina od ME
DNK metilacija, modifikacija histona, RNK interferencija ME su preferencijalno metilovani u genomima
Metilacija ME (CpG) – regulacija aktivnosti susednih gena Npr. kod Anthropoidea – metilacija Alu elementa → regulacija CCRK gena
(uloga u odlaganju apoptoze)
Viši stepen metilacije u neokorteksu ljudi nego šimpanzi → veća aktivnost CCRK → više neurona

Ugrađivanje u gametsku liniju ćelija
Transgeneracijski mutageni efekti → evolvabilnost populacije
Kod sisara globalna demetilacija u ranim gametima
demetilacija u fazi preimplantacione embriogeneze
Transpozicija ME, aktivnost reverzne transkriptaze (L1) i retrotranskripcija aktivnih gena → retrokopije
gena

Retrogeni Reverznom transkripcijom najčešće nastaju “obrađeni pseudogeni”
(retropseudogeni) – odsustvo introna, ostaci poli(A) repića, veliki broj mutacija i odsustvo cis-regulatora.
Neki postaju aktivni geni:
Vretenasti plod paradajza: Retrotranspozon kopirao deo genoma domaćina (25Kb) sa kodirajućom
oblašću IQD12 gena (učestvuje u oblikovanju ploda)
Ugrađivanje u drugi hromozom u blizinu tuđih regulatornih elemenata →SUN retrogen
SUN aktivan tokom ranijih faza razvića ploda
U ljudskom genomu – 163 retrogena 25 retrogena su “siročići” – roditeljski geni izgubljeni

Retrogeni
“Izvoz” gena sa X na autozome kod sisara: GLUD2 (glutamat dehidrogenaza 2) – na 10. hromozomu
Nastao od GLUD1 (na X hromozomu) pre 18 – 25 mil.god.
2 nove mutacije u našoj evolucionoj liniji → povećana enzimska aktivnost → povećana aktivnost neurona
PGK 2 (fosfogliceratna kinaza) – na autozomu – eksprimira se u testisima tokom poslednjih stupnjeva spermatogeneze → korišćenje fruktoze iz semene tečnosti
Nastao od PGK1 (na X hromozomu) – eksprimira se u telesnim ćelijama

Molekulska domestikacija ME
Uloga u nastanku novih gena domaćina Npr. u proteinu PTPN1 (proteinska tirozinska fosfataza) – fragment
poreklom od L3 elementa
Enzim telomeraza – reverzna transkriptaza (kod organizama bez telomeraze, funkciju preuzimaju dva retrotranspozona)
Evolucija placente kod Eutheria i Metatheria
Peg10 gen – izveden iz gag gena jednog retrotranspozona
Peg11 gen – izveden iz gag gena drugog retrotranspozona (kod Eutheria – održavanje embrionskih kapilara u kasnijoj trudnoći)
sincicin1 i sincicin2 - izvedeni iz env gena ERV (sinteza glukoproteina)
MER20 transpozoni – sadrže cis elemente, direktno vezuju transkripcione faktore Preoblikovanje regulatorne mreže ekspresije gena u endometrijumu
Povećana ekspresija prolaktina (LTH)

Molekulska domestikacija ME – V(D)J
RSS – kratki invertovani ponovci – analogni terminalnim invertovanim ponovcima DNK transpozonaRAG1 i RAG2 – imobilizovane autonomne verzije Transcrib familije DNK transpozona

Uloga ME u specijaciji
Mutageni efekti povećavaju plastičnost genoma (genetičku varijabilnost)
Stres pojačava aktivnost ME – ili ih direktno aktivira ili inhibira mehanizme zaštite → adaptivna radijacija
Korelacija između broja vrsta i aktivnosti ME u evol. linijama Npr. ljiljci (adaptivna radijacija počela pre 55 – 44 mil.god) – veliki broj i
aktivnost transpozona (retro – i DNK)
Npr. glodari (1814 vrsta) – brojni i aktivni retro-ME (ERV)
Ribe Cichlidae iz jezera Afrike (oko 3000 vrsta)
Pre 5 mil.god. nastalo 1000-2000 vrsta u jezerima Malavi, Tanganjika i Viktorija
Veliki morfološki diverzitet, a mali genetički
Analiza SINE → višestruki ciklusi insercija SINE praćeni specijacijama (drastične fluktuacije nivoa vode)

Periodi intenzivne aktivnosti ME praćeni specijacijom DNK transpozoni – u vreme nastanka primata (pre 80 – 65 mil.god)
74000 do 98300 fiksiranih DNK transpozona u genomu čoveka potiče iz tog perioda
Pre 40 mil.god. prestaju aktivnosti DNK transpozona
Alu elementi (SINE) karakteristični za primate
AluJ pre oko 65 mil.god. (vreme razdvajanja polumajmuna i majmuna)
AluS ekspanzija pre oko 45 mil.god (vreme razdvajanja majmuna novog i starog sveta)
AluY ekspanzija pre oko 30 mil.god (vreme razdvajanja majmuna starogsveta i hominoidea)
Kod čoveka – broj Alu 3.4 puta veći od šimpanze
Najviše Alu ugrađeno pre oko 2 mil.god (pojava prvih Homo)
Uloga ME u specijaciji primata