evaluacin de la abundancia del calamar gigante … · de lm más pequeña en el mes de julio. el...
TRANSCRIPT

i
LA PESQUERÍA DEL CALAMAR GIGANTE (Dosidicus gigas) EN EL GOLFO DE CALIFORNIA. RECOMENDACIONES DE INVESTIGACIÓN Y TÁCTICAS
DE REGULACIÓN
Autores:
Susana Martínez-Aguilar 1, Enrique Morales-Bojórquez1, Juan Gabriel Díaz-Uribe1, Ma. Del Carmen Suárez-Higuera1, Agustín Hernández-Herrera2.
ISBN 970-9892-01-0
1 Instituto Nacional de la Pesca. Centro Regional de Investigación Pesquera Unidad La Paz, Carretera a Pichilingue Km.1 s/n. CP 23020. La Paz, Baja California Sur, México. 2 Centro Interdisciplinario de Ciencias Marinas. Instituto Politécnico Nacional, Av. I.P.N. s/n, Colonia Palo de Santa Rita. Apartado Postal 592, CP 23000. La Paz, Baja California Sur, México.

DIRECTORIO
JAVIER BERNARDO USABIAGA ARROYO Secretario de Agricultura, Ganadería, Desarrollo
Rural, Pesca y Alimentación
COMISION NACIONAL DE ACUACULTURA Y PESCA
Ramón Corral Avila
Comisionado Nacional
Prisciliano Meléndrez Barrios Director General de Ordenamiento
Pesquero y Acuícola
Raúl Villaseñor Talavera Director de Normalización
INSTITUTO NACIONAL DE LA
PESCA
Guillermo Compeán Jiménez Director en Jefe del Instituto Nacional
de la Pesca
Luis Beléndez Moreno Director General de Investigación
Pesquera del Pacífico Norte
Claudia de la Garza Montaño Directora del Centro Regional de
Investigación Pesquera de La Paz, B.C.S.
ii

PREPARACION DE ESTE DOCUMENTO Este documento fue elaborado en el Centro Regional de Investigación Pesquera de La Paz, Baja California Sur y sometido a revisión y edición en las instalaciones de la CONAPESCA en Mazatlán, Sinaloa. La revisión editorial de este documento se efectuó en la Dirección General de Ordenamiento Pesquero y Acuícola de la CONAPESCA, y estuvo a cargo de Raúl Villaseñor y Omar C. Carrillo Sandoval. Las citas bibliográficas, así como la referencia de los autores en la cita de este documento, se presentan tal y como fueron proporcionadas por los autores.
CITA DE ESTE DOCUMENTO
Martínez-Aguilar , S., Morales-Bojórquez, E., Díaz-Uribe, J. G., Suárez-Higuera, M. C. y Hernández-Herrera, A., 2004. La Pesquería del Calamar Gigante (Dosidicus gigas) en el Golfo de California. Recomendaciones de Investigación y Tácticas de Regulación. Comisión Nacional de Acuacultura y Pesca, Instituto Nacional de la Pesca, Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación. México. 77 p.
ii

Contenido
PRÓLOGO ...........................................................................................................5
RESUMEN............................................................................................................6
INTRODUCCIÓN..................................................................................................7
ANTECEDENTES.................................................................................................9
La pesquería .................................................................................................................... 9
Evaluación de stock .................................................................................................... 12
Biología reproductiva ................................................................................................. 13
ÁREA DE ESTUDIO...........................................................................................14
MATERIAL Y MÉTODOS...................................................................................15
Fuente de datos............................................................................................................ 15
Modelos .......................................................................................................................... 16 Reclutamiento y escape proporcional..................................................................... 16 Cambios del coeficiente de capturabilidad ( ), en función de la longitud........ 17 qCambios relativos de con el tiempo ..................................................................... 18 qAbundancia de la población ..................................................................................... 19
Ciclo de reproducción ................................................................................................ 19
Definición de grupos de longitud de manto ......................................................... 21
RESULTADOS ...................................................................................................21
Reclutamiento y escape proporcional ................................................................... 22
Coeficiente de capturabilidad ( )............................................................................ 22 q
Ciclo de reproducción ................................................................................................ 23
DISCUSIÓN Y CONCLUSIONES.......................................................................26
iii

RECOMENDACIONES DE MANEJO ................................................................32
RECOMENDACIONES DE INVESTIGACIÓN....................................................33
AGRADECIMIENTOS ........................................................................................33
LITERATURA CITADA ......................................................................................35
LISTA DE FIGURAS. .........................................................................................39
FIGURAS............................................................................................................41
TABLAS .............................................................................................................62
iv

PRÓLOGO La pesquería de calamar gigante (Dosidicus gigas) en el Golfo de California inicia en
1974 y actualmente es capturado en las zonas de Santa Rosalía, B.C.S., Guaymas,
Son. y Mazatlán, Sin.. El área de pesca de Santa Rosalía, B.C.S., constituye el 70% del
volumen nacional de captura de esta especie en peso fresco. Por esta razón, también
tiene la flota ribereña más importante en cuanto al número de licencias que participan
de la pesca, con casi 1200 embarcaciones menores distribuidas desde Loreto hasta
Santa Rosalía, B.C.S.. Desde el punto de vista económico, el recurso ha representado
una importante entrada de divisas a la pesca artesanal; la población de Santa Rosalía,
B.C.S. ha reiniciado una recuperación económica por la oferta de empleos directos e
indirectos, debido a las altas abundancias del recurso, ya que el principal mercado es
de exportación, principalmente hacia Oriente, donde Corea es el consumidor más
importante. En cuanto al manejo, la principal dificultad para regular la pesquería radica
en los cambios anuales que presenta en abundancia y composición de tallas. La
abundancia de estos organismos es variable, pero su distribución cercana a la costa de
la península, hace al recurso disponible durante casi todo el año. En esta zona se
observan continuamente organismos tanto de tallas (longitud de manto) pequeñas como
grandes, debido a la entrada y salida a la zona de pesca de calamares provenientes de
diferentes épocas de reproducción. La composición de la estructura de tallas observada
en esta zona no ha sido reportada para Guaymas, Son.. Existen indicios de que la zona
de pesca de Santa Rosalía es un importante núcleo de reproducción para la especie,
diversos cruceros de investigación han colectado información sobre individuos de D.
gigas con tallas pequeñas. La producción de calamares de la zona de Santa Rosalía,
mantiene la pesquería en el área de Guaymas, Son. durante toda la temporada. Cuando
la captura en Guaymas se agota, los barcos camaroneros adaptados a la pesca del
calamar gigante viajan a los caladeros de Santa Rosalía, ocasionando algunos
conflictos con la flota ribereña de la región. De esta manera, la importancia del recurso
calamar gigante (D. gigas) en la zona de Santa Rosalía, B.C.S., es debido a que
existen las condiciones que favorecen su ciclo de vida y a que funciona como centro de
dispersión hacia otras zonas del Golfo de California, razón por la cual podría constituir
una unidad de manejo independiente de Guaymas, Son.

RESUMEN Para el análisis de la pesquería de calamar gigante (Dosidicus gigas) del Golfo de California se
realizaron muestreos quincenales en la zona de Santa Rosalía, B.C.S., durante el periodo
comprendido entre Mayo y Octubre del 2002. Se observó la tendencia de la distribución de tallas
y la relación peso longitud encontrada en las capturas comerciales fue , con un
factor de correlación de 0.94. El intervalo de longitud de manto (LM) de la captura comercial
estuvo entre los 30 y 90 cm. Del total de organismos obtenidos de las capturas comerciales el
44% eran machos y el 56% hembras, con una proporción de sexos variable durante el periodo
de muestreo. Los resultados de la estimación del reclutamiento mostraron una subestimación,
debido a que la captura por unidad de esfuerzo ( ) exhibió dos picos a lo largo de la
temporada. Este comportamiento no se había observado antes y todo indica que la pesquería
está sustentada por dos cohortes, por lo que la estimación del reclutamiento requiere de
consideraciones para poblaciones con cohortes múltiples. El rendimiento también fue analizado a
través de estimaciones quincenales del coeficiente de capturabilidad ( q ) en función de la LM,
con el método de proyección de matrices. El patrón promedio de q no muestra una tendencia
definida, pero el comportamiento sugiere que la captura en la primera mitad de la temporada de
pesca estuvo sustentada por una cohorte. La variación de en función de LM con el tiempo, fue
determinada con la tasa de cambio de la quincenal respecto al promedio del tiempo total
analizado. La tendencia fue muy variable en el periodo de estudio observándose a los calamares
de LM más pequeña en el mes de julio. El proceso de maduración sexual de las hembras de
calamar gigante, se examinó por métodos histológicos, para validar las fases de madurez
gonadal morfocromática. Las muestras fueron obtenidas quincenalmente en el área de Santa
Rosalía, B.C.S., durante el periodo comprendido entre Junio del 2001 a Noviembre del 2002. Los
distintos tipos de ovocitos observados en el tejido, fueron cuantificados, encontrándose una
composición diferente para cada una de las fases morfocromáticas. La proporción ovocitos, se
utilizó para determinar estadísticamente las fases de madurez gonadal aplicando un estimador
denominado índice de madurez (IM). El IM permitió diferenciar claramente las gónadas
reproductivamente inactivas (fases I, II y III.) de las activas (fases IV, V y VI). De acuerdo a los
resultados del IM, la mayoría de hembras de Dosidicus gigas son maduras entre los 70 y 75 cm
de LM, con dos probables picos de actividad reproductiva durante verano y otoño.
9.200003.0 LP =
CPUE
q
CPUE

Introducción
as de calamar y camarón, (Morales-
ojórquez et al. 2001d).
nua recuperación de
cuerdo con los patrones de explotación (Rosenberg et al., 1990).
En los últimos años, la abundancia del recurso calamar gigante (Dosidicus gigas) en el
Golfo de California (Figura 1), se ha incrementado. Caddy y Rodhouse (1998),
mencionan que la abundancia de algunos recursos de peces ha sido reducida por
sobrepesca y los calamares que son de vida corta y ecológicamente oportunistas, se
han incrementado debido a la reducción en la presión por depredación y la debilitada
competencia por alimento. La variación en la abundancia de calamares también ha sido
explicada por la variabilidad en los parámetros ambientales (Roberts, 1998; Robin y
Denis, 1999; Waluda et al., 1999, 2001; Agnew et al, 2000; Sakurai et al, 2000). En el
Golfo de California las variaciones interanuales de la captura (Figura 2), han sido
relacionadas con las fluctuaciones ambientales, migraciones extensivas del recurso y
con la interacción existente entre las pesquerí
B
La información biológica y pesquera de Dosidicus gigas en el Golfo de California es
todavía escasa. El problema con mayor atención ha sido la estimación de biomasa del
recurso (Hernández-Herrera et al. 1998, Nevárez-Martínez et al. 2000, Morales-
Bojórquez et al. 2001b). Estas estimaciones se han basado en el modelo de Rosenberg
et al. (1990). El modelo asume que la población explotada está compuesta por una
cohorte que se agota a través del tiempo (Leslie y Davis 1939, De Lury 1947, Ricker
1975), de tal forma, que resulta relativamente simple evaluar la abundancia poblacional
a partir de un estimado del tamaño inicial (reclutamiento). Sin embargo, el valor de la
pendiente asociada a la capturabilidad, no permanece constante (Hilborn y Walters
1992), en algunos casos el modelo lineal se cuelga, mostrando una curvatura en lugar
de describir una línea (Gould y Pollock 1997, Gould et al. 1997, Morales-Bojórquez y
Nevárez-Martínez 2002). En los modelos de extracción sucesiva, las medidas del
esfuerzo pesquero y la captura son fundamentales en la estimación del reclutamiento y
la capturabilidad, ya que con ellos se define la estrategia de explotación del recurso,
calculando el escape proporcional constante, que se asume es necesario para
mantener a la población dentro de límites que permitan su conti
a
7

En recientes análisis, los resultados indican la presencia de más de una cohorte en la
población (Morales-Bojórquez et al. 2001d). Esta situación ha creado dudas sobre la
dinámica del recurso y la forma de evaluarlo, cambiando la perspectiva de
administración del calamar, ya que se debe estimar con precisión la magnitud de cada
horte así como su contribución a la reproducción, lo cual influye sobre la tasa de
s (longitud de manto), lo que supone la
xistencia de distintas cohortes. Esta composición en la estructura de longitud de manto
ra
valuar la abundancia del calamar gigante (Dosidicus gigas) en el Golfo de California,
ciones del coeficiente de capturabilidad y el desarrollo gonádico.
co
reemplazo de la población.
El porcentaje más alto de la captura total en peso fresco obtenida en el Golfo de
California es aportado por la pesca realizada en la zona de Santa Rosalía, B.C.S.,
seguido por Guaymas, Son, y Mazatlán, Sin.. Desde el punto de vista económico, el
recurso ha representado una importante entrada de divisas a la pesca artesanal;
poblaciones como Santa Rosalía, B.C.S. han reiniciado una recuperación económica
por la oferta de empleos directos e indirectos, debido a las altas abundancias del
recurso, ya que el principal mercado es de exportación, principalmente hacia Oriente,
donde Corea es el consumidor más importante. Santa Rosalía concentra la más
importante flota ribereña, con casi las 1,200 embarcaciones menores (pangas) con
motor fuera de borda, distribuidas desde Loreto hasta Santa Rosalía, B.C.S. En cuanto
al manejo, la principal dificultad para regular la pesquería radica en los cambios anuales
que presenta en abundancia y estructura de tallas. En Santa Rosalía se observan
continuamente organismos de diferentes talla
e
no ha sido reportado para Guaymas, Sonora.
Por otro lado, la zona costera de Baja California Sur es considerada como área de
desove, debido a que se han registrado altas tasas de apareamiento, cuantificadas por
la presencia de espermatóforos adheridos a la membrana oral de las hembras (Sato,
1975, 1976; Markaida y Sosa-Nishizaki, 2001; Tafur et al. 2001). La importancia que
tiene la zona de Santa Rosalía, en cuanto a volumen de captura y su probable
contribución al reclutamiento, así como, las fuertes implicaciones de manejo que tiene la
existencia de más de una cohorte en la pesquería de calamar gigante, fue la base pa
e
analizando las varia
8

ANTECEDENTES
La pesquería
En el Golfo de California, los primeros estudios sobre calamar gigante se realizaron al
principio de la década de los 80’s por Ehrhardt et al. (1983) y Klett (1996). Se intentó
explicar la dinámica de la población a partir de datos de captura y esfuerzo, cruceros de
investigación e información biológica básica. Sin embrago, el recurso desapareció
repentinamente y la hipótesis que pareció más razonable es que el calentamiento de la
corriente de California (El Niño) fue la causa (Klett 1996). El recurso no fue estudiado de
nuevo hasta que en 1995 la pesquería resurgió (Morales-Bojórquez et al. 1997, 2001c).
El recurso ha mostrado variaciones importantes en disponibilidad, observando en la
mporada de 1997-1998 una nueva caída que se asoció con la presencia de otro
ticas de manejo del
curso son controlar el esfuerzo de pesca a través de la asignación de licencias o
ones y recomendaciones sobre análisis de datos de captura y esfuerzo, así
omo de la información obtenida por cruceros de investigación (Nevárez-Martínez et al.
el 15 de Marzo del 2004; segunda sección), menciona el uso de talla mínima,
te
evento El Niño (Morales-Bojórquez et al. 2001c).
El esquema de manejo se ha basado en mantener una biomasa del 40% al final de
cada temporada de pesca, la cual es calculada en el momento mismo que el
reclutamiento es observado durante los meses de mayo y junio (Morales-Bojórquez et
al. 1997, 2001b, 2001d; Hernández-Herrera et al. 1998). Así las tác
re
permisos de pesca (Nevárez-Martínez y Morales-Bojórquez 1997).
Adicionalmente, cuando no se obtiene información veraz sobre el tamaño del
reclutamiento a partir de datos de captura y esfuerzo, se utiliza un sistema de cuotas de
captura que permiten simultáneamente obtener información de la pesquería y plantear
recomendaciones de administración. El Instituto Nacional de la Pesca, basa sus
estimaci
c
2000).
Las medidas de regulación establecidas a través del Diario Oficial de la Federación,
publicado
9

que no es útil en esta pesquería. Incluso, el mismo documento no hace referencia a talla
alguna.
A diferencia de Sonora (en donde ha habido un ambiente de cooperación reflejado no
sólo en la aceptación de las propuestas de manejo, sino en el apoyo económico de los
industriales y Gobierno del Estado de Sonora a la investigación pesquera), en la parte
de Santa Rosalía, B.C.S. no existen comités, sectores gubernamentales o grupos de
productores que en colaboración con el Instituto Nacional de la Pesca conformen
rupos de trabajo que permitan analizar, discutir o sugerir las bases de manejo del
s barcos camaroneros se adaptan con sistemas de pesca para calamar
igante, y a su vez, cuando la temporada de camarón del Pacífico Mexicano inicia,
minuye (Morales-Bojórquez et al.
001d).
oras, con rendimientos promedios de 400 o 500 kilos de
anto en los meses de mayor abundancia (primavera). Los sistemas de luz de las
embarcaciones se les acoplan carretes manuales de poteras, o bien, la pesca es
g
recurso. Aunque existe buena disposición hacia compartir sus bases de datos tanto de
captura como sus principales indicadores económicos.
La elevada abundancia del recurso durante 1996 creó expectativas sobre la pesquería
(Hernández-Herrera et al. 1998), ya que representó una alternativa rentable que se
complementó bien con la pesquería del camarón del Pacífico Mexicano, prácticamente
cuando los rendimientos del camarón disminuyen hacia los meses de diciembre y
enero, lo
g
entonces la presión de pesca sobre el calamar dis
2
Las flotas que explotan el recurso son de dos tipos:
La flota de pangas caracterizada por embarcaciones con motor fuera de borda de 40 a
75 caballos de fuerza, y dos pescadores por panga. La actividad de la flota en la zona
de Santa Rosalía. B.C.S. inicia a partir de las 16:00 horas y puede concluir hasta las
2:00 horas del día siguiente, sin embargo el tiempo de pesca efectivo de cada
embarcación varía entre 4 y 5 h
m
pangas son variables, usan desde focos de automóvil o tipo “árbol de Navidad” hasta
focos caseros de pocos watss.
La flota de barcos camaroneros adaptados a la pesca de calamar gigante. A estas
10

manual con solo una línea y su potera correspondiente. También se les adaptan
sistemas de iluminación variables. Pueden ser linternas que tienen cuatro lámparas de
00 wats y 100 volts, con luminosidad de 6,840 lúmenes por linterna, o bien lámparas
centro del Golfo de California hasta Santa Rosalía, B.C.S. lo que
romueve el desplazamiento de los barcos de Sonora hacia las zonas de pesca de
o de mayor captura 160,000 t (1996), lo
ue coincidió con la caída en el precio del recurso en 1999 para Santa Rosalía, B.C.S. y
de captura y esfuerzo, los cuales no son de fácil acceso en la
1
de 2,000 wats y 200 volts, con luminosidad de 30,000 lúmenes.
Morales-Bojórquez et al. 1997, sugieren que las zonas de Santa Rosalía, B.C.S. y
Guaymas, Son., explotan “un stock” localizado en la parte central del Golfo de
California, el cual está conformado por una cohorte o generación dominante. Los
individuos de esa cohorte también han mostrado un movimiento entre ambas costas, lo
cual beneficia la pesca en las dos localidades. También se ha observado que el recurso
se concentra desde el
p
Santa Rosalía, B.C.S.
La pesquería tiene una variabilidad en las capturas que hasta el momento no se ha
podido explicar de manera concluyente. Al parecer no hay evidencias de
sobreexplotación y la disminución de 1998-1999 se asoció con la presencia de un
calentamiento de la corriente de California, donde incluso el recurso mostró un patrón
de movimiento que lo llevó de la parte centro norte del Golfo de California hacia el
Pacífico (Bahía Magdalena, BCS). En la temporada 1999-2000 se observaron
problemas de precio en el recurso, ya que se pagaba barato el kilo de calamar, así los
patrones de captura a través del tiempo siguieron las tendencias del mercado
internacional del calamar. En algunos años, aunque la abundancia fue alta, la captura
permaneció baja en comparación con el period
q
de Sonora en el 2000.
Desde el punto de vista de la evaluación del stock explotado, la pesquería cuenta con
modelos explícitos para la estimación de biomasa, los cuales se apoyan en modelos de
extracción sucesiva. Las predicciones de captura pueden hacerse puesto que la escala
de tiempo usada son periodos semanales o quincenales. El recurso capturado a lo largo
de la temporada depende del reclutamiento a la pesquería y de la eficiencia de las flotas
correspondientes (capturabilidad). Así, la evaluación del recurso depende de la
veracidad de los registros
11

zona de Santa Rosalía, B.C.S., debido principalmente a la existencia de varios sitios de
o.
ctores que
ralmente utilizado es el análisis de reducción de stock de Delury (Rosenberg et al.,
1990), , descrito en términos de la captura por unidad de esfuerzo, ,
por:
desembarco del recurs
Evaluación de stock
La mayoría de las especies de calamar tienen un ciclo de vida anual y su abundancia
presenta grandes variaciones de un año al siguiente. Para evaluar la población, se
asume que la pesquería es sostenida por una sola cohorte, proveniente de un pulso
máximo de reclutamiento. El número de animales sobrevivientes al final del año afectan
la pesquería del año siguiente sólo a través de la cantidad de los productos del desove.
La evaluación se orienta a determinar el número de reclutas al inicio de la temporada de
pesca, la tasa de explotación durante la temporada y el número de organismos
reprodu sobreviven hasta el final de la temporada de pesca. El modelo
gene
tt ANoN −= CPUE
totE
t t
en número de individuos, tE es el esfuerzo pesquero, tA es la captura acumulada
desde el inicio de la estación de pesca al tiempo ,t No es la abundancia inicial en
número de individuos (reclutamiento) al inicio de la temporada de pesca y q , es el
coeficiente de capturabilidad. Esta relación supone proporcionalidad entre la captura por
unidad de esfuerzo y la abundancia del recurso y que la capturabilidad es constante a
través del tiempo. Con esta función, Rosenberg et al. (1990), plantearon el escape
proporcional de reproductores como estrategia de manejo del calamar Illex argentinus
en las islas Falkland, con un nivel permisible de K = 40% y desde entonces el escape
proporcional tam ha consid rado en otras pesquerías de calamar en el mundo
(Beddington
t qAqNCCPUE −=
Donde, es la abundancia en número de individuos en el tiempo , es la captura
bién se e
et al., 1990, Hernández-Herrera et al. 1998). El escape proporcional es
efinido como es la tasa instantánea de mort
=
N t C
FeK −= , donde F alidad por pescad
definida por EqF ⋅= .
12

La principal limitación observada para la utilización del método de Delury, es cuando la
CPUE , se incrementa al final de la estación incrementando también la abundancia
estimada del stock, esto descarta, el supuesto de que cada tem de pesca
representa una cohorte separada. Además del incremento de CPUE , se ha observado
disminución de la longitud media de las capturas y picos
porada
durante la
mporada de pesca, que son evidencias del ingreso de más de una cohorte a la
(2001d
e la pesquería de calamar, no obstante, los valores de
anteniendo su consideración de constancia a través del tiempo, dentro de la
debido a las altas tasas de apareamiento que encontró en las hembras de calamar
de la CPUEte
pesquería (Agnew et al. 1998, Hatfield and Des Clers, 1998).
Para probar la existencia de más de una cohorte, Agnew et al. (1998) utilizó los valores
del coeficiente de capturabilidad y señala que grupos de valores de q , evaluados con el
método de Delury, corresponden a cohortes individuales. Para la pesquería de calamar
gigante (Dosidicus gigas) del Golfo de California, Morales-Bojórquez et al. ),
evaluaron el coeficiente de capturabilidad en función de la longitud del manto, para una
temporada de pesca a través del método determinístico de capturabilidad )(MDC ,
(Arreguín-Sánchez, 1996, Martínez-Aguilar, et al. 1997) observando grupos de valores
de q en diferentes intervalos de longitud de manto considerando que dichos grupos
corresponden a distintas cohortes. Los trabajos de Agnew et al. y Morales-Bojórquez et
al. op cit., aportan información muy importante, que tiene implicaciones fuertes en la
evaluación y manejo d q , siguen
m
temporada de pesca.
Biología reproductiva
El proceso de maduración en Dosidicus gigas ha sido estudiado en sus dos áreas
principales de distribución: en el Pacífico Sur (Nesis 1970; Tafur y Rabí 1997; Tafur et
al. 2001) y en el Pacífico Norte incluyendo el Golfo de California (Michel et al. 1986;
Markaida y Sosa-Nishisaki 2000). Nesis (1970), define la primera escala de madurez
específica para Dosidicus gigas y propone la presencia de tres grupos en función de la
distribución de los fotóforos como respuesta al aislamiento geográfico entre el grupo de
Sudamérica y los grupos de Centroamérica y México. Sato (1976) sugiere que las
costas de la península de Baja California pueden funcionar como áreas de desove
13

gigante. Michel et al. (1986) describen por primera vez las fases de desarrollo de los
gametos del calamar gigante, sin embargo no describen los cambios en la composición
e gametos conforme la gónada madura.
s maduras con respecto a los machos
aduros es muy baja a lo largo de todo el año.
o fácil definir los períodos reproductivos del calamar
arkaida y Sosa-Nishisaki 2000).
REA DE ESTUDIO
d
Según Ehrhardt et al. (1983, 1986) en una temporada, el ciclo reproductivo de
Dosidicus gigas, no está fijo, varía con las condiciones oceanográficas. En 1980
encontraron hembras maduras a lo largo de todo el año con tres picos de reproducción,
el más importante en Diciembre/Enero en el Noreste del Golfo de California (cerca de
Guaymas) y el talud continental al Oeste de la península de Baja California. Este pico
genera los individuos que se reclutan a la pesquería en Marzo/Abril y sostienen la
pesquería en Septiembre. Los otros dos picos ocurren en Mayo/Junio y en Septiembre
cerca de Santa Rosalía, observando reclutamientos a la pesquería en Septiembre y
Enero/Febrero respectivamente. En contraste, Markaida y Sosa-Nishisaki (2000) no
encontraron ningún pico reproductivo entre 1995-1997 y argumentan que la época de
reproducción podría extenderse todo el año. Además, plantean que Dosidicus gigas
utiliza la región central del Golfo de California como área de alimentación y no de
reproducción, ya que la proporción de hembra
m
Los análisis de madurez gonadal y ciclo reproductivo en Dosidicus gigas se han basado
primordialmente en las características morfocromáticas de las gónadas y para ello se
utilizan distintas escalas de madurez, sin que estas se hayan contrastado con los
cambios que ocurren a nivel de tejido. La disparidad de escalas es probablemente una
de las razones por las que no ha sid
(M
Á
La localidad de Santa Rosalía, B.C.S. se localiza en los 27o 20’ LN y los 112o 16’ de LO
(Figura 1). Se ubica a 552.5 Km al Norte de La Paz, B.C.S., por la carretera
transpeninsular con una elevación sobre el nivel medio del mar de 40 m, presenta un
14

clima BW(h’) que corresponde al tipo muy seco, muy cálido y cálido con una
mperatura media anual de 22.6o C.
ODOS
tos de
s ovarios para su posterior identificación. El estatolito y los parásitos se sumergieron
La Paz, Baja California Sur. La captura total por clase de LM y el esfuerzo de
esca se usaron para estimar el índice de abundancia relativo de la pesquería, en este
aso la .
te
MATERIAL Y MÉT
Fuente de datos.
Se realizaron diez muestreos en la captura comercial de la flota artesanal ubicada en
Santa Rosalía, B.C.S., con una periodicidad quincenal correspondientes al periodo del 6
de mayo al 22 de octubre del 2002. En cada muestra se procesaron aproximadamente
cien calamares, registrando para cada organismo: peso total del cuerpo, longitud y peso
del manto. De una pequeña fracción de la muestra (∼30%) se determinó el sexo y
estadio de madurez sexual, con base en escala reportada por Lipinski (1995), (Tabla 1).
Las gónadas fueron fijadas en solución de Davidson (Bell y Lighter 1988) para ser
transportadas y almacenadas hasta su procesamiento histológico. Además, se
colectaron estatolitos, para análisis de edad, picos para el estudio de relaciones tróficas
y estómagos para identificar hábitos alimenticios. Así mismo, se extrajeron parási
lo
en alcohol al 70%, y el estómago y pico fueron congelados para su preservación.
La captura total por clase de longitud de manto (LM), en número de calamares por cada
2 cm de clase de LM, fue obtenida de la combinación de las estadísticas diarias de
captura (en kilogramos), agrupada en periodos quincenales y la estructura de LM,
obtenida de los muestreos quincenales de las capturas comerciales. Los registros del
esfuerzo pesquero (noches de pesca), fueron proporcionados por la Subdelegación de
pesca de
p
CPUEc
15

Modelos
Reclutamiento y escape proporcional
Se utilizó el modelo de extracción propuesto por Rosenberg et al. (1990) y aplicado por
Morales-Bojórquez et al. (1997) en la pesquería de calamar gigante. El modelo tiene la
siguiente forma:
( ) ( )∑−
=
1t
i la captura acumulada en número de
dividuos para cada unidad de tiempo t. Para este análisis M = 0.202 por quincena
n lognormal del mismo (Hilborn y Walters, 1992).
on este enfoque, se analizó una hipótesis de error de observación (Hilborn y Walters,
1992; Polacheck et al., 1993). El error de observación fue simulado 2,000 veces de
acuerdo con el siguiente estimador:
−−−− −=0
15.00
i
Mti
Mtt eCqeqNCPUE
donde q representa el coeficiente de capturabilidad, N0 el reclutamiento en número de
individuos, M la mortalidad natural (constante), C
in
(Hernández-Herrera et al.,1998), que fue estimado con el método Silliman descrito por
Ricker (1975) y Morales-Bojórquez et al. (1997).
Dado que la información de esfuerzo estuvo incompleta se simularon valores de
esfuerzo, asumiendo una distribució
C
⎟⎟⎠
⎞⎜⎜⎝
⎛−
= 2
2δσ
δ
eff isim
Así el valor estimado de CPUE que
se utilizó para fines de parametrizar el modelo fue
l valor promedio de los datos simulados (Hilborn y Walters, 1992). Finalmente la
fue estimada con el d to observado de captura (en número de individuos) y el
esfuerzo simulado, de acuerdo con:
e
CPUE a
16

( ) ( )∑−
−−−−
=⎠⎝
−=1
15.0t
MtMt
i
i eCqeqN
ef
C
se en el análisis de la ca
manto se realizaron estimac
r Arreguín-Sánchez (1
scrita
como NAN ,⋅= donde es el vector del tamaño del stock en número al tiempo
N
tra
⎟⎟⎞
⎜⎜⎛
− 00
2
2i
iδσ
δ
Cambios del coeficiente de capturabilidad ( q ), en función de la longitud
Con ba ptura por unidad de esfuerzo por clase de longitud de
iones de q con el método determinístico de capturabilidad
(MDC), de la forma propuesta po 996). El modelo estima la
capturabilidad por clase de longitud ( ), con base en una matriz de transición de
tl 1, +
(quincenas). tl , y 1,tl son representados por la ( ). es la matriz de
nsición expresada por: SG = A
l
: tl tlN ,
t N + CPUE A
⋅ donde G es la matriz de probabilidad de
crecimiento y S es la matriz de supervivencia. Cada uno de los elementos de S son
obtenidos de )(, expexp , ttl EqMZtiS ⋅+−− == don ti, es la mortalidad total y de Z M es la
mortalidad natural para el i th grupo de longitud al tiempo t . La matriz de crecimiento,
G , es definida suponiendo que los organismos crecen de acuerdo al modelo de von-
Bertalanffy, las probabilidades son calculadas r Shepherd
(1987), (Morales-Bojórq 2001d). 1 se resuelve con el algoritmo de
mínimos cuadrados utilizando dos rutinas de búsqueda directa no lineal. La primera
lcula valores aproximados de
con el método descrito po
uez et al. , +tlN
ca A. Los valores de A son acepta os uan las
diferencias uadrado del valor obs y el calculado 1,
d c
al o de
do
c ervad 1, +tlN ~+tlN alcanzan su
valor mínimo Como G es conocid . a y SGA ⋅= , la segunda rutina calcula valores
s de valores son aceptados cuando las diferencias al e S y sus cuadrado daproximado
A y su nueva estimación A~ , son mínimas. Como los valores de , S M y E son
s tLq , fue despejada de )(, exp tEqMtiS ⋅+−= . Finalmente se estimó un valor
promedio de q por clase de longitud
conocido
tLq , .
17

Los parámetros de crecimiento del modelo de von Bertalanffy ( /quincena
ernández-Herrera (1998). La mortalidad natural
con el método de Silliman (Ricker 1975), fue considerada constante con un
Cambios relativos d
de la capturabilidad en función de la longitud del manto en el
expresados por la relación de la tasa anual de por clase de longitud
de la quincena al tiempo , , respecto al promedio de la temporada analizada
PUE , . De acuerdo con Arreguín-Sánchez (1996), la relación en su forma lineal es
xpresada como:
033.0=k
87=∞L cm) fueron tomados de H
estimada
valor de, 2.0=M por quincena.
e q con el tiempo
Los cambios relativos
tiempo, son CPUE
t tLCPUE ,
yLC
e
L CPUECPU[
y L,
⋅βαLn
Donde: tL,
+ = ]EtL,t L,
t L,
β , representa las diferencias relativas en el patrón de cambio de q con la
itud de la quincena t respecto a yLCPUE , ; tL,long α representa las diferencias relativas
en la abundancia de org presentes en la captura de la cena
. El signo de la pendiente y de la ordenada al origen
los grupos de lon itud que son más abundantes. Entonces si
anismos más pequeños quin
, respecto a yLCPUE , tL,β tL,α t
g tL,βindica es positivo y
tL,α negativo, los organismos grandes son más abundantes que los pequeños y
iceversa.
v
18

Ab
a mortalidad por pesca ( ), fue calculada de la expresión general (Ricker
1975, Gulland 1983). Para las estimaciones de abund ia en número de calamares,
fue determinada con la misma ecuación pero expresada para cada clase de LM y para
a
undancia de la población
F EqF ⋅=L
( tN )anc
Fcad quincena ( t ).
ttLtL EqF ⋅= ,,
a , se obtuvo de la siguiente expresión general: tNL
( )( ) tLMFtL
tLtL F
MFtL ,
,
,, .exp1
⋅⎟⎟⎠
⎞⎜⎜⎝
⎛
−⋅
+= +−
CN
onde: representa la captura en número de individuos por clase de LM, para cada
El peso promedio por clase de LM obtenido de la relación peso-
ngitud se utilizó para transformar la abundancia en número de calamares a biomasa en
n las características morfológicas y del
licular que rodea a los ovocitos, se definieron sus fases de desarrollo. Las
diferencias entre los estadios de madurez macroscópica asignados en el campo a partir
tLC ,D
tiempo t correspondiente.
lo
toneladas.
Ciclo de reproducción
En el laboratorio, de cada gónada se extrajo una muestra de tejido de 1 cc
aproximadamente. La muestra se deshidrató en alcohol etílico, se transparentó con
Hemo-De y se incluyó en parafina de punto de fusión alto. Para obtener las laminillas se
hicieron cortes transversales de 7 μm de espesor con un micrótomo de rotación manual
y fueron teñidas con la técnica hematoxilina-eosina (H&E). La lectura de cada laminilla
consistió en seleccionar un campo visual de 0.5 cm2 para contar los ovocitos en cada
una de sus fases de desarrollo. Con base e
epitelio fo
19

de las propiedades morfocromáticas de las gónadas, fueron evaluadas con la ecuación
descrita por Sauer y Lipinski (1990):
∑=
= 5
0ssn
IM ∑
=
×5
0
)(s
s sn
ador del nivel de desarrollo de los ovocitos en
s gónadas, entonces, cada estadio macroscópico tendrá un IM significativamente
el análisis de la proporción de hembras maduras por intervalo de
M. Para ello, se estimó la frecuencia acumulada de hembras maduras en cada
intervalo de talla, y se ajustó por el método de mínimos cuadrados a una curva logística
de la siguiente forma:
donde: IM es el índice de madurez, s es un número discreto entre 0 y 5 asignado a cada
nivel de desarrollo del ovocito y ns es el número de ovocitos contados en el estadio s.
Se aplicó un análisis de varianza de una vía y pruebas de Tukey para comparación
múltiple de factores. La hipótesis planteada para esta prueba fue: si los estadios de
madurez macroscópica son un buen indic
la
distinto al siguiente estadio. Además, el IM se utilizó para evaluar los cambios en el
nivel de maduración a través del tiempo.
La talla de primera madurez (Lm ) en la cual el 50% de las hembras son sexualmente
maduras, se obtuvo d
L
)(11
ibLai eP −+
=
donde: Pi es la frecuencia relativa acumulada de hembras maduras en el intervalo de
ngitud i, Li es la marca de clase del intervalo i en términos de LM, a y b son las
onstantes de regresión y cumplen con la condición a/b = Lm.
lo
c
20

Definición de grupos de longitud de manto
La división de la longitud de los calamares en grupos, fue usada como marco de
referencia, en la descripción de los diferentes procesos que involucra el estudio de este
recurso. Por esta razón, se estableció la división de la estructura de longitud del manto,
para machos y hembras mezclados en tres grupos: pequeños, medianos y grandes,
definiendo los intervalos de talla de13-34cm, 36-60 cm y >62-120 cm de LM, para cada
grupo respectivamente. Nigmatullin et al. (2001) proporcionaron la base para la
definición de los grupos. Los autores dividieron la estructura de longitud de manto por
separado de machos y hembras adultos de esta especie. Los machos pequeños los
definen entre los 13 y 26 cm de LM, los medianos entre los 24 y 42 cm de LM y los
grandes los >40-50 cm de LM. Para hembras pequeñas los organismos de 14-34 cm de
LM, las medianas de 28-60 cm de LM y las grandes entre los 55 y 65 a los 100 y 120
cm de LM. En este estudio la división fue el resultado de la integración de la estructura
e longitud de manto de machos y hembras, con intervalos de longitud del manto de los
lapamiento.
ructura de longit
el periodo en estudio. La
roporción macho:hembra se presenta en la Tabla 2. En el mes de septiembre se
bservó la mayor proporción de hembras por cada macho.
d
grupos sin sobre
RESULTADOS
El intervalo de LM en las muestras de las capturas comerciales estuvo entre los 30 cm y
los 90 cm de LM, (Figura 3). La estructura mostró una gran variabilidad en su
composición, observándose alternadamente presencia de organismos pequeños y
grandes. En el mes de Octubre se registraron los calamares de longitud de manto más
pequeña. Para extrapolar la est ud de las muestras a la captura total
(Figura 4) y obtener la captura por unidad de esfuerzo por clase de LM (CPUE), se
calculó la relación peso longitud: 91.200003.0 LP ⋅= con un factor de correlación de 0.94,
(Figura 5). De las muestras de las capturas comerciales el 44% eran machos y 56%
hembras con una proporción de sexos variable durante
p
o
21

Reclutamiento y escape proporcional
Los resultados mostraron una subestimación del reclutamiento 2,302,284 calamares
con error estándar ( ES ) de 1,118,363 (P< .375). El valor de la capturabilidad fue de
0.00015 con ES = 0.00014 (P< .375) y r2 = 0.21 (Figura 6). La tendencia de la captura
por unidad de esfuerzo a través del tiempo mostró una caída a partir de la quincena 4
(Figura 7), esto implicó que el aparente pico de reclutamiento se observara en Julio, es
decir, dos meses después del tiempo esperado en el cual el reclutamiento sucede.
demás se observó una mezcla de distintas tallas a lo largo de toda la temporada,
ión anual. La subestimación del reclutamiento y la falta
e ajuste del modelo, solo mostraron que la serie de tiempo tiene al menos dos picos de
apturabilidad
uestra el valor más alto a los 72 cm de LM. La desviación estándar es alta debido a
A
indicando el flujo continuo de organismos hacia la estructura de la población (Figura 3).
La variabilidad en tallas observada en el periodo de julio a octubre del 2002 es un
comportamiento muy extraño, al parecer los organismos pequeños (entre 30 y 36 cm de
longitud de manto) aparecen hasta octubre, mientras que en julio su presencia es
escasa, aunque suficiente para tener impacto sobre la CPUE . Sin embargo la
presencia de organismos grandes (mayores a 66 cm de LM ) que aparecieron en
agosto, puede ser que incorporaran una alta biomasa a la población que es difícil de
medir. Los cambios repentinos en la CPUE no son una tendencia común en recursos
con una sola cohorte o generac
d
reclutamiento (Julio y Octubre).
Coeficiente de capturabilidad ( q ).
La variación del valor promedio del coeficiente de capturabilidad por clase de LM se
presenta en la Figura 8. El promedio general fue de 0.0014, (d.s.=0.003) y los valores
mínimo y máximo observados fueron 0.0003 y 0.014 respectivamente. La c
m
las variaciones del coeficiente de capturabilidad con la LM y con el tiempo.
Las tendencias en los cambios relativos quincenales de la tasa de captura en función de
LM, respecto al promedio periodo analizado (6 de Mayo al 26 de Julio), presentaron una
gran variabilidad (Figura 9). En los meses de mayo y julio se capturaron calamares de
intervalos de longitud mediana (36-60 cm de LM), con predominio en la primera
22

quincena del mes de Mayo y la segunda de Julio. Con excepción de las quincenas
antes mencionadas, la presencia de calamares de LM mediana es escasa. En Julio la
abundancia de calamares pequeños y grandes en la captura fue alta, observándose las
longitudes más pequeñas (30-36 cm de LM) en la primera quincena. La Tabla 3
presenta las constante del modelo lineal ajustado a las tendencias de los cambios
relativos observados de la CPUE por clase de LM y el coeficiente de correlación. La
pendiente del modelo
s
β , indica el cambio relativo de capturabilidad en función de la LM
y α , la abundancia de los organismos más pequeños presentes en la captura. Los
alores duplicados de las constantes, con distinto signo, en la misma quincena, indica la v
presencia simultánea de calamares pequeños y grandes en las capturas.
El índice de capturabilidad β , en función de la cantidad de pesca mostró una tendencia
descendente con una disminución aproximada del esfuerzo pesquero del 35% entre el
29 de Mayo y 26 de Ju 10). Los valores negativos de lio (Figura β en la figura indican
ue la captura estuvo sustentada principalmente por calamares de longitudes
n base en las estimaciones de y el esfuerzo pesquero
jercido en cada quincena, el valor total de biomasa para el periodo analizado fue de
a 11).
a actividad ciclónica no permitió que la pesca de calamar se
alizara con regularidad, por lo que no hay registros en la segunda quincena de agosto
q
pequeñas.
La abundancia fue calculada co q
e
31, 244 toneladas (Figur
Ciclo de reproducción
El análisis histológico se hizo con las bases de datos obtenidas de Junio a Noviembre
del 2001 y de Mayo a Octubre del 2002. Durante estos período se colectaron un total de
428 organismos, de los cuales el 71% fueron hembras, el 27% machos y el 2%
indiferenciados (Tabla 4). En 2001 el muestreo se dirigió casi exclusivamente a las
hembras, mientras que en el 2002 se colectaron tanto hembras como machos. A finales
de la temporada 2001, l
re
y el mes de septiembre.
23

Con los cortes histológicos se identificaron cinco tipos de ovocitos en el tejido que
representan diferentes estadios de desarrollo, desde inmaduros hasta ovocitos a punto
de ser expulsados (Tabla 5). Los primeros dos estadios describieron no consecutivas,
en las que el ovocito no está disponible para la actividad reproductiva. Para el cálculo
del IM a estos dos estadios se les asignó S= 1 y 2 respectivamente. Los otros tres
stadios representaron etapas de maduración consecutivas en las que se consideró que
1 disminuyen al 60% y el resto se encuentran madurando,
rincipalmente en estadios 3 y 4. Cuando las gónadas alcanzan la fase IV de madurez,
a los organismos que se encontraron en fase de
producción activa de la inactiva. Por esta razón, el análisis temporal de la maduración
e
el ovocito estaba en preparación o disponible para la actividad reproductiva. A estos
estadios se les asignaron valores de S= 3, 4 y 5 respectivamente.
En términos generales, las categorías de maduración establecidas de acuerdo a las
propiedades morfocromáticas de las gónadas estuvieron correlacionadas con las
características histológicas del propio órgano. Se pudo observar que la composición de
los distintos tipos de ovocitos en la gónada cambia conforme el organismo madura
sexualmente (Tabla 6). En las gónadas de juveniles (Fase I), solamente se observaron
ovocitos 1, sin embargo, un 7% de los organismos clasificados como juveniles fueron
organismos que ya habían desovado o se encontraban en reposo, y por lo tanto no eran
propiamente juveniles. En la fase II, los ovocitos 2 y 3 empiezan a ser más abundantes
(~10% cada uno), sin embargo los ovocitos 1 siguen predominando (~75%). En la fase
III, los ovocitos
p
alrededor del 90% de los ovocitos se han desarrollado hasta los estadios 3 y 4 en una
proporción 0.4:1.
En la fase V, se observó la mayor abundancia de ovocitos 5 con un 18% de las células
contadas. El resto de los ovocitos se distribuyó homogéneamente en comparación con
los otros estadios de madurez. En la fase VI, se observaron dos casos, uno en donde el
tejido presentó restos de ovocitos maduros y tejido conjuntivo en abundancia y el otro
en el que más del 80 % de los ovocitos estuvieron en el estadio 1 ó 3. Aunque estos
cambios también se reflejaron en el IM (Figura 13) y en el análisis de varianza cuyo
valor de F fue significativo, la prueba de comparaciones múltiples de Tukey solo detectó
una diferencia significativa entre las fases III y IV (Tabla 7). Es decir, la composición de
ovocitos en las gónadas fue tan variable que la escala de madurez macroscópica solo
permitió identificar con confianza
re
24

y la determinación de la LM de primera madurez se trabajó con las lecturas histológicas
del tejido de la gónada.
Durante 2001, el promedio más alto del IM de las hembras se presentó en la primera
quincena de octubre, con un valor significativamente >2. En esa quincena más del 90%
de los ovocitos estaban desarrollados entre los tipos 3 y 5 con una predominancia de
los ovocitos 4 (~62%). En la segunda quincena de Junio y primera de Julio el índice
promedio es igual a 2, sin embargo fueron los periodos de mayor dispersión del IM . En
estas quincenas la predominancia de ovocitos 1 (~60-70%) está compensada por la
presencia de ovocitos 4 y 5 (~20-25%). Lo anterior indica una transición un
eriodo reproductivamente activo a uno inactivo. La primera quincena de agosto, el
entre
IM p
es similar al de las dos quincenas anteriores, sin embargo la composición de ovocitos
es diferente, en la que predominan los ovocitos 2.
En 2002, se observaron dos picos en los que el IM fue significativamente >2 (Figura
13). En la segunda quincena de mayo los ovocitos en el tejido fueron del 3 al 5 de
estadio de desarrollo comprendieron el 57% , en la que los de tipo 3 fueron los
predominantes (38%). En la segunda quincena de agosto los ovocitos 3 a 5
omprendieron el 73% de los ovocitos en el tejido, de los cuales, los ovocitos 4 fueron
de LM, principalmente entre 75 y 80 cm de
M. La hembra madura más pequeña que se registro fue de 33 cm de LM. El análisis
e frecuencia acumulada indica que Dosidicus gigas alcanza la talla de primera
adurez a los 72 cm de LM, (Figura 14b).
c
los más abundantes (31%). En los otros meses el IM fue <2 debido a la predominancia
de ovocitos 1 y a la ausencia propiamente de ovocitos 5.
Las tallas de las hembras en los muestreos oscilaron entre 33 y 91 cm de LM,
observándose la mayor frecuencia entre 50 y 80 cm (Figura 14a). Más del 90% de las
hembras adultas fueron mayores de 50 cm
L
d
m
25

DISCUSIÓN Y CONCLUSIONES
La captura comercial de calamar gigante (Dosidicus gigas) en el área de Santa Rosalía,
B.C.S. mostró gran variabilidad, con el pico más alto en el mes de Julio. En estudios
previos (Roberts 1998. Robin y Denis, 1999; Waluda et al., 1999 y 2001; Agnew et al,
2000; Sakurai et al, 2000, Morales-Bojórquez et al. 2001d), se han planteado diferentes
factores que influyen en la variación de la abundancia, sin embargo, en la variación de
la captura comercial en el Golfo de California, influyen también factores tales como los
desacuerdos entre la industria y los pescadores en el precio por kilo y la capacidad de
procesamiento que tienen las plantas, que en ocasiones, cuando el recurso es muy
abundante, limitan la cantidad de captura que reciben, provocando problemas en la
valuación del recurso pues esta depende primordialmente de los datos de captura y
Golfo de California. Sin embargo, este tipo de estudios se hacen
on métodos de marcaje y recaptura, que resultan muy costosos y a la fecha no existen
en la estructura de
eso y longitud de manto, es conveniente hacer el estimado con una periodicidad anual.
e
esfuerzo.
El calamar gigante es una especie migratoria y las variaciones de disponibilidad del
recurso dependen de este comportamiento, por lo que es importante describir el patrón
migratorio dentro del
c
reportes al respecto.
En las muestras obtenidas la proporción de sexos fue variable durante el periodo de
estudio, con una dominancia de hembras de 9:1 en el mes de Septiembre. Markaida y
Sosa-Nishizaki (2001), mencionan que la proporción sexual es casi igual durante la
estación de desove y en el pico de desove, las hembras maduras dominan entre un
70% y 80% del total. En este trabajo la proporción sexual observada fue considerada
como un elemento auxiliar en la determinación del pico de desove. En cuanto a la
relación peso-longitud, debido a las variaciones que se presentan
p
La relación peso-longitud en este estudio es una estimación anual.
Los muestreos quincenales del recurso calamar proporcionan información sobre la
estructura de longitud del manto y su variación en el tiempo y permiten observar el
periodo de reclutamiento y hacer el seguimiento de la cohorte o cohortes presentes en
la población. La estrategia de manejo establecida para esta especie se basa en un
26

modelo que asume la existencia de una sola cohorte anual, lo que hace importante, el
monitoreo de la estructura de longitud del manto en periodos cortos de tiempo y la
determinación del ciclo reproductivo. En este trabajo la estructura de longitud de manto
registrada, presentó una gran variabilidad, observándose en las frecuencias de longitud,
odas alternantes de organismos pequeños y grandes, que no permitieron, por este
parame
entras que en el
eriodo de agosto a octubre dominaron los organismos de talla pequeña, con una
dancia
undan
del
avés de cruceros stigación y otros métodos alternos de análisis de ,
m
medio, identificar la cohorte o cohortes presentes durante el periodo de estudio.
El patrón de cambio observado en los picos de abundancia relativa de la CPUE ,
sugieren la presencia de dos cohortes de calamar en la temporada 2002. La
trización del modelo de extracción sucesiva no fue estadísticamente exitosa, ya
que el valor de N0 estimado en julio fue insuficiente para soportar el incremento en
CPUE debido a la presencia de la cohorte de octubre (Figura 3), además del patrón
irregular observado de los organismos grandes dentro de la zona de estudio. Morales-
Bojórquez et al. (2001b) registraron la presencia de tres cohortes de calamar gigante en
el Golfo de California, sin embargo, para la zona de Guaymas la explotación del recurso
se sustenta en una cohorte principal (Hernández-Herrera et al. 1998). Los datos
mostraron la presencia de organismos medianos de mayo a julio, mi
p
mezcla de baja abun para tallas medianas y grandes (Figura 3).
Los cambios en la CPUE de la pesquería del calamar gigante parecen tener relación
con los cambios en la composición de tallas de la captura, pero no con la ab cia.
La pesquería muestra signos de hiperestabilidad, definida por la concentración de
esfuerzo en zonas donde el calamar es más abundante, de tal forma que la CPUE se
mantiene mientras que la abundancia declina (Hilborn y Walters 1992), sin embargo, los
cambios repentinos de abundancia por la entrada o salida de una nueva cohorte,
pueden provocar hiperdepleción, es decir, que la CPUE decline más rápido que la
abundancia (Hilborn y Walters 1992). La dinámica del recurso no permite por ahora,
conocer con precisión qué fenómeno se está presentando, pero el tama
reclutamiento y el tamaño de la población a diferentes tiempos es posible evaluarlos a
ño
de inve CPUEtr
como el método determinístico de capturabilidad ( MDC ) que se aplicó en este estudio.
27

Los cam epentinos en la estructura de la población puede tener relación con
cambios en las tasa de mortalidad y de la capturabilidad, el comportamiento de la flota
depende de la disponibilidad, de mercado y precio, y esta variación tiene su impacto
sobre la CPUE comercial. Al margen de esta observación, la población de calamar que
se pesca en Santa Rosalía parece tener dos cohortes que son explotadas,
comportamiento que no había sido observado y que dificulta la estimación de los
parámetros del modelo. La posibilidad de tener dos cohortes sugiere éxito
bios r
en la
upervivencia, aunque cabe la posibilidad de que sea el resultado de la mezcla de
de una cohorte principal en la porción de tiempo de la
mporada de pesca analizada con el MDC. Basson et al. 1996 y Agnew et al. (1998)
p
s
stocks diferentes, la cual es una hipótesis a comprobar. En cualquier caso, esto puede
implicar que la población sea más grande de lo que las estimaciones mostraron.
Aunque la estructura de tallas no permitió identificar claramente el número de cohortes
presentes en esta temporada de pesca, la presencia de picos de q indican el ingreso
de más de una cohorte a la pesquería. Sin embargo el grupo más conspicuo de valores
de q estuvo entre los 54 y 82 cm de LM con el valor más alto en los 72 cm de LM, lo
cual indico la presencia
te
señalaron que diferentes grupos del coeficiente de capturabilidad indican dos o más
cohortes en la pesquería.
Con el propósito de comprobar si el comportamiento observado en cuanto a estructura
de LM y coeficiente de capturabilidad en la temporada 2002, era un atrón repetido, se
analizó la temporada 2001 de la misma forma que la del 2002 (Figura 15). Además, se
determinó la estructura de LM total para cada una de las temporadas y se evalúo q por
clase de LM de la temporada 2001 (Figura 16). En ambos casos q mostró una gran
variabilidad respecto a la LM. No obstante, la tendencia general observada para los
organismos pequeños a medianos es que la capturabilidad decrece conforme se
incrementa la LM, y en los calamares medianos a grandes la capturabilidad se
incrementa conforme se incrementa la LM. El cambio en la tendencia está precisamente
en los organismos de LM mediana, los cuales son escasos en la estructura de la
captura comercial y son precisamente las tallas en donde las hembras empiezan a
madurar aunque no se trata propiamente de la talla crítica o talla de primera madurez,
que corresponde al 50% de hembras maduras. En la evaluación de calamares, la
28

suposición general es que la capturabilidad es constante para una estación de pesca
determinada, sin embargo, dado que se pescan durante gran parte de su ciclo de vida y
el comportamiento cambia conforme crece de juvenil a adulto era de esperarse
encontrar cambios en capturabilidad entre organismos de tallas pequeñas y grandes,
pero el flujo de calamares hacia adentro o fuera del área produce una gran variabilidad
en los valores de q . En el área de Santa Rosalía este comportamiento es diferente a lo
ue hasta ahora se conoce, y seguramente está relacionado con la dinámica del
diferencial de
alamares por L por una dominancia en la población de calamares de LM grande.
q
recurso. Esto merece más atención por las fuertes implicaciones que tiene el manejo del
recurso, ya que esta zona es la que mayor volumen aporta a la captura total.
Los cambios relativos de la capturabilidad en función de la LM, con el tiempo
presentaron una gran variabilidad en su tendencia. En los gráficos, los valores negativos
denotan cantidades bajas de calamares en las capturas de la LM correspondiente, y los
positivos cantidades altas. La variación en la tendencia se debe a la presencia
alternada, en la captura, de calamares de tallas pequeñas (13-34 cm de LM) y grandes
(62-120 cm de LM). Con excepción de la primera quincena de Mayo y la segunda de
Julio , es notoria la ausencia o baja presencia de calamares de LM medianas (36-60 cm
de LM). Markaida y Sosa-Nishizaki (2001) en su análisis de distribución por LM, sexo y
estadio de madurez, mencionan que la escasez de calamares de tallas medianas,
puede ser efecto de la selectividad de la potera, de la segregación
c M, o
Nosotros consideramos que son necesarios estudios específicos para determinar la
razón de la ausencia de este grupo de intervalos de LM en las capturas.
En el gráfico de β en función del esfuerzo pesquero (Figura 10), se observó que la
captura, durante el tiempo analizado, está sustentada principalmente de tallas
pequeñas, con una disminución del esfuerzo pesquero conforme se incrementa el
empo. La disminución del esfuerzo puede estar asociado a la presencia de calamares
s de la variación de respecto a la LM,
sí como, el seguimiento de sus cambios relativos en función del tiempo y del esfuerzo
ti
grandes, ya que existe una preferencia por calamares de tallas pequeñas en las plantas
procesadoras.
La escasez de datos de esfuerzo limitó el análisi q
a
29

pesquero. No obstante, los resultados permiten concluir que el coeficiente de
capturabilidad varía con la talla y con el tiempo.
La presencia de más de una cohorte en la pesquería de calamar produce una mezcla
en la estructura de tallas y considerando el tamaño y peso de especie Dosidicus gigas,
sto provocó que la captura en número de calamares no reflejara la captura total en
válida para Dosidicus gigas, ya que de los seis
iveles de madurez sólo se pudieron distinguir dos grupos: reproductivamente activos e
iderar que para
nalizar el ciclo reproductivo, es necesario segregar a los organismos maduros de los
e
peso, aún cuando la transformación se hizo con el peso promedio correspondiente a
cada intervalo de LM. Esto no pasaría de existir sólo una cohorte en la pesquería.
En cuanto a madurez gonádica, de las diversas escalas de madurez descritas para
calamares (Juanicó 1983) varias han sido utilizadas para Dosidicus gigas (Michel et al.
1986; Ehrhardt et al. 1986; Tafur y Rabí 1997). Sin embargo ninguna de ellas ha sido
validada histológicamente. La escala de madurez utilizada en este trabajo fue validada
para Loligo vulgaris (Cephalopoda: Liliginidae) (Sauer y Lipinski 1990) Sin embargo,
esta misma escala solo es parcialmente
n
inactivos. identificaron dos grupos, los cuales fueron separados en dos grupos:
reproductivamente activos e inactivos.
La reproducción es un proceso continuo y bien definido, pero la escala de madurez es
en realidad un punto de referencia arbitrario. Dependiendo de los objetivos del análisis,
se puede elegir una escala de dos niveles (maduros e inmaduros) o una escala de
varios niveles (varias etapas inmaduras, varias etapas maduras) (Jaunicó 1983).
Aunque una escala multinivel proporciona información más fina del proceso
reproductivo, requiere también de propiedades morfocromáticas definidas con más
precisión y fácilmente identificables. De no cumplirse esta condición la probabilidad de
traslapo entre los distintos niveles aumenta considerablemente. Dado que la escala
empleada en este estudio es una de las que tiene más niveles de maduración, es
probable que el uso de una escala con menor número de niveles se correlacione mejor
con los cambios histológicos de la gónada. Este cambio debe cons
a
que ya han desovado, mientras que para el análisis de la talla de maduración es
necesario distinguir a los juveniles de los adultos que están en reposo.
30

Aunque existen huecos de información en la serie de tiempo analizada, los resultados
obtenidos del seguimiento de los estadios de maduración en función del desarrollo los
ovocitos fueron consistentes con los reportes de otros autores. Ehrhardt et al. (1983)
identificaron tres picos reproductivos en 1980: 1) Diciembre-Enero, basado en la
presencia de hembras maduras; 2) Mayo-Junio, basado en la presencia de hembras
desovadas y 3) Septiembre, basado en la presencia de hembras maduras. Ramírez y
lett (1985) definen también un pico en Mayo-Junio durante 1981 y una predominancia
l hueco de
ormación en agosto y septiembre, no es fácil de interpretar. Bien puede representar
La constancia de la talla de primera madurez en
un período de 5 años puede ser reflejo de estabilidad en la población, sin embargo sería
recomendable evaluar de manera más precisa el impacto que la captura de juveniles
puede tener en la dinámica del stock.
K
de hembras maduras en Agosto-Septiembre. Estos picos mantienen cierta
correspondencia con los valores altos del IM encontrados en mayo y agosto del 2002.
Con respecto a 2001, el período de transición descrito para junio y julio hacen suponer
que debió haber un pico reproductivo en mayo, lo cual coincide con los estudios previos
y el 2002. Sin embargo, el pico observado en octubre de 2001, dado e
inf
un período reproductivo prolongado, si el calamar empezó su temporada en agosto-
septiembre, o bien puede representar un retraso en el período reproductivo.
Considerando que hasta antes de la década de los 80s, los calamares mayores de 40
cm LM no habían sido reconocidos en el hemisferio norte (Markaida 2001), la frecuencia
de tallas registrada en este estudio demuestra que la pesquería de calamar en el Golfo
de California se sostiene por un intervalo amplio de tallas. Sin embargo, la talla de
primera madurez estimada en 72 cm de LM, implica que el 60% de las hembras
capturadas son juveniles. Durante 1996, Markaida y Sosa-Nishisaki, encontraron que la
mayor parte de las hembras capturadas en Santa. Rosalía maduraban en los 74 cm de
LM (Markaida y Sosa-Nishisaki 2001).
31

RECOMENDACIONES DE MANEJO
1.-Sería favorable para el manejo del recurso, que el permiso de pesca se autorice por
panga o embarcación, esto permitiría tener un mejor control del esfuerzo pesquero y la
vigilancia seria mas eficiente, pues cada panga traería su documentación.
2.-Realizar un sistema de libretas de campo en los sitios pesqueros. Esta libreta puede
ser solicitada a los permisionarios o pescadores cada vez que entreguen los avisos de
arribo y mientras se procesa la información darles otra y así irlas rotando con una
periodicidad preferentemente semanal o quincenal. Generalmente, en cada sitio los
permisionarios anotan en un cuaderno la captura en peso que descarga cada pescador
por día de pesca para después pagarles. Los registros en el cuaderno, hechos por el
encargado del sitio pesquero, proporcionan información de captura y de esfuerzo que
son esenciales para la evaluación de la población de calamar y en consecuencia para el
manejo.
3.-Mantener la dimensión actual del esfuerzo pesquero, es decir, no incrementar el
número de permisos de pesca hasta que se tengan resultados del sistema de libretas
de campo. Con esta medida, la variabilidad en la dimensión del esfuerzo se mantendría
dentro de un intervalo que sería más fácil de medir. Sostener el esfuerzo pesquero no
afectaría social y económicamente la región, pues cuando la abundancia del recurso ha
sido alta, la captura se ve limitada por la capacidad de las plantas procesadoras de
calamar.
El sistema de libretas de campo aplicado al calamar gigante (Dosidicus gigas) en el
área de Santa Rosalía, B.C.S. sería una prueba para medir la captura y el esfuerzo
pesquero y de funcionar podría aplicarse a otras pesquerías artesanales en las que este
tipo de información es difícil de obtener.
3.-El proyecto del sistema de libretas de campo podría iniciarse en el área de Santa
Rosalía, B.C.S. por el personal del proyecto calamar del Centro Regional de
Investigación Pesquera, La Paz, B.C.S., quienes proporcionarían la información
obtenida a la oficina de Administración de pesquerías de la subdelegación estatal de la

SAGARPA. Para hacer esto posible, la solicitud de la libreta a los permisionarios,
tendría que ser un requisito oficial para renovar los permisos de pesca y hacerse a
través de la autoridad correspondiente con el apoyo de la oficina de pesca, donde se
reciben los avisos de arribo.
RECOMENDACIONES DE INVESTIGACIÓN
Realizar estudios bio-económicos, puesto que, en la variación de la captura influyen
factores tales como, el precio del producto y la capacidad de procesamiento del recurso
en las plantas procesadoras. Esto produce problemas en la evaluación de la pesquería
y en consecuencia en el manejo.
Ejecutar estudios para describir los movimientos migratorios de la especie, dentro del
Golfo de California, pues de esto depende la disponibilidad del recurso a la pesquería.
Efectuar por lo menos, otro ciclo de análisis histológico, que incluya la época de
invierno, para completar o corroborar las fases de madurez identificadas, y establecer
una escala de madurez gonádica particular para el calamar gigante.
Continuar con el monitoreo de la pesquería de calamar gigante (Dosidicus gigas), en la
zona de Santa Rosalía, B.C.S., pues los cambios repentinos en la CPUE no son una
tendencia común en recursos de una sola cohorte o generación anual y la alta variación
observada en la zona de Santa Rosalía, B.C.S., no había sido reportada. Esto tiene
impacto en las medidas de administración del recurso y es probable que se necesite
implementar medidas de manejo propias para esta zona.
AGRADECIMIENTOS Los autores agradecen a la Comisión Nacional de Acuacultura y Pesca (CONAPESCA),
Dirección General de Ordenamiento Pesquero y Acuícola, por el financiamiento para
llevar a cabo este estudio. A las Plantas procesadoras de calamar, por la información y
el apoyo brindado durante los muestreos, especialmente a sus gerentes: Ing. Regulo
33

López, C.P. Miguel Tito Cota y C.P. Etelvina Carvajal. A la Subdelegación de Pesca de
la SAGARPA por la información brindada, especialmente al Biól. José Hernández
Lizardi, Jjefe del Departamento de Administración de Pesquerías. Al personal de
campo: Raúl Zamarrón Daniels, Leonardo Méndez López, Luis Arturo Robles Vázquez y
Armando Hernández López.
34

LITERATURA CITADA Agnew, D.J., R. Baranowsky, J.R. Beddington, S. Des Clers, C.P. Nolangneew. 1998. Approaches to assessing stocks of Loligo gahi around the Falkland Island. Fish. Res. 35,155-169. Agnew, D.J., S. Hill, J.R. Beddington. 2000. Predicting recruitment strength of annual squid stock: Loligo gahi around the Falkland Islands. Can. J. Fish. Aquat. Sci. 57, 2479-2487. Arreguín-Sánchez, F. 1996. Catchability a key parameter for fish stocks assessment. Rev. Fish Biol. Fish. 6, 221-242. Basson, M., J. Beddington, J. Crombie, S. Holden, L. Purchase, G. Tingley. 1996. Assessment and management technique for migratory annual squid stocks: The Illex argentinus fishery in the Southwest Atlantic s an example. Fish. Res. 28, 3-27. Bell, T.A., D.V. Lighter. 1988, A handbook of normal penaeid shrimp histology. Allen Press. Kansas, USA. 114 pp. Caddy, J.F., P.G. Rodhouse. 1998. Cephalopod and groundfish landings: evidence for ecological change in global fisheries. Rev. Fish Biol. Fish. 8, 431-444. Ehrhardt, N. M., P.S. Jaquemin, B.F. García, D.G. González, B.J.M. López, C.J. Ortíz, N. A. Solis. 1983. On the fishery and biology of the giant squid Dosidicus gigas in the Gulf of California, Mexico. In: Caddy, J. F. (Ed.), Advances in assessment of world cephalopods resources. FAO Fish. Tech. Papers. 231, 306-340. Ehrhardt N.M., P.S. Jaquemin, B.F. García, D.G. González, B.J.M. López, C.J. Ortíz, N.A. Solís. 1983. On the fishery and biology of the gigant squid Dosidicus gigas in the Gulf of California, Mexico. En: Caddy, J.F.(Ed.), Advances in assessment of world cephalopod resources. FAO Fish. Tech. Papers. 231, 306-340. Ehrthardt, N.M., N.A. Solís, P.S.Jaquemin, C.J. Ortíz, R.P. Ulloa, D.G. González, B.F. García. 1986. Análisis de la biología y condiciones del stock del calamar gigante Dosidicus gigas en el Golfo de California, México, durante 1980. Ciencia Pesquera 5, 63-76. Gould, W.R., K.M. Pollock. 1997. Catch-effort maximum likelihood estimation of important population parameters. Can. J. Fish. Aquat. Sci.. 54, 890-897. Gould, W.R., L.A. Stefanski, K.M. Pollock. 1997. Effect of measurement error on catch-effort estimation. Can. J. Fish. Aquat. Sci. 54, 898-906. Gulland, J.A. 1983. Fish stock assessment. A manual for basic methods. John Wiley and Sons, New York. Hatfield, E.M.C., S. Des Clers. 1998. Fisheries management and research for Loligo gahi in the Falkland Islands. Calif. Coop. Oceanic Fish. Invest. Rep. No. 39,81-91.

Hernández-Herrera, A., E. Morales-Bojórquez, M.A. Cisneros-Mata, M.O. Nevárez-Martínez and G.I. Rivera-Parra. 1998. Management strategy for the giant squid (Dosidicus gigas) from Gulf of California, México. CalCOFI Reports 39, 212-218. Hilborn, R., C. Walters. 1992. Quantitative fisheries stock assessment. Choice, dynamics and uncertainty. Chapman-Hall, New York. 572 pp. Juanicó, M., 1983. Squid maturity scales for population analysis. In: Caddy, J.F.(Ed.), Advances in assessment of world cephalopod resources. FAO Fish. Tech. Pap. 231, 341-378. Klett, T. A., 1996. Pesquería del calamar gigante Dosidicus gigas, In: Casas-Valdez, M., Ponce-Díaz, G., (Eds.), Estudio del potencial pesquero y acuícola de Baja California Sur, Vol. I. CIB-CICIMAR-UABCS. 127-149 p. Lipinski M. 1995. Sexual maturation in squid: quantum or continuum?. S. Afr. J. Mar. Sci. 15, 207-223. Markaida, U., O. Sosa-Nishizaki. 2001. Reproductive biology of jumbo squid Dosidicus gigas in the Gulf of California, 1995-1997. Fish. Res. 54, 63-82. Markaida, U., 2001. Biología del calamar gigante Dosidicus gigas orbigny, 1835 (Cephalopoda: Ommastrephidae) en el Golfo de California, México. Tesis Doctoral. Centro de Investigación Científica y de Educación Superior de Ensenada (CICESE). 381 pp. Martínez-Aguilar, S., J.A. De Anda-Montañez, F. Arreguín-Sánchez. 1997. Densidad y capturabilidad de la sardina Monterrey (Sardinops sagax) (Pises: Clupeidae) del Golfo de California, México. Rev. Biol. Trop. 45:1, 527-535. Michel, E., A. Klett, R.I. Ochoa. 1986. Estudio preliminar para la determinación de madurez gonádica del calamar gigante Dosidicus gigas (D’Orbigny 1835). Ciencia Pesquera 5, 77-89. Morales-Bojórquez, E. A. Hernández-Herrera, M.O. Nevárez-Martínez, G.I. Rivera-Parra, A. Balmori-Ramírez, C. Cervantes-Valle. 2001a. Calamar gigante. En: Sustentabilidad y Pesca responsable en México. Evaluación y manejo. Editores. M.A. Cisneros-Mata, L.F. Beléndez Moreno, E. Zarate-Becerra, M.T. Gapar-Dillanes, L.C. López-González, C. Saucedo-Ruíz y J. Tovar-Avila. Instituto Nacional de la Pesca-SAGARPA. p. 98-117. Morales-Bojórquez, E., M.O. Nevárez-Martínez. 2002. Estimación estocástica de la capturabilidad y reclutamiento del calamar gigante Dosidicus gigas (D’Orbigny, 1835) del Golfo de California, México. Ciencias Marinas. 28:2, 193-204. Morales-Bojórquez, E., A. Hernández-Herrera, M. Nevárez-Martínez, M. A. Cisneros-Mata, F. Guerrero. 2001b. Population size and exploitation of giant squid (Dosidicus gigas) in the Gulf of California México. Scientia Marina, 65:1, 75-80.
36

Morales-Bojórquez, E., A. Hernández-Herrera, M.O. Nevárez-Martínez, A. J. Díaz de León, G.I. Rivera-Parra, A. Ramos. 1997. Abundancia poblacional del calamar gigante (Dosidicus gigas) en las costas de Sonora, México. Oceánides, 12:2, 91-96. Morales-Bojórquez, E., M.A. Cisneros-Mata, A. Hernández-Herrera, M. O. Nevárez-Martínez. 2001c. Review of stock assessment and fisheries research for Dosidicus gigas in the Gulf of California, Mexico. Fish. Res. 54, 393-404. Morales-Bojórquez, E., S. Martínez-Aguilar, F. Arreguín-Sánchez, M.O. Nevárez-Martínez. 2001d. Estimations of catchability-at-length for the jumbo squid (Dosidicus gigas) in the Gulf of California, Mexico. CalCOFI Reports. 42, 167-171. Nesis, K.N., 1970. Biology of the Peru-Chilean gigant squid, Dosidicus gigas. Okeanology. 10, 140-152. Nevárez-Martínez, M.O., A. Hernández-Herrera, E. Morales-Bojórquez, A. Balmori-Ramírez, M. A. Cisneros-Mata, R. Morales. 2000. Distribution and abundance of jumbo squid (Dosidicus gigas) in the Gulf of California, México. Fish. Res. 49:2, 129-140. Nevárez-Martínez, M.O., E. Morales-Bojórquez. 1997. El escape proporcional y el uso del punto de referencia biológico F%BR, para la explotacion del calamar gigante Dosidicus gigas, del Golfo de California, México. Oceánides, 12:2, 97-105. Nigmatullin, CH.M., K.N. Nesis, A.I. Arkhipkin. 2001. A review of the biology of the jumbo squid Dosidicus gigas (Cephalopoda: Omamastrephidae). Fish. Res. 54, 9-19. Ramírez, R.M., A. Klett., 1985. Composición de la captura del calamar gigante en el Golfo de California durante 1981. Trans. CIBCASIO X: 123-127. Ricker, W.E. 1975. Computation and interpretation of biological statistics of fish populations. Bull. Fish. Res. Board Can. 191, 382p. Roberts, M.J. 1998. The influence of the environment on chokka squid Loligo vulgaris reynaudii spawning aggregations: steps towards a quantified model. S. Afr. J. Mar. Sci. 20, 267-284. Robin, J.P., V. Denis. 1999. Squid stock fluctuations and water temperature: temporal analysis of English Channel Loliginidae. J. Appl. Ecol. 36, 101-110. Rosenberg, A. A., G.P. Kirkwood., J.A. Crombie, J.R. Beddington.1990. The assessment of stocks of annual squid species. Fish. Res. 8, 335-350. Sakurai, Y., H. Kiyofuji, S. Sautoh, T. Goto, Y. Hiyama. 2000. Changes in inferred spawning areas of Todarodes pacificus (Cephalopoda: Ommastrephidae) due to changing environmental conditions. ICES J. Mar. Sci. 57, 24-30. Sato, T., 1975. II 3 C. Higashitaiheiyo karihorunia Hanto Oki no America Ooakaikda Dosidicus gigas (Dorbigny) gyogyo kaihatsu no Genjo (Can. Yransl. Fish. Aquat. Sci. No. 4586, 22pp). Prog. Rep. Squid Fish. Survey World 5, 147-154. (In Japanese).
37

Sato, T., 1976. Results of exploratory fishing for Dosidicus gigas (D’Orbigny) off California and Mexico. FAO Fish. Rep. 170: 1, 61-67. Sauer, W.H., M.R. Lipinski. 1990. Histological validation of morphological stages of sexual maturity in chokker squid Loligo vulgaris reynaudii D’Orb (Cephalopoda: Loliginidae). S. Afr. J. Mar. Sci. 9, 189-200. Sherpherd, J.G. 1987. Towards a method for short-term forecasting of catch rates based on length compositions. In Length-based methods in fisheries research. D. Pauly and G.R. Morgan. Eds. International Center for Living Aquatic Resources Management. Manila, Philippines and Kuwait. ICLARM conference proceedings 13, 468 pp. Tafur, R., M. Rabí. 1997. Reproduction of the jumbo squid, Dosidicus gigas (Orbigny, 1835) (Cephalopoda: Ommastrephidae) off Peruvian coasts. Sci. Mar. 61, 2, 33-37. Tafur, R., P. Villegas, M. Rabí, C. Yamashiro. 2001. Dynamics of maturation, seasonality of reproduction and spawning grounds of the jumbo squid Dosidicus gigas (Cephalopoda: Ommastrephidae) in Peruvian waters. Fish. Res. 54, 33-49. Taipe, A., C. Yamashiro, L. Maritegui , P. Rojas, C. Roque. 2001. Distribution and concentration of jumbo flying squid (Dosidicus gigas) off the Peruvian coast between 1991 and 1999. Fish. Res. 54, 22-32. Waluda, C.M., P.G. Rodhouse, G.P. Podestá, P.N. Tratan, G.P. Pierce. 2001. Oceanography of Illex argentinus (Cepahlopod:Ommastrephidae) hatching grounds an influences on recruitment variability. Mar. Biol. In press. Waluda, C.M., P.N. Tratan, P.G. Rodhouse. 1999. Influence of oceanographic variability on recruitment in the genus Illex argentinus (Cephalopoda: Ommastrephidae) fishery in the South Atlantic. Mar. Ecol. Prog. Ser. 183, 159-167.
38

LISTA DE FIGURAS.
Figura.1. Tendencia anual de las capturas de calamar gigante Dosidicus gigas. Modificado de Morales-Bojórquez et al. 2001. Figura 2. Área de estudio en Santa Rosalía, B.C.S. Figura 3.- Distribución de longitudes de manto de Dosidicus gigas durante la temporada 2002. Figura. 4. Captura comercial de calamar gigante (Dosidicus gigas), descargada en las plantas procesadoras de la zona de Santa Rosalía, B.C.S. Temporada de pesca 2002. Figura 5. Relación peso-longitud del calamar gigante (Dosidicus gigas) de la zona de Santa Rosalía, B.C.S. Temporada 2002. Figura 6.- Ajuste del modelo lineal de extracción sucesiva. Figura 7.- Tendencia de la CPUE de calamar gigante Dosidicus gigas. Figura 8. Estimaciones quincenales promedio del coeficiente de capturabilidad en función de la longitud de manto para el periodo comprendido entre el 6 de Mayo al 26 Julio del 2002. Figura 9. Cambios relativos quincenales de la tasa de captura en función de la longitud de manto, respecto al promedio del periodo analizado (6 de mayo al 26 de Julio). Figura 10. Diferencias relativas quincenales de capturabilidad ( tβ ) en función del esfuerzo pesquero (número de viajes). Figura 11. Estimaciones de abundancia en número de calamares y abundancia en toneladas. Figura 12. Índice de madurez estimado en cada fase de madurez gonádica. Cada barra representa el valor promedio y su intervalo al 95% de confianza. La línea horizontal representa el valor promedio estimado para el total de los calamares analizados. Figura 13. Composición de ovocitos (izquierda) e Índice de Madurez quincenal (derecha). Ovocitos 1; Ovocitos 2; Ovocitos 3; Ovocitos 4; Ovocitos 5. Las barras representan el promedio, el intervalo de confianza y el intervalo máximo y mínimo de los índices de madurez estimados en cada quincena. Figura 14. (a) Proporción de organismos maduros por clase longitud de manto. Las barras negras representan el total de organismos y las grises el número de organismos maduros.(b) Frecuencia acumulada de organismos maduros. Se muestra la longitud de manto de primera madurez.

Figura 15. Estimaciones quincenales promedio del coeficiente de capturabilidad en función de la longitud de manto para el periodo comprendido entre el 9 de Mayo a 19 de Julio del 2001. Figura 16. Estimaciones anuales promedio del coeficiente de capturabilidad en función de la longitud de manto para la temporada 2001. : 9 de Mayo a 19 de Julio del 2001 y : 6 de Mayo al 26 Julio del 2002.
tN
1+tN Figura 17. Estadio microscópico del desarrollo de los ovocitos. Primer estadio: ovocito inmaduro o primario. Figura 18. Estadio microscópico del desarrollo de los ovocitos. Segundo estadio: reposo o postdesove, ovocito primario cuboidal. Figura 19. Estadio microscópico del desarrollo de los ovocitos. Tercer estadio: madurando, epitelio invadiendo el ovocito. Figura 20. Estadio microscópico del desarrollo de los ovocitos. Cuarto estadio: maduro, desplazamiento del epitelio folicular. Figura 21. Estadio microscópico del desarrollo de los ovocitos. Quinto estadio: ovocito listo para ser liberado.
40

FIGURAS
116o 115o 114o 113o 112o 111o 110o 109o 108o 107o 106o
23o
24o
25o
26o
27o
28o
29o
30o
31o
32o
Océano Pacífico Golfo de California
MÉXICO
Guaymas
SantaRosalía
Figura 1.
41

0
20
40
60
80
100
120
140
160
Cap
tura
(mile
s de
tone
lada
s)
Tiem po (años)
TotalBaja Cal. Sur.
Figura 2.
42

Julio-2
05
10152025
30 36 42 48 54 60 66 72 78 84 90
Frec
uenc
ia (%
)
Julio-26
0
5
10
15
20
25
30 36 42 48 54 60 66 72 78 84 90
Longitud de manto (cm)
Frec
uenc
ia (%
)
Agosto-30
05
10152025
30 36 42 48 54 60 66 72 78 84 90
Agosto-7
05
10152025
30 36 42 48 54 60 66 72 78 84 90
Mayo-6
05
10152025
30 36 42 48 54 60 66 72 78 84 90
Frec
uenc
ia (%
)
Mayo-29
05
10152025
30 36 42 48 54 60 66 72 78 84 90
Frec
uenc
ia (%
)
Junio-12
05
10152025
30 36 42 48 54 60 66 72 78 84 90
Frec
uenc
ia (%
)
`
Octubre-8
0
5
10
15
20
30 36 42 48 54 60 66 72 78 84 90
Septiembre-12
0
5
10
15
20
30 36 42 48 54 60 66 72 78 84 90
Octubre-22
05
101520
30 36 42 48 54 60 66 72 78 84 90
Longitud de manto (cm)
Figura 3.
43

0
100
200
300
400
500
1 2 3 4 5 6 7 8 9 10
Tiempo (quincenas)
Cap
tura
(num
ero
de c
alam
ares
) x
10 3
Figura 4.
44

Figura 5.
050
100150200250300350400
0 1 2 3 4 5 6 7 8 9 10 11
Tiempo (quincenas)
CP
UE
0
5
10
15
20
25
22 26 30 34 38 42 46 50 54 58 62 66 70 74 78 82 86 90
Longitud de manto (cm).
Pes
o de
man
to (K
g)
45

y = -0.0001x + 343.36R2 = 0.2155
0
50
100
150
200
250
300
600,000 800,000 1,000,000 1,200,000 1,400,000
Captura acumulada
CP
UE
Figura 6.
46

0
100
200
300
400
0 1 2 3 4 5 6 7 8 9 10 11
Tiempo (quincenas)
CP
UE
Figura 7.
47

Figura 8.
0
0.0005
0.001
0.0015
0.002
0.0025
0.003
28 32 36 40 44 48 52 56 60 64 68 72 76 80 84 88
Longitud de manto (cm)
Cap
tura
bilid
ad
48
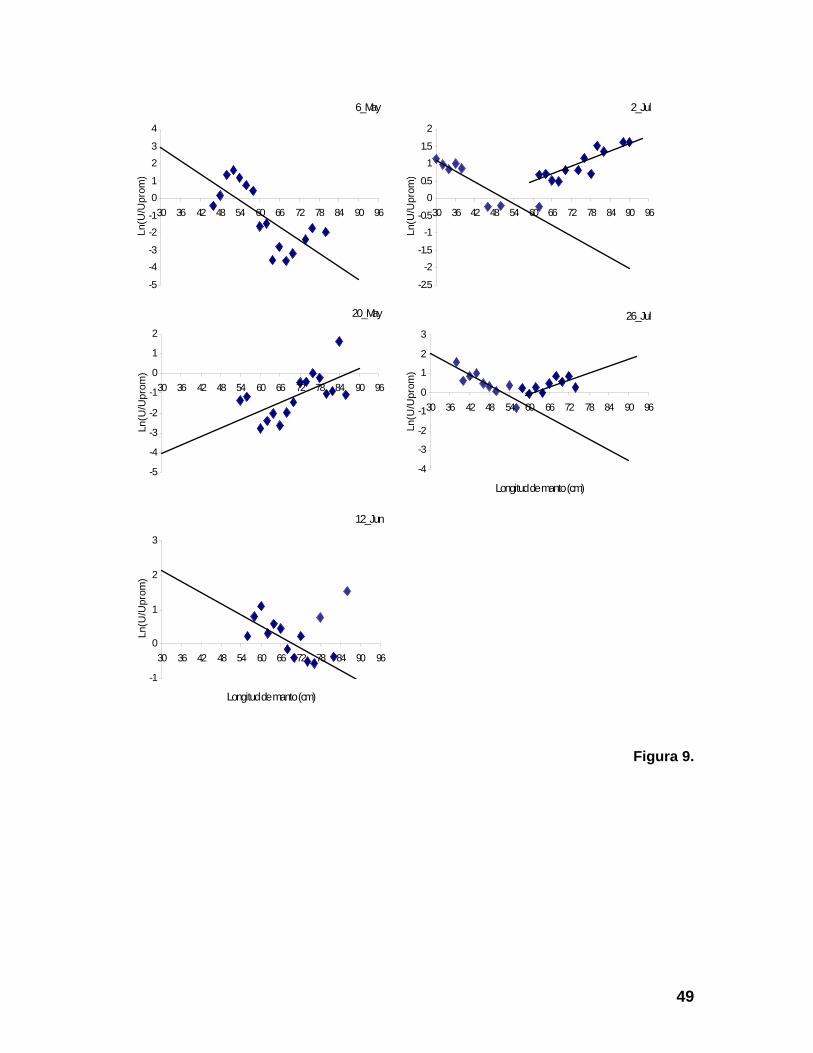
-5
-4-3
-2-1
01
23
4
30 36 42 48 54 60 66 72 78 84 90 96
Ln(U
/Upr
om)
6_May
-5
-4
-3
-2
-1
0
1
2
30 36 42 48 54 60 66 72 78 84 90 96
Ln(U
/Upr
om)
20_May
-1
0
1
2
3
30 36 42 48 54 60 66 72 78 84 90 96
Longitud de manto (cm)
Ln(U
/Upr
om)
12_Jun
-2.5
-2-1.5
-1-0.5
00.5
11.5
2
30 36 42 48 54 60 66 72 78 84 90 96
Ln(U
/Upr
om)
2_Jul
-4
-3
-2
-1
0
1
2
3
30 36 42 48 54 60 66 72 78 84 90 96
Longitud de manto (cm)
Ln(U
/Upr
om)
26_Jul
Figura 9.
49

6_May
26_Jul
12_Jun2_Jul
29_May
26_Jul
2_Jul
-0.15
-0.1
-0.05
0
0.05
0.1
0 500 1000 1500 2000 2500
Esfuerzo pesquero
βt
Figura 10.
50

0
500,000
1,000,000
1,500,000
2,000,000
2,500,000
1 2 3 4
Tiempo (quincenas)
Nt (
num
ero
de c
alm
ares
)
-3,000
2,000
7,000
12,000
17,000
22,000
Bt (
tone
lada
s)
Nt
Bt
Figura 11.
51

0
1
2
3
4
5
I II III IV V VI
Madurez Macroscópica
Indi
ce d
e M
adur
ez (I
M)
Figura 12
52

0
1
2
3
4
5
MAY JUN JUL AGO SEP OCT NOVÍn
dice
de
Mad
urez
2001
0.0
0.2
0.4
0.6
0.8
1.0
MAY JUN JUL AGO SEP OCT NOV
Frec
uenc
ia R
elat
iva
0
1
2
3
4
5
MAY JUN JUL AGO SEP OCT NOV
Índi
ce d
e M
adur
ez
2002
0.0
0.2
0.4
0.6
0.8
1.0
MAY JUN JUL AGO SEP OCT NOV
Frec
uenc
ia R
elat
iva
Figura 13
53

Figura 14
0
4
8
12
16
20
33 43 53 63 73 83 93
Longitud del Manto (cm)
Frec
uenc
ia (%
)
0.0
0.2
0.4
0.6
0.8
1.0
0 10 20 30 40 50 60 70 80 90 100
Longitud del Manto (cm)
Hem
bras
Mad
uras
(%A
cum
)
72
54

0
0.0005
0.001
0.0015
0.002
0.0025
28 32 36 40 44 48 52 56 60 64 68 72 76 80 84 88
Longitud de manto (cm)
Cap
tura
bilid
ad
Figura 15
55

00.000020.000040.000060.000080.0001
0.000120.000140.00016
30 36 42 48 54 60 66 72 78 84 90
Longitud de manto (cm)
Cap
tura
bilid
ad
Figura 16
56

Figura 17
57

Figura 18
58

Figura 19
59

Figura 20
60

Figura 21
61

TABLAS Tabla 1. Descripción morfológica de las fases de maduración gonádica usada en calamares (Sauer y Lipinski 1990). Fase de Maduración Características morfológicas I Juvenil Glándulas nidamentales como bandas transparentes. El resto de las
estructuras no son visibles a simple vista II Reposo Conducto ovárico visible. El ovario es homogéneo III Preparatorio Conducto ovárico extendido. Huevos inmaduros visibles IV Madurando Glándulas nidamentales grandes. Se observan algunos huevos
maduros (amarillos) en el ovario. No existen huevos maduros en el oviducto
V Maduro Huevos maduros en el oviducto. Hay secreción en las glándulas nidamentales
VI Desovado Indefinido
62

Tabla 2. Proporción macho:hembra, registrado en los muestreos masivos de la capturas comerciales de calamar gigante (Dosidicus gigas) en la zona de Santa Rosalía, B.C.S., durante el periodo comprendido entre Mayo y Octubre del 2002.
Quincena Macho-Hembra Mayo-6 1:1.2 Mayo-29 1:3.5 Junio-12 1:1 Julio-2 1:1 Julio-26 3:1 Agosto-7 1:1.2 Agosto-30 1:1.2 Septiembre-26 1:9 Octubre-8 1:1 Octubre-22 1:3.5
Tabla 3. Índice de los cambios relativos de capturabilidad en función de la longitud de manto ( β ) y el índice de la abundancia de los calamares más pequeños presentes en lcaptura (
a α ) para el periodo de tiempo analizado. R (factor de correlación). ,
Quincena � � R Mayo-6 -0.1267 6.7285 0.7 Mayo-29 0.0715 -6.197 0.6 Junio-12 -0.0533 3.7225 0.8 Julio-2 -0.0515 2.6294 0.9 0.0407 -2.0446 0.9 Julio-26 -0.0923 4.7932 0.9 -0.0371 -2.0548 0.6
63

Tabla 4. Número de machos y hembras registrados en lo muestreos biológicos de la capturas comerciales de calamar gigante (Dosidicus gigas) en la zona de Santa Rosalía, B.C.S., durante el periodo comprendido entre junio y noviembre del 2001 y de mayo a noviembre del 2002. QNA (quincena).
Número de calamares Mes Quincena Indiferenciados Hembras Machos JUN/2001 1 0 11 0 2 2 10 7 JUL 1 0 16 1 2 0 16 0 AGO 1 0 6 0 2 SEP 1 2 OCT 1 0 18 0 2 NOV 1 0 20 0 2 0 20 0 MAY/2002 1 0 17 9 2 0 36 10 JUN 1 0 21 25 2 JUL 1 0 11 16 2 0 9 20 AGO 1 3 16 6 2 0 22 7 SEP 1 0 21 9 2 2 17 0 OCT 1 1 3 2 2 2 14 2 TOTAL 19 10 304 114
64

Tabla 5. Descripción de las etapas de desarrollo de los ovocitos de Dosidicus gigas. Estadio del Ovocito
Nivel de Desarrollo (S) Características celulares
Inmaduro 1
Ovocito con núcleo esférico con uno o varios nucleolos, rodeados por un citoplasma eosinófilo. Se observa una capa delgada de células germinales alrededor del ovocito. (Figura 17)
Reposo o Postdesove 2
Solo se observan restos de ovocitos maduros. Tejido conjuntivo abundante y desorganizado, lo cual dificulta el conteo de ovocitos (Figura 18).
Madurando 3
Ovocitos de mayor tamaño con nucleolos menos aparentes. Proliferan las células germinales que se adhieren al ovocito y junto con el epitelio folicular, comienzan a invadir el ovocito formando interdigitaciones. Su morfología cambia de cuadrada a cúbica (Figura 19).
Maduro 4
El núcleo migra hacia uno de los polos del ovocito. Las interdigitaciones se retiran hacia la periferia y su morfología cambia de cúbica a columnar (Figura 20).
A punto de expulsión 5
El núcleo y los nucleolos disminuyen de talla hasta desaparecer. Las células foliculares se retraen y forman una estructura laminar alrededor del óvulo. Hay formación de vitelo y se incrementa el citoplasma eosinófilo (Figura 21).
65

Tabla 6. Composición de ovocitos y promedio del índice de madures (IM) en cada fase de madurez gonádica. Tamaño de la muestra (N).
Madurez Nivel de desarrollo de los ovocitos
Gonadal 1 2 3 4 5 IM N
I 93.0 7.0 - - - 1.070 43
II 74.3 12.1 8.8 4.6 0.2 1.443 91
III 61.8 - 28.6 9.5 0.2 1.863 35
IV 1.9 5.9 25.3 62.3 4.6 3.618 17
V 17.7 - 30.9 33.0 18.4 3.345 39
33.3 - 48.0 12.5 6.1 2.581 3 VI
- 100.0 - - - 2.000 3
66

Tabla 7. Resultados del análisis de varianza del promedio del índice de madurez (IM), y de la prueba de Tukey para comparación múltiple de factores que se aplicó después de que el análisis de varianza resultó significativo. El nivel de confianza (1-α) de referencia es del 95%. Pruebas altamente significativas con P ≤ 0.01 (***) y no significativas (NS), (P > 0.05).
Suma de Cuadrado Fuente Cuadrados g.l. Medio F Prob.
IM 180.777 5 36.155 49.783 6.99x10-30 ***
Error 163.408 225 0.726
Total 344.185 230
Prueba de Tukey para diferentes tamaños de muestra
Etapas
comparadas P
I-II 0.3232 NS
II-III 0.3103 NS
III-IV 2.03x10-5 ***
IV-V 0.9370 NS
V-VI 0.2653 NS
67

“La pesquería del calamar gigante (Dosidicus gigas) en el Golfo de California. Recomendaciones de Investigación y Tácticas de Regulación “ se terminó de imprimir en Diciembre del 2004 en Talleres Gráficos de México. Se tiraron 500 ejemplares.
68