estudo da interferência de fármacos, que atuam via receptores … · gaba no metabolismo do t....
TRANSCRIPT

FLAacuteVIA SILVA DAMASCENO
Estudo da interferecircncia de faacutermacos que atuam via
receptores eou transportadores de GABA e Glutamato
no ciclo de vida do Trypanosoma cruzi
Dissertaccedilatildeo apresentada ao Programa de Poacutes-
Graduaccedilatildeo em Biologia da Relaccedilatildeo Patoacutegeno-
Hospedeiro do Instituto de Ciecircncias Biomeacutedicas da
Universidade de Satildeo Paulo para obtenccedilatildeo do Tiacutetulo
de Mestre em Ciecircncias
Satildeo Paulo
2013
FLAacuteVIA SILVA DAMASCENO
Estudo da interferecircncia de faacutermacos que atuam via
receptores eou transportadores de GABA e Glutamato
no ciclo de vida do Trypanosoma cruzi
Dissertaccedilatildeo apresentada ao Programa de Poacutes-
Graduaccedilatildeo em Biologia da Relaccedilatildeo Patoacutegeno-
Hospedeiro do Instituto de Ciecircncias Biomeacutedicas da
Universidade de Satildeo Paulo para obtenccedilatildeo do Tiacutetulo
de Mestre em Ciecircncias
Aacuterea de concentraccedilatildeo Biologia da Relaccedilatildeo
Patoacutegeno-Hospedeiro
Orientador Prof Dr Ariel Mariano Silber
Versatildeo corrigida A versatildeo original eletrocircnica
encontra-se disponiacutevel tanto na Biblioteca do ICB
quanto na Biblioteca Digital de Teses e Dissertaccedilotildees
da USP (BDTD)
Satildeo Paulo
2013
Trabalho realizado com auxiacutelios financeiros
concedido pela Fundaccedilatildeo de Amparo a
Pesquisa do Estado de Satildeo Paulo
(FAPESP) Instituto Nacional de
Biotecnologia Estrutural e Quiacutemica
Medicinal em Doenccedilas Infecciosas
(INBEQMeDI) e Conselho Nacional de
Desenvolvimento Cientiacutefico e Tecnoloacutegico
(CNPq)
Aos meus Pais Flamarion e Maria Nasareacute pelo
apoio amor e compreensatildeo incondicionais
AGRADECIMENTOS
Ao Prof Dr Ariel Mariano Silber pela grande oportunidade que me concedeu pela
orientaccedilatildeo paciecircncia apoio e amizade Obrigada por tudo
Agrave Elizabeth Pral teacutecnica do laboratoacuterio pelo apoio amizade ajuda e todo o suporte teacutecnico
Agrave minha amiga Mariacutea Julia Bariacuteson pela amizade apoio ajuda nos experimentos e nas
discussotildees dos resultados
Agrave minha famiacutelia meus pais Flamarion e Maria Nasareacute meus irmatildeos(atildes) Gilberto Gilmar
Gilson Gilsa Gildo Gildete e Jacoacute meus sobrinhos(as) Mariane Mateus Gisely Ceciacutelia
Nataacutelia Isabele Laviacutenia Maria e Venacircncio pelo apoio compreensatildeo e incentivo em todos os
momentos
Agrave minha amiga Valnia Pita por me fazer acreditar que era possiacutevel a realizaccedilatildeo desse sonho e
pelo apoio e estiacutemulo nessa caminhada
Aos meus amigos(as) do laboratoacuterio Raiacutessa Melo Marcell Crispim Sandra Carla Higo
Fernando Inga Eva Jean Douglas Carolina Manchola Ludmila Nakamura Brian Suarez
Rossana Duque e Letiacutecia Marchese pelo apoio ajuda e amizade
Ao Prof Dr Carsten Wrenger e seu grupo Flaacutevia Zimbres Kamila Meissner Jasmin
Lindner Thales Kronenberger
A Dra Lisvane Paes pelo apoio amizade e colaboraccedilatildeo nesse trabalho
Ao Prof Dr Joseacute Roberto Meyer-Fernandes (UFRJ) e aos integrantes do seu laboratoacuterio pela
disponibilidade e ajuda em alguns experimentos
Agrave Profa Dra Alicia J Kowaltowski do Instituto de Quiacutemica e aos integrantes do seu
laboratoacuterio pela disponibilidade do uso de equipamentos
Ao Prof Dr Gerhard Wunderlich e integrantes do seu laboratoacuterio em especial ao doutorando
Wesley Fotoran pela disponibilidade do uso de equipamentos e ajuda em alguns
experimentos
Agrave Fundaccedilatildeo de Amparo a Pesquisa do Estado de Satildeo Paulo (FAPESP) pelo suporte
financeiro
Ao Conselho Nacional de Desenvolvimento Cientiacutefico e Tecnoloacutegico (CNPq) pelo suporte
financeiro no primeiro ano desse trabalho
Agrave todos os professores funcionaacuterios e colegas do Departamento de Parasitologia (ICB-USP)
pelo apoio e convivecircncia
ldquoNunca deixe que lhe digam
que natildeo vale a pena
acreditar no sonho que se tem
ou que seus planos nunca vatildeo dar certo
ou que vocecirc nunca vai ser algueacutem ()
Confie em si mesmordquo
Renato Russo e Flaacutevio Venturini 1987
RESUMO
Damasceno FS Estudo da interferecircncia de faacutermacos que atuam via receptores eou
transportadores de GABA e Glutamato no ciclo de vida do Trypanosoma cruzi [dissertaccedilatildeo
(Mestrado em Biologia da Relaccedilatildeo Patoacutegeno-Hospedeiro)] Satildeo Paulo Instituto de Ciecircncias
Biomeacutedicas Universidade de Satildeo Paulo 2013
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica central e do Sul A quimioterapia disponiacutevel atualmente
limita-se a dois compostos Nifurtimox e o Benzonidazol Ambos os faacutermacos reduzem os
sintomas da doenccedila e a mortalidade das pessoas infectadas quando utilizadas na fase aguda
mas sua eficaacutecia na fase crocircnica eacute controversa Aleacutem do mais apresentam vaacuterios efeitos
colaterais O T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico como tambeacutem a importacircncia do aminoaacutecido
GABA no metabolismo do T cruzi visto que este tambeacutem eacute transportado pelo parasita Nesse
contexto o objetivo desse trabalho foi avaliar a interferecircncia de faacutermacos que atuam via
receptores ou transportadores de GABA e glutamato no ciclo de vida do T cruzi Os
faacutermacos selecionados foram Vigabatrina Pregabalina MK-801 MTEP e Memantina Os
resultados demonstram que apenas Memantina apresentou um melhor efeito tripanocida
Inibiu a proliferaccedilatildeo de formas epimastigotas interferiu na metaciclogecircnese como tambeacutem
afetou o metabolismo energeacutetico do parasita com a diminuiccedilatildeo dos niacuteveis de ATP aleacutem de
desencadear mecanismos que levam a apoptose das formas epimastigotas Memantina
tambeacutem interferiu no ciclo intracelular do parasita mais especificamente no estaacutegio
amastigota Interessantemente as fases do ciclo do parasita que satildeo mais afetadas satildeo as fases
que requerem mais energia estaacutegios replicativos (amastigota e epimastigota) como tambeacutem
os processos de diferenciaccedilatildeo e invasatildeo celular Apesar de apresentar valores de IC50 que
podem ser considerados altos em relaccedilatildeo aos valores descritos para formas epimastigotas
tratadas com Benzonidazol esses resultados mostram perspectivas promissoras visto que o
Memantina poderaacute ser utilizado como composto liacuteder para o desenho de derivados com
atividade tripanocida otimizada
Palavras-chave Trypanosoma cruzi Memantina Receptor de glutamato
ABSTRACT
Damasceno FS Study of interference of drugs that act via receptors andor transport of
GABA and glutamate in the life cycle of Trypanosoma cruzi [Masters thesis (Host-Pathogen
Relationship Biology)] Satildeo Paulo Instituto de Ciecircncias Biomeacutedicas Universidade de Satildeo
Paulo 2013
Chagas disease also known as American trypanosomiasis is caused by the flagellate
protozoan Trypanosoma cruzi and affects approximately 10 million people in endemic areas
of Mexico Central America and South America with about 25 million people living in areas
at risk of infection The currently available chemotherapy is limited to two compounds which
are Nifurtimox and Benznidazole Both drugs reduce symptoms of the disease and mortality
of infected people when used in the acute phase but its efficacy in chronic phase (phase in
which the majority of cases are diagnosed) is still controversial Moreover these drugs have
several side effects T cruzi is able to utilize carbohydrate and amino acids as carbon and
energy source Our group has studied the involvement of glutamate in parasite resistance to
thermal oxidative and metabolic stress as well as the importance of the amino acid GABA
metabolism in T cruzi The aim of this study was to evaluate the interference of drugs that
act in GABA and glutamate transporters or receptors in the life cycle of T cruzi The drugs
selected were vigabatrin pregabalin MK-801 memantine and MTEP The results showed
that only memantine has a better trypanocidal effect Memantine inhibited the proliferation of
epimastigotes interfered in metacyclogenesis and affected the energy metabolism of the
parasite with decreased levels of ATP and trigger mechanisms that lead to apoptosis of
epimastigotes Moreover interferes with intracellular cycle of the parasite specifically in
amastigote stage Interesting that stages of the parasite that require more energy are more
affected replicative stages (epimastigote and amastigote) as well as the processes of
differentiation and cell invasion IC50 value is more than that described to epimastigotes
treateds with Benznidazole but Memantine could be used as leader compound for the design
of drugs with optimized trypanocidal activity
Keywords Trypanosoma cruzi Memantine Glutamate receptor
LISTA DE ILUSTRACcedilOtildeES
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi 19
Figura 2 - Estruturas quiacutemicas do Benzonidazol e Nifurtimox 24
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica 28
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica 29
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse 40
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os
tratamentos com MK-801 ou Memantina 41
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina 42
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico ou oxidativo 44
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na
presenccedila do Memantina 45
Figura 10 - Efeito da Memantina na metaciclogecircnese 46
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo
de morte celular 48
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2 49
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
50
Figura 14 - Formas epimastigotas de T cruzi 51
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi 53
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade
de formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo
celular 54
LISTA DE ABREVIATURAS
ANOVA Anaacutelise de variacircncia
ATP Adenosina trifosfato
CHO-K1 Chinese Hamster Ovary (linhagem celular proveniente de ovaacuterio de
hamester chinecircs)
Ca2+
Caacutelcio
CO2 Dioacutexido de carbono
DNA Aacutecido desoxirribonucleacuteico
DO Densidade oacutetica
DTUs Discrete Typing Units
EGTA Aacutecido etilenoglicol-bis-aminoetiletertetraceacutetico
EROs Espeacutecies reativas de oxigecircnio
GABA Aminoaacutecido γ-aminobutiacuterico
GABA T Aminoaacutecido γ-aminobutiacuterico transaminase
GAPDH Gliceraldeiacutedo -3-fosfato desidrogenase
H2O2 Peroacutexido de hidrogecircnio
IC50 Metade da concentraccedilatildeo maacutexima inibitoacuteria
KCl Cloreto de potaacutessio
LIT Liver Infusion-Tryptose (Meio de cultura)
MCP Morte celular programada
mg Miligramas
mGluR Receptor metabotroacutepico de glutamato
MTT (3-(45-dimetiltiazol-2-il)-25 difenil bromo tetrazolio)
Na2HPO4 Fosfato de soacutedio monobaacutesico anidro
NaCl Cloreto de soacutedio
NaHCO3 Bicarbonato de soacutedio
NCCD Nomenclature Comittee on Cell Death (Comite de Nomeclatura de Morte
Celular)
NH4+
Ion amocircnio
NMDA N-metil-D-aspartato
OH Radical hidroxila
PBS Soluccedilatildeo salina tamponada com fosfato
pH Potencial hidrogeniocircnico
RPMI Roswell Park Memorial Institute
SDS Soacutedio dodecil sulfat
SFB Soro fetal bovino
U Unidades
μg Micrograma
μl Microlitro
μm Microcircmetro
SUMAacuteRIO
1 INTRODUCcedilAtildeO 16
11 Aspectos gerais da doenccedila de chagas 16
12 Trypanosoma cruzi 18
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos 20
14 Morte celular em Trypanosoma cruzi 21
15 Terapia na doenccedila de Chagas 23
16 Perspectivas de novos faacutermacos 25
2 OBJETIVOS 30
21 Objetivo geral 30
22 Objetivos especiacuteficos 30
3 MATERIAIS E MEacuteTODOS 31
31 Microorganismos utilizados no trabalho 31
311 Ceacutelulas hospedeiras 31
312 Trypanosoma cruzi 31
32 Manutenccedilatildeo das ceacutelulas hospedeiras 31
33 Obtenccedilatildeo dos parasitas 31
331 Epimastigotas 31
332 Curvas de crescimento de epimastigotas de T cruzi 31
333 Formas tripomastigotas 32
34 Resistecircncia a diferentes tipos de estresse 32
341 Ensaio de estresse teacutermico em formas epimastigotas 32
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico 33
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular 33
343 Ensaio de estresse oxidativo em formas epimastigotas 33
35 Tipo de morte celular em parasitas tratados com Memantina 34
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita 34
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas
epimastigotas 35
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio 35
36 Efeito sobre a metaciclogecircnese 35
37 Anaacutelise do efeito dos compostos no ciclo intracelular do T cruzi 36
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos 36
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas 36
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas
tripomastigotas 37
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os
compostos 37
38 Anaacutelises estatiacutesticas 38
4 RESULTADOS 39
41 Curvas de crescimento do T cruzi na presenccedila dos compostos 39
42 Interaccedilatildeo com condiccedilotildees de estresse 41
43 Efeito da Memantina na metaciclogecircnese 46
44 Tipo de morte celular sofrida pelo parasita tratado com Memantina 47
45 Efeito da Memantina no ciclo intracelular do T cruzi 52
5 DISCUSSAtildeO 55
6 CONCLUSOtildeES 59
REFEREcircNCIAS 61
16
1 INTRODUCcedilAtildeO
11 Aspectos gerais da doenccedila de chagas
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana foi
descoberta pelo meacutedico brasileiro Carlos Chagas (Chagas 1909) Esta doenccedila eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica Central e do Sul com cerca de 25 milhotildees de pessoas
vivendo em aacutereas de risco de infecccedilatildeo Ocorrem por ano aproximadamente 40 mil novos
casos e 12 mil mortes devido a complicaccedilotildees cardiacuteacas e digestivas (World Health
Organization 2012) Programas de controle do vetor desenvolvidos nos uacuteltimos 20 anos
como tambeacutem um maior controle nos bancos de sangue tem reduzido o nuacutemero de novos
casos da doenccedila de Chagas na Ameacuterica Latina (Rassi Marin-Neto 2010) Devido as intensas
migraccedilotildees de pessoas de paiacuteses endecircmicos o nuacutemero de casos de indiviacuteduos infectados tem
aumentado em aacutereas consideradas natildeo endecircmicas tais como Estados Unidos Canadaacute e
algumas regiotildees da Europa (Bern et al 2009 Maguire 2006 Schmunis 2007) A doenccedila eacute
uma zoonose que tem como reservatoacuterio natural do T cruzi uma ampla variedade de
mamiacuteferos do continente americano (Urbina Docampo 2003) Eacute transmitida para os seres
humanos e para mais de 150 espeacutecies de animais domeacutesticos e animais selvagens
principalmente por insetos hematoacutefagos da famiacutelia Reduviidae Apesar de existirem mais de
130 espeacutecies de triatomiacuteneos satildeo trecircs as mais importantes na transmissatildeo do T cruzi para o
homem Triatoma infestans Rhodnius prolixus e Triatoma dimidiata (Rassi Marin-Neto
2010) O parasita tambeacutem pode ser transmitido por transfusatildeo de sangue congenitamente (de
matildees infectadas para receacutem-nascidos) por transplante de oacutergatildeos ou medula oacutessea Tambeacutem
pode ocorrer por acidentes em laboratoacuterios que trabalham com o parasita vivo via esta que
epidemiologicamente eacute pouco relevante Outra forma de transmissatildeo eacute por via oral atraveacutes de
alimentos contaminados essa via vecircm sendo considerada como relevante na manutenccedilatildeo do
ciclo de transmissatildeo natural (Pereira et al 2009 Rassi Marin-Neto 2010)
A evoluccedilatildeo cliacutenica da doenccedila de Chagas pode ser dividida em duas fases aguda e
crocircnica A fase aguda normalmente eacute assintomaacutetica ou apresenta sintomatologia discreta que
na maioria dos casos passa despercebida pelo paciente Os sintomas quando aparecem o
fazem uma ou duas semanas apoacutes a exposiccedilatildeo ao triatomiacuteneo infectado ou poucos meses apoacutes
a transfusatildeo com sangue contaminado (Rassi Marin-Neto 2010) Os sintomas mais
frequentemente apresentados pelos pacientes agudos satildeo febre prolongada mal-estar
17
distensatildeo do fiacutegado baccedilo e linfonodos Pode haver tambeacutem edema subcutacircneo localizado ou
generalizado No caso particular de transmissatildeo vetorial o paciente pode apresentar um sinal
da porta de entrada do T cruzi na pele (chagoma) ou via mucosa ocular (sinal de Romantildea) As
manifestaccedilotildees da fase aguda se resolvem espontaneamente em aproximadamente 90 dos
indiviacuteduos infectados mesmo sem o tratamento com drogas tripanocidas Essa fase eacute
caracterizada pela alta parasitemia e ausecircncia de anticorpos especiacuteficos Nessa fase os
parasitas podem ser encontrados em praticamente todos os tecidos estando majoritariamente
presentes no sangue Por ser uma fase de curta duraccedilatildeo sem sintomatologia aparente ou com
sintomatologia pouco definida haacute um pequeno nuacutemero de casos detectados sendo portanto a
fase menos estudada (Rassi Marin-Neto 2010 Silber et al 2005b)
Logo apoacutes a fase aguda tem iniacutecio a fase crocircnica caracterizada pelos altos tiacutetulos de
anticorpos no sangue do paciente e pelo fato de que a parasitemia eacute inaparente O paciente
pode permanecer por vaacuterios anos ou ateacute mesmo pelo resto da vida sem apresentar sinais ou
sintomas Essa forma da fase crocircnica eacute chamada de indeterminada Aproximadamente 30-40
dos pacientes crocircnicos indeterminados desenvolvem a sintomatologia caracteriacutestica da
doenccedila evoluindo para a chamada fase crocircnica sintomaacutetica Nesta fase as trecircs principais
formas satildeo a cardiacuteaca a digestiva e a cardiodigestiva (Acquatella 2007 Marin-Neto et al
1999) Na forma cardiacuteaca o coraccedilatildeo eacute um dos primeiros oacutergatildeos afetados natildeo necessariamente
o uacutenico desenvolvendo hipertrofia e dilataccedilatildeo Na forma digestiva o trato digestivo apresenta
alteraccedilotildees anatocircmicas e fisioloacutegicas importantes sendo a principal altereccedilatildeo a dilataccedilatildeo do
intestino e do esocircfago que podem apresentar aspecto de megaviacutescera (Marin-Neto et al
1999 Rassi et al 2000) revisado por (Boscardin et al 2009 Rassi Marin-Neto 2010)
18
12 Trypanosoma cruzi
Trypanosoma cruzi eacute um parasita intracelular obrigatoacuterio classicamente localizado na
ordem kinetoplastida gecircnero Trypanosoma que causa uma infecccedilatildeo crocircnica em humanos e
em grande nuacutemero de outras espeacutecies de mamiacuteferos (Hoare 1972) Recentemente devido agrave
alta variabilidade bioloacutegica bioquiacutemica e molecular a espeacutecie Trypanosoma cruzi foi
dividida em seis linhagens subsespeciacuteficas (Discrete Typing Units ndash DTUs) numerados de I a
VI Esta classificaccedilatildeo foi realizada com base em caracteriacutesticas geneacuteticas moleculares e
imunoloacutegicas com o objetivo de unificar a nomenclatura e facilitar a comunicaccedilatildeo dentro da
comunidade cientiacutefica (Zingales et al 2009)
O T cruzi tem um ciclo de vida complexo que alterna entre o inseto vetor e o
hospedeiro mamiacutefero O parasita apresenta diferentes formas correspondentes a diferentes
estaacutegios do seu ciclo de vida Estes diferentes estaacutegios tecircm sido descritos com base em
criteacuterios morfoloacutegicos como a forma fusiforme ou arredondados a regiatildeo onde emerge o
flagelo e a posiccedilatildeo do cinetoplasto (estrutura contendo o DNA da uacutenica mitococircndria presente
no parasita) em relaccedilatildeo ao nuacutecleo No hospedeiro mamiacutefero foram descritas as formas
amastigota epimastigota intracelular e tripomastigota sanguiacutecola A forma amastigota eacute um
estaacutegio replicativo e infectivo Satildeo ceacutelulas arredondadas medindo de 2-6 microm de diacircmetro
encontradas no citoplasma das ceacutelulas infectadas Os epimastigotas intracelulares
(epimastigote-like) satildeo formas replicativas intermediaacuterias entre amastigotas e tripomastigotas
tambeacutem encontrados dentro das ceacutelulas medindo aproximadamente 54 microm Essas formas satildeo
menores que as formas epimastigotas encontradas no inseto vetor poreacutem compartilha com as
mesmas padrotildees morfomeacutetricos e diversas caracteriacutesticas bioquiacutemicas e bioloacutegicas (Almeida-
De-Faria et al 1999) O tripomastigota sanguiacutecola eacute um estaacutegio flagelado infectivo natildeo
replicativo encontrado principalmente na corrente sanguiacutenea do hospedeiro mamiacutefero Mede
aproximadamente 18 microm de comprimento incluindo um extremo livre do flagelo de
aproximadamente 6 microm e 2-3 microm de largura
No inseto vetor encontram-se predominantemente as formas epimastigota e
tripomastigota metaciacuteclico Epimastigota eacute uma forma flagelada replicativa e natildeo infectiva
Mede aproximadamente 20-40 microm de comprimento e 2-5 microm de largura essas formas estatildeo
presentes tipicamente na porccedilatildeo meacutedia do intestino do hospedeiro invertebrado
Tripomastigota metaciacuteclico eacute uma forma infectiva natildeo replicativa encontrada na porccedilatildeo final
do intestino do inseto vetor (Brener 1971) revisado por (Silber et al 2005a Tyler Engman
2001)
19
O ciclo de vida do T cruzi inicia-se quando os parasitas presentes na corrente
sanguiacutenea do hospedeiro vertebrado infectado passam ao inseto vetor durante o repasto
sanguiacuteneo No intestino do inseto estas formas se diferenciam em formas epimastigotas (De
Souza et al 2002) que apoacutes multiplicaccedilatildeo por fissatildeo binaacuteria colonizam a porccedilatildeo terminal do
intestino e se diferenciam novamente em tripomastigotas metaciacuteclicos Quando o inseto
infectado pica o hospedeiro vertebrado e inicia o repasto sanguiacuteneo concomitantemente
elimina as formas tripomastigotas metaciacuteclicos junto com as fezes e urina Uma vez em
contato com a pele ou mucosas do hospedeiro mamiacutefero os tripomastigotas satildeo internalizados
atraveacutes de pequenas lesotildees na pele causadas pelo proacuteprio hospedeiro (Alves Colli 2008) Os
tripomastigotas invadem as ceacutelulas do hospedeiro mamiacutefero e se diferenciam em formas
replicativas denominadas amastigotas estabelecendo assim a infecccedilatildeo Apoacutes
aproximadamente 96 horas os amastigotas comeccedilam a se diferenciar em tripomastigotas
passando pela forma intermediaacuteria epimastigota intracelular Estas formas tripomastigotas satildeo
liberadas na corrente sanguiacutenea podendo invadir novas ceacutelulas ou serem ingeridas pelo inseto
vetor em um novo repasto sanguiacuteneo fechando assim o seu ciclo de vida O esquema do ciclo
de vida do Trypanosoma cruzi estaacute representado na Figura 1
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi
20
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos
T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono As formas epimastigotas preferencialmente catabolizam glicose e apoacutes a exaustatildeo
do carboidrato ocorre uma mudanccedila para o metabolismo de aminoaacutecidos A prolina e a
glutamina satildeo os principais aminoaacutecidos metabolizados pelas formas epimastigotas visto que
satildeo abudantes na hemolinfa e nos fluiacutedos dos tecidos do inseto vetor Esse metabolismo
ocorre com produccedilatildeo de NH4+
(produto da deaminaccedilatildeo dos aminoaacutecidos) que eacute excretada para
o meio extracelular (Bringaud et al 2006 Cazzulo 1994) As formas tripomastigotas
utilizam principalmente a glicose que eacute abundante nos fluiacutedos de seus hospedeiros
vertebrados Jaacute as formas amastigotas encontradas no citoplasma das ceacutelulas do hospedeiro
mamiacutefero ambiente pobre em glicose livre usam os aminoaacutecidos como principal fonte de
carbono e energia apresentando maior quantidade de prolina intracelular do que as formas
tripomastigotas (Silber et al 2009)
Entre os aminoaacutecidos metabolizados pelas formas epimastigotas estatildeo prolina glicina
asparagina glutamina aspartato glutamato leucina e isoleucina (Mancilla et al 1967 Odaly
et al 1983 Silber et al 2005a Sylvester Krassner 1976 Zeledon 1960) Os aminoaacutecidos
estatildeo envolvidos tambeacutem em outros processos importantes do ciclo de vida do T cruzi O
aminoaacutecido L-prolina eacute essencial na diferenciaccedilatildeo das formas epimastigota intracelular para
tripomastigotas aleacutem de estarem envolvidos nos processos de osmorregulaccedilatildeo crescimento e
resistecircncia do parasita ao estresse oxidativo e nutricional (Homsy et al 1989 Magdaleno et
al 2009 Magdaleno et al 2011 Pereira et al 2002 Pereira et al 2003 Tonelli et al
2004) Glutamato prolina e aspartato satildeo importantes na metaciclogecircnese ou seja na
diferenciaccedilatildeo de formas epimastigotas para tripomastigotas metaciacuteclicos (Contreras et al
1985) Prolina glicina alanina e glutamato satildeo descritos como importantes nos processos de
osmorregulaccedilatildeo e controle do volume celular (Rohloff et al 2003 Rohloff et al 2004a)
Os aminoaacutecidos para que sejam metabolizados ou cumprirem outras funccedilotildees devem
ser obtidos a partir da degradaccedilatildeo de proteiacutenas sintetizados ou incorporados pelo parasita
atraveacutes de transportadores localizados na membrana (Silber et al 2005a) O primeiro
transportador de aminoaacutecidos em T cruzi foi descrito bioquimicamente em 1970 o
transportador de lisina (Hampton 1970) Outros aminoaacutecidos tambeacutem jaacute tiveram a sua
cineacutetica e seu mecanismo de transporte descrito para T cruzi A incorporaccedilatildeo de arginina
pelo parasita apresenta pelo menos dois sistemas de transporte sendo um de alta afinidade e
outro de baixa afinidade Esse transporte eacute regulado ao longo do ciclo de vida do parasita
21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

FLAacuteVIA SILVA DAMASCENO
Estudo da interferecircncia de faacutermacos que atuam via
receptores eou transportadores de GABA e Glutamato
no ciclo de vida do Trypanosoma cruzi
Dissertaccedilatildeo apresentada ao Programa de Poacutes-
Graduaccedilatildeo em Biologia da Relaccedilatildeo Patoacutegeno-
Hospedeiro do Instituto de Ciecircncias Biomeacutedicas da
Universidade de Satildeo Paulo para obtenccedilatildeo do Tiacutetulo
de Mestre em Ciecircncias
Aacuterea de concentraccedilatildeo Biologia da Relaccedilatildeo
Patoacutegeno-Hospedeiro
Orientador Prof Dr Ariel Mariano Silber
Versatildeo corrigida A versatildeo original eletrocircnica
encontra-se disponiacutevel tanto na Biblioteca do ICB
quanto na Biblioteca Digital de Teses e Dissertaccedilotildees
da USP (BDTD)
Satildeo Paulo
2013
Trabalho realizado com auxiacutelios financeiros
concedido pela Fundaccedilatildeo de Amparo a
Pesquisa do Estado de Satildeo Paulo
(FAPESP) Instituto Nacional de
Biotecnologia Estrutural e Quiacutemica
Medicinal em Doenccedilas Infecciosas
(INBEQMeDI) e Conselho Nacional de
Desenvolvimento Cientiacutefico e Tecnoloacutegico
(CNPq)
Aos meus Pais Flamarion e Maria Nasareacute pelo
apoio amor e compreensatildeo incondicionais
AGRADECIMENTOS
Ao Prof Dr Ariel Mariano Silber pela grande oportunidade que me concedeu pela
orientaccedilatildeo paciecircncia apoio e amizade Obrigada por tudo
Agrave Elizabeth Pral teacutecnica do laboratoacuterio pelo apoio amizade ajuda e todo o suporte teacutecnico
Agrave minha amiga Mariacutea Julia Bariacuteson pela amizade apoio ajuda nos experimentos e nas
discussotildees dos resultados
Agrave minha famiacutelia meus pais Flamarion e Maria Nasareacute meus irmatildeos(atildes) Gilberto Gilmar
Gilson Gilsa Gildo Gildete e Jacoacute meus sobrinhos(as) Mariane Mateus Gisely Ceciacutelia
Nataacutelia Isabele Laviacutenia Maria e Venacircncio pelo apoio compreensatildeo e incentivo em todos os
momentos
Agrave minha amiga Valnia Pita por me fazer acreditar que era possiacutevel a realizaccedilatildeo desse sonho e
pelo apoio e estiacutemulo nessa caminhada
Aos meus amigos(as) do laboratoacuterio Raiacutessa Melo Marcell Crispim Sandra Carla Higo
Fernando Inga Eva Jean Douglas Carolina Manchola Ludmila Nakamura Brian Suarez
Rossana Duque e Letiacutecia Marchese pelo apoio ajuda e amizade
Ao Prof Dr Carsten Wrenger e seu grupo Flaacutevia Zimbres Kamila Meissner Jasmin
Lindner Thales Kronenberger
A Dra Lisvane Paes pelo apoio amizade e colaboraccedilatildeo nesse trabalho
Ao Prof Dr Joseacute Roberto Meyer-Fernandes (UFRJ) e aos integrantes do seu laboratoacuterio pela
disponibilidade e ajuda em alguns experimentos
Agrave Profa Dra Alicia J Kowaltowski do Instituto de Quiacutemica e aos integrantes do seu
laboratoacuterio pela disponibilidade do uso de equipamentos
Ao Prof Dr Gerhard Wunderlich e integrantes do seu laboratoacuterio em especial ao doutorando
Wesley Fotoran pela disponibilidade do uso de equipamentos e ajuda em alguns
experimentos
Agrave Fundaccedilatildeo de Amparo a Pesquisa do Estado de Satildeo Paulo (FAPESP) pelo suporte
financeiro
Ao Conselho Nacional de Desenvolvimento Cientiacutefico e Tecnoloacutegico (CNPq) pelo suporte
financeiro no primeiro ano desse trabalho
Agrave todos os professores funcionaacuterios e colegas do Departamento de Parasitologia (ICB-USP)
pelo apoio e convivecircncia
ldquoNunca deixe que lhe digam
que natildeo vale a pena
acreditar no sonho que se tem
ou que seus planos nunca vatildeo dar certo
ou que vocecirc nunca vai ser algueacutem ()
Confie em si mesmordquo
Renato Russo e Flaacutevio Venturini 1987
RESUMO
Damasceno FS Estudo da interferecircncia de faacutermacos que atuam via receptores eou
transportadores de GABA e Glutamato no ciclo de vida do Trypanosoma cruzi [dissertaccedilatildeo
(Mestrado em Biologia da Relaccedilatildeo Patoacutegeno-Hospedeiro)] Satildeo Paulo Instituto de Ciecircncias
Biomeacutedicas Universidade de Satildeo Paulo 2013
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica central e do Sul A quimioterapia disponiacutevel atualmente
limita-se a dois compostos Nifurtimox e o Benzonidazol Ambos os faacutermacos reduzem os
sintomas da doenccedila e a mortalidade das pessoas infectadas quando utilizadas na fase aguda
mas sua eficaacutecia na fase crocircnica eacute controversa Aleacutem do mais apresentam vaacuterios efeitos
colaterais O T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico como tambeacutem a importacircncia do aminoaacutecido
GABA no metabolismo do T cruzi visto que este tambeacutem eacute transportado pelo parasita Nesse
contexto o objetivo desse trabalho foi avaliar a interferecircncia de faacutermacos que atuam via
receptores ou transportadores de GABA e glutamato no ciclo de vida do T cruzi Os
faacutermacos selecionados foram Vigabatrina Pregabalina MK-801 MTEP e Memantina Os
resultados demonstram que apenas Memantina apresentou um melhor efeito tripanocida
Inibiu a proliferaccedilatildeo de formas epimastigotas interferiu na metaciclogecircnese como tambeacutem
afetou o metabolismo energeacutetico do parasita com a diminuiccedilatildeo dos niacuteveis de ATP aleacutem de
desencadear mecanismos que levam a apoptose das formas epimastigotas Memantina
tambeacutem interferiu no ciclo intracelular do parasita mais especificamente no estaacutegio
amastigota Interessantemente as fases do ciclo do parasita que satildeo mais afetadas satildeo as fases
que requerem mais energia estaacutegios replicativos (amastigota e epimastigota) como tambeacutem
os processos de diferenciaccedilatildeo e invasatildeo celular Apesar de apresentar valores de IC50 que
podem ser considerados altos em relaccedilatildeo aos valores descritos para formas epimastigotas
tratadas com Benzonidazol esses resultados mostram perspectivas promissoras visto que o
Memantina poderaacute ser utilizado como composto liacuteder para o desenho de derivados com
atividade tripanocida otimizada
Palavras-chave Trypanosoma cruzi Memantina Receptor de glutamato
ABSTRACT
Damasceno FS Study of interference of drugs that act via receptors andor transport of
GABA and glutamate in the life cycle of Trypanosoma cruzi [Masters thesis (Host-Pathogen
Relationship Biology)] Satildeo Paulo Instituto de Ciecircncias Biomeacutedicas Universidade de Satildeo
Paulo 2013
Chagas disease also known as American trypanosomiasis is caused by the flagellate
protozoan Trypanosoma cruzi and affects approximately 10 million people in endemic areas
of Mexico Central America and South America with about 25 million people living in areas
at risk of infection The currently available chemotherapy is limited to two compounds which
are Nifurtimox and Benznidazole Both drugs reduce symptoms of the disease and mortality
of infected people when used in the acute phase but its efficacy in chronic phase (phase in
which the majority of cases are diagnosed) is still controversial Moreover these drugs have
several side effects T cruzi is able to utilize carbohydrate and amino acids as carbon and
energy source Our group has studied the involvement of glutamate in parasite resistance to
thermal oxidative and metabolic stress as well as the importance of the amino acid GABA
metabolism in T cruzi The aim of this study was to evaluate the interference of drugs that
act in GABA and glutamate transporters or receptors in the life cycle of T cruzi The drugs
selected were vigabatrin pregabalin MK-801 memantine and MTEP The results showed
that only memantine has a better trypanocidal effect Memantine inhibited the proliferation of
epimastigotes interfered in metacyclogenesis and affected the energy metabolism of the
parasite with decreased levels of ATP and trigger mechanisms that lead to apoptosis of
epimastigotes Moreover interferes with intracellular cycle of the parasite specifically in
amastigote stage Interesting that stages of the parasite that require more energy are more
affected replicative stages (epimastigote and amastigote) as well as the processes of
differentiation and cell invasion IC50 value is more than that described to epimastigotes
treateds with Benznidazole but Memantine could be used as leader compound for the design
of drugs with optimized trypanocidal activity
Keywords Trypanosoma cruzi Memantine Glutamate receptor
LISTA DE ILUSTRACcedilOtildeES
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi 19
Figura 2 - Estruturas quiacutemicas do Benzonidazol e Nifurtimox 24
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica 28
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica 29
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse 40
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os
tratamentos com MK-801 ou Memantina 41
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina 42
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico ou oxidativo 44
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na
presenccedila do Memantina 45
Figura 10 - Efeito da Memantina na metaciclogecircnese 46
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo
de morte celular 48
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2 49
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
50
Figura 14 - Formas epimastigotas de T cruzi 51
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi 53
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade
de formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo
celular 54
LISTA DE ABREVIATURAS
ANOVA Anaacutelise de variacircncia
ATP Adenosina trifosfato
CHO-K1 Chinese Hamster Ovary (linhagem celular proveniente de ovaacuterio de
hamester chinecircs)
Ca2+
Caacutelcio
CO2 Dioacutexido de carbono
DNA Aacutecido desoxirribonucleacuteico
DO Densidade oacutetica
DTUs Discrete Typing Units
EGTA Aacutecido etilenoglicol-bis-aminoetiletertetraceacutetico
EROs Espeacutecies reativas de oxigecircnio
GABA Aminoaacutecido γ-aminobutiacuterico
GABA T Aminoaacutecido γ-aminobutiacuterico transaminase
GAPDH Gliceraldeiacutedo -3-fosfato desidrogenase
H2O2 Peroacutexido de hidrogecircnio
IC50 Metade da concentraccedilatildeo maacutexima inibitoacuteria
KCl Cloreto de potaacutessio
LIT Liver Infusion-Tryptose (Meio de cultura)
MCP Morte celular programada
mg Miligramas
mGluR Receptor metabotroacutepico de glutamato
MTT (3-(45-dimetiltiazol-2-il)-25 difenil bromo tetrazolio)
Na2HPO4 Fosfato de soacutedio monobaacutesico anidro
NaCl Cloreto de soacutedio
NaHCO3 Bicarbonato de soacutedio
NCCD Nomenclature Comittee on Cell Death (Comite de Nomeclatura de Morte
Celular)
NH4+
Ion amocircnio
NMDA N-metil-D-aspartato
OH Radical hidroxila
PBS Soluccedilatildeo salina tamponada com fosfato
pH Potencial hidrogeniocircnico
RPMI Roswell Park Memorial Institute
SDS Soacutedio dodecil sulfat
SFB Soro fetal bovino
U Unidades
μg Micrograma
μl Microlitro
μm Microcircmetro
SUMAacuteRIO
1 INTRODUCcedilAtildeO 16
11 Aspectos gerais da doenccedila de chagas 16
12 Trypanosoma cruzi 18
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos 20
14 Morte celular em Trypanosoma cruzi 21
15 Terapia na doenccedila de Chagas 23
16 Perspectivas de novos faacutermacos 25
2 OBJETIVOS 30
21 Objetivo geral 30
22 Objetivos especiacuteficos 30
3 MATERIAIS E MEacuteTODOS 31
31 Microorganismos utilizados no trabalho 31
311 Ceacutelulas hospedeiras 31
312 Trypanosoma cruzi 31
32 Manutenccedilatildeo das ceacutelulas hospedeiras 31
33 Obtenccedilatildeo dos parasitas 31
331 Epimastigotas 31
332 Curvas de crescimento de epimastigotas de T cruzi 31
333 Formas tripomastigotas 32
34 Resistecircncia a diferentes tipos de estresse 32
341 Ensaio de estresse teacutermico em formas epimastigotas 32
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico 33
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular 33
343 Ensaio de estresse oxidativo em formas epimastigotas 33
35 Tipo de morte celular em parasitas tratados com Memantina 34
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita 34
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas
epimastigotas 35
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio 35
36 Efeito sobre a metaciclogecircnese 35
37 Anaacutelise do efeito dos compostos no ciclo intracelular do T cruzi 36
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos 36
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas 36
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas
tripomastigotas 37
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os
compostos 37
38 Anaacutelises estatiacutesticas 38
4 RESULTADOS 39
41 Curvas de crescimento do T cruzi na presenccedila dos compostos 39
42 Interaccedilatildeo com condiccedilotildees de estresse 41
43 Efeito da Memantina na metaciclogecircnese 46
44 Tipo de morte celular sofrida pelo parasita tratado com Memantina 47
45 Efeito da Memantina no ciclo intracelular do T cruzi 52
5 DISCUSSAtildeO 55
6 CONCLUSOtildeES 59
REFEREcircNCIAS 61
16
1 INTRODUCcedilAtildeO
11 Aspectos gerais da doenccedila de chagas
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana foi
descoberta pelo meacutedico brasileiro Carlos Chagas (Chagas 1909) Esta doenccedila eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica Central e do Sul com cerca de 25 milhotildees de pessoas
vivendo em aacutereas de risco de infecccedilatildeo Ocorrem por ano aproximadamente 40 mil novos
casos e 12 mil mortes devido a complicaccedilotildees cardiacuteacas e digestivas (World Health
Organization 2012) Programas de controle do vetor desenvolvidos nos uacuteltimos 20 anos
como tambeacutem um maior controle nos bancos de sangue tem reduzido o nuacutemero de novos
casos da doenccedila de Chagas na Ameacuterica Latina (Rassi Marin-Neto 2010) Devido as intensas
migraccedilotildees de pessoas de paiacuteses endecircmicos o nuacutemero de casos de indiviacuteduos infectados tem
aumentado em aacutereas consideradas natildeo endecircmicas tais como Estados Unidos Canadaacute e
algumas regiotildees da Europa (Bern et al 2009 Maguire 2006 Schmunis 2007) A doenccedila eacute
uma zoonose que tem como reservatoacuterio natural do T cruzi uma ampla variedade de
mamiacuteferos do continente americano (Urbina Docampo 2003) Eacute transmitida para os seres
humanos e para mais de 150 espeacutecies de animais domeacutesticos e animais selvagens
principalmente por insetos hematoacutefagos da famiacutelia Reduviidae Apesar de existirem mais de
130 espeacutecies de triatomiacuteneos satildeo trecircs as mais importantes na transmissatildeo do T cruzi para o
homem Triatoma infestans Rhodnius prolixus e Triatoma dimidiata (Rassi Marin-Neto
2010) O parasita tambeacutem pode ser transmitido por transfusatildeo de sangue congenitamente (de
matildees infectadas para receacutem-nascidos) por transplante de oacutergatildeos ou medula oacutessea Tambeacutem
pode ocorrer por acidentes em laboratoacuterios que trabalham com o parasita vivo via esta que
epidemiologicamente eacute pouco relevante Outra forma de transmissatildeo eacute por via oral atraveacutes de
alimentos contaminados essa via vecircm sendo considerada como relevante na manutenccedilatildeo do
ciclo de transmissatildeo natural (Pereira et al 2009 Rassi Marin-Neto 2010)
A evoluccedilatildeo cliacutenica da doenccedila de Chagas pode ser dividida em duas fases aguda e
crocircnica A fase aguda normalmente eacute assintomaacutetica ou apresenta sintomatologia discreta que
na maioria dos casos passa despercebida pelo paciente Os sintomas quando aparecem o
fazem uma ou duas semanas apoacutes a exposiccedilatildeo ao triatomiacuteneo infectado ou poucos meses apoacutes
a transfusatildeo com sangue contaminado (Rassi Marin-Neto 2010) Os sintomas mais
frequentemente apresentados pelos pacientes agudos satildeo febre prolongada mal-estar
17
distensatildeo do fiacutegado baccedilo e linfonodos Pode haver tambeacutem edema subcutacircneo localizado ou
generalizado No caso particular de transmissatildeo vetorial o paciente pode apresentar um sinal
da porta de entrada do T cruzi na pele (chagoma) ou via mucosa ocular (sinal de Romantildea) As
manifestaccedilotildees da fase aguda se resolvem espontaneamente em aproximadamente 90 dos
indiviacuteduos infectados mesmo sem o tratamento com drogas tripanocidas Essa fase eacute
caracterizada pela alta parasitemia e ausecircncia de anticorpos especiacuteficos Nessa fase os
parasitas podem ser encontrados em praticamente todos os tecidos estando majoritariamente
presentes no sangue Por ser uma fase de curta duraccedilatildeo sem sintomatologia aparente ou com
sintomatologia pouco definida haacute um pequeno nuacutemero de casos detectados sendo portanto a
fase menos estudada (Rassi Marin-Neto 2010 Silber et al 2005b)
Logo apoacutes a fase aguda tem iniacutecio a fase crocircnica caracterizada pelos altos tiacutetulos de
anticorpos no sangue do paciente e pelo fato de que a parasitemia eacute inaparente O paciente
pode permanecer por vaacuterios anos ou ateacute mesmo pelo resto da vida sem apresentar sinais ou
sintomas Essa forma da fase crocircnica eacute chamada de indeterminada Aproximadamente 30-40
dos pacientes crocircnicos indeterminados desenvolvem a sintomatologia caracteriacutestica da
doenccedila evoluindo para a chamada fase crocircnica sintomaacutetica Nesta fase as trecircs principais
formas satildeo a cardiacuteaca a digestiva e a cardiodigestiva (Acquatella 2007 Marin-Neto et al
1999) Na forma cardiacuteaca o coraccedilatildeo eacute um dos primeiros oacutergatildeos afetados natildeo necessariamente
o uacutenico desenvolvendo hipertrofia e dilataccedilatildeo Na forma digestiva o trato digestivo apresenta
alteraccedilotildees anatocircmicas e fisioloacutegicas importantes sendo a principal altereccedilatildeo a dilataccedilatildeo do
intestino e do esocircfago que podem apresentar aspecto de megaviacutescera (Marin-Neto et al
1999 Rassi et al 2000) revisado por (Boscardin et al 2009 Rassi Marin-Neto 2010)
18
12 Trypanosoma cruzi
Trypanosoma cruzi eacute um parasita intracelular obrigatoacuterio classicamente localizado na
ordem kinetoplastida gecircnero Trypanosoma que causa uma infecccedilatildeo crocircnica em humanos e
em grande nuacutemero de outras espeacutecies de mamiacuteferos (Hoare 1972) Recentemente devido agrave
alta variabilidade bioloacutegica bioquiacutemica e molecular a espeacutecie Trypanosoma cruzi foi
dividida em seis linhagens subsespeciacuteficas (Discrete Typing Units ndash DTUs) numerados de I a
VI Esta classificaccedilatildeo foi realizada com base em caracteriacutesticas geneacuteticas moleculares e
imunoloacutegicas com o objetivo de unificar a nomenclatura e facilitar a comunicaccedilatildeo dentro da
comunidade cientiacutefica (Zingales et al 2009)
O T cruzi tem um ciclo de vida complexo que alterna entre o inseto vetor e o
hospedeiro mamiacutefero O parasita apresenta diferentes formas correspondentes a diferentes
estaacutegios do seu ciclo de vida Estes diferentes estaacutegios tecircm sido descritos com base em
criteacuterios morfoloacutegicos como a forma fusiforme ou arredondados a regiatildeo onde emerge o
flagelo e a posiccedilatildeo do cinetoplasto (estrutura contendo o DNA da uacutenica mitococircndria presente
no parasita) em relaccedilatildeo ao nuacutecleo No hospedeiro mamiacutefero foram descritas as formas
amastigota epimastigota intracelular e tripomastigota sanguiacutecola A forma amastigota eacute um
estaacutegio replicativo e infectivo Satildeo ceacutelulas arredondadas medindo de 2-6 microm de diacircmetro
encontradas no citoplasma das ceacutelulas infectadas Os epimastigotas intracelulares
(epimastigote-like) satildeo formas replicativas intermediaacuterias entre amastigotas e tripomastigotas
tambeacutem encontrados dentro das ceacutelulas medindo aproximadamente 54 microm Essas formas satildeo
menores que as formas epimastigotas encontradas no inseto vetor poreacutem compartilha com as
mesmas padrotildees morfomeacutetricos e diversas caracteriacutesticas bioquiacutemicas e bioloacutegicas (Almeida-
De-Faria et al 1999) O tripomastigota sanguiacutecola eacute um estaacutegio flagelado infectivo natildeo
replicativo encontrado principalmente na corrente sanguiacutenea do hospedeiro mamiacutefero Mede
aproximadamente 18 microm de comprimento incluindo um extremo livre do flagelo de
aproximadamente 6 microm e 2-3 microm de largura
No inseto vetor encontram-se predominantemente as formas epimastigota e
tripomastigota metaciacuteclico Epimastigota eacute uma forma flagelada replicativa e natildeo infectiva
Mede aproximadamente 20-40 microm de comprimento e 2-5 microm de largura essas formas estatildeo
presentes tipicamente na porccedilatildeo meacutedia do intestino do hospedeiro invertebrado
Tripomastigota metaciacuteclico eacute uma forma infectiva natildeo replicativa encontrada na porccedilatildeo final
do intestino do inseto vetor (Brener 1971) revisado por (Silber et al 2005a Tyler Engman
2001)
19
O ciclo de vida do T cruzi inicia-se quando os parasitas presentes na corrente
sanguiacutenea do hospedeiro vertebrado infectado passam ao inseto vetor durante o repasto
sanguiacuteneo No intestino do inseto estas formas se diferenciam em formas epimastigotas (De
Souza et al 2002) que apoacutes multiplicaccedilatildeo por fissatildeo binaacuteria colonizam a porccedilatildeo terminal do
intestino e se diferenciam novamente em tripomastigotas metaciacuteclicos Quando o inseto
infectado pica o hospedeiro vertebrado e inicia o repasto sanguiacuteneo concomitantemente
elimina as formas tripomastigotas metaciacuteclicos junto com as fezes e urina Uma vez em
contato com a pele ou mucosas do hospedeiro mamiacutefero os tripomastigotas satildeo internalizados
atraveacutes de pequenas lesotildees na pele causadas pelo proacuteprio hospedeiro (Alves Colli 2008) Os
tripomastigotas invadem as ceacutelulas do hospedeiro mamiacutefero e se diferenciam em formas
replicativas denominadas amastigotas estabelecendo assim a infecccedilatildeo Apoacutes
aproximadamente 96 horas os amastigotas comeccedilam a se diferenciar em tripomastigotas
passando pela forma intermediaacuteria epimastigota intracelular Estas formas tripomastigotas satildeo
liberadas na corrente sanguiacutenea podendo invadir novas ceacutelulas ou serem ingeridas pelo inseto
vetor em um novo repasto sanguiacuteneo fechando assim o seu ciclo de vida O esquema do ciclo
de vida do Trypanosoma cruzi estaacute representado na Figura 1
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi
20
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos
T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono As formas epimastigotas preferencialmente catabolizam glicose e apoacutes a exaustatildeo
do carboidrato ocorre uma mudanccedila para o metabolismo de aminoaacutecidos A prolina e a
glutamina satildeo os principais aminoaacutecidos metabolizados pelas formas epimastigotas visto que
satildeo abudantes na hemolinfa e nos fluiacutedos dos tecidos do inseto vetor Esse metabolismo
ocorre com produccedilatildeo de NH4+
(produto da deaminaccedilatildeo dos aminoaacutecidos) que eacute excretada para
o meio extracelular (Bringaud et al 2006 Cazzulo 1994) As formas tripomastigotas
utilizam principalmente a glicose que eacute abundante nos fluiacutedos de seus hospedeiros
vertebrados Jaacute as formas amastigotas encontradas no citoplasma das ceacutelulas do hospedeiro
mamiacutefero ambiente pobre em glicose livre usam os aminoaacutecidos como principal fonte de
carbono e energia apresentando maior quantidade de prolina intracelular do que as formas
tripomastigotas (Silber et al 2009)
Entre os aminoaacutecidos metabolizados pelas formas epimastigotas estatildeo prolina glicina
asparagina glutamina aspartato glutamato leucina e isoleucina (Mancilla et al 1967 Odaly
et al 1983 Silber et al 2005a Sylvester Krassner 1976 Zeledon 1960) Os aminoaacutecidos
estatildeo envolvidos tambeacutem em outros processos importantes do ciclo de vida do T cruzi O
aminoaacutecido L-prolina eacute essencial na diferenciaccedilatildeo das formas epimastigota intracelular para
tripomastigotas aleacutem de estarem envolvidos nos processos de osmorregulaccedilatildeo crescimento e
resistecircncia do parasita ao estresse oxidativo e nutricional (Homsy et al 1989 Magdaleno et
al 2009 Magdaleno et al 2011 Pereira et al 2002 Pereira et al 2003 Tonelli et al
2004) Glutamato prolina e aspartato satildeo importantes na metaciclogecircnese ou seja na
diferenciaccedilatildeo de formas epimastigotas para tripomastigotas metaciacuteclicos (Contreras et al
1985) Prolina glicina alanina e glutamato satildeo descritos como importantes nos processos de
osmorregulaccedilatildeo e controle do volume celular (Rohloff et al 2003 Rohloff et al 2004a)
Os aminoaacutecidos para que sejam metabolizados ou cumprirem outras funccedilotildees devem
ser obtidos a partir da degradaccedilatildeo de proteiacutenas sintetizados ou incorporados pelo parasita
atraveacutes de transportadores localizados na membrana (Silber et al 2005a) O primeiro
transportador de aminoaacutecidos em T cruzi foi descrito bioquimicamente em 1970 o
transportador de lisina (Hampton 1970) Outros aminoaacutecidos tambeacutem jaacute tiveram a sua
cineacutetica e seu mecanismo de transporte descrito para T cruzi A incorporaccedilatildeo de arginina
pelo parasita apresenta pelo menos dois sistemas de transporte sendo um de alta afinidade e
outro de baixa afinidade Esse transporte eacute regulado ao longo do ciclo de vida do parasita
21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

Trabalho realizado com auxiacutelios financeiros
concedido pela Fundaccedilatildeo de Amparo a
Pesquisa do Estado de Satildeo Paulo
(FAPESP) Instituto Nacional de
Biotecnologia Estrutural e Quiacutemica
Medicinal em Doenccedilas Infecciosas
(INBEQMeDI) e Conselho Nacional de
Desenvolvimento Cientiacutefico e Tecnoloacutegico
(CNPq)
Aos meus Pais Flamarion e Maria Nasareacute pelo
apoio amor e compreensatildeo incondicionais
AGRADECIMENTOS
Ao Prof Dr Ariel Mariano Silber pela grande oportunidade que me concedeu pela
orientaccedilatildeo paciecircncia apoio e amizade Obrigada por tudo
Agrave Elizabeth Pral teacutecnica do laboratoacuterio pelo apoio amizade ajuda e todo o suporte teacutecnico
Agrave minha amiga Mariacutea Julia Bariacuteson pela amizade apoio ajuda nos experimentos e nas
discussotildees dos resultados
Agrave minha famiacutelia meus pais Flamarion e Maria Nasareacute meus irmatildeos(atildes) Gilberto Gilmar
Gilson Gilsa Gildo Gildete e Jacoacute meus sobrinhos(as) Mariane Mateus Gisely Ceciacutelia
Nataacutelia Isabele Laviacutenia Maria e Venacircncio pelo apoio compreensatildeo e incentivo em todos os
momentos
Agrave minha amiga Valnia Pita por me fazer acreditar que era possiacutevel a realizaccedilatildeo desse sonho e
pelo apoio e estiacutemulo nessa caminhada
Aos meus amigos(as) do laboratoacuterio Raiacutessa Melo Marcell Crispim Sandra Carla Higo
Fernando Inga Eva Jean Douglas Carolina Manchola Ludmila Nakamura Brian Suarez
Rossana Duque e Letiacutecia Marchese pelo apoio ajuda e amizade
Ao Prof Dr Carsten Wrenger e seu grupo Flaacutevia Zimbres Kamila Meissner Jasmin
Lindner Thales Kronenberger
A Dra Lisvane Paes pelo apoio amizade e colaboraccedilatildeo nesse trabalho
Ao Prof Dr Joseacute Roberto Meyer-Fernandes (UFRJ) e aos integrantes do seu laboratoacuterio pela
disponibilidade e ajuda em alguns experimentos
Agrave Profa Dra Alicia J Kowaltowski do Instituto de Quiacutemica e aos integrantes do seu
laboratoacuterio pela disponibilidade do uso de equipamentos
Ao Prof Dr Gerhard Wunderlich e integrantes do seu laboratoacuterio em especial ao doutorando
Wesley Fotoran pela disponibilidade do uso de equipamentos e ajuda em alguns
experimentos
Agrave Fundaccedilatildeo de Amparo a Pesquisa do Estado de Satildeo Paulo (FAPESP) pelo suporte
financeiro
Ao Conselho Nacional de Desenvolvimento Cientiacutefico e Tecnoloacutegico (CNPq) pelo suporte
financeiro no primeiro ano desse trabalho
Agrave todos os professores funcionaacuterios e colegas do Departamento de Parasitologia (ICB-USP)
pelo apoio e convivecircncia
ldquoNunca deixe que lhe digam
que natildeo vale a pena
acreditar no sonho que se tem
ou que seus planos nunca vatildeo dar certo
ou que vocecirc nunca vai ser algueacutem ()
Confie em si mesmordquo
Renato Russo e Flaacutevio Venturini 1987
RESUMO
Damasceno FS Estudo da interferecircncia de faacutermacos que atuam via receptores eou
transportadores de GABA e Glutamato no ciclo de vida do Trypanosoma cruzi [dissertaccedilatildeo
(Mestrado em Biologia da Relaccedilatildeo Patoacutegeno-Hospedeiro)] Satildeo Paulo Instituto de Ciecircncias
Biomeacutedicas Universidade de Satildeo Paulo 2013
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica central e do Sul A quimioterapia disponiacutevel atualmente
limita-se a dois compostos Nifurtimox e o Benzonidazol Ambos os faacutermacos reduzem os
sintomas da doenccedila e a mortalidade das pessoas infectadas quando utilizadas na fase aguda
mas sua eficaacutecia na fase crocircnica eacute controversa Aleacutem do mais apresentam vaacuterios efeitos
colaterais O T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico como tambeacutem a importacircncia do aminoaacutecido
GABA no metabolismo do T cruzi visto que este tambeacutem eacute transportado pelo parasita Nesse
contexto o objetivo desse trabalho foi avaliar a interferecircncia de faacutermacos que atuam via
receptores ou transportadores de GABA e glutamato no ciclo de vida do T cruzi Os
faacutermacos selecionados foram Vigabatrina Pregabalina MK-801 MTEP e Memantina Os
resultados demonstram que apenas Memantina apresentou um melhor efeito tripanocida
Inibiu a proliferaccedilatildeo de formas epimastigotas interferiu na metaciclogecircnese como tambeacutem
afetou o metabolismo energeacutetico do parasita com a diminuiccedilatildeo dos niacuteveis de ATP aleacutem de
desencadear mecanismos que levam a apoptose das formas epimastigotas Memantina
tambeacutem interferiu no ciclo intracelular do parasita mais especificamente no estaacutegio
amastigota Interessantemente as fases do ciclo do parasita que satildeo mais afetadas satildeo as fases
que requerem mais energia estaacutegios replicativos (amastigota e epimastigota) como tambeacutem
os processos de diferenciaccedilatildeo e invasatildeo celular Apesar de apresentar valores de IC50 que
podem ser considerados altos em relaccedilatildeo aos valores descritos para formas epimastigotas
tratadas com Benzonidazol esses resultados mostram perspectivas promissoras visto que o
Memantina poderaacute ser utilizado como composto liacuteder para o desenho de derivados com
atividade tripanocida otimizada
Palavras-chave Trypanosoma cruzi Memantina Receptor de glutamato
ABSTRACT
Damasceno FS Study of interference of drugs that act via receptors andor transport of
GABA and glutamate in the life cycle of Trypanosoma cruzi [Masters thesis (Host-Pathogen
Relationship Biology)] Satildeo Paulo Instituto de Ciecircncias Biomeacutedicas Universidade de Satildeo
Paulo 2013
Chagas disease also known as American trypanosomiasis is caused by the flagellate
protozoan Trypanosoma cruzi and affects approximately 10 million people in endemic areas
of Mexico Central America and South America with about 25 million people living in areas
at risk of infection The currently available chemotherapy is limited to two compounds which
are Nifurtimox and Benznidazole Both drugs reduce symptoms of the disease and mortality
of infected people when used in the acute phase but its efficacy in chronic phase (phase in
which the majority of cases are diagnosed) is still controversial Moreover these drugs have
several side effects T cruzi is able to utilize carbohydrate and amino acids as carbon and
energy source Our group has studied the involvement of glutamate in parasite resistance to
thermal oxidative and metabolic stress as well as the importance of the amino acid GABA
metabolism in T cruzi The aim of this study was to evaluate the interference of drugs that
act in GABA and glutamate transporters or receptors in the life cycle of T cruzi The drugs
selected were vigabatrin pregabalin MK-801 memantine and MTEP The results showed
that only memantine has a better trypanocidal effect Memantine inhibited the proliferation of
epimastigotes interfered in metacyclogenesis and affected the energy metabolism of the
parasite with decreased levels of ATP and trigger mechanisms that lead to apoptosis of
epimastigotes Moreover interferes with intracellular cycle of the parasite specifically in
amastigote stage Interesting that stages of the parasite that require more energy are more
affected replicative stages (epimastigote and amastigote) as well as the processes of
differentiation and cell invasion IC50 value is more than that described to epimastigotes
treateds with Benznidazole but Memantine could be used as leader compound for the design
of drugs with optimized trypanocidal activity
Keywords Trypanosoma cruzi Memantine Glutamate receptor
LISTA DE ILUSTRACcedilOtildeES
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi 19
Figura 2 - Estruturas quiacutemicas do Benzonidazol e Nifurtimox 24
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica 28
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica 29
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse 40
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os
tratamentos com MK-801 ou Memantina 41
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina 42
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico ou oxidativo 44
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na
presenccedila do Memantina 45
Figura 10 - Efeito da Memantina na metaciclogecircnese 46
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo
de morte celular 48
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2 49
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
50
Figura 14 - Formas epimastigotas de T cruzi 51
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi 53
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade
de formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo
celular 54
LISTA DE ABREVIATURAS
ANOVA Anaacutelise de variacircncia
ATP Adenosina trifosfato
CHO-K1 Chinese Hamster Ovary (linhagem celular proveniente de ovaacuterio de
hamester chinecircs)
Ca2+
Caacutelcio
CO2 Dioacutexido de carbono
DNA Aacutecido desoxirribonucleacuteico
DO Densidade oacutetica
DTUs Discrete Typing Units
EGTA Aacutecido etilenoglicol-bis-aminoetiletertetraceacutetico
EROs Espeacutecies reativas de oxigecircnio
GABA Aminoaacutecido γ-aminobutiacuterico
GABA T Aminoaacutecido γ-aminobutiacuterico transaminase
GAPDH Gliceraldeiacutedo -3-fosfato desidrogenase
H2O2 Peroacutexido de hidrogecircnio
IC50 Metade da concentraccedilatildeo maacutexima inibitoacuteria
KCl Cloreto de potaacutessio
LIT Liver Infusion-Tryptose (Meio de cultura)
MCP Morte celular programada
mg Miligramas
mGluR Receptor metabotroacutepico de glutamato
MTT (3-(45-dimetiltiazol-2-il)-25 difenil bromo tetrazolio)
Na2HPO4 Fosfato de soacutedio monobaacutesico anidro
NaCl Cloreto de soacutedio
NaHCO3 Bicarbonato de soacutedio
NCCD Nomenclature Comittee on Cell Death (Comite de Nomeclatura de Morte
Celular)
NH4+
Ion amocircnio
NMDA N-metil-D-aspartato
OH Radical hidroxila
PBS Soluccedilatildeo salina tamponada com fosfato
pH Potencial hidrogeniocircnico
RPMI Roswell Park Memorial Institute
SDS Soacutedio dodecil sulfat
SFB Soro fetal bovino
U Unidades
μg Micrograma
μl Microlitro
μm Microcircmetro
SUMAacuteRIO
1 INTRODUCcedilAtildeO 16
11 Aspectos gerais da doenccedila de chagas 16
12 Trypanosoma cruzi 18
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos 20
14 Morte celular em Trypanosoma cruzi 21
15 Terapia na doenccedila de Chagas 23
16 Perspectivas de novos faacutermacos 25
2 OBJETIVOS 30
21 Objetivo geral 30
22 Objetivos especiacuteficos 30
3 MATERIAIS E MEacuteTODOS 31
31 Microorganismos utilizados no trabalho 31
311 Ceacutelulas hospedeiras 31
312 Trypanosoma cruzi 31
32 Manutenccedilatildeo das ceacutelulas hospedeiras 31
33 Obtenccedilatildeo dos parasitas 31
331 Epimastigotas 31
332 Curvas de crescimento de epimastigotas de T cruzi 31
333 Formas tripomastigotas 32
34 Resistecircncia a diferentes tipos de estresse 32
341 Ensaio de estresse teacutermico em formas epimastigotas 32
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico 33
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular 33
343 Ensaio de estresse oxidativo em formas epimastigotas 33
35 Tipo de morte celular em parasitas tratados com Memantina 34
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita 34
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas
epimastigotas 35
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio 35
36 Efeito sobre a metaciclogecircnese 35
37 Anaacutelise do efeito dos compostos no ciclo intracelular do T cruzi 36
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos 36
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas 36
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas
tripomastigotas 37
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os
compostos 37
38 Anaacutelises estatiacutesticas 38
4 RESULTADOS 39
41 Curvas de crescimento do T cruzi na presenccedila dos compostos 39
42 Interaccedilatildeo com condiccedilotildees de estresse 41
43 Efeito da Memantina na metaciclogecircnese 46
44 Tipo de morte celular sofrida pelo parasita tratado com Memantina 47
45 Efeito da Memantina no ciclo intracelular do T cruzi 52
5 DISCUSSAtildeO 55
6 CONCLUSOtildeES 59
REFEREcircNCIAS 61
16
1 INTRODUCcedilAtildeO
11 Aspectos gerais da doenccedila de chagas
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana foi
descoberta pelo meacutedico brasileiro Carlos Chagas (Chagas 1909) Esta doenccedila eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica Central e do Sul com cerca de 25 milhotildees de pessoas
vivendo em aacutereas de risco de infecccedilatildeo Ocorrem por ano aproximadamente 40 mil novos
casos e 12 mil mortes devido a complicaccedilotildees cardiacuteacas e digestivas (World Health
Organization 2012) Programas de controle do vetor desenvolvidos nos uacuteltimos 20 anos
como tambeacutem um maior controle nos bancos de sangue tem reduzido o nuacutemero de novos
casos da doenccedila de Chagas na Ameacuterica Latina (Rassi Marin-Neto 2010) Devido as intensas
migraccedilotildees de pessoas de paiacuteses endecircmicos o nuacutemero de casos de indiviacuteduos infectados tem
aumentado em aacutereas consideradas natildeo endecircmicas tais como Estados Unidos Canadaacute e
algumas regiotildees da Europa (Bern et al 2009 Maguire 2006 Schmunis 2007) A doenccedila eacute
uma zoonose que tem como reservatoacuterio natural do T cruzi uma ampla variedade de
mamiacuteferos do continente americano (Urbina Docampo 2003) Eacute transmitida para os seres
humanos e para mais de 150 espeacutecies de animais domeacutesticos e animais selvagens
principalmente por insetos hematoacutefagos da famiacutelia Reduviidae Apesar de existirem mais de
130 espeacutecies de triatomiacuteneos satildeo trecircs as mais importantes na transmissatildeo do T cruzi para o
homem Triatoma infestans Rhodnius prolixus e Triatoma dimidiata (Rassi Marin-Neto
2010) O parasita tambeacutem pode ser transmitido por transfusatildeo de sangue congenitamente (de
matildees infectadas para receacutem-nascidos) por transplante de oacutergatildeos ou medula oacutessea Tambeacutem
pode ocorrer por acidentes em laboratoacuterios que trabalham com o parasita vivo via esta que
epidemiologicamente eacute pouco relevante Outra forma de transmissatildeo eacute por via oral atraveacutes de
alimentos contaminados essa via vecircm sendo considerada como relevante na manutenccedilatildeo do
ciclo de transmissatildeo natural (Pereira et al 2009 Rassi Marin-Neto 2010)
A evoluccedilatildeo cliacutenica da doenccedila de Chagas pode ser dividida em duas fases aguda e
crocircnica A fase aguda normalmente eacute assintomaacutetica ou apresenta sintomatologia discreta que
na maioria dos casos passa despercebida pelo paciente Os sintomas quando aparecem o
fazem uma ou duas semanas apoacutes a exposiccedilatildeo ao triatomiacuteneo infectado ou poucos meses apoacutes
a transfusatildeo com sangue contaminado (Rassi Marin-Neto 2010) Os sintomas mais
frequentemente apresentados pelos pacientes agudos satildeo febre prolongada mal-estar
17
distensatildeo do fiacutegado baccedilo e linfonodos Pode haver tambeacutem edema subcutacircneo localizado ou
generalizado No caso particular de transmissatildeo vetorial o paciente pode apresentar um sinal
da porta de entrada do T cruzi na pele (chagoma) ou via mucosa ocular (sinal de Romantildea) As
manifestaccedilotildees da fase aguda se resolvem espontaneamente em aproximadamente 90 dos
indiviacuteduos infectados mesmo sem o tratamento com drogas tripanocidas Essa fase eacute
caracterizada pela alta parasitemia e ausecircncia de anticorpos especiacuteficos Nessa fase os
parasitas podem ser encontrados em praticamente todos os tecidos estando majoritariamente
presentes no sangue Por ser uma fase de curta duraccedilatildeo sem sintomatologia aparente ou com
sintomatologia pouco definida haacute um pequeno nuacutemero de casos detectados sendo portanto a
fase menos estudada (Rassi Marin-Neto 2010 Silber et al 2005b)
Logo apoacutes a fase aguda tem iniacutecio a fase crocircnica caracterizada pelos altos tiacutetulos de
anticorpos no sangue do paciente e pelo fato de que a parasitemia eacute inaparente O paciente
pode permanecer por vaacuterios anos ou ateacute mesmo pelo resto da vida sem apresentar sinais ou
sintomas Essa forma da fase crocircnica eacute chamada de indeterminada Aproximadamente 30-40
dos pacientes crocircnicos indeterminados desenvolvem a sintomatologia caracteriacutestica da
doenccedila evoluindo para a chamada fase crocircnica sintomaacutetica Nesta fase as trecircs principais
formas satildeo a cardiacuteaca a digestiva e a cardiodigestiva (Acquatella 2007 Marin-Neto et al
1999) Na forma cardiacuteaca o coraccedilatildeo eacute um dos primeiros oacutergatildeos afetados natildeo necessariamente
o uacutenico desenvolvendo hipertrofia e dilataccedilatildeo Na forma digestiva o trato digestivo apresenta
alteraccedilotildees anatocircmicas e fisioloacutegicas importantes sendo a principal altereccedilatildeo a dilataccedilatildeo do
intestino e do esocircfago que podem apresentar aspecto de megaviacutescera (Marin-Neto et al
1999 Rassi et al 2000) revisado por (Boscardin et al 2009 Rassi Marin-Neto 2010)
18
12 Trypanosoma cruzi
Trypanosoma cruzi eacute um parasita intracelular obrigatoacuterio classicamente localizado na
ordem kinetoplastida gecircnero Trypanosoma que causa uma infecccedilatildeo crocircnica em humanos e
em grande nuacutemero de outras espeacutecies de mamiacuteferos (Hoare 1972) Recentemente devido agrave
alta variabilidade bioloacutegica bioquiacutemica e molecular a espeacutecie Trypanosoma cruzi foi
dividida em seis linhagens subsespeciacuteficas (Discrete Typing Units ndash DTUs) numerados de I a
VI Esta classificaccedilatildeo foi realizada com base em caracteriacutesticas geneacuteticas moleculares e
imunoloacutegicas com o objetivo de unificar a nomenclatura e facilitar a comunicaccedilatildeo dentro da
comunidade cientiacutefica (Zingales et al 2009)
O T cruzi tem um ciclo de vida complexo que alterna entre o inseto vetor e o
hospedeiro mamiacutefero O parasita apresenta diferentes formas correspondentes a diferentes
estaacutegios do seu ciclo de vida Estes diferentes estaacutegios tecircm sido descritos com base em
criteacuterios morfoloacutegicos como a forma fusiforme ou arredondados a regiatildeo onde emerge o
flagelo e a posiccedilatildeo do cinetoplasto (estrutura contendo o DNA da uacutenica mitococircndria presente
no parasita) em relaccedilatildeo ao nuacutecleo No hospedeiro mamiacutefero foram descritas as formas
amastigota epimastigota intracelular e tripomastigota sanguiacutecola A forma amastigota eacute um
estaacutegio replicativo e infectivo Satildeo ceacutelulas arredondadas medindo de 2-6 microm de diacircmetro
encontradas no citoplasma das ceacutelulas infectadas Os epimastigotas intracelulares
(epimastigote-like) satildeo formas replicativas intermediaacuterias entre amastigotas e tripomastigotas
tambeacutem encontrados dentro das ceacutelulas medindo aproximadamente 54 microm Essas formas satildeo
menores que as formas epimastigotas encontradas no inseto vetor poreacutem compartilha com as
mesmas padrotildees morfomeacutetricos e diversas caracteriacutesticas bioquiacutemicas e bioloacutegicas (Almeida-
De-Faria et al 1999) O tripomastigota sanguiacutecola eacute um estaacutegio flagelado infectivo natildeo
replicativo encontrado principalmente na corrente sanguiacutenea do hospedeiro mamiacutefero Mede
aproximadamente 18 microm de comprimento incluindo um extremo livre do flagelo de
aproximadamente 6 microm e 2-3 microm de largura
No inseto vetor encontram-se predominantemente as formas epimastigota e
tripomastigota metaciacuteclico Epimastigota eacute uma forma flagelada replicativa e natildeo infectiva
Mede aproximadamente 20-40 microm de comprimento e 2-5 microm de largura essas formas estatildeo
presentes tipicamente na porccedilatildeo meacutedia do intestino do hospedeiro invertebrado
Tripomastigota metaciacuteclico eacute uma forma infectiva natildeo replicativa encontrada na porccedilatildeo final
do intestino do inseto vetor (Brener 1971) revisado por (Silber et al 2005a Tyler Engman
2001)
19
O ciclo de vida do T cruzi inicia-se quando os parasitas presentes na corrente
sanguiacutenea do hospedeiro vertebrado infectado passam ao inseto vetor durante o repasto
sanguiacuteneo No intestino do inseto estas formas se diferenciam em formas epimastigotas (De
Souza et al 2002) que apoacutes multiplicaccedilatildeo por fissatildeo binaacuteria colonizam a porccedilatildeo terminal do
intestino e se diferenciam novamente em tripomastigotas metaciacuteclicos Quando o inseto
infectado pica o hospedeiro vertebrado e inicia o repasto sanguiacuteneo concomitantemente
elimina as formas tripomastigotas metaciacuteclicos junto com as fezes e urina Uma vez em
contato com a pele ou mucosas do hospedeiro mamiacutefero os tripomastigotas satildeo internalizados
atraveacutes de pequenas lesotildees na pele causadas pelo proacuteprio hospedeiro (Alves Colli 2008) Os
tripomastigotas invadem as ceacutelulas do hospedeiro mamiacutefero e se diferenciam em formas
replicativas denominadas amastigotas estabelecendo assim a infecccedilatildeo Apoacutes
aproximadamente 96 horas os amastigotas comeccedilam a se diferenciar em tripomastigotas
passando pela forma intermediaacuteria epimastigota intracelular Estas formas tripomastigotas satildeo
liberadas na corrente sanguiacutenea podendo invadir novas ceacutelulas ou serem ingeridas pelo inseto
vetor em um novo repasto sanguiacuteneo fechando assim o seu ciclo de vida O esquema do ciclo
de vida do Trypanosoma cruzi estaacute representado na Figura 1
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi
20
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos
T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono As formas epimastigotas preferencialmente catabolizam glicose e apoacutes a exaustatildeo
do carboidrato ocorre uma mudanccedila para o metabolismo de aminoaacutecidos A prolina e a
glutamina satildeo os principais aminoaacutecidos metabolizados pelas formas epimastigotas visto que
satildeo abudantes na hemolinfa e nos fluiacutedos dos tecidos do inseto vetor Esse metabolismo
ocorre com produccedilatildeo de NH4+
(produto da deaminaccedilatildeo dos aminoaacutecidos) que eacute excretada para
o meio extracelular (Bringaud et al 2006 Cazzulo 1994) As formas tripomastigotas
utilizam principalmente a glicose que eacute abundante nos fluiacutedos de seus hospedeiros
vertebrados Jaacute as formas amastigotas encontradas no citoplasma das ceacutelulas do hospedeiro
mamiacutefero ambiente pobre em glicose livre usam os aminoaacutecidos como principal fonte de
carbono e energia apresentando maior quantidade de prolina intracelular do que as formas
tripomastigotas (Silber et al 2009)
Entre os aminoaacutecidos metabolizados pelas formas epimastigotas estatildeo prolina glicina
asparagina glutamina aspartato glutamato leucina e isoleucina (Mancilla et al 1967 Odaly
et al 1983 Silber et al 2005a Sylvester Krassner 1976 Zeledon 1960) Os aminoaacutecidos
estatildeo envolvidos tambeacutem em outros processos importantes do ciclo de vida do T cruzi O
aminoaacutecido L-prolina eacute essencial na diferenciaccedilatildeo das formas epimastigota intracelular para
tripomastigotas aleacutem de estarem envolvidos nos processos de osmorregulaccedilatildeo crescimento e
resistecircncia do parasita ao estresse oxidativo e nutricional (Homsy et al 1989 Magdaleno et
al 2009 Magdaleno et al 2011 Pereira et al 2002 Pereira et al 2003 Tonelli et al
2004) Glutamato prolina e aspartato satildeo importantes na metaciclogecircnese ou seja na
diferenciaccedilatildeo de formas epimastigotas para tripomastigotas metaciacuteclicos (Contreras et al
1985) Prolina glicina alanina e glutamato satildeo descritos como importantes nos processos de
osmorregulaccedilatildeo e controle do volume celular (Rohloff et al 2003 Rohloff et al 2004a)
Os aminoaacutecidos para que sejam metabolizados ou cumprirem outras funccedilotildees devem
ser obtidos a partir da degradaccedilatildeo de proteiacutenas sintetizados ou incorporados pelo parasita
atraveacutes de transportadores localizados na membrana (Silber et al 2005a) O primeiro
transportador de aminoaacutecidos em T cruzi foi descrito bioquimicamente em 1970 o
transportador de lisina (Hampton 1970) Outros aminoaacutecidos tambeacutem jaacute tiveram a sua
cineacutetica e seu mecanismo de transporte descrito para T cruzi A incorporaccedilatildeo de arginina
pelo parasita apresenta pelo menos dois sistemas de transporte sendo um de alta afinidade e
outro de baixa afinidade Esse transporte eacute regulado ao longo do ciclo de vida do parasita
21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

Aos meus Pais Flamarion e Maria Nasareacute pelo
apoio amor e compreensatildeo incondicionais
AGRADECIMENTOS
Ao Prof Dr Ariel Mariano Silber pela grande oportunidade que me concedeu pela
orientaccedilatildeo paciecircncia apoio e amizade Obrigada por tudo
Agrave Elizabeth Pral teacutecnica do laboratoacuterio pelo apoio amizade ajuda e todo o suporte teacutecnico
Agrave minha amiga Mariacutea Julia Bariacuteson pela amizade apoio ajuda nos experimentos e nas
discussotildees dos resultados
Agrave minha famiacutelia meus pais Flamarion e Maria Nasareacute meus irmatildeos(atildes) Gilberto Gilmar
Gilson Gilsa Gildo Gildete e Jacoacute meus sobrinhos(as) Mariane Mateus Gisely Ceciacutelia
Nataacutelia Isabele Laviacutenia Maria e Venacircncio pelo apoio compreensatildeo e incentivo em todos os
momentos
Agrave minha amiga Valnia Pita por me fazer acreditar que era possiacutevel a realizaccedilatildeo desse sonho e
pelo apoio e estiacutemulo nessa caminhada
Aos meus amigos(as) do laboratoacuterio Raiacutessa Melo Marcell Crispim Sandra Carla Higo
Fernando Inga Eva Jean Douglas Carolina Manchola Ludmila Nakamura Brian Suarez
Rossana Duque e Letiacutecia Marchese pelo apoio ajuda e amizade
Ao Prof Dr Carsten Wrenger e seu grupo Flaacutevia Zimbres Kamila Meissner Jasmin
Lindner Thales Kronenberger
A Dra Lisvane Paes pelo apoio amizade e colaboraccedilatildeo nesse trabalho
Ao Prof Dr Joseacute Roberto Meyer-Fernandes (UFRJ) e aos integrantes do seu laboratoacuterio pela
disponibilidade e ajuda em alguns experimentos
Agrave Profa Dra Alicia J Kowaltowski do Instituto de Quiacutemica e aos integrantes do seu
laboratoacuterio pela disponibilidade do uso de equipamentos
Ao Prof Dr Gerhard Wunderlich e integrantes do seu laboratoacuterio em especial ao doutorando
Wesley Fotoran pela disponibilidade do uso de equipamentos e ajuda em alguns
experimentos
Agrave Fundaccedilatildeo de Amparo a Pesquisa do Estado de Satildeo Paulo (FAPESP) pelo suporte
financeiro
Ao Conselho Nacional de Desenvolvimento Cientiacutefico e Tecnoloacutegico (CNPq) pelo suporte
financeiro no primeiro ano desse trabalho
Agrave todos os professores funcionaacuterios e colegas do Departamento de Parasitologia (ICB-USP)
pelo apoio e convivecircncia
ldquoNunca deixe que lhe digam
que natildeo vale a pena
acreditar no sonho que se tem
ou que seus planos nunca vatildeo dar certo
ou que vocecirc nunca vai ser algueacutem ()
Confie em si mesmordquo
Renato Russo e Flaacutevio Venturini 1987
RESUMO
Damasceno FS Estudo da interferecircncia de faacutermacos que atuam via receptores eou
transportadores de GABA e Glutamato no ciclo de vida do Trypanosoma cruzi [dissertaccedilatildeo
(Mestrado em Biologia da Relaccedilatildeo Patoacutegeno-Hospedeiro)] Satildeo Paulo Instituto de Ciecircncias
Biomeacutedicas Universidade de Satildeo Paulo 2013
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica central e do Sul A quimioterapia disponiacutevel atualmente
limita-se a dois compostos Nifurtimox e o Benzonidazol Ambos os faacutermacos reduzem os
sintomas da doenccedila e a mortalidade das pessoas infectadas quando utilizadas na fase aguda
mas sua eficaacutecia na fase crocircnica eacute controversa Aleacutem do mais apresentam vaacuterios efeitos
colaterais O T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico como tambeacutem a importacircncia do aminoaacutecido
GABA no metabolismo do T cruzi visto que este tambeacutem eacute transportado pelo parasita Nesse
contexto o objetivo desse trabalho foi avaliar a interferecircncia de faacutermacos que atuam via
receptores ou transportadores de GABA e glutamato no ciclo de vida do T cruzi Os
faacutermacos selecionados foram Vigabatrina Pregabalina MK-801 MTEP e Memantina Os
resultados demonstram que apenas Memantina apresentou um melhor efeito tripanocida
Inibiu a proliferaccedilatildeo de formas epimastigotas interferiu na metaciclogecircnese como tambeacutem
afetou o metabolismo energeacutetico do parasita com a diminuiccedilatildeo dos niacuteveis de ATP aleacutem de
desencadear mecanismos que levam a apoptose das formas epimastigotas Memantina
tambeacutem interferiu no ciclo intracelular do parasita mais especificamente no estaacutegio
amastigota Interessantemente as fases do ciclo do parasita que satildeo mais afetadas satildeo as fases
que requerem mais energia estaacutegios replicativos (amastigota e epimastigota) como tambeacutem
os processos de diferenciaccedilatildeo e invasatildeo celular Apesar de apresentar valores de IC50 que
podem ser considerados altos em relaccedilatildeo aos valores descritos para formas epimastigotas
tratadas com Benzonidazol esses resultados mostram perspectivas promissoras visto que o
Memantina poderaacute ser utilizado como composto liacuteder para o desenho de derivados com
atividade tripanocida otimizada
Palavras-chave Trypanosoma cruzi Memantina Receptor de glutamato
ABSTRACT
Damasceno FS Study of interference of drugs that act via receptors andor transport of
GABA and glutamate in the life cycle of Trypanosoma cruzi [Masters thesis (Host-Pathogen
Relationship Biology)] Satildeo Paulo Instituto de Ciecircncias Biomeacutedicas Universidade de Satildeo
Paulo 2013
Chagas disease also known as American trypanosomiasis is caused by the flagellate
protozoan Trypanosoma cruzi and affects approximately 10 million people in endemic areas
of Mexico Central America and South America with about 25 million people living in areas
at risk of infection The currently available chemotherapy is limited to two compounds which
are Nifurtimox and Benznidazole Both drugs reduce symptoms of the disease and mortality
of infected people when used in the acute phase but its efficacy in chronic phase (phase in
which the majority of cases are diagnosed) is still controversial Moreover these drugs have
several side effects T cruzi is able to utilize carbohydrate and amino acids as carbon and
energy source Our group has studied the involvement of glutamate in parasite resistance to
thermal oxidative and metabolic stress as well as the importance of the amino acid GABA
metabolism in T cruzi The aim of this study was to evaluate the interference of drugs that
act in GABA and glutamate transporters or receptors in the life cycle of T cruzi The drugs
selected were vigabatrin pregabalin MK-801 memantine and MTEP The results showed
that only memantine has a better trypanocidal effect Memantine inhibited the proliferation of
epimastigotes interfered in metacyclogenesis and affected the energy metabolism of the
parasite with decreased levels of ATP and trigger mechanisms that lead to apoptosis of
epimastigotes Moreover interferes with intracellular cycle of the parasite specifically in
amastigote stage Interesting that stages of the parasite that require more energy are more
affected replicative stages (epimastigote and amastigote) as well as the processes of
differentiation and cell invasion IC50 value is more than that described to epimastigotes
treateds with Benznidazole but Memantine could be used as leader compound for the design
of drugs with optimized trypanocidal activity
Keywords Trypanosoma cruzi Memantine Glutamate receptor
LISTA DE ILUSTRACcedilOtildeES
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi 19
Figura 2 - Estruturas quiacutemicas do Benzonidazol e Nifurtimox 24
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica 28
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica 29
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse 40
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os
tratamentos com MK-801 ou Memantina 41
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina 42
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico ou oxidativo 44
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na
presenccedila do Memantina 45
Figura 10 - Efeito da Memantina na metaciclogecircnese 46
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo
de morte celular 48
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2 49
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
50
Figura 14 - Formas epimastigotas de T cruzi 51
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi 53
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade
de formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo
celular 54
LISTA DE ABREVIATURAS
ANOVA Anaacutelise de variacircncia
ATP Adenosina trifosfato
CHO-K1 Chinese Hamster Ovary (linhagem celular proveniente de ovaacuterio de
hamester chinecircs)
Ca2+
Caacutelcio
CO2 Dioacutexido de carbono
DNA Aacutecido desoxirribonucleacuteico
DO Densidade oacutetica
DTUs Discrete Typing Units
EGTA Aacutecido etilenoglicol-bis-aminoetiletertetraceacutetico
EROs Espeacutecies reativas de oxigecircnio
GABA Aminoaacutecido γ-aminobutiacuterico
GABA T Aminoaacutecido γ-aminobutiacuterico transaminase
GAPDH Gliceraldeiacutedo -3-fosfato desidrogenase
H2O2 Peroacutexido de hidrogecircnio
IC50 Metade da concentraccedilatildeo maacutexima inibitoacuteria
KCl Cloreto de potaacutessio
LIT Liver Infusion-Tryptose (Meio de cultura)
MCP Morte celular programada
mg Miligramas
mGluR Receptor metabotroacutepico de glutamato
MTT (3-(45-dimetiltiazol-2-il)-25 difenil bromo tetrazolio)
Na2HPO4 Fosfato de soacutedio monobaacutesico anidro
NaCl Cloreto de soacutedio
NaHCO3 Bicarbonato de soacutedio
NCCD Nomenclature Comittee on Cell Death (Comite de Nomeclatura de Morte
Celular)
NH4+
Ion amocircnio
NMDA N-metil-D-aspartato
OH Radical hidroxila
PBS Soluccedilatildeo salina tamponada com fosfato
pH Potencial hidrogeniocircnico
RPMI Roswell Park Memorial Institute
SDS Soacutedio dodecil sulfat
SFB Soro fetal bovino
U Unidades
μg Micrograma
μl Microlitro
μm Microcircmetro
SUMAacuteRIO
1 INTRODUCcedilAtildeO 16
11 Aspectos gerais da doenccedila de chagas 16
12 Trypanosoma cruzi 18
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos 20
14 Morte celular em Trypanosoma cruzi 21
15 Terapia na doenccedila de Chagas 23
16 Perspectivas de novos faacutermacos 25
2 OBJETIVOS 30
21 Objetivo geral 30
22 Objetivos especiacuteficos 30
3 MATERIAIS E MEacuteTODOS 31
31 Microorganismos utilizados no trabalho 31
311 Ceacutelulas hospedeiras 31
312 Trypanosoma cruzi 31
32 Manutenccedilatildeo das ceacutelulas hospedeiras 31
33 Obtenccedilatildeo dos parasitas 31
331 Epimastigotas 31
332 Curvas de crescimento de epimastigotas de T cruzi 31
333 Formas tripomastigotas 32
34 Resistecircncia a diferentes tipos de estresse 32
341 Ensaio de estresse teacutermico em formas epimastigotas 32
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico 33
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular 33
343 Ensaio de estresse oxidativo em formas epimastigotas 33
35 Tipo de morte celular em parasitas tratados com Memantina 34
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita 34
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas
epimastigotas 35
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio 35
36 Efeito sobre a metaciclogecircnese 35
37 Anaacutelise do efeito dos compostos no ciclo intracelular do T cruzi 36
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos 36
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas 36
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas
tripomastigotas 37
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os
compostos 37
38 Anaacutelises estatiacutesticas 38
4 RESULTADOS 39
41 Curvas de crescimento do T cruzi na presenccedila dos compostos 39
42 Interaccedilatildeo com condiccedilotildees de estresse 41
43 Efeito da Memantina na metaciclogecircnese 46
44 Tipo de morte celular sofrida pelo parasita tratado com Memantina 47
45 Efeito da Memantina no ciclo intracelular do T cruzi 52
5 DISCUSSAtildeO 55
6 CONCLUSOtildeES 59
REFEREcircNCIAS 61
16
1 INTRODUCcedilAtildeO
11 Aspectos gerais da doenccedila de chagas
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana foi
descoberta pelo meacutedico brasileiro Carlos Chagas (Chagas 1909) Esta doenccedila eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica Central e do Sul com cerca de 25 milhotildees de pessoas
vivendo em aacutereas de risco de infecccedilatildeo Ocorrem por ano aproximadamente 40 mil novos
casos e 12 mil mortes devido a complicaccedilotildees cardiacuteacas e digestivas (World Health
Organization 2012) Programas de controle do vetor desenvolvidos nos uacuteltimos 20 anos
como tambeacutem um maior controle nos bancos de sangue tem reduzido o nuacutemero de novos
casos da doenccedila de Chagas na Ameacuterica Latina (Rassi Marin-Neto 2010) Devido as intensas
migraccedilotildees de pessoas de paiacuteses endecircmicos o nuacutemero de casos de indiviacuteduos infectados tem
aumentado em aacutereas consideradas natildeo endecircmicas tais como Estados Unidos Canadaacute e
algumas regiotildees da Europa (Bern et al 2009 Maguire 2006 Schmunis 2007) A doenccedila eacute
uma zoonose que tem como reservatoacuterio natural do T cruzi uma ampla variedade de
mamiacuteferos do continente americano (Urbina Docampo 2003) Eacute transmitida para os seres
humanos e para mais de 150 espeacutecies de animais domeacutesticos e animais selvagens
principalmente por insetos hematoacutefagos da famiacutelia Reduviidae Apesar de existirem mais de
130 espeacutecies de triatomiacuteneos satildeo trecircs as mais importantes na transmissatildeo do T cruzi para o
homem Triatoma infestans Rhodnius prolixus e Triatoma dimidiata (Rassi Marin-Neto
2010) O parasita tambeacutem pode ser transmitido por transfusatildeo de sangue congenitamente (de
matildees infectadas para receacutem-nascidos) por transplante de oacutergatildeos ou medula oacutessea Tambeacutem
pode ocorrer por acidentes em laboratoacuterios que trabalham com o parasita vivo via esta que
epidemiologicamente eacute pouco relevante Outra forma de transmissatildeo eacute por via oral atraveacutes de
alimentos contaminados essa via vecircm sendo considerada como relevante na manutenccedilatildeo do
ciclo de transmissatildeo natural (Pereira et al 2009 Rassi Marin-Neto 2010)
A evoluccedilatildeo cliacutenica da doenccedila de Chagas pode ser dividida em duas fases aguda e
crocircnica A fase aguda normalmente eacute assintomaacutetica ou apresenta sintomatologia discreta que
na maioria dos casos passa despercebida pelo paciente Os sintomas quando aparecem o
fazem uma ou duas semanas apoacutes a exposiccedilatildeo ao triatomiacuteneo infectado ou poucos meses apoacutes
a transfusatildeo com sangue contaminado (Rassi Marin-Neto 2010) Os sintomas mais
frequentemente apresentados pelos pacientes agudos satildeo febre prolongada mal-estar
17
distensatildeo do fiacutegado baccedilo e linfonodos Pode haver tambeacutem edema subcutacircneo localizado ou
generalizado No caso particular de transmissatildeo vetorial o paciente pode apresentar um sinal
da porta de entrada do T cruzi na pele (chagoma) ou via mucosa ocular (sinal de Romantildea) As
manifestaccedilotildees da fase aguda se resolvem espontaneamente em aproximadamente 90 dos
indiviacuteduos infectados mesmo sem o tratamento com drogas tripanocidas Essa fase eacute
caracterizada pela alta parasitemia e ausecircncia de anticorpos especiacuteficos Nessa fase os
parasitas podem ser encontrados em praticamente todos os tecidos estando majoritariamente
presentes no sangue Por ser uma fase de curta duraccedilatildeo sem sintomatologia aparente ou com
sintomatologia pouco definida haacute um pequeno nuacutemero de casos detectados sendo portanto a
fase menos estudada (Rassi Marin-Neto 2010 Silber et al 2005b)
Logo apoacutes a fase aguda tem iniacutecio a fase crocircnica caracterizada pelos altos tiacutetulos de
anticorpos no sangue do paciente e pelo fato de que a parasitemia eacute inaparente O paciente
pode permanecer por vaacuterios anos ou ateacute mesmo pelo resto da vida sem apresentar sinais ou
sintomas Essa forma da fase crocircnica eacute chamada de indeterminada Aproximadamente 30-40
dos pacientes crocircnicos indeterminados desenvolvem a sintomatologia caracteriacutestica da
doenccedila evoluindo para a chamada fase crocircnica sintomaacutetica Nesta fase as trecircs principais
formas satildeo a cardiacuteaca a digestiva e a cardiodigestiva (Acquatella 2007 Marin-Neto et al
1999) Na forma cardiacuteaca o coraccedilatildeo eacute um dos primeiros oacutergatildeos afetados natildeo necessariamente
o uacutenico desenvolvendo hipertrofia e dilataccedilatildeo Na forma digestiva o trato digestivo apresenta
alteraccedilotildees anatocircmicas e fisioloacutegicas importantes sendo a principal altereccedilatildeo a dilataccedilatildeo do
intestino e do esocircfago que podem apresentar aspecto de megaviacutescera (Marin-Neto et al
1999 Rassi et al 2000) revisado por (Boscardin et al 2009 Rassi Marin-Neto 2010)
18
12 Trypanosoma cruzi
Trypanosoma cruzi eacute um parasita intracelular obrigatoacuterio classicamente localizado na
ordem kinetoplastida gecircnero Trypanosoma que causa uma infecccedilatildeo crocircnica em humanos e
em grande nuacutemero de outras espeacutecies de mamiacuteferos (Hoare 1972) Recentemente devido agrave
alta variabilidade bioloacutegica bioquiacutemica e molecular a espeacutecie Trypanosoma cruzi foi
dividida em seis linhagens subsespeciacuteficas (Discrete Typing Units ndash DTUs) numerados de I a
VI Esta classificaccedilatildeo foi realizada com base em caracteriacutesticas geneacuteticas moleculares e
imunoloacutegicas com o objetivo de unificar a nomenclatura e facilitar a comunicaccedilatildeo dentro da
comunidade cientiacutefica (Zingales et al 2009)
O T cruzi tem um ciclo de vida complexo que alterna entre o inseto vetor e o
hospedeiro mamiacutefero O parasita apresenta diferentes formas correspondentes a diferentes
estaacutegios do seu ciclo de vida Estes diferentes estaacutegios tecircm sido descritos com base em
criteacuterios morfoloacutegicos como a forma fusiforme ou arredondados a regiatildeo onde emerge o
flagelo e a posiccedilatildeo do cinetoplasto (estrutura contendo o DNA da uacutenica mitococircndria presente
no parasita) em relaccedilatildeo ao nuacutecleo No hospedeiro mamiacutefero foram descritas as formas
amastigota epimastigota intracelular e tripomastigota sanguiacutecola A forma amastigota eacute um
estaacutegio replicativo e infectivo Satildeo ceacutelulas arredondadas medindo de 2-6 microm de diacircmetro
encontradas no citoplasma das ceacutelulas infectadas Os epimastigotas intracelulares
(epimastigote-like) satildeo formas replicativas intermediaacuterias entre amastigotas e tripomastigotas
tambeacutem encontrados dentro das ceacutelulas medindo aproximadamente 54 microm Essas formas satildeo
menores que as formas epimastigotas encontradas no inseto vetor poreacutem compartilha com as
mesmas padrotildees morfomeacutetricos e diversas caracteriacutesticas bioquiacutemicas e bioloacutegicas (Almeida-
De-Faria et al 1999) O tripomastigota sanguiacutecola eacute um estaacutegio flagelado infectivo natildeo
replicativo encontrado principalmente na corrente sanguiacutenea do hospedeiro mamiacutefero Mede
aproximadamente 18 microm de comprimento incluindo um extremo livre do flagelo de
aproximadamente 6 microm e 2-3 microm de largura
No inseto vetor encontram-se predominantemente as formas epimastigota e
tripomastigota metaciacuteclico Epimastigota eacute uma forma flagelada replicativa e natildeo infectiva
Mede aproximadamente 20-40 microm de comprimento e 2-5 microm de largura essas formas estatildeo
presentes tipicamente na porccedilatildeo meacutedia do intestino do hospedeiro invertebrado
Tripomastigota metaciacuteclico eacute uma forma infectiva natildeo replicativa encontrada na porccedilatildeo final
do intestino do inseto vetor (Brener 1971) revisado por (Silber et al 2005a Tyler Engman
2001)
19
O ciclo de vida do T cruzi inicia-se quando os parasitas presentes na corrente
sanguiacutenea do hospedeiro vertebrado infectado passam ao inseto vetor durante o repasto
sanguiacuteneo No intestino do inseto estas formas se diferenciam em formas epimastigotas (De
Souza et al 2002) que apoacutes multiplicaccedilatildeo por fissatildeo binaacuteria colonizam a porccedilatildeo terminal do
intestino e se diferenciam novamente em tripomastigotas metaciacuteclicos Quando o inseto
infectado pica o hospedeiro vertebrado e inicia o repasto sanguiacuteneo concomitantemente
elimina as formas tripomastigotas metaciacuteclicos junto com as fezes e urina Uma vez em
contato com a pele ou mucosas do hospedeiro mamiacutefero os tripomastigotas satildeo internalizados
atraveacutes de pequenas lesotildees na pele causadas pelo proacuteprio hospedeiro (Alves Colli 2008) Os
tripomastigotas invadem as ceacutelulas do hospedeiro mamiacutefero e se diferenciam em formas
replicativas denominadas amastigotas estabelecendo assim a infecccedilatildeo Apoacutes
aproximadamente 96 horas os amastigotas comeccedilam a se diferenciar em tripomastigotas
passando pela forma intermediaacuteria epimastigota intracelular Estas formas tripomastigotas satildeo
liberadas na corrente sanguiacutenea podendo invadir novas ceacutelulas ou serem ingeridas pelo inseto
vetor em um novo repasto sanguiacuteneo fechando assim o seu ciclo de vida O esquema do ciclo
de vida do Trypanosoma cruzi estaacute representado na Figura 1
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi
20
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos
T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono As formas epimastigotas preferencialmente catabolizam glicose e apoacutes a exaustatildeo
do carboidrato ocorre uma mudanccedila para o metabolismo de aminoaacutecidos A prolina e a
glutamina satildeo os principais aminoaacutecidos metabolizados pelas formas epimastigotas visto que
satildeo abudantes na hemolinfa e nos fluiacutedos dos tecidos do inseto vetor Esse metabolismo
ocorre com produccedilatildeo de NH4+
(produto da deaminaccedilatildeo dos aminoaacutecidos) que eacute excretada para
o meio extracelular (Bringaud et al 2006 Cazzulo 1994) As formas tripomastigotas
utilizam principalmente a glicose que eacute abundante nos fluiacutedos de seus hospedeiros
vertebrados Jaacute as formas amastigotas encontradas no citoplasma das ceacutelulas do hospedeiro
mamiacutefero ambiente pobre em glicose livre usam os aminoaacutecidos como principal fonte de
carbono e energia apresentando maior quantidade de prolina intracelular do que as formas
tripomastigotas (Silber et al 2009)
Entre os aminoaacutecidos metabolizados pelas formas epimastigotas estatildeo prolina glicina
asparagina glutamina aspartato glutamato leucina e isoleucina (Mancilla et al 1967 Odaly
et al 1983 Silber et al 2005a Sylvester Krassner 1976 Zeledon 1960) Os aminoaacutecidos
estatildeo envolvidos tambeacutem em outros processos importantes do ciclo de vida do T cruzi O
aminoaacutecido L-prolina eacute essencial na diferenciaccedilatildeo das formas epimastigota intracelular para
tripomastigotas aleacutem de estarem envolvidos nos processos de osmorregulaccedilatildeo crescimento e
resistecircncia do parasita ao estresse oxidativo e nutricional (Homsy et al 1989 Magdaleno et
al 2009 Magdaleno et al 2011 Pereira et al 2002 Pereira et al 2003 Tonelli et al
2004) Glutamato prolina e aspartato satildeo importantes na metaciclogecircnese ou seja na
diferenciaccedilatildeo de formas epimastigotas para tripomastigotas metaciacuteclicos (Contreras et al
1985) Prolina glicina alanina e glutamato satildeo descritos como importantes nos processos de
osmorregulaccedilatildeo e controle do volume celular (Rohloff et al 2003 Rohloff et al 2004a)
Os aminoaacutecidos para que sejam metabolizados ou cumprirem outras funccedilotildees devem
ser obtidos a partir da degradaccedilatildeo de proteiacutenas sintetizados ou incorporados pelo parasita
atraveacutes de transportadores localizados na membrana (Silber et al 2005a) O primeiro
transportador de aminoaacutecidos em T cruzi foi descrito bioquimicamente em 1970 o
transportador de lisina (Hampton 1970) Outros aminoaacutecidos tambeacutem jaacute tiveram a sua
cineacutetica e seu mecanismo de transporte descrito para T cruzi A incorporaccedilatildeo de arginina
pelo parasita apresenta pelo menos dois sistemas de transporte sendo um de alta afinidade e
outro de baixa afinidade Esse transporte eacute regulado ao longo do ciclo de vida do parasita
21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

AGRADECIMENTOS
Ao Prof Dr Ariel Mariano Silber pela grande oportunidade que me concedeu pela
orientaccedilatildeo paciecircncia apoio e amizade Obrigada por tudo
Agrave Elizabeth Pral teacutecnica do laboratoacuterio pelo apoio amizade ajuda e todo o suporte teacutecnico
Agrave minha amiga Mariacutea Julia Bariacuteson pela amizade apoio ajuda nos experimentos e nas
discussotildees dos resultados
Agrave minha famiacutelia meus pais Flamarion e Maria Nasareacute meus irmatildeos(atildes) Gilberto Gilmar
Gilson Gilsa Gildo Gildete e Jacoacute meus sobrinhos(as) Mariane Mateus Gisely Ceciacutelia
Nataacutelia Isabele Laviacutenia Maria e Venacircncio pelo apoio compreensatildeo e incentivo em todos os
momentos
Agrave minha amiga Valnia Pita por me fazer acreditar que era possiacutevel a realizaccedilatildeo desse sonho e
pelo apoio e estiacutemulo nessa caminhada
Aos meus amigos(as) do laboratoacuterio Raiacutessa Melo Marcell Crispim Sandra Carla Higo
Fernando Inga Eva Jean Douglas Carolina Manchola Ludmila Nakamura Brian Suarez
Rossana Duque e Letiacutecia Marchese pelo apoio ajuda e amizade
Ao Prof Dr Carsten Wrenger e seu grupo Flaacutevia Zimbres Kamila Meissner Jasmin
Lindner Thales Kronenberger
A Dra Lisvane Paes pelo apoio amizade e colaboraccedilatildeo nesse trabalho
Ao Prof Dr Joseacute Roberto Meyer-Fernandes (UFRJ) e aos integrantes do seu laboratoacuterio pela
disponibilidade e ajuda em alguns experimentos
Agrave Profa Dra Alicia J Kowaltowski do Instituto de Quiacutemica e aos integrantes do seu
laboratoacuterio pela disponibilidade do uso de equipamentos
Ao Prof Dr Gerhard Wunderlich e integrantes do seu laboratoacuterio em especial ao doutorando
Wesley Fotoran pela disponibilidade do uso de equipamentos e ajuda em alguns
experimentos
Agrave Fundaccedilatildeo de Amparo a Pesquisa do Estado de Satildeo Paulo (FAPESP) pelo suporte
financeiro
Ao Conselho Nacional de Desenvolvimento Cientiacutefico e Tecnoloacutegico (CNPq) pelo suporte
financeiro no primeiro ano desse trabalho
Agrave todos os professores funcionaacuterios e colegas do Departamento de Parasitologia (ICB-USP)
pelo apoio e convivecircncia
ldquoNunca deixe que lhe digam
que natildeo vale a pena
acreditar no sonho que se tem
ou que seus planos nunca vatildeo dar certo
ou que vocecirc nunca vai ser algueacutem ()
Confie em si mesmordquo
Renato Russo e Flaacutevio Venturini 1987
RESUMO
Damasceno FS Estudo da interferecircncia de faacutermacos que atuam via receptores eou
transportadores de GABA e Glutamato no ciclo de vida do Trypanosoma cruzi [dissertaccedilatildeo
(Mestrado em Biologia da Relaccedilatildeo Patoacutegeno-Hospedeiro)] Satildeo Paulo Instituto de Ciecircncias
Biomeacutedicas Universidade de Satildeo Paulo 2013
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica central e do Sul A quimioterapia disponiacutevel atualmente
limita-se a dois compostos Nifurtimox e o Benzonidazol Ambos os faacutermacos reduzem os
sintomas da doenccedila e a mortalidade das pessoas infectadas quando utilizadas na fase aguda
mas sua eficaacutecia na fase crocircnica eacute controversa Aleacutem do mais apresentam vaacuterios efeitos
colaterais O T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico como tambeacutem a importacircncia do aminoaacutecido
GABA no metabolismo do T cruzi visto que este tambeacutem eacute transportado pelo parasita Nesse
contexto o objetivo desse trabalho foi avaliar a interferecircncia de faacutermacos que atuam via
receptores ou transportadores de GABA e glutamato no ciclo de vida do T cruzi Os
faacutermacos selecionados foram Vigabatrina Pregabalina MK-801 MTEP e Memantina Os
resultados demonstram que apenas Memantina apresentou um melhor efeito tripanocida
Inibiu a proliferaccedilatildeo de formas epimastigotas interferiu na metaciclogecircnese como tambeacutem
afetou o metabolismo energeacutetico do parasita com a diminuiccedilatildeo dos niacuteveis de ATP aleacutem de
desencadear mecanismos que levam a apoptose das formas epimastigotas Memantina
tambeacutem interferiu no ciclo intracelular do parasita mais especificamente no estaacutegio
amastigota Interessantemente as fases do ciclo do parasita que satildeo mais afetadas satildeo as fases
que requerem mais energia estaacutegios replicativos (amastigota e epimastigota) como tambeacutem
os processos de diferenciaccedilatildeo e invasatildeo celular Apesar de apresentar valores de IC50 que
podem ser considerados altos em relaccedilatildeo aos valores descritos para formas epimastigotas
tratadas com Benzonidazol esses resultados mostram perspectivas promissoras visto que o
Memantina poderaacute ser utilizado como composto liacuteder para o desenho de derivados com
atividade tripanocida otimizada
Palavras-chave Trypanosoma cruzi Memantina Receptor de glutamato
ABSTRACT
Damasceno FS Study of interference of drugs that act via receptors andor transport of
GABA and glutamate in the life cycle of Trypanosoma cruzi [Masters thesis (Host-Pathogen
Relationship Biology)] Satildeo Paulo Instituto de Ciecircncias Biomeacutedicas Universidade de Satildeo
Paulo 2013
Chagas disease also known as American trypanosomiasis is caused by the flagellate
protozoan Trypanosoma cruzi and affects approximately 10 million people in endemic areas
of Mexico Central America and South America with about 25 million people living in areas
at risk of infection The currently available chemotherapy is limited to two compounds which
are Nifurtimox and Benznidazole Both drugs reduce symptoms of the disease and mortality
of infected people when used in the acute phase but its efficacy in chronic phase (phase in
which the majority of cases are diagnosed) is still controversial Moreover these drugs have
several side effects T cruzi is able to utilize carbohydrate and amino acids as carbon and
energy source Our group has studied the involvement of glutamate in parasite resistance to
thermal oxidative and metabolic stress as well as the importance of the amino acid GABA
metabolism in T cruzi The aim of this study was to evaluate the interference of drugs that
act in GABA and glutamate transporters or receptors in the life cycle of T cruzi The drugs
selected were vigabatrin pregabalin MK-801 memantine and MTEP The results showed
that only memantine has a better trypanocidal effect Memantine inhibited the proliferation of
epimastigotes interfered in metacyclogenesis and affected the energy metabolism of the
parasite with decreased levels of ATP and trigger mechanisms that lead to apoptosis of
epimastigotes Moreover interferes with intracellular cycle of the parasite specifically in
amastigote stage Interesting that stages of the parasite that require more energy are more
affected replicative stages (epimastigote and amastigote) as well as the processes of
differentiation and cell invasion IC50 value is more than that described to epimastigotes
treateds with Benznidazole but Memantine could be used as leader compound for the design
of drugs with optimized trypanocidal activity
Keywords Trypanosoma cruzi Memantine Glutamate receptor
LISTA DE ILUSTRACcedilOtildeES
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi 19
Figura 2 - Estruturas quiacutemicas do Benzonidazol e Nifurtimox 24
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica 28
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica 29
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse 40
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os
tratamentos com MK-801 ou Memantina 41
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina 42
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico ou oxidativo 44
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na
presenccedila do Memantina 45
Figura 10 - Efeito da Memantina na metaciclogecircnese 46
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo
de morte celular 48
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2 49
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
50
Figura 14 - Formas epimastigotas de T cruzi 51
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi 53
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade
de formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo
celular 54
LISTA DE ABREVIATURAS
ANOVA Anaacutelise de variacircncia
ATP Adenosina trifosfato
CHO-K1 Chinese Hamster Ovary (linhagem celular proveniente de ovaacuterio de
hamester chinecircs)
Ca2+
Caacutelcio
CO2 Dioacutexido de carbono
DNA Aacutecido desoxirribonucleacuteico
DO Densidade oacutetica
DTUs Discrete Typing Units
EGTA Aacutecido etilenoglicol-bis-aminoetiletertetraceacutetico
EROs Espeacutecies reativas de oxigecircnio
GABA Aminoaacutecido γ-aminobutiacuterico
GABA T Aminoaacutecido γ-aminobutiacuterico transaminase
GAPDH Gliceraldeiacutedo -3-fosfato desidrogenase
H2O2 Peroacutexido de hidrogecircnio
IC50 Metade da concentraccedilatildeo maacutexima inibitoacuteria
KCl Cloreto de potaacutessio
LIT Liver Infusion-Tryptose (Meio de cultura)
MCP Morte celular programada
mg Miligramas
mGluR Receptor metabotroacutepico de glutamato
MTT (3-(45-dimetiltiazol-2-il)-25 difenil bromo tetrazolio)
Na2HPO4 Fosfato de soacutedio monobaacutesico anidro
NaCl Cloreto de soacutedio
NaHCO3 Bicarbonato de soacutedio
NCCD Nomenclature Comittee on Cell Death (Comite de Nomeclatura de Morte
Celular)
NH4+
Ion amocircnio
NMDA N-metil-D-aspartato
OH Radical hidroxila
PBS Soluccedilatildeo salina tamponada com fosfato
pH Potencial hidrogeniocircnico
RPMI Roswell Park Memorial Institute
SDS Soacutedio dodecil sulfat
SFB Soro fetal bovino
U Unidades
μg Micrograma
μl Microlitro
μm Microcircmetro
SUMAacuteRIO
1 INTRODUCcedilAtildeO 16
11 Aspectos gerais da doenccedila de chagas 16
12 Trypanosoma cruzi 18
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos 20
14 Morte celular em Trypanosoma cruzi 21
15 Terapia na doenccedila de Chagas 23
16 Perspectivas de novos faacutermacos 25
2 OBJETIVOS 30
21 Objetivo geral 30
22 Objetivos especiacuteficos 30
3 MATERIAIS E MEacuteTODOS 31
31 Microorganismos utilizados no trabalho 31
311 Ceacutelulas hospedeiras 31
312 Trypanosoma cruzi 31
32 Manutenccedilatildeo das ceacutelulas hospedeiras 31
33 Obtenccedilatildeo dos parasitas 31
331 Epimastigotas 31
332 Curvas de crescimento de epimastigotas de T cruzi 31
333 Formas tripomastigotas 32
34 Resistecircncia a diferentes tipos de estresse 32
341 Ensaio de estresse teacutermico em formas epimastigotas 32
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico 33
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular 33
343 Ensaio de estresse oxidativo em formas epimastigotas 33
35 Tipo de morte celular em parasitas tratados com Memantina 34
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita 34
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas
epimastigotas 35
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio 35
36 Efeito sobre a metaciclogecircnese 35
37 Anaacutelise do efeito dos compostos no ciclo intracelular do T cruzi 36
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos 36
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas 36
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas
tripomastigotas 37
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os
compostos 37
38 Anaacutelises estatiacutesticas 38
4 RESULTADOS 39
41 Curvas de crescimento do T cruzi na presenccedila dos compostos 39
42 Interaccedilatildeo com condiccedilotildees de estresse 41
43 Efeito da Memantina na metaciclogecircnese 46
44 Tipo de morte celular sofrida pelo parasita tratado com Memantina 47
45 Efeito da Memantina no ciclo intracelular do T cruzi 52
5 DISCUSSAtildeO 55
6 CONCLUSOtildeES 59
REFEREcircNCIAS 61
16
1 INTRODUCcedilAtildeO
11 Aspectos gerais da doenccedila de chagas
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana foi
descoberta pelo meacutedico brasileiro Carlos Chagas (Chagas 1909) Esta doenccedila eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica Central e do Sul com cerca de 25 milhotildees de pessoas
vivendo em aacutereas de risco de infecccedilatildeo Ocorrem por ano aproximadamente 40 mil novos
casos e 12 mil mortes devido a complicaccedilotildees cardiacuteacas e digestivas (World Health
Organization 2012) Programas de controle do vetor desenvolvidos nos uacuteltimos 20 anos
como tambeacutem um maior controle nos bancos de sangue tem reduzido o nuacutemero de novos
casos da doenccedila de Chagas na Ameacuterica Latina (Rassi Marin-Neto 2010) Devido as intensas
migraccedilotildees de pessoas de paiacuteses endecircmicos o nuacutemero de casos de indiviacuteduos infectados tem
aumentado em aacutereas consideradas natildeo endecircmicas tais como Estados Unidos Canadaacute e
algumas regiotildees da Europa (Bern et al 2009 Maguire 2006 Schmunis 2007) A doenccedila eacute
uma zoonose que tem como reservatoacuterio natural do T cruzi uma ampla variedade de
mamiacuteferos do continente americano (Urbina Docampo 2003) Eacute transmitida para os seres
humanos e para mais de 150 espeacutecies de animais domeacutesticos e animais selvagens
principalmente por insetos hematoacutefagos da famiacutelia Reduviidae Apesar de existirem mais de
130 espeacutecies de triatomiacuteneos satildeo trecircs as mais importantes na transmissatildeo do T cruzi para o
homem Triatoma infestans Rhodnius prolixus e Triatoma dimidiata (Rassi Marin-Neto
2010) O parasita tambeacutem pode ser transmitido por transfusatildeo de sangue congenitamente (de
matildees infectadas para receacutem-nascidos) por transplante de oacutergatildeos ou medula oacutessea Tambeacutem
pode ocorrer por acidentes em laboratoacuterios que trabalham com o parasita vivo via esta que
epidemiologicamente eacute pouco relevante Outra forma de transmissatildeo eacute por via oral atraveacutes de
alimentos contaminados essa via vecircm sendo considerada como relevante na manutenccedilatildeo do
ciclo de transmissatildeo natural (Pereira et al 2009 Rassi Marin-Neto 2010)
A evoluccedilatildeo cliacutenica da doenccedila de Chagas pode ser dividida em duas fases aguda e
crocircnica A fase aguda normalmente eacute assintomaacutetica ou apresenta sintomatologia discreta que
na maioria dos casos passa despercebida pelo paciente Os sintomas quando aparecem o
fazem uma ou duas semanas apoacutes a exposiccedilatildeo ao triatomiacuteneo infectado ou poucos meses apoacutes
a transfusatildeo com sangue contaminado (Rassi Marin-Neto 2010) Os sintomas mais
frequentemente apresentados pelos pacientes agudos satildeo febre prolongada mal-estar
17
distensatildeo do fiacutegado baccedilo e linfonodos Pode haver tambeacutem edema subcutacircneo localizado ou
generalizado No caso particular de transmissatildeo vetorial o paciente pode apresentar um sinal
da porta de entrada do T cruzi na pele (chagoma) ou via mucosa ocular (sinal de Romantildea) As
manifestaccedilotildees da fase aguda se resolvem espontaneamente em aproximadamente 90 dos
indiviacuteduos infectados mesmo sem o tratamento com drogas tripanocidas Essa fase eacute
caracterizada pela alta parasitemia e ausecircncia de anticorpos especiacuteficos Nessa fase os
parasitas podem ser encontrados em praticamente todos os tecidos estando majoritariamente
presentes no sangue Por ser uma fase de curta duraccedilatildeo sem sintomatologia aparente ou com
sintomatologia pouco definida haacute um pequeno nuacutemero de casos detectados sendo portanto a
fase menos estudada (Rassi Marin-Neto 2010 Silber et al 2005b)
Logo apoacutes a fase aguda tem iniacutecio a fase crocircnica caracterizada pelos altos tiacutetulos de
anticorpos no sangue do paciente e pelo fato de que a parasitemia eacute inaparente O paciente
pode permanecer por vaacuterios anos ou ateacute mesmo pelo resto da vida sem apresentar sinais ou
sintomas Essa forma da fase crocircnica eacute chamada de indeterminada Aproximadamente 30-40
dos pacientes crocircnicos indeterminados desenvolvem a sintomatologia caracteriacutestica da
doenccedila evoluindo para a chamada fase crocircnica sintomaacutetica Nesta fase as trecircs principais
formas satildeo a cardiacuteaca a digestiva e a cardiodigestiva (Acquatella 2007 Marin-Neto et al
1999) Na forma cardiacuteaca o coraccedilatildeo eacute um dos primeiros oacutergatildeos afetados natildeo necessariamente
o uacutenico desenvolvendo hipertrofia e dilataccedilatildeo Na forma digestiva o trato digestivo apresenta
alteraccedilotildees anatocircmicas e fisioloacutegicas importantes sendo a principal altereccedilatildeo a dilataccedilatildeo do
intestino e do esocircfago que podem apresentar aspecto de megaviacutescera (Marin-Neto et al
1999 Rassi et al 2000) revisado por (Boscardin et al 2009 Rassi Marin-Neto 2010)
18
12 Trypanosoma cruzi
Trypanosoma cruzi eacute um parasita intracelular obrigatoacuterio classicamente localizado na
ordem kinetoplastida gecircnero Trypanosoma que causa uma infecccedilatildeo crocircnica em humanos e
em grande nuacutemero de outras espeacutecies de mamiacuteferos (Hoare 1972) Recentemente devido agrave
alta variabilidade bioloacutegica bioquiacutemica e molecular a espeacutecie Trypanosoma cruzi foi
dividida em seis linhagens subsespeciacuteficas (Discrete Typing Units ndash DTUs) numerados de I a
VI Esta classificaccedilatildeo foi realizada com base em caracteriacutesticas geneacuteticas moleculares e
imunoloacutegicas com o objetivo de unificar a nomenclatura e facilitar a comunicaccedilatildeo dentro da
comunidade cientiacutefica (Zingales et al 2009)
O T cruzi tem um ciclo de vida complexo que alterna entre o inseto vetor e o
hospedeiro mamiacutefero O parasita apresenta diferentes formas correspondentes a diferentes
estaacutegios do seu ciclo de vida Estes diferentes estaacutegios tecircm sido descritos com base em
criteacuterios morfoloacutegicos como a forma fusiforme ou arredondados a regiatildeo onde emerge o
flagelo e a posiccedilatildeo do cinetoplasto (estrutura contendo o DNA da uacutenica mitococircndria presente
no parasita) em relaccedilatildeo ao nuacutecleo No hospedeiro mamiacutefero foram descritas as formas
amastigota epimastigota intracelular e tripomastigota sanguiacutecola A forma amastigota eacute um
estaacutegio replicativo e infectivo Satildeo ceacutelulas arredondadas medindo de 2-6 microm de diacircmetro
encontradas no citoplasma das ceacutelulas infectadas Os epimastigotas intracelulares
(epimastigote-like) satildeo formas replicativas intermediaacuterias entre amastigotas e tripomastigotas
tambeacutem encontrados dentro das ceacutelulas medindo aproximadamente 54 microm Essas formas satildeo
menores que as formas epimastigotas encontradas no inseto vetor poreacutem compartilha com as
mesmas padrotildees morfomeacutetricos e diversas caracteriacutesticas bioquiacutemicas e bioloacutegicas (Almeida-
De-Faria et al 1999) O tripomastigota sanguiacutecola eacute um estaacutegio flagelado infectivo natildeo
replicativo encontrado principalmente na corrente sanguiacutenea do hospedeiro mamiacutefero Mede
aproximadamente 18 microm de comprimento incluindo um extremo livre do flagelo de
aproximadamente 6 microm e 2-3 microm de largura
No inseto vetor encontram-se predominantemente as formas epimastigota e
tripomastigota metaciacuteclico Epimastigota eacute uma forma flagelada replicativa e natildeo infectiva
Mede aproximadamente 20-40 microm de comprimento e 2-5 microm de largura essas formas estatildeo
presentes tipicamente na porccedilatildeo meacutedia do intestino do hospedeiro invertebrado
Tripomastigota metaciacuteclico eacute uma forma infectiva natildeo replicativa encontrada na porccedilatildeo final
do intestino do inseto vetor (Brener 1971) revisado por (Silber et al 2005a Tyler Engman
2001)
19
O ciclo de vida do T cruzi inicia-se quando os parasitas presentes na corrente
sanguiacutenea do hospedeiro vertebrado infectado passam ao inseto vetor durante o repasto
sanguiacuteneo No intestino do inseto estas formas se diferenciam em formas epimastigotas (De
Souza et al 2002) que apoacutes multiplicaccedilatildeo por fissatildeo binaacuteria colonizam a porccedilatildeo terminal do
intestino e se diferenciam novamente em tripomastigotas metaciacuteclicos Quando o inseto
infectado pica o hospedeiro vertebrado e inicia o repasto sanguiacuteneo concomitantemente
elimina as formas tripomastigotas metaciacuteclicos junto com as fezes e urina Uma vez em
contato com a pele ou mucosas do hospedeiro mamiacutefero os tripomastigotas satildeo internalizados
atraveacutes de pequenas lesotildees na pele causadas pelo proacuteprio hospedeiro (Alves Colli 2008) Os
tripomastigotas invadem as ceacutelulas do hospedeiro mamiacutefero e se diferenciam em formas
replicativas denominadas amastigotas estabelecendo assim a infecccedilatildeo Apoacutes
aproximadamente 96 horas os amastigotas comeccedilam a se diferenciar em tripomastigotas
passando pela forma intermediaacuteria epimastigota intracelular Estas formas tripomastigotas satildeo
liberadas na corrente sanguiacutenea podendo invadir novas ceacutelulas ou serem ingeridas pelo inseto
vetor em um novo repasto sanguiacuteneo fechando assim o seu ciclo de vida O esquema do ciclo
de vida do Trypanosoma cruzi estaacute representado na Figura 1
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi
20
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos
T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono As formas epimastigotas preferencialmente catabolizam glicose e apoacutes a exaustatildeo
do carboidrato ocorre uma mudanccedila para o metabolismo de aminoaacutecidos A prolina e a
glutamina satildeo os principais aminoaacutecidos metabolizados pelas formas epimastigotas visto que
satildeo abudantes na hemolinfa e nos fluiacutedos dos tecidos do inseto vetor Esse metabolismo
ocorre com produccedilatildeo de NH4+
(produto da deaminaccedilatildeo dos aminoaacutecidos) que eacute excretada para
o meio extracelular (Bringaud et al 2006 Cazzulo 1994) As formas tripomastigotas
utilizam principalmente a glicose que eacute abundante nos fluiacutedos de seus hospedeiros
vertebrados Jaacute as formas amastigotas encontradas no citoplasma das ceacutelulas do hospedeiro
mamiacutefero ambiente pobre em glicose livre usam os aminoaacutecidos como principal fonte de
carbono e energia apresentando maior quantidade de prolina intracelular do que as formas
tripomastigotas (Silber et al 2009)
Entre os aminoaacutecidos metabolizados pelas formas epimastigotas estatildeo prolina glicina
asparagina glutamina aspartato glutamato leucina e isoleucina (Mancilla et al 1967 Odaly
et al 1983 Silber et al 2005a Sylvester Krassner 1976 Zeledon 1960) Os aminoaacutecidos
estatildeo envolvidos tambeacutem em outros processos importantes do ciclo de vida do T cruzi O
aminoaacutecido L-prolina eacute essencial na diferenciaccedilatildeo das formas epimastigota intracelular para
tripomastigotas aleacutem de estarem envolvidos nos processos de osmorregulaccedilatildeo crescimento e
resistecircncia do parasita ao estresse oxidativo e nutricional (Homsy et al 1989 Magdaleno et
al 2009 Magdaleno et al 2011 Pereira et al 2002 Pereira et al 2003 Tonelli et al
2004) Glutamato prolina e aspartato satildeo importantes na metaciclogecircnese ou seja na
diferenciaccedilatildeo de formas epimastigotas para tripomastigotas metaciacuteclicos (Contreras et al
1985) Prolina glicina alanina e glutamato satildeo descritos como importantes nos processos de
osmorregulaccedilatildeo e controle do volume celular (Rohloff et al 2003 Rohloff et al 2004a)
Os aminoaacutecidos para que sejam metabolizados ou cumprirem outras funccedilotildees devem
ser obtidos a partir da degradaccedilatildeo de proteiacutenas sintetizados ou incorporados pelo parasita
atraveacutes de transportadores localizados na membrana (Silber et al 2005a) O primeiro
transportador de aminoaacutecidos em T cruzi foi descrito bioquimicamente em 1970 o
transportador de lisina (Hampton 1970) Outros aminoaacutecidos tambeacutem jaacute tiveram a sua
cineacutetica e seu mecanismo de transporte descrito para T cruzi A incorporaccedilatildeo de arginina
pelo parasita apresenta pelo menos dois sistemas de transporte sendo um de alta afinidade e
outro de baixa afinidade Esse transporte eacute regulado ao longo do ciclo de vida do parasita
21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

ldquoNunca deixe que lhe digam
que natildeo vale a pena
acreditar no sonho que se tem
ou que seus planos nunca vatildeo dar certo
ou que vocecirc nunca vai ser algueacutem ()
Confie em si mesmordquo
Renato Russo e Flaacutevio Venturini 1987
RESUMO
Damasceno FS Estudo da interferecircncia de faacutermacos que atuam via receptores eou
transportadores de GABA e Glutamato no ciclo de vida do Trypanosoma cruzi [dissertaccedilatildeo
(Mestrado em Biologia da Relaccedilatildeo Patoacutegeno-Hospedeiro)] Satildeo Paulo Instituto de Ciecircncias
Biomeacutedicas Universidade de Satildeo Paulo 2013
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica central e do Sul A quimioterapia disponiacutevel atualmente
limita-se a dois compostos Nifurtimox e o Benzonidazol Ambos os faacutermacos reduzem os
sintomas da doenccedila e a mortalidade das pessoas infectadas quando utilizadas na fase aguda
mas sua eficaacutecia na fase crocircnica eacute controversa Aleacutem do mais apresentam vaacuterios efeitos
colaterais O T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico como tambeacutem a importacircncia do aminoaacutecido
GABA no metabolismo do T cruzi visto que este tambeacutem eacute transportado pelo parasita Nesse
contexto o objetivo desse trabalho foi avaliar a interferecircncia de faacutermacos que atuam via
receptores ou transportadores de GABA e glutamato no ciclo de vida do T cruzi Os
faacutermacos selecionados foram Vigabatrina Pregabalina MK-801 MTEP e Memantina Os
resultados demonstram que apenas Memantina apresentou um melhor efeito tripanocida
Inibiu a proliferaccedilatildeo de formas epimastigotas interferiu na metaciclogecircnese como tambeacutem
afetou o metabolismo energeacutetico do parasita com a diminuiccedilatildeo dos niacuteveis de ATP aleacutem de
desencadear mecanismos que levam a apoptose das formas epimastigotas Memantina
tambeacutem interferiu no ciclo intracelular do parasita mais especificamente no estaacutegio
amastigota Interessantemente as fases do ciclo do parasita que satildeo mais afetadas satildeo as fases
que requerem mais energia estaacutegios replicativos (amastigota e epimastigota) como tambeacutem
os processos de diferenciaccedilatildeo e invasatildeo celular Apesar de apresentar valores de IC50 que
podem ser considerados altos em relaccedilatildeo aos valores descritos para formas epimastigotas
tratadas com Benzonidazol esses resultados mostram perspectivas promissoras visto que o
Memantina poderaacute ser utilizado como composto liacuteder para o desenho de derivados com
atividade tripanocida otimizada
Palavras-chave Trypanosoma cruzi Memantina Receptor de glutamato
ABSTRACT
Damasceno FS Study of interference of drugs that act via receptors andor transport of
GABA and glutamate in the life cycle of Trypanosoma cruzi [Masters thesis (Host-Pathogen
Relationship Biology)] Satildeo Paulo Instituto de Ciecircncias Biomeacutedicas Universidade de Satildeo
Paulo 2013
Chagas disease also known as American trypanosomiasis is caused by the flagellate
protozoan Trypanosoma cruzi and affects approximately 10 million people in endemic areas
of Mexico Central America and South America with about 25 million people living in areas
at risk of infection The currently available chemotherapy is limited to two compounds which
are Nifurtimox and Benznidazole Both drugs reduce symptoms of the disease and mortality
of infected people when used in the acute phase but its efficacy in chronic phase (phase in
which the majority of cases are diagnosed) is still controversial Moreover these drugs have
several side effects T cruzi is able to utilize carbohydrate and amino acids as carbon and
energy source Our group has studied the involvement of glutamate in parasite resistance to
thermal oxidative and metabolic stress as well as the importance of the amino acid GABA
metabolism in T cruzi The aim of this study was to evaluate the interference of drugs that
act in GABA and glutamate transporters or receptors in the life cycle of T cruzi The drugs
selected were vigabatrin pregabalin MK-801 memantine and MTEP The results showed
that only memantine has a better trypanocidal effect Memantine inhibited the proliferation of
epimastigotes interfered in metacyclogenesis and affected the energy metabolism of the
parasite with decreased levels of ATP and trigger mechanisms that lead to apoptosis of
epimastigotes Moreover interferes with intracellular cycle of the parasite specifically in
amastigote stage Interesting that stages of the parasite that require more energy are more
affected replicative stages (epimastigote and amastigote) as well as the processes of
differentiation and cell invasion IC50 value is more than that described to epimastigotes
treateds with Benznidazole but Memantine could be used as leader compound for the design
of drugs with optimized trypanocidal activity
Keywords Trypanosoma cruzi Memantine Glutamate receptor
LISTA DE ILUSTRACcedilOtildeES
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi 19
Figura 2 - Estruturas quiacutemicas do Benzonidazol e Nifurtimox 24
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica 28
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica 29
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse 40
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os
tratamentos com MK-801 ou Memantina 41
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina 42
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico ou oxidativo 44
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na
presenccedila do Memantina 45
Figura 10 - Efeito da Memantina na metaciclogecircnese 46
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo
de morte celular 48
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2 49
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
50
Figura 14 - Formas epimastigotas de T cruzi 51
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi 53
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade
de formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo
celular 54
LISTA DE ABREVIATURAS
ANOVA Anaacutelise de variacircncia
ATP Adenosina trifosfato
CHO-K1 Chinese Hamster Ovary (linhagem celular proveniente de ovaacuterio de
hamester chinecircs)
Ca2+
Caacutelcio
CO2 Dioacutexido de carbono
DNA Aacutecido desoxirribonucleacuteico
DO Densidade oacutetica
DTUs Discrete Typing Units
EGTA Aacutecido etilenoglicol-bis-aminoetiletertetraceacutetico
EROs Espeacutecies reativas de oxigecircnio
GABA Aminoaacutecido γ-aminobutiacuterico
GABA T Aminoaacutecido γ-aminobutiacuterico transaminase
GAPDH Gliceraldeiacutedo -3-fosfato desidrogenase
H2O2 Peroacutexido de hidrogecircnio
IC50 Metade da concentraccedilatildeo maacutexima inibitoacuteria
KCl Cloreto de potaacutessio
LIT Liver Infusion-Tryptose (Meio de cultura)
MCP Morte celular programada
mg Miligramas
mGluR Receptor metabotroacutepico de glutamato
MTT (3-(45-dimetiltiazol-2-il)-25 difenil bromo tetrazolio)
Na2HPO4 Fosfato de soacutedio monobaacutesico anidro
NaCl Cloreto de soacutedio
NaHCO3 Bicarbonato de soacutedio
NCCD Nomenclature Comittee on Cell Death (Comite de Nomeclatura de Morte
Celular)
NH4+
Ion amocircnio
NMDA N-metil-D-aspartato
OH Radical hidroxila
PBS Soluccedilatildeo salina tamponada com fosfato
pH Potencial hidrogeniocircnico
RPMI Roswell Park Memorial Institute
SDS Soacutedio dodecil sulfat
SFB Soro fetal bovino
U Unidades
μg Micrograma
μl Microlitro
μm Microcircmetro
SUMAacuteRIO
1 INTRODUCcedilAtildeO 16
11 Aspectos gerais da doenccedila de chagas 16
12 Trypanosoma cruzi 18
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos 20
14 Morte celular em Trypanosoma cruzi 21
15 Terapia na doenccedila de Chagas 23
16 Perspectivas de novos faacutermacos 25
2 OBJETIVOS 30
21 Objetivo geral 30
22 Objetivos especiacuteficos 30
3 MATERIAIS E MEacuteTODOS 31
31 Microorganismos utilizados no trabalho 31
311 Ceacutelulas hospedeiras 31
312 Trypanosoma cruzi 31
32 Manutenccedilatildeo das ceacutelulas hospedeiras 31
33 Obtenccedilatildeo dos parasitas 31
331 Epimastigotas 31
332 Curvas de crescimento de epimastigotas de T cruzi 31
333 Formas tripomastigotas 32
34 Resistecircncia a diferentes tipos de estresse 32
341 Ensaio de estresse teacutermico em formas epimastigotas 32
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico 33
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular 33
343 Ensaio de estresse oxidativo em formas epimastigotas 33
35 Tipo de morte celular em parasitas tratados com Memantina 34
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita 34
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas
epimastigotas 35
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio 35
36 Efeito sobre a metaciclogecircnese 35
37 Anaacutelise do efeito dos compostos no ciclo intracelular do T cruzi 36
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos 36
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas 36
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas
tripomastigotas 37
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os
compostos 37
38 Anaacutelises estatiacutesticas 38
4 RESULTADOS 39
41 Curvas de crescimento do T cruzi na presenccedila dos compostos 39
42 Interaccedilatildeo com condiccedilotildees de estresse 41
43 Efeito da Memantina na metaciclogecircnese 46
44 Tipo de morte celular sofrida pelo parasita tratado com Memantina 47
45 Efeito da Memantina no ciclo intracelular do T cruzi 52
5 DISCUSSAtildeO 55
6 CONCLUSOtildeES 59
REFEREcircNCIAS 61
16
1 INTRODUCcedilAtildeO
11 Aspectos gerais da doenccedila de chagas
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana foi
descoberta pelo meacutedico brasileiro Carlos Chagas (Chagas 1909) Esta doenccedila eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica Central e do Sul com cerca de 25 milhotildees de pessoas
vivendo em aacutereas de risco de infecccedilatildeo Ocorrem por ano aproximadamente 40 mil novos
casos e 12 mil mortes devido a complicaccedilotildees cardiacuteacas e digestivas (World Health
Organization 2012) Programas de controle do vetor desenvolvidos nos uacuteltimos 20 anos
como tambeacutem um maior controle nos bancos de sangue tem reduzido o nuacutemero de novos
casos da doenccedila de Chagas na Ameacuterica Latina (Rassi Marin-Neto 2010) Devido as intensas
migraccedilotildees de pessoas de paiacuteses endecircmicos o nuacutemero de casos de indiviacuteduos infectados tem
aumentado em aacutereas consideradas natildeo endecircmicas tais como Estados Unidos Canadaacute e
algumas regiotildees da Europa (Bern et al 2009 Maguire 2006 Schmunis 2007) A doenccedila eacute
uma zoonose que tem como reservatoacuterio natural do T cruzi uma ampla variedade de
mamiacuteferos do continente americano (Urbina Docampo 2003) Eacute transmitida para os seres
humanos e para mais de 150 espeacutecies de animais domeacutesticos e animais selvagens
principalmente por insetos hematoacutefagos da famiacutelia Reduviidae Apesar de existirem mais de
130 espeacutecies de triatomiacuteneos satildeo trecircs as mais importantes na transmissatildeo do T cruzi para o
homem Triatoma infestans Rhodnius prolixus e Triatoma dimidiata (Rassi Marin-Neto
2010) O parasita tambeacutem pode ser transmitido por transfusatildeo de sangue congenitamente (de
matildees infectadas para receacutem-nascidos) por transplante de oacutergatildeos ou medula oacutessea Tambeacutem
pode ocorrer por acidentes em laboratoacuterios que trabalham com o parasita vivo via esta que
epidemiologicamente eacute pouco relevante Outra forma de transmissatildeo eacute por via oral atraveacutes de
alimentos contaminados essa via vecircm sendo considerada como relevante na manutenccedilatildeo do
ciclo de transmissatildeo natural (Pereira et al 2009 Rassi Marin-Neto 2010)
A evoluccedilatildeo cliacutenica da doenccedila de Chagas pode ser dividida em duas fases aguda e
crocircnica A fase aguda normalmente eacute assintomaacutetica ou apresenta sintomatologia discreta que
na maioria dos casos passa despercebida pelo paciente Os sintomas quando aparecem o
fazem uma ou duas semanas apoacutes a exposiccedilatildeo ao triatomiacuteneo infectado ou poucos meses apoacutes
a transfusatildeo com sangue contaminado (Rassi Marin-Neto 2010) Os sintomas mais
frequentemente apresentados pelos pacientes agudos satildeo febre prolongada mal-estar
17
distensatildeo do fiacutegado baccedilo e linfonodos Pode haver tambeacutem edema subcutacircneo localizado ou
generalizado No caso particular de transmissatildeo vetorial o paciente pode apresentar um sinal
da porta de entrada do T cruzi na pele (chagoma) ou via mucosa ocular (sinal de Romantildea) As
manifestaccedilotildees da fase aguda se resolvem espontaneamente em aproximadamente 90 dos
indiviacuteduos infectados mesmo sem o tratamento com drogas tripanocidas Essa fase eacute
caracterizada pela alta parasitemia e ausecircncia de anticorpos especiacuteficos Nessa fase os
parasitas podem ser encontrados em praticamente todos os tecidos estando majoritariamente
presentes no sangue Por ser uma fase de curta duraccedilatildeo sem sintomatologia aparente ou com
sintomatologia pouco definida haacute um pequeno nuacutemero de casos detectados sendo portanto a
fase menos estudada (Rassi Marin-Neto 2010 Silber et al 2005b)
Logo apoacutes a fase aguda tem iniacutecio a fase crocircnica caracterizada pelos altos tiacutetulos de
anticorpos no sangue do paciente e pelo fato de que a parasitemia eacute inaparente O paciente
pode permanecer por vaacuterios anos ou ateacute mesmo pelo resto da vida sem apresentar sinais ou
sintomas Essa forma da fase crocircnica eacute chamada de indeterminada Aproximadamente 30-40
dos pacientes crocircnicos indeterminados desenvolvem a sintomatologia caracteriacutestica da
doenccedila evoluindo para a chamada fase crocircnica sintomaacutetica Nesta fase as trecircs principais
formas satildeo a cardiacuteaca a digestiva e a cardiodigestiva (Acquatella 2007 Marin-Neto et al
1999) Na forma cardiacuteaca o coraccedilatildeo eacute um dos primeiros oacutergatildeos afetados natildeo necessariamente
o uacutenico desenvolvendo hipertrofia e dilataccedilatildeo Na forma digestiva o trato digestivo apresenta
alteraccedilotildees anatocircmicas e fisioloacutegicas importantes sendo a principal altereccedilatildeo a dilataccedilatildeo do
intestino e do esocircfago que podem apresentar aspecto de megaviacutescera (Marin-Neto et al
1999 Rassi et al 2000) revisado por (Boscardin et al 2009 Rassi Marin-Neto 2010)
18
12 Trypanosoma cruzi
Trypanosoma cruzi eacute um parasita intracelular obrigatoacuterio classicamente localizado na
ordem kinetoplastida gecircnero Trypanosoma que causa uma infecccedilatildeo crocircnica em humanos e
em grande nuacutemero de outras espeacutecies de mamiacuteferos (Hoare 1972) Recentemente devido agrave
alta variabilidade bioloacutegica bioquiacutemica e molecular a espeacutecie Trypanosoma cruzi foi
dividida em seis linhagens subsespeciacuteficas (Discrete Typing Units ndash DTUs) numerados de I a
VI Esta classificaccedilatildeo foi realizada com base em caracteriacutesticas geneacuteticas moleculares e
imunoloacutegicas com o objetivo de unificar a nomenclatura e facilitar a comunicaccedilatildeo dentro da
comunidade cientiacutefica (Zingales et al 2009)
O T cruzi tem um ciclo de vida complexo que alterna entre o inseto vetor e o
hospedeiro mamiacutefero O parasita apresenta diferentes formas correspondentes a diferentes
estaacutegios do seu ciclo de vida Estes diferentes estaacutegios tecircm sido descritos com base em
criteacuterios morfoloacutegicos como a forma fusiforme ou arredondados a regiatildeo onde emerge o
flagelo e a posiccedilatildeo do cinetoplasto (estrutura contendo o DNA da uacutenica mitococircndria presente
no parasita) em relaccedilatildeo ao nuacutecleo No hospedeiro mamiacutefero foram descritas as formas
amastigota epimastigota intracelular e tripomastigota sanguiacutecola A forma amastigota eacute um
estaacutegio replicativo e infectivo Satildeo ceacutelulas arredondadas medindo de 2-6 microm de diacircmetro
encontradas no citoplasma das ceacutelulas infectadas Os epimastigotas intracelulares
(epimastigote-like) satildeo formas replicativas intermediaacuterias entre amastigotas e tripomastigotas
tambeacutem encontrados dentro das ceacutelulas medindo aproximadamente 54 microm Essas formas satildeo
menores que as formas epimastigotas encontradas no inseto vetor poreacutem compartilha com as
mesmas padrotildees morfomeacutetricos e diversas caracteriacutesticas bioquiacutemicas e bioloacutegicas (Almeida-
De-Faria et al 1999) O tripomastigota sanguiacutecola eacute um estaacutegio flagelado infectivo natildeo
replicativo encontrado principalmente na corrente sanguiacutenea do hospedeiro mamiacutefero Mede
aproximadamente 18 microm de comprimento incluindo um extremo livre do flagelo de
aproximadamente 6 microm e 2-3 microm de largura
No inseto vetor encontram-se predominantemente as formas epimastigota e
tripomastigota metaciacuteclico Epimastigota eacute uma forma flagelada replicativa e natildeo infectiva
Mede aproximadamente 20-40 microm de comprimento e 2-5 microm de largura essas formas estatildeo
presentes tipicamente na porccedilatildeo meacutedia do intestino do hospedeiro invertebrado
Tripomastigota metaciacuteclico eacute uma forma infectiva natildeo replicativa encontrada na porccedilatildeo final
do intestino do inseto vetor (Brener 1971) revisado por (Silber et al 2005a Tyler Engman
2001)
19
O ciclo de vida do T cruzi inicia-se quando os parasitas presentes na corrente
sanguiacutenea do hospedeiro vertebrado infectado passam ao inseto vetor durante o repasto
sanguiacuteneo No intestino do inseto estas formas se diferenciam em formas epimastigotas (De
Souza et al 2002) que apoacutes multiplicaccedilatildeo por fissatildeo binaacuteria colonizam a porccedilatildeo terminal do
intestino e se diferenciam novamente em tripomastigotas metaciacuteclicos Quando o inseto
infectado pica o hospedeiro vertebrado e inicia o repasto sanguiacuteneo concomitantemente
elimina as formas tripomastigotas metaciacuteclicos junto com as fezes e urina Uma vez em
contato com a pele ou mucosas do hospedeiro mamiacutefero os tripomastigotas satildeo internalizados
atraveacutes de pequenas lesotildees na pele causadas pelo proacuteprio hospedeiro (Alves Colli 2008) Os
tripomastigotas invadem as ceacutelulas do hospedeiro mamiacutefero e se diferenciam em formas
replicativas denominadas amastigotas estabelecendo assim a infecccedilatildeo Apoacutes
aproximadamente 96 horas os amastigotas comeccedilam a se diferenciar em tripomastigotas
passando pela forma intermediaacuteria epimastigota intracelular Estas formas tripomastigotas satildeo
liberadas na corrente sanguiacutenea podendo invadir novas ceacutelulas ou serem ingeridas pelo inseto
vetor em um novo repasto sanguiacuteneo fechando assim o seu ciclo de vida O esquema do ciclo
de vida do Trypanosoma cruzi estaacute representado na Figura 1
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi
20
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos
T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono As formas epimastigotas preferencialmente catabolizam glicose e apoacutes a exaustatildeo
do carboidrato ocorre uma mudanccedila para o metabolismo de aminoaacutecidos A prolina e a
glutamina satildeo os principais aminoaacutecidos metabolizados pelas formas epimastigotas visto que
satildeo abudantes na hemolinfa e nos fluiacutedos dos tecidos do inseto vetor Esse metabolismo
ocorre com produccedilatildeo de NH4+
(produto da deaminaccedilatildeo dos aminoaacutecidos) que eacute excretada para
o meio extracelular (Bringaud et al 2006 Cazzulo 1994) As formas tripomastigotas
utilizam principalmente a glicose que eacute abundante nos fluiacutedos de seus hospedeiros
vertebrados Jaacute as formas amastigotas encontradas no citoplasma das ceacutelulas do hospedeiro
mamiacutefero ambiente pobre em glicose livre usam os aminoaacutecidos como principal fonte de
carbono e energia apresentando maior quantidade de prolina intracelular do que as formas
tripomastigotas (Silber et al 2009)
Entre os aminoaacutecidos metabolizados pelas formas epimastigotas estatildeo prolina glicina
asparagina glutamina aspartato glutamato leucina e isoleucina (Mancilla et al 1967 Odaly
et al 1983 Silber et al 2005a Sylvester Krassner 1976 Zeledon 1960) Os aminoaacutecidos
estatildeo envolvidos tambeacutem em outros processos importantes do ciclo de vida do T cruzi O
aminoaacutecido L-prolina eacute essencial na diferenciaccedilatildeo das formas epimastigota intracelular para
tripomastigotas aleacutem de estarem envolvidos nos processos de osmorregulaccedilatildeo crescimento e
resistecircncia do parasita ao estresse oxidativo e nutricional (Homsy et al 1989 Magdaleno et
al 2009 Magdaleno et al 2011 Pereira et al 2002 Pereira et al 2003 Tonelli et al
2004) Glutamato prolina e aspartato satildeo importantes na metaciclogecircnese ou seja na
diferenciaccedilatildeo de formas epimastigotas para tripomastigotas metaciacuteclicos (Contreras et al
1985) Prolina glicina alanina e glutamato satildeo descritos como importantes nos processos de
osmorregulaccedilatildeo e controle do volume celular (Rohloff et al 2003 Rohloff et al 2004a)
Os aminoaacutecidos para que sejam metabolizados ou cumprirem outras funccedilotildees devem
ser obtidos a partir da degradaccedilatildeo de proteiacutenas sintetizados ou incorporados pelo parasita
atraveacutes de transportadores localizados na membrana (Silber et al 2005a) O primeiro
transportador de aminoaacutecidos em T cruzi foi descrito bioquimicamente em 1970 o
transportador de lisina (Hampton 1970) Outros aminoaacutecidos tambeacutem jaacute tiveram a sua
cineacutetica e seu mecanismo de transporte descrito para T cruzi A incorporaccedilatildeo de arginina
pelo parasita apresenta pelo menos dois sistemas de transporte sendo um de alta afinidade e
outro de baixa afinidade Esse transporte eacute regulado ao longo do ciclo de vida do parasita
21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

RESUMO
Damasceno FS Estudo da interferecircncia de faacutermacos que atuam via receptores eou
transportadores de GABA e Glutamato no ciclo de vida do Trypanosoma cruzi [dissertaccedilatildeo
(Mestrado em Biologia da Relaccedilatildeo Patoacutegeno-Hospedeiro)] Satildeo Paulo Instituto de Ciecircncias
Biomeacutedicas Universidade de Satildeo Paulo 2013
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica central e do Sul A quimioterapia disponiacutevel atualmente
limita-se a dois compostos Nifurtimox e o Benzonidazol Ambos os faacutermacos reduzem os
sintomas da doenccedila e a mortalidade das pessoas infectadas quando utilizadas na fase aguda
mas sua eficaacutecia na fase crocircnica eacute controversa Aleacutem do mais apresentam vaacuterios efeitos
colaterais O T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico como tambeacutem a importacircncia do aminoaacutecido
GABA no metabolismo do T cruzi visto que este tambeacutem eacute transportado pelo parasita Nesse
contexto o objetivo desse trabalho foi avaliar a interferecircncia de faacutermacos que atuam via
receptores ou transportadores de GABA e glutamato no ciclo de vida do T cruzi Os
faacutermacos selecionados foram Vigabatrina Pregabalina MK-801 MTEP e Memantina Os
resultados demonstram que apenas Memantina apresentou um melhor efeito tripanocida
Inibiu a proliferaccedilatildeo de formas epimastigotas interferiu na metaciclogecircnese como tambeacutem
afetou o metabolismo energeacutetico do parasita com a diminuiccedilatildeo dos niacuteveis de ATP aleacutem de
desencadear mecanismos que levam a apoptose das formas epimastigotas Memantina
tambeacutem interferiu no ciclo intracelular do parasita mais especificamente no estaacutegio
amastigota Interessantemente as fases do ciclo do parasita que satildeo mais afetadas satildeo as fases
que requerem mais energia estaacutegios replicativos (amastigota e epimastigota) como tambeacutem
os processos de diferenciaccedilatildeo e invasatildeo celular Apesar de apresentar valores de IC50 que
podem ser considerados altos em relaccedilatildeo aos valores descritos para formas epimastigotas
tratadas com Benzonidazol esses resultados mostram perspectivas promissoras visto que o
Memantina poderaacute ser utilizado como composto liacuteder para o desenho de derivados com
atividade tripanocida otimizada
Palavras-chave Trypanosoma cruzi Memantina Receptor de glutamato
ABSTRACT
Damasceno FS Study of interference of drugs that act via receptors andor transport of
GABA and glutamate in the life cycle of Trypanosoma cruzi [Masters thesis (Host-Pathogen
Relationship Biology)] Satildeo Paulo Instituto de Ciecircncias Biomeacutedicas Universidade de Satildeo
Paulo 2013
Chagas disease also known as American trypanosomiasis is caused by the flagellate
protozoan Trypanosoma cruzi and affects approximately 10 million people in endemic areas
of Mexico Central America and South America with about 25 million people living in areas
at risk of infection The currently available chemotherapy is limited to two compounds which
are Nifurtimox and Benznidazole Both drugs reduce symptoms of the disease and mortality
of infected people when used in the acute phase but its efficacy in chronic phase (phase in
which the majority of cases are diagnosed) is still controversial Moreover these drugs have
several side effects T cruzi is able to utilize carbohydrate and amino acids as carbon and
energy source Our group has studied the involvement of glutamate in parasite resistance to
thermal oxidative and metabolic stress as well as the importance of the amino acid GABA
metabolism in T cruzi The aim of this study was to evaluate the interference of drugs that
act in GABA and glutamate transporters or receptors in the life cycle of T cruzi The drugs
selected were vigabatrin pregabalin MK-801 memantine and MTEP The results showed
that only memantine has a better trypanocidal effect Memantine inhibited the proliferation of
epimastigotes interfered in metacyclogenesis and affected the energy metabolism of the
parasite with decreased levels of ATP and trigger mechanisms that lead to apoptosis of
epimastigotes Moreover interferes with intracellular cycle of the parasite specifically in
amastigote stage Interesting that stages of the parasite that require more energy are more
affected replicative stages (epimastigote and amastigote) as well as the processes of
differentiation and cell invasion IC50 value is more than that described to epimastigotes
treateds with Benznidazole but Memantine could be used as leader compound for the design
of drugs with optimized trypanocidal activity
Keywords Trypanosoma cruzi Memantine Glutamate receptor
LISTA DE ILUSTRACcedilOtildeES
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi 19
Figura 2 - Estruturas quiacutemicas do Benzonidazol e Nifurtimox 24
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica 28
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica 29
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse 40
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os
tratamentos com MK-801 ou Memantina 41
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina 42
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico ou oxidativo 44
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na
presenccedila do Memantina 45
Figura 10 - Efeito da Memantina na metaciclogecircnese 46
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo
de morte celular 48
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2 49
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
50
Figura 14 - Formas epimastigotas de T cruzi 51
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi 53
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade
de formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo
celular 54
LISTA DE ABREVIATURAS
ANOVA Anaacutelise de variacircncia
ATP Adenosina trifosfato
CHO-K1 Chinese Hamster Ovary (linhagem celular proveniente de ovaacuterio de
hamester chinecircs)
Ca2+
Caacutelcio
CO2 Dioacutexido de carbono
DNA Aacutecido desoxirribonucleacuteico
DO Densidade oacutetica
DTUs Discrete Typing Units
EGTA Aacutecido etilenoglicol-bis-aminoetiletertetraceacutetico
EROs Espeacutecies reativas de oxigecircnio
GABA Aminoaacutecido γ-aminobutiacuterico
GABA T Aminoaacutecido γ-aminobutiacuterico transaminase
GAPDH Gliceraldeiacutedo -3-fosfato desidrogenase
H2O2 Peroacutexido de hidrogecircnio
IC50 Metade da concentraccedilatildeo maacutexima inibitoacuteria
KCl Cloreto de potaacutessio
LIT Liver Infusion-Tryptose (Meio de cultura)
MCP Morte celular programada
mg Miligramas
mGluR Receptor metabotroacutepico de glutamato
MTT (3-(45-dimetiltiazol-2-il)-25 difenil bromo tetrazolio)
Na2HPO4 Fosfato de soacutedio monobaacutesico anidro
NaCl Cloreto de soacutedio
NaHCO3 Bicarbonato de soacutedio
NCCD Nomenclature Comittee on Cell Death (Comite de Nomeclatura de Morte
Celular)
NH4+
Ion amocircnio
NMDA N-metil-D-aspartato
OH Radical hidroxila
PBS Soluccedilatildeo salina tamponada com fosfato
pH Potencial hidrogeniocircnico
RPMI Roswell Park Memorial Institute
SDS Soacutedio dodecil sulfat
SFB Soro fetal bovino
U Unidades
μg Micrograma
μl Microlitro
μm Microcircmetro
SUMAacuteRIO
1 INTRODUCcedilAtildeO 16
11 Aspectos gerais da doenccedila de chagas 16
12 Trypanosoma cruzi 18
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos 20
14 Morte celular em Trypanosoma cruzi 21
15 Terapia na doenccedila de Chagas 23
16 Perspectivas de novos faacutermacos 25
2 OBJETIVOS 30
21 Objetivo geral 30
22 Objetivos especiacuteficos 30
3 MATERIAIS E MEacuteTODOS 31
31 Microorganismos utilizados no trabalho 31
311 Ceacutelulas hospedeiras 31
312 Trypanosoma cruzi 31
32 Manutenccedilatildeo das ceacutelulas hospedeiras 31
33 Obtenccedilatildeo dos parasitas 31
331 Epimastigotas 31
332 Curvas de crescimento de epimastigotas de T cruzi 31
333 Formas tripomastigotas 32
34 Resistecircncia a diferentes tipos de estresse 32
341 Ensaio de estresse teacutermico em formas epimastigotas 32
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico 33
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular 33
343 Ensaio de estresse oxidativo em formas epimastigotas 33
35 Tipo de morte celular em parasitas tratados com Memantina 34
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita 34
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas
epimastigotas 35
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio 35
36 Efeito sobre a metaciclogecircnese 35
37 Anaacutelise do efeito dos compostos no ciclo intracelular do T cruzi 36
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos 36
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas 36
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas
tripomastigotas 37
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os
compostos 37
38 Anaacutelises estatiacutesticas 38
4 RESULTADOS 39
41 Curvas de crescimento do T cruzi na presenccedila dos compostos 39
42 Interaccedilatildeo com condiccedilotildees de estresse 41
43 Efeito da Memantina na metaciclogecircnese 46
44 Tipo de morte celular sofrida pelo parasita tratado com Memantina 47
45 Efeito da Memantina no ciclo intracelular do T cruzi 52
5 DISCUSSAtildeO 55
6 CONCLUSOtildeES 59
REFEREcircNCIAS 61
16
1 INTRODUCcedilAtildeO
11 Aspectos gerais da doenccedila de chagas
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana foi
descoberta pelo meacutedico brasileiro Carlos Chagas (Chagas 1909) Esta doenccedila eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica Central e do Sul com cerca de 25 milhotildees de pessoas
vivendo em aacutereas de risco de infecccedilatildeo Ocorrem por ano aproximadamente 40 mil novos
casos e 12 mil mortes devido a complicaccedilotildees cardiacuteacas e digestivas (World Health
Organization 2012) Programas de controle do vetor desenvolvidos nos uacuteltimos 20 anos
como tambeacutem um maior controle nos bancos de sangue tem reduzido o nuacutemero de novos
casos da doenccedila de Chagas na Ameacuterica Latina (Rassi Marin-Neto 2010) Devido as intensas
migraccedilotildees de pessoas de paiacuteses endecircmicos o nuacutemero de casos de indiviacuteduos infectados tem
aumentado em aacutereas consideradas natildeo endecircmicas tais como Estados Unidos Canadaacute e
algumas regiotildees da Europa (Bern et al 2009 Maguire 2006 Schmunis 2007) A doenccedila eacute
uma zoonose que tem como reservatoacuterio natural do T cruzi uma ampla variedade de
mamiacuteferos do continente americano (Urbina Docampo 2003) Eacute transmitida para os seres
humanos e para mais de 150 espeacutecies de animais domeacutesticos e animais selvagens
principalmente por insetos hematoacutefagos da famiacutelia Reduviidae Apesar de existirem mais de
130 espeacutecies de triatomiacuteneos satildeo trecircs as mais importantes na transmissatildeo do T cruzi para o
homem Triatoma infestans Rhodnius prolixus e Triatoma dimidiata (Rassi Marin-Neto
2010) O parasita tambeacutem pode ser transmitido por transfusatildeo de sangue congenitamente (de
matildees infectadas para receacutem-nascidos) por transplante de oacutergatildeos ou medula oacutessea Tambeacutem
pode ocorrer por acidentes em laboratoacuterios que trabalham com o parasita vivo via esta que
epidemiologicamente eacute pouco relevante Outra forma de transmissatildeo eacute por via oral atraveacutes de
alimentos contaminados essa via vecircm sendo considerada como relevante na manutenccedilatildeo do
ciclo de transmissatildeo natural (Pereira et al 2009 Rassi Marin-Neto 2010)
A evoluccedilatildeo cliacutenica da doenccedila de Chagas pode ser dividida em duas fases aguda e
crocircnica A fase aguda normalmente eacute assintomaacutetica ou apresenta sintomatologia discreta que
na maioria dos casos passa despercebida pelo paciente Os sintomas quando aparecem o
fazem uma ou duas semanas apoacutes a exposiccedilatildeo ao triatomiacuteneo infectado ou poucos meses apoacutes
a transfusatildeo com sangue contaminado (Rassi Marin-Neto 2010) Os sintomas mais
frequentemente apresentados pelos pacientes agudos satildeo febre prolongada mal-estar
17
distensatildeo do fiacutegado baccedilo e linfonodos Pode haver tambeacutem edema subcutacircneo localizado ou
generalizado No caso particular de transmissatildeo vetorial o paciente pode apresentar um sinal
da porta de entrada do T cruzi na pele (chagoma) ou via mucosa ocular (sinal de Romantildea) As
manifestaccedilotildees da fase aguda se resolvem espontaneamente em aproximadamente 90 dos
indiviacuteduos infectados mesmo sem o tratamento com drogas tripanocidas Essa fase eacute
caracterizada pela alta parasitemia e ausecircncia de anticorpos especiacuteficos Nessa fase os
parasitas podem ser encontrados em praticamente todos os tecidos estando majoritariamente
presentes no sangue Por ser uma fase de curta duraccedilatildeo sem sintomatologia aparente ou com
sintomatologia pouco definida haacute um pequeno nuacutemero de casos detectados sendo portanto a
fase menos estudada (Rassi Marin-Neto 2010 Silber et al 2005b)
Logo apoacutes a fase aguda tem iniacutecio a fase crocircnica caracterizada pelos altos tiacutetulos de
anticorpos no sangue do paciente e pelo fato de que a parasitemia eacute inaparente O paciente
pode permanecer por vaacuterios anos ou ateacute mesmo pelo resto da vida sem apresentar sinais ou
sintomas Essa forma da fase crocircnica eacute chamada de indeterminada Aproximadamente 30-40
dos pacientes crocircnicos indeterminados desenvolvem a sintomatologia caracteriacutestica da
doenccedila evoluindo para a chamada fase crocircnica sintomaacutetica Nesta fase as trecircs principais
formas satildeo a cardiacuteaca a digestiva e a cardiodigestiva (Acquatella 2007 Marin-Neto et al
1999) Na forma cardiacuteaca o coraccedilatildeo eacute um dos primeiros oacutergatildeos afetados natildeo necessariamente
o uacutenico desenvolvendo hipertrofia e dilataccedilatildeo Na forma digestiva o trato digestivo apresenta
alteraccedilotildees anatocircmicas e fisioloacutegicas importantes sendo a principal altereccedilatildeo a dilataccedilatildeo do
intestino e do esocircfago que podem apresentar aspecto de megaviacutescera (Marin-Neto et al
1999 Rassi et al 2000) revisado por (Boscardin et al 2009 Rassi Marin-Neto 2010)
18
12 Trypanosoma cruzi
Trypanosoma cruzi eacute um parasita intracelular obrigatoacuterio classicamente localizado na
ordem kinetoplastida gecircnero Trypanosoma que causa uma infecccedilatildeo crocircnica em humanos e
em grande nuacutemero de outras espeacutecies de mamiacuteferos (Hoare 1972) Recentemente devido agrave
alta variabilidade bioloacutegica bioquiacutemica e molecular a espeacutecie Trypanosoma cruzi foi
dividida em seis linhagens subsespeciacuteficas (Discrete Typing Units ndash DTUs) numerados de I a
VI Esta classificaccedilatildeo foi realizada com base em caracteriacutesticas geneacuteticas moleculares e
imunoloacutegicas com o objetivo de unificar a nomenclatura e facilitar a comunicaccedilatildeo dentro da
comunidade cientiacutefica (Zingales et al 2009)
O T cruzi tem um ciclo de vida complexo que alterna entre o inseto vetor e o
hospedeiro mamiacutefero O parasita apresenta diferentes formas correspondentes a diferentes
estaacutegios do seu ciclo de vida Estes diferentes estaacutegios tecircm sido descritos com base em
criteacuterios morfoloacutegicos como a forma fusiforme ou arredondados a regiatildeo onde emerge o
flagelo e a posiccedilatildeo do cinetoplasto (estrutura contendo o DNA da uacutenica mitococircndria presente
no parasita) em relaccedilatildeo ao nuacutecleo No hospedeiro mamiacutefero foram descritas as formas
amastigota epimastigota intracelular e tripomastigota sanguiacutecola A forma amastigota eacute um
estaacutegio replicativo e infectivo Satildeo ceacutelulas arredondadas medindo de 2-6 microm de diacircmetro
encontradas no citoplasma das ceacutelulas infectadas Os epimastigotas intracelulares
(epimastigote-like) satildeo formas replicativas intermediaacuterias entre amastigotas e tripomastigotas
tambeacutem encontrados dentro das ceacutelulas medindo aproximadamente 54 microm Essas formas satildeo
menores que as formas epimastigotas encontradas no inseto vetor poreacutem compartilha com as
mesmas padrotildees morfomeacutetricos e diversas caracteriacutesticas bioquiacutemicas e bioloacutegicas (Almeida-
De-Faria et al 1999) O tripomastigota sanguiacutecola eacute um estaacutegio flagelado infectivo natildeo
replicativo encontrado principalmente na corrente sanguiacutenea do hospedeiro mamiacutefero Mede
aproximadamente 18 microm de comprimento incluindo um extremo livre do flagelo de
aproximadamente 6 microm e 2-3 microm de largura
No inseto vetor encontram-se predominantemente as formas epimastigota e
tripomastigota metaciacuteclico Epimastigota eacute uma forma flagelada replicativa e natildeo infectiva
Mede aproximadamente 20-40 microm de comprimento e 2-5 microm de largura essas formas estatildeo
presentes tipicamente na porccedilatildeo meacutedia do intestino do hospedeiro invertebrado
Tripomastigota metaciacuteclico eacute uma forma infectiva natildeo replicativa encontrada na porccedilatildeo final
do intestino do inseto vetor (Brener 1971) revisado por (Silber et al 2005a Tyler Engman
2001)
19
O ciclo de vida do T cruzi inicia-se quando os parasitas presentes na corrente
sanguiacutenea do hospedeiro vertebrado infectado passam ao inseto vetor durante o repasto
sanguiacuteneo No intestino do inseto estas formas se diferenciam em formas epimastigotas (De
Souza et al 2002) que apoacutes multiplicaccedilatildeo por fissatildeo binaacuteria colonizam a porccedilatildeo terminal do
intestino e se diferenciam novamente em tripomastigotas metaciacuteclicos Quando o inseto
infectado pica o hospedeiro vertebrado e inicia o repasto sanguiacuteneo concomitantemente
elimina as formas tripomastigotas metaciacuteclicos junto com as fezes e urina Uma vez em
contato com a pele ou mucosas do hospedeiro mamiacutefero os tripomastigotas satildeo internalizados
atraveacutes de pequenas lesotildees na pele causadas pelo proacuteprio hospedeiro (Alves Colli 2008) Os
tripomastigotas invadem as ceacutelulas do hospedeiro mamiacutefero e se diferenciam em formas
replicativas denominadas amastigotas estabelecendo assim a infecccedilatildeo Apoacutes
aproximadamente 96 horas os amastigotas comeccedilam a se diferenciar em tripomastigotas
passando pela forma intermediaacuteria epimastigota intracelular Estas formas tripomastigotas satildeo
liberadas na corrente sanguiacutenea podendo invadir novas ceacutelulas ou serem ingeridas pelo inseto
vetor em um novo repasto sanguiacuteneo fechando assim o seu ciclo de vida O esquema do ciclo
de vida do Trypanosoma cruzi estaacute representado na Figura 1
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi
20
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos
T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono As formas epimastigotas preferencialmente catabolizam glicose e apoacutes a exaustatildeo
do carboidrato ocorre uma mudanccedila para o metabolismo de aminoaacutecidos A prolina e a
glutamina satildeo os principais aminoaacutecidos metabolizados pelas formas epimastigotas visto que
satildeo abudantes na hemolinfa e nos fluiacutedos dos tecidos do inseto vetor Esse metabolismo
ocorre com produccedilatildeo de NH4+
(produto da deaminaccedilatildeo dos aminoaacutecidos) que eacute excretada para
o meio extracelular (Bringaud et al 2006 Cazzulo 1994) As formas tripomastigotas
utilizam principalmente a glicose que eacute abundante nos fluiacutedos de seus hospedeiros
vertebrados Jaacute as formas amastigotas encontradas no citoplasma das ceacutelulas do hospedeiro
mamiacutefero ambiente pobre em glicose livre usam os aminoaacutecidos como principal fonte de
carbono e energia apresentando maior quantidade de prolina intracelular do que as formas
tripomastigotas (Silber et al 2009)
Entre os aminoaacutecidos metabolizados pelas formas epimastigotas estatildeo prolina glicina
asparagina glutamina aspartato glutamato leucina e isoleucina (Mancilla et al 1967 Odaly
et al 1983 Silber et al 2005a Sylvester Krassner 1976 Zeledon 1960) Os aminoaacutecidos
estatildeo envolvidos tambeacutem em outros processos importantes do ciclo de vida do T cruzi O
aminoaacutecido L-prolina eacute essencial na diferenciaccedilatildeo das formas epimastigota intracelular para
tripomastigotas aleacutem de estarem envolvidos nos processos de osmorregulaccedilatildeo crescimento e
resistecircncia do parasita ao estresse oxidativo e nutricional (Homsy et al 1989 Magdaleno et
al 2009 Magdaleno et al 2011 Pereira et al 2002 Pereira et al 2003 Tonelli et al
2004) Glutamato prolina e aspartato satildeo importantes na metaciclogecircnese ou seja na
diferenciaccedilatildeo de formas epimastigotas para tripomastigotas metaciacuteclicos (Contreras et al
1985) Prolina glicina alanina e glutamato satildeo descritos como importantes nos processos de
osmorregulaccedilatildeo e controle do volume celular (Rohloff et al 2003 Rohloff et al 2004a)
Os aminoaacutecidos para que sejam metabolizados ou cumprirem outras funccedilotildees devem
ser obtidos a partir da degradaccedilatildeo de proteiacutenas sintetizados ou incorporados pelo parasita
atraveacutes de transportadores localizados na membrana (Silber et al 2005a) O primeiro
transportador de aminoaacutecidos em T cruzi foi descrito bioquimicamente em 1970 o
transportador de lisina (Hampton 1970) Outros aminoaacutecidos tambeacutem jaacute tiveram a sua
cineacutetica e seu mecanismo de transporte descrito para T cruzi A incorporaccedilatildeo de arginina
pelo parasita apresenta pelo menos dois sistemas de transporte sendo um de alta afinidade e
outro de baixa afinidade Esse transporte eacute regulado ao longo do ciclo de vida do parasita
21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

ABSTRACT
Damasceno FS Study of interference of drugs that act via receptors andor transport of
GABA and glutamate in the life cycle of Trypanosoma cruzi [Masters thesis (Host-Pathogen
Relationship Biology)] Satildeo Paulo Instituto de Ciecircncias Biomeacutedicas Universidade de Satildeo
Paulo 2013
Chagas disease also known as American trypanosomiasis is caused by the flagellate
protozoan Trypanosoma cruzi and affects approximately 10 million people in endemic areas
of Mexico Central America and South America with about 25 million people living in areas
at risk of infection The currently available chemotherapy is limited to two compounds which
are Nifurtimox and Benznidazole Both drugs reduce symptoms of the disease and mortality
of infected people when used in the acute phase but its efficacy in chronic phase (phase in
which the majority of cases are diagnosed) is still controversial Moreover these drugs have
several side effects T cruzi is able to utilize carbohydrate and amino acids as carbon and
energy source Our group has studied the involvement of glutamate in parasite resistance to
thermal oxidative and metabolic stress as well as the importance of the amino acid GABA
metabolism in T cruzi The aim of this study was to evaluate the interference of drugs that
act in GABA and glutamate transporters or receptors in the life cycle of T cruzi The drugs
selected were vigabatrin pregabalin MK-801 memantine and MTEP The results showed
that only memantine has a better trypanocidal effect Memantine inhibited the proliferation of
epimastigotes interfered in metacyclogenesis and affected the energy metabolism of the
parasite with decreased levels of ATP and trigger mechanisms that lead to apoptosis of
epimastigotes Moreover interferes with intracellular cycle of the parasite specifically in
amastigote stage Interesting that stages of the parasite that require more energy are more
affected replicative stages (epimastigote and amastigote) as well as the processes of
differentiation and cell invasion IC50 value is more than that described to epimastigotes
treateds with Benznidazole but Memantine could be used as leader compound for the design
of drugs with optimized trypanocidal activity
Keywords Trypanosoma cruzi Memantine Glutamate receptor
LISTA DE ILUSTRACcedilOtildeES
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi 19
Figura 2 - Estruturas quiacutemicas do Benzonidazol e Nifurtimox 24
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica 28
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica 29
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse 40
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os
tratamentos com MK-801 ou Memantina 41
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina 42
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico ou oxidativo 44
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na
presenccedila do Memantina 45
Figura 10 - Efeito da Memantina na metaciclogecircnese 46
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo
de morte celular 48
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2 49
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
50
Figura 14 - Formas epimastigotas de T cruzi 51
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi 53
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade
de formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo
celular 54
LISTA DE ABREVIATURAS
ANOVA Anaacutelise de variacircncia
ATP Adenosina trifosfato
CHO-K1 Chinese Hamster Ovary (linhagem celular proveniente de ovaacuterio de
hamester chinecircs)
Ca2+
Caacutelcio
CO2 Dioacutexido de carbono
DNA Aacutecido desoxirribonucleacuteico
DO Densidade oacutetica
DTUs Discrete Typing Units
EGTA Aacutecido etilenoglicol-bis-aminoetiletertetraceacutetico
EROs Espeacutecies reativas de oxigecircnio
GABA Aminoaacutecido γ-aminobutiacuterico
GABA T Aminoaacutecido γ-aminobutiacuterico transaminase
GAPDH Gliceraldeiacutedo -3-fosfato desidrogenase
H2O2 Peroacutexido de hidrogecircnio
IC50 Metade da concentraccedilatildeo maacutexima inibitoacuteria
KCl Cloreto de potaacutessio
LIT Liver Infusion-Tryptose (Meio de cultura)
MCP Morte celular programada
mg Miligramas
mGluR Receptor metabotroacutepico de glutamato
MTT (3-(45-dimetiltiazol-2-il)-25 difenil bromo tetrazolio)
Na2HPO4 Fosfato de soacutedio monobaacutesico anidro
NaCl Cloreto de soacutedio
NaHCO3 Bicarbonato de soacutedio
NCCD Nomenclature Comittee on Cell Death (Comite de Nomeclatura de Morte
Celular)
NH4+
Ion amocircnio
NMDA N-metil-D-aspartato
OH Radical hidroxila
PBS Soluccedilatildeo salina tamponada com fosfato
pH Potencial hidrogeniocircnico
RPMI Roswell Park Memorial Institute
SDS Soacutedio dodecil sulfat
SFB Soro fetal bovino
U Unidades
μg Micrograma
μl Microlitro
μm Microcircmetro
SUMAacuteRIO
1 INTRODUCcedilAtildeO 16
11 Aspectos gerais da doenccedila de chagas 16
12 Trypanosoma cruzi 18
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos 20
14 Morte celular em Trypanosoma cruzi 21
15 Terapia na doenccedila de Chagas 23
16 Perspectivas de novos faacutermacos 25
2 OBJETIVOS 30
21 Objetivo geral 30
22 Objetivos especiacuteficos 30
3 MATERIAIS E MEacuteTODOS 31
31 Microorganismos utilizados no trabalho 31
311 Ceacutelulas hospedeiras 31
312 Trypanosoma cruzi 31
32 Manutenccedilatildeo das ceacutelulas hospedeiras 31
33 Obtenccedilatildeo dos parasitas 31
331 Epimastigotas 31
332 Curvas de crescimento de epimastigotas de T cruzi 31
333 Formas tripomastigotas 32
34 Resistecircncia a diferentes tipos de estresse 32
341 Ensaio de estresse teacutermico em formas epimastigotas 32
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico 33
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular 33
343 Ensaio de estresse oxidativo em formas epimastigotas 33
35 Tipo de morte celular em parasitas tratados com Memantina 34
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita 34
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas
epimastigotas 35
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio 35
36 Efeito sobre a metaciclogecircnese 35
37 Anaacutelise do efeito dos compostos no ciclo intracelular do T cruzi 36
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos 36
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas 36
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas
tripomastigotas 37
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os
compostos 37
38 Anaacutelises estatiacutesticas 38
4 RESULTADOS 39
41 Curvas de crescimento do T cruzi na presenccedila dos compostos 39
42 Interaccedilatildeo com condiccedilotildees de estresse 41
43 Efeito da Memantina na metaciclogecircnese 46
44 Tipo de morte celular sofrida pelo parasita tratado com Memantina 47
45 Efeito da Memantina no ciclo intracelular do T cruzi 52
5 DISCUSSAtildeO 55
6 CONCLUSOtildeES 59
REFEREcircNCIAS 61
16
1 INTRODUCcedilAtildeO
11 Aspectos gerais da doenccedila de chagas
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana foi
descoberta pelo meacutedico brasileiro Carlos Chagas (Chagas 1909) Esta doenccedila eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica Central e do Sul com cerca de 25 milhotildees de pessoas
vivendo em aacutereas de risco de infecccedilatildeo Ocorrem por ano aproximadamente 40 mil novos
casos e 12 mil mortes devido a complicaccedilotildees cardiacuteacas e digestivas (World Health
Organization 2012) Programas de controle do vetor desenvolvidos nos uacuteltimos 20 anos
como tambeacutem um maior controle nos bancos de sangue tem reduzido o nuacutemero de novos
casos da doenccedila de Chagas na Ameacuterica Latina (Rassi Marin-Neto 2010) Devido as intensas
migraccedilotildees de pessoas de paiacuteses endecircmicos o nuacutemero de casos de indiviacuteduos infectados tem
aumentado em aacutereas consideradas natildeo endecircmicas tais como Estados Unidos Canadaacute e
algumas regiotildees da Europa (Bern et al 2009 Maguire 2006 Schmunis 2007) A doenccedila eacute
uma zoonose que tem como reservatoacuterio natural do T cruzi uma ampla variedade de
mamiacuteferos do continente americano (Urbina Docampo 2003) Eacute transmitida para os seres
humanos e para mais de 150 espeacutecies de animais domeacutesticos e animais selvagens
principalmente por insetos hematoacutefagos da famiacutelia Reduviidae Apesar de existirem mais de
130 espeacutecies de triatomiacuteneos satildeo trecircs as mais importantes na transmissatildeo do T cruzi para o
homem Triatoma infestans Rhodnius prolixus e Triatoma dimidiata (Rassi Marin-Neto
2010) O parasita tambeacutem pode ser transmitido por transfusatildeo de sangue congenitamente (de
matildees infectadas para receacutem-nascidos) por transplante de oacutergatildeos ou medula oacutessea Tambeacutem
pode ocorrer por acidentes em laboratoacuterios que trabalham com o parasita vivo via esta que
epidemiologicamente eacute pouco relevante Outra forma de transmissatildeo eacute por via oral atraveacutes de
alimentos contaminados essa via vecircm sendo considerada como relevante na manutenccedilatildeo do
ciclo de transmissatildeo natural (Pereira et al 2009 Rassi Marin-Neto 2010)
A evoluccedilatildeo cliacutenica da doenccedila de Chagas pode ser dividida em duas fases aguda e
crocircnica A fase aguda normalmente eacute assintomaacutetica ou apresenta sintomatologia discreta que
na maioria dos casos passa despercebida pelo paciente Os sintomas quando aparecem o
fazem uma ou duas semanas apoacutes a exposiccedilatildeo ao triatomiacuteneo infectado ou poucos meses apoacutes
a transfusatildeo com sangue contaminado (Rassi Marin-Neto 2010) Os sintomas mais
frequentemente apresentados pelos pacientes agudos satildeo febre prolongada mal-estar
17
distensatildeo do fiacutegado baccedilo e linfonodos Pode haver tambeacutem edema subcutacircneo localizado ou
generalizado No caso particular de transmissatildeo vetorial o paciente pode apresentar um sinal
da porta de entrada do T cruzi na pele (chagoma) ou via mucosa ocular (sinal de Romantildea) As
manifestaccedilotildees da fase aguda se resolvem espontaneamente em aproximadamente 90 dos
indiviacuteduos infectados mesmo sem o tratamento com drogas tripanocidas Essa fase eacute
caracterizada pela alta parasitemia e ausecircncia de anticorpos especiacuteficos Nessa fase os
parasitas podem ser encontrados em praticamente todos os tecidos estando majoritariamente
presentes no sangue Por ser uma fase de curta duraccedilatildeo sem sintomatologia aparente ou com
sintomatologia pouco definida haacute um pequeno nuacutemero de casos detectados sendo portanto a
fase menos estudada (Rassi Marin-Neto 2010 Silber et al 2005b)
Logo apoacutes a fase aguda tem iniacutecio a fase crocircnica caracterizada pelos altos tiacutetulos de
anticorpos no sangue do paciente e pelo fato de que a parasitemia eacute inaparente O paciente
pode permanecer por vaacuterios anos ou ateacute mesmo pelo resto da vida sem apresentar sinais ou
sintomas Essa forma da fase crocircnica eacute chamada de indeterminada Aproximadamente 30-40
dos pacientes crocircnicos indeterminados desenvolvem a sintomatologia caracteriacutestica da
doenccedila evoluindo para a chamada fase crocircnica sintomaacutetica Nesta fase as trecircs principais
formas satildeo a cardiacuteaca a digestiva e a cardiodigestiva (Acquatella 2007 Marin-Neto et al
1999) Na forma cardiacuteaca o coraccedilatildeo eacute um dos primeiros oacutergatildeos afetados natildeo necessariamente
o uacutenico desenvolvendo hipertrofia e dilataccedilatildeo Na forma digestiva o trato digestivo apresenta
alteraccedilotildees anatocircmicas e fisioloacutegicas importantes sendo a principal altereccedilatildeo a dilataccedilatildeo do
intestino e do esocircfago que podem apresentar aspecto de megaviacutescera (Marin-Neto et al
1999 Rassi et al 2000) revisado por (Boscardin et al 2009 Rassi Marin-Neto 2010)
18
12 Trypanosoma cruzi
Trypanosoma cruzi eacute um parasita intracelular obrigatoacuterio classicamente localizado na
ordem kinetoplastida gecircnero Trypanosoma que causa uma infecccedilatildeo crocircnica em humanos e
em grande nuacutemero de outras espeacutecies de mamiacuteferos (Hoare 1972) Recentemente devido agrave
alta variabilidade bioloacutegica bioquiacutemica e molecular a espeacutecie Trypanosoma cruzi foi
dividida em seis linhagens subsespeciacuteficas (Discrete Typing Units ndash DTUs) numerados de I a
VI Esta classificaccedilatildeo foi realizada com base em caracteriacutesticas geneacuteticas moleculares e
imunoloacutegicas com o objetivo de unificar a nomenclatura e facilitar a comunicaccedilatildeo dentro da
comunidade cientiacutefica (Zingales et al 2009)
O T cruzi tem um ciclo de vida complexo que alterna entre o inseto vetor e o
hospedeiro mamiacutefero O parasita apresenta diferentes formas correspondentes a diferentes
estaacutegios do seu ciclo de vida Estes diferentes estaacutegios tecircm sido descritos com base em
criteacuterios morfoloacutegicos como a forma fusiforme ou arredondados a regiatildeo onde emerge o
flagelo e a posiccedilatildeo do cinetoplasto (estrutura contendo o DNA da uacutenica mitococircndria presente
no parasita) em relaccedilatildeo ao nuacutecleo No hospedeiro mamiacutefero foram descritas as formas
amastigota epimastigota intracelular e tripomastigota sanguiacutecola A forma amastigota eacute um
estaacutegio replicativo e infectivo Satildeo ceacutelulas arredondadas medindo de 2-6 microm de diacircmetro
encontradas no citoplasma das ceacutelulas infectadas Os epimastigotas intracelulares
(epimastigote-like) satildeo formas replicativas intermediaacuterias entre amastigotas e tripomastigotas
tambeacutem encontrados dentro das ceacutelulas medindo aproximadamente 54 microm Essas formas satildeo
menores que as formas epimastigotas encontradas no inseto vetor poreacutem compartilha com as
mesmas padrotildees morfomeacutetricos e diversas caracteriacutesticas bioquiacutemicas e bioloacutegicas (Almeida-
De-Faria et al 1999) O tripomastigota sanguiacutecola eacute um estaacutegio flagelado infectivo natildeo
replicativo encontrado principalmente na corrente sanguiacutenea do hospedeiro mamiacutefero Mede
aproximadamente 18 microm de comprimento incluindo um extremo livre do flagelo de
aproximadamente 6 microm e 2-3 microm de largura
No inseto vetor encontram-se predominantemente as formas epimastigota e
tripomastigota metaciacuteclico Epimastigota eacute uma forma flagelada replicativa e natildeo infectiva
Mede aproximadamente 20-40 microm de comprimento e 2-5 microm de largura essas formas estatildeo
presentes tipicamente na porccedilatildeo meacutedia do intestino do hospedeiro invertebrado
Tripomastigota metaciacuteclico eacute uma forma infectiva natildeo replicativa encontrada na porccedilatildeo final
do intestino do inseto vetor (Brener 1971) revisado por (Silber et al 2005a Tyler Engman
2001)
19
O ciclo de vida do T cruzi inicia-se quando os parasitas presentes na corrente
sanguiacutenea do hospedeiro vertebrado infectado passam ao inseto vetor durante o repasto
sanguiacuteneo No intestino do inseto estas formas se diferenciam em formas epimastigotas (De
Souza et al 2002) que apoacutes multiplicaccedilatildeo por fissatildeo binaacuteria colonizam a porccedilatildeo terminal do
intestino e se diferenciam novamente em tripomastigotas metaciacuteclicos Quando o inseto
infectado pica o hospedeiro vertebrado e inicia o repasto sanguiacuteneo concomitantemente
elimina as formas tripomastigotas metaciacuteclicos junto com as fezes e urina Uma vez em
contato com a pele ou mucosas do hospedeiro mamiacutefero os tripomastigotas satildeo internalizados
atraveacutes de pequenas lesotildees na pele causadas pelo proacuteprio hospedeiro (Alves Colli 2008) Os
tripomastigotas invadem as ceacutelulas do hospedeiro mamiacutefero e se diferenciam em formas
replicativas denominadas amastigotas estabelecendo assim a infecccedilatildeo Apoacutes
aproximadamente 96 horas os amastigotas comeccedilam a se diferenciar em tripomastigotas
passando pela forma intermediaacuteria epimastigota intracelular Estas formas tripomastigotas satildeo
liberadas na corrente sanguiacutenea podendo invadir novas ceacutelulas ou serem ingeridas pelo inseto
vetor em um novo repasto sanguiacuteneo fechando assim o seu ciclo de vida O esquema do ciclo
de vida do Trypanosoma cruzi estaacute representado na Figura 1
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi
20
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos
T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono As formas epimastigotas preferencialmente catabolizam glicose e apoacutes a exaustatildeo
do carboidrato ocorre uma mudanccedila para o metabolismo de aminoaacutecidos A prolina e a
glutamina satildeo os principais aminoaacutecidos metabolizados pelas formas epimastigotas visto que
satildeo abudantes na hemolinfa e nos fluiacutedos dos tecidos do inseto vetor Esse metabolismo
ocorre com produccedilatildeo de NH4+
(produto da deaminaccedilatildeo dos aminoaacutecidos) que eacute excretada para
o meio extracelular (Bringaud et al 2006 Cazzulo 1994) As formas tripomastigotas
utilizam principalmente a glicose que eacute abundante nos fluiacutedos de seus hospedeiros
vertebrados Jaacute as formas amastigotas encontradas no citoplasma das ceacutelulas do hospedeiro
mamiacutefero ambiente pobre em glicose livre usam os aminoaacutecidos como principal fonte de
carbono e energia apresentando maior quantidade de prolina intracelular do que as formas
tripomastigotas (Silber et al 2009)
Entre os aminoaacutecidos metabolizados pelas formas epimastigotas estatildeo prolina glicina
asparagina glutamina aspartato glutamato leucina e isoleucina (Mancilla et al 1967 Odaly
et al 1983 Silber et al 2005a Sylvester Krassner 1976 Zeledon 1960) Os aminoaacutecidos
estatildeo envolvidos tambeacutem em outros processos importantes do ciclo de vida do T cruzi O
aminoaacutecido L-prolina eacute essencial na diferenciaccedilatildeo das formas epimastigota intracelular para
tripomastigotas aleacutem de estarem envolvidos nos processos de osmorregulaccedilatildeo crescimento e
resistecircncia do parasita ao estresse oxidativo e nutricional (Homsy et al 1989 Magdaleno et
al 2009 Magdaleno et al 2011 Pereira et al 2002 Pereira et al 2003 Tonelli et al
2004) Glutamato prolina e aspartato satildeo importantes na metaciclogecircnese ou seja na
diferenciaccedilatildeo de formas epimastigotas para tripomastigotas metaciacuteclicos (Contreras et al
1985) Prolina glicina alanina e glutamato satildeo descritos como importantes nos processos de
osmorregulaccedilatildeo e controle do volume celular (Rohloff et al 2003 Rohloff et al 2004a)
Os aminoaacutecidos para que sejam metabolizados ou cumprirem outras funccedilotildees devem
ser obtidos a partir da degradaccedilatildeo de proteiacutenas sintetizados ou incorporados pelo parasita
atraveacutes de transportadores localizados na membrana (Silber et al 2005a) O primeiro
transportador de aminoaacutecidos em T cruzi foi descrito bioquimicamente em 1970 o
transportador de lisina (Hampton 1970) Outros aminoaacutecidos tambeacutem jaacute tiveram a sua
cineacutetica e seu mecanismo de transporte descrito para T cruzi A incorporaccedilatildeo de arginina
pelo parasita apresenta pelo menos dois sistemas de transporte sendo um de alta afinidade e
outro de baixa afinidade Esse transporte eacute regulado ao longo do ciclo de vida do parasita
21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

LISTA DE ILUSTRACcedilOtildeES
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi 19
Figura 2 - Estruturas quiacutemicas do Benzonidazol e Nifurtimox 24
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica 28
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica 29
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse 40
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os
tratamentos com MK-801 ou Memantina 41
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina 42
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico ou oxidativo 44
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na
presenccedila do Memantina 45
Figura 10 - Efeito da Memantina na metaciclogecircnese 46
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo
de morte celular 48
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2 49
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
50
Figura 14 - Formas epimastigotas de T cruzi 51
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi 53
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade
de formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo
celular 54
LISTA DE ABREVIATURAS
ANOVA Anaacutelise de variacircncia
ATP Adenosina trifosfato
CHO-K1 Chinese Hamster Ovary (linhagem celular proveniente de ovaacuterio de
hamester chinecircs)
Ca2+
Caacutelcio
CO2 Dioacutexido de carbono
DNA Aacutecido desoxirribonucleacuteico
DO Densidade oacutetica
DTUs Discrete Typing Units
EGTA Aacutecido etilenoglicol-bis-aminoetiletertetraceacutetico
EROs Espeacutecies reativas de oxigecircnio
GABA Aminoaacutecido γ-aminobutiacuterico
GABA T Aminoaacutecido γ-aminobutiacuterico transaminase
GAPDH Gliceraldeiacutedo -3-fosfato desidrogenase
H2O2 Peroacutexido de hidrogecircnio
IC50 Metade da concentraccedilatildeo maacutexima inibitoacuteria
KCl Cloreto de potaacutessio
LIT Liver Infusion-Tryptose (Meio de cultura)
MCP Morte celular programada
mg Miligramas
mGluR Receptor metabotroacutepico de glutamato
MTT (3-(45-dimetiltiazol-2-il)-25 difenil bromo tetrazolio)
Na2HPO4 Fosfato de soacutedio monobaacutesico anidro
NaCl Cloreto de soacutedio
NaHCO3 Bicarbonato de soacutedio
NCCD Nomenclature Comittee on Cell Death (Comite de Nomeclatura de Morte
Celular)
NH4+
Ion amocircnio
NMDA N-metil-D-aspartato
OH Radical hidroxila
PBS Soluccedilatildeo salina tamponada com fosfato
pH Potencial hidrogeniocircnico
RPMI Roswell Park Memorial Institute
SDS Soacutedio dodecil sulfat
SFB Soro fetal bovino
U Unidades
μg Micrograma
μl Microlitro
μm Microcircmetro
SUMAacuteRIO
1 INTRODUCcedilAtildeO 16
11 Aspectos gerais da doenccedila de chagas 16
12 Trypanosoma cruzi 18
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos 20
14 Morte celular em Trypanosoma cruzi 21
15 Terapia na doenccedila de Chagas 23
16 Perspectivas de novos faacutermacos 25
2 OBJETIVOS 30
21 Objetivo geral 30
22 Objetivos especiacuteficos 30
3 MATERIAIS E MEacuteTODOS 31
31 Microorganismos utilizados no trabalho 31
311 Ceacutelulas hospedeiras 31
312 Trypanosoma cruzi 31
32 Manutenccedilatildeo das ceacutelulas hospedeiras 31
33 Obtenccedilatildeo dos parasitas 31
331 Epimastigotas 31
332 Curvas de crescimento de epimastigotas de T cruzi 31
333 Formas tripomastigotas 32
34 Resistecircncia a diferentes tipos de estresse 32
341 Ensaio de estresse teacutermico em formas epimastigotas 32
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico 33
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular 33
343 Ensaio de estresse oxidativo em formas epimastigotas 33
35 Tipo de morte celular em parasitas tratados com Memantina 34
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita 34
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas
epimastigotas 35
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio 35
36 Efeito sobre a metaciclogecircnese 35
37 Anaacutelise do efeito dos compostos no ciclo intracelular do T cruzi 36
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos 36
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas 36
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas
tripomastigotas 37
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os
compostos 37
38 Anaacutelises estatiacutesticas 38
4 RESULTADOS 39
41 Curvas de crescimento do T cruzi na presenccedila dos compostos 39
42 Interaccedilatildeo com condiccedilotildees de estresse 41
43 Efeito da Memantina na metaciclogecircnese 46
44 Tipo de morte celular sofrida pelo parasita tratado com Memantina 47
45 Efeito da Memantina no ciclo intracelular do T cruzi 52
5 DISCUSSAtildeO 55
6 CONCLUSOtildeES 59
REFEREcircNCIAS 61
16
1 INTRODUCcedilAtildeO
11 Aspectos gerais da doenccedila de chagas
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana foi
descoberta pelo meacutedico brasileiro Carlos Chagas (Chagas 1909) Esta doenccedila eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica Central e do Sul com cerca de 25 milhotildees de pessoas
vivendo em aacutereas de risco de infecccedilatildeo Ocorrem por ano aproximadamente 40 mil novos
casos e 12 mil mortes devido a complicaccedilotildees cardiacuteacas e digestivas (World Health
Organization 2012) Programas de controle do vetor desenvolvidos nos uacuteltimos 20 anos
como tambeacutem um maior controle nos bancos de sangue tem reduzido o nuacutemero de novos
casos da doenccedila de Chagas na Ameacuterica Latina (Rassi Marin-Neto 2010) Devido as intensas
migraccedilotildees de pessoas de paiacuteses endecircmicos o nuacutemero de casos de indiviacuteduos infectados tem
aumentado em aacutereas consideradas natildeo endecircmicas tais como Estados Unidos Canadaacute e
algumas regiotildees da Europa (Bern et al 2009 Maguire 2006 Schmunis 2007) A doenccedila eacute
uma zoonose que tem como reservatoacuterio natural do T cruzi uma ampla variedade de
mamiacuteferos do continente americano (Urbina Docampo 2003) Eacute transmitida para os seres
humanos e para mais de 150 espeacutecies de animais domeacutesticos e animais selvagens
principalmente por insetos hematoacutefagos da famiacutelia Reduviidae Apesar de existirem mais de
130 espeacutecies de triatomiacuteneos satildeo trecircs as mais importantes na transmissatildeo do T cruzi para o
homem Triatoma infestans Rhodnius prolixus e Triatoma dimidiata (Rassi Marin-Neto
2010) O parasita tambeacutem pode ser transmitido por transfusatildeo de sangue congenitamente (de
matildees infectadas para receacutem-nascidos) por transplante de oacutergatildeos ou medula oacutessea Tambeacutem
pode ocorrer por acidentes em laboratoacuterios que trabalham com o parasita vivo via esta que
epidemiologicamente eacute pouco relevante Outra forma de transmissatildeo eacute por via oral atraveacutes de
alimentos contaminados essa via vecircm sendo considerada como relevante na manutenccedilatildeo do
ciclo de transmissatildeo natural (Pereira et al 2009 Rassi Marin-Neto 2010)
A evoluccedilatildeo cliacutenica da doenccedila de Chagas pode ser dividida em duas fases aguda e
crocircnica A fase aguda normalmente eacute assintomaacutetica ou apresenta sintomatologia discreta que
na maioria dos casos passa despercebida pelo paciente Os sintomas quando aparecem o
fazem uma ou duas semanas apoacutes a exposiccedilatildeo ao triatomiacuteneo infectado ou poucos meses apoacutes
a transfusatildeo com sangue contaminado (Rassi Marin-Neto 2010) Os sintomas mais
frequentemente apresentados pelos pacientes agudos satildeo febre prolongada mal-estar
17
distensatildeo do fiacutegado baccedilo e linfonodos Pode haver tambeacutem edema subcutacircneo localizado ou
generalizado No caso particular de transmissatildeo vetorial o paciente pode apresentar um sinal
da porta de entrada do T cruzi na pele (chagoma) ou via mucosa ocular (sinal de Romantildea) As
manifestaccedilotildees da fase aguda se resolvem espontaneamente em aproximadamente 90 dos
indiviacuteduos infectados mesmo sem o tratamento com drogas tripanocidas Essa fase eacute
caracterizada pela alta parasitemia e ausecircncia de anticorpos especiacuteficos Nessa fase os
parasitas podem ser encontrados em praticamente todos os tecidos estando majoritariamente
presentes no sangue Por ser uma fase de curta duraccedilatildeo sem sintomatologia aparente ou com
sintomatologia pouco definida haacute um pequeno nuacutemero de casos detectados sendo portanto a
fase menos estudada (Rassi Marin-Neto 2010 Silber et al 2005b)
Logo apoacutes a fase aguda tem iniacutecio a fase crocircnica caracterizada pelos altos tiacutetulos de
anticorpos no sangue do paciente e pelo fato de que a parasitemia eacute inaparente O paciente
pode permanecer por vaacuterios anos ou ateacute mesmo pelo resto da vida sem apresentar sinais ou
sintomas Essa forma da fase crocircnica eacute chamada de indeterminada Aproximadamente 30-40
dos pacientes crocircnicos indeterminados desenvolvem a sintomatologia caracteriacutestica da
doenccedila evoluindo para a chamada fase crocircnica sintomaacutetica Nesta fase as trecircs principais
formas satildeo a cardiacuteaca a digestiva e a cardiodigestiva (Acquatella 2007 Marin-Neto et al
1999) Na forma cardiacuteaca o coraccedilatildeo eacute um dos primeiros oacutergatildeos afetados natildeo necessariamente
o uacutenico desenvolvendo hipertrofia e dilataccedilatildeo Na forma digestiva o trato digestivo apresenta
alteraccedilotildees anatocircmicas e fisioloacutegicas importantes sendo a principal altereccedilatildeo a dilataccedilatildeo do
intestino e do esocircfago que podem apresentar aspecto de megaviacutescera (Marin-Neto et al
1999 Rassi et al 2000) revisado por (Boscardin et al 2009 Rassi Marin-Neto 2010)
18
12 Trypanosoma cruzi
Trypanosoma cruzi eacute um parasita intracelular obrigatoacuterio classicamente localizado na
ordem kinetoplastida gecircnero Trypanosoma que causa uma infecccedilatildeo crocircnica em humanos e
em grande nuacutemero de outras espeacutecies de mamiacuteferos (Hoare 1972) Recentemente devido agrave
alta variabilidade bioloacutegica bioquiacutemica e molecular a espeacutecie Trypanosoma cruzi foi
dividida em seis linhagens subsespeciacuteficas (Discrete Typing Units ndash DTUs) numerados de I a
VI Esta classificaccedilatildeo foi realizada com base em caracteriacutesticas geneacuteticas moleculares e
imunoloacutegicas com o objetivo de unificar a nomenclatura e facilitar a comunicaccedilatildeo dentro da
comunidade cientiacutefica (Zingales et al 2009)
O T cruzi tem um ciclo de vida complexo que alterna entre o inseto vetor e o
hospedeiro mamiacutefero O parasita apresenta diferentes formas correspondentes a diferentes
estaacutegios do seu ciclo de vida Estes diferentes estaacutegios tecircm sido descritos com base em
criteacuterios morfoloacutegicos como a forma fusiforme ou arredondados a regiatildeo onde emerge o
flagelo e a posiccedilatildeo do cinetoplasto (estrutura contendo o DNA da uacutenica mitococircndria presente
no parasita) em relaccedilatildeo ao nuacutecleo No hospedeiro mamiacutefero foram descritas as formas
amastigota epimastigota intracelular e tripomastigota sanguiacutecola A forma amastigota eacute um
estaacutegio replicativo e infectivo Satildeo ceacutelulas arredondadas medindo de 2-6 microm de diacircmetro
encontradas no citoplasma das ceacutelulas infectadas Os epimastigotas intracelulares
(epimastigote-like) satildeo formas replicativas intermediaacuterias entre amastigotas e tripomastigotas
tambeacutem encontrados dentro das ceacutelulas medindo aproximadamente 54 microm Essas formas satildeo
menores que as formas epimastigotas encontradas no inseto vetor poreacutem compartilha com as
mesmas padrotildees morfomeacutetricos e diversas caracteriacutesticas bioquiacutemicas e bioloacutegicas (Almeida-
De-Faria et al 1999) O tripomastigota sanguiacutecola eacute um estaacutegio flagelado infectivo natildeo
replicativo encontrado principalmente na corrente sanguiacutenea do hospedeiro mamiacutefero Mede
aproximadamente 18 microm de comprimento incluindo um extremo livre do flagelo de
aproximadamente 6 microm e 2-3 microm de largura
No inseto vetor encontram-se predominantemente as formas epimastigota e
tripomastigota metaciacuteclico Epimastigota eacute uma forma flagelada replicativa e natildeo infectiva
Mede aproximadamente 20-40 microm de comprimento e 2-5 microm de largura essas formas estatildeo
presentes tipicamente na porccedilatildeo meacutedia do intestino do hospedeiro invertebrado
Tripomastigota metaciacuteclico eacute uma forma infectiva natildeo replicativa encontrada na porccedilatildeo final
do intestino do inseto vetor (Brener 1971) revisado por (Silber et al 2005a Tyler Engman
2001)
19
O ciclo de vida do T cruzi inicia-se quando os parasitas presentes na corrente
sanguiacutenea do hospedeiro vertebrado infectado passam ao inseto vetor durante o repasto
sanguiacuteneo No intestino do inseto estas formas se diferenciam em formas epimastigotas (De
Souza et al 2002) que apoacutes multiplicaccedilatildeo por fissatildeo binaacuteria colonizam a porccedilatildeo terminal do
intestino e se diferenciam novamente em tripomastigotas metaciacuteclicos Quando o inseto
infectado pica o hospedeiro vertebrado e inicia o repasto sanguiacuteneo concomitantemente
elimina as formas tripomastigotas metaciacuteclicos junto com as fezes e urina Uma vez em
contato com a pele ou mucosas do hospedeiro mamiacutefero os tripomastigotas satildeo internalizados
atraveacutes de pequenas lesotildees na pele causadas pelo proacuteprio hospedeiro (Alves Colli 2008) Os
tripomastigotas invadem as ceacutelulas do hospedeiro mamiacutefero e se diferenciam em formas
replicativas denominadas amastigotas estabelecendo assim a infecccedilatildeo Apoacutes
aproximadamente 96 horas os amastigotas comeccedilam a se diferenciar em tripomastigotas
passando pela forma intermediaacuteria epimastigota intracelular Estas formas tripomastigotas satildeo
liberadas na corrente sanguiacutenea podendo invadir novas ceacutelulas ou serem ingeridas pelo inseto
vetor em um novo repasto sanguiacuteneo fechando assim o seu ciclo de vida O esquema do ciclo
de vida do Trypanosoma cruzi estaacute representado na Figura 1
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi
20
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos
T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono As formas epimastigotas preferencialmente catabolizam glicose e apoacutes a exaustatildeo
do carboidrato ocorre uma mudanccedila para o metabolismo de aminoaacutecidos A prolina e a
glutamina satildeo os principais aminoaacutecidos metabolizados pelas formas epimastigotas visto que
satildeo abudantes na hemolinfa e nos fluiacutedos dos tecidos do inseto vetor Esse metabolismo
ocorre com produccedilatildeo de NH4+
(produto da deaminaccedilatildeo dos aminoaacutecidos) que eacute excretada para
o meio extracelular (Bringaud et al 2006 Cazzulo 1994) As formas tripomastigotas
utilizam principalmente a glicose que eacute abundante nos fluiacutedos de seus hospedeiros
vertebrados Jaacute as formas amastigotas encontradas no citoplasma das ceacutelulas do hospedeiro
mamiacutefero ambiente pobre em glicose livre usam os aminoaacutecidos como principal fonte de
carbono e energia apresentando maior quantidade de prolina intracelular do que as formas
tripomastigotas (Silber et al 2009)
Entre os aminoaacutecidos metabolizados pelas formas epimastigotas estatildeo prolina glicina
asparagina glutamina aspartato glutamato leucina e isoleucina (Mancilla et al 1967 Odaly
et al 1983 Silber et al 2005a Sylvester Krassner 1976 Zeledon 1960) Os aminoaacutecidos
estatildeo envolvidos tambeacutem em outros processos importantes do ciclo de vida do T cruzi O
aminoaacutecido L-prolina eacute essencial na diferenciaccedilatildeo das formas epimastigota intracelular para
tripomastigotas aleacutem de estarem envolvidos nos processos de osmorregulaccedilatildeo crescimento e
resistecircncia do parasita ao estresse oxidativo e nutricional (Homsy et al 1989 Magdaleno et
al 2009 Magdaleno et al 2011 Pereira et al 2002 Pereira et al 2003 Tonelli et al
2004) Glutamato prolina e aspartato satildeo importantes na metaciclogecircnese ou seja na
diferenciaccedilatildeo de formas epimastigotas para tripomastigotas metaciacuteclicos (Contreras et al
1985) Prolina glicina alanina e glutamato satildeo descritos como importantes nos processos de
osmorregulaccedilatildeo e controle do volume celular (Rohloff et al 2003 Rohloff et al 2004a)
Os aminoaacutecidos para que sejam metabolizados ou cumprirem outras funccedilotildees devem
ser obtidos a partir da degradaccedilatildeo de proteiacutenas sintetizados ou incorporados pelo parasita
atraveacutes de transportadores localizados na membrana (Silber et al 2005a) O primeiro
transportador de aminoaacutecidos em T cruzi foi descrito bioquimicamente em 1970 o
transportador de lisina (Hampton 1970) Outros aminoaacutecidos tambeacutem jaacute tiveram a sua
cineacutetica e seu mecanismo de transporte descrito para T cruzi A incorporaccedilatildeo de arginina
pelo parasita apresenta pelo menos dois sistemas de transporte sendo um de alta afinidade e
outro de baixa afinidade Esse transporte eacute regulado ao longo do ciclo de vida do parasita
21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

LISTA DE ABREVIATURAS
ANOVA Anaacutelise de variacircncia
ATP Adenosina trifosfato
CHO-K1 Chinese Hamster Ovary (linhagem celular proveniente de ovaacuterio de
hamester chinecircs)
Ca2+
Caacutelcio
CO2 Dioacutexido de carbono
DNA Aacutecido desoxirribonucleacuteico
DO Densidade oacutetica
DTUs Discrete Typing Units
EGTA Aacutecido etilenoglicol-bis-aminoetiletertetraceacutetico
EROs Espeacutecies reativas de oxigecircnio
GABA Aminoaacutecido γ-aminobutiacuterico
GABA T Aminoaacutecido γ-aminobutiacuterico transaminase
GAPDH Gliceraldeiacutedo -3-fosfato desidrogenase
H2O2 Peroacutexido de hidrogecircnio
IC50 Metade da concentraccedilatildeo maacutexima inibitoacuteria
KCl Cloreto de potaacutessio
LIT Liver Infusion-Tryptose (Meio de cultura)
MCP Morte celular programada
mg Miligramas
mGluR Receptor metabotroacutepico de glutamato
MTT (3-(45-dimetiltiazol-2-il)-25 difenil bromo tetrazolio)
Na2HPO4 Fosfato de soacutedio monobaacutesico anidro
NaCl Cloreto de soacutedio
NaHCO3 Bicarbonato de soacutedio
NCCD Nomenclature Comittee on Cell Death (Comite de Nomeclatura de Morte
Celular)
NH4+
Ion amocircnio
NMDA N-metil-D-aspartato
OH Radical hidroxila
PBS Soluccedilatildeo salina tamponada com fosfato
pH Potencial hidrogeniocircnico
RPMI Roswell Park Memorial Institute
SDS Soacutedio dodecil sulfat
SFB Soro fetal bovino
U Unidades
μg Micrograma
μl Microlitro
μm Microcircmetro
SUMAacuteRIO
1 INTRODUCcedilAtildeO 16
11 Aspectos gerais da doenccedila de chagas 16
12 Trypanosoma cruzi 18
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos 20
14 Morte celular em Trypanosoma cruzi 21
15 Terapia na doenccedila de Chagas 23
16 Perspectivas de novos faacutermacos 25
2 OBJETIVOS 30
21 Objetivo geral 30
22 Objetivos especiacuteficos 30
3 MATERIAIS E MEacuteTODOS 31
31 Microorganismos utilizados no trabalho 31
311 Ceacutelulas hospedeiras 31
312 Trypanosoma cruzi 31
32 Manutenccedilatildeo das ceacutelulas hospedeiras 31
33 Obtenccedilatildeo dos parasitas 31
331 Epimastigotas 31
332 Curvas de crescimento de epimastigotas de T cruzi 31
333 Formas tripomastigotas 32
34 Resistecircncia a diferentes tipos de estresse 32
341 Ensaio de estresse teacutermico em formas epimastigotas 32
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico 33
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular 33
343 Ensaio de estresse oxidativo em formas epimastigotas 33
35 Tipo de morte celular em parasitas tratados com Memantina 34
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita 34
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas
epimastigotas 35
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio 35
36 Efeito sobre a metaciclogecircnese 35
37 Anaacutelise do efeito dos compostos no ciclo intracelular do T cruzi 36
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos 36
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas 36
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas
tripomastigotas 37
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os
compostos 37
38 Anaacutelises estatiacutesticas 38
4 RESULTADOS 39
41 Curvas de crescimento do T cruzi na presenccedila dos compostos 39
42 Interaccedilatildeo com condiccedilotildees de estresse 41
43 Efeito da Memantina na metaciclogecircnese 46
44 Tipo de morte celular sofrida pelo parasita tratado com Memantina 47
45 Efeito da Memantina no ciclo intracelular do T cruzi 52
5 DISCUSSAtildeO 55
6 CONCLUSOtildeES 59
REFEREcircNCIAS 61
16
1 INTRODUCcedilAtildeO
11 Aspectos gerais da doenccedila de chagas
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana foi
descoberta pelo meacutedico brasileiro Carlos Chagas (Chagas 1909) Esta doenccedila eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica Central e do Sul com cerca de 25 milhotildees de pessoas
vivendo em aacutereas de risco de infecccedilatildeo Ocorrem por ano aproximadamente 40 mil novos
casos e 12 mil mortes devido a complicaccedilotildees cardiacuteacas e digestivas (World Health
Organization 2012) Programas de controle do vetor desenvolvidos nos uacuteltimos 20 anos
como tambeacutem um maior controle nos bancos de sangue tem reduzido o nuacutemero de novos
casos da doenccedila de Chagas na Ameacuterica Latina (Rassi Marin-Neto 2010) Devido as intensas
migraccedilotildees de pessoas de paiacuteses endecircmicos o nuacutemero de casos de indiviacuteduos infectados tem
aumentado em aacutereas consideradas natildeo endecircmicas tais como Estados Unidos Canadaacute e
algumas regiotildees da Europa (Bern et al 2009 Maguire 2006 Schmunis 2007) A doenccedila eacute
uma zoonose que tem como reservatoacuterio natural do T cruzi uma ampla variedade de
mamiacuteferos do continente americano (Urbina Docampo 2003) Eacute transmitida para os seres
humanos e para mais de 150 espeacutecies de animais domeacutesticos e animais selvagens
principalmente por insetos hematoacutefagos da famiacutelia Reduviidae Apesar de existirem mais de
130 espeacutecies de triatomiacuteneos satildeo trecircs as mais importantes na transmissatildeo do T cruzi para o
homem Triatoma infestans Rhodnius prolixus e Triatoma dimidiata (Rassi Marin-Neto
2010) O parasita tambeacutem pode ser transmitido por transfusatildeo de sangue congenitamente (de
matildees infectadas para receacutem-nascidos) por transplante de oacutergatildeos ou medula oacutessea Tambeacutem
pode ocorrer por acidentes em laboratoacuterios que trabalham com o parasita vivo via esta que
epidemiologicamente eacute pouco relevante Outra forma de transmissatildeo eacute por via oral atraveacutes de
alimentos contaminados essa via vecircm sendo considerada como relevante na manutenccedilatildeo do
ciclo de transmissatildeo natural (Pereira et al 2009 Rassi Marin-Neto 2010)
A evoluccedilatildeo cliacutenica da doenccedila de Chagas pode ser dividida em duas fases aguda e
crocircnica A fase aguda normalmente eacute assintomaacutetica ou apresenta sintomatologia discreta que
na maioria dos casos passa despercebida pelo paciente Os sintomas quando aparecem o
fazem uma ou duas semanas apoacutes a exposiccedilatildeo ao triatomiacuteneo infectado ou poucos meses apoacutes
a transfusatildeo com sangue contaminado (Rassi Marin-Neto 2010) Os sintomas mais
frequentemente apresentados pelos pacientes agudos satildeo febre prolongada mal-estar
17
distensatildeo do fiacutegado baccedilo e linfonodos Pode haver tambeacutem edema subcutacircneo localizado ou
generalizado No caso particular de transmissatildeo vetorial o paciente pode apresentar um sinal
da porta de entrada do T cruzi na pele (chagoma) ou via mucosa ocular (sinal de Romantildea) As
manifestaccedilotildees da fase aguda se resolvem espontaneamente em aproximadamente 90 dos
indiviacuteduos infectados mesmo sem o tratamento com drogas tripanocidas Essa fase eacute
caracterizada pela alta parasitemia e ausecircncia de anticorpos especiacuteficos Nessa fase os
parasitas podem ser encontrados em praticamente todos os tecidos estando majoritariamente
presentes no sangue Por ser uma fase de curta duraccedilatildeo sem sintomatologia aparente ou com
sintomatologia pouco definida haacute um pequeno nuacutemero de casos detectados sendo portanto a
fase menos estudada (Rassi Marin-Neto 2010 Silber et al 2005b)
Logo apoacutes a fase aguda tem iniacutecio a fase crocircnica caracterizada pelos altos tiacutetulos de
anticorpos no sangue do paciente e pelo fato de que a parasitemia eacute inaparente O paciente
pode permanecer por vaacuterios anos ou ateacute mesmo pelo resto da vida sem apresentar sinais ou
sintomas Essa forma da fase crocircnica eacute chamada de indeterminada Aproximadamente 30-40
dos pacientes crocircnicos indeterminados desenvolvem a sintomatologia caracteriacutestica da
doenccedila evoluindo para a chamada fase crocircnica sintomaacutetica Nesta fase as trecircs principais
formas satildeo a cardiacuteaca a digestiva e a cardiodigestiva (Acquatella 2007 Marin-Neto et al
1999) Na forma cardiacuteaca o coraccedilatildeo eacute um dos primeiros oacutergatildeos afetados natildeo necessariamente
o uacutenico desenvolvendo hipertrofia e dilataccedilatildeo Na forma digestiva o trato digestivo apresenta
alteraccedilotildees anatocircmicas e fisioloacutegicas importantes sendo a principal altereccedilatildeo a dilataccedilatildeo do
intestino e do esocircfago que podem apresentar aspecto de megaviacutescera (Marin-Neto et al
1999 Rassi et al 2000) revisado por (Boscardin et al 2009 Rassi Marin-Neto 2010)
18
12 Trypanosoma cruzi
Trypanosoma cruzi eacute um parasita intracelular obrigatoacuterio classicamente localizado na
ordem kinetoplastida gecircnero Trypanosoma que causa uma infecccedilatildeo crocircnica em humanos e
em grande nuacutemero de outras espeacutecies de mamiacuteferos (Hoare 1972) Recentemente devido agrave
alta variabilidade bioloacutegica bioquiacutemica e molecular a espeacutecie Trypanosoma cruzi foi
dividida em seis linhagens subsespeciacuteficas (Discrete Typing Units ndash DTUs) numerados de I a
VI Esta classificaccedilatildeo foi realizada com base em caracteriacutesticas geneacuteticas moleculares e
imunoloacutegicas com o objetivo de unificar a nomenclatura e facilitar a comunicaccedilatildeo dentro da
comunidade cientiacutefica (Zingales et al 2009)
O T cruzi tem um ciclo de vida complexo que alterna entre o inseto vetor e o
hospedeiro mamiacutefero O parasita apresenta diferentes formas correspondentes a diferentes
estaacutegios do seu ciclo de vida Estes diferentes estaacutegios tecircm sido descritos com base em
criteacuterios morfoloacutegicos como a forma fusiforme ou arredondados a regiatildeo onde emerge o
flagelo e a posiccedilatildeo do cinetoplasto (estrutura contendo o DNA da uacutenica mitococircndria presente
no parasita) em relaccedilatildeo ao nuacutecleo No hospedeiro mamiacutefero foram descritas as formas
amastigota epimastigota intracelular e tripomastigota sanguiacutecola A forma amastigota eacute um
estaacutegio replicativo e infectivo Satildeo ceacutelulas arredondadas medindo de 2-6 microm de diacircmetro
encontradas no citoplasma das ceacutelulas infectadas Os epimastigotas intracelulares
(epimastigote-like) satildeo formas replicativas intermediaacuterias entre amastigotas e tripomastigotas
tambeacutem encontrados dentro das ceacutelulas medindo aproximadamente 54 microm Essas formas satildeo
menores que as formas epimastigotas encontradas no inseto vetor poreacutem compartilha com as
mesmas padrotildees morfomeacutetricos e diversas caracteriacutesticas bioquiacutemicas e bioloacutegicas (Almeida-
De-Faria et al 1999) O tripomastigota sanguiacutecola eacute um estaacutegio flagelado infectivo natildeo
replicativo encontrado principalmente na corrente sanguiacutenea do hospedeiro mamiacutefero Mede
aproximadamente 18 microm de comprimento incluindo um extremo livre do flagelo de
aproximadamente 6 microm e 2-3 microm de largura
No inseto vetor encontram-se predominantemente as formas epimastigota e
tripomastigota metaciacuteclico Epimastigota eacute uma forma flagelada replicativa e natildeo infectiva
Mede aproximadamente 20-40 microm de comprimento e 2-5 microm de largura essas formas estatildeo
presentes tipicamente na porccedilatildeo meacutedia do intestino do hospedeiro invertebrado
Tripomastigota metaciacuteclico eacute uma forma infectiva natildeo replicativa encontrada na porccedilatildeo final
do intestino do inseto vetor (Brener 1971) revisado por (Silber et al 2005a Tyler Engman
2001)
19
O ciclo de vida do T cruzi inicia-se quando os parasitas presentes na corrente
sanguiacutenea do hospedeiro vertebrado infectado passam ao inseto vetor durante o repasto
sanguiacuteneo No intestino do inseto estas formas se diferenciam em formas epimastigotas (De
Souza et al 2002) que apoacutes multiplicaccedilatildeo por fissatildeo binaacuteria colonizam a porccedilatildeo terminal do
intestino e se diferenciam novamente em tripomastigotas metaciacuteclicos Quando o inseto
infectado pica o hospedeiro vertebrado e inicia o repasto sanguiacuteneo concomitantemente
elimina as formas tripomastigotas metaciacuteclicos junto com as fezes e urina Uma vez em
contato com a pele ou mucosas do hospedeiro mamiacutefero os tripomastigotas satildeo internalizados
atraveacutes de pequenas lesotildees na pele causadas pelo proacuteprio hospedeiro (Alves Colli 2008) Os
tripomastigotas invadem as ceacutelulas do hospedeiro mamiacutefero e se diferenciam em formas
replicativas denominadas amastigotas estabelecendo assim a infecccedilatildeo Apoacutes
aproximadamente 96 horas os amastigotas comeccedilam a se diferenciar em tripomastigotas
passando pela forma intermediaacuteria epimastigota intracelular Estas formas tripomastigotas satildeo
liberadas na corrente sanguiacutenea podendo invadir novas ceacutelulas ou serem ingeridas pelo inseto
vetor em um novo repasto sanguiacuteneo fechando assim o seu ciclo de vida O esquema do ciclo
de vida do Trypanosoma cruzi estaacute representado na Figura 1
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi
20
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos
T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono As formas epimastigotas preferencialmente catabolizam glicose e apoacutes a exaustatildeo
do carboidrato ocorre uma mudanccedila para o metabolismo de aminoaacutecidos A prolina e a
glutamina satildeo os principais aminoaacutecidos metabolizados pelas formas epimastigotas visto que
satildeo abudantes na hemolinfa e nos fluiacutedos dos tecidos do inseto vetor Esse metabolismo
ocorre com produccedilatildeo de NH4+
(produto da deaminaccedilatildeo dos aminoaacutecidos) que eacute excretada para
o meio extracelular (Bringaud et al 2006 Cazzulo 1994) As formas tripomastigotas
utilizam principalmente a glicose que eacute abundante nos fluiacutedos de seus hospedeiros
vertebrados Jaacute as formas amastigotas encontradas no citoplasma das ceacutelulas do hospedeiro
mamiacutefero ambiente pobre em glicose livre usam os aminoaacutecidos como principal fonte de
carbono e energia apresentando maior quantidade de prolina intracelular do que as formas
tripomastigotas (Silber et al 2009)
Entre os aminoaacutecidos metabolizados pelas formas epimastigotas estatildeo prolina glicina
asparagina glutamina aspartato glutamato leucina e isoleucina (Mancilla et al 1967 Odaly
et al 1983 Silber et al 2005a Sylvester Krassner 1976 Zeledon 1960) Os aminoaacutecidos
estatildeo envolvidos tambeacutem em outros processos importantes do ciclo de vida do T cruzi O
aminoaacutecido L-prolina eacute essencial na diferenciaccedilatildeo das formas epimastigota intracelular para
tripomastigotas aleacutem de estarem envolvidos nos processos de osmorregulaccedilatildeo crescimento e
resistecircncia do parasita ao estresse oxidativo e nutricional (Homsy et al 1989 Magdaleno et
al 2009 Magdaleno et al 2011 Pereira et al 2002 Pereira et al 2003 Tonelli et al
2004) Glutamato prolina e aspartato satildeo importantes na metaciclogecircnese ou seja na
diferenciaccedilatildeo de formas epimastigotas para tripomastigotas metaciacuteclicos (Contreras et al
1985) Prolina glicina alanina e glutamato satildeo descritos como importantes nos processos de
osmorregulaccedilatildeo e controle do volume celular (Rohloff et al 2003 Rohloff et al 2004a)
Os aminoaacutecidos para que sejam metabolizados ou cumprirem outras funccedilotildees devem
ser obtidos a partir da degradaccedilatildeo de proteiacutenas sintetizados ou incorporados pelo parasita
atraveacutes de transportadores localizados na membrana (Silber et al 2005a) O primeiro
transportador de aminoaacutecidos em T cruzi foi descrito bioquimicamente em 1970 o
transportador de lisina (Hampton 1970) Outros aminoaacutecidos tambeacutem jaacute tiveram a sua
cineacutetica e seu mecanismo de transporte descrito para T cruzi A incorporaccedilatildeo de arginina
pelo parasita apresenta pelo menos dois sistemas de transporte sendo um de alta afinidade e
outro de baixa afinidade Esse transporte eacute regulado ao longo do ciclo de vida do parasita
21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

RPMI Roswell Park Memorial Institute
SDS Soacutedio dodecil sulfat
SFB Soro fetal bovino
U Unidades
μg Micrograma
μl Microlitro
μm Microcircmetro
SUMAacuteRIO
1 INTRODUCcedilAtildeO 16
11 Aspectos gerais da doenccedila de chagas 16
12 Trypanosoma cruzi 18
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos 20
14 Morte celular em Trypanosoma cruzi 21
15 Terapia na doenccedila de Chagas 23
16 Perspectivas de novos faacutermacos 25
2 OBJETIVOS 30
21 Objetivo geral 30
22 Objetivos especiacuteficos 30
3 MATERIAIS E MEacuteTODOS 31
31 Microorganismos utilizados no trabalho 31
311 Ceacutelulas hospedeiras 31
312 Trypanosoma cruzi 31
32 Manutenccedilatildeo das ceacutelulas hospedeiras 31
33 Obtenccedilatildeo dos parasitas 31
331 Epimastigotas 31
332 Curvas de crescimento de epimastigotas de T cruzi 31
333 Formas tripomastigotas 32
34 Resistecircncia a diferentes tipos de estresse 32
341 Ensaio de estresse teacutermico em formas epimastigotas 32
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico 33
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular 33
343 Ensaio de estresse oxidativo em formas epimastigotas 33
35 Tipo de morte celular em parasitas tratados com Memantina 34
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita 34
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas
epimastigotas 35
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio 35
36 Efeito sobre a metaciclogecircnese 35
37 Anaacutelise do efeito dos compostos no ciclo intracelular do T cruzi 36
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos 36
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas 36
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas
tripomastigotas 37
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os
compostos 37
38 Anaacutelises estatiacutesticas 38
4 RESULTADOS 39
41 Curvas de crescimento do T cruzi na presenccedila dos compostos 39
42 Interaccedilatildeo com condiccedilotildees de estresse 41
43 Efeito da Memantina na metaciclogecircnese 46
44 Tipo de morte celular sofrida pelo parasita tratado com Memantina 47
45 Efeito da Memantina no ciclo intracelular do T cruzi 52
5 DISCUSSAtildeO 55
6 CONCLUSOtildeES 59
REFEREcircNCIAS 61
16
1 INTRODUCcedilAtildeO
11 Aspectos gerais da doenccedila de chagas
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana foi
descoberta pelo meacutedico brasileiro Carlos Chagas (Chagas 1909) Esta doenccedila eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica Central e do Sul com cerca de 25 milhotildees de pessoas
vivendo em aacutereas de risco de infecccedilatildeo Ocorrem por ano aproximadamente 40 mil novos
casos e 12 mil mortes devido a complicaccedilotildees cardiacuteacas e digestivas (World Health
Organization 2012) Programas de controle do vetor desenvolvidos nos uacuteltimos 20 anos
como tambeacutem um maior controle nos bancos de sangue tem reduzido o nuacutemero de novos
casos da doenccedila de Chagas na Ameacuterica Latina (Rassi Marin-Neto 2010) Devido as intensas
migraccedilotildees de pessoas de paiacuteses endecircmicos o nuacutemero de casos de indiviacuteduos infectados tem
aumentado em aacutereas consideradas natildeo endecircmicas tais como Estados Unidos Canadaacute e
algumas regiotildees da Europa (Bern et al 2009 Maguire 2006 Schmunis 2007) A doenccedila eacute
uma zoonose que tem como reservatoacuterio natural do T cruzi uma ampla variedade de
mamiacuteferos do continente americano (Urbina Docampo 2003) Eacute transmitida para os seres
humanos e para mais de 150 espeacutecies de animais domeacutesticos e animais selvagens
principalmente por insetos hematoacutefagos da famiacutelia Reduviidae Apesar de existirem mais de
130 espeacutecies de triatomiacuteneos satildeo trecircs as mais importantes na transmissatildeo do T cruzi para o
homem Triatoma infestans Rhodnius prolixus e Triatoma dimidiata (Rassi Marin-Neto
2010) O parasita tambeacutem pode ser transmitido por transfusatildeo de sangue congenitamente (de
matildees infectadas para receacutem-nascidos) por transplante de oacutergatildeos ou medula oacutessea Tambeacutem
pode ocorrer por acidentes em laboratoacuterios que trabalham com o parasita vivo via esta que
epidemiologicamente eacute pouco relevante Outra forma de transmissatildeo eacute por via oral atraveacutes de
alimentos contaminados essa via vecircm sendo considerada como relevante na manutenccedilatildeo do
ciclo de transmissatildeo natural (Pereira et al 2009 Rassi Marin-Neto 2010)
A evoluccedilatildeo cliacutenica da doenccedila de Chagas pode ser dividida em duas fases aguda e
crocircnica A fase aguda normalmente eacute assintomaacutetica ou apresenta sintomatologia discreta que
na maioria dos casos passa despercebida pelo paciente Os sintomas quando aparecem o
fazem uma ou duas semanas apoacutes a exposiccedilatildeo ao triatomiacuteneo infectado ou poucos meses apoacutes
a transfusatildeo com sangue contaminado (Rassi Marin-Neto 2010) Os sintomas mais
frequentemente apresentados pelos pacientes agudos satildeo febre prolongada mal-estar
17
distensatildeo do fiacutegado baccedilo e linfonodos Pode haver tambeacutem edema subcutacircneo localizado ou
generalizado No caso particular de transmissatildeo vetorial o paciente pode apresentar um sinal
da porta de entrada do T cruzi na pele (chagoma) ou via mucosa ocular (sinal de Romantildea) As
manifestaccedilotildees da fase aguda se resolvem espontaneamente em aproximadamente 90 dos
indiviacuteduos infectados mesmo sem o tratamento com drogas tripanocidas Essa fase eacute
caracterizada pela alta parasitemia e ausecircncia de anticorpos especiacuteficos Nessa fase os
parasitas podem ser encontrados em praticamente todos os tecidos estando majoritariamente
presentes no sangue Por ser uma fase de curta duraccedilatildeo sem sintomatologia aparente ou com
sintomatologia pouco definida haacute um pequeno nuacutemero de casos detectados sendo portanto a
fase menos estudada (Rassi Marin-Neto 2010 Silber et al 2005b)
Logo apoacutes a fase aguda tem iniacutecio a fase crocircnica caracterizada pelos altos tiacutetulos de
anticorpos no sangue do paciente e pelo fato de que a parasitemia eacute inaparente O paciente
pode permanecer por vaacuterios anos ou ateacute mesmo pelo resto da vida sem apresentar sinais ou
sintomas Essa forma da fase crocircnica eacute chamada de indeterminada Aproximadamente 30-40
dos pacientes crocircnicos indeterminados desenvolvem a sintomatologia caracteriacutestica da
doenccedila evoluindo para a chamada fase crocircnica sintomaacutetica Nesta fase as trecircs principais
formas satildeo a cardiacuteaca a digestiva e a cardiodigestiva (Acquatella 2007 Marin-Neto et al
1999) Na forma cardiacuteaca o coraccedilatildeo eacute um dos primeiros oacutergatildeos afetados natildeo necessariamente
o uacutenico desenvolvendo hipertrofia e dilataccedilatildeo Na forma digestiva o trato digestivo apresenta
alteraccedilotildees anatocircmicas e fisioloacutegicas importantes sendo a principal altereccedilatildeo a dilataccedilatildeo do
intestino e do esocircfago que podem apresentar aspecto de megaviacutescera (Marin-Neto et al
1999 Rassi et al 2000) revisado por (Boscardin et al 2009 Rassi Marin-Neto 2010)
18
12 Trypanosoma cruzi
Trypanosoma cruzi eacute um parasita intracelular obrigatoacuterio classicamente localizado na
ordem kinetoplastida gecircnero Trypanosoma que causa uma infecccedilatildeo crocircnica em humanos e
em grande nuacutemero de outras espeacutecies de mamiacuteferos (Hoare 1972) Recentemente devido agrave
alta variabilidade bioloacutegica bioquiacutemica e molecular a espeacutecie Trypanosoma cruzi foi
dividida em seis linhagens subsespeciacuteficas (Discrete Typing Units ndash DTUs) numerados de I a
VI Esta classificaccedilatildeo foi realizada com base em caracteriacutesticas geneacuteticas moleculares e
imunoloacutegicas com o objetivo de unificar a nomenclatura e facilitar a comunicaccedilatildeo dentro da
comunidade cientiacutefica (Zingales et al 2009)
O T cruzi tem um ciclo de vida complexo que alterna entre o inseto vetor e o
hospedeiro mamiacutefero O parasita apresenta diferentes formas correspondentes a diferentes
estaacutegios do seu ciclo de vida Estes diferentes estaacutegios tecircm sido descritos com base em
criteacuterios morfoloacutegicos como a forma fusiforme ou arredondados a regiatildeo onde emerge o
flagelo e a posiccedilatildeo do cinetoplasto (estrutura contendo o DNA da uacutenica mitococircndria presente
no parasita) em relaccedilatildeo ao nuacutecleo No hospedeiro mamiacutefero foram descritas as formas
amastigota epimastigota intracelular e tripomastigota sanguiacutecola A forma amastigota eacute um
estaacutegio replicativo e infectivo Satildeo ceacutelulas arredondadas medindo de 2-6 microm de diacircmetro
encontradas no citoplasma das ceacutelulas infectadas Os epimastigotas intracelulares
(epimastigote-like) satildeo formas replicativas intermediaacuterias entre amastigotas e tripomastigotas
tambeacutem encontrados dentro das ceacutelulas medindo aproximadamente 54 microm Essas formas satildeo
menores que as formas epimastigotas encontradas no inseto vetor poreacutem compartilha com as
mesmas padrotildees morfomeacutetricos e diversas caracteriacutesticas bioquiacutemicas e bioloacutegicas (Almeida-
De-Faria et al 1999) O tripomastigota sanguiacutecola eacute um estaacutegio flagelado infectivo natildeo
replicativo encontrado principalmente na corrente sanguiacutenea do hospedeiro mamiacutefero Mede
aproximadamente 18 microm de comprimento incluindo um extremo livre do flagelo de
aproximadamente 6 microm e 2-3 microm de largura
No inseto vetor encontram-se predominantemente as formas epimastigota e
tripomastigota metaciacuteclico Epimastigota eacute uma forma flagelada replicativa e natildeo infectiva
Mede aproximadamente 20-40 microm de comprimento e 2-5 microm de largura essas formas estatildeo
presentes tipicamente na porccedilatildeo meacutedia do intestino do hospedeiro invertebrado
Tripomastigota metaciacuteclico eacute uma forma infectiva natildeo replicativa encontrada na porccedilatildeo final
do intestino do inseto vetor (Brener 1971) revisado por (Silber et al 2005a Tyler Engman
2001)
19
O ciclo de vida do T cruzi inicia-se quando os parasitas presentes na corrente
sanguiacutenea do hospedeiro vertebrado infectado passam ao inseto vetor durante o repasto
sanguiacuteneo No intestino do inseto estas formas se diferenciam em formas epimastigotas (De
Souza et al 2002) que apoacutes multiplicaccedilatildeo por fissatildeo binaacuteria colonizam a porccedilatildeo terminal do
intestino e se diferenciam novamente em tripomastigotas metaciacuteclicos Quando o inseto
infectado pica o hospedeiro vertebrado e inicia o repasto sanguiacuteneo concomitantemente
elimina as formas tripomastigotas metaciacuteclicos junto com as fezes e urina Uma vez em
contato com a pele ou mucosas do hospedeiro mamiacutefero os tripomastigotas satildeo internalizados
atraveacutes de pequenas lesotildees na pele causadas pelo proacuteprio hospedeiro (Alves Colli 2008) Os
tripomastigotas invadem as ceacutelulas do hospedeiro mamiacutefero e se diferenciam em formas
replicativas denominadas amastigotas estabelecendo assim a infecccedilatildeo Apoacutes
aproximadamente 96 horas os amastigotas comeccedilam a se diferenciar em tripomastigotas
passando pela forma intermediaacuteria epimastigota intracelular Estas formas tripomastigotas satildeo
liberadas na corrente sanguiacutenea podendo invadir novas ceacutelulas ou serem ingeridas pelo inseto
vetor em um novo repasto sanguiacuteneo fechando assim o seu ciclo de vida O esquema do ciclo
de vida do Trypanosoma cruzi estaacute representado na Figura 1
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi
20
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos
T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono As formas epimastigotas preferencialmente catabolizam glicose e apoacutes a exaustatildeo
do carboidrato ocorre uma mudanccedila para o metabolismo de aminoaacutecidos A prolina e a
glutamina satildeo os principais aminoaacutecidos metabolizados pelas formas epimastigotas visto que
satildeo abudantes na hemolinfa e nos fluiacutedos dos tecidos do inseto vetor Esse metabolismo
ocorre com produccedilatildeo de NH4+
(produto da deaminaccedilatildeo dos aminoaacutecidos) que eacute excretada para
o meio extracelular (Bringaud et al 2006 Cazzulo 1994) As formas tripomastigotas
utilizam principalmente a glicose que eacute abundante nos fluiacutedos de seus hospedeiros
vertebrados Jaacute as formas amastigotas encontradas no citoplasma das ceacutelulas do hospedeiro
mamiacutefero ambiente pobre em glicose livre usam os aminoaacutecidos como principal fonte de
carbono e energia apresentando maior quantidade de prolina intracelular do que as formas
tripomastigotas (Silber et al 2009)
Entre os aminoaacutecidos metabolizados pelas formas epimastigotas estatildeo prolina glicina
asparagina glutamina aspartato glutamato leucina e isoleucina (Mancilla et al 1967 Odaly
et al 1983 Silber et al 2005a Sylvester Krassner 1976 Zeledon 1960) Os aminoaacutecidos
estatildeo envolvidos tambeacutem em outros processos importantes do ciclo de vida do T cruzi O
aminoaacutecido L-prolina eacute essencial na diferenciaccedilatildeo das formas epimastigota intracelular para
tripomastigotas aleacutem de estarem envolvidos nos processos de osmorregulaccedilatildeo crescimento e
resistecircncia do parasita ao estresse oxidativo e nutricional (Homsy et al 1989 Magdaleno et
al 2009 Magdaleno et al 2011 Pereira et al 2002 Pereira et al 2003 Tonelli et al
2004) Glutamato prolina e aspartato satildeo importantes na metaciclogecircnese ou seja na
diferenciaccedilatildeo de formas epimastigotas para tripomastigotas metaciacuteclicos (Contreras et al
1985) Prolina glicina alanina e glutamato satildeo descritos como importantes nos processos de
osmorregulaccedilatildeo e controle do volume celular (Rohloff et al 2003 Rohloff et al 2004a)
Os aminoaacutecidos para que sejam metabolizados ou cumprirem outras funccedilotildees devem
ser obtidos a partir da degradaccedilatildeo de proteiacutenas sintetizados ou incorporados pelo parasita
atraveacutes de transportadores localizados na membrana (Silber et al 2005a) O primeiro
transportador de aminoaacutecidos em T cruzi foi descrito bioquimicamente em 1970 o
transportador de lisina (Hampton 1970) Outros aminoaacutecidos tambeacutem jaacute tiveram a sua
cineacutetica e seu mecanismo de transporte descrito para T cruzi A incorporaccedilatildeo de arginina
pelo parasita apresenta pelo menos dois sistemas de transporte sendo um de alta afinidade e
outro de baixa afinidade Esse transporte eacute regulado ao longo do ciclo de vida do parasita
21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

SUMAacuteRIO
1 INTRODUCcedilAtildeO 16
11 Aspectos gerais da doenccedila de chagas 16
12 Trypanosoma cruzi 18
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos 20
14 Morte celular em Trypanosoma cruzi 21
15 Terapia na doenccedila de Chagas 23
16 Perspectivas de novos faacutermacos 25
2 OBJETIVOS 30
21 Objetivo geral 30
22 Objetivos especiacuteficos 30
3 MATERIAIS E MEacuteTODOS 31
31 Microorganismos utilizados no trabalho 31
311 Ceacutelulas hospedeiras 31
312 Trypanosoma cruzi 31
32 Manutenccedilatildeo das ceacutelulas hospedeiras 31
33 Obtenccedilatildeo dos parasitas 31
331 Epimastigotas 31
332 Curvas de crescimento de epimastigotas de T cruzi 31
333 Formas tripomastigotas 32
34 Resistecircncia a diferentes tipos de estresse 32
341 Ensaio de estresse teacutermico em formas epimastigotas 32
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico 33
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular 33
343 Ensaio de estresse oxidativo em formas epimastigotas 33
35 Tipo de morte celular em parasitas tratados com Memantina 34
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita 34
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas
epimastigotas 35
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio 35
36 Efeito sobre a metaciclogecircnese 35
37 Anaacutelise do efeito dos compostos no ciclo intracelular do T cruzi 36
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos 36
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas 36
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas
tripomastigotas 37
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os
compostos 37
38 Anaacutelises estatiacutesticas 38
4 RESULTADOS 39
41 Curvas de crescimento do T cruzi na presenccedila dos compostos 39
42 Interaccedilatildeo com condiccedilotildees de estresse 41
43 Efeito da Memantina na metaciclogecircnese 46
44 Tipo de morte celular sofrida pelo parasita tratado com Memantina 47
45 Efeito da Memantina no ciclo intracelular do T cruzi 52
5 DISCUSSAtildeO 55
6 CONCLUSOtildeES 59
REFEREcircNCIAS 61
16
1 INTRODUCcedilAtildeO
11 Aspectos gerais da doenccedila de chagas
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana foi
descoberta pelo meacutedico brasileiro Carlos Chagas (Chagas 1909) Esta doenccedila eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica Central e do Sul com cerca de 25 milhotildees de pessoas
vivendo em aacutereas de risco de infecccedilatildeo Ocorrem por ano aproximadamente 40 mil novos
casos e 12 mil mortes devido a complicaccedilotildees cardiacuteacas e digestivas (World Health
Organization 2012) Programas de controle do vetor desenvolvidos nos uacuteltimos 20 anos
como tambeacutem um maior controle nos bancos de sangue tem reduzido o nuacutemero de novos
casos da doenccedila de Chagas na Ameacuterica Latina (Rassi Marin-Neto 2010) Devido as intensas
migraccedilotildees de pessoas de paiacuteses endecircmicos o nuacutemero de casos de indiviacuteduos infectados tem
aumentado em aacutereas consideradas natildeo endecircmicas tais como Estados Unidos Canadaacute e
algumas regiotildees da Europa (Bern et al 2009 Maguire 2006 Schmunis 2007) A doenccedila eacute
uma zoonose que tem como reservatoacuterio natural do T cruzi uma ampla variedade de
mamiacuteferos do continente americano (Urbina Docampo 2003) Eacute transmitida para os seres
humanos e para mais de 150 espeacutecies de animais domeacutesticos e animais selvagens
principalmente por insetos hematoacutefagos da famiacutelia Reduviidae Apesar de existirem mais de
130 espeacutecies de triatomiacuteneos satildeo trecircs as mais importantes na transmissatildeo do T cruzi para o
homem Triatoma infestans Rhodnius prolixus e Triatoma dimidiata (Rassi Marin-Neto
2010) O parasita tambeacutem pode ser transmitido por transfusatildeo de sangue congenitamente (de
matildees infectadas para receacutem-nascidos) por transplante de oacutergatildeos ou medula oacutessea Tambeacutem
pode ocorrer por acidentes em laboratoacuterios que trabalham com o parasita vivo via esta que
epidemiologicamente eacute pouco relevante Outra forma de transmissatildeo eacute por via oral atraveacutes de
alimentos contaminados essa via vecircm sendo considerada como relevante na manutenccedilatildeo do
ciclo de transmissatildeo natural (Pereira et al 2009 Rassi Marin-Neto 2010)
A evoluccedilatildeo cliacutenica da doenccedila de Chagas pode ser dividida em duas fases aguda e
crocircnica A fase aguda normalmente eacute assintomaacutetica ou apresenta sintomatologia discreta que
na maioria dos casos passa despercebida pelo paciente Os sintomas quando aparecem o
fazem uma ou duas semanas apoacutes a exposiccedilatildeo ao triatomiacuteneo infectado ou poucos meses apoacutes
a transfusatildeo com sangue contaminado (Rassi Marin-Neto 2010) Os sintomas mais
frequentemente apresentados pelos pacientes agudos satildeo febre prolongada mal-estar
17
distensatildeo do fiacutegado baccedilo e linfonodos Pode haver tambeacutem edema subcutacircneo localizado ou
generalizado No caso particular de transmissatildeo vetorial o paciente pode apresentar um sinal
da porta de entrada do T cruzi na pele (chagoma) ou via mucosa ocular (sinal de Romantildea) As
manifestaccedilotildees da fase aguda se resolvem espontaneamente em aproximadamente 90 dos
indiviacuteduos infectados mesmo sem o tratamento com drogas tripanocidas Essa fase eacute
caracterizada pela alta parasitemia e ausecircncia de anticorpos especiacuteficos Nessa fase os
parasitas podem ser encontrados em praticamente todos os tecidos estando majoritariamente
presentes no sangue Por ser uma fase de curta duraccedilatildeo sem sintomatologia aparente ou com
sintomatologia pouco definida haacute um pequeno nuacutemero de casos detectados sendo portanto a
fase menos estudada (Rassi Marin-Neto 2010 Silber et al 2005b)
Logo apoacutes a fase aguda tem iniacutecio a fase crocircnica caracterizada pelos altos tiacutetulos de
anticorpos no sangue do paciente e pelo fato de que a parasitemia eacute inaparente O paciente
pode permanecer por vaacuterios anos ou ateacute mesmo pelo resto da vida sem apresentar sinais ou
sintomas Essa forma da fase crocircnica eacute chamada de indeterminada Aproximadamente 30-40
dos pacientes crocircnicos indeterminados desenvolvem a sintomatologia caracteriacutestica da
doenccedila evoluindo para a chamada fase crocircnica sintomaacutetica Nesta fase as trecircs principais
formas satildeo a cardiacuteaca a digestiva e a cardiodigestiva (Acquatella 2007 Marin-Neto et al
1999) Na forma cardiacuteaca o coraccedilatildeo eacute um dos primeiros oacutergatildeos afetados natildeo necessariamente
o uacutenico desenvolvendo hipertrofia e dilataccedilatildeo Na forma digestiva o trato digestivo apresenta
alteraccedilotildees anatocircmicas e fisioloacutegicas importantes sendo a principal altereccedilatildeo a dilataccedilatildeo do
intestino e do esocircfago que podem apresentar aspecto de megaviacutescera (Marin-Neto et al
1999 Rassi et al 2000) revisado por (Boscardin et al 2009 Rassi Marin-Neto 2010)
18
12 Trypanosoma cruzi
Trypanosoma cruzi eacute um parasita intracelular obrigatoacuterio classicamente localizado na
ordem kinetoplastida gecircnero Trypanosoma que causa uma infecccedilatildeo crocircnica em humanos e
em grande nuacutemero de outras espeacutecies de mamiacuteferos (Hoare 1972) Recentemente devido agrave
alta variabilidade bioloacutegica bioquiacutemica e molecular a espeacutecie Trypanosoma cruzi foi
dividida em seis linhagens subsespeciacuteficas (Discrete Typing Units ndash DTUs) numerados de I a
VI Esta classificaccedilatildeo foi realizada com base em caracteriacutesticas geneacuteticas moleculares e
imunoloacutegicas com o objetivo de unificar a nomenclatura e facilitar a comunicaccedilatildeo dentro da
comunidade cientiacutefica (Zingales et al 2009)
O T cruzi tem um ciclo de vida complexo que alterna entre o inseto vetor e o
hospedeiro mamiacutefero O parasita apresenta diferentes formas correspondentes a diferentes
estaacutegios do seu ciclo de vida Estes diferentes estaacutegios tecircm sido descritos com base em
criteacuterios morfoloacutegicos como a forma fusiforme ou arredondados a regiatildeo onde emerge o
flagelo e a posiccedilatildeo do cinetoplasto (estrutura contendo o DNA da uacutenica mitococircndria presente
no parasita) em relaccedilatildeo ao nuacutecleo No hospedeiro mamiacutefero foram descritas as formas
amastigota epimastigota intracelular e tripomastigota sanguiacutecola A forma amastigota eacute um
estaacutegio replicativo e infectivo Satildeo ceacutelulas arredondadas medindo de 2-6 microm de diacircmetro
encontradas no citoplasma das ceacutelulas infectadas Os epimastigotas intracelulares
(epimastigote-like) satildeo formas replicativas intermediaacuterias entre amastigotas e tripomastigotas
tambeacutem encontrados dentro das ceacutelulas medindo aproximadamente 54 microm Essas formas satildeo
menores que as formas epimastigotas encontradas no inseto vetor poreacutem compartilha com as
mesmas padrotildees morfomeacutetricos e diversas caracteriacutesticas bioquiacutemicas e bioloacutegicas (Almeida-
De-Faria et al 1999) O tripomastigota sanguiacutecola eacute um estaacutegio flagelado infectivo natildeo
replicativo encontrado principalmente na corrente sanguiacutenea do hospedeiro mamiacutefero Mede
aproximadamente 18 microm de comprimento incluindo um extremo livre do flagelo de
aproximadamente 6 microm e 2-3 microm de largura
No inseto vetor encontram-se predominantemente as formas epimastigota e
tripomastigota metaciacuteclico Epimastigota eacute uma forma flagelada replicativa e natildeo infectiva
Mede aproximadamente 20-40 microm de comprimento e 2-5 microm de largura essas formas estatildeo
presentes tipicamente na porccedilatildeo meacutedia do intestino do hospedeiro invertebrado
Tripomastigota metaciacuteclico eacute uma forma infectiva natildeo replicativa encontrada na porccedilatildeo final
do intestino do inseto vetor (Brener 1971) revisado por (Silber et al 2005a Tyler Engman
2001)
19
O ciclo de vida do T cruzi inicia-se quando os parasitas presentes na corrente
sanguiacutenea do hospedeiro vertebrado infectado passam ao inseto vetor durante o repasto
sanguiacuteneo No intestino do inseto estas formas se diferenciam em formas epimastigotas (De
Souza et al 2002) que apoacutes multiplicaccedilatildeo por fissatildeo binaacuteria colonizam a porccedilatildeo terminal do
intestino e se diferenciam novamente em tripomastigotas metaciacuteclicos Quando o inseto
infectado pica o hospedeiro vertebrado e inicia o repasto sanguiacuteneo concomitantemente
elimina as formas tripomastigotas metaciacuteclicos junto com as fezes e urina Uma vez em
contato com a pele ou mucosas do hospedeiro mamiacutefero os tripomastigotas satildeo internalizados
atraveacutes de pequenas lesotildees na pele causadas pelo proacuteprio hospedeiro (Alves Colli 2008) Os
tripomastigotas invadem as ceacutelulas do hospedeiro mamiacutefero e se diferenciam em formas
replicativas denominadas amastigotas estabelecendo assim a infecccedilatildeo Apoacutes
aproximadamente 96 horas os amastigotas comeccedilam a se diferenciar em tripomastigotas
passando pela forma intermediaacuteria epimastigota intracelular Estas formas tripomastigotas satildeo
liberadas na corrente sanguiacutenea podendo invadir novas ceacutelulas ou serem ingeridas pelo inseto
vetor em um novo repasto sanguiacuteneo fechando assim o seu ciclo de vida O esquema do ciclo
de vida do Trypanosoma cruzi estaacute representado na Figura 1
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi
20
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos
T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono As formas epimastigotas preferencialmente catabolizam glicose e apoacutes a exaustatildeo
do carboidrato ocorre uma mudanccedila para o metabolismo de aminoaacutecidos A prolina e a
glutamina satildeo os principais aminoaacutecidos metabolizados pelas formas epimastigotas visto que
satildeo abudantes na hemolinfa e nos fluiacutedos dos tecidos do inseto vetor Esse metabolismo
ocorre com produccedilatildeo de NH4+
(produto da deaminaccedilatildeo dos aminoaacutecidos) que eacute excretada para
o meio extracelular (Bringaud et al 2006 Cazzulo 1994) As formas tripomastigotas
utilizam principalmente a glicose que eacute abundante nos fluiacutedos de seus hospedeiros
vertebrados Jaacute as formas amastigotas encontradas no citoplasma das ceacutelulas do hospedeiro
mamiacutefero ambiente pobre em glicose livre usam os aminoaacutecidos como principal fonte de
carbono e energia apresentando maior quantidade de prolina intracelular do que as formas
tripomastigotas (Silber et al 2009)
Entre os aminoaacutecidos metabolizados pelas formas epimastigotas estatildeo prolina glicina
asparagina glutamina aspartato glutamato leucina e isoleucina (Mancilla et al 1967 Odaly
et al 1983 Silber et al 2005a Sylvester Krassner 1976 Zeledon 1960) Os aminoaacutecidos
estatildeo envolvidos tambeacutem em outros processos importantes do ciclo de vida do T cruzi O
aminoaacutecido L-prolina eacute essencial na diferenciaccedilatildeo das formas epimastigota intracelular para
tripomastigotas aleacutem de estarem envolvidos nos processos de osmorregulaccedilatildeo crescimento e
resistecircncia do parasita ao estresse oxidativo e nutricional (Homsy et al 1989 Magdaleno et
al 2009 Magdaleno et al 2011 Pereira et al 2002 Pereira et al 2003 Tonelli et al
2004) Glutamato prolina e aspartato satildeo importantes na metaciclogecircnese ou seja na
diferenciaccedilatildeo de formas epimastigotas para tripomastigotas metaciacuteclicos (Contreras et al
1985) Prolina glicina alanina e glutamato satildeo descritos como importantes nos processos de
osmorregulaccedilatildeo e controle do volume celular (Rohloff et al 2003 Rohloff et al 2004a)
Os aminoaacutecidos para que sejam metabolizados ou cumprirem outras funccedilotildees devem
ser obtidos a partir da degradaccedilatildeo de proteiacutenas sintetizados ou incorporados pelo parasita
atraveacutes de transportadores localizados na membrana (Silber et al 2005a) O primeiro
transportador de aminoaacutecidos em T cruzi foi descrito bioquimicamente em 1970 o
transportador de lisina (Hampton 1970) Outros aminoaacutecidos tambeacutem jaacute tiveram a sua
cineacutetica e seu mecanismo de transporte descrito para T cruzi A incorporaccedilatildeo de arginina
pelo parasita apresenta pelo menos dois sistemas de transporte sendo um de alta afinidade e
outro de baixa afinidade Esse transporte eacute regulado ao longo do ciclo de vida do parasita
21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas 36
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas
tripomastigotas 37
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os
compostos 37
38 Anaacutelises estatiacutesticas 38
4 RESULTADOS 39
41 Curvas de crescimento do T cruzi na presenccedila dos compostos 39
42 Interaccedilatildeo com condiccedilotildees de estresse 41
43 Efeito da Memantina na metaciclogecircnese 46
44 Tipo de morte celular sofrida pelo parasita tratado com Memantina 47
45 Efeito da Memantina no ciclo intracelular do T cruzi 52
5 DISCUSSAtildeO 55
6 CONCLUSOtildeES 59
REFEREcircNCIAS 61
16
1 INTRODUCcedilAtildeO
11 Aspectos gerais da doenccedila de chagas
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana foi
descoberta pelo meacutedico brasileiro Carlos Chagas (Chagas 1909) Esta doenccedila eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica Central e do Sul com cerca de 25 milhotildees de pessoas
vivendo em aacutereas de risco de infecccedilatildeo Ocorrem por ano aproximadamente 40 mil novos
casos e 12 mil mortes devido a complicaccedilotildees cardiacuteacas e digestivas (World Health
Organization 2012) Programas de controle do vetor desenvolvidos nos uacuteltimos 20 anos
como tambeacutem um maior controle nos bancos de sangue tem reduzido o nuacutemero de novos
casos da doenccedila de Chagas na Ameacuterica Latina (Rassi Marin-Neto 2010) Devido as intensas
migraccedilotildees de pessoas de paiacuteses endecircmicos o nuacutemero de casos de indiviacuteduos infectados tem
aumentado em aacutereas consideradas natildeo endecircmicas tais como Estados Unidos Canadaacute e
algumas regiotildees da Europa (Bern et al 2009 Maguire 2006 Schmunis 2007) A doenccedila eacute
uma zoonose que tem como reservatoacuterio natural do T cruzi uma ampla variedade de
mamiacuteferos do continente americano (Urbina Docampo 2003) Eacute transmitida para os seres
humanos e para mais de 150 espeacutecies de animais domeacutesticos e animais selvagens
principalmente por insetos hematoacutefagos da famiacutelia Reduviidae Apesar de existirem mais de
130 espeacutecies de triatomiacuteneos satildeo trecircs as mais importantes na transmissatildeo do T cruzi para o
homem Triatoma infestans Rhodnius prolixus e Triatoma dimidiata (Rassi Marin-Neto
2010) O parasita tambeacutem pode ser transmitido por transfusatildeo de sangue congenitamente (de
matildees infectadas para receacutem-nascidos) por transplante de oacutergatildeos ou medula oacutessea Tambeacutem
pode ocorrer por acidentes em laboratoacuterios que trabalham com o parasita vivo via esta que
epidemiologicamente eacute pouco relevante Outra forma de transmissatildeo eacute por via oral atraveacutes de
alimentos contaminados essa via vecircm sendo considerada como relevante na manutenccedilatildeo do
ciclo de transmissatildeo natural (Pereira et al 2009 Rassi Marin-Neto 2010)
A evoluccedilatildeo cliacutenica da doenccedila de Chagas pode ser dividida em duas fases aguda e
crocircnica A fase aguda normalmente eacute assintomaacutetica ou apresenta sintomatologia discreta que
na maioria dos casos passa despercebida pelo paciente Os sintomas quando aparecem o
fazem uma ou duas semanas apoacutes a exposiccedilatildeo ao triatomiacuteneo infectado ou poucos meses apoacutes
a transfusatildeo com sangue contaminado (Rassi Marin-Neto 2010) Os sintomas mais
frequentemente apresentados pelos pacientes agudos satildeo febre prolongada mal-estar
17
distensatildeo do fiacutegado baccedilo e linfonodos Pode haver tambeacutem edema subcutacircneo localizado ou
generalizado No caso particular de transmissatildeo vetorial o paciente pode apresentar um sinal
da porta de entrada do T cruzi na pele (chagoma) ou via mucosa ocular (sinal de Romantildea) As
manifestaccedilotildees da fase aguda se resolvem espontaneamente em aproximadamente 90 dos
indiviacuteduos infectados mesmo sem o tratamento com drogas tripanocidas Essa fase eacute
caracterizada pela alta parasitemia e ausecircncia de anticorpos especiacuteficos Nessa fase os
parasitas podem ser encontrados em praticamente todos os tecidos estando majoritariamente
presentes no sangue Por ser uma fase de curta duraccedilatildeo sem sintomatologia aparente ou com
sintomatologia pouco definida haacute um pequeno nuacutemero de casos detectados sendo portanto a
fase menos estudada (Rassi Marin-Neto 2010 Silber et al 2005b)
Logo apoacutes a fase aguda tem iniacutecio a fase crocircnica caracterizada pelos altos tiacutetulos de
anticorpos no sangue do paciente e pelo fato de que a parasitemia eacute inaparente O paciente
pode permanecer por vaacuterios anos ou ateacute mesmo pelo resto da vida sem apresentar sinais ou
sintomas Essa forma da fase crocircnica eacute chamada de indeterminada Aproximadamente 30-40
dos pacientes crocircnicos indeterminados desenvolvem a sintomatologia caracteriacutestica da
doenccedila evoluindo para a chamada fase crocircnica sintomaacutetica Nesta fase as trecircs principais
formas satildeo a cardiacuteaca a digestiva e a cardiodigestiva (Acquatella 2007 Marin-Neto et al
1999) Na forma cardiacuteaca o coraccedilatildeo eacute um dos primeiros oacutergatildeos afetados natildeo necessariamente
o uacutenico desenvolvendo hipertrofia e dilataccedilatildeo Na forma digestiva o trato digestivo apresenta
alteraccedilotildees anatocircmicas e fisioloacutegicas importantes sendo a principal altereccedilatildeo a dilataccedilatildeo do
intestino e do esocircfago que podem apresentar aspecto de megaviacutescera (Marin-Neto et al
1999 Rassi et al 2000) revisado por (Boscardin et al 2009 Rassi Marin-Neto 2010)
18
12 Trypanosoma cruzi
Trypanosoma cruzi eacute um parasita intracelular obrigatoacuterio classicamente localizado na
ordem kinetoplastida gecircnero Trypanosoma que causa uma infecccedilatildeo crocircnica em humanos e
em grande nuacutemero de outras espeacutecies de mamiacuteferos (Hoare 1972) Recentemente devido agrave
alta variabilidade bioloacutegica bioquiacutemica e molecular a espeacutecie Trypanosoma cruzi foi
dividida em seis linhagens subsespeciacuteficas (Discrete Typing Units ndash DTUs) numerados de I a
VI Esta classificaccedilatildeo foi realizada com base em caracteriacutesticas geneacuteticas moleculares e
imunoloacutegicas com o objetivo de unificar a nomenclatura e facilitar a comunicaccedilatildeo dentro da
comunidade cientiacutefica (Zingales et al 2009)
O T cruzi tem um ciclo de vida complexo que alterna entre o inseto vetor e o
hospedeiro mamiacutefero O parasita apresenta diferentes formas correspondentes a diferentes
estaacutegios do seu ciclo de vida Estes diferentes estaacutegios tecircm sido descritos com base em
criteacuterios morfoloacutegicos como a forma fusiforme ou arredondados a regiatildeo onde emerge o
flagelo e a posiccedilatildeo do cinetoplasto (estrutura contendo o DNA da uacutenica mitococircndria presente
no parasita) em relaccedilatildeo ao nuacutecleo No hospedeiro mamiacutefero foram descritas as formas
amastigota epimastigota intracelular e tripomastigota sanguiacutecola A forma amastigota eacute um
estaacutegio replicativo e infectivo Satildeo ceacutelulas arredondadas medindo de 2-6 microm de diacircmetro
encontradas no citoplasma das ceacutelulas infectadas Os epimastigotas intracelulares
(epimastigote-like) satildeo formas replicativas intermediaacuterias entre amastigotas e tripomastigotas
tambeacutem encontrados dentro das ceacutelulas medindo aproximadamente 54 microm Essas formas satildeo
menores que as formas epimastigotas encontradas no inseto vetor poreacutem compartilha com as
mesmas padrotildees morfomeacutetricos e diversas caracteriacutesticas bioquiacutemicas e bioloacutegicas (Almeida-
De-Faria et al 1999) O tripomastigota sanguiacutecola eacute um estaacutegio flagelado infectivo natildeo
replicativo encontrado principalmente na corrente sanguiacutenea do hospedeiro mamiacutefero Mede
aproximadamente 18 microm de comprimento incluindo um extremo livre do flagelo de
aproximadamente 6 microm e 2-3 microm de largura
No inseto vetor encontram-se predominantemente as formas epimastigota e
tripomastigota metaciacuteclico Epimastigota eacute uma forma flagelada replicativa e natildeo infectiva
Mede aproximadamente 20-40 microm de comprimento e 2-5 microm de largura essas formas estatildeo
presentes tipicamente na porccedilatildeo meacutedia do intestino do hospedeiro invertebrado
Tripomastigota metaciacuteclico eacute uma forma infectiva natildeo replicativa encontrada na porccedilatildeo final
do intestino do inseto vetor (Brener 1971) revisado por (Silber et al 2005a Tyler Engman
2001)
19
O ciclo de vida do T cruzi inicia-se quando os parasitas presentes na corrente
sanguiacutenea do hospedeiro vertebrado infectado passam ao inseto vetor durante o repasto
sanguiacuteneo No intestino do inseto estas formas se diferenciam em formas epimastigotas (De
Souza et al 2002) que apoacutes multiplicaccedilatildeo por fissatildeo binaacuteria colonizam a porccedilatildeo terminal do
intestino e se diferenciam novamente em tripomastigotas metaciacuteclicos Quando o inseto
infectado pica o hospedeiro vertebrado e inicia o repasto sanguiacuteneo concomitantemente
elimina as formas tripomastigotas metaciacuteclicos junto com as fezes e urina Uma vez em
contato com a pele ou mucosas do hospedeiro mamiacutefero os tripomastigotas satildeo internalizados
atraveacutes de pequenas lesotildees na pele causadas pelo proacuteprio hospedeiro (Alves Colli 2008) Os
tripomastigotas invadem as ceacutelulas do hospedeiro mamiacutefero e se diferenciam em formas
replicativas denominadas amastigotas estabelecendo assim a infecccedilatildeo Apoacutes
aproximadamente 96 horas os amastigotas comeccedilam a se diferenciar em tripomastigotas
passando pela forma intermediaacuteria epimastigota intracelular Estas formas tripomastigotas satildeo
liberadas na corrente sanguiacutenea podendo invadir novas ceacutelulas ou serem ingeridas pelo inseto
vetor em um novo repasto sanguiacuteneo fechando assim o seu ciclo de vida O esquema do ciclo
de vida do Trypanosoma cruzi estaacute representado na Figura 1
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi
20
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos
T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono As formas epimastigotas preferencialmente catabolizam glicose e apoacutes a exaustatildeo
do carboidrato ocorre uma mudanccedila para o metabolismo de aminoaacutecidos A prolina e a
glutamina satildeo os principais aminoaacutecidos metabolizados pelas formas epimastigotas visto que
satildeo abudantes na hemolinfa e nos fluiacutedos dos tecidos do inseto vetor Esse metabolismo
ocorre com produccedilatildeo de NH4+
(produto da deaminaccedilatildeo dos aminoaacutecidos) que eacute excretada para
o meio extracelular (Bringaud et al 2006 Cazzulo 1994) As formas tripomastigotas
utilizam principalmente a glicose que eacute abundante nos fluiacutedos de seus hospedeiros
vertebrados Jaacute as formas amastigotas encontradas no citoplasma das ceacutelulas do hospedeiro
mamiacutefero ambiente pobre em glicose livre usam os aminoaacutecidos como principal fonte de
carbono e energia apresentando maior quantidade de prolina intracelular do que as formas
tripomastigotas (Silber et al 2009)
Entre os aminoaacutecidos metabolizados pelas formas epimastigotas estatildeo prolina glicina
asparagina glutamina aspartato glutamato leucina e isoleucina (Mancilla et al 1967 Odaly
et al 1983 Silber et al 2005a Sylvester Krassner 1976 Zeledon 1960) Os aminoaacutecidos
estatildeo envolvidos tambeacutem em outros processos importantes do ciclo de vida do T cruzi O
aminoaacutecido L-prolina eacute essencial na diferenciaccedilatildeo das formas epimastigota intracelular para
tripomastigotas aleacutem de estarem envolvidos nos processos de osmorregulaccedilatildeo crescimento e
resistecircncia do parasita ao estresse oxidativo e nutricional (Homsy et al 1989 Magdaleno et
al 2009 Magdaleno et al 2011 Pereira et al 2002 Pereira et al 2003 Tonelli et al
2004) Glutamato prolina e aspartato satildeo importantes na metaciclogecircnese ou seja na
diferenciaccedilatildeo de formas epimastigotas para tripomastigotas metaciacuteclicos (Contreras et al
1985) Prolina glicina alanina e glutamato satildeo descritos como importantes nos processos de
osmorregulaccedilatildeo e controle do volume celular (Rohloff et al 2003 Rohloff et al 2004a)
Os aminoaacutecidos para que sejam metabolizados ou cumprirem outras funccedilotildees devem
ser obtidos a partir da degradaccedilatildeo de proteiacutenas sintetizados ou incorporados pelo parasita
atraveacutes de transportadores localizados na membrana (Silber et al 2005a) O primeiro
transportador de aminoaacutecidos em T cruzi foi descrito bioquimicamente em 1970 o
transportador de lisina (Hampton 1970) Outros aminoaacutecidos tambeacutem jaacute tiveram a sua
cineacutetica e seu mecanismo de transporte descrito para T cruzi A incorporaccedilatildeo de arginina
pelo parasita apresenta pelo menos dois sistemas de transporte sendo um de alta afinidade e
outro de baixa afinidade Esse transporte eacute regulado ao longo do ciclo de vida do parasita
21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

16
1 INTRODUCcedilAtildeO
11 Aspectos gerais da doenccedila de chagas
A doenccedila de Chagas tambeacutem conhecida como tripanossomiacutease americana foi
descoberta pelo meacutedico brasileiro Carlos Chagas (Chagas 1909) Esta doenccedila eacute causada pelo
protozoaacuterio flagelado Trypanosoma cruzi e afeta aproximadamente 10 milhotildees de pessoas em
aacutereas endecircmicas do Meacutexico Ameacuterica Central e do Sul com cerca de 25 milhotildees de pessoas
vivendo em aacutereas de risco de infecccedilatildeo Ocorrem por ano aproximadamente 40 mil novos
casos e 12 mil mortes devido a complicaccedilotildees cardiacuteacas e digestivas (World Health
Organization 2012) Programas de controle do vetor desenvolvidos nos uacuteltimos 20 anos
como tambeacutem um maior controle nos bancos de sangue tem reduzido o nuacutemero de novos
casos da doenccedila de Chagas na Ameacuterica Latina (Rassi Marin-Neto 2010) Devido as intensas
migraccedilotildees de pessoas de paiacuteses endecircmicos o nuacutemero de casos de indiviacuteduos infectados tem
aumentado em aacutereas consideradas natildeo endecircmicas tais como Estados Unidos Canadaacute e
algumas regiotildees da Europa (Bern et al 2009 Maguire 2006 Schmunis 2007) A doenccedila eacute
uma zoonose que tem como reservatoacuterio natural do T cruzi uma ampla variedade de
mamiacuteferos do continente americano (Urbina Docampo 2003) Eacute transmitida para os seres
humanos e para mais de 150 espeacutecies de animais domeacutesticos e animais selvagens
principalmente por insetos hematoacutefagos da famiacutelia Reduviidae Apesar de existirem mais de
130 espeacutecies de triatomiacuteneos satildeo trecircs as mais importantes na transmissatildeo do T cruzi para o
homem Triatoma infestans Rhodnius prolixus e Triatoma dimidiata (Rassi Marin-Neto
2010) O parasita tambeacutem pode ser transmitido por transfusatildeo de sangue congenitamente (de
matildees infectadas para receacutem-nascidos) por transplante de oacutergatildeos ou medula oacutessea Tambeacutem
pode ocorrer por acidentes em laboratoacuterios que trabalham com o parasita vivo via esta que
epidemiologicamente eacute pouco relevante Outra forma de transmissatildeo eacute por via oral atraveacutes de
alimentos contaminados essa via vecircm sendo considerada como relevante na manutenccedilatildeo do
ciclo de transmissatildeo natural (Pereira et al 2009 Rassi Marin-Neto 2010)
A evoluccedilatildeo cliacutenica da doenccedila de Chagas pode ser dividida em duas fases aguda e
crocircnica A fase aguda normalmente eacute assintomaacutetica ou apresenta sintomatologia discreta que
na maioria dos casos passa despercebida pelo paciente Os sintomas quando aparecem o
fazem uma ou duas semanas apoacutes a exposiccedilatildeo ao triatomiacuteneo infectado ou poucos meses apoacutes
a transfusatildeo com sangue contaminado (Rassi Marin-Neto 2010) Os sintomas mais
frequentemente apresentados pelos pacientes agudos satildeo febre prolongada mal-estar
17
distensatildeo do fiacutegado baccedilo e linfonodos Pode haver tambeacutem edema subcutacircneo localizado ou
generalizado No caso particular de transmissatildeo vetorial o paciente pode apresentar um sinal
da porta de entrada do T cruzi na pele (chagoma) ou via mucosa ocular (sinal de Romantildea) As
manifestaccedilotildees da fase aguda se resolvem espontaneamente em aproximadamente 90 dos
indiviacuteduos infectados mesmo sem o tratamento com drogas tripanocidas Essa fase eacute
caracterizada pela alta parasitemia e ausecircncia de anticorpos especiacuteficos Nessa fase os
parasitas podem ser encontrados em praticamente todos os tecidos estando majoritariamente
presentes no sangue Por ser uma fase de curta duraccedilatildeo sem sintomatologia aparente ou com
sintomatologia pouco definida haacute um pequeno nuacutemero de casos detectados sendo portanto a
fase menos estudada (Rassi Marin-Neto 2010 Silber et al 2005b)
Logo apoacutes a fase aguda tem iniacutecio a fase crocircnica caracterizada pelos altos tiacutetulos de
anticorpos no sangue do paciente e pelo fato de que a parasitemia eacute inaparente O paciente
pode permanecer por vaacuterios anos ou ateacute mesmo pelo resto da vida sem apresentar sinais ou
sintomas Essa forma da fase crocircnica eacute chamada de indeterminada Aproximadamente 30-40
dos pacientes crocircnicos indeterminados desenvolvem a sintomatologia caracteriacutestica da
doenccedila evoluindo para a chamada fase crocircnica sintomaacutetica Nesta fase as trecircs principais
formas satildeo a cardiacuteaca a digestiva e a cardiodigestiva (Acquatella 2007 Marin-Neto et al
1999) Na forma cardiacuteaca o coraccedilatildeo eacute um dos primeiros oacutergatildeos afetados natildeo necessariamente
o uacutenico desenvolvendo hipertrofia e dilataccedilatildeo Na forma digestiva o trato digestivo apresenta
alteraccedilotildees anatocircmicas e fisioloacutegicas importantes sendo a principal altereccedilatildeo a dilataccedilatildeo do
intestino e do esocircfago que podem apresentar aspecto de megaviacutescera (Marin-Neto et al
1999 Rassi et al 2000) revisado por (Boscardin et al 2009 Rassi Marin-Neto 2010)
18
12 Trypanosoma cruzi
Trypanosoma cruzi eacute um parasita intracelular obrigatoacuterio classicamente localizado na
ordem kinetoplastida gecircnero Trypanosoma que causa uma infecccedilatildeo crocircnica em humanos e
em grande nuacutemero de outras espeacutecies de mamiacuteferos (Hoare 1972) Recentemente devido agrave
alta variabilidade bioloacutegica bioquiacutemica e molecular a espeacutecie Trypanosoma cruzi foi
dividida em seis linhagens subsespeciacuteficas (Discrete Typing Units ndash DTUs) numerados de I a
VI Esta classificaccedilatildeo foi realizada com base em caracteriacutesticas geneacuteticas moleculares e
imunoloacutegicas com o objetivo de unificar a nomenclatura e facilitar a comunicaccedilatildeo dentro da
comunidade cientiacutefica (Zingales et al 2009)
O T cruzi tem um ciclo de vida complexo que alterna entre o inseto vetor e o
hospedeiro mamiacutefero O parasita apresenta diferentes formas correspondentes a diferentes
estaacutegios do seu ciclo de vida Estes diferentes estaacutegios tecircm sido descritos com base em
criteacuterios morfoloacutegicos como a forma fusiforme ou arredondados a regiatildeo onde emerge o
flagelo e a posiccedilatildeo do cinetoplasto (estrutura contendo o DNA da uacutenica mitococircndria presente
no parasita) em relaccedilatildeo ao nuacutecleo No hospedeiro mamiacutefero foram descritas as formas
amastigota epimastigota intracelular e tripomastigota sanguiacutecola A forma amastigota eacute um
estaacutegio replicativo e infectivo Satildeo ceacutelulas arredondadas medindo de 2-6 microm de diacircmetro
encontradas no citoplasma das ceacutelulas infectadas Os epimastigotas intracelulares
(epimastigote-like) satildeo formas replicativas intermediaacuterias entre amastigotas e tripomastigotas
tambeacutem encontrados dentro das ceacutelulas medindo aproximadamente 54 microm Essas formas satildeo
menores que as formas epimastigotas encontradas no inseto vetor poreacutem compartilha com as
mesmas padrotildees morfomeacutetricos e diversas caracteriacutesticas bioquiacutemicas e bioloacutegicas (Almeida-
De-Faria et al 1999) O tripomastigota sanguiacutecola eacute um estaacutegio flagelado infectivo natildeo
replicativo encontrado principalmente na corrente sanguiacutenea do hospedeiro mamiacutefero Mede
aproximadamente 18 microm de comprimento incluindo um extremo livre do flagelo de
aproximadamente 6 microm e 2-3 microm de largura
No inseto vetor encontram-se predominantemente as formas epimastigota e
tripomastigota metaciacuteclico Epimastigota eacute uma forma flagelada replicativa e natildeo infectiva
Mede aproximadamente 20-40 microm de comprimento e 2-5 microm de largura essas formas estatildeo
presentes tipicamente na porccedilatildeo meacutedia do intestino do hospedeiro invertebrado
Tripomastigota metaciacuteclico eacute uma forma infectiva natildeo replicativa encontrada na porccedilatildeo final
do intestino do inseto vetor (Brener 1971) revisado por (Silber et al 2005a Tyler Engman
2001)
19
O ciclo de vida do T cruzi inicia-se quando os parasitas presentes na corrente
sanguiacutenea do hospedeiro vertebrado infectado passam ao inseto vetor durante o repasto
sanguiacuteneo No intestino do inseto estas formas se diferenciam em formas epimastigotas (De
Souza et al 2002) que apoacutes multiplicaccedilatildeo por fissatildeo binaacuteria colonizam a porccedilatildeo terminal do
intestino e se diferenciam novamente em tripomastigotas metaciacuteclicos Quando o inseto
infectado pica o hospedeiro vertebrado e inicia o repasto sanguiacuteneo concomitantemente
elimina as formas tripomastigotas metaciacuteclicos junto com as fezes e urina Uma vez em
contato com a pele ou mucosas do hospedeiro mamiacutefero os tripomastigotas satildeo internalizados
atraveacutes de pequenas lesotildees na pele causadas pelo proacuteprio hospedeiro (Alves Colli 2008) Os
tripomastigotas invadem as ceacutelulas do hospedeiro mamiacutefero e se diferenciam em formas
replicativas denominadas amastigotas estabelecendo assim a infecccedilatildeo Apoacutes
aproximadamente 96 horas os amastigotas comeccedilam a se diferenciar em tripomastigotas
passando pela forma intermediaacuteria epimastigota intracelular Estas formas tripomastigotas satildeo
liberadas na corrente sanguiacutenea podendo invadir novas ceacutelulas ou serem ingeridas pelo inseto
vetor em um novo repasto sanguiacuteneo fechando assim o seu ciclo de vida O esquema do ciclo
de vida do Trypanosoma cruzi estaacute representado na Figura 1
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi
20
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos
T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono As formas epimastigotas preferencialmente catabolizam glicose e apoacutes a exaustatildeo
do carboidrato ocorre uma mudanccedila para o metabolismo de aminoaacutecidos A prolina e a
glutamina satildeo os principais aminoaacutecidos metabolizados pelas formas epimastigotas visto que
satildeo abudantes na hemolinfa e nos fluiacutedos dos tecidos do inseto vetor Esse metabolismo
ocorre com produccedilatildeo de NH4+
(produto da deaminaccedilatildeo dos aminoaacutecidos) que eacute excretada para
o meio extracelular (Bringaud et al 2006 Cazzulo 1994) As formas tripomastigotas
utilizam principalmente a glicose que eacute abundante nos fluiacutedos de seus hospedeiros
vertebrados Jaacute as formas amastigotas encontradas no citoplasma das ceacutelulas do hospedeiro
mamiacutefero ambiente pobre em glicose livre usam os aminoaacutecidos como principal fonte de
carbono e energia apresentando maior quantidade de prolina intracelular do que as formas
tripomastigotas (Silber et al 2009)
Entre os aminoaacutecidos metabolizados pelas formas epimastigotas estatildeo prolina glicina
asparagina glutamina aspartato glutamato leucina e isoleucina (Mancilla et al 1967 Odaly
et al 1983 Silber et al 2005a Sylvester Krassner 1976 Zeledon 1960) Os aminoaacutecidos
estatildeo envolvidos tambeacutem em outros processos importantes do ciclo de vida do T cruzi O
aminoaacutecido L-prolina eacute essencial na diferenciaccedilatildeo das formas epimastigota intracelular para
tripomastigotas aleacutem de estarem envolvidos nos processos de osmorregulaccedilatildeo crescimento e
resistecircncia do parasita ao estresse oxidativo e nutricional (Homsy et al 1989 Magdaleno et
al 2009 Magdaleno et al 2011 Pereira et al 2002 Pereira et al 2003 Tonelli et al
2004) Glutamato prolina e aspartato satildeo importantes na metaciclogecircnese ou seja na
diferenciaccedilatildeo de formas epimastigotas para tripomastigotas metaciacuteclicos (Contreras et al
1985) Prolina glicina alanina e glutamato satildeo descritos como importantes nos processos de
osmorregulaccedilatildeo e controle do volume celular (Rohloff et al 2003 Rohloff et al 2004a)
Os aminoaacutecidos para que sejam metabolizados ou cumprirem outras funccedilotildees devem
ser obtidos a partir da degradaccedilatildeo de proteiacutenas sintetizados ou incorporados pelo parasita
atraveacutes de transportadores localizados na membrana (Silber et al 2005a) O primeiro
transportador de aminoaacutecidos em T cruzi foi descrito bioquimicamente em 1970 o
transportador de lisina (Hampton 1970) Outros aminoaacutecidos tambeacutem jaacute tiveram a sua
cineacutetica e seu mecanismo de transporte descrito para T cruzi A incorporaccedilatildeo de arginina
pelo parasita apresenta pelo menos dois sistemas de transporte sendo um de alta afinidade e
outro de baixa afinidade Esse transporte eacute regulado ao longo do ciclo de vida do parasita
21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

17
distensatildeo do fiacutegado baccedilo e linfonodos Pode haver tambeacutem edema subcutacircneo localizado ou
generalizado No caso particular de transmissatildeo vetorial o paciente pode apresentar um sinal
da porta de entrada do T cruzi na pele (chagoma) ou via mucosa ocular (sinal de Romantildea) As
manifestaccedilotildees da fase aguda se resolvem espontaneamente em aproximadamente 90 dos
indiviacuteduos infectados mesmo sem o tratamento com drogas tripanocidas Essa fase eacute
caracterizada pela alta parasitemia e ausecircncia de anticorpos especiacuteficos Nessa fase os
parasitas podem ser encontrados em praticamente todos os tecidos estando majoritariamente
presentes no sangue Por ser uma fase de curta duraccedilatildeo sem sintomatologia aparente ou com
sintomatologia pouco definida haacute um pequeno nuacutemero de casos detectados sendo portanto a
fase menos estudada (Rassi Marin-Neto 2010 Silber et al 2005b)
Logo apoacutes a fase aguda tem iniacutecio a fase crocircnica caracterizada pelos altos tiacutetulos de
anticorpos no sangue do paciente e pelo fato de que a parasitemia eacute inaparente O paciente
pode permanecer por vaacuterios anos ou ateacute mesmo pelo resto da vida sem apresentar sinais ou
sintomas Essa forma da fase crocircnica eacute chamada de indeterminada Aproximadamente 30-40
dos pacientes crocircnicos indeterminados desenvolvem a sintomatologia caracteriacutestica da
doenccedila evoluindo para a chamada fase crocircnica sintomaacutetica Nesta fase as trecircs principais
formas satildeo a cardiacuteaca a digestiva e a cardiodigestiva (Acquatella 2007 Marin-Neto et al
1999) Na forma cardiacuteaca o coraccedilatildeo eacute um dos primeiros oacutergatildeos afetados natildeo necessariamente
o uacutenico desenvolvendo hipertrofia e dilataccedilatildeo Na forma digestiva o trato digestivo apresenta
alteraccedilotildees anatocircmicas e fisioloacutegicas importantes sendo a principal altereccedilatildeo a dilataccedilatildeo do
intestino e do esocircfago que podem apresentar aspecto de megaviacutescera (Marin-Neto et al
1999 Rassi et al 2000) revisado por (Boscardin et al 2009 Rassi Marin-Neto 2010)
18
12 Trypanosoma cruzi
Trypanosoma cruzi eacute um parasita intracelular obrigatoacuterio classicamente localizado na
ordem kinetoplastida gecircnero Trypanosoma que causa uma infecccedilatildeo crocircnica em humanos e
em grande nuacutemero de outras espeacutecies de mamiacuteferos (Hoare 1972) Recentemente devido agrave
alta variabilidade bioloacutegica bioquiacutemica e molecular a espeacutecie Trypanosoma cruzi foi
dividida em seis linhagens subsespeciacuteficas (Discrete Typing Units ndash DTUs) numerados de I a
VI Esta classificaccedilatildeo foi realizada com base em caracteriacutesticas geneacuteticas moleculares e
imunoloacutegicas com o objetivo de unificar a nomenclatura e facilitar a comunicaccedilatildeo dentro da
comunidade cientiacutefica (Zingales et al 2009)
O T cruzi tem um ciclo de vida complexo que alterna entre o inseto vetor e o
hospedeiro mamiacutefero O parasita apresenta diferentes formas correspondentes a diferentes
estaacutegios do seu ciclo de vida Estes diferentes estaacutegios tecircm sido descritos com base em
criteacuterios morfoloacutegicos como a forma fusiforme ou arredondados a regiatildeo onde emerge o
flagelo e a posiccedilatildeo do cinetoplasto (estrutura contendo o DNA da uacutenica mitococircndria presente
no parasita) em relaccedilatildeo ao nuacutecleo No hospedeiro mamiacutefero foram descritas as formas
amastigota epimastigota intracelular e tripomastigota sanguiacutecola A forma amastigota eacute um
estaacutegio replicativo e infectivo Satildeo ceacutelulas arredondadas medindo de 2-6 microm de diacircmetro
encontradas no citoplasma das ceacutelulas infectadas Os epimastigotas intracelulares
(epimastigote-like) satildeo formas replicativas intermediaacuterias entre amastigotas e tripomastigotas
tambeacutem encontrados dentro das ceacutelulas medindo aproximadamente 54 microm Essas formas satildeo
menores que as formas epimastigotas encontradas no inseto vetor poreacutem compartilha com as
mesmas padrotildees morfomeacutetricos e diversas caracteriacutesticas bioquiacutemicas e bioloacutegicas (Almeida-
De-Faria et al 1999) O tripomastigota sanguiacutecola eacute um estaacutegio flagelado infectivo natildeo
replicativo encontrado principalmente na corrente sanguiacutenea do hospedeiro mamiacutefero Mede
aproximadamente 18 microm de comprimento incluindo um extremo livre do flagelo de
aproximadamente 6 microm e 2-3 microm de largura
No inseto vetor encontram-se predominantemente as formas epimastigota e
tripomastigota metaciacuteclico Epimastigota eacute uma forma flagelada replicativa e natildeo infectiva
Mede aproximadamente 20-40 microm de comprimento e 2-5 microm de largura essas formas estatildeo
presentes tipicamente na porccedilatildeo meacutedia do intestino do hospedeiro invertebrado
Tripomastigota metaciacuteclico eacute uma forma infectiva natildeo replicativa encontrada na porccedilatildeo final
do intestino do inseto vetor (Brener 1971) revisado por (Silber et al 2005a Tyler Engman
2001)
19
O ciclo de vida do T cruzi inicia-se quando os parasitas presentes na corrente
sanguiacutenea do hospedeiro vertebrado infectado passam ao inseto vetor durante o repasto
sanguiacuteneo No intestino do inseto estas formas se diferenciam em formas epimastigotas (De
Souza et al 2002) que apoacutes multiplicaccedilatildeo por fissatildeo binaacuteria colonizam a porccedilatildeo terminal do
intestino e se diferenciam novamente em tripomastigotas metaciacuteclicos Quando o inseto
infectado pica o hospedeiro vertebrado e inicia o repasto sanguiacuteneo concomitantemente
elimina as formas tripomastigotas metaciacuteclicos junto com as fezes e urina Uma vez em
contato com a pele ou mucosas do hospedeiro mamiacutefero os tripomastigotas satildeo internalizados
atraveacutes de pequenas lesotildees na pele causadas pelo proacuteprio hospedeiro (Alves Colli 2008) Os
tripomastigotas invadem as ceacutelulas do hospedeiro mamiacutefero e se diferenciam em formas
replicativas denominadas amastigotas estabelecendo assim a infecccedilatildeo Apoacutes
aproximadamente 96 horas os amastigotas comeccedilam a se diferenciar em tripomastigotas
passando pela forma intermediaacuteria epimastigota intracelular Estas formas tripomastigotas satildeo
liberadas na corrente sanguiacutenea podendo invadir novas ceacutelulas ou serem ingeridas pelo inseto
vetor em um novo repasto sanguiacuteneo fechando assim o seu ciclo de vida O esquema do ciclo
de vida do Trypanosoma cruzi estaacute representado na Figura 1
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi
20
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos
T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono As formas epimastigotas preferencialmente catabolizam glicose e apoacutes a exaustatildeo
do carboidrato ocorre uma mudanccedila para o metabolismo de aminoaacutecidos A prolina e a
glutamina satildeo os principais aminoaacutecidos metabolizados pelas formas epimastigotas visto que
satildeo abudantes na hemolinfa e nos fluiacutedos dos tecidos do inseto vetor Esse metabolismo
ocorre com produccedilatildeo de NH4+
(produto da deaminaccedilatildeo dos aminoaacutecidos) que eacute excretada para
o meio extracelular (Bringaud et al 2006 Cazzulo 1994) As formas tripomastigotas
utilizam principalmente a glicose que eacute abundante nos fluiacutedos de seus hospedeiros
vertebrados Jaacute as formas amastigotas encontradas no citoplasma das ceacutelulas do hospedeiro
mamiacutefero ambiente pobre em glicose livre usam os aminoaacutecidos como principal fonte de
carbono e energia apresentando maior quantidade de prolina intracelular do que as formas
tripomastigotas (Silber et al 2009)
Entre os aminoaacutecidos metabolizados pelas formas epimastigotas estatildeo prolina glicina
asparagina glutamina aspartato glutamato leucina e isoleucina (Mancilla et al 1967 Odaly
et al 1983 Silber et al 2005a Sylvester Krassner 1976 Zeledon 1960) Os aminoaacutecidos
estatildeo envolvidos tambeacutem em outros processos importantes do ciclo de vida do T cruzi O
aminoaacutecido L-prolina eacute essencial na diferenciaccedilatildeo das formas epimastigota intracelular para
tripomastigotas aleacutem de estarem envolvidos nos processos de osmorregulaccedilatildeo crescimento e
resistecircncia do parasita ao estresse oxidativo e nutricional (Homsy et al 1989 Magdaleno et
al 2009 Magdaleno et al 2011 Pereira et al 2002 Pereira et al 2003 Tonelli et al
2004) Glutamato prolina e aspartato satildeo importantes na metaciclogecircnese ou seja na
diferenciaccedilatildeo de formas epimastigotas para tripomastigotas metaciacuteclicos (Contreras et al
1985) Prolina glicina alanina e glutamato satildeo descritos como importantes nos processos de
osmorregulaccedilatildeo e controle do volume celular (Rohloff et al 2003 Rohloff et al 2004a)
Os aminoaacutecidos para que sejam metabolizados ou cumprirem outras funccedilotildees devem
ser obtidos a partir da degradaccedilatildeo de proteiacutenas sintetizados ou incorporados pelo parasita
atraveacutes de transportadores localizados na membrana (Silber et al 2005a) O primeiro
transportador de aminoaacutecidos em T cruzi foi descrito bioquimicamente em 1970 o
transportador de lisina (Hampton 1970) Outros aminoaacutecidos tambeacutem jaacute tiveram a sua
cineacutetica e seu mecanismo de transporte descrito para T cruzi A incorporaccedilatildeo de arginina
pelo parasita apresenta pelo menos dois sistemas de transporte sendo um de alta afinidade e
outro de baixa afinidade Esse transporte eacute regulado ao longo do ciclo de vida do parasita
21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

18
12 Trypanosoma cruzi
Trypanosoma cruzi eacute um parasita intracelular obrigatoacuterio classicamente localizado na
ordem kinetoplastida gecircnero Trypanosoma que causa uma infecccedilatildeo crocircnica em humanos e
em grande nuacutemero de outras espeacutecies de mamiacuteferos (Hoare 1972) Recentemente devido agrave
alta variabilidade bioloacutegica bioquiacutemica e molecular a espeacutecie Trypanosoma cruzi foi
dividida em seis linhagens subsespeciacuteficas (Discrete Typing Units ndash DTUs) numerados de I a
VI Esta classificaccedilatildeo foi realizada com base em caracteriacutesticas geneacuteticas moleculares e
imunoloacutegicas com o objetivo de unificar a nomenclatura e facilitar a comunicaccedilatildeo dentro da
comunidade cientiacutefica (Zingales et al 2009)
O T cruzi tem um ciclo de vida complexo que alterna entre o inseto vetor e o
hospedeiro mamiacutefero O parasita apresenta diferentes formas correspondentes a diferentes
estaacutegios do seu ciclo de vida Estes diferentes estaacutegios tecircm sido descritos com base em
criteacuterios morfoloacutegicos como a forma fusiforme ou arredondados a regiatildeo onde emerge o
flagelo e a posiccedilatildeo do cinetoplasto (estrutura contendo o DNA da uacutenica mitococircndria presente
no parasita) em relaccedilatildeo ao nuacutecleo No hospedeiro mamiacutefero foram descritas as formas
amastigota epimastigota intracelular e tripomastigota sanguiacutecola A forma amastigota eacute um
estaacutegio replicativo e infectivo Satildeo ceacutelulas arredondadas medindo de 2-6 microm de diacircmetro
encontradas no citoplasma das ceacutelulas infectadas Os epimastigotas intracelulares
(epimastigote-like) satildeo formas replicativas intermediaacuterias entre amastigotas e tripomastigotas
tambeacutem encontrados dentro das ceacutelulas medindo aproximadamente 54 microm Essas formas satildeo
menores que as formas epimastigotas encontradas no inseto vetor poreacutem compartilha com as
mesmas padrotildees morfomeacutetricos e diversas caracteriacutesticas bioquiacutemicas e bioloacutegicas (Almeida-
De-Faria et al 1999) O tripomastigota sanguiacutecola eacute um estaacutegio flagelado infectivo natildeo
replicativo encontrado principalmente na corrente sanguiacutenea do hospedeiro mamiacutefero Mede
aproximadamente 18 microm de comprimento incluindo um extremo livre do flagelo de
aproximadamente 6 microm e 2-3 microm de largura
No inseto vetor encontram-se predominantemente as formas epimastigota e
tripomastigota metaciacuteclico Epimastigota eacute uma forma flagelada replicativa e natildeo infectiva
Mede aproximadamente 20-40 microm de comprimento e 2-5 microm de largura essas formas estatildeo
presentes tipicamente na porccedilatildeo meacutedia do intestino do hospedeiro invertebrado
Tripomastigota metaciacuteclico eacute uma forma infectiva natildeo replicativa encontrada na porccedilatildeo final
do intestino do inseto vetor (Brener 1971) revisado por (Silber et al 2005a Tyler Engman
2001)
19
O ciclo de vida do T cruzi inicia-se quando os parasitas presentes na corrente
sanguiacutenea do hospedeiro vertebrado infectado passam ao inseto vetor durante o repasto
sanguiacuteneo No intestino do inseto estas formas se diferenciam em formas epimastigotas (De
Souza et al 2002) que apoacutes multiplicaccedilatildeo por fissatildeo binaacuteria colonizam a porccedilatildeo terminal do
intestino e se diferenciam novamente em tripomastigotas metaciacuteclicos Quando o inseto
infectado pica o hospedeiro vertebrado e inicia o repasto sanguiacuteneo concomitantemente
elimina as formas tripomastigotas metaciacuteclicos junto com as fezes e urina Uma vez em
contato com a pele ou mucosas do hospedeiro mamiacutefero os tripomastigotas satildeo internalizados
atraveacutes de pequenas lesotildees na pele causadas pelo proacuteprio hospedeiro (Alves Colli 2008) Os
tripomastigotas invadem as ceacutelulas do hospedeiro mamiacutefero e se diferenciam em formas
replicativas denominadas amastigotas estabelecendo assim a infecccedilatildeo Apoacutes
aproximadamente 96 horas os amastigotas comeccedilam a se diferenciar em tripomastigotas
passando pela forma intermediaacuteria epimastigota intracelular Estas formas tripomastigotas satildeo
liberadas na corrente sanguiacutenea podendo invadir novas ceacutelulas ou serem ingeridas pelo inseto
vetor em um novo repasto sanguiacuteneo fechando assim o seu ciclo de vida O esquema do ciclo
de vida do Trypanosoma cruzi estaacute representado na Figura 1
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi
20
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos
T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono As formas epimastigotas preferencialmente catabolizam glicose e apoacutes a exaustatildeo
do carboidrato ocorre uma mudanccedila para o metabolismo de aminoaacutecidos A prolina e a
glutamina satildeo os principais aminoaacutecidos metabolizados pelas formas epimastigotas visto que
satildeo abudantes na hemolinfa e nos fluiacutedos dos tecidos do inseto vetor Esse metabolismo
ocorre com produccedilatildeo de NH4+
(produto da deaminaccedilatildeo dos aminoaacutecidos) que eacute excretada para
o meio extracelular (Bringaud et al 2006 Cazzulo 1994) As formas tripomastigotas
utilizam principalmente a glicose que eacute abundante nos fluiacutedos de seus hospedeiros
vertebrados Jaacute as formas amastigotas encontradas no citoplasma das ceacutelulas do hospedeiro
mamiacutefero ambiente pobre em glicose livre usam os aminoaacutecidos como principal fonte de
carbono e energia apresentando maior quantidade de prolina intracelular do que as formas
tripomastigotas (Silber et al 2009)
Entre os aminoaacutecidos metabolizados pelas formas epimastigotas estatildeo prolina glicina
asparagina glutamina aspartato glutamato leucina e isoleucina (Mancilla et al 1967 Odaly
et al 1983 Silber et al 2005a Sylvester Krassner 1976 Zeledon 1960) Os aminoaacutecidos
estatildeo envolvidos tambeacutem em outros processos importantes do ciclo de vida do T cruzi O
aminoaacutecido L-prolina eacute essencial na diferenciaccedilatildeo das formas epimastigota intracelular para
tripomastigotas aleacutem de estarem envolvidos nos processos de osmorregulaccedilatildeo crescimento e
resistecircncia do parasita ao estresse oxidativo e nutricional (Homsy et al 1989 Magdaleno et
al 2009 Magdaleno et al 2011 Pereira et al 2002 Pereira et al 2003 Tonelli et al
2004) Glutamato prolina e aspartato satildeo importantes na metaciclogecircnese ou seja na
diferenciaccedilatildeo de formas epimastigotas para tripomastigotas metaciacuteclicos (Contreras et al
1985) Prolina glicina alanina e glutamato satildeo descritos como importantes nos processos de
osmorregulaccedilatildeo e controle do volume celular (Rohloff et al 2003 Rohloff et al 2004a)
Os aminoaacutecidos para que sejam metabolizados ou cumprirem outras funccedilotildees devem
ser obtidos a partir da degradaccedilatildeo de proteiacutenas sintetizados ou incorporados pelo parasita
atraveacutes de transportadores localizados na membrana (Silber et al 2005a) O primeiro
transportador de aminoaacutecidos em T cruzi foi descrito bioquimicamente em 1970 o
transportador de lisina (Hampton 1970) Outros aminoaacutecidos tambeacutem jaacute tiveram a sua
cineacutetica e seu mecanismo de transporte descrito para T cruzi A incorporaccedilatildeo de arginina
pelo parasita apresenta pelo menos dois sistemas de transporte sendo um de alta afinidade e
outro de baixa afinidade Esse transporte eacute regulado ao longo do ciclo de vida do parasita
21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

19
O ciclo de vida do T cruzi inicia-se quando os parasitas presentes na corrente
sanguiacutenea do hospedeiro vertebrado infectado passam ao inseto vetor durante o repasto
sanguiacuteneo No intestino do inseto estas formas se diferenciam em formas epimastigotas (De
Souza et al 2002) que apoacutes multiplicaccedilatildeo por fissatildeo binaacuteria colonizam a porccedilatildeo terminal do
intestino e se diferenciam novamente em tripomastigotas metaciacuteclicos Quando o inseto
infectado pica o hospedeiro vertebrado e inicia o repasto sanguiacuteneo concomitantemente
elimina as formas tripomastigotas metaciacuteclicos junto com as fezes e urina Uma vez em
contato com a pele ou mucosas do hospedeiro mamiacutefero os tripomastigotas satildeo internalizados
atraveacutes de pequenas lesotildees na pele causadas pelo proacuteprio hospedeiro (Alves Colli 2008) Os
tripomastigotas invadem as ceacutelulas do hospedeiro mamiacutefero e se diferenciam em formas
replicativas denominadas amastigotas estabelecendo assim a infecccedilatildeo Apoacutes
aproximadamente 96 horas os amastigotas comeccedilam a se diferenciar em tripomastigotas
passando pela forma intermediaacuteria epimastigota intracelular Estas formas tripomastigotas satildeo
liberadas na corrente sanguiacutenea podendo invadir novas ceacutelulas ou serem ingeridas pelo inseto
vetor em um novo repasto sanguiacuteneo fechando assim o seu ciclo de vida O esquema do ciclo
de vida do Trypanosoma cruzi estaacute representado na Figura 1
Figura 1 - Esquema do ciclo de vida do Trypanosoma cruzi
20
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos
T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono As formas epimastigotas preferencialmente catabolizam glicose e apoacutes a exaustatildeo
do carboidrato ocorre uma mudanccedila para o metabolismo de aminoaacutecidos A prolina e a
glutamina satildeo os principais aminoaacutecidos metabolizados pelas formas epimastigotas visto que
satildeo abudantes na hemolinfa e nos fluiacutedos dos tecidos do inseto vetor Esse metabolismo
ocorre com produccedilatildeo de NH4+
(produto da deaminaccedilatildeo dos aminoaacutecidos) que eacute excretada para
o meio extracelular (Bringaud et al 2006 Cazzulo 1994) As formas tripomastigotas
utilizam principalmente a glicose que eacute abundante nos fluiacutedos de seus hospedeiros
vertebrados Jaacute as formas amastigotas encontradas no citoplasma das ceacutelulas do hospedeiro
mamiacutefero ambiente pobre em glicose livre usam os aminoaacutecidos como principal fonte de
carbono e energia apresentando maior quantidade de prolina intracelular do que as formas
tripomastigotas (Silber et al 2009)
Entre os aminoaacutecidos metabolizados pelas formas epimastigotas estatildeo prolina glicina
asparagina glutamina aspartato glutamato leucina e isoleucina (Mancilla et al 1967 Odaly
et al 1983 Silber et al 2005a Sylvester Krassner 1976 Zeledon 1960) Os aminoaacutecidos
estatildeo envolvidos tambeacutem em outros processos importantes do ciclo de vida do T cruzi O
aminoaacutecido L-prolina eacute essencial na diferenciaccedilatildeo das formas epimastigota intracelular para
tripomastigotas aleacutem de estarem envolvidos nos processos de osmorregulaccedilatildeo crescimento e
resistecircncia do parasita ao estresse oxidativo e nutricional (Homsy et al 1989 Magdaleno et
al 2009 Magdaleno et al 2011 Pereira et al 2002 Pereira et al 2003 Tonelli et al
2004) Glutamato prolina e aspartato satildeo importantes na metaciclogecircnese ou seja na
diferenciaccedilatildeo de formas epimastigotas para tripomastigotas metaciacuteclicos (Contreras et al
1985) Prolina glicina alanina e glutamato satildeo descritos como importantes nos processos de
osmorregulaccedilatildeo e controle do volume celular (Rohloff et al 2003 Rohloff et al 2004a)
Os aminoaacutecidos para que sejam metabolizados ou cumprirem outras funccedilotildees devem
ser obtidos a partir da degradaccedilatildeo de proteiacutenas sintetizados ou incorporados pelo parasita
atraveacutes de transportadores localizados na membrana (Silber et al 2005a) O primeiro
transportador de aminoaacutecidos em T cruzi foi descrito bioquimicamente em 1970 o
transportador de lisina (Hampton 1970) Outros aminoaacutecidos tambeacutem jaacute tiveram a sua
cineacutetica e seu mecanismo de transporte descrito para T cruzi A incorporaccedilatildeo de arginina
pelo parasita apresenta pelo menos dois sistemas de transporte sendo um de alta afinidade e
outro de baixa afinidade Esse transporte eacute regulado ao longo do ciclo de vida do parasita
21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

20
13 Trypanosoma cruzi e o metabolismo de aminoaacutecidos
T cruzi eacute capaz de utilizar carboidratos e aminoaacutecidos como fonte de energia e
carbono As formas epimastigotas preferencialmente catabolizam glicose e apoacutes a exaustatildeo
do carboidrato ocorre uma mudanccedila para o metabolismo de aminoaacutecidos A prolina e a
glutamina satildeo os principais aminoaacutecidos metabolizados pelas formas epimastigotas visto que
satildeo abudantes na hemolinfa e nos fluiacutedos dos tecidos do inseto vetor Esse metabolismo
ocorre com produccedilatildeo de NH4+
(produto da deaminaccedilatildeo dos aminoaacutecidos) que eacute excretada para
o meio extracelular (Bringaud et al 2006 Cazzulo 1994) As formas tripomastigotas
utilizam principalmente a glicose que eacute abundante nos fluiacutedos de seus hospedeiros
vertebrados Jaacute as formas amastigotas encontradas no citoplasma das ceacutelulas do hospedeiro
mamiacutefero ambiente pobre em glicose livre usam os aminoaacutecidos como principal fonte de
carbono e energia apresentando maior quantidade de prolina intracelular do que as formas
tripomastigotas (Silber et al 2009)
Entre os aminoaacutecidos metabolizados pelas formas epimastigotas estatildeo prolina glicina
asparagina glutamina aspartato glutamato leucina e isoleucina (Mancilla et al 1967 Odaly
et al 1983 Silber et al 2005a Sylvester Krassner 1976 Zeledon 1960) Os aminoaacutecidos
estatildeo envolvidos tambeacutem em outros processos importantes do ciclo de vida do T cruzi O
aminoaacutecido L-prolina eacute essencial na diferenciaccedilatildeo das formas epimastigota intracelular para
tripomastigotas aleacutem de estarem envolvidos nos processos de osmorregulaccedilatildeo crescimento e
resistecircncia do parasita ao estresse oxidativo e nutricional (Homsy et al 1989 Magdaleno et
al 2009 Magdaleno et al 2011 Pereira et al 2002 Pereira et al 2003 Tonelli et al
2004) Glutamato prolina e aspartato satildeo importantes na metaciclogecircnese ou seja na
diferenciaccedilatildeo de formas epimastigotas para tripomastigotas metaciacuteclicos (Contreras et al
1985) Prolina glicina alanina e glutamato satildeo descritos como importantes nos processos de
osmorregulaccedilatildeo e controle do volume celular (Rohloff et al 2003 Rohloff et al 2004a)
Os aminoaacutecidos para que sejam metabolizados ou cumprirem outras funccedilotildees devem
ser obtidos a partir da degradaccedilatildeo de proteiacutenas sintetizados ou incorporados pelo parasita
atraveacutes de transportadores localizados na membrana (Silber et al 2005a) O primeiro
transportador de aminoaacutecidos em T cruzi foi descrito bioquimicamente em 1970 o
transportador de lisina (Hampton 1970) Outros aminoaacutecidos tambeacutem jaacute tiveram a sua
cineacutetica e seu mecanismo de transporte descrito para T cruzi A incorporaccedilatildeo de arginina
pelo parasita apresenta pelo menos dois sistemas de transporte sendo um de alta afinidade e
outro de baixa afinidade Esse transporte eacute regulado ao longo do ciclo de vida do parasita
21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

21
(Canepa et al 2004 Pereira et al 1999) A prolina tambeacutem eacute incorporada atraveacutes de dois
sistemas de transporte O sistema ldquoArdquo de alta afinidade eacute dependente do gradiente de H+ e o
sistema ldquoBrdquo de baixa afinidade eacute dependente de ATP (Silber et al 2002) Jaacute o transporte de
glutamato ocorre atraveacutes do gradiente de H+
na membrana do parasita mas natildeo eacute dependente
da concentraccedilatildeo de soacutedio ou potaacutessio (Silber et al 2006) Recentemente o nosso grupo
descreveu uma atividade de transporte do aacutecido gamma-aminobutiacuterico (GABA) em T cruzi
aminoaacutecido que aleacutem de ser um conhecido neurotransmissor eacute um anaacutelogo estrutural do
glutamato (Rojas 2007) A existecircncia de um transportador de GABA pode indicar a
participaccedilatildeo deste aminoaacutecido em processos bioloacutegicos e no metabolismo do T cruzi
A caracterizaccedilatildeo molecular e bioquiacutemica de vias metaboacutelicas e transportadores de
aminoaacutecidos em tripanossomatiacutedeos satildeo de grande importacircncia pois tem revelado uma
variedade de rotas que permitem a siacutentese de novo de compostos cruciais para a sua
sobrevivecircncia Ou tambeacutem a sua incorporaccedilatildeo e aproveitamento a partir de compostos
sintetizados pelos organismos hospedeiros O estudo de diferentes vias metaboacutelicas de
aminoaacutecidos tambeacutem permite obter informaccedilotildees relevantes sobre a importacircncia de cada um
desses metaboacutelitos para a biologia do parasita Desta forma tem se identificado enzimas e
transportadores que podem ser alvos interessantes para a busca de novas drogas para novos
tratamentos contra a doenccedila de Chagas
14 Morte celular em Trypanosoma cruzi
O termo morte celular programada (MCP) foi introduzido em 1964 por Lockshin e
Williams (Lockshin 1964) Esse termo descreve eventos que de forma orquestrada e bem
definida em etapas controladas e natildeo acidentalmente levam a morte celular (Duszenko et al
2006) Durante muito tempo acreditou-se que esse processo ocorria somente em organismos
multicelulares com o objetivo de evitar malformaccedilotildees na embriogecircnese evitar doenccedilas
autoimunes durante a diferenciaccedilatildeo linfocitaacuteria ou para manter a integridade dos tecidos e
evitar o cacircncer
O Comitecirc de Nomeclatura de Morte Celular (Nomenclature Comittee on Cell Death ndash
NCCD) define os diferentes tipos de morte celular baseados em seus mecanismos (Kroemer et
al 2005) Os trecircs principais tipos satildeo apoptose necrose e autofagia
- Apoptose eacute caracterizada pelo arredondamento da ceacutelula retraccedilatildeo de pseudoacutepodes
reduccedilatildeo do volume celular ativaccedilatildeo das caspases condensaccedilatildeo da cromatima fragmentaccedilatildeo
22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

22
do nuacutecleo manutenccedilatildeo da integridade da membrana celular ateacute os uacuteltimos estaacutegios do
processo
- Autofagia eacute o tipo de morte celular que ocorre sem condensaccedilatildeo da cromatina
acompanhada por vacuolizaccedilatildeo autofaacutegica do citoplasma Esses vacuacuteolos apresentam dupla
membrana e conteacutem organelas citoplasmaacuteticas em processo de degeneraccedilatildeo o que permite
que sejam destinguidas de outras vesiacuteculas como endossomos e lisossomos por microscopia
eletrocircnica
- Necrose eacute considerada um tipo de morte celular que natildeo apresenta sinais de
apoptose ou autofagia Caracteriza-se principalmente por ruptura da membrana celular e
dilataccedilatildeo de organelas citoplasmaacuteticas como mitococircndria retiacuteculo endoplasmaacutetico e
complexo de Golgi (Kroemer et al 2005)
Mais recentemente foram relatadas vaacuterias evidecircncias de MCP em eucariotos
unicelulares incluindo tripanossomatiacutedeos dos gecircneros Leishmania e Trypanosoma
(Debrabant et al 2003) Vaacuterias caracteriacutesticas de MCP encontradas em organismos
multicelulares tecircm sido relatadas para tripanossomatiacutedeos Estas caracteriacutesticas incluem
despolarizaccedilatildeo da membrana mitocondrial liberaccedilatildeo de citocromo C ativaccedilatildeo de proteases
exposiccedilatildeo de fosfatidilserina encolhimento ou arredondamento das ceacutelulas manutenccedilatildeo da
integridade da membrana e fragmentaccedilatildeo de DNA (Debrabant et al 2003 Duszenko et al
2006 Kaczanowski et al 2011) Tambeacutem foi observado que o uso de inibidores de caspases
bloqueia algumas caracteriacutesticas associadas com MCP em tripanossomatiacutedeos (Irigoin et al
2009)
Um estudo realizado por Ameisen e colaboradores demonstrou que formas
epimastigotas cultivadas por mais de uma semana no mesmo meio se diferenciavam para
metaciacuteclicos ou morriam apresentando caracteriacutesticas de apoptose (Ameisen et al 1995)
Muitas proteiacutenas descritas como participantes da apoptose em metazoaacuterios natildeo estatildeo presentes
em tripanossomatiacutedeos Haacute evidecircncias de que metacaspases um grupo de proteases com
estrutura similar agraves caspases mas com diferentes atividades enzimaacuteticas podem estar
envolvidas no processo de apoptose desses organismos (Smirlis Soteriadou 2011) Essas
proteiacutenas estatildeo envolvidas em fases importantes do ciclo de vida do parasita como progressatildeo
do ciclo celular proteccedilatildeo para que formas epimastigotas natildeo sofram morte celular e
diferenciem para tripomastigota metaciacuteclico (Laverriere et al 2012) O fato de que as
metacaspases natildeo estatildeo presentes em humanos e desempenham um importante papel em
tripanossomatiacutedeos faz com que apresentem perspectivas promissoras como potenciais alvos
terapecircuticos (Berg et al 2010) Alguns autores tecircm descrito a participaccedilatildeo da mitococircndria no
23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

23
processo de apoptose em T cruzi Como acontece em metazoaacuterios a mitococircndria eacute a principal
organela envolvida nesse processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando
ocorre a produccedilatildeo de espeacutecies reativas de oxigecircnio (EROs) foram identificadas como o
principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-
Barreto et al 2009 Smirlis Soteriadou 2011) A produccedilatildeo de EROs e a desregulaccedilatildeo da
homeostase de Ca2+
pode contribuir para a apoptose nessas ceacutelulas (Smirlis Soteriadou
2011) Os motivos pelos quais os tripanossomatiacutedeos sofrem MCP ainda natildeo estatildeo bem
esclarecidos mas alguns autores sugerem que apesar das ceacutelulas viverem de forma
independente competem por nutrientes e fatores de crescimento Este seria um mecanismo de
sobrevivecircncia relacionado com a regulaccedilatildeo da proliferaccedilatildeo celular aleacutem da manutenccedilatildeo de
uma infecccedilatildeo controlada e persistente afetando a resposta imunoloacutegica do hospedeiro Este
mecanismo tambeacutem poderia estar envolvido no controle da populaccedilatildeo de parasitas no
hospedeiro aumentando a sobrevivecircncia do mesmo e favorecendo a sua transmissatildeo para o
proacuteximo hospedeiro (Debrabant et al 2003 Duszenko et al 2006)
15 Terapia na doenccedila de Chagas
Nas uacuteltimas deacutecadas muitas medidas foram colocadas em praacutetica visando o controle da
transmissatildeo vetorial da doenccedila de Chagas Apesar de terem sido bem sucedidas os resultados
dessas medidas natildeo beneficiam diretamente os milhotildees de indiviacuteduos jaacute infectados tatildeo pouco
os pacientes chagaacutesicos com manifestaccedilotildees cliacutenicas da doenccedila (Coura et al 2002) Embora a
doenccedila tenha mais de um seacuteculo da sua descoberta a quimioterapia disponiacutevel atualmente
limita-se a dois compostos que comeccedilaram a ser utilizados em humanos na deacutecada de 1970
que satildeo Nifurtimox [3-methyl-4(5rsquo-nitrofurfurylideneamine) tetrahydro-4H-tiazine-11-
dioxide] fabricado pela Bayer com o nome comercial de Lampitreg
e o Benzonidazol (N-
benzyl-2-nitroimidazole acetamide) fabricado pela Roche com o nome comercial de
Rochaganreg
(Dias et al 2009 Senkovich et al 2005) As estruturas quiacutemicas desses
compostos estatildeo representadas na Figura 2
24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

24
Figura 2 ndash Estruturas quiacutemicas do Benzonidazol e Nifurtimox
(1) Benzonidazol e (2) Nifurtimox
A dose recomendada na fase aguda eacute de 8-10 mgkgdia de Nifurtimox ou 5 mgkgdia
de Bezonidazol A meacutedia do tempo de tratamento eacute de 60 dias mas quando haacute uma reativaccedilatildeo
da fase crocircnica em pacientes imunocomprometidos por exemplo o tratamento pode durar
cinco meses ou mais Em casos da possibilidade de infecccedilatildeo como no caso de transfusatildeo com
sangue supostamente contaminado ou acidentes em laboratoacuterios eacute feito um tratamento
profilaacutetico com duraccedilatildeo de dez dias (Maya et al 2007 Rassi Marin-Neto 2010) O
mecanismo de accedilatildeo de ambos os compostos ainda natildeo estaacute bem claro mas acredita-se que
ocorra por meio da formaccedilatildeo de radicais livres como tambeacutem pela formaccedilatildeo de metaboacutelitos
eletrofiacutelicos
Ao radical hidroxila (OH) resultante do metabolismo desses faacutermacos eacute atribuiacutedo o
efeito tripanocida por mecanismos complexos que envolvem ligaccedilatildeo com lipiacutedeos proteiacutenas e
ao DNA do T cruzi Os metaboacutelitos eletrofiacutelicos formados pelo mecanismo de accedilatildeo de ambos
os compostos podem atuar tambeacutem em outros sistemas especialmente do hospedeiro humano
devido a sua alta reatividade (Dias et al 2009 Docampo 1990 Maya et al 2007 Urbina
2002)
Ambas as drogas reduzem os sintomas da doenccedila e a mortalidade das pessoas
infectadas quando utilizadas na fase aguda (Cancado 1999) mas sua eficaacutecia varia de acordo
com a duraccedilatildeo do tratamento como tambeacutem com a idade e distribuiccedilatildeo geograacutefica dos
pacientes provavelmente devido agrave susceptibilidade das diferentes cepas de T cruzi ao
tratamento (Dias et al 2009 Filardi Brener 1987) Eacute importante ressaltar que vaacuterios estudos
sugerem o tratamento com Benzonidazol de pacientes com a forma crocircnica da doenccedila de
Chagas Poreacutem nesta fase (em que a maioria dos casos satildeo diagnosticados) a eficaacutecia deste
tratamento eacute controversa (Boscardin et al 2009 Silber et al 2005b) Aleacutem do mais
25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

25
apresentam vaacuterios efeitos colaterais como por exemplo anorexia nauacutesea vocircmito dor de
cabeccedila depressatildeo do sistema nervoso central parestesia dermatite entre outros sintomas e
requerem longo tempo de tratamento que pode durar ateacute 90 dias (Maya et al 2007 Urbina
Docampo 2003) Esse longo periacuteodo pode levar ao abandono do tratamento pelos pacientes o
que resulta na seleccedilatildeo (dentro da populaccedilatildeo tratada) de parasitas mais resistentes
O Benzonidazol o uacutenico tripanocida disponiacutevel no mercado brasileiro atualmente eacute
indicado nas seguintes situaccedilotildees infecccedilatildeo na fase aguda reativaccedilatildeo da infecccedilatildeo por drogas
imunossupressoras infecccedilotildees adquiridas recentemente e para pacientes com oacutergatildeos
transplantados procedentes de pacientes chagaacutesicos (Silber et al 2005b)
16 Perspectivas de novos faacutermacos
Eacute urgente a necessidade do desenvolvimento de drogas mais potentes e seguras para o
tratamento especiacutefico da doenccedila de Chagas (Urbina Docampo 2003) As pesquisas por novos
faacutermacos com atividade tripanocida tecircm evoluiacutedo sensivelmente nas uacuteltimas deacutecadas
principalmente apoacutes o sequenciamento dos genomas do T cruzi T brucei e Leishmania
(Moreira et al 2009) Mesmo assim ainda natildeo foi desenvolvido um faacutermaco seguro e eficaz
contra esses parasitas Atualmente alguns alvos macromoleculares com funccedilotildees bioloacutegicas
importantes para a viabilidade do T cruzi tecircm sido explorados para o desenvolvimento de
novos faacutermacos
Entre eles estatildeo
Via glicoliacutetica
As formas infectivas do parasita utilizam preferencialmente a glicose como fonte de
carbono e energia Esta via eacute importante para geraccedilatildeo de energia atraveacutes da produccedilatildeo de
ATP Trecircs enzimas dessa via tecircm sido bastante exploradas como alvos bioloacutegicos a
gliceraldeiacutedo-3-fosfato desidrogenase (GAPDH EC12112) a hexoquinase (EC 27111) e a
fosfofrutoquinase (EC 2711) (Dias et al 2009) Contudo haacute indiacutecios de que essa via natildeo
estaacute ativa no estaacutegio intracelular amastigota (Silber et al 2009) o que limitaria a utilizaccedilatildeo
desses alvos
26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

26
Metabolismo dependente de grupos tioacuteis
O T cruzi natildeo apresenta as vias claacutessicas descritas para defesa contra o estresse
oxidativo jaacute que nenhuma catalase ou glutationa peroxidase eacute encontrada no parasita e a
atividade da superoacutexido dismutase eacute reduzida (Maya et al 2003) Desta forma o principal
mecanismo de defesa envolve os tioacuteis tripanotiona homotripanotiona glutationa e ovotiol
Uma reduccedilatildeo nos niacuteveis dos tioacuteis antioxidantes tornaria o parasita altamente vulneraacutevel aos
efeitos de radicais livres As enzimas que se destacam na via de biossiacutentese de tioacuteis satildeo
tripanotiona redutase (EC 18112) e tripanotiona sintase (EC 3619) A primeira eacute
responsaacutevel pela manutenccedilatildeo dos niacuteveis de tripanotiona reduzida e a segunda pela adiccedilatildeo do
composto glutationa ao substrato glutationil espermidina formando tripanotiona A enzima
triparedoxina peroxidase tambeacutem apresenta importacircncia consideraacutevel na exposiccedilatildeo do T cruzi
agraves espeacutecies reativas de oxigecircnio (Dias et al 2009 Urbina 2002 Wilkinson 2003)
Biossiacutentese de esteroacuteis
Os esteroacuteis satildeo componentes essenciais agraves membranas das ceacutelulas Em organismos
unicelulares os esteroacuteis satildeo essenciais ao crescimento celular T cruzi natildeo sintetiza colesterol
sintezando portanto o ergosterol (Urbina et al 1995) o qual eacute importante na proliferaccedilatildeo e
viabilidade celular (Maya et al 2007) Essa via tem sido um alvo atrativo para o
desenvolvimento de faacutermacos Inibidores de biossiacutentese de esteroacuteis satildeo os uacutenicos que
alcanccedilaram triagens cliacutenicas avanccediladas como candidatos a faacutermacos anti- T cruzi (Urbina
2010) Dentre os inibidores mais representativos estatildeo os derivados azoacutelicos inibidores da
esterol 14-demitilase cetoconazol fluconazol e itraconazol aleacutem do tipifarnib que eacute um
inibidor da farnesiltransferase (Dias et al 2009)
Proteases
Em T cruzi as proteases possuem muacuteltiplas funccedilotildees que envolvem desde a invasatildeo
celular ateacute o escape do parasita do sistema imune do hospedeiro A cruzipaiacutena (gp5157) eacute
responsaacutevel pela principal atividade proteoliacutetica em todos os estaacutegios do ciclo de vida desse
parasita (Urbina 2002) Dada a sua importacircncia algumas proteases tecircm sido selecionadas
como alvos para o desenvolvimento de novos agentes antichagaacutesicos Serinoproteases
27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

27
metaloproteases cisteiacutenoproteases e treoninoproteases satildeo exemplos de proteases amplamente
estudadas (Duschak Couto 2007 Soeiro De Castro 2009)
Vias metaboacutelicas de aminoaacutecidos
O estudo das vias metaboacutelicas dos aminoaacutecidos em tripanosomatiacutedeos tambeacutem tem
revelado uma variedade de compostos cruciais para a sobrevivecircncia do T cruzi Assim
transportadores e enzimas relacionados ao seu metabolismo podem ser alvos interessantes
para o desenho de novos faacutermacos Jaacute foi descrito a importacircncia de muitos aminoaacutecidos no
ciclo de vida do T cruzi Como exemplo o glutamato estaacute envolvido em processos
importantes como metaciclogecircnese osmorregulaccedilatildeo e controle do volume celular (Contreras
et al 1985 Rohloff et al 2003 Rohloff et al 2004b)
O nosso grupo tem estudado o envolvimento do glutamato na resistecircncia do parasita
ao estresse teacutermico oxidativo e metaboacutelico (Magdaleno et al 2011) como tambeacutem iniciamos
estudos sobre a importacircncia do aminoaacutecido GABA no metabolismo do T cruzi visto que
este tambeacutem eacute transportado pelo parasita (Rojas 2007) Esses metaboacutelitos satildeo relatados na
literatura neurobioloacutegica como os principais neurotransmissores excitatoacuterio e inibitoacuterio
respectivamente em mamiacuteferos
Diante do exposto foi considerado relevante o estudo da possiacutevel atividade anti-T
cruzi de compostos com atuaccedilatildeo nas vias glutamateacutergica e GABAeacutergica do sistema nervoso
central jaacute disponiacuteveis para uso em seres humanos com o intuito de aproveitar o
conhecimento da sua farmacocineacutetica biodisponibilidade toxicidade efeitos secundaacuterios
dentre outros paracircmetros Existem no mercado alguns faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato e que satildeo utilizados amplamente para o tratamento
cliacutenico de vaacuterias patologias de acometimento do sistema nervoso central Para este estudo
foram selecionados inicialmente dois faacutermacos com accedilatildeo sobre a via GABAeacutergica cujas
estruturas quiacutemicas estatildeo representadas na Figura 3 e estatildeo descritos a seguir
28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

28
Figura 3 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo GABAeacutergica
(1) Vigabatrina e (2) Pregabalina
A Vigabatrina eacute um inibidor irreversiacutevel do receptor GABA Transaminase (GABA T)
Esta inibiccedilatildeo foi demonstrada em um estudo realizado in vitro com neurocircnios e ceacutelulas da glia
(Gran Borchgrevink 1989) A ligaccedilatildeo da Vigabatrina ao receptor eacute irreversiacutevel soacute uma nova
siacutentese da enzima pode restabelecer a atividade de GABA T podendo tambeacutem bloquear o
transporte de GABA (Jung et al 1977)
Pregabalina eacute um anaacutelogo de GABA com uma substituiccedilatildeo na posiccedilatildeo C-3 (Gajcy et
al 2010) atua ligando-se a subunidade α2-δ do canal de Ca2+
controlado por voltagem A
potente inibiccedilatildeo desse siacutetio reduz o influxo de Ca2+
para os terminais nervosos preacute-sinaacutepticos
reduzindo assim a liberaccedilatildeo de alguns neurotransmissores como glutamato e noradrenalina
(Ha et al 2008)
Tambeacutem foram selecionados trecircs faacutermacos que atuam via receptores de glutamato
cujas estruturas quiacutemicas estatildeo representadas na Figura 4 e estatildeo descritos a seguir
(1) (2)
29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

29
Figura 4 - Estruturas quiacutemicas dos faacutermacos com accedilatildeo glutamateacutergica
(1) MK-801 (2) (MTEP) e (3) Memantina
O MK-801 eacute um antagonista do receptor de glutamato N-metil-D-aspartato (NMDA)
(Liu et al 2009) Atua ligando-se no receptor impedindo o fluxo de iacuteons tais como Ca2+
(Coan et al 1987) O MTEP 3-((2-Metil-4-thiazolil) etinil)piridina atua como um
antagonista seletivo natildeo competitivo do receptor metabotroacutepico de glutamato mGluR5 (Iso et
al 2006) Uma caracterizaccedilatildeo in vivo e in vitro indicou que o MTEP eacute altamente seletivo
para mGluR5 e natildeo tem efeito significativo em outros subtipos mGluR Jaacute a Memantina eacute
uma amina triciacuteclica a primeira da classe de novos medicamentos para a doenccedila de
Alzheimer atuando no sistema glutamateacutergico com o bloqueio da atividade de receptores de
glutamato do tipo NMDA (Lipton 2005) Estes faacutermacos tanto os de accedilatildeo GABAeacutergica
quanto os de accedilatildeo glutamateacutergica jaacute satildeo utilizados em seres humanos Apresentam efeitos
terapecircuticos beneacuteficos em vaacuterias desordens neuroloacutegicas agudas e crocircnicas como
anticonvulsivantes e ateacute mesmo no tratamento da esquizofrenia Portanto satildeo faacutermacos jaacute
aprovados para uso em seres humanos com efeitos colaterais e toxicidade conhecidos como
tambeacutem a farmacocineacutetica e farmacodinacircmica
30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

30
2 OBJETIVOS
21 Objetivo geral
Analisar as possiacuteveis interferecircncias de faacutermacos que atuam via receptores ou
transportadores de GABA e glutamato no ciclo de vida do T cruzi
22 Objetivos especiacuteficos
Avaliar o efeito dos faacutermacos selecionados sobre as curvas de crescimento da forma
epimastigota e sobre a diferenciaccedilatildeo de formas epimastigotas para tripomastigotas
(metaciclogecircnese)
Avaliar a resistecircncia da forma epimastigota a diferentes tipos de estresse (metaboacutelico
teacutermico e oxidativo) na presenccedila dos faacutermacos em estudo
Comparar os efeitos inibitoacuterios desses faacutermacos na progressatildeo do ciclo de vida nos
diferentes estaacutegios parasitaacuterios com ecircnfase nos estaacutegios correspondentes ao ciclo de
infecccedilatildeo em mamiacuteferos (amastigotas epimastigotas intracelular e tripomastigotas)
31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

31
3 MATERIAIS E MEacuteTODOS
31 Microorganismos utilizados no trabalho
311 Ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary)
312 Trypanosoma cruzi
Cepa CL clone 14 (Brener Chiari 1963)
32 Manutenccedilatildeo das ceacutelulas hospedeiras
Ceacutelulas da linhagem CHO-K1 (Chinese Hamster Ovary) foram cultivadas a 37 degC em
meio RPMI 1640 (Roswell Park Memorial Institute) acrescido com 10 de SFB (Soro Fetal
Bovino) 015 (mv) de NaHCO3 sob atmosfera uacutemida a 5 de CO2 (Tonelli et al 2004)
33 Obtenccedilatildeo dos parasitas
331 Epimastigotas
As formas epimastigotas extracelulares foram mantidas em fase exponencial de
crescimento por passagens sucessivas (cada 48 horas) em meio LIT (Liver Infusion-Tryptose)
que conteacutem infusatildeo de fiacutegado 50 gL Triptose 50 gL NaCl 40 gL KCL 04 gL
Na2HPO4 80 gL glicose 2 gL hemina 10 gL suplementado com 10 de SBF pH 72 a
28 ordmC (Camargo 1964)
332 Curvas de crescimento de epimastigotas de T cruzi
Foram utilizadas formas epimastigotas de T cruzi em fase de crescimento exponencial
(50 a 60 x 107 ceacutelulas mL
-1) As ceacutelulas foram tratadas com diferentes concentraccedilotildees dos
compostos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo foi utilizado uma
combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) As ceacutelulas (25 x 106 ceacutelulas mL
-1)
32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

32
foram transferidas para placas de cultura de 96 poccedilos (200 microlpoccedilo) A placa foi mantida na
estufa a 28 degC A proliferaccedilatildeo celular foi estimada por leitura da absorbacircncia da densidade
oacutetica (DO) em λ 620 nm durante 8 dias A absorbacircncia foi transformada em valores de
densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que inibiu 50 da
proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de crescimento (quinto
dia) mediante ajuste dos dados na curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
(Magdaleno et al 2009) Cada composto foi avaliado em quadruplicata em cada experimento
sendo que os resultados apresentados correspondem a trecircs experimentos independentes
333 Formas tripomastigotas
As ceacutelulas CHO-K1 foram infectadas com 30 x 107 tripomastigotas por garrafa (75
cm2) em meio RPMI 1640 contendo 10 de SFB Os parasitas ficaram em contato com as
ceacutelulas durante 4 horas mantidas a 37 degC sob atmosfera uacutemida com 5 de CO2 como
descrito por (Tonelli et al 2004) Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com
PBS (Phosphate Buffered Saline) e acrescidas de meio RPMI 1640 contendo 10 de SFB e
incubadas a 37 degC Apoacutes 12 horas as ceacutelulas foram lavadas com PBS acrescidas de meio
RPMI 1640 (2 SFB) e incubadas a 33 degC Os parasitas foram coletados no quinto sexto e
seacutetimo dia poacutes-infecccedilatildeo contados na cacircmara de Neubauer e utilizados em experimentos
posteriores
34 Resistecircncia a diferentes tipos de estresse
341 Ensaio de estresse teacutermico em formas epimastigotas
Os ensaios foram realizados conforme descrito na seccedilatildeo 332 modificando-se apenas
a temperatura de incubaccedilatildeo As placas foram mantidas na estufa a 33 degC ou 37 degC
(Magdaleno et al 2009)
33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

33
342 Avaliaccedilatildeo da interaccedilatildeo dos faacutermacos com o estresse metaboacutelico
Para avaliar a interaccedilatildeo entre os faacutermacos e o estresse metaboacutelico epimastigotas (50 x
106 ceacutelulas mL
-1) foram lavadas duas vezes com PBS e ressuspendidos em meio LIT ou PBS
e tratados ou natildeo (controle negativo) com a concentraccedilatildeo correspondente ao seu IC50 As
ceacutelulas foram incubadas a 28 degC durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de
ATP intracelular
3421 Avaliaccedilatildeo dos niacuteveis de ATP intracelular
Para avaliar os niacuteveis de ATP intracelular em formas epimastigotas de T cruzi
utilizamos o kit bioluminescente para ceacutelulas somaacuteticas comprado da Sigma-Alderich
(Sigma-Alderich Saint Louis MO USA) conforme instruccedilotildees do fabricante Brevemente 50
microL de PBS foi adicionado a 100 microL do reagente de lise e acrescentou-se 50 microL da suspensatildeo
de parasitas contendo 50 x 106 ceacutelulas mL
-1 tratadas ou natildeo (controle) A concentraccedilatildeo de
ATP foi determinada utilizando-se a curva padratildeo das diferentes concentraccedilotildees de ATP A
luminescecircncia foi obtida pela reaccedilatildeo entre a luciferase e o ATP que foi liberado apoacutes a lise
celular sendo determinada pela utilizaccedilatildeo do luminocircmetro Lumat LB 9507 utilizando-se um
λ 570 nm (Martins et al 2009)
343 Ensaio de estresse oxidativo em formas epimastigotas
Inicialmente foi determinado o valor de IC50 para H2O2 nas formas epimastigotas de T
cruzi o qual foi utilizado posteriormente para os ensaios de estresse oxidativo Para isso as
ceacutelulas (50 x 106
ceacutelulas mL-1
) foram lavadas duas vezes com PBS e incubadas em PBS com
diferentes concentraccedilotildees de H2O2 (10 a 100 microM) durante 60 90 e 120 minutos a 28 degC Apoacutes
esse tempo as ceacutelulas foram centrifugadas ressuspensas em meio LIT e distribuiacutedas em placa
de 96 poccedilos A placa foi mantida na estufa a 28 degC A proliferaccedilatildeo celular foi estimada por
leitura da DO a λ 620 nm a cada 24 horas durante 6 dias A DO foi transformada em valores
de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo linear que foi obtida
previamente sob as mesmas condiccedilotildees A concentraccedilatildeo de H2O2 correspondente ao valor do
IC50 foi determinada na fase exponencial de crescimento (quinto dia) mediante ajuste dos
dados da curva dose-resposta com a equaccedilatildeo claacutessica sigmoacuteide
34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

34
Para avaliar uma possiacutevel interaccedilatildeo entre os faacutermacos e o estresse oxidativo os
parasitas foram lavados duas vezes com PBS ressuspensos em PBS com a concentraccedilatildeo de
H2O2 correspondente ao IC50 determinado anteriormente e incubados por 1 hora a 28 degC Em
seguida foram centrifugadas ressuspensas em meio LIT e tratadas com diferentes
concentraccedilotildees dos faacutermacos ou natildeo (controle negativo) Como controle positivo de inibiccedilatildeo
foi utilizado uma combinaccedilatildeo de Rotenona (60 microM) e Antimicina (05 microM) Os parasitas
foram incubados em placas de cultura de 96 poccedilos em estufa (28 degC) A proliferaccedilatildeo celular
foi estimada por leitura da DO em λ 620 nm a cada 24 horas durante 8 dias A DO foi
transformada em valores de densidade celular (ceacutelulasmL) usando uma equaccedilatildeo de regressatildeo
linear que foi obtida previamente sob as mesmas condiccedilotildees A concentraccedilatildeo do faacutermaco que
inibiu 50 da proliferaccedilatildeo dos parasitas (IC50) foi determinada na fase exponencial de
crescimento (quinto dia) mediante ajuste dos dados da curva dose-resposta com a equaccedilatildeo
claacutessica sigmoacuteide Cada concentraccedilatildeo foi avaliada em quadruplicata sendo que os resultados
apresentados correspondem a trecircs experimentos independentes
35 Avaliaccedilatildeo do tipo de morte celular em parasitas tratados com Memantina
351 Avaliaccedilatildeo da exposiccedilatildeo de fosfatidilserina na membrana do parasita
Formas epimastigota (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT a 28 degC e
tratadas com a concentraccedilatildeo correspondente ao IC50 de cada composto determinado nas
curvas de crescimento dos parasitas ou natildeo (controle) Parasitas tratados com estaurosporina
(1microM) 24 horas antes da realizaccedilatildeo do ensaio foram utilizados como controle positivo para
apoptose O controle positivo para necrose foi realizado incubando os parasitas durante 30
minutos na presenccedila de 150 microM de digitonina e 1 microgmL de iodeto de propiacutedio No quarto dia
apoacutes o iniacutecio do tratamento 10 x 106 ceacutelulas mL
-1 de cada tratamento como tambeacutem dos
controles foram lavados uma vez com o tampatildeo de anexina (10 mM HEPES 140 mM NaCl e
25 mM CaCl2 pH 74) e ressuspensos em 50 microl do mesmo tampatildeo Os parasitas foram
incubados em gelo e protegidos da luz por 15 minutos na presenccedila de anexina-V FITC
Invitrogen (Invitrogen Eugene Oregon USA) conforme instruccedilotildees do fabricante e iodeto de
propiacutedio (1microgmL) Apoacutes esse periacuteodo adicionou-se 450 microl do tampatildeo de anexina e os
parasitas foram analisados por citometria de fluxo no equipamento Guava cytometer
(General Electric)(Jimenez et al 2008)
35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

35
352 Ensaio para quantificaccedilatildeo de Ca2+
intracelular em formas epimastigotas
Parasitas foram cultivados em LIT tratados com a concentraccedilatildeo correspondente ao
IC50 de cada composto ou natildeo (controle) No quarto dia apoacutes o iniacutecio do tratamento 10 x 108
ceacutelulas foram incubadas na presenccedila de 5 microM de Fluo-4 AM (Invitrogen) por uma hora a 28
ordmC Apoacutes esse periacuteodo as ceacutelulas foram lavadas duas vezes com o tampatildeo Hepes -glicose (50
mM Hepes (pH 74) 116 mM NaCl 54 mM KCl 08 mM MgSO4 55 mM glicose e 2 mM
CaCl2) ressuspensas no mesmo tampatildeo e distribuiacutedas em placa de 96 poccedilos (25 x 107 por
poccedilo) em triplicata A leitura foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 490 nm e λ emissatildeo 518 nm (Dolai et al 2009)
353 Ensaio para verificaccedilatildeo da geraccedilatildeo de espeacutecies reativas de oxigecircnio
Para quantificar a produccedilatildeo de H2O2 pelos parasitas formas epimastigotas (50 x 106
ceacutelulas mL-1
) foram cultivadas em meio LIT a 28 degC e tratadas com a concentraccedilatildeo
correspondente ao IC50 de cada composto determinado na curva de crescimento do parasita
ou natildeo (controle) Apoacutes 24 horas do iniacutecio do tratamento 10 x 107 ceacutelulas mL
-1 tratadas ou
natildeo foram lavadas por centrifugaccedilatildeo e ressuspensas em PBS (5 mM de succinato)
Adicionou-se 12 microM do reagente Amplex Red e 005 UmL de proxidase (Horseradish
Peroxidase) A leitura das amostras foi realizada no fluoriacutemetro Spectra Max M3 Molecular
Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm
36 Efeito sobre a metaciclogecircnese
Formas epimastigotas (50 x 106 ceacutelulas mL
-1) foram cultivadas em meio LIT e apoacutes
trecircs dias foram lavadas duas vezes em PBS e transferidas para o meio Grace (Martins et al
2009) tratados ou natildeo (controle) com a concentraccedilatildeo correspondente ao IC50 determinado nas
curvas de crescimento das formas epimastigotas No sexto dia apoacutes a transferecircncia dos
parasitas para o meio Grace foi avaliado o efeito dos faacutermacos sobre a metaciclogecircnese Os
parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no estaacutegio
metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas metaciacuteclicas) foi
avaliado
36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

36
37 Anaacutelises do efeito dos compostos no ciclo intracelular do T cruzi
371 Toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos
A toxicidade dos faacutermacos em ceacutelulas de mamiacuteferos foi avaliada incubando ceacutelulas
CHO-K1 (50 x 105 ceacutelulas por poccedilo) em placas de 24 poccedilos em meio RPMI suplementado
com SFB (10) na presenccedila de diferentes concentraccedilotildees dos faacutermacos ou natildeo (controle) As
placas foram mantidas em estufa a 37 ordmC 5 de CO2 A viabilidade celular foi avaliada 48
horas apoacutes o iniacutecio do tratamento utilizando-se o meacutetodo de MTT (3-(45-dimetiltiazol-2-il)-
25-difenil tretazolio bromide) (Mosmann 1983) As ceacutelulas foram lavadas duas vezes com
PBS e adicionou-se em cada poccedilo 300 μl de PBS e 60 μl MTT (5mgmL) As placas foram
incubadas a 37 degC durante 3 horas (protegidas da luz) A reaccedilatildeo foi interrompida com a
adiccedilatildeo de 200 μl de SDS 10 A viabilidade das ceacutelulas foi observada mediante o
aparecimento da cor azul de formazan homogecircneo ldquothiazolidinrdquo As placas foram lidas em
espectofotocircmetro de placa utilizando-se λ 595 nm e como referecircncia λ 690 nm O IC50 foi
determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
372 Anaacutelise do efeito dos compostos na infectividade das formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) e tratadas com diferentes
concentraccedilotildees dos compostos Os parasitas ficaram em contato com as ceacutelulas por um periacuteodo
de quatro horas Apoacutes esse periacuteodo os parasitas que ainda estavam no sobrenadante foram
removidos juntamente com os compostos As ceacutelulas foram lavadas duas vezes com PBS e
acrescido meio RPMI 10 SFB A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes
esse periacuteodo o meio RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada
de 37 ordmC para 33 ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os
tripomastigotas que estavam no sobrenadante foram contados em cacircmara de Neubauer O IC50
foi determinado pelo ajuste dos dados em uma equaccedilatildeo sigmoidal dose-resposta
37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

37
373 Anaacutelise do efeito dos compostos apoacutes a invasatildeo celular pelas formas tripomastigotas
Ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de 24 poccedilos em meio
RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave placa as ceacutelulas foram
infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os parasitas ficaram em contato
com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse periacuteodo os parasitas que ainda
estavam no sobrenadante foram removidos As ceacutelulas foram lavadas duas vezes com PBS
adicionou-se meio RPMI 10 SFB e diferentes concentraccedilotildees dos compostos ou natildeo
(controle) A placa foi mantida na estufa a 37 ordmC por 16 horas Apoacutes esse periacuteodo o meio
RPMI 10 foi substituiacutedo por RPMI 2 de SFB e a temperatura alterada de 37 ordmC para 33
ordmC A placa foi mantida a 33 ordmC e no quinto dia poacutes-infecccedilatildeo os tripomastigotas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O IC50 foi determinado pelo ajuste
dos dados em uma equaccedilatildeo sigmoidal dose-resposta
374 Avaliaccedilatildeo da fase intracelular mais susceptiacutevel ao tratamento com os compostos
Para verificarmos em qual fase do ciclo intracelular o parasita eacute mais susceptiacutevel ao
tratamento com o faacutermaco ceacutelulas CHO-K1 (50 x 104
por poccedilo) foram mantidas em placas de
24 poccedilos em meio RPMI 10 SFB a 37 degC Apoacutes 24 horas quando jaacute estavam aderidas agrave
placa as ceacutelulas foram infectadas com formas tripomastigotas (25 x 106 por poccedilo) Os
parasitas ficaram em contato com as ceacutelulas por um periacuteodo de quatro horas Apoacutes esse
periacuteodo os parasitas que ainda estavam no sobrenadante foram removidos as ceacutelulas foram
lavadas duas vezes com PBS e adicionou-se meio RPMI 10 SFB O tratamento foi realizado
em diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado
anteriormente Diferentes poccedilos foram tratados em diferentes periacuteodos do ciclo durante o
periacuteodo da invasatildeo celular pelos parasitas apoacutes 24 horas ateacute 60 horas (fase amastigota) e apoacutes
60 horas (fase epimastigota intracelular) No quinto dia poacutes-infecccedilatildeo os parasitas que estavam
no sobrenadante foram contados em cacircmara de Neubauer O nuacutemero de parasitas de cada
tratamento foi comparado com o controle ceacutelulas infectadas natildeo tratadas
38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

38
38 Anaacutelises estatiacutesticas
O programa Excel 2007 foi utilizado para calcular meacutedia e desvio padratildeo das reacuteplicas
dos ensaios Os programas OriginPro8 e GraphPadPrism5 foram utilizados para construccedilatildeo
dos graacuteficos e ajustes dos mesmos o meacutetodo ANOVA de uma via seguido do teste Tukey foi
utilizado nas anaacutelises estatiacutesticas dos tratamentos em relaccedilatildeo ao controle Para anaacutelise de
diferenccedila entre grupos foi utilizado o teste T O valor de p lt 005 foi considerado
estatisticamente significativo
39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

39
4 RESULTADOS
41 Curvas de crescimento do T cruzi na presenccedila dos compostos
Os faacutermacos selecionados para verificaccedilatildeo de uma possiacutevel atividade anti- T cruzi
foram avaliados inicialmente em parasitas no estaacutegio epimastigota como mostram as curvas de
crescimento na Figura 5 O faacutermaco Vigabatrina que eacute um inibidor irreversiacutevel do receptor
GABA T foi avaliado em diferentes concentraccedilotildees (50 microM a 1 mM) (Figura 5 (a)) Jaacute a
Pregabalina anaacutelogo de GABA foi avaliada nas concentraccedilotildees (01 a 1 mM) (Figura 5 (b))
Os dois faacutermacos natildeo apresentaram efeito inibitoacuterio na proliferaccedilatildeo das formas epimastigota O
MTEP 3-((2-metil-4-thiazolil) etinil)piridine que atua como um antagonista seletivo natildeo
competitivo do receptor metabotroacutepico de glutamato mGluR5 foi avaliado nas concentraccedilotildees
(01 a 1 mM) (Figura 5 (c)) Observa-se que apenas a concentraccedilatildeo 1 mM apresenta efeito
inibitoacuterio na proliferaccedilatildeo de formas epimastigotas visto que todas as outras concentraccedilotildees
utilizadas inibem menos que o controle realizado com DMSO solvente utilizado na diluiccedilatildeo do
composto O MK-801 antagonista do receptor de glutamato do tipo NMDA apresentou efeito
inibitoacuterio na proliferaccedilatildeo das formas epimastigotas do T cruzi Esse composto foi avaliado nas
concentraccedilotildees (01 a 1 mM) como observado na Figura 5 (d) O IC50 determinado para o
tratamento realizado a 28 degC pH 75 foi de 03 plusmn 0028 mM conforme Figura 6 (a) A
Memantina que atua no sistema glutamateacutergico bloqueando a atividade de receptores de
glutamato do tipo NMDA tambeacutem foi avaliada nas curvas de crescimento do T cruzi
utilizando-se as concentraccedilotildees (30 a 300 microM) como pode ser obeservado na Figura 5 (e) Esse
composto tambeacutem apresentou efeito anti-proliferativo no crescimento do parasita Conforme
observado na Figura 6 (b) o IC50 determinado a 28 degC pH 75 foi de 1726 plusmn 9 microM
Memantina foi o faacutermaco que apresentou melhor efeito inibitoacuterio nas curvas de crescimento das
formas epimastigotas
40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

40
Figura 5 - Curvas de crescimento de T cruzi estaacutegio epimastigota tratados com os
faacutermacos de interesse
(a) Vigabatrina (50 microM a 1 mM) (b) Pregabalina (01a 1 mM) (c) MTEP (01 a 1 mM) (d) MK-801 (01 a
1 mM) e (e) Memantina (30 a 300 microM) Os parasitas foram mantidos a 28degC cultivados em meio LIT pH 75 O
controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o controle de inibiccedilatildeo (C
+)
utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

41
Figura 6 - Curvas dose-resposta do T cruzi estaacutegio epimastigota para os tratamentos
com MK-801 ou Memantina
(a) (b)
(a) parasitas tratados com MK-801 (01 a 1 mM) o IC50 determinado foi 03 plusmn 0028 mM (b) parasitas tratados
com Memantina (30 a 300 microM) o IC50 determinado foi 1726 plusmn 9 microM A anaacutelise foi realizada no quinto dia de
crescimento fase exponencial da cultura utilizando-se o programa OriginPro8
42 Interaccedilatildeo com condiccedilotildees de estresse
Estresse teacutermico e oxidativo
Baseados nos resultados apresentados anteriormente avaliamos um possiacutevel efeito
sineacutergico entre estresse teacutermico e o MK-801 e entre o estresse teacutermico ou oxidativo e a
Memantina (Figura 7) Como observado na Figura 7 (a) e (b) o MK-801 foi avaliado nas
concentraccedilotildees (01 a 045 mM) a 33 degC e a 37 degC respectivamente Jaacute a Memantina foi avaliada
nas concentraccedilotildees (100 a 500 microM) a 33 degC e (30 a 300 microM) a 37 degC conforme a Figura 7 (c) e
(d) respectivamente
A Memantina tambeacutem foi avaliada em parasitas estressados oxidativamente Os parasitas
foram tratados com o IC50 previamente estabelecido para H2O2 (726 microM) por uma hora Apoacutes
esse periacuteodo foram ressuspensos em LIT e tratados com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) e cultivados a 28 degC como observado na Figura 7 (e)
0 100 200 300
0
50
100
d
e in
ibiccedil
ao
Memantina (uM)0 100 200 300
0
50
100
d
e inib
iccedilao
Memantina (uM)
42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

42
Figura 7 - Curvas de crescimento do T cruzi estaacutegio epimastigota submetidos ao
estresse teacutermico e oxidativo e tratados com MK-801 ou Memantina
Os parasitas foram cultivados em meio LIT pH 75 em diferentes temperaturas e tratados com (a) MK-801 (01
a 045 mM) a 33 degC (b) MK-801 (01 a 045 mM) a 37 degC (c) Memantina (100 a 500 microM) a 33degC (d) Memantina (30 a
300 microM) a 37degC (e) tratados com 726 microM de H2O2 por 1 hora e com diferentes concentraccedilotildees de Memantina
(30 a 300 microM) a 28 degC O controle negativo (C-) foi realizado sem adiccedilatildeo de composto na cultura Para o
controle de inibiccedilatildeo (C+) utilizamos uma combinaccedilatildeo de antimicina (05 microM) e rotenona (60 microM)
43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

43
Ao avaliarmos os resultados observamos que natildeo haacute um efeito sineacutergico entre o MK-
801 e diferentes temperaturas visto que o IC50 determinado a 33 degC foi de 036 plusmn 006 mM e a
37 degC foi de 029 plusmn 0025 mM conforme Figura 8 (a) e (b) respectivamente Enquanto que
o IC50 determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) foi
03 plusmn 0028 mM
A Memantina tambeacutem natildeo apresentou efeito sineacutergico com as diferentes temperaturas
ou estresse oxidativo visto que o IC50 determinado nessas condiccedilotildees eacute maior do que o IC50
determinado em condiccedilotildees normais de cultivo (parasitas cultivados em LIT a 28 degC) Para os
parasitas cultivados a 33 degC observou-se um IC50 de 402 plusmn 60 microM conforme observado na
Figura 8 (c) e a 37 degC o IC50 foi de 228 plusmn 8 microM como mostra a Figura 8 (d) Em relaccedilatildeo
ao parasitas submetidos ao estresse oxidativo quando avaliamos o efeito do tratamento
simultacircneo com a concentraccedilatildeo correspondente ao IC25 da Memantina (142 plusmn 8 microM) e o IC50
do H2O2 (726 microM) foi observado uma diminuiccedilatildeo na proliferaccedilatildeo dos parasitas de 63 plusmn
4 mostrando que natildeo haacute um efeito sineacutergico e sim uma soma dos efeitos dos tratamentos
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4
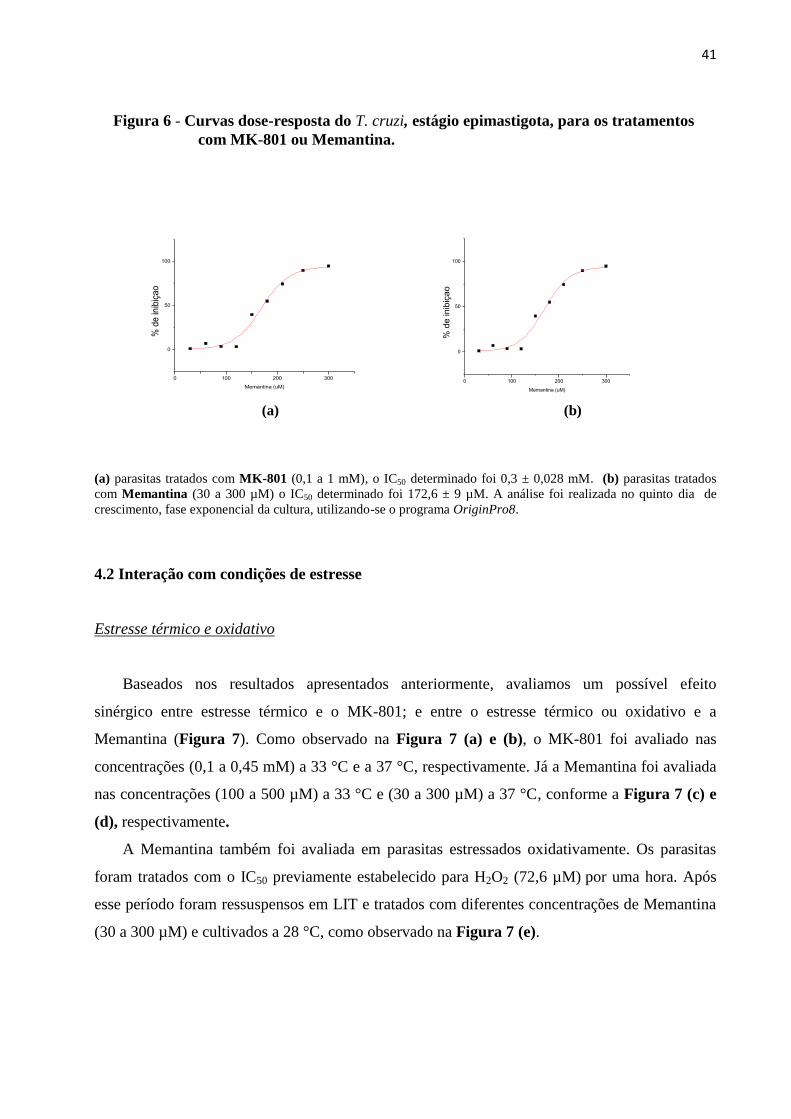
44
Figura 8 - Curvas dose-resposta do crescimento de T cruzi estaacutegio epimastigota
submetidos ao estresse teacutermico
Os parasitas foram tratados com (a) MK-801 (01 a 045 mM) a 33 degC IC50 036 plusmn 006 mM (b) MK-801 (01 a 045
mM) a 37 degC IC50 029 plusmn 0025 mM (c) Memantina (100 a 500 microM) a 33 degC IC50 402 plusmn 60 microM (d) Memantina (30
a 300 microM) a 37 degC IC50 228 plusmn 8 microM A anaacutelise foi realizada no quinto dia de crescimento fase exponencial da
cultura utilizando-se o programa OriginPro8
45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

45
O MK-801 e a Memantina apresentam o mesmo mecanismo de accedilatildeo por meio de
receptores de glutamato do tipo NMDA Devido o fato da Memantina ter apresentado melhor
efeito antiproliferativo em relaccedilatildeo ao MK-801 e quando administrado aos pacientes apresenta
pouco ou nenhum efeito colateral descrito decidimos escolher esse composto para prosseguir
os experimentos
Estresse metaboacutelico
Para verificar a resposta de epimastigotas ao estresse nutricional na presenccedila de 1726
microM de Memantina (IC50 determinado anteriormente) 50 x 106 ceacutelulas mL
-1 foram cultivadas
em LIT ou PBS e tratados ou natildeo (controle negativo) As ceacutelulas foram incubadas a 28 degC
durante 30 horas e apoacutes esse periacuteodo foi avaliado o niacutevel de ATP intracelular representado na
Figura 9 Observa-se uma diminuiccedilatildeo nos niacuteveis de ATP dos parasitas cultivados em LIT e
tratados com Memantina sugerindo que haacute uma interferecircncia no metabolismo energeacutetico do
parasita No entanto natildeo haacute um efeito sineacutergico entre o estresse metaboacutelico e o tratamento
visto que natildeo haacute diferenccedila estatisticamente significativa entre os parasitas que foram mantidos
em PBS e os parasitas que foram mantidos em PBS e tratados
Figura 9 - Resposta do Tcruzi estaacutegio epimastigota ao estresse nutricional na presenccedila
da Memantina
Parasitas submetidos ao estresse nutricional e ao tratamento com Memantina (1726 microM) IC50 determinado nas
curvas de crescimento Os parasitas foram incubados por 30 horas em LIT LIT+Memantina PBS
PBS+Memantina Apoacutes esse periacuteodo os niacuteveis de ATP intracelular foram avaliados Teste Tukey plt 005 MM
Memantina ns natildeo significativo
46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

46
43 Efeito da Memantina na metaciclogecircnese
Avaliamos o efeito da Memantina na metaciclogecircnese utilizando parasitas no estaacutegio
epimastigota (50 x 106 ceacutelulas mL
-1) cultivadas em meio LIT transferidos para o meio Grace
e tratados ou natildeo (controle) com 1726 microM de Memantina (concentraccedilatildeo correspondente ao
IC50 determinado nas curvas de crescimento) No sexto dia apoacutes a transferecircncia para meio
Grace os parasitas foram contados em cacircmara de Neubauer e a porcentagem de parasitas no
estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Como demonstrado na Figura 10 Memantina interfere na
metaciclogecircnese pois foi observado que culturas de parasitas tratados apresentam uma
diminuiccedilatildeo na diferenciaccedilatildeo de aproximadamente 30 em relaccedilatildeo ao controle
Figura 10 - Efeito da Memantina na metaciclogecircnese
Os parasitas foram cultivados em meio LIT e transferidos para o meio Grace e tratados ou natildeo (controle) com
1726 microM No sexto dia apoacutes a transferecircncia os parasitas foram contados em cacircmara de Neubauer e a
porcentagem de parasitas no estaacutegio metaciacuteclico em relaccedilatildeo ao nuacutemero total de parasitas (epimastigotas + formas
metaciacuteclicas) foi avaliado Resultado apresentado eacute a sobreposiccedilatildeo de trecircs ensaios independentes Meta
metaciacuteclico Epiepimastigota
47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

47
44 Tipo de morte celular em parasitas tratados com Memantina
Avaliamos tambeacutem o tipo de morte celular dos parasitas submetidos ao tratamento
com Memantina Os parasitas foram incubados na presenccedila de anexina V-FITC para
avaliarmos a exposiccedilatildeo de fosfatidilserina na membrana caracteriacutestica de apoptose como
tambeacutem na presenccedila de iodeto de propiacutedio para verificarmos uma possiacutevel ruptura da
membrana do parasita caracteriacutestica de necrose conforme descrito em materiais e meacutetodos
Os parasitas foram avaliados por citometria de fluxo como mostra a Figura 11 Como
observado na Figura 11(a) os parasitas controle (parasitas natildeo tratados) apresentaram 10
de positividade para exposiccedilatildeo de fosfatidilserina enquanto que os parasitas tratados com
Memantina apresentaram 42 conforme Figura 11(b) Outra caracteriacutestica de apoptose
observada nesse experimento eacute a manutenccedilatildeo da integridade da membrana do parasita visto
que as ceacutelulas praticamente natildeo foram marcadas pelo iodeto de propiacutedio Como observado na
Figura 11 (a) e (b) o nuacutemero de ceacutelulas marcadas dentre as que foram tratadas com
Memantina e no controle foi 046 e 078 respectivamente Esses resultados sugerem que
os parasitas tratados com Memantina apresentam caracteriacutesticas de morte do tipo apoptose e
natildeo necrose
48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

48
Figura 11 - Representaccedilatildeo da anaacutelise por citometria de fluxo para detecccedilatildeo do tipo de
morte celular
Os parasitas foram tratados com Memantina (1726 microM) ou natildeo (controle) durante 4 dias Apoacutes esse periacuteodo os
parasitas foram marcados com anexina V-FITC e Iodeto de Propiacutedio (PI) e avaliados (a) parasitas natildeo tratados
marcados com anexina e PI (b) parasitas tratados com Memantina marcados com anexina e PI (c) representaccedilatildeo
graacutefica de trecircs experimentos independentes Controles (d) parasitas natildeo tratados marcados apenas com anexina
(e) parasitas natildeo tratados marcados apenas com PI (f) controle positivo para necrose parasitas natildeo tratados
permeabilizados com digitonina e marcados com PI e (g) controle positivo para apoptose parasitas tratados com
estaurosporina 24 horas antes do ensaio e marcados com anexina
49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

49
Baseados no resultado anterior avaliamos outras caracteriacutestas descritas para apoptose
em tripanossomatiacutedeos Tem sido descrito que o aumento da produccedilatildeo de EROs eacute o principal
evento de sinalizaccedilatildeo e execuccedilatildeo desse processo (Irigoin et al 2009 Menna-Barreto et al
2009 Smirlis Soteriadou 2011) Portanto avaliamos a produccedilatildeo de H2O2 em parasitas
tratados com Memantina Formas epimastigotas foram tratadas com a concentraccedilatildeo
correspondente ao IC50 (1726 microM) e apoacutes 24 horas de tratamento os parasitas foram
incubados na presenccedila de amplex red e peroxidase (horsedish peroxidase) conforme descrito
em materiais e meacutetodos Como obeserva-se na Figura 12 os parasitas tratados produzem
mais H2O2 que os parasitas do grupo controle
Figura 12 - Representaccedilatildeo graacutefica da quantificaccedilatildeo de H2O2
Formas epimastigotas foram tratadas com Memantina (1726 microM) ou natildeo (controle) durante 24 horas Apoacutes esse
periacuteodo os parasitas (10 x 107) foram lavados por centrifugaccedilatildeo ressuspensos em PBS (5mM de succinato) e
adicionou-se 25 microM de Amplex Red e 005 UmL de Horseradish Peroxidase As amostras foram analisadas no
fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ excitaccedilatildeo 563 nm e λ emissatildeo 587 nm Teste T
plt 005
50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

50
Jaacute foi descrito que aleacutem da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem contribui para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010) Portanto
avaliamos tambeacutem a concentraccedilatildeo intracelular desse caacutetion em formas epimastigotas expostas
ao tratamento com Memantina conforme descrito em materiais e meacutetodos Como se pode
observar na Figura 13 a exposiccedilatildeo dos parasitas ao tratamento com Memantina aumenta a
concentraccedilatildeo intracelular de Ca2+
quando comparados aos parasitas do grupo controle
sugerindo mais uma vez que o parasita estaacute sofrendo apoptose
Figura 13 - Representaccedilatildeo graacutefica da concentraccedilatildeo intracelular de Ca2+
Formas epimastigotas foram tratados com Memantina (1726 microM ) ou natildeo (controle) durante 4 dias Apoacutes esse
periacuteodo os parasitas (25 x 107) foram incubados com 5 microM de Fluo-4 AM por 1 hora a 28 ordmC lavadas duas
vezes em tampatildeo hepes-glicose e avaliadas no fluoriacutemetro Spectra Max M3 Molecular Devices utilizando-se λ
excitaccedilatildeo 490 nm e λ emissatildeo 518 nm Teste T plt 005
Observamos tambeacutem a morfologia dos parasitas tratados com Memantina em relaccedilatildeo
ao controle Comforme podemos observar na Figura 14 os parasitas tratados apresentam
mudanccedila na sua morfologia apresentando forma arredondada (Figura 14 (c) e (d)) em
relaccedilatildeo ao controle (parasitas que natildeo foram tratados) como mostra a Figura 14 (a) e (b)
Uma caracteriacutestica tambeacutem descrita para ceacutelulas apoptoacuteticas (Kaczanowski et al 2011
Smirlis et al 2010)
51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

51
Figura 14 - Formas epimastigotas de T cruzi
Formas epimastigotas no quarto dia de crescimento (a) e (b) parasitas sem tratamento (controle) (c) e (d)
parasitas tratados com Memantina (1726 microM ) Coloraccedilatildeo Panoacutetico Resoluccedilatildeo (a) e (c) 40x (b) e (d) 100x
52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

52
45 Efeito da Memantina no ciclo intracelular do T cruzi
Verificamos tambeacutem o efeito da Memantina no ciclo intracelular do parasita
conforme Figura 15 Inicialmente foi avaliada a toxicidade do composto em ceacutelulas CHO-K1
modelo de infecccedilatildeo utilizado por nosso grupo As ceacutelulas CHO-K1 foram incubadas por 48
horas na presenccedila de diferentes concentraccedilotildees (50 microM a 1 mM) ou natildeo (controle) conforme
descrito em materiais e meacutetodos Como se pode observar na Figura 15 (a) Memantina eacute bem
tolerada pelas ceacutelulas CHO-K1 pois nas concentraccedilotildees abaixo de 04 mM (valor acima do
IC50 determinado para o estaacutegio epimastigota) natildeo apresenta diferenccedila em relaccedilatildeo ao controle
Como observado na Figura 16 (a) o IC50 determinado foi 6245 plusmn 46 microM Com base neste
resultado realizamos os ensaios de infecccedilatildeo tratando com concentraccedilotildees de Memantina natildeo
toacutexicas para as ceacutelulas CHO-K1 em diferentes periacuteodos da infecccedilatildeo
Para verificarmos o efeito do faacutermaco na infectividade de formas tripomastigotas as
ceacutelulas foram infectadas como descrito em materiais e meacutetodos e tratadas ou natildeo (controle)
durante somente o momento da invasatildeo celular com diferentes concentraccedilotildees de Memantina
(50 a 400 microM) conforme representado na Figura 15 (b) O IC50 determinado nesse tratamento
foi de 2063 microM como pode ser observado na Figura 16 (b) Em todos os tratamentos foi
observada uma diminuiccedilatildeo estatisticamente significativa do nuacutemero de formas tripomastigotas
que eclodiram das ceacutelulas tratadas em relaccedilatildeo ao controle conforme Figura 15 (b)
Avaliamos tambeacutem o efeito no tratamento apoacutes a infecccedilatildeo pelo T cruzi Ceacutelulas CHO-
K1 foram infectadas com formas tripomastigotas e apoacutes a invasatildeo celular pelos parasitas
iniciamos o tratamento com diferentes concentraccedilotildees de Memantina (5 a 300 microM) conforme
descrito em materiais e meacutetodos Como observado na Figura 15 (c) todos os tratamentos
mostraram uma diminuiccedilatildeo estatisticamente significativa no nuacutemero de formas
tripomastigotas O IC50 determinado foi de 31 microM conforme Figura 16 (c)
Apoacutes a verificaccedilatildeo do efeito da Memantina durante a invasatildeo celular e apoacutes a
infecccedilatildeo consideramos relevante a avaliaccedilatildeo de qual fase do ciclo intracelular (amastigota ou
epimastigota intracelular) seria mais susceptiacutevel ao tratamento Neste experiemento como
descrito em materiais e meacutetodos o tratamento foi realizado em diferentes periacuteodos da
infecccedilatildeo com a concentraccedilatildeo correspondente ao IC50 (31 microM) O nuacutemero de parasitas de cada
periacuteodo de tratamento foi comparado com o controle ceacutelulas infectadas e natildeo tratadas Como
observado na Figura 15 (d) a fase mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota
forma intracelular replicativa e infectiva
53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

53
Figura 15 - Efeito da Memantina no ciclo intracelular do T cruzi
(a) Viabilidade das ceacutelulas CHO-K1 tratadas com diferentes concentraccedilotildees de Memantina (50 a 1 mM) As
ceacutelulas foram incubadas por 48 horas e a viabilidade celular foi avaliada utilizando-se o meacutetodo de MTT (b)
Efeito na infectividade de formas tripomastigotas Os parasitas foram tratados somente durante o periacuteodo da
infecccedilatildeo (c) Efeito no tratamento apoacutes a invasatildeo dos parasitas nas ceacutelulas CHO-K1 (d) Avaliaccedilatildeo de qual fase
do ciclo intracelular do T cruzi eacute mais susceptiacutevel ao tratamento com Memantina As ceacutelulas foram tratadas em
diferentes periacuteodos com a concentraccedilatildeo correspondente ao IC50 determinado anteriormente (31 microM) T
tripomastigota A amastigota EI epimastigota Em todos os experimentos foi avaliada a eclosatildeo de formas
tripomastigotas no quinto dia poacutes-infecccedilatildeo por contagem dos parasitas que se encontravam no sobrenadante em
cacircmara de Neubauer Teste Tukey plt 005
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4
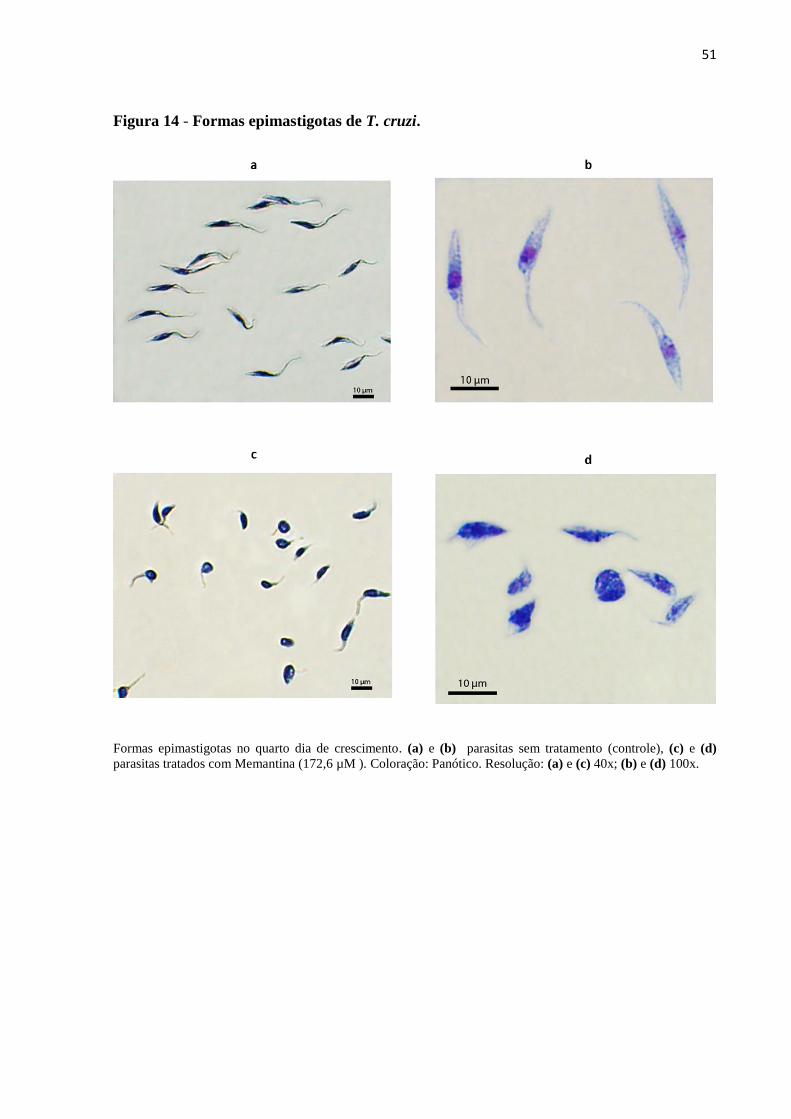
54
Figura 16 - Curvas dose-resposta da toxicidade em ceacutelulas CHO-K1 infectividade de
formas tripomastigostas e tratamento da infecccedilatildeo apoacutes a invasatildeo celular
(a) Resposta das ceacutelulas CHO-k1 ao tratamento com diferentes concentraccedilotildees do Memantina (50 microM a 1 mM)
durante 48 horas e avaliados pelo ensaio de MTT IC50 6245 plusmn 46 microM (b) tratamento de parasitas no estaacutegio
tripomastigota durante o periacuteodo da invasatildeo celular com diferentes concentraccedilotildees de Memantina (50 a 400 microM)
IC50 2063 microM (c) tratamento da infecccedilatildeo apoacutes a invasatildeo pelos parasitas durante todo o periacuteodo da infecccedilatildeo
com diferentes concentraccedilotildees de Memantina (5 a 300 microM) IC50 31 microM A anaacutelise foi realizada no quinto dia
apoacutes a infecccedilatildeo pela contagem em cacircmara de Neubauer dos parasitas que encontravam-se no sobrenadante A
determinaccedilatildeo do valor de IC50 foi realizada utilizando-se o programa OriginPro8
55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

55
5 DISCUSSAtildeO
Como demonstrado nos resultados os faacutermacos Vigabatrina (inibidor do receptor
GABA T) e Pregabalina (anaacutelogo do GABA) natildeo apresentaram efeito inibitoacuterio na
proliferaccedilatildeo de formas epimastigotas Poreacutem jaacute havia sido relatado a atividade tripanocida de
faacutermacos anaacutelogos de GABA Por exemplo o acamprosato que eacute utilizado no tratamento da
dependecircncia alcooacutelica foi avaliado pelo nosso grupo em curvas de crescimento de formas
epimastigotas e apresentou efeito inibitoacuterio (IC50 250 microM) (Lange 2012)
Dentre os faacutermacos com accedilatildeo glutamateacutergica o MTEP que atua como um antagonista
seletivo natildeo competitivo do receptor metabotroacutepico mGluR5 tambeacutem natildeo apresentou efeito
inibitoacuterio nas curvas de crescimento das formas epimastigotas Jaacute o MK-801 e a Memantina
demonstraram efeito inibitoacuterio na proliferaccedilatildeo dos parasitas Ambos atuam em mamiacuteferos
bloqueando receptores de glutamato do tipo NMDA principalmente em humanos onde esses
receptores de glutamato estatildeo bem caracterizados e associados a canais iocircnicos
principalmente Ca2+
(Coan et al 1987 Lipton 2005 Xia et al 2010) Em T cruzi
receptores de glutamato do tipo NMDA ainda natildeo foram caracterizados Uma anaacutelise feita na
sequecircncia do genoma de T cruzi demonstrou que natildeo haacute uma sequecircncia compatiacutevel com
receptores do tipo NMDA de humanos (TriTrypDB) No entanto Paveto e colaboradores tecircm
sugerido que T cruzi pode apresentar receptores de glutamato similar ao receptor NMDA
descrito para o tecido nervoso e como ocorre em ceacutelulas neurais receptores NMDA de T
cruzi seriam provavelmente os principais responsaacuteveis por controlar os niacuteveis de Ca2+
citosoacutelico do parasita (Paveto et al 1995)
MK-801 e Memantina satildeo utilizados no controle de doenccedilas neurodegenerativas ou
ligadas ao sistema nervoso central Jaacute foi demonstrado que o MK-801 apresenta propriedades
anesteacutesicas e anticonvulsivantes (Ameer et al 2010) Em formas epimastigotas de T cruzi foi
observado que a conversatildeo de L-[3H]arginina para [
3H]citrulina eacute estimulada por L-glutamato
e NMDA e eacute bloqueada por EGTA MK-801 e ketamina (Paveto et al 1995) Tambeacutem foi
verificada uma dimuniccedilatildeo na motilidade de epimastigotas tratados com MK-801
provavelmente por interferecircncia nos niacuteveis de GMP ciacuteclico mediado por oacutexido niacutetrico (Pereira
et al 1997) Jaacute a Memantina eacute uma amina triciacuteclica o primeiro da classe de novos
medicamentos para a doenccedila de Alzheimer (Lipton 2005 Xia et al 2010) Jaacute foi descrita
atividade tripanocida para outras aminas O Trypanosoma brucei por exemplo eacute sensiacutevel ao
tratamento com Rimantadina uma droga utilizada para o tratamento e profilaxia da infecccedilatildeo
pelo viacuterus da influenza A (Kelly et al 1999) Kelly e colaboradores tambeacutem sintetizaram e
56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

56
testaram vaacuterios compostos derivados da aminoadamantana e aminoalquilciclohexano e
observaram que vaacuterios desses compostos apresentam atividade consideraacutevel contra as formas
sanguiacuteneas do T brucei tanto in vitro como tambeacutem in vivo (Kelly et al 2001)
Recentemente o mesmo grupo sintetizou compostos anaacutelogos de amantadina e memantina
que continham caracteriacutesticas estruturais necessaacuterias para atividade antagonista de receptores
NMDA e atividade tripanocida Esses compostos foram avaliados em T brucei e foi
observado que a introduccedilatildeo de um grupo etil em C-5 aumenta a atividade tripanocida (Duque
et al 2010)
Ao avaliarmos a interaccedilatildeo do MK-801 ou da Memantina com diferentes situaccedilotildees de
estresse (metaboacutelico teacutermico e oxidativo) observamos que natildeo haacute interaccedilatildeo visto que os
valores de IC50 obtidos nessas condiccedilotildees foram maiores ou iguais do que foi obtido quando os
parasitas foram mantidos em condiccedilotildees normais de cultivo Uma interaccedilatildeo entre os faacutermacos
e as condiccedilotildees de estresse seria interessante do ponto de vista terapecircutico uma vez que o T
cruzi estaacute exposto a situaccedilotildees de estresse teacutermico metaboacutelico e oxidativo durante o seu ciclo
de vida jaacute que transita em diferentes regiotildees do intestino do inseto vetor e no meio intra e
extracelular no hospedeiro mamiacutefero (Silber et al 2005a) O parasita eacute submetido ao estresse
teacutermico quando passa pelo inseto vetor visto que o triatomiacuteneo natildeo regula sua temperatura
interna podendo variar de acordo com a temperatura no ambiente externo Como tambeacutem o
parasita alterna entre o hospedeiro mamiacutefero que apresenta uma temperatura meacutedia de 37 ordmC e
o inseto vetor no qual a meacutedia de temperatura eacute 28 ordmC (Magdaleno et al 2009) O estresse
metaboacutelico ocorre principalmente na porccedilatildeo final do intestino do triatomiacuteneo onde ocorre a
diferenciaccedilatildeo das formas epimastigotas para tripomastigotas metacicliacutecos (Contreras et al
1985) e no estabelecimento da infecccedilatildeo no citoplasma das ceacutelulas do hospedeiro mamiacutefero
onde devem competir por nutrientes com a proacutepia ceacutelula hospedeira Aleacutem do mais os
parasitas precisam adaptar-se a disponibilidade de diferentes nutrientes nos diversos
ambientes (diferentes meios extracelulares e intracelulares no hospedeiro mamiacutefero e
diferentes porccedilotildees do tubo digestivo no inseto vetor) (Silber et al 2009) Jaacute o estresse
oxidativo pode ocorrer tanto pelo metabolismo do proacuteprio parasita ou pela resposta imune do
hospedeiro mamiacutefero (Turrens 2004)
A Memantina tambeacutem interferiu na metaciclogecircnese processo de diferenciaccedilatildeo em
que o parasita passa do estaacutegio epimastigota (estaacutegio replicativo e natildeo infectivo) para
tripomastigota metaciacuteclico (estaacutegio infectivo e natildeo replicativo) Esse processo ocorre
naturalmente na porccedilatildeo final do intestino de triatomiacuteneos onde o parasita encontra
aminoaacutecidos como prolina glutamato e aspartato que satildeo importantes nesse processo
57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

57
(Contreras et al 1985) pois permitem ao parasita restabelecer os niacuteveis de ATP intracelular
necessaacuterios agrave metaciclogecircnese (Cazzulo 1992) Os parasitas que foram submetidos ao
tratamento apresentaram uma diminuiccedilatildeo no nuacutemero de formas metaciacuteclicas em
aproximadamente 30 em relaccedilatildeo ao controle Como jaacute mencionado tambeacutem obeservamos
uma diminuiccedilatildeo dos niacuteveis de ATP intracelular dos parasitas quando desafiados com
Memantina Provavelmente essa inibiccedilatildeo na metaciclogecircnese pode estar relacionada com a
diminuiccedilatildeo nos niacuteveis de ATP
T cruzi eacute um parasita intracelular obrigatoacuterio como mencionado anteriormente no
hospedeiro mamiacutefero o parasita passa por diferenciaccedilatildeo do estaacutegio tripomastigota para
amastigota e novamente para tripomastigota passando por um estaacutegio intermediaacuterio chamado
epimastigota intracelular (Almeida-De-Faria et al 1999) Como observado nos resultados a
Memantina interfere no ciclo intracelular do T cruzi Tanto na infectividade de formas
tripomastigotas como no tratamento ao longo do periacuteodo de infeccedilatildeo diminuindo o nuacutemero de
formas tripomastigotas que eclodem no quinto dia apoacutes a infecccedilatildeo A fase do ciclo intracelular
mais susceptiacutevel ao tratamento eacute o estaacutegio amastigota uacutenico estaacutegio replicativo do ciclo
intracelular Esse dado eacute muito importante visto que na fase crocircnica da doenccedila de Chagas o
parasita forma ninhos de amastigotas em tecidos do hospedeiro mamiacutefero principalmente no
tecido cardiacuteaco (Rassi Marin-Neto 2010) e nesta fase da doenccedila a eficaacutecia da terapia
disponiacutevel atualmente eacute controrversa (Boscardin et al 2009 Silber et al 2005b)
Nossos resultados tambeacutem sugerem que o tratamento com Memantina desencadea o
mecanismo de morte celular programada com caracteriacutesticas de apoptose em formas
epimastigotas de T cruzi Como foi demonstrado formas epimastigotas tratadas com
Memantina apresentam aumento da produccedilatildeo de EROs diminuiccedilatildeo dos niacuteveis de ATP
intracelular aumento da concentraccedilatildeo de Ca2+
intracelular exposiccedilatildeo de fosfatidilserina na
membrana do parasita aleacutem de mudanccedilas morfoloacutegicas Esse tipo de MCD tem sido descrita
para protistas unicelulares incluindo T cruzi Plasmodium e Leishmania (Al-Olayan et al
2002 Ameisen et al 1995 Das et al 2001 Duszenko et al 2006) Alguns autores tem
descrito a participaccedilatildeo da mitococircndria no processo de apoptose em T cruzi semelhante ao
que ocorre em metazoaacuterios onde a mitococircndria eacute a principal organela envolvida nesse
processo Mudanccedilas na funccedilatildeo mitocondrial principalmente quando ocorre a produccedilatildeo de
(EROs) foram identificadas como o principal evento de sinalizaccedilatildeo e execuccedilatildeo desse processo
(Irigoin et al 2009 Menna-Barreto et al 2009 Smirlis et al 2010) Foi descrito tambeacutem
que aleacutem do aumento da produccedilatildeo de EROs a desregulaccedilatildeo da homeostase de Ca2+
tambeacutem
pode contribuir para a apoptose em tripanossomatiacutedeos (Smirlis et al 2010)
58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

58
Os dados apresentados demonstram que dos faacutermacos estudados apenas Memantina
apresentou um melhor efeito tripanocida Inibe a proliferaccedilatildeo de formas epimastigotas
interfere na metaciclogecircnese como tambeacutem afeta o metabolismo energeacutetico do parasita com a
diminuiccedilatildeo dos niacuteveis de ATP aleacutem de desencadear mecanismos que levam a apoptose das
formas epimastigotas Aleacutem do mais interfere no ciclo intracelular do parasita mais
especificamente no estaacutegio amastigota Interessantemente as fases do ciclo do parasita que
satildeo mais afetadas satildeo as fases que requerem mais energia estaacutegios replicativos (amastigota e
epimastigota) como tambeacutem os processos de diferenciaccedilatildeo e invasatildeo celular Estes dados
sugerem que o metabolismo energeacutetico do parasita estaacute sendo afetado como tambeacutem
provalvelmente ocorra uma dimuniccedilatildeo na motilidade do parasita pela interferecircncia nos niacuteveis
de GMP ciacuteclico mediado por oacutexido niacutetrico conforme sugerido por Pereira e colaboradores
(Pereira et al 1997)
Apesar de apresentar valores de IC50 que podem ser considerados altos em relaccedilatildeo aos
valores descritos para formas epimastigotas tratadas com Benzonidazol (76 a 32 microM)
variando de acordo com a cepa do T cruzi (Moreno et al 2010) esses resultados mostram
perspectivas promissoras visto que a Memantina poderaacute ser utilizado como composto liacuteder
para o desenho de derivados com atividade anti-T cruzi otimizada como jaacute estaacute sendo
trabalhado em relaccedilatildeo ao tratamento do Trypanosoma brucei (Duque et al 2010) Aleacutem do
mais jaacute eacute utilizado em humanos para doenccedila de Alzheimer sendo bem tolerado pelos
pacientes
59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

59
6 CONCLUSOtildeES
Diante do exposto concluimos que
1- Os faacutermacos Vigabatrina Pregabalina e MTEP natildeo apresentaram efeito inibitoacuterio nas
curvas de crescimento de T cruzi
2- Os faacutermacos MK-801 e Memantina apresentaram efeito antiproliferativo em formas
epimastigotas sendo que Memantina apresentou um valor de IC50 mais baixo sendo portanto
mais eficaz Interessantemente os dois compostos tem o mesmo mecanismo de accedilatildeo descrito
na literatura para mamiacuteferos receptores de glutamato do tipo NMDA
3- Memantina foi capaz de desencadear mecanismos de morte celular programada do
tipo apoptose em formas epimastigotas as quais apresentaram exposiccedilatildeo de fosfatidilserina
na membrana externa aumento de Ca2+
intracelular aumento da geraccedilatildeo de EROs
diminuiccedilatildeo dos niacuteveis de ATP e mudanccedilas morfoloacutegicas
4- O processo de metaciclogecircnese tambeacutem foi afetado pelo tratamento com Memantina
visto que ocorreu uma diminuiccedilatildeo no nuacutemero de parasitas que diferenciam quando tratados
5- Memantina eacute bem tolerada pelas ceacutelulas CHO-K1 visto que o valor de IC50
determinado quando as ceacutelulas foram desafiadas eacute quase trecircs vezes maior que o determinado
nas curvas de crescimento de formas epimastigotas e mais de dez vezes maior do que o valor
determinado no tratamento do ciclo intracelular (CHO-K1 6245 plusmn 46 microM formas
epimastigotas 1726 microM e ciclo intracelular 31 microM)
6- A capacidade de infecccedilatildeo de formas tripomastigotas foi dimuiacuteda quando os parasitas
foram tratados com Memantina durante o periacuteodo da invasatildeo celular como tambeacutem
observou-se uma diminuiccedilatildeo do nuacutemero de parasitas que eclodiram quando as ceacutelulas CHO-
K1 infectadas foram tratadas durante todo periacuteodo de infecccedilatildeo
7- O estaacutegio amastigota eacute a forma mais susceptiacutevel ao tratamento com Memantina
60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

60
8- As fases do ciclo de vida do parasita mais afetadas satildeo as fases que requerem mais
energia estaacutegios replicativos (amastigota e epimastigota) Como tambeacutem os processos de
diferenciaccedilatildeo e invasatildeo celular Esses dados em conjunto sugerem que o tratamento com
Memantina interfere no metabolismo energeacutetico do parasita
61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

61
REFEREcircNCIAS
Acquatella H Echocardiography in Chagas heart disease Circulation 2007115(9)1124-31
Almeida-de-Faria M Freymuller E Colli W Alves MJ Trypanosoma cruzi characterization
of an intracellular epimastigote-like form Exp Parasitol 199992(4)263-74
Al-Olayan EM Williams GT Hurd H Apoptosis in the malaria protozoan Plasmodium
berghei a possible mechanism for limiting intensity of infection in the mosquito Int J
Parasitol 200232(9)1133-43
Alves MJ Colli W Role of the gp85trans-sialidase superfamily of glycoproteins in the
interaction of Trypanosoma cruzi with host structures Subcell Biochem 20084758-69
Ameer NH Lee JH Choi MA Jin GS Kim MS Park BR The Effects of Glutamate NMDA
Receptor Antagonist MK-801 on Gastrointestinal Motility after Middle Cerebral Artery
Occlusion in Rats Korean J Physiol Pharmacol 201014(3)151-6
Ameisen JC Idziorek T Billaut-Mulot O Loyens M Tissier JP Potentier A et al Apoptosis
in a unicellular eukaryote (Trypanosoma cruzi) implications for the evolutionary origin and
role of programmed cell death in the control of cell proliferation differentiation and survival
Cell Death Differ 19952(4)285-300
Berg M Van der Veken P Joossens J Muthusamy V Breugelmans M Moss CX et al
Design and evaluation of Trypanosoma brucei metacaspase inhibitors Bioorg Med Chem
Lett 201020(6)2001-6
Bern C Verastegui M Gilman RH Lafuente C Galdos-Cardenas G Calderon M et al
Congenital Trypanosoma cruzi transmission in Santa Cruz Bolivia Clin Infect Dis
200949(11)1667-74
Boscardin SB Torrecilhas AC Manarin R Revelli S Rey EG Tonelli RR et al Chagas
disease an update on immune mechanisms and therapeutic strategies J Cell Mol Med
200914(6B)1373-84
Brener Z Chiari E Morphological variations observed in different strains of Trypanosoma
cruzi Rev Inst Med Trop Sao Paulo 196319220-4
Brener Z Life cycle of Trypanosoma cruzi Rev Inst Med Trop Sao Paulo 197113(3)171-8
Bringaud F Riviere L Coustou V Energy metabolism of trypanosomatids adaptation to
available carbon sources Mol Biochem Parasitol 2006149(1)1-9
De acordo com
International Committee of Medical Journal Editors [Internet] Uniform requirements for manuscripts
submitted to Biomedical Journal sample references [updated 2011 Jul 15] Available
from httpwwwicmjeorg
62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

62
Camargo EP Growth and Differentiation in Trypanosoma cruzi I Origin of Metacyclic
Trypanosomes in Liquid Media Rev Inst Med Trop Sao Paulo 19641293-100
Cancado JR Criteria of Chagas disease cure Mem Inst Oswaldo Cruz 199994(1)331-5
Canepa GE Silber AM Bouvier LA Pereira CA Biochemical characterization of a low-
affinity arginine permease from the parasite Trypanosoma cruzi FEMS Microbiol Lett
2004236(1)79-84
Cazzulo JJ Energy metabolism in Trypanosoma cruzi Subcell Biochem 199218235-57
Cazzulo JJ Intermediate metabolism in Trypanosoma cruzi J Bioenerg Biomembr
199426(2)157-65
Coan EJ Saywood W Collingridge GL MK-801 blocks NMDA receptor-mediated synaptic
transmission and long term potentiation in rat hippocampal slices Neurosci Lett
198780(1)111-4
Contreras VT Salles JM Thomas N Morel CM Goldenberg S In vitro differentiation of
Trypanosoma cruzi under chemically defined conditions Mol Biochem Parasitol
198516(3)315-27
Coura JR Junqueira AC Fernandes O Valente SA Miles MA Emerging Chagas disease in
Amazonian Brazil Trends Parasitol 200218(4)171-6
Das M Mukherjee SB Shaha C Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes J Cell Sci 2001114(Pt 13)2461-9
de Souza AP Melo de Oliveira G Neve J Vanderpas J Pirmez C de Castro SL et al
Trypanosoma cruzi host selenium deficiency leads to higher mortality but similar parasitemia
in mice Exp Parasitol 2002101(4)193-9
Debrabant A Lee N Bertholet S Duncan R Nakhasi HL Programmed cell death in
trypanosomatids and other unicellular organisms Int J Parasitol 200333(3)257-67
Dias LC Dessoy MA Silva JJN Thiemann OH Oliva G Andricopulo AD Chemotherapy of
Chagas disease state of the art and perspectives for the development of new drugs Quiacutem
Nova 200932
Docampo R Sensitivity of parasites to free radical damage by antiparasitic drugs Chem Biol
Interact 199073(1)1-27
Dolai S Yadav RK Pal S Adak S Overexpression of mitochondrial Leishmania major
ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death
and protein damage Eukaryot Cell 20098(11)1721-31
Duque MD Camps P Torres E Valverde E Sureda FX Lopez-Querol M et al New
oxapolycyclic cage amines with NMDA receptor antagonist and trypanocidal activities
Bioorg Med Chem 201018(1)46-57
63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

63
Duschak VG Couto AS An insight on targets and patented drugs for chemotherapy of
Chagas disease Recent Pat Antiinfect Drug Discov 20072(1)19-51
Duszenko M Figarella K Macleod ET Welburn SC Death of a trypanosome a selfish
altruism Trends Parasitol 200622(11)536-42
Filardi LS Brener Z Susceptibility and natural resistance of Trypanosoma cruzi strains to
drugs used clinically in Chagas disease Trans R Soc Trop Med Hyg 198781(5)755-9
Gajcy K Lochynski S Librowski T A role of GABA analogues in the treatment of
neurological diseases Curr Med Chem 201017(22)2338-47
Gran L Borchgrevink CF Pain treatment of patients with cancer Tidsskr Nor Laegeforen
1989109(33)3424-6
Ha KY Kim YH Rhyu KW Kwon SE Pregabalin as a neuroprotector after spinal cord
injury in rats Eur Spine J 200817(6)864-72
Hampton JR Lysine uptake in cultured Trypanosoma cruzi interactions of competitive
inhibitors J Protozool 197017(4)597-600
Hoare C The Trypanosomes of Mammals Blackwell Scientific Publication Oxford 1972
Homsy JJ Granger B Krassner SM Some factors inducing formation of metacyclic stages of
Trypanosoma cruzi J Protozool 198936(2)150-3
Irigoin F Inada NM Fernandes MP Piacenza L Gadelha FR Vercesi AE et al
Mitochondrial calcium overload triggers complement-dependent superoxide-mediated
programmed cell death in Trypanosoma cruzi Biochem J 2009418(3)595-604
Iso Y Grajkowska E Wroblewski JT Davis J Goeders NE Johnson KM et al Synthesis and
structure-activity relationships of 3-[(2-methyl-13-thiazol-4-yl)ethynyl]pyridine analogues as
potent noncompetitive metabotropic glutamate receptor subtype 5 antagonists search for
cocaine medications J Med Chem 200649(3)1080-100
Jimenez V Paredes R Sosa MA Galanti N Natural programmed cell death in T cruzi
epimastigotes maintained in axenic cultures J Cell Biochem 2008105(3)688-98
Jung MJ Lippert B Metcalf BW Bohlen P Schechter PJ gamma-Vinyl GABA (4-amino-
hex-5-enoic acid) a new selective irreversible inhibitor of GABA-T effects on brain GABA
metabolism in mice J Neurochem 197729(5)797-802
Kaczanowski S Sajid M Reece SE Evolution of apoptosis-like programmed cell death in
unicellular protozoan parasites Parasit Vectors 2011444
Kelly JM Miles MA Skinner AC The anti-influenza virus drug rimantadine has trypanocidal
activity Antimicrob Agents Chemother 199943(4)985-7
64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

64
Kelly JM Quack G Miles MM In vitro and in vivo activities of aminoadamantane and
aminoalkylcyclohexane derivatives against Trypanosoma brucei Antimicrob Agents
Chemother 200145(5)1360-6
Kroemer G El-Deiry WS Golstein P Peter ME Vaux D Vandenabeele P et al
Classification of cell death recommendations of the Nomenclature Committee on Cell Death
Cell Death Differ 200512 Suppl 21463-7
Lange IES Estudo dos efeitos dos psicofaacutermacos acamprosato e levomepromazina sobre as
formas epimastigotas e tripomastigotas do Trypanosoma cruzi in vitro Satildeo Paulo
Universidade de Satildeo Paulo 2012
Laverriere M Cazzulo JJ Alvarez VE Antagonic activities of Trypanosoma cruzi
metacaspases affect the balance between cell proliferation death and differentiation Cell
Death Differ 201219(8)1358-69
Lipton SA The molecular basis of memantine action in Alzheimers disease and other
neurologic disorders low-affinity uncompetitive antagonism Curr Alzheimer Res
20052(2)155-65
Liu JL Li M Dang XR Wang ZH Rao ZR Wu SX et al A NMDA receptor antagonist
MK-801 impairs consolidating extinction of auditory conditioned fear responses in a
Pavlovian model PLoS One 20094(10)e7548
Lockshin RA and Williams CM Programmed Cell Death J Insect Physiol 1964643
Magdaleno A Ahn IY Paes LS Silber AM Actions of a proline analogue L-thiazolidine-4-
carboxylic acid (T4C) on Trypanosoma cruzi PLoS One 20094(2)e4534
Magdaleno A Mantilla BS Rocha SC Pral EMF Silber AM The involvement of glutamate
metabolism in the resistance to thermal nutritional and oxidative stress in Trypanosoma cruzi
Enzyme Research 2011
Maguire JH Chagas disease--can we stop the deaths N Engl J Med 2006355(8)760-1
Mancilla R Naquira C Lanas C Protein biosynthesis in trypanosomidae II The metabolic
fate of DL-leucine-1-C14 in Trypanosoma cruzi Exp Parasitol 196721(2)154-9
Marin Neto JA Simoes MV Sarabanda AV Chagas heart disease Arq Bras Cardiol
199972(3)247-80
Martins RM Covarrubias C Rojas RG Silber AM Yoshida N Use of L-proline and ATP
production by Trypanosoma cruzi metacyclic forms as requirements for host cell invasion
Infect Immun 200977(7)3023-32
Maya JD Bollo S Nunez-Vergara LJ Squella JA Repetto Y Morello A et al Trypanosoma
cruzi effect and mode of action of nitroimidazole and nitrofuran derivatives Biochem
Pharmacol 200365(6)999-1006
65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

65
Maya JD Cassels BK Iturriaga-Vasquez P Ferreira J Faundez M Galanti N et al Mode of
action of natural and synthetic drugs against Trypanosoma cruzi and their interaction with the
mammalian host Comp Biochem Physiol A Mol Integr Physiol 2007146(4)601-20
Menna-Barreto RF Correa JR Cascabulho CM Fernandes MC Pinto AV Soares MJ et al
Naphthoimidazoles promote different death phenotypes in Trypanosoma cruzi Parasitology
2009136(5)499-510
Moreira DR Leite AC dos Santos RR Soares MB Approaches for the development of new
anti-Trypanosoma cruzi agents Curr Drug Targets 200910(3)212-31
Moreno M DAvila D A Silva MN Galvao LM Macedo AM Chiari E et al Trypanosoma
cruzi benznidazole susceptibility in vitro does not predict the therapeutic outcome of human
Chagas disease Mem Inst Oswaldo Cruz 2010105(7)918-24
Mosmann T Rapid colorimetric assay for cellular growth and survival application to
proliferation and cytotoxicity assays J Immunol Methods 198365(1-2)55-63
ODaly JA Serrano LE Rodriguez MB Free aminoacid pool and proteolytic enzymes in
Trypanosoma cruzi cultured in vitro Int J Parasitol 198313(5)433-40
Paveto C Pereira C Espinosa J Montagna AE Farber M Esteva M et al The nitric oxide
transduction pathway in Trypanosoma cruzi J Biol Chem 1995270(28)16576-9
Pereira C Paveto C Espinosa J Alonso G Flawia MM Torres HN Control of Trypanosoma
cruzi epimastigote motility through the nitric oxide pathway J Eukaryot Microbiol
199744(2)155-6
Pereira CA Alonso GD Ivaldi S Silber A Alves MJ Bouvier LA et al Arginine
metabolism in Trypanosoma cruzi is coupled to parasite stage and replication FEBS Lett
2002526(1-3)111-4
Pereira CA Alonso GD Ivaldi S Silber AM Alves MJ Torres HN et al Arginine kinase
overexpression improves Trypanosoma cruzi survival capability FEBS Lett 2003554(1-
2)201-5
Pereira CA Alonso GD Paveto MC Flawia MM Torres HN L-arginine uptake and L-
phosphoarginine synthesis in Trypanosoma cruzi J Eukaryot Microbiol 199946(6)566-70
Pereira KS Schmidt FL Guaraldo AM Franco RM Dias VL Passos LA Chagas disease as
a foodborne illness J Food Prot 200972(2)441-6
Rassi A Jr Rassi A Little WC Chagas heart disease Clin Cardiol 200023(12)883-9
Rassi A Marin-Neto JA Chagas disease The Lancet 2010375(9723)1388-402
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

66
Rohloff P Montalvetti A Docampo R Acidocalcisomes and the contractile vacuole complex
are involved in osmoregulation in Trypanosoma cruzi J Biol Chem 2004279(50)52270-81
Rohloff P Rodrigues CO Docampo R Regulatory volume decrease in Trypanosoma cruzi
involves amino acid efflux and changes in intracellular calcium Mol Biochem Parasitol
2003126(2)219-30
Rojas RLG Anaacutelise de Transporte de Glutamato e GABA em epimastigotas de Trypanosoma
cruzi Satildeo Paulo Universidade de Satildeo Paulo 2007
Schmunis GA Epidemiology of Chagas disease in non-endemic countries the role of
international migration Mem Inst Oswaldo Cruz 2007102(1)75-85
Senkovich O Bhatia V Garg N Chattopadhyay D Lipophilic antifolate trimetrexate is a
potent inhibitor of Trypanosoma cruzi prospect for chemotherapy of Chagas disease
Antimicrob Agents Chemother 200549(8)3234-8
Silber A Tonelli R Martinelli M Colli W Alves M Active transport of L-proline in
Trypanosoma cruzi J Eukaryot Microbiol 200249(6)441-6
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Colli W Ulrich H Alves MJ Pereira CA Amino acid metabolic routes in
Trypanosoma cruzi possible therapeutic targets against Chagas disease Curr Drug Targets
Infect Disord 20055(1)53-64
Silber AM Rojas RL Urias U Colli W Alves MJ Biochemical characterization of the
glutamate transport in Trypanosoma cruzi Int J Parasitol 200636(2)157-63
Silber AM Tonelli RR Lopes CG Cunha-e-Silva N Torrecilhas AC Schumacher RI et al
Glucose uptake in the mammalian stages of Trypanosoma cruzi Mol Biochem Parasitol
2009168(1)102-8
Smirlis D Duszenko M Ruiz AJ Scoulica E Bastien P Fasel N et al Targeting essential
pathways in trypanosomatids gives insights into protozoan mechanisms of cell death Parasit
Vectors 20103107
Smirlis D Soteriadou K Trypanosomatid apoptosis Apoptosis without the canonical
regulators Virulence 20112(3)253-6
Soeiro MN de Castro SL Trypanosoma cruzi targets for new chemotherapeutic approaches
Expert Opin Ther Targets 200913(1)105-21
Sylvester D Krassner SM Proline metabolism in Trypanosoma cruzi epimastigotes Comp
Biochem Physiol B 197655(3B)443-7
67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4

67
Tonelli RR Silber AM Almeida-de-Faria M Hirata IY Colli W Alves MJ L-proline is
essential for the intracellular differentiation of Trypanosoma cruzi Cell Microbiol
20046(8)733-41
Turrens JF Oxidative stress and antioxidant defenses a target for the treatment of diseases
caused by parasitic protozoa Mol Aspects Med 200425(1-2)211-20
Tyler KM Engman DM The life cycle of Trypanosoma cruzi revisited Int J Parasitol
200131(5-6)472-81
Urbina JA Docampo R Specific chemotherapy of Chagas disease controversies and
advances Trends Parasitol 200319(11)495-501
Urbina JA Vivas J Visbal G Contreras LM Modification of the sterol composition of
Trypanosoma (Schizotrypanum) cruzi epimastigotes by delta 24(25)-sterol methyl transferase
inhibitors and their combinations with ketoconazole Mol Biochem Parasitol 199573(1-
2)199-210
Urbina JA Chemotherapy of Chagas disease Curr Pharm Des 20028(4)287-95
Urbina JA Specific chemotherapy of Chagas disease relevance current limitations and new
approaches Acta Trop 2010115(1-2)55-68
Wilkinson SR Kelly JM The role of glutathione peroxidases in trypanosomatids Biol Chem
2003 Apr384(4)517-25
World Health Organization ndash WHO Chagas disease (American Trypanosomiasis) 2012
[updated 2012 cited 2013 fevereiro] Available from wwwwhoint
Xia P Chen HS Zhang D Lipton SA Memantine preferentially blocks extrasynaptic over
synaptic NMDA receptor currents in hippocampal autapses J Neurosci 2010
1830(33)11246-50
Zeledon R Comparative physiological studies on four species of hemoflagellates in culture
II Effect of carbohydrates and related substances and some amino compounds on the
respiration J Parasitol 196046541-51
Zingales B Andrade SG Briones MR Campbell DA Chiari E Fernandes O et al A new
consensus for Trypanosoma cruzi intraspecific nomenclature second revision meeting
recommends TcI to TcVI Mem Inst Oswaldo Cruz 2009104(7)1051-4