establecimiento de protocolos de micropropagaciÓn …
TRANSCRIPT

ESTABLECIMIENTO DE PROTOCOLOS DE
MICROPROPAGACIÓN E IDENTIFICACIÓN DE BIOPRODUCTOS EN ESPECIES NATIVAS
DE ALTO VALOR ECOLÓGICO DEL NORTE
CHICO
Este proyecto fue financiado por el Fondo de Investigación del Bosque Nativo, proyecto n° 037/2011
Investigador responsable
Dr. Cristian Ibáñez, Departamento de Biología, Universidad de La Serena
Investigador asociado
MSc. Fabiola Jamett, Departamento de Química, Universidad de La Serena
Octubre 2013

2
ÍNDICE
1. INTRODUCCIÓN 6
1.1 Chile, hábitat natural de una gran biodiversidad vegetal 6
1.2 Biodiversidad en las zonas áridas y semiáridas de Chile 6
1.3 Especies de interés 7
1.3.1 Garra de León (Leontochir ovallei) 7
1.3.2 Lucumillo (Myrcianthes coquimbensis) 8
1.3.3 Algarrobo dulce (Prosopis flexuosa) 8
1.3.4 Algarrobo (Prosopis chilensis) 8
1.4 Propagación vegetativa: una alternativa para resguardar el patrimonio
genético nativo 8
1.5 Biosprospección de compuestos metabólicos: potenciando un uso alternativo de la biodiversidad nativa 10
1.6 Perfiles metabolómicos 11
1.7 Propósito de la investigación 12
2. OBJETIVOS 14
3. METODOLOGÍA 16
3.1 Propagación in vitro de tipo organizado 16
3.2 Propagación vegetativa por esquejes en cama de propagación 17
3.3 Propagación por semillas 17
3.4 Tratamiento Preliminar de las Muestras 19
3.5 Análisis de Humedad total y remanente después del secado inicial
antes de la molienda 19
3.6 Análisis de Cenizas 19
3.7 Determinación Analítica del extracto etéreo (EE) 19

3
3.8 Análisis de la proteina bruta 19
3.9 Determinación de azúcares totales, azúcares reductores y no reductores 19
3.10 Análisis de Fibra. 21
3.11 Análisis de los componeetes minerales 21
3.12 Determinación de Boro 21
3.13 Determinación de Fósforo 22
3.14 Polifenoles Totales 22
3.15 Perfil de Ácidos Grasos 22
3.16 Identificación de metabolitos secundarios por TLC. 23
3.17 Preparación del infuso agua 23
3.18 Prueba de susceptibilidad antimicrobiana en los extractos de hojas. 25
3.19 Análisis Estadísticos. 25
4. RESULTADOS 26
4.1 Parte Propagación
4.1.1 Garra de León (Leontochir ovallei) 26
4.1.2 Lucumillo (Myrcianthes coquimbensis) 26
4.1.3 Algarrobo dulce (Prosopis flexuosa) 26
4.1.3 Algarrobo (Prosopis chilensis) 26
4.2 Parte Fitoquímica
4.2.1 Garra de León (L. ovallei) 27
4.2.2 Lucumillo (M. coquimbensis) 29
4.2.3 Algarrobo dulce (P. flexuosa) 30
4.2.4 Algarrobo (P. chilensis) 32
5. CONCLUSIONES Y RECOMENDACIONES 33
6. PRODUCTOS OBTENIDOS 37

4
6.1 Actividades de difusión 37
7. BIBLIOGRAFÍA 38
8. ANEXOS 42
ANEXO n° 1. Colecta de semillas de Garra de León (L. ovallei) 42
ANEXO n° 2. Protocolo de Germinación in vitro de Garra de León (L. ovallei) 46
ANEXO n° 3: Protocolo de multiplicación de rizomas de Garra de León (L. ovallei) 50
ANEXO n° 4: Protocolo de germinación semillas de Lucumillo (M. coquimbensis) 52
ANEXO n° 5: Protocolo de propagación ex vitro por esquejes en Lucumillo (M. coquimbensis) 56
ANEXO n° 6: Protocolo de propagación in vitro de Lucumillo (M. coquimbensis) 62
ANEXO n° 7: Protocolo de germinación semillas de Algarrobo dulce (P. flexuosa) 67
ANEXO n° 8: Protocolo de propagación ex vitro por esquejes de Algarrobo dulce (P. flexuosa) 70
ANEXO n° 9: Protocolo de propagación in vitro de Algarrobo dulce (P. flexuosa) 71
ANEXO n° 10: Protocolo de germinación semillas de Algarrobo (P. chilensis) 75
ANEXO n° 11: Protocolo de propagación ex vitro por esquejes de Algarrobo (P. chilensis) 77
ANEXO n° 12: Protocolo de propagación in vitro de Algarrobo (P. chilensis) 80
ANEXO n° 13: Materiales usados en investigación 82
ANEXO n° 14: Poster 1 presentado en la VII Reunión de Biología Vegetal. Pucón
2012. 84
ANEXO n° 15: Poster 2 presentado en la VII Reunión de Biología Vegetal. Pucón 2012. 85
ANEXO n° 16: Poster 3 presentado en la VII Reunión de Biología Vegetal. Pucón 2012. 86
ANEXO n° 17: Programa II Encuentro temático del FIBN, Iquique 2013. 87
ANEXO n° 18: Certificado de aceptación trabajo oral Cristian Ibáñez. Tercer Congreso de Flora Nativa. Santiago 2013. 88

5
ANEXO n° 19: Certificado de aceptación trabajo oral Fabiola Jamett. Tercer
Congreso de Flora Nativa. Santiago 2013. 89
ANEXO n° 20: Certificado Poster 4 presentado al Tercer Congreso de Flora Nativa. Santiago 2013. 90
ANEXO n° 21: Poster 5 presentado en la VIIU Reunión de Biología Vegetal. Pucón 2013. 91
9. ANEXOS - TABLAS 92
ANEXO Tabla 1. Esquema y posterior detalle de los tratamientos de germinación de semillas de L. ovallei probados durante esta investigación. 92

6
1. INTRODUCCIÓN
1.1 Chile, hábitat natural de una gran biodiversidad vegetal. La sorprendente
variabilidad genética de las plantas, ha posibilitado que éstas conquisten hábitat de la
más diversa índole y complejidad, capaces de adoptar y adaptar formas de vida en
ecosistemas dinámicos, demandantes y a veces, muy limitantes. Como resultado, se ha
generado una flora que desconoce fronteras y que está constituida por especies con
características únicas y diferenciadoras (CBD, 2006; Taiz & Zeiger, 2007). Chile, por su
gran variedad de climas y suelos, está constituido por una flora nativa, que si bien no es
tan abundante en cantidad comparado a sus países vecinos, sí es fértil en endemismos
de particular interés ecológico que la hacen única (Squeo et al, 2001; Lazo et al, 2008;
Santibáñez et al, 2008). Además, la interacción entre la herencia genética de esta flora
nativa con su entorno ambiental ha dado origen a versiones fenotípicas que han llamado
la atención de la sociedad tanto por su valor ornamental como por sus características
aromáticas, sus propiedades medicinales o simplemente por su cualidades nutritivas o
funciones ecológicas. Lamentablemente, muchas de estas especies se encuentran
fuertemente amenazadas por las actividades humanas, por el dinamismo secular propio
del medioambiente o por el escaso conocimiento que tenemos de ellas.
En 1992 se creó la Convención Internacional para la Diversidad Biológica (CBD) que
reúne a 193 países de todo el orbe (CBD, 2011). Su principal objetivo es trabajar
conjuntamente para evitar la pérdida de hábitat, de especies y de genes, contribuyendo
así a mitigar la pobreza y mejorar la calidad de vida de todas las formas de vida que
habitan la tierra (CBD, 2006). Dentro de sus programas de protección de la biodiversidad
mundial, la CBD señala a las zonas áridas y semiáridas del mundo como zonas
prioritarias de conservación debido a la fragilidad de sus ecosistemas, su riqueza natural
y cultural (CBD, 2011). Los principales factores que amenazan las zonas áridas son el
pastoreo desmesurado, el agotamiento del suelo por el uso excesivo de fertilizantes, el
consumo excesivo del siempre escaso recurso hídrico, la pérdida de la estructura del
suelo, la pérdida de especies leñosas para producir leña o carbón, y sobre todo, la
deforestación a la que son sometidos los terrenos para habilitar zonas agrícolas (CBD,
2006). Recientemente, un estudio realizado por el Centro de Información de Recursos
Naturales (CIREN) determinó que el 49.1% (36.8 millones de hectáreas) del territorio
nacional se encuentra con algún grado de erosión (CIREN, 2011). Entre éstas, la Región
de Coquimbo presenta un alarmante 84% de sus suelos erosionados, de los cuales un
65.3% se encuentra en la categoría de erosión “severa” y “muy severa” (CIREN, 2011).
1.2 Biodiversidad en las zonas áridas y semiáridas de Chile. En Chile, las zonas
áridas y semiáridas están situadas aproximadamente entre los 18º y 33º Latitud Sur
(Astaburuaga, 2004) y corresponden al tipo macro-bioclimático tropical que incluye las
variantes bioclimáticas tropical pluviestacional, antitropical, xérico, desértico e
hiperdesértico (Amigo & Ramírez, 1998). Por lo general, las zonas áridas y semiáridas se
caracterizan por tener veranos largos y calurosos, poseer suelos con bajo contenido en
materia orgánica, pedregosos y fácilmente erosionables por lluvias que, aunque
esporádicas, suelen dañar bastante la estructura del suelo (Luebert & Priscoff, 2006). A
pesar de tener una escasez importante de agua, la disponibilidad de ésta en la zonas
áridas y semiáridas es posible caracterizarla en componentes espaciales y temporales
(Stoll et al, 2011). Existen dos gradientes: a) latitudinal, que influyen en la cantidad de
agua (van Husen 1967; Luebert & Pliscoff 2006) y b) altitudinal, que diversifica las
fuentes de agua (neblina, lluvia, agua subterránea, nieve y glaciares (Favier et al, 2009).
Además de esta excepcional heterogeneidad espacial, el ciclo hidrológico también es muy
variable en el tiempo: a) ciclos diurnos de evaporación y evapotranspiración (Kalthoff et
al, 2006), b) ciclos estacionales de precipitación (vientos alisios – lluvias de verano en el
altiplano entre 13º y 27º S, vientos del oeste – lluvias de invierno entre 27º y 38ºS)
(Garreaud et al, 2009), c) neblina costera con ciclos diurnos y anuales (maximo en
primavera) (Garreaud et al, 2008), d) ciclos interanuales como El Niño - Southern

7
Oscillation (ENSO) (Gutiérrez et al, 2007; Garreaud et al, 2009; Gutiérrez et al, 2010).
Probablemente sea esta excepcional heterogeneidad temporal y espacial del recurso
hídrico lo que ha posibilitado la presencia de un interesante conjunto de especies
xerofíticas y otras de tipo esclerófilo que han logrado exitosamente adaptarse a las duras
condiciones áridas (Donoso et al, 1993; Squeo et al, 2001; Luebert & Priscoff, 2006).
El Norte Chico de Chile (Regiones de Atacama y Coquimbo) es una zona de extrema
riqueza en plantas vasculares y con un elevado porcentaje de endemismos tanto
regionales como nacionales (Squeo et al, 2001, 2008). Entre las especies que dominan
los paisajes áridos del Norte Chico figuran especies leñosas del género Porlieria spp.,
Caesalpinia spp., Cordia decandra, Monttea spp., Vasconcellea spp., Prosopis spp., varios
tipos de cactáceas como Erioscyse spp., Eulychnia spp., Opuntia spp., Copiapoa spp., y
herbáceas como Fabiana spp., Leucocoryne spp., Calceolaria spp. y Adesmia spp. (Squeo
et al, 2001, 2008). Muchas de estas plantas cumplen un importante rol para las
comunidades que habitan las zonas áridas pues tienden a ser fuente de alimento para
ellas, su ganado y la fauna silvestre del lugar. Pueden además actuar como contenedoras
de la erosión, protegiendo contra la desertificación y contribuyendo a contrarrestar la
sequedad característica del desierto y las zonas áridas (Pardos, 1984). No obstante
aquello, la biodiversidad de las zonas áridas y semiáridas se está viendo muy afectada
por la creciente intervención humana (Squeo et al, 2001). La sobreexplotación de
recursos naturales, alteración y destrucción de los hábitat son unos de los factores mas
importantes. Contribuye también la escasa conciencia sobre la conservación del ambiente
y los beneficios que ello trae (por ejemplo, los servicios ecosistémicos) a las
comunidades, lo cual genera un manejo poco sustentable de los recursos naturales,
provocando así la degradación de la diversidad biótica y abiótica. La degradación y/o
pérdida de biodiversidad puede tener serias consecuencias en la producción de alimentos
y sustentabilidad medioambiental, además de tener efectos globales a nivel ecosistémico
y la continuación de prácticas no sustentables (Sala et al, 2000; Chapin III et al, 2000).
La propuesta presentada tuvo como objetivo enfrentar este problema global en una
escala regional, considerando tres aspectos importantes de la conservación de
biodiversidad:
(1) generación de conocimiento científico,
(2) sensibilización y capacitación de la población local, y
(3) manejo y usos alternativos de la biodiversidad.
1.3 Especies de interés
Para abordar estos objetivos, se eligió un grupo de cuatro especies nativas del Norte
Chico, que, a pesar de su innegable valor ecológico e interesante potencial económico, se
encuentran pobremente estudiadas en nuestro país. Estas especies son:
1.3.1 Garra de León (Leontochir ovallei)
Descripción: Planta rizomatosa con tallos herbáceos, rastreros, rodeados de hojas
oblongo-lanceoladas (hasta 8 cm de largo y 4,5 cm de ancho). Inflorescencia con flores
rojas o raro amarillas, agrupadas de 18-20 flores en una gran cabezuela globosa de 8-10
cm de diámetro. Cada flor tiene 6 tépalos, espatulados en dos series. El fruto es una
cápsula de 3-4 lados, con los tépalos persistentes. En el año 2000 se describe la variedad
L. ovallei var. luteus M. Muñoz, de flores amarillas, que es muy rara entre la población de
flores rojas (Muñoz, 2000).
Área de distribución: Región de Atacama, endémica en la costa e interior entre Carrizal
Bajo (28° 05’ -71° 09’ W) y Totoral (27° 54’ S/ 70° 57’ W) (Muñoz, 1966).
Estado de conservación: En Peligro (Ravenna et al, 1998), Amenazada (Squeo et al,
2008).
Amenazas: Falta de regeneración natural (colecta de las flores por los lugareños y
turistas durante los períodos de desierto Florido), baja productividad de semillas, planta

8
consumida por guanacos y cabras, destrucción de hábitat, extracción intensiva para
comercialización interna y exportación (Ravenna et al, 1998).
Comentarios: Su reproducción con métodos de micropropagación puede disminuir
considerablemente la presión de colecta sobre sus poblaciones naturales.
1.3.2 Lucumillo (Myrcianthes coquimbensis)
Descripción: Arbusto de copa globosa y densa, que presenta alturas de 1 hasta 1,5 m,
las hojas perennes, simples, enteras y aromáticas. Las flores son hermafroditas y
blancas, compuestas en pequeños grupos. El fruto es una baya carnosa, de un color
intenso rojo en la madurez.
Área de distribución: Región de Coquimbo, entre Barrancones (limite norte) y Las Tacas
(límite sur). La extensión es una delgada franja de aprox. 83 km de largo en la costa y
aprox. 3 km hacia el interior (García et al, 2011).
Estado de conservación: En Peligro (Hechenleitner et al, 2005), Amenazada (Squeo et al,
2001).
Amenazas: Falta de regeneración natural (baja productividad de semillas, depredación de
semillas), destrucción de hábitat y extracción de hojarasca.
Comentarios: La especie presenta problemas de conservación en bancos de semilla,
donde ésta no puede ser mantenida viable (información verbal. Pedro León, Banco de
Germoplasma-INIA Intihuasi).
1.3.3 Algarrobo dulce (Prosopis flexuosa)
Descripción: Árbol o arbusto, con máx. 8 m de altura. Hojas compuestas, flores en
espigas de 20 a 30 cm de largo y de un color amarillo-verdoso, melífera. Los frutos son
vainas de color oscuro, planas, cinturadas parecido a un rosario y largas (aprox. 12 cm).
Área de distribución: Región de Atacama y Coquimbo, muy escasa, muy pocas
poblaciones conocidas. Además, varias lugares de colectas históricas han desaparecido
por actividad humana (cultivos de vid) y movimiento de dunas (cercanía de Copiapó).
También habita en Argentina y Bolivia (Zuloaga et al, 2008).
Estado de conservación: En Peligro (Squeo et al, 2001, 2008).
Amenazas: Falta de regeneración natural (depredación de semillas por cabras y
brúquidos), destrucción de hábitat, extracción de para producción de leña y carbón.
Comentarios: Taxonómicamente su distinción de P. flexuosa es muy complicada y sólo se
puede realizar con certeza en los frutos. Además, ambas especies forman híbridos de
manera natural, lo que dificulta aún más su identificación.
1.3.4 Algarrobo (Prosopis chilensis)
Descripción: Árbol caducifolio en su límite sur de distribución y siempreverde hacia el
límite norte, con aprox. 10 m de altura. Hojas compuestas, flores en espigas de 20 a 30
cm de largo y de un color amarillo-verdoso, melífera. Los frutos son vainas de color
claro, planas, de borde recto y largas (aprox. 12 cm).
Área de distribución: Región de Atacama hasta Región del Libertador O´Higgins,
preferentemente en suelos arenosos en el interior y la pre-cordillera. Acompaña la
vegetación local, en ocasiones forma bosquetes. También se encuentra en Argentina,
Perú, Bolivia y Paraguay (Zuloaga et al, 2008).
Estado de conservación: En Peligro (Squeo et al, 2001, 2008).
Amenazas: Falta de regeneración natural (depredación de semillas por cabras y
brúquidos), destrucción de hábitat, extracción para producir leña y carbón.
Comentarios: Taxonómicamente su distinción de P. flexuosa es muy complicada y sólo se
puede realizar con certeza con los frutos. Además, ambas especies forman híbridos de
manera natural, lo que dificulta aún más su identificación.
1.4 Propagación vegetativa: una alternativa para resguardar el patrimonio
genético nativo. No cabe duda que el método mas extendido para preservar la
existencia de una especie son sus semillas (Taiz & Zeiger, 2007). En ellas no solo se
transporta una parte importante de la información genética de la especie sino también se
adosan una serie de mecanismos que mantienen viable la semilla por un largo periodo de

9
tiempo y mecanismos que permiten sincronizar el proceso de germinación con las
condiciones mas favorables para que aquello ocurra (Engelman, 1997; Razdan &
Cocking, 1997; Taiz & Zeiger, 2007). En algunas plantas, sin embargo, la germinación
por semillas puede no ser un proceso sencillo e incluso llegar a ser inviable. Existen
especies cuyas semillas son incapaces de ser almacenadas puesto que pierden
rápidamente su humedad interna o porque no toleran las condiciones frías de
almacenamiento (Barbedo & Cicero, 2000). Estas semillas, conocidas como
recalcitrantes, son típicas de zonas tropicales y subtropicales (como el Lucumillo)
(Pammenter et al, 1994; Barbedo & Cicero, 2000). Existen también otro grupo de plantas
que carecen de semillas o éstas de encuentras atrofiadas o sufren de germinación
precoz, generando patrones anormales en el desarrollo. Bananas y algunos tipos de uvas
se encuentran dentro de este grupo de plantas (Finkelstein & Crouch, 1984). Cuando se
presenta alguna de las situaciones anteriores, la reproducción vegetativa por esquejes o
a través de injertación, ha sido la forma tradicional de propagación utilizada (Matthews,
1999). Sin embargo, no siempre es factible desarrollar este tipo de propagación, ya sea
por factores endógenos de las plantas, por requerimientos de asociación con
microorganismos específicos o porque fito-sanitariamente no son un material vegetal
adecuado de propagar (Jain & Häggman, 2007). Afortunadamente, la biotecnología de
plantas y sus áreas afines, han contribuido con una serie de métodos que permiten
resguardar los acervos genéticos nativos mediante la propagación masiva de una planta
a partir de una muestra muy pequeña de ella en un medio estéril que contiene todo lo
necesario para el crecimiento de la planta. A este tipo de micropropagación se le ha
llamado cultivo in vitro y ha sido amplia y exitosamente utilizado en especies herbáceas
y leñosas (Conger 1981; Caro et al, 2002; Gomes & Canhoto, 2003; Sabja et al, 2008).
Entre las ventajas del cultivo in vitro está la producción de plantas libre de virus y otras
enfermedades, la propagación clonal de individuos de características superiores (árboles
elite), la aceleración de los programas de mejoramiento al permitir una rápida
regeneración de las plantas, eliminación de factores de inhibición endógena de ciertas
semillas, lo cual aumenta las posibilidades de regeneración exitosamente. Entre las
desventajas está la posibilidad latente de contaminación del cultivo causada por hongos,
bacterias u otros microorganismos. Afortunadamente, la utilización de campanas de flujo
laminar que expulsan aire desde la campana al exterior reducen notablemente que
esporas u otros agentes patógenos contaminen los cultivos al interior de ellas. La
vitrificación o endurecimiento de ciertas partes de la planta es otra de las desventajas. La
solución a este problema se realiza tomando las partes de la plántula no vitrificadas y
que son nuevamente propagadas (Dodds, 1991).
En general los protocolos de micropropagación in vitro incluyen seis etapas principales
(Dodds, 1991):
1) Obtención del explante o muestra a micropropagar (hoja, tallo, raíz, ápice, etc).
2) Establecimiento del cultivo en un medio semi-sólido rico en nutrientes.
3) Multiplicación de explantes establecidos y almacenamiento del cultivo en condiciones
controladas.
4) Recuperación de cultivo viables.
5) Regeneración de las plantas a través de sucesivas multiplicaciones.
6) Fases de aclimatación in vitro y ex vitro (invernaderos).
En plantas superiores, el cultivo in vitro se puede desarrollar a través de un cultivo
organizado y a través de un cultivo no organizado (De Fossard, 1977). En el tipo
organizado, se cultivan plantas completas o casi completas por medio de embriones o
semillas. Este tipo de cultivo in vitro es similar al cultivo vegetativo pues incluye la
propagación de especies vegetales vía esquejes, estolones o yemas, conduciendo a una
descendencia idéntica al material vegetal original. El tipo no organizado, por su parte,
involucra el cultivo de células o tejidos que se aíslan desde una zona organizada de la
planta (por ejemplo, un trozo de hoja), la cual es desdiferenciada con el uso de
hormonas vegetales. Esta acción da origen a un masa embriogénica o callo, el cual es

10
dividido en múltiples fragmentos, los cuales mediante una combinación de hormonas de
crecimiento permiten la transformación del callo en plantas completas, idéntica a la
original.
Algunos ejemplos del uso de técnicas de micropropagación in vitro lo tenemos en nuestro
país. Iniciativas del Núcleo Milenio lideradas por científicos de la Pontificia Universidad
Católica de Chile, han realizado con éxito la micropropagación del Toromiro (Shopora
toromiro), un árbol nativo de la Isla de Pascua y que lamentablemente había sido
declarado como extinto en 1978.
(http://www.iniciativamilenio.cl/centros/actividades_det.php?id=1109). Según lo
informado por este grupo de investigadores, la propagación clonal de Toromiros ya
habría alcanzado las 700 plantas y se espera contar con 5 mil ejemplares dentro de los
próximos años. Estas árboles serán reinsertados en su hábitat natural de forma paulatina
contribuyendo a rescatar el patrimonio genético propio de la Isla y aportará a
reestablecer el equilibrio ecológico de la misma. Otros ejemplos de micropropagación de
especies leñosas con problemas de conservación o de interés productivo incluye la Palma
Chilena (Jubaea chilensis), Lingue (Persea lingue), Queule (Gomortega Keule), Luma del
Norte (Legrandia concinna) y Raulí (Nothofagus alpina (Infante, 1989; Calderón-Baltierra
et al, 1993; Sánchez-Olate et al, 2004; Uribe-Moraga & Cifuentes, 2004; Cob et al,
2010). Esto demuestra que aunque no ausente de complejidades, la propagación in vitro
de especies leñosas es posible de realizar.
Como previamente se indicó, el cultivo in vitro se utiliza preferentemente en especies
donde la propagación organizada es ineficiente o prácticamente nula. Sin embargo, la
propagación in vitro no carece de complejidades y varios son los factores que interceden
para lograr una exitosa propagación. Su estandarización es crucial para lograrlo (Conger,
1981). Entre ellos, el genotipo de la planta parece ser clave. Plantas dicotiledóneas
suelen presentar una mejor regeneración in vitro que plantas monocotiledóneas (Zhao et
al, 2009). Idéntica situación se ha observado cuando se comparan angiospermas con
gimnospermas, pues éstas últimas suelen presentar menor capacidad de regeneración in
vitro que las primeras (Deb & Tandom, 2004). La edad de la planta y su estadio de
desarrollo también parecen constituir un factor importante al momento de implementar
esta técnica. Los tejidos jóvenes, no lignificados, por lo general son más apropiados para
el cultivo in vitro, sin embargo, la capacidad regenerativa de algunas especies,
independiente de la edad de éstas, aumenta considerablemente en etapas posteriores de
desarrollo, por ejemplo, durante el periodo de floración (Pierik & Steegmans, 1986).
Otros factores a considerar cuando se desea regenerar un planta de manera in vitro son
el estado fisiológico de la planta, el cual tiene un fuerte efecto sobre la división celular y
la regeneración (Zhao et al, 2009); el estado sanitario de la planta también es
importante pues incide en la vigorosidad de la misma (Morini, 2000); la concentración de
reguladores de crecimiento, sacarosa y minerales; y las condiciones usadas de
temperatura, luz, humedad, potencial osmótico, pH son otros aspectos a evaluar (Wang
& Charles, 1991).
Considerando lo anteriormente expuesto, una de las líneas de trabajo que se realizó en
esta propuesta fue estandarizar métodos de cultivos in vitro para las cuatro especies
nativas de alto interés ecológico y que son consideradas prioritarias en los programas
actuales de conservación (Squeo et al, 2001; 2008). El interés responde a la acción
imperante de contar con sistemas eficientes para estudiar, mantener y resguardar el
patrimonio nativo, lo cual a su vez podría generar nuevas estrategias coactivas en pro de
su protección y desarrollo.
1.5 Biosprospección de compuestos metabólicos: potenciando un uso
alternativo de la biodiversidad nativa. El desciframiento de la estructura del ADN en
la década del 50 (Watson & Crick, 1953), supuso un avance cualitativo y cuantitativo- sin
precedentes en la historia de la humanidad- en lo que se refiere a la compresión del
funcionamiento biológico de los organismos (Machuka, 2004). Desde una perspectiva

11
genética, estos avances tuvieron un punto de inflexión importante en el año 2000 cuando
se anunció la secuenciación completa del primer genoma vegetal de una planta. El honor
correspondió a la planta Arabidopsis thaliana (The Arabidopsis Genome Initiative, 2000).
Una año mas tarde, la ciencia dio otro paso gigantesco y anunció la secuenciación del
genoma Humano (Venter Craig et al, 2001). A partir de ese momento, las técnicas y
metodologías no han parado de perfeccionarse y la secuenciación de genomas vegetales,
de animales, de peces, de insectos, de bacterias, de levaduras, prácticamente se ha
vuelto algo rutinario. Actualmente, la base datos pública que reúne la información
genómica ya cuenta con 1000 genomas secuenciados de organismos procarióticos y
aproximadamente 12 genomas vegetales (http://www.ncbi.nlm.nih.gov/genome).
Esta explosiva disponibilidad de información ha dado origen a una nueva era dentro la
genética bautizada como la era “Post-Genómica”. Áreas como la genómica estructural,
genómica evolutiva, epigenómica, transcriptómica, proteómica, metabolómica,
metagenómica, y otras en permanente desarrollo, comparten el objetivo común de
aprovechar y procesar los grandes volúmenes de información genética que hoy es posible
generar, estableciendo correlaciones intra e inter-genoma inimaginables hace un par de
años atrás (Brenner, 2001; Montpetit, 2003). Entre estas nuevas áreas de investigación,
el estudio de los perfiles bioquímicos o metabólicos de las plantas se presenta como una
de las ramas más atractivas e influyentes dentro del actual siglo 21 (Oliver, 2002).
Históricamente, los estudios metabolómicos se centraron en el diagnóstico de patologías
o en la caracterización de cepas bacterianas (Shiloach et al, 2010). Con el correr del
tiempo, su utilización se amplió para describir los perfiles metabólicos de hongos y
plantas aunque su desarrollo se consideraba limitado (Kehr, 2001). Esta situación cambió
radicalmente con el avance tecnológico notable de técnicas y tecnologías destinadas a la
caracterización bioquímica de las plantas como son la Cromatografía de Gases (GC),
Cromatografía liquida (LC), Espectrofotometría de Masas (MS), Electroforesis de
Capilaridad (CE) y Resonancia Magnética Nuclear (NMR). Si a este avance técnico le
unimos el mencionado aumento de la secuenciación de genomas y una asociación
estratégica con poderosas herramientas bioinformáticas, podemos señalar que hoy en día
contamos con herramientas biotecnológicas que nos permiten identificar y caracterizar
masivamente un grupo total de metabolitos presentes en un organismo expuesto a una
condición dada, de una manera mucho más precisa y en un rango de tiempo mucho
menor que hace tan solo 10 años atrás (Moco et al, 2007).
El reto de la metabolómica reside, por lo tanto, en encontrar cambios en las cantidades
de metabolitos que se puedan correlacionar con el estado fisiológico y desarrollo de una
célula, tejido u organismo adaptado a sus condiciones de hábitat particular. Lo es
también, el interés y la capacidad de las Universidades y Centros de Investigación por
adquirir prontamente estas nuevas tecnologías e instrumentación disponible. Como
primer paso para desarrollar la metabolómica en plenitud, es requisito fundamental
contar con perfiles metabolómicos “básicos” que orienten hacia donde debería ser
dirigida la secuenciación de su metaboloma en un sentido amplio y preciso (Schwab,
2003). Una vez conocida la cantidad y calidad de sus componentes bioquímicos, la
naturaleza propia de las plantas hará que se generen nuevas oportunidades excitantes
para aplicar y aprovechar sus bio-productos.
1.6 Perfiles metabolómicos. Los metabolitos se definen como el producto final de un
proceso celular y corresponden a la última respuesta que los sistemas biológicos hacen
de acuerdo a su constitución genética o influencia medioambiental (Oliver, 2002). Los
perfiles metabólicos corresponden a una “radiografía” de lo que está contenido en el
metaboloma de esa planta, es decir, el conjunto de metabolitos totales presentes en un
organismo bajo un cierto tipo de condición biológica, de tiempo o de espacio y la ciencia
encargada de estudiarlos se conoce como metabolómica (Montpetit, 2003).
Los metabolitos se dividen en metabolitos primarios (azúcares, aminoácidos, ácidos
orgánicos, lípidos) y metabolitos secundarios (terpenos, compuestos fenólicos y

12
compuestos que contienen Nitrógeno) (Taiz & Zeiger, 2007). En el caso de las plantas,
los metabolitos tanto primarios comos secundarios son muy importantes pues actúan en
respuestas al ataque de patógenos, colaboran a contrarrestar condiciones de estrés
abiótico, contribuyen a mantener el color, sabor y olor tanto de flores como de frutos,
potencian o arruinan la calidad de algunos productos y finalmente, son los constituyentes
básicos de muchas de las propiedades químicas que hacen que una planta sea o no de
interés popular (Oliver, 2002). En papa (Solanum tuberosum), se ha logrado caracterizar
los perfiles metabólicos específicos de seis fases particulares del desarrollo de esta planta
(Shephard et al, 2010). La información generada ha permitido no sólo entender el ciclo
de vida de Solanum sino también ha impulsado el diseño de nuevas estrategias para su
mejoramiento nutricional. Otro ejemplo notable ocurre con el aceite de oliva. Desde hace
un tiempo que es conocida la presencia del oleocanthal, un metabolito secundario con
efecto antiinflamatorio presente exclusivamente en el aceite de oliva “extra virgen” y que
inhibe la actividad de las enzimas de la ciclooxigenasa (COX), causante de la inflamación
y cuya actividad es similar a la realizada por el ibuprofeno (Beauchamp et al, 2005). Las
tremendas propiedades de los ácidos grasos del aceite de oliva no terminan ahí.
Recientemente, se ha comprobado que el oleocanthal también ayudaría a contrarrestar la
enfermedad de Alzheimer (Pitt et al, 2010). Otro enfoque acerca de cómo los perfiles
metabolómicos pueden contribuir al conocimientos de las especies, ocurre en estudios
metabólicos comparativos realizados en leguminosas. Aquí, comunidades que contenían
plantas altas-leguminosas (AL), plantas altas-no leguminosas (ANL), plantas bajas-
leguminosas (BL) y plantas bajas-no leguminosas (BNL), encontraron que la carencia de
nutrientes necesarios para sintetizar ciertos compuestos primarios podría actuar como un
estimulador de la diversidad de las plantas (Scherling, 2010). Así también, existen
innumerables ejemplos que demuestran que los metabolitos o compuestos naturales
presentes en una planta específica, pueden verse potenciados por las características
particulares de su hábitat. Es así como el déficit hídrico en vides aumenta el contenido de
fenoles y antocianinas, los cuales tienen un impacto positivo sobre la calidad final del
vino (Ferreyra & Sellés, 1997). Idéntico tratamiento hídrico se traduce en un incremento
de los azúcares contenido en las bayas de vides de mesa, mejorando su apreciación final
por el consumidor (Peacock & Dokoozlian, 1997). La sequía también se ha visto que
aumenta el contenido final de polifenoles y tocoferoles en el aceite de oliva, lo cual
incrementa el picor y el ardor que son dos de las características que mas contribuyen a
incrementar la calidad final de él (Troncoso et al, 2006). Por su parte, caracterizaciones
realizadas en plantas medicinales demuestran la existencia de altas cantidades de
metabolitos primarios y secundarios que pueden tener efectos gastroprotectores
(antocianinas), antitumorales (diterpenos) y nutricionales (aminoácidos esenciales)
(Schmeda-Hirschmann et al, 1992; Schmeda-Hirschmann et al, 1993; Shirley et al, 2007;
Theoduloz et al, 2009). Todos estos antecedentes refuerzan la idea que los compuestos
metabólicos de las plantas pueden impactar significativamente tanto la industria
alimenticia como aquella con fines fármaco-ecológicos. Sin embargo, a pesar de los
esfuerzos por conocer la estructura química que les constituye, su alcance es aún
insuficiente, limitado a ciertas especies, o como en la mayoría de los casos, inexistente.
Si a esto le sumamos que hay especies ecológicamente importantes cuyas propiedades
químicas se desconocen, entonces nuestra labor y políticas de conservación deberían
verse potenciadas con el fin de preservar un material que ecológica y genéticamente
podría ser relevante para el desarrollo del país.
1.7 Propósito de la investigación. Las regiones de Atacama y Coquimbo pertenecen al
llamado “Hot spot” chileno de biodiversidad mundial (Arroyo et al, 2004). En ellas
conviven un número importante de especies nativas de excepcional belleza e incalculable
valor ecológico. No obstante aquello, las escasas precipitaciones, suelos pobre en
nutrientes, procesos severos de desertificación, elevada intervención antropogénica y
presencia de ciclos climáticos (ENSO) que alteran la biodinámica de sus ecosistemas, ha
puesto en peligro de conservación un número importante de especies nativas (Luebert &
Priscoff, 2006). Con el fin de contribuir a su protección e identificación de nuevos usos
biotecnológicos, esta propuesta consideró la aplicación de técnicas de micropropagación

13
in vitro y caracterización bioquímica de cuatro especies nativas de alto valor ecológico
adaptadas a las zonas áridas y semiáridas de Chile. Estas especies fueron Garra de Léon
(Leontochir ovallei), Lucumillo (Myrcianthes coquimbensis), Algarrobo Dulce (Prosopis
flexuosa) y Algarrobo (Prosopis chilensis). Para lograr la conservación de este material la
propuesta consideró la estandarización de protocolos de propagación sexual (por
semillas) y propagación clonal in vitro de tipo organizado (a partir de tejidos completos
como puede ser yemas, hojas, tallos), ambos con el fin de sustentar posibles programas
de forestación y reforestación con éstas cuatro especies nativas. Esto último de gran
importancia y necesarios de implementar luego de conocer los preocupantes niveles de
erosión de los suelos de Chile y en especial, los de la Región de Coquimbo (CIREN,
2011).

14
2. OBJETIVOS
2.1 Objetivo General
Evaluar y resguardar el patrimonio genético de plantas nativas de zonas áridas y
semiáridas mediante el establecimiento de protocolos de propagación vegetativa
de tipo in vitro y determinación de compuestos naturales presentes en el
metaboloma de hojas, frutos o raíces.
2.2 Objetivos Específicos
1. Estandarizar protocolos de propagación por semillas, por esquejes y de tipo in
Vitro para Garra de Léon (Leontochir ovallei), Lucumillo (Myrcianthes
coquimbensis), Algarrobo dulce (Prosopis flexuosa) y Algarrobo (Prosopis
chilensis).
2. Estandarizar protocolos para identificar y cuantificar -mediante herramientas
metabolómicas- compuestos metabólicos naturales presentes en hojas, frutos o
raíces de Garra de Léon (Leontochir ovallei), Lucumillo (Myrcianthes
coquimbensis), Algarrobo dulce (Prosopis flexuosa) y Algarrobo (Prosopis
chilensis).
3. Generar un registro de los metabolitos presentes en Garra de Léon (Leontochir
ovallei), Lucumillo (Myrcianthes coquimbensis), Algarrobo dulce (Prosopis
flexuosa), Algarrobo (Prosopis chilensis) y cuyo potencial bioquímico podría ser
usado en la industria agroalimentaria y/o biomédica nacional.

15
3. METODOLOGÍA
Estandarización de protocolos de propagación clonal in vitro, por esquejes y por
semillas para Garra de Léon (L. ovallei), Lucumillo (M. coquimbensis), Algarrobo
dulce (P. flexuosa) y Algarrobo (P. chilensis).
Con el objetivo de aumentar las posibilidades de micropropagar exitosamente las cuatro
especies, se optimizaron protocolos de propagación sexual y asexual. El Cuadro 1
muestra el diseño experimental, el Cuadro 2 indica el número de metodologías probadas,
y el Cuadro 3 se muestra el tiempo requerido para optimizar cada protocolo.
Cuadro 1. Metodología de trabajo
Cuadro 2. Cantidad de metodologías probadas.
Propagación Prosopis
flexuosa
Leontochir
ovallei
Myrcianthes.
coquimbensis
Prosopis
chilensis
Semillas 2 3 (108) 6 2
Ex vitro 24 2 24 24
In vitro 6 2 27 6
Cuadro 3. Tiempo requerido para optimizar protocolos de propagación.
Propagación Prosopis
flexuosa
Leontochir
ovallei
Myrcianthes.
coquimbensis
Prosopis
chilensis
Semillas 3 días 17 meses 6 meses 3 días
Ex vitro 2 meses 8 meses 2 meses 2 meses
In vitro 4 meses 2 meses 17 meses 4 meses

16
3.1. Propagación in vitro de tipo organizado.
3.1.1 Colecta de material: Se utilizaron explantes (o esquejes) de plantas jóvenes. Si
bien se intentó utilizar explantes desde plantas adultas colectadas en terreno, la elevada
carga de hongos, virus y bacterias que arrastraban hizo inviable su utilización. Las
plantas jóvenes se obtuvieron a partir de la germinación de semillas de cada especie.
3.1.2 Esterilización de explantes: El material colectado fue lavado con agua potable
para eliminar el exceso de polvo, y luego sumergido en una solución fungicida (2,5 g
CAPTAN+0,6 g BENLATE /1 litro de agua destilada esterilizada (H20 dE) y agitación
durante 15-20 minutos. Luego, los esquejes fueron profusamente enguajados con H20dE
y dos gotas de Tween-20 y mantenidos en agitación por 10 minutos. Luego, los
explantes fueron sumergidos en una solución de hipoclorito de sodio 1.5% + 2 gotas de
Tween-20 durante 10 minutos en agitación. A continuación se realizaron tres lavados con
H2O destilada estéril en agitación por 5 minutos cada uno.
3.1.3 Cultivo de explantes: Los explantes estériles fueron transferidos a un medio de
crecimiento básico de Murashige & Skoog rico en macro y micro nutrientes,
suplementado con vitaminas, fitohormonas (auxinas), azúcar y agar-agar. Para cada
especie se desarrolló un medio básico específico. Los segmentos uninodales se cultivaron
en condiciones de asepsia por 30 días a 21°C, fotoperíodo largo (16 h luz y 8 h
oscuridad), 135 µmol m-2 s-1, 70% de humedad relativa.
3.1.4 Multiplicación nuevos explantes: Las plantas establecidas in vitro fueron
usadas como nuevos explantes. Así, los vástagos de estos explantes fueron segmentados
en pequeños trozos de una longitud mínima de 0,5 cm. Su cultivo se hizo como se
describió en el punto 1.3.
3.1.5 Enraizamiento: Posterior al establecimiento de los plantines (plántula sin raíz), se
inició el enraizamiento de los mismos. Para ello, los plantines fueron crecidos sobre un
medio de crecimiento básico de Murashige & Skoog rico en macro y micro nutrientes,
suplementado con vitaminas, una combinación de auxinas (Ácido Indol Butírico) y
citoquininas (Kinetina), azúcar, agar-agar y Carbón activado. Nuevamente, los
segmentos uninodales se cultivaron en condiciones de asepsia por 30 días a 21°C,
fotoperíodo largo (16 h luz y 8 h oscuridad), 135 µmol m-2 s-1, 70% de humedad relativa.
3.1.6 Aclimatación: Una vez que las plántulas (plantines con raíz) lograron un buen
desarrollado de raíces, se inició su aclimatación para ser transferidas a un invernadero.
Este proceso se realizó en una cámara de crecimiento a 21°C, fotoperíodo largo (16 h luz
y 8 h oscuridad), 70% de humedad relativa. El proceso se inició sacando las plantas de
los frascos cuidando no romper las raíces y lavándolas con agua potable eliminado los
restos de agar. Luego, las plantas fueron trasplantadas en vasos de plástico que
contenían una mezcla esterilizada de arena: vermiculita (2:1). El sustrato fue
humedecido con agua destilada y fertilizado levemente con un abono granulado
previamente diluido. Para evitar la deshidratación de las plántulas, cada vaso de plástico
fue cubierto con otro vaso de plástico invertido para generar un mini invernadero, en el
cual ambos vasos fueron agujereados en sus extremos para permitir la absorción de
agua (vaso inferior) e intercambio gaseoso (vaso superior). Luego, y con el fin de
aclimatar a la planta a las condiciones de la cámara de crecimiento, los vasos fueron
paulatinamente abiertos cada 7 días, proceso que terminó al cabo de 1 mes. Durante
todo este período, las plantas fueron permanente regadas. Una vez completado el mes
de aclimatación, las plantas fueron trasladadas a un invernadero a 21°C ± 5°C, donde la
luz natural fue complementado con luz artificial desde las 18:00h a las 24:00h mediante
la utilización de luces halógenas de sodio (150W) que permitían generar una intensidad
lumínica de 250-350 µmol m-2 s-1 y establecer así un fotoperíodo largo (18 h luz y 6 h
oscuridad). Las plantas se mantuvieron en estas condiciones durante 2 meses, con riegos
diarios durante las dos primeras semanas, luego tres por semana, y finalizar con dos
riegos por semana. Una vez cumplido los dos meses, las plantas se transfirieron a un
sombreadero dispuesto especialmente para este proyecto y expuesto a condiciones
climáticas naturales de La Serena 29°54’53” (S), 71°14’31” (O), con el fin de ir
reforzando su aclimatación y adaptación a las condiciones áridas.
3.1.7 Análisis estadísticos: La significancia estadística entre tratamientos o entre las
distintas pruebas que se hicieron a lo largo del estudio, se realizaron mediante análisis de

17
varianza (ANOVA) y Test de Tukey, ambos con un 95% de confianza (p<0.05) usando el
programa estadístico R (R, 2005).
3.2. Propagación vegetativa por esquejes en cama de propagación
3.2.1. Colecta de material: Al igual que en A.1, se utilizaron esquejes de plantas
jóvenes previamente germinadas por nosotros mismos pues los esquejes provenientes de
plantas adultas colectadas en terreno traían la ya mencionada elevada carga de
microorganismos que imposibilitaban su utilización. En cada esqueje se mantuvieron al
menos 6 yemas laterales, y todas sus hojas.
3.2.2 Esterilización de esquejes: Idem punto A.2
3.2.3 Enraizamiento de esquejes: Una vez desinfectados los esquejes, estos fueron
nuevamente cortados dejando solo dos yemas laterales, manteniendo la yema superior y
cortando las hojas de la yema inferior. Luego, los explantes fueron sumergidos por 5
segundos en Ácido Indol Butírico (auxina) en concentraciones que variaron entre 5000 y
7500 mg/L según cada especie. Luego, los explantes fueron puestos en una cama de
propagación (o cama caliente) mantenida a 21°C constante y con cuatro sustratos
distintos (arena, vermiculita, tierra de hojas, y una mezcla de todas ellas). La cama
caliente se encontraba al interior de un invernadero con condiciones ya indicadas en A.6
El enraizamiento de los esquejes de las plantas se evaluó después de 60 días.
3.2.4 Aclimatación: Aquellos esquejes enraizados fueron traspasados cuidadosamente
desde la cama caliente a bolsas de vivero. El sustrato elegido para este proceso de
aclimatación fue tierra de hoja comercial. Las plántulas fueron mantenidas en el
invernadero por 1 mes después del cual fueron trasladadas hasta un sombreadero (ver
A.6).
3.2.5 Análisis estadísticos: La significancia estadística entre tratamientos o entre las
distintas pruebas que se hicieron a lo largo del estudio, se realizaron mediante análisis de
varianza (ANOVA) y Test de Tukey, ambos con un 95% de confianza (p<0.05) usando el
programa estadístico R (R, 2005).
3.3. Propagación por semillas
3.3.1 Colecta de material: Para L. ovallei se tuvo la suerte que la especie floreció en el
año 2011, lo que permitió obtener semillas para conducir el estudio en esta planta. Si
bien se consiguió un permiso especial para colectar las semillas en el Parque Nacional
Llanos de Challe (28°11’07” S, 71°09’25” O) hasta el año 2014, solo hubo floración el
año indicado por lo que se desconoce si podremos volver a colectar semillas dentro del
Parque Nacional, que es donde se concentra la mayor población de L. ovallei. Para
colectar sus semillas se utilizó un método simple pero muy eficiente. Este consistió en
poner bolsas de papel adosadas en cada inflorescencia de la cual se quiso obtener
semillas. Esto no solo logró evitar la depredación de las semillas por parte de la fauna
local sino además permitió controlar la indehiscencia explosiva que presenta esta
inflorescencia. Un total de 120 inflorescencias fueron seleccionadas, lo que equivale a
tener aproximadamente 21.000 semillas. Para M. coquimbensis, un total de 5000
semillas fueron colectadas a lo largo de su rango de distribución (Punta de Choros hasta
el Norte de Tongoy), concentrando la colecta en la zona del Arrayán (29°42’48” S,
71°18’02” O), que es el lugar donde se concentra la población más numerosa de la
especie. Para Prosopis chilensis se colectaron 300 vainas (1200 semillas) colectadas a lo
largo de su distribución en la Región de Coquimbo (Valle del Elqui hasta Valle del
Choapa). Se evaluaron dos tratamientos de escarificación, con ácido sulfúrico y sin él,
siendo tan robustos los resultados que no fue necesario seguir probando nuevos
tratamientos. Para P. flexuosa, se colectaron 600 (2400 semillas) colectadas en la Región
de Atacama, mayoritariamente en unos bosquetes ubicados al costado de la carretera
que une Caldera con Copiapó (27°21’22” S, 70°39’59” S). Al igual que en P. chilensis,
aquí no fue necesario evaluar más que los dos tratamientos de escarificación ácida.
3.3.1 Tratamientos germinativos: L. ovallei se probaron 108 tratamientos (ver ANEXO
Tabla 1). Para M. coquimbensis se evaluaron tratamientos de germinación con y sin
pericarpio. Para P. chilensis y P. flexuosa se evaluaron tratamientos de escarificación, con
ácido sulfúrico y sin él.

18
3.3.2 Análisis estadísticos: La significancia estadística entre tratamientos o entre las
distintas pruebas que se hicieron a lo largo del estudio, se realizaron mediante análisis de
varianza (ANOVA) y Test de Tukey, ambos con un 95% de confianza (p<0.05) usando el
programa estadístico R (R, 2005).
Estandarización de protocolos para identificar y cuantificar -mediante
herramientas metabolómicas- compuestos biológicos naturales presentes en
hojas, frutos o raíces de las cuatro especies nativas seleccionadas.
Se aplicaron protocolos de análisis químicos para evaluar los principios inmediatos
(grupos nutritivos del metabolismo primario) e identificación/cuantificación de algunos
metabolitos secundarios en hojas y semillas de L. ovallei, hojas, frutos y semillas en M.
coquimbensis, hojas y vainas de P. chilensis, y en hojas y vainas de P. flexuosa. Los
grupos nutritivos del metabolismo primario (análisis proximal, centecimal o de weende)
aplicados al estudio de las especies vegetales mencionadas y los análisis de metabolitos
secundarios aplicados se resumen en el Cuadro 4.
Cuadro 4: Resumen de las Técnicas analíticas (Protocolos de análisis) empleados aplicados al estudio de las cuatro especies vegetales.
3.4 Tratamiento Preliminar de las Muestras (Preparación de la muestra
ingresada a Laboratorio para ser transformada en muestra analítica): Las
muestras de hojas de las especies en estudio, se recogieron al azar y se guardaron en
bolsas de papel para posteriormente ser llevadas a Laboratorio. Una vez en éste, se
procedió a separar las hojas de los tallos. La hojas se lavaron con detergente no iónico, y
enjuagues sucesivos con agua destilada y se secaron en estufa de aire forzado, marca
memmert, a 60º C hasta alcanzar peso constante, por un periodo de 48 a 72 horas. Una
vez secas se molieron en molino de cuchillas, se pasaron por un tamiz de 40 mallas y se
almacenaron en un tubo falcón a -80ºC para su conservación y posterior análisis. En el
Humedad (Técnica gravimetría: Secado en estufa convencional) Proximal
Cenizas (Técnica gravimetría: método calcinación en mufla)
Extracto Etéreo (Técnica Gravimetría: método extracción Soxhlet)
Azúcares Totales/Reductores (Técnica EAM: método Dubois/ Somogyi-Nelson)
Proteínas (Técnica volumetría: método Kjeldahl)
Mineral Cenizas
Macronutrientes: Ca, Mg : Técnica Espectroscopía de Absorción Atómica
Na y K : Técnica Espectroscopía de Emisión Atómica
N: Técnica Volumétrica; método de Kjeldahl P: Técnica EAM; método del fosfovanadomolibdato.
Micronutrientes
(Fe, Cu, Mn, Zn): Técnica EAA; método digestión ácida
B: Técnica EAM; método Azometina H.
Polifenoles Espectroscopía de Absorción Molecular: método de Folin Ciocalteau
Perfil ácidos Grasos Cromatografía de Gases- Detector FID
Otros metabolitos secundarios
Cromatografía en capa final (TLC) y HPLC

19
caso de las vainas de Prosopis chilensis y Prosopis flexuosa se sometieron al mismo
proceso de secado de las hojas, una vez secas las vainas se molieron. Para la especie
Myrcianthes coquimbensis se retiró la semilla del fruto y ambos fueron analizados por
separado bajo los mismos procedimientos previos de secado y molienda. Los análisis de
las muestras analíticas de las especies de interés se describen en los párrafos sguientes.
3.5 Análisis de Humedad total y remanente después del secado inicial antes de
la molienda.
Para la determinación de la humedad se emplearon cápsulas de petri, taradas después de
someterlas a secado en estufa a 60° por 24 horas. Luego se masaron 10 g de muestra
con sensibilidad de 0,1 mg en balanza analítica y se dejaron en estufa a 60º C
(aproximadamente 24 horas) hasta obtener peso constante. El porcentaje de humedad
en la muestra se calculó por diferencia de pérdida de masa, expresada en porcentaje. A
partir de este valor de humedad se determinó la materia Seca. Sobre la materia seca se
realizaron los análisis químicos correspondientes.
3.6 Análisis de Cenizas.
Para el análisis de cenizas, que corresponden a la fracción Total del componente mineral
del vegetal, se empleó una porción de la muestra previamente seca, la cual fue masada y
luego combustionada en una mufla a 550°. Toda la materia orgánica se incineró y sólo
quedó el residuo inorgánico. Para la determinación se emplearon crisoles de porcelana
tarados, los cuales fueron previamente puesto en una mufla por 3 horas a 550 0C. Luego
se masaron 0,5 g de muestra con sensibilidad de 0,1 mg en balanza analítica. Se calcinó
la muestra empleando un mechero, y posteriormente se sometió a 550°C en mufla por 3
horas. El residuo mineral obtenido que representó el contenido de cenizas en las
muestras se expresó en porcentaje.
3.7 Determinación Analítica del extracto etéreo (EE).
La fracción denominada extracto etéreo, que corresponde a la destilación de una muestra
con un solvente apolar, y que incluye, además de los lípidos, ceras, pigmentos, alcoholes
y ácidos grasos orgánicos, se analizó empleando el método Soxhlet, empleando como
extractante hexano. Para ello se masaron 5 g de muestra anhidra, con precisión de 0,1
mg, la que se colocó en un cartuchohecho con papel filtro previamente masado, de tal
forma de que no se pudiese perder muestra durante la extracción. El sobre se colocó
dentro de la cámara de extracción del extractor soxhlet y se agregó el hexano hasta el
primer sifón. Bajo calentamiento regulado y constante, la muestra se sometió a
extracciones continuas por un lapso de 3 horas. Una vez terminado este tiempo, se retiró
el cartucho y se dejó secar en estufa hasta eliminación completa del hexano remanente,
se enfrió en desecador y se masó. El contenido, expresado en porcentaje de extracto
etéreo, se calculó mediante la pérdida de peso de la muestra seca en función de la
muestra desgrasada, en porcentaje.
3.8 Análisis de la proteina bruta.
El método empleado para el análisis de proteína bruta fue el método de Kjeldahl por
medio de un sistema digestor y destilador Kjeldahl semi automático Büchi. Para ello se
masaron 0.5 gramos, con precisión de 0,1 mg de muestra seca y se llevaron a tubos
Kjeldahl a los cuales se le agregaron 10 mL de H2SO4 (96%, 18 mol/L, d= 1.84 kg/L) con
acelerador de selenio y 5 mL de H2O2, dejándose en digestión por 1 hora a 420 oC. Una
vez terminado el tiempo de digestión, para realizar el destilado del amonio formado, se
adicionó al tubo de digestión NaOH 40% p/v, hasta reacción alcalina, la cual se verificó
por aparición de un tono rosado del indicador fenolftaleína. El amoníaco destilado se
recogió en un matraz erlenmeyer conteniendo 25 mL de H3BO3 al 4%, al cual se le
adicionó previamente indicador mixto Tashiro. Posteriormente el amoníaco recogido en
ácido bórico, como borato amónico, se tituló con una solución estándarizada de H2SO4
0.1 N hasta nuevo viraje del indicador. Se expresó el nitrógeno en % y se aplicó el factor
5,7 para conversión del nitrógeno en % de proteina.

20
3.9 Determinación de azúcares totales, azúcares reductores y no reductores.
Para la determinación de azúcares totales y reductores se procedió a masar 0.1 g de
muestra en balanza analítica con precisión de 0,1 mg en un vaso precipitado de 50 mL.
Se adicionó 5 mL de etanol al 80% y se cubrió con vidrio de reloj, para luego ser
sometido a ebullición por un periodo de 15 minutos en plancha calefactora a 150 oC. Una
vez cumplido el tiempo de ebullición y frias las muestras, se filtró empleando papel
whatman 41, recogiéndose el filtrado directamente en un matraz de aforo de 10,0 mL.
Los lavados y enrase a volumen se efectuaron empleando etanol al 80%. Se rotuló como
extracto 1. Luego del extracto 1, bien homogeneizado, en un tubo de ensayos, se colocó
una alícuota de 1,0 mL, para ser llevada a sequedad por calentamiento a baño maría.
Una vez eliminado completamente el solvente, el residuo seco se disolvió en 3 mL de
agua destilada, con agitación intensa de un minuto por medio de un vortex. Se filtró
empleando papel whatman 41, recibiéndose el filtrado directamente en matraz de aforo
de 10,0 mL. Los lavados y enrase a volumen se efectuaron empleando agua destilada. Se
rotuló como extracto 2.
a) Determinación de azúcares reductores: Método de Somonyi-Nelson
Se realizó empleando una alícuota de 2 mL del extracto 2 a la cual se le adicionó 1 mL de
reactivo de Somonyi. Se llevó a ebullición en baño maría por 10 a 15 minutos, se enfrió
en baño de hielo y se agregó 1 mL de reactivo de Nelson con agitación continua. Una vez
desarrollado el color, se midió la absorbancia a 620 nm en un Espectrofotométro de
Absorción Molecular UV-VIS, Marca Jasco, modelo V530. El contenido de azúcares
reductores se obtuvo frente a una curva de calibración externa de glucosa en un rango
de 0 a 100 mg glucosa/L, expresándose el resultado en porcentaje.
Reactivo de Nelson: Se disolvieron 25 g de molibdato de amonio en 450 mL de agua
destilada, luego se agregaron 21 mL de H2SO4 (96%, 18 mol/L, d= 1.84 kg/L), después
de mezclar se agregaron 3 g de Na2HAsO4.7H2O disueltos en 25 mL de agua destilada. Se
homogenizó bien y se incubó a 37° por 48 horas. La solución se guardó en frasco color
topacio.
Reactivo de Somogyi: Se disolvieron 28 g de Na2HPO4 anhidro y 4 g de tartrato de
sodio y potasio en aproximadamente 700 mL de agua destilada. Se agregaron luego 100
mL de NaOH 1N agitando, y luego 80 mL de CuSO4 10% (p/v). Una vez disuelto todo se
gregaron 180 g de Na2SO4 anhidro y se diluyó a 1 litro con agua destilada. La solución se
dejó en reposo durante un día y se decantó el sobrenadante clarificado. Este reactivo es
estable indefinidamente.
b) Determinación de azúcares totales (Método de Dubois)
Se tomó una alícuota de 1,0 mL de muestra del extracto 2 en tubo de ensayos
introducido en un baño de hielo y se adicionó 1 mL de fenol al 5% más 5 mL de H2SO4
(96% , 18 mol/L , d= 1.84 kg/L). La solución enfriada se agitó posteriormente en un
vortex, para ser llevada luego a baño maría por 10-15 minutos. Una vez fría la solución
se midió la absorbancia a 422 nm en un Espectrofotométro de Absorción Molecular UV-
VIS, Marca Jasco, modelo V530. El contenido de azúcares totales se obtuvo frente a una
curva de calibración externa de glucosa en un rango de 0 a 100 mg glucosa/L,
expresándose el resultado en porcentaje de glucosa.
c) Azúcares No- Reductores.
Se obtienen por diferencia entre los azúcares totales y los no- reductores.
3.10 Análisis de Fibra.
Para la determinación de fibra se masaron, con precisión de 0,1 mg, 2 g de muestra seca
y desgrasada, residuo proveniente de la extracción soxhlet, muestra la cual se colocó
dentro de un vaso pp de 500 mL al cual se le adicionó 250 mL de solución de H2SO4 1.25
% (0,255N) más 0.5 mL de alcohol amílico, para evitar formación de espumas. Se
agregaron perlas de ebullición, el sistema se cerró mediante un sistema de refrigeración
redondo sobre el vaso y se sometió a reflujo por 30 minutos, controlando la temperatura
de la ebullición. Una vez completado el tiempo de reflujo, en caliente, se filtró a presión
reducida la solución, empleando lino como material filtrante. El residuo se lavó con agua
caliente hasta cese de la reacción ácida del filtrado, ayudándose para ello con papel

21
indicador universal de pH. Al vaso en el cual se realizó la primera hidrólisis se traspasó el
residuo del medio filtrante, utlizando 250 mL de solución alcalina de NaOH 1.25%
(0,313N) más 0.5 mL de alcohol amílico y perlas de ebullición. Se sometió a reflujo de la
misma manera que la señalada anteriormente para la hidrólisis ácida. Finalizado el
tiempo de reflujo, se filtró en caliente, empleando como medio filtrante un papel de filtro
de poro grueso, previamente tarado. El papel, conteniendo el residuo, correspondiente a
la fibra bruta, se llevó a estufa, a 60 oC, hasta total sequedad y se masó. Expresándose
el residuo en porcentaje respecto a la muestra inicial.
3.11 Análisis de los componentes minerales.
La preparación de la muestra para el análisis de los componentes minerales se realizó
por mineralización vía húmeda, para lo cual se masó, con sensibilidad de 0.1 mg, 0.5 g
de muestra seca y finamente pulverizada que se colocó en un vaso de precipitados de
borosilicato de 250 mL. Se adicionó 8.0 mL de HNO3 (d= 1.40 g/mL 65 % p/p) y 3.0 mL
de HClO4 (d= 1.67 g/mL 72% p/p), se cubrió con un vidrio de reloj y se llevó a digestión
en placa calefactora (300oC), hasta aparición de humos blancos, etapa que duró
aproximadamente 10 minutos. Pasado este lapso de tiempo se retiró el vaso de la placa,
se dejó enfriar y se adicionó 15.0 mL de agua destilada. Se llevó a ebullición para
disolución de las sales formadas. Una vez retirado el vaso del calor y enfriado se procedió
a filtrar cuantitativamente con papel filtro Whatman 40 recogiéndose el filtrado en
matraces de aforo de 100,0 mL. El lavado y enrase se realizó empleando agua destilada.
Se rotuló como solución 1. En esta solución se analizaron por Espectroscopía de
absorción atómica (EAA) en un equipo Varian, modelo SpectrAA 220, los elementos
minoritarios presentes en las hojas: Fe, Cu, Mn y Zn, bajo las condiciones estándares
establecidas por el fabricante del equipo. Para el análisis de Na, K, Ca y Mg, elementos
mayoritarios se procedió a tomar una alícuota de 10.0 mL de la solución 1, la cual se
llevó a un matraz de aforo de 100.0 mL, agregándose como amortiguador de
interferencias y supresor de ionización una alícuota de 10.0 mL de solución de La 2% y
Cs 1%. En esta solución se analizaron por Espectroscopía de absorción atómica (EAA) en
un equipo Varian, modelo SpectrAA 220, los elementos mayoritarios: Na, K, Ca y Mg,
bajo las condiciones estándares establecidas por el fabricante del equipo. Los patrones
empleados en la preparación de las curvas de calibración para cada elemento fueron
obtenidos de titrisoles de 1,00 g/L marca Merck. Las diluciones para las curvas se
realizaron empleando como solución de enrase HCl 2% y agregando en las mismas
proporciones la solución de La 2% y Cs 1%.
3.12 Determinación de Boro.
La determinación Boro se realizó a partir de las cenizas obtenidas luego de la calcinación
en mufla. Para el análisis se procedió a untar las cenizas con gotas de agua destilada,
para luego adicionar 10,0 mL de HCl 2 M. y calentar en plancha calefactora hasta
ebullición. Luego de enfriar la solución se filtró utilizando papel filtro whatman 40 y
recogiéndose el filtrado en un matraz de 100 mL, enrasándose con agua destilada. Se
llevó a cabo mediante el método espectrofotométrico empleando el agente acomplejante
azometina-H. Para ello se procedió a tomar una alícuota de 1 mL del filtrado en un tubo
de ensayos, se agregó 4 mL de solución tampón, una vez mezclado se adicionó 2 mL de
solución de azometina-H. Se dejó en reposo por 30 a 60 minutos pero no más de 90
minutos y se procedió a leer a 430 nm en un Espectrofotométro de Absorción Molecular
UV-VIS, Marca Jasco, modelo V530, aplicando el método de calibración externa con un
patrón de Boro, calidad Patrón Primario. Para la preparación de la Solución Tampón se
disolvió 100 g de acetato de amonio, (CH3COONH4), en 160 mL de agua destilada,
solución a la cual se le agregó posteriormente 50 mL de ácido acético glacial, se
homogeneizó bien y se agregó, hasta disolución, 2.68 g de sal di sódica de ácido
etilendiamino-tetraacético (Na2EDTA ∙2H2O). Enseguida se agregó 2.4 mL de ácido
tioglicólico (ácido mercaptoacético, C2H4O2S, d= 1.32 Kg/L) y se homogeneizó
correctamente, para dejar la solución en reposo durante la noche, almacenándose
posteriormente en frascos de polietileno. (La solución es estable sólo una semana). Para
la preparación de la solución de Azometina-H/ácido ascórbico, reactivo colorimétrico, se

22
masaron 0,9 g de azometina-H, (C17H12NNaO8S2) más 2 g de ácido ascórbico (C6H8O6)
disueltos en 100 mL de agua destilada. (Nota: la solución guardada en frasco de
polietileno, refrigerada, dura 15 días).
3.13 Determinacion de Fósforo.
La determinación Fósforo se realizó a partir de las cenizas obtenidas luego de la
calcinación en mufla, en la determinación de cenizas señaladas en párrafos anteriores.
Para el análisis se procedió a untar las cenizas con gotas de agua destilada, para luego
adicionar 10,0 mL de HCl 2 M. y calentar en plancha calefactora hasta ebullición. Luego
de enfriar la solución se filtró utilizando papel filtro whatman 40 y recogiéndose el filtrado
en un matraz de 100 mL, enrasándose con agua destilada. Se aplicó el método
espectrofotométrico del fosfo-vanadomolibdato, para ello se tomó una alícuota de 1,0 mL
del filtrado, se agregó 4 mL de la solución de nitro-vanadomolibdato, se mezcló y se dejó
en reposo por 1 hora, al cabo de la cual se midió la absorbancia a 450 nm en un
Espectrofotométro de Absorción Molecular UV-VIS, Marca Jasco, modelo V530. Para la
cuantificación del fósforo se aplicó el método de calibración externa por medio de una
serie de patrones de fósforo en un rango de concentraciones entre 0 y 200 mg/L de P.
Para la preparación de la solución de nitro-vanado-molibdato, se mezclaron en partes
iguales las siguientes soluciones: Solución de vanadato de amonio 0,9 g/L, preparada por
disolución de 0,9 g de vanadato de amonio NH4VO3 en alrededor de 500 mL de agua
hirviendo, adicionada una vez fría de 24 mL de ácido nítrico concentrado y diluida con
agua destilada a 1L. Solución de molibdato de amonio 19 g/L, preparada por disolución
de 19,0 g de molibdato de amonio (NH4)6Mo7O24 ∙4 H2O en agua a 50 oC, enfriada y
diluida a 1 L con agua destilada y Ácido nítrico 1,5 mol/L, preparado por disolución en
agua de 105 mL de HNO3 (65%, 14 mol/L, d= 1.39 Kg/L) enrasado a 1L.
3.14 Polifenoles Totales. Método de Folin Ciocalteau.
La determinación de polifenoles se realizó en tres etapas. La primera fue una extracción
con solvente para la cual se pesaron 0,100 g de muestra seca en tubos de centrífuga y
se le adicionaron 2 mL de metanol al 80%. Esta muestra se agitó en vortex por 1 minuto
y se centrifugó a 3000 rpm por 5 minutos. Luego se separó el sobrenadante y se guardó
en un frasco ámbar. En el residuo se repitió el proceso de extracción hasta completar
tres veces que juntos constituyeron el total de polifenoles extraídos. La segunda etapa
consistió en el desarrollo del color. En matraces de aforo de 10,0 mL se adicionaron 200
uL de extracto (resultado de la primera etapa), 1 mL de metanol (p.a) y 500uL de
reactivo de folin. Esta mezcla fue agitada por 3 minutos y luego se le adicionó 1 mL de
Na2CO3 al 36%. Luego la solución se aforó con agua destilada y se dejó por 1 hora en
oscuridad, para luego proceder a su centrifugación por 5 minutos a 3000 rpm. La tercera
etapa fue la medición de las muestras en un espectrofotómetro a 725 nm junto con los
patrones de calibración, en un rango de concentraciones de 0 a 20,0 ug de ácido
cafeico/mL tratados de la misma forma mencionada en la segunda etapa.
3.15 Perfil de ácidos grasos.
Los ácidos grasos, salvo contadas excepciones, son los componentes fundamentales de
un aceite o grasa. No se encuentran normalmente como ácidos grasos libres, cuando lo
están es tan sólo en pequeñas cantidades, comunicando a la grasa cierta acidez.
Normalmente los ácidos grasos están formando ésteres, habitualmente con la glicerina,
para dar lugar a los glicéridos (mono, di y triaciglicerol) y fosfátidos. También pueden
formar ésteres con alcoholes grasos de estructura lineal (ceras) o terpénica (ésteres de
terpenos y ésteres se esteroles). Los ácidos grasos pueden ser saturados y no saturados.
Ejemplos de ácidos grasos insaturados son el ácido oleico y el linoleico, ambos presentes
en los aceites vegetales. El perfil de ácidos grasos, que caracteriza la ocurrencia de
ácidos grasos de origen vegetal en los triglicéridos, se analizó empleando un
cromatógrafo de gases marca Agilent Technologies modelo 6890N con detector FID,
empleando una columna capilar de 60 metros, aplicando la metodología International
Standard ISO 5508 Animal and vegetable fats and oils_ Analysis by gas chomatografhy

23
of methyl esters of fatty acids, e ISO 5509 Preparation of methyl esters of fatty acids.
Los ácidos grasos mayoritarios suelen ser los saturados palmítico (C16:0) y esteárico
(C18:0); monoinsaturados oleico (C18:1); y poliinsaturados linoleico (C18:2) y linolénico
(C18:3), variando estos contenidos y con aparición de otros ácidos grasos según el
origen de la semilla o especie vegetal.
3.16 Identificación de metabolitos secundarios por cromatografía en capa fina
(TLC).
El procedimiento llevado a cabo fue una adaptación del trabajo realizado por Pedrasa, M
(2004) en los estudios químicos y cromatográficos en Lophopappus tarapacanus. El
trabajo se realizó en dos etapas. La primera fue una extracción y preparación de un
infuso y la segunda una cromatografía en capa fina. Todos los reactivos utilizados fueron
de la marca Merck grado p.a.; como equipamiento se empleó un rotavapor Buchi modelo
R-200, agitador magnético marca Fisotom y equipamiento para cromatografía en capa
fina constituido por dos tanques de vidrio de 4L con tapa, sistema de pulverización y
micropipetas de sembrado de 1 uL y un sistema revelador-lámpara Spectroline modelo
ENF-260C/FE en una cabina de la misma marca modelo CM-10. En los diagramas
presentados en las figuras 2 y 3 se detallan los procedimientos seguidos para las
extracciones de las muestras e identificación de metabolitos secundarios.
3.17 Preparación del infuso en agua.
Se pesaron 16 g de muestra fresca en un vaso de precipitados de 250 mL y se adicionó
150 mL de agua hirviente. Se dejó macerar por 30 minutos para luego ser filtrada. El
filtrado se guardó en el congelador para su posterior uso en el análisis cromatográfico.
3.17.1 Análisis Cromatográfico.
Mediante cromatografía en capa fina se realizó un análisis de los diferentes extractos y
del infuso con el fin de investigar la presencia o liberación de metabolitos secundarios de
esta infusión. En este proceso se empleó como fase estacionaria cromatoláminas de
silicagel tipo 60 F 254 (Merck). El sembrado de los extractos metanólico de los exudados
fue realizado en el siguiente orden: EXDCM; EXHEX; EXDCM; EXMET de planta seca,
EXDCM; EXHEX; EXDCM; EXMET de planta fresca e Infuso metanólico en planta fresca.
Además se utilizaron dos fases móviles, que correspondieron a mezclas de disolventes
que permitían una mejor separación de los constituyentes de la infusión, resinas y
extractos metanólicos. Las cromatografías fueron analizadas con luz UV, δ de 254 y 366
nm con una lámpara Spectroline modelo ENF-260C/FE en una cabina de la misma marca
modelo CM-10 y por aspersión de reactivos químicos reveladores específicos, usando las
siguientes soluciones:
a. Reactivo de Dragendorff. Identificación de alcaloides Constituido por yoduro doble
de bismuto y potasio, solución lista para usar marca Merck.
b. Reactivo de Lieberman-Burchard: Utilizada para identificar triterpenos y esteroles,
los primeros dan coloración violeta, mientras que los esteroles dan coloraciones que
varían del rosado al verde y en algunas ocasiones pardas. En su preparación se
mezclan con cuidado y en frío 5 mL de anhídrido acético con 5 mL de ácido sulfúrico
concentrado, se le adicionan 50 mL de etanol absoluto, debe ser utilizado fresco, esta
solución es vaporizada directamente sobre la placa y luego es sometida a 105°C por 5
minutos.
c. Reactivo de Borntrager: Identificación de grupos antraquinónicos. Consistente en
una solución de KOH al 5% la cual generará una coloración rojiza.
d. Reactivo de p-Anisaldehido sulfúrico: Este reactivo que permite identificar
terpenos, se utiliza aplicándolo directamente sobre la placa, y luego aplicando
temperatura para una mejor revelación, 105°C por 5 minutos. La reacción positiva
corresponde a coloraciones que va desde el azul al violeta. Se prepara mezclando 1
mL de ácido sulfúrico concentrado. Sobre una solución que contiene 0.5 mL de p-
Anisaldehido en 50 mL de ácido acético glacial se debe utilizar fresco.
e. Reactivo de AlCl3 al 2% en metanol: Empleada para la identificación de flavonoides
y cumarinas, ambos presentan fluorescencia a la luz UV, a longitudes de onda de 254
24
y 366 nm. En el caso de los flavonoides cambian o intensifican su coloración, mientras
que en presencia del reactivo revelador, las cumarinas presentan coloración celeste.
f. Reactivo de FeCl3: Utilizado para identificar flavonoides los cuales presentan
coloraciones verdes cuando la reacción es positiva, posee una concentración de 1-5%
de FeCl3 en HCl 0.5 N.
g. Vapores de NH3: Empleada para identificar flavonoides. Las placas cromatográficas
expuestas a los vapores de NH3 generan una coloración amarilla en presencia de
flavonoides.
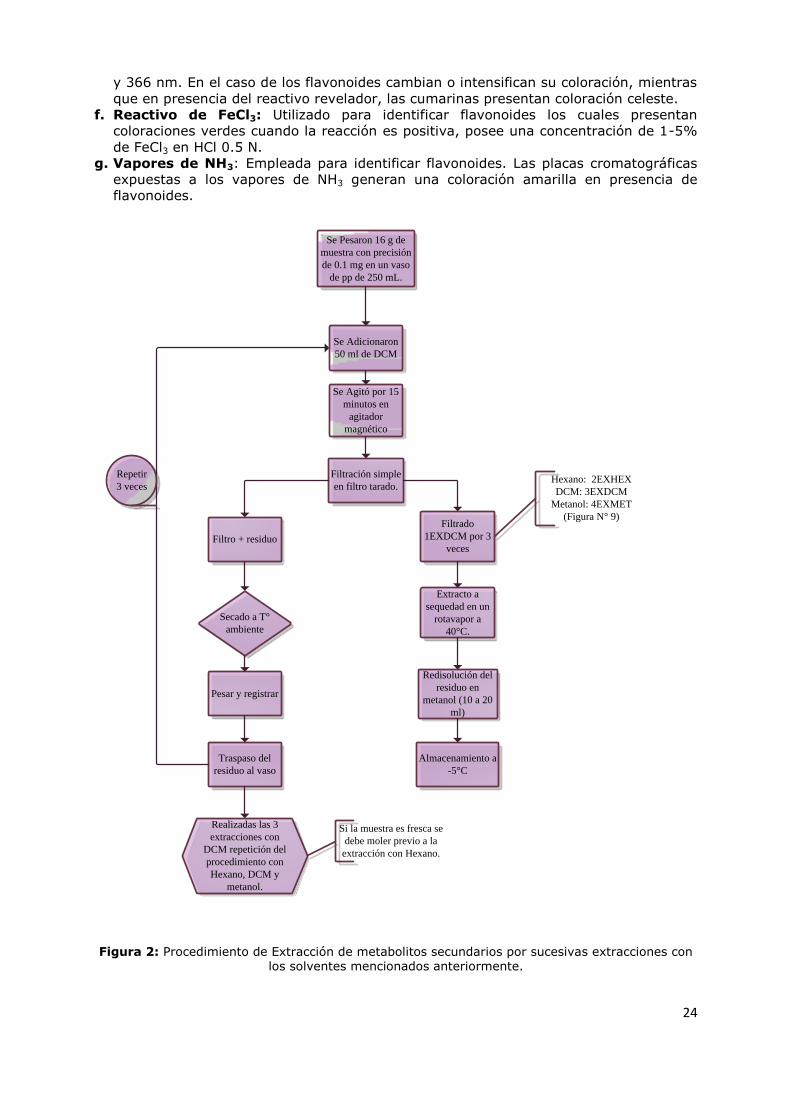
Figura 2: Procedimiento de Extracción de metabolitos secundarios por sucesivas extracciones con los solventes mencionados anteriormente.
Se Pesaron 16 g de
muestra con precisión
de 0.1 mg en un vaso
de pp de 250 mL.
Se Adicionaron
50 ml de DCM
Se Agitó por 15
minutos en
agitador
magnético
Filtración simple
en filtro tarado.
Filtro + residuo
Filtrado
1EXDCM por 3
veces
Pesar y registrar
Secado a T°
ambiente
Traspaso del
residuo al vaso
Repetir
3 veces
Extracto a
sequedad en un
rotavapor a
40°C.
Redisolución del
residuo en
metanol (10 a 20
ml)
Almacenamiento a
-5°C
Realizadas las 3
extracciones con
DCM repetición del
procedimiento con
Hexano, DCM y
metanol.
Hexano: 2EXHEX
DCM: 3EXDCM
Metanol: 4EXMET
(Figura N° 9)
Si la muestra es fresca se
debe moler previo a la
extracción con Hexano.

25
Fase móvil
DCM:EtAc
(1:1)
Flavonoides
Cumarinas
MetabolitoReactivo
revelador
Vapores de NH3
AlCl3 al 2%
metanol
FeCl3
Observar bajo luz UV a
254 y 366 nm para
intensificar los colores
Figura 3: Diagrama resumen de identificación de metabolitos secundarios, fase móvil y revelador
correspondiente y Diagrama de procedimiento de identificación de flavonoides y cumarinas.
3.18. Prueba de susceptibilidad antimicrobiana de metabolitos secundarios en
diferentes extractos de hojas.
Las pruebas se realizaron sobre 8 extractos de hojas frescas, 4 obtenidos de extracciones
sucesivas de 16 g de hojas, con cuatro distintos solventes, en orden con diclorometano
(1-EXDCM), seguido de hexano (2-EXHEX), diclorometano (3-EXDCM), y, finalmente
metanol (4-EXMET). Los 4 restantes extractos, se obtuvieron directamente sobre 16 g de
hojas con los siguientes solventes: agua (5-INF.H20), macerado con etanol (6M-ETOH),
macerado con hexano(7M.HEX) y por último el extracto obtenido del macerado con agua
(8-M.H2O). Los solventes de extracción se eliminaron a presión reducida a baja
temperatura, empleando el sistema rotavapor mencionado en párrafos anteriores, para
evitar destrucción de metabolitos secundarios. Los 8 extractos obtenidos se emplearon
en la evaluación de actividad antimicrobiana. El procedimiento o prueba de
susceptibilidad antimicrobiana se realizó por difusión en agar según ISP Chile (2005).
3.19 Análisis estadísticos: Para establecer diferencias significativas entre tratamientos
(hojas, tallos, semillas y frutos) se realizará una prueba de separación de medias
(ANDEVA) y prueba de medias de Tukey, además, para establecer posibles correlaciones
entre diferentes variables (componentes químicos) se realizará análisis de regresión
(PLS) y un análisis estadístico multivariante a través de análisis de componentes
principales (PCA).
Fase móvil
DCM:EtAc
(9:1)
Alcaloides
Tr iterpenos –
Esteroles
Terpenos
Antraquinonas
MetabolitoReactivo
revelador
Reactivo de
Dragendorf
Reactivo de
Liebermann-
Burchard
Reactivo
p-Anisaldehido
Reactivo de
Borntrager

26
4. RESULTADOS y DISCUSIÓN
4.1. PARTE DE PROPAGACIÓN (Ver mayores detalles en ANEXOS-propagación y
Difusión)
4.1.1 Garra de León (Leontochir ovallei): Se optimizaron protocolos de germinación
in vitro de semillas, propagación de rizomas y propagación in vitro de plántulas, cada uno
de los cuales se entregan con detalles en los ANEXOS n° 1, 2, 3 y 4, respectivamente. La
colecta de semillas se realizó entre Diciembre de 2010 y Febrero de 2011. Utilizando
bolsas de papel adosadas a la inflorescencia se logró contener la indehescencia explosiva
y recuperar el 100% de semillas (ANEXO n° 1). Una vez desinfectadas con Captan (u
otro antifúngico similar), las semillas fueron secadas hasta alcanzar peso constante y
luego almacenadas en frío (4°C) sin que ello afectara su viabilidad. En términos
generales, podemos indicar que las semillas de L. ovallei son muy difíciles de germinar. A
lo largo del proyecto se probaron 108 metodologías distintas. Entre otros, se probó la
germinación sobre distintos sustratos (arena, vermiculita, agar), varios tipos de
desinfecciones (con etanol, hipoclorito de sodio), varios medios de cultivo (MS, Gambord,
Hoagland), escarificación química (ácido sulfúrico, ác. clorhídrico), mecánica (rayado y
corte de testas) y térmica (remojo en agua caliente); diferentes tipos de hormonas
(auxinas, citoquininas, giberelinas), oxigenación de semillas, germinación con y sin luz,
medios con y sin sacarosa, a 21°C y 30°C. Finalmente, la combinación que mejor
germinación dio (13%) fue la desinfección de semillas con Hipoclorito de sodio, seguida
por cortes superficiales sobre las semillas y siembra en agar con medio MS suplementado
con sacarosa (2%), ácido giberélico (5mg/L), 6-Bencilaminopurina (0,5 mg/L), Cloruro
de Sodio (20 uM) y agar (8 g/L). Ver ANEXO n° 2. Si bien este % no es tan alto, es
bastante mejor que el 2% obtenido al inicio del proyecto. Al ser traspasadas a sustrato
de arena:vermiculita (2:1) estéril y después de 8 meses de crecimiento, se logró obtener
en promedio, cinco rizomas por planta (ANEXO n°3). En total, al término de este
proyecto se lograron tener 98 plantas de L. ovallei en proceso de aclimatación, que a su
vez suponen un total aproximado de 490 rizomas, y sumando.
4.1.2 Lucumillo (Myrcianthes coquimbensis): Se han optimizado protocolos de
germinación de semillas (Anexo n° 4), propagación ex vitro por esquejes (Anexo n° 5) y
propagación in vitro (Anexo n° 6). La germinación de semillas no es difícil aunque es
fundamental usar semillas maduras y eliminar el mesocarpio (ANEXO n° 4). En
promedio, se obtuvo 80% de germinación (ANEXO n° 5). En cuanto a la propagación ex
vitro por esquejes, el protocolo que logro el más alto porcentaje de enraizamiento (85%)
fue la combinación de vermiculita o tierra de hojas, ambas con AIB a 10000 ppm (ANEXO
n° 5). En cuanto a la propagación in vitro, se logró optimizar un protocolo formado por
Polivinilpirrolidona (PVP, 1mg/L), 6-bencilaminopurina (BAP, 14,1 µM) y enraizamiento de
explantes con Ácido Indolbutírico (AIB, 5000ppm). Con este protocolo (ANEXO n° 6), se
obtuvo una propagación del 80%.
4.1.3. Algarrobo dulce (Prosopis flexuosa): Se han optimizado protocolos de
germinación de semillas (Anexo n° 7), propagación ex vitro por esquejes (Anexo n° 8) y
propagación in vitro (Anexo n° 9). La germinación de semillas utilizó ácido sulfúrico, que
permitió obtener un 95% de germinación (ANEXO n° 7). En cuanto a la propagación ex
vitro por esquejes, éstos enraizaron sin necesidad de agregar enraizante aunque el
sistema de cama caliente a 21 ± 4 °C fue relevante (ANEXO n° 8). Con estas condiciones
se obtuvo un 70% de enraizamiento. Finalmente, la propagación in vitro utilizó un
protocolo con 6-bencilaminopurina (4,4 µM) y enraizamiento con Ácido Indolbutírico
(AIB, 10.000 mg/L) como productos relevantes (ANEXO n° 10). Se logró un 70% de
propagación.

27
4.1.4. Algarrobo (Prosopis chilensis): Se han optimizado protocolos de germinación
de semillas (Anexo n° 10), propagación ex vitro por esquejes (Anexo n° 11) y
propagación in vitro (Anexo n° 12). La germinación también se basó en ácido sulfúrico
con 95% de germinación (ANEXO n° 10). La propagación ex vitro por esquejes utilizó AIB
a 7500ppm como concentración óptima para enraizamiento y tierra de hojas como mejor
sustrato (ANEXO n° 11). Con estas condiciones se obtuvo un 70% de enraizamiento.
Finalmente, la propagación in vitro de esta planta fue posible utilizando casi el mismo
protocolo in vitro desarrollado en P. flexuosa, siendo diferente solo la menor
concentración de AIB (5000 mg/L) que se requirió para enraizar los esquejes de P.
chilensis (ANEXO n° 12). Con este protocolo, se obtuvo un 30% de propagación.
4.2 PARTE FITOQUÍMICA
4.2.1. Garra de León (L. ovallei): En la tabla 1 se resumen los valores obtenidos de
los análisis llevado a cabo en las hojas y semillas de L. ovallei, fenómeno del desierto
Florido Año 2011. Las hojas fueron tomadas al azar de 40 plantas para hacer un
compósito de 100 hojas representativas aproximadamente, en triplicado, y las semillas
colectadas correspondieron a un universo de 30 inflorescencia (c/u entre 10 a 12 flores)
de las cuales se colectó una sola flor (aproximadamente 20 a 30 semillas). Los lugares de
los cuales se tomaron las muestras fueron: Carrizal Bajo (Sector Desembarco), Quebrada
del Mono (Km 54, al interior del Parque Nacional Llanos del Challe) y Mina Oriente. Las
hojas y semillas fueron tratadas según se señala en el punto 3. Tratamiento Preliminar
de las Muestras. Las semillas una vez molidas se sometieron nuevamente a secado para
determinar el contenido de agua al interior de éstas que resultó ser significativo.
Dentro de los análisis realizados destaca en las hojas L. ovallei un contenido de agua
significativo de 71%, correspondiendo a materia seca sólo un 29%, del cual un 16,56%
corresponde a cenizas (Componente mineral inorgánico) y un 4,83 % a polifenoles
(componente sintetizados por las plantas en el metabolismo secundario y relacionado
directamente a respuestas de estress hídrico, entre otros factores de carencia). Si bien
no fue posible realizar el análisis de nitrógeno para estimar el contenido de proteínas por
agotamiento de la muestra recolectada y la fibra (que en su conjunto representarían el
72,6 % de la materia seca, lo que supondría un alto contenido en fibra) las hojas
manifestaron ser ricas en componentes lipídicos saponificables y no saponificables,
representados por el extracto etéreo con un 5,26 %. Dentro de los macro elementos
esenciales se observó que el contenido de fósforo obtenido en el análisis, 0,14%, se
presentó al límite. Valores entre 0,10 a 0,15 % es señal de déficit en las plantas y entre
los microelementos destacó el bajo contenido de Fe (56 mg/Kg) cuyo valor normal en
plantas es de 80 a 320 mg/Kg.
Respecto a las semillas es importante hacer notar que al ser secadas presentaron un
54% de humedad, quedando reducidas de tamaño significativamente. En este estado
mantuvieron en el interior un 25% de humedad que permanece y que sólo puede ser
eliminada al pulverizar mecánicamente la semilla. El contenido de agua total en la semilla
finalmente fue de un 79%. La materia seca que representa un 21% del total de la semilla
contiene un 2,01% de cenizas (componente mineral), siendo ricas en Hierro y Zinc, 73 y
35 mg/kg, respectivamente, mayor contenido que el encontrado en las hojas de estos
dos microelementos, un 52 % de azúcares totales solubles, siendo mayoritarios los
azúcares reductores con un aporte del 38 % en la materia seca. Se destaca además su
alto contenido en compuestos polifenólicos (antioxidantes) con un 6,9%. El aceite de la
semilla (11,7% como Extracto Etéreo), extraído en frío, resultó ser rico en ácido Linoleico
(ácido graso esencial y omega 6 ) alcanzando un 49,7 % y de ácido oleico (omega 9)
con un 34,1%. Semejante al aceite de soja, sólo que con un contenido superior en ácido
oleico. El aceite se describiría como rico en poliinsaturados (50,1%) y monoinsaturados
(34,1%) y bajo en grasas saturadas (14,0%). Se hace notar que el aceite aportaría con
un 0,1% de alfa-linolénico, ácido graso esencial (omega 3).

28
TABLA 1: COMPONENTES QUÍMICOS DE Leontochir ovallei (GARRA DE LEÓN)
PARÁMETROS QUÍMICOS HOJAS SEMILLAS
HUMEDAD (%) 71 ± 3 Primer secado: 54 ± 2 Pulverizadas: 25± 2
Total en semilla: 79 ± 2
MATERIA SECA (%) 29 ± 3 21 ± 2
ANÁLISIS DE LA MATERIA SECA: Cenizas (%)
Extracto etéreo (%) Azúcares Totales (%) Azúcares Reductores
Azúcares No-Reductores Proteína Bruta (%) Fibra (%) Polifenoles totales (metabolismo secundario)
16,6 ± 0,3
5,26 ± 0,2 0,67 ± 0,06
NA
NA NA NA
4,83 ±
2,01 ± 0,2
11,7 ± 0,3 52 ± 2
14,1 ± 0,9
38 ± 3 NA NA
6,9 ± 0,8
ANÁLISIS DE LAS CENIZAS
Macronutrientes Calcio (%) 2,74 ± 0,3 0,11 ±0,02
Magnesio (%) 0,74 ± 0,09 NA
Sodio (%) 1,1 ± 0,5 0,07 ± 0,04
Potasio (%) 2,8 ± 0,1 0,1 ± 0,0
Fósforo (%) 0,14 ± 0,03 NA
ANÁLISIS DE LAS CENIZAS Micronutrientes Hierro (mg/Kg) 56 ± 7 73 ± 11
Cobre (mg/Kg) 26 ± 3 6 ± 3
Manganeso (mg/Kg) 82 ± 11 7,7 ± 0,1
Zinc (mg/Kg) 21 ± 6 35 ± 1
Boro (mg/Kg) 37 ± 18 NA
ANÁLISIS DEL EXTRACTO ETÉREO (perfil de ácidos grasos : % en el aceite) Ácido palmítico (C16:0)
Ácido Esteárico (C18:0) Ácido Oleico (C18:1) omega 9 Ácido Linoleico (C18:2) omega 6
Ácido Linoleico (C18:3) omega 3 Ácido Araquídico (C20:0) Ácido Eicosanoico (C20:1) Ácido Behénico (C22:0) Otros S/identificación:
NA
10,3
3,1 34,1 49,7
0,1 0,4 0,2 0,2 2,0
Nota: NA: componente no analizado químicamente.

29
4.2.2. Lucumillo (M. coquimbensis): En la tabla 2 se resumen los valores obtenidos
de los análisis llevado a cabo en las hojas, frutos y semillas de M. coquimbensis. Las
hojas y frutos fueron tomadas en un sólo sector, por lo que las plantas se agruparon
originando tres muestras formadas por aproximadamente 100 hojas, las cuales fueron
tratadas según se señala en el punto 3. Tratamiento Preliminar de las Muestras.
Las hojas de M. coquimbensis dieron bajo contenido de K, Fe y Zn, mientras que el Cu y
Mn fueron elevados, los demás nutrientes foliares se presentaron dentro de los rangos de
suficiencia. Es interesante en esta planta, con un aporte de 94% de materia seca, la gran
riqueza polifenólica de sus hojas, 19% (en el Té verde rico en polifenoles se reporta un
componente polifenólico de entre 15 a 20%). Este resultado abre una puerta interesante
de investigar para establecer capacidades antioxidantes e identificar polifenoles
individuales que podrían tener aplicaciones nutraceútica interesante de elucidar. Con una
riqueza en el extracto etéreo (12%) y Azúcares reductores y No reductores en
proporciones semejantes (5%). M. coquimbensis dio positivo la identificación de
terpenos, taninos, triterpenos, antraquinonas, flavonoides y cumarinas, corroborando el
alto valor encontrado en el contenido de polifenoles. Por otro lado el extracto que
presentó actividad antimicrobianas en ambas cepas bacterianas estudiadas, Escherichia
coli (gram -) y Staphylococus aureus (gram +) fue el macerado en agua (8M.H2O). En
pruebas preliminares la presencia de cumarinas gálicas se asoció al efecto antibacteriano.
Por otro lado el estudio químico del mesocarpio del fruto y de la semilla resultó, en
magnitudes semejantes, rico en carbohidratos solubles totales (76%) siendo los azúcares
no-reductores los mayoritarios (63%), seguido de un 9% de extracto etéreo (lípidos,
pigmentos, alcoholes y ácidos grasos orgánicos) y un aporte en proteinas de 4 a 5 %. El
ácido graso mayoritario en el mesocarpio fue el ácido graso saturado Palmítico (C16:0),
34%, seguido por linoleico (omega 6), 30%, oleico (omega 9), 13% y linolénico (omega
3), este último con una riqueza destacable de un 10%. La semilla sin embargo fue baja
en ác. linoleico (2,5 %) , y rica en ácido oleico (36%), ácido palmítico (27%) y ácido
linoleico (25%).
TABLA 2: COMPONENTES QUÍMICOS DE Myrcianthes coquimbensis (LUCUMILLO)
Hojas Fruto Semillas
HUMEDAD (%) 6,43 0,2 24,5 0,2 38,5 0,2
MATERIA SECA (%) 93,6 0,2 75,5 0,2 61,5 0,2
ANÁLISIS DE LA MATERIA SECA:
Cenizas (%) Extracto etéreo (%)
Azúcares Totales (%) Azúcares no reductores (%) Azúcares Reductores (%) Proteína Bruta (%) Fibra (%) Polifenoles totales (%) (metabolismo
secundario)
8,99 0,07
12,3 0,6
10,1 0,7
5,0 0,5
5,1 0,6
NA NA
19 3
3,7 1
9,2 0,2
76 1
62 1
14,54 0,04
5,39 0,003
5,8 2 6,4 0,4
2,7 0,3
8,6 0,2
76,2 0,9
63 2
13 1
3,9 0,2
9 2 1,7 0,1
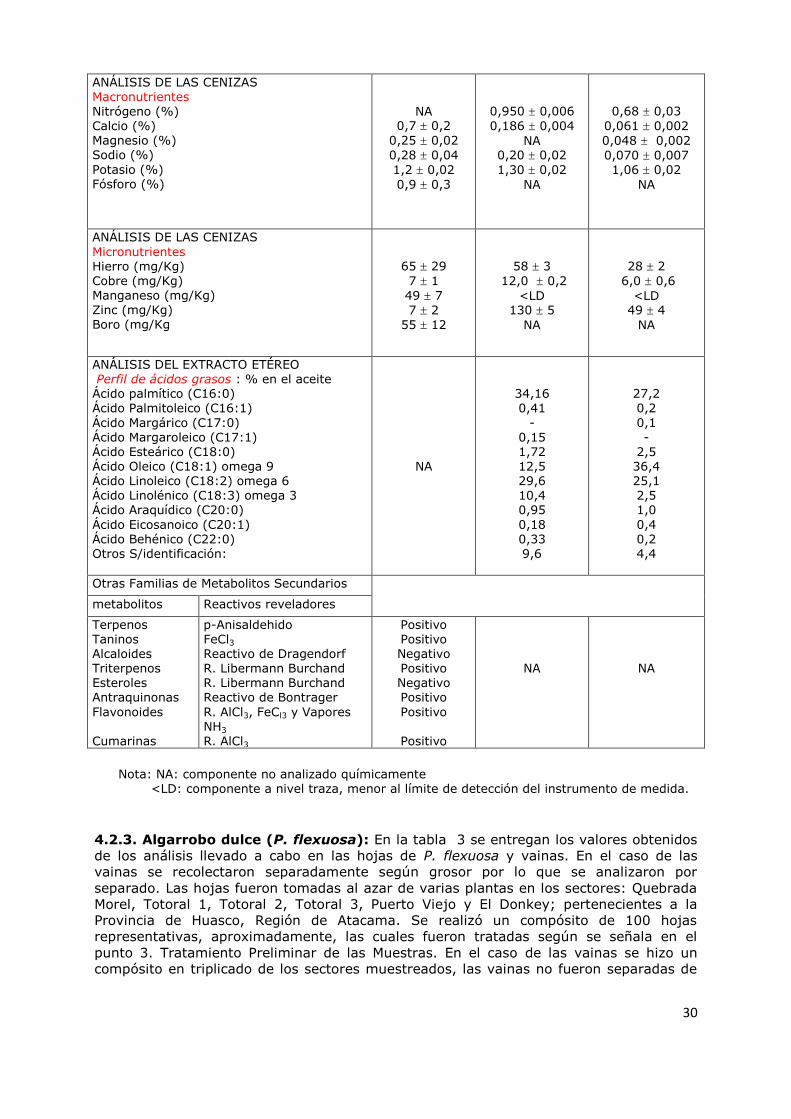
30
ANÁLISIS DE LAS CENIZAS Macronutrientes
Nitrógeno (%) Calcio (%) Magnesio (%) Sodio (%)
Potasio (%) Fósforo (%)
NA 0,7 0,2
0,25 0,02
0,28 0,04
1,2 0,02
0,9 0,3
0,950 0,006
0,186 0,004
NA 0,20 0,02
1,30 0,02
NA
0,68 0,03
0,061 0,002
0,048 0,002
0,070 0,007 1,06 0,02
NA
ANÁLISIS DE LAS CENIZAS
Micronutrientes Hierro (mg/Kg) Cobre (mg/Kg) Manganeso (mg/Kg)
Zinc (mg/Kg) Boro (mg/Kg
65 29
7 1
49 7
7 2
55 12
58 3
12,0 0,2
<LD 130 5
NA
28 2
6,0 0,6
<LD 49 4
NA
ANÁLISIS DEL EXTRACTO ETÉREO Perfil de ácidos grasos : % en el aceite Ácido palmítico (C16:0) Ácido Palmitoleico (C16:1)
Ácido Margárico (C17:0) Ácido Margaroleico (C17:1) Ácido Esteárico (C18:0) Ácido Oleico (C18:1) omega 9 Ácido Linoleico (C18:2) omega 6 Ácido Linolénico (C18:3) omega 3 Ácido Araquídico (C20:0)
Ácido Eicosanoico (C20:1) Ácido Behénico (C22:0) Otros S/identificación:
NA
34,16 0,41
- 0,15 1,72 12,5 29,6 10,4 0,95
0,18 0,33 9,6
27,2 0,2
0,1 -
2,5 36,4 25,1 2,5 1,0
0,4 0,2 4,4
Otras Familias de Metabolitos Secundarios
metabolitos Reactivos reveladores
Terpenos Taninos Alcaloides Triterpenos Esteroles Antraquinonas
Flavonoides
Cumarinas
p-Anisaldehido FeCl3 Reactivo de Dragendorf R. Libermann Burchand R. Libermann Burchand Reactivo de Bontrager
R. AlCl3, FeCl3 y Vapores
NH3
R. AlCl3
Positivo Positivo Negativo Positivo Negativo Positivo
Positivo
Positivo
NA
NA
Nota: NA: componente no analizado químicamente
<LD: componente a nivel traza, menor al límite de detección del instrumento de medida.
4.2.3. Algarrobo dulce (P. flexuosa): En la tabla 3 se entregan los valores obtenidos
de los análisis llevado a cabo en las hojas de P. flexuosa y vainas. En el caso de las
vainas se recolectaron separadamente según grosor por lo que se analizaron por
separado. Las hojas fueron tomadas al azar de varias plantas en los sectores: Quebrada
Morel, Totoral 1, Totoral 2, Totoral 3, Puerto Viejo y El Donkey; pertenecientes a la
Provincia de Huasco, Región de Atacama. Se realizó un compósito de 100 hojas
representativas, aproximadamente, las cuales fueron tratadas según se señala en el
punto 3. Tratamiento Preliminar de las Muestras. En el caso de las vainas se hizo un
compósito en triplicado de los sectores muestreados, las vainas no fueron separadas de
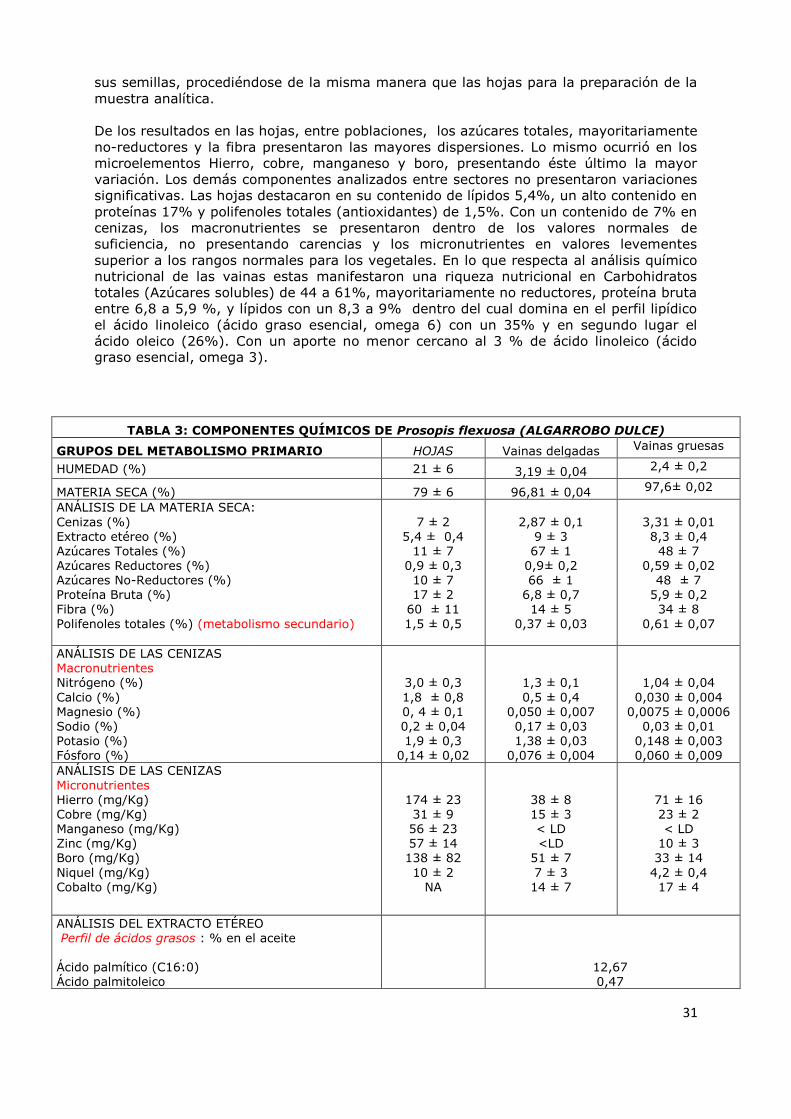
31
sus semillas, procediéndose de la misma manera que las hojas para la preparación de la
muestra analítica.
De los resultados en las hojas, entre poblaciones, los azúcares totales, mayoritariamente
no-reductores y la fibra presentaron las mayores dispersiones. Lo mismo ocurrió en los
microelementos Hierro, cobre, manganeso y boro, presentando éste último la mayor
variación. Los demás componentes analizados entre sectores no presentaron variaciones
significativas. Las hojas destacaron en su contenido de lípidos 5,4%, un alto contenido en
proteínas 17% y polifenoles totales (antioxidantes) de 1,5%. Con un contenido de 7% en
cenizas, los macronutrientes se presentaron dentro de los valores normales de
suficiencia, no presentando carencias y los micronutrientes en valores levementes
superior a los rangos normales para los vegetales. En lo que respecta al análisis químico
nutricional de las vainas estas manifestaron una riqueza nutricional en Carbohidratos
totales (Azúcares solubles) de 44 a 61%, mayoritariamente no reductores, proteína bruta
entre 6,8 a 5,9 %, y lípidos con un 8,3 a 9% dentro del cual domina en el perfil lipídico
el ácido linoleico (ácido graso esencial, omega 6) con un 35% y en segundo lugar el
ácido oleico (26%). Con un aporte no menor cercano al 3 % de ácido linoleico (ácido
graso esencial, omega 3).
TABLA 3: COMPONENTES QUÍMICOS DE Prosopis flexuosa (ALGARROBO DULCE)
GRUPOS DEL METABOLISMO PRIMARIO HOJAS Vainas delgadas Vainas gruesas
HUMEDAD (%) 21 ± 6 3,19 ± 0,04 2,4 ± 0,2
MATERIA SECA (%) 79 ± 6 96,81 ± 0,04 97,6± 0,02
ANÁLISIS DE LA MATERIA SECA:
Cenizas (%) Extracto etéreo (%) Azúcares Totales (%) Azúcares Reductores (%) Azúcares No-Reductores (%) Proteína Bruta (%) Fibra (%)
Polifenoles totales (%) (metabolismo secundario)
7 ± 2 5,4 ± 0,4
11 ± 7 0,9 ± 0,3 10 ± 7 17 ± 2
60 ± 11
1,5 ± 0,5
2,87 ± 0,1 9 ± 3 67 ± 1
0,9± 0,2 66 ± 1
6,8 ± 0,7 14 ± 5
0,37 ± 0,03
3,31 ± 0,01 8,3 ± 0,4 48 ± 7
0,59 ± 0,02 48 ± 7
5,9 ± 0,2 34 ± 8
0,61 ± 0,07
ANÁLISIS DE LAS CENIZAS Macronutrientes Nitrógeno (%)
Calcio (%) Magnesio (%)
Sodio (%) Potasio (%) Fósforo (%)
3,0 ± 0,3
1,8 ± 0,8 0, 4 ± 0,1
0,2 ± 0,04 1,9 ± 0,3
0,14 ± 0,02
1,3 ± 0,1
0,5 ± 0,4 0,050 ± 0,007
0,17 ± 0,03 1,38 ± 0,03
0,076 ± 0,004
1,04 ± 0,04
0,030 ± 0,004 0,0075 ± 0,0006
0,03 ± 0,01 0,148 ± 0,003 0,060 ± 0,009
ANÁLISIS DE LAS CENIZAS
Micronutrientes Hierro (mg/Kg) Cobre (mg/Kg) Manganeso (mg/Kg) Zinc (mg/Kg) Boro (mg/Kg)
Niquel (mg/Kg) Cobalto (mg/Kg)
174 ± 23 31 ± 9 56 ± 23 57 ± 14 138 ± 82
10 ± 2 NA
38 ± 8 15 ± 3 < LD <LD
51 ± 7
7 ± 3 14 ± 7
71 ± 16 23 ± 2 < LD
10 ± 3 33 ± 14
4,2 ± 0,4 17 ± 4
ANÁLISIS DEL EXTRACTO ETÉREO
Perfil de ácidos grasos : % en el aceite Ácido palmítico (C16:0) Ácido palmitoleico
12,67 0,47
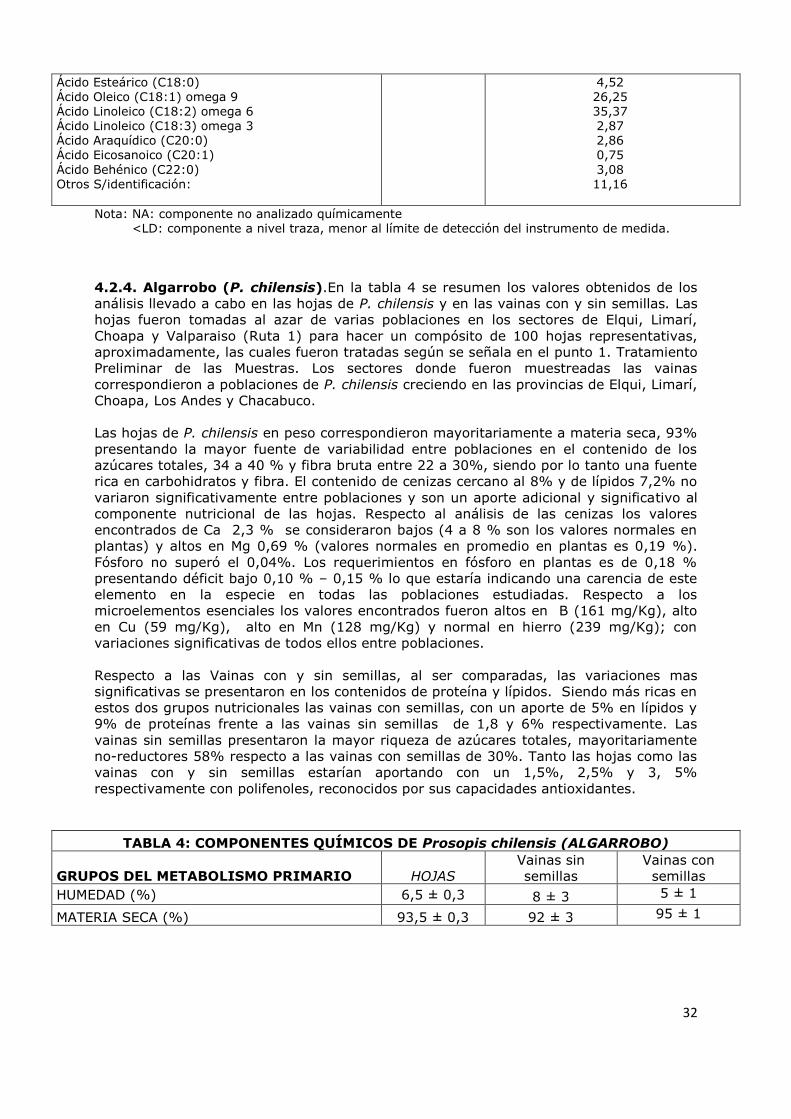
32
Ácido Esteárico (C18:0) Ácido Oleico (C18:1) omega 9
Ácido Linoleico (C18:2) omega 6 Ácido Linoleico (C18:3) omega 3 Ácido Araquídico (C20:0) Ácido Eicosanoico (C20:1)
Ácido Behénico (C22:0) Otros S/identificación:
4,52 26,25
35,37 2,87 2,86 0,75
3,08 11,16
Nota: NA: componente no analizado químicamente <LD: componente a nivel traza, menor al límite de detección del instrumento de medida.
4.2.4. Algarrobo (P. chilensis).En la tabla 4 se resumen los valores obtenidos de los
análisis llevado a cabo en las hojas de P. chilensis y en las vainas con y sin semillas. Las
hojas fueron tomadas al azar de varias poblaciones en los sectores de Elqui, Limarí,
Choapa y Valparaiso (Ruta 1) para hacer un compósito de 100 hojas representativas,
aproximadamente, las cuales fueron tratadas según se señala en el punto 1. Tratamiento
Preliminar de las Muestras. Los sectores donde fueron muestreadas las vainas
correspondieron a poblaciones de P. chilensis creciendo en las provincias de Elqui, Limarí,
Choapa, Los Andes y Chacabuco.
Las hojas de P. chilensis en peso correspondieron mayoritariamente a materia seca, 93%
presentando la mayor fuente de variabilidad entre poblaciones en el contenido de los
azúcares totales, 34 a 40 % y fibra bruta entre 22 a 30%, siendo por lo tanto una fuente
rica en carbohidratos y fibra. El contenido de cenizas cercano al 8% y de lípidos 7,2% no
variaron significativamente entre poblaciones y son un aporte adicional y significativo al
componente nutricional de las hojas. Respecto al análisis de las cenizas los valores
encontrados de Ca 2,3 % se consideraron bajos (4 a 8 % son los valores normales en
plantas) y altos en Mg 0,69 % (valores normales en promedio en plantas es 0,19 %).
Fósforo no superó el 0,04%. Los requerimientos en fósforo en plantas es de 0,18 %
presentando déficit bajo 0,10 % – 0,15 % lo que estaría indicando una carencia de este
elemento en la especie en todas las poblaciones estudiadas. Respecto a los
microelementos esenciales los valores encontrados fueron altos en B (161 mg/Kg), alto
en Cu (59 mg/Kg), alto en Mn (128 mg/Kg) y normal en hierro (239 mg/Kg); con
variaciones significativas de todos ellos entre poblaciones.
Respecto a las Vainas con y sin semillas, al ser comparadas, las variaciones mas
significativas se presentaron en los contenidos de proteína y lípidos. Siendo más ricas en
estos dos grupos nutricionales las vainas con semillas, con un aporte de 5% en lípidos y
9% de proteínas frente a las vainas sin semillas de 1,8 y 6% respectivamente. Las
vainas sin semillas presentaron la mayor riqueza de azúcares totales, mayoritariamente
no-reductores 58% respecto a las vainas con semillas de 30%. Tanto las hojas como las
vainas con y sin semillas estarían aportando con un 1,5%, 2,5% y 3, 5%
respectivamente con polifenoles, reconocidos por sus capacidades antioxidantes.
TABLA 4: COMPONENTES QUÍMICOS DE Prosopis chilensis (ALGARROBO)
GRUPOS DEL METABOLISMO PRIMARIO HOJAS
Vainas sin
semillas
Vainas con
semillas
HUMEDAD (%) 6,5 ± 0,3 8 ± 3 5 ± 1
MATERIA SECA (%) 93,5 ± 0,3 92 ± 3 95 ± 1
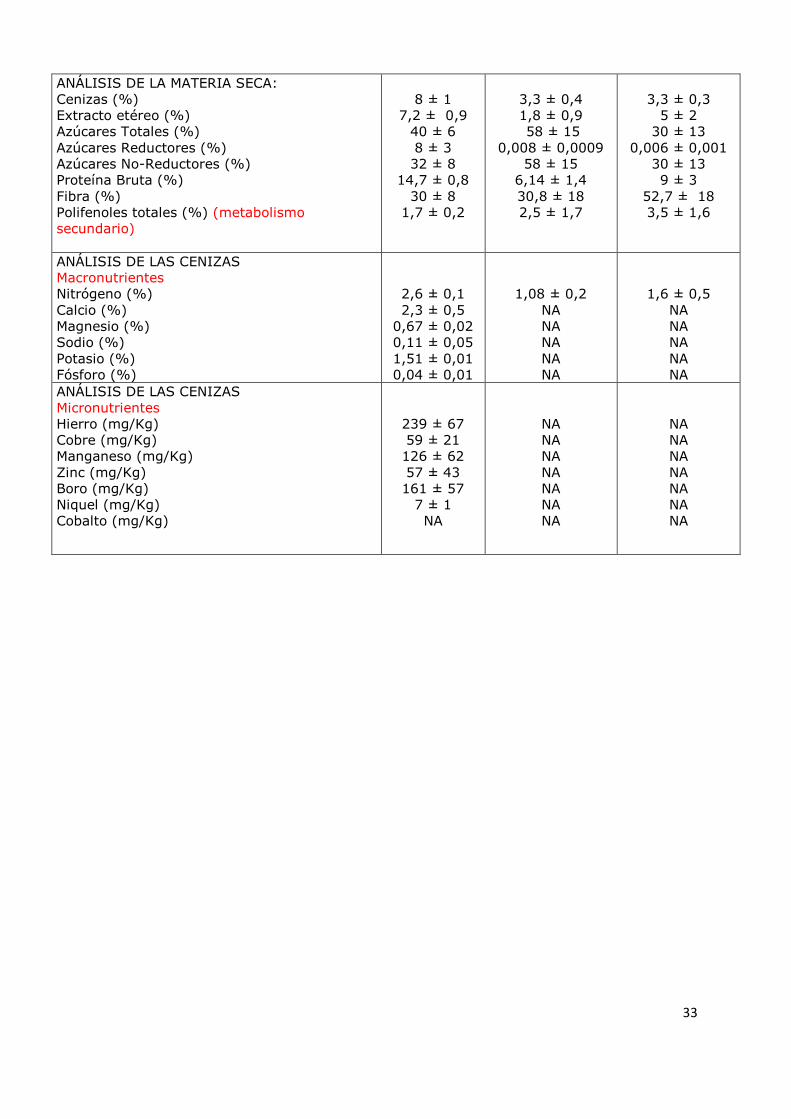
33
ANÁLISIS DE LA MATERIA SECA:
Cenizas (%)
Extracto etéreo (%)
Azúcares Totales (%)
Azúcares Reductores (%)
Azúcares No-Reductores (%)
Proteína Bruta (%)
Fibra (%)
Polifenoles totales (%) (metabolismo
secundario)
8 ± 1
7,2 ± 0,9
40 ± 6
8 ± 3
32 ± 8
14,7 ± 0,8
30 ± 8
1,7 ± 0,2
3,3 ± 0,4
1,8 ± 0,9
58 ± 15
0,008 ± 0,0009
58 ± 15
6,14 ± 1,4
30,8 ± 18
2,5 ± 1,7
3,3 ± 0,3
5 ± 2
30 ± 13
0,006 ± 0,001
30 ± 13
9 ± 3
52,7 ± 18
3,5 ± 1,6
ANÁLISIS DE LAS CENIZAS
Macronutrientes
Nitrógeno (%)
Calcio (%)
Magnesio (%)
Sodio (%)
Potasio (%)
Fósforo (%)
2,6 ± 0,1
2,3 ± 0,5
0,67 ± 0,02
0,11 ± 0,05
1,51 ± 0,01
0,04 ± 0,01
1,08 ± 0,2
NA
NA
NA
NA
NA
1,6 ± 0,5
NA
NA
NA
NA
NA
ANÁLISIS DE LAS CENIZAS
Micronutrientes
Hierro (mg/Kg)
Cobre (mg/Kg)
Manganeso (mg/Kg)
Zinc (mg/Kg)
Boro (mg/Kg)
Niquel (mg/Kg)
Cobalto (mg/Kg)
239 ± 67
59 ± 21
126 ± 62
57 ± 43
161 ± 57
7 ± 1
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA

34
5. CONCLUSIONES y RECOMENDACIONES
5.1. Garra de Léon (Leontochir ovallei):
Se logró desarrollar exitosamente un protocolo de propagación sexual (semillas) y
asexual (rizomas) para esta especie. Recomendamos a CONAF utilizar bolsas de
papel adosadas a las inflorescencias cada vez que haya desierto florido. Este
simple método permite colectar el 100% de semillas de cada inflorescencia.
Además recomendamos una desinfección simple con antifúngico tipo Captan,
deshidratación hasta peso constante y conservación en frío (4ºC), pues se verificó
que viabilidad de las semillas se mantiene intacta (semillas no recalcitrantes).
Se logró obtener una cantidad importante de rizomas de L. ovallei, que
continuaremos propagando y una parte importante de ellos, serán devueltos al
Parque Nacional Llanos de Challe.
En relación al análisis fitoquímico, dado que no existen analisis químicos
reportados en la literatura, estos resutados corresponderían al Primer Estandar
Nutricional de la especie L. ovallei realizados en hojas y semillas,. Hay que hacer
notar que estos resultados se basan en la última aparición ocurrida en esta planta
ocurrida en el año 2011, por lo cual sería recomendable repetir estos análisis con
plantas florecidas en años posteriores con el find e establecer un patrón químico d
ela especie.
En virtud del alto contenido de extracto etéreo (componentes lipídicos) y de
polifenoles observados tanto en hojas como en semillas, se sugiere realizar un
fraccionamiento para identificar presencia de otros metabolitos secundarios como
Alcaloides, Triterpenos, Esteroles, Antraquinonas, Flavonoides y Cumarinas.
Se sugiere determinar en el aceite las vitaminas liposolubles tales como
tocoferoles (Vitamina E), vitamina A y K, entre otras. En este estudio no se pudo
realizar este procedimiento debido a que se requería mayor cantidad de material y
las limitaciones propias de la planta (protegida) no posibilitaron colectar
demasiadas hojas. Lo que se encontró es que de 100 g de hojas, solo 20 gramos
corresponderán a materia seca. Con esta información se podría estimar la
cantidad de hojas que deberán ser recolectadas para futuros análisis.
5.2. Lucumillo (Myrcianthes coquimbensis):
Se logró optimizar un protocolo de germinación muy exitoso, donde resultó
fundamental remover el mesocarpio (pulpa) del fruto y colectar frutos maduros.
Aunque la siembra se puede realizar desde Agosto hasta Enero, el momento
óptimo es entre Octubre y Diciembre.
Se logró establecer un interesante protocolo de propagación de esquejes in vitro y
propagación de esquejes en cama caliente. En ambos casos es importante utilizar
individuos jóvenes, ojalá provenientes de germinaciones en vivero para disminuir
la presencia de microorganismos patógenos.
Los protocolos de propagación asexual posibilitaría la propagación masiva de M.
coquimbensis, el cuál dado su delicado estado de conservación e interesante
potencial como especie ornamental podría abstacer la demanda de viveristas u
otros interesados en utilizarla con este u otros fines comerciales.

35
En relación al análisis fitoquímico, los resultados obtenidos corresponderían al
Primer Estándar Nutricional reportado en esta especie, tanto de hojas, como de
mesocarpio y semillas.
La especie se presenta como un verdadero reservorio nutraceútico que debe
seguir estudiándose, sobre la base de los metabolitos secundarios identificados y
la riqueza polifenólica de sus hojas y frutos.
5.3. Algarrobo dulce (P. flexuosa):
Se logró desarrollar exitosamente un protocolo de germinación de semillas, donde
se recomienda implementar el sistema de colecta de semillas usando bolsas de
papel utilizado en L. ovallei, dado que son muy pocos los ejemplares que quedan
en pie, y las amenzas sobre esos individuos son muy elevadas, sobre todo la
ganadería no controlada y eventos cada vez mas prolongados de sequía. Por lo
mismo, creemos necesario que CONAF refuerce sus estrategias para colectar
semillas de esta especie. Se aconseja colectar semillas en el punto indicado en la
metodología (carretera Caldera-Copiaó) dado el excelente estado sanitario en que
se encuentran dichos individuos.
Para la propagación asexual se recomienda usar individuos jóvenes, o estimular la
generación de brotes nuevos mediante podas in situ de los árboles adultos a partir
de los cuales luego se puedan colectar esquejes tiernos. Si esto se realiza, hay
que asegurar el traslado de los esquejes sumergidos en agua y luego realizar una
desinfección de los esquejes de moderada a fuerte. Es importante hacer notar que
P. flexuosa no requiere el uso de enraizantes pues tiene buan capacidad para
enraizar naturalmente.
En relación al análisis fitoquímico, los resultados obtenidos corresponderían al
Primer Estandar Nutricional de la especie en Chile (existen reportes de análisis de
la especie en Argentina, pero sólo en vainas).
Se destaca en las vainas un potencial alimentario interesante por su alto
contenido en Azúcares totales solubles, mayoritariamente no reductores, y
proteína, con un perfil lipídico que aporta ácido oleico, linoleico y linolénico, estos
dos últimos ácidos grasos esenciales.
Se recomienda estudiar la digestibilidad y estabilidad química de las vainas, junto
con identificar la presencia de metabolitos secundarios que pudiesen generar
toxicidad, así como completar los estudios en función de los aportes nutricionales
en vitaminas.
5.4. Algarrobo (P. chilensis):
El comportamiento de P. chilensis fue muy similar a P. flexuosa. Se logró
desarrollar un protocolo óptimo para germinar tanto sus semillas como propagarlo
asexualmente.
Se aconseja mantener el riego periódico de las plántulas aclimatadas de P.
chilensis pues aunque algunas plantas muestren signos de deshidratación severa,
la capacidad de recuperación de este árbol en condiciones de vivero es
extraordinaria. Un signo evidente que las plantas aún están vivas es el color de su
tallo. Si este se encuentre verde la planta está viva mas allá que carezca
completamente de hojas. Esta característica de recuperación también la presenta
P. flexuosa por lo que podríamos estar ante una estrategia de supervivencia
conservada en el género Prosopis spp.
En relación al análisis fitoquímico, los resultados obtenidos corresponderían a una
actualización de los estándares nutricionales conocidos hasta ahora. El último
reporte químico, con datos algo más completos de esta especie es el reportado
por F.M. Galera (2000), texto en los Depósitos de Documentos de la FAO.

36
En acuerdo con F.M Galera (2000), en P. chilensis se destaca en las vainas un
potencial alimentario interesante por su alto contenido en Azúcares totales
solubles, mayoritariamente no reductores, y proteína. Estudios futuros de esta
especie debieran enforcarse a una posible industrialización de sus vainas con fines
alimentarios.
Se recomienda completar los estudios en función de los aportes nutricionales en
vitaminas, polifenoles y otros componentes del metabolismo secundario que
podrían estar presentes en el extracto etéreo, así como la caracterización de los
azúcares individuales con fines nutraceúticos.

37
6. PRODUCTOS OBTENIDOS
6.1 Actividades de Difusión
6.1.1 DEVELOPMENT OF PROTOCOLS TO PROPAGATE FOUR NATIVE PLANTS
ENDANGERED BY HUMAN ACTIVITY AND HARSH ARID ENVIRONMENTAL
CONDITIONS OF NORTHERN CHILE. Marta Vargas, Elda Jofré, Catherine Rojas,
Paloma Gachón, Carlos Navarrete, Jaime Bravo and Cristian Ibáñez. Poster
presentado en la VII Reunión de Biología Vegetal. Pucón, 3-6 Diciembre
2012 (Ver Anexo n° 14)
6.1.2 PROPAGATION STRATEGY FOR Myrcianthes coquimbensis, AN ENDANGERED
SPECIE OF COQUIMBO COASTAL REGION. Marta Vargas, Catherine Rojas, Paloma
Gachón, Carlos Navarrete, Jaime Bravo and Cristian Ibáñez. Poster presentado
en la VII Reunión de Biología Vegetal. Pucón, 3-6 Diciembre 2012 (Ver Anexo
n° 15).
6.1.3 FIRST CHEMICAL APPROACHES TO ANALYZE FOUR ENDANGERED PLANTS FROM
NORTH OF CHILE. Fabiola Jamett, Manuel Tapia, Esthefany Muñoz, Jessenia
Velásquez, Carlos Navarrete, Jaime Bravo, and Cristian Ibáñez. Poster
presentado en la VII Reunión de Biología Vegetal. Pucón, 3-6 Diciembre
2012 (Ver Anexo n° 16).
6.1.4 ESTABLECIMIENTO DE PROTOCOLOS DE PROPAGACIÓN IN VITRO E
IDENTIFICACIÓN DE BIOCOMPUESTOS EN ESPECIES NATIVAS DE ALTO VALOR
ECOLÓGICO DEL NORTE CHICO. Crisitian Ibáñez y Fabiola Jamett. Oradores en
el II Encuentro Temático del Fondo de Investigación del Bosque Nativo
“Aportando al conocimiento de Formaciones Xerofíticas”. Iquique, 13 de
Agosto 2013 (Anexo n° 17).
6.1.5 EVALUACIÓN DE LA GERMINACIÓN DE GARRA DE LEÓN (Leontochir ovallei), UNA
ESPECIE EN PELIGRO DE EXTINCIÓN. Elda Jofré, Marta Vargas, Alejandra Araya,
Maricela Rojas, Ana María Vásquez, Carlos Navarrete, Fabiola Jamett y Cristian
Ibáñez. Poster presentado en el III Congreso Nacional de Flora Nativa.
Santiago, 5-7 Septiembre 2013 (Anexo n° 18).
6.1.6 CARACTERIZACIÓN QUÍMICA DE Bomarea ovallei (Garra de León), Myrcianthes
coquimbensis (Lucumillo), Prosopis flexuosa (Algarrobo dulce) y Prosopis chilensis
(Algarrobo). Fabiola Jamett. Orador en el III Congreso Nacional de Flora
Nativa. Santiago, 5-7 Septiembre 2013 (Anexo n° 19).
6.1.7 ESTABLECIMIENTO DE PROTOCOLOS DE PROPAGACIÓN DE CUATRO ESPECIES
NATIVAS DE ALTO VALOR ECOLÓGICO DE LA REGIÓN ÁRIDA y SEMIÁRIDA DE
CHILE. Cristian Ibáñez. Orador en el III Congreso Nacional de Flora Nativa.
Santiago, 5-7 Septiembre 2013 (Anexo n° 20).
6.1.8 DEVELOPING A CUTTING PROTOCOL FOR PROPAGATION OF Prosopis flexuosa, AN
ENDANGERED SPECIE FROM NORTH OF CHILE. Marta Vargas, Elda Jofré,
Alejandra Araya, Maricela Rojas, Ana María Vásquez, Carlos Navarrete y Cristian
Ibáñez. Poster presentado en la VIII Reunión de Biología Vegetal. Pucón,
2-5 Diciembre 2013 (Ver Anexo n° 21).

38
7. BIBLIOGRAFÍA
Abugoch JLE (2009). Chapter 1. Quinoa (Chenopodium quinoa Willd.): Composition,
Chemistry, Nutritional, and Functional Properties. Advances in Food and Nutrition
Research, 58:1-31.
Amigo J & Ramírez C (1998). A bioclimatic classification of Chile: woodland communities
in the temperate zone. Plant Ecology 136: 9-26.
Astaburuaga R (2004). El agua en las zonas áridas de Chile. AQR. Pontificia Universidad
Católica de Chile. 57:68-72.
Barbedo CJ & Cicero SM (2000). Effecs of initial quality, low temperature and ABA on the
storage of seeds of Inga uruguensis a tropical species with recalcitrant seed. Seed
Science and Technology 28:793-808.
Beauchamp GK, Keast RSJ, Morel D, Lin J, Pika J, Han Q, Lee C-H, Smith AB & Breslin
PAS (2005). Ibuprofen-like activity in extra-virgin olive oil. Nature 437: 1-2.
Brenner SE (2001). A tour structural genomics. Nature Reviews Genetics 2:801-809.
Calderón-Baltierra X, Pérez F & Rotella A (1993) Micropropagación de una especie chilena
en peligro de extinción: Gomortega keule (Mol.) Baillon (Magnoliopsidae,
Gomortegaceae). BOSQUE 14 (1): 23-28.
Caro LA, Polci PA, Lindstrom LI, Echenique CV & Harnández LF (2002). Micropropagation
of Prosopis chilensis (Mol.) Stuntz from young and mature plants. Biocell, 26 (1):
25-33.
CBD (2011). About the Convention. http://cbd.int/convention/about.shtml. Visitada el
16.02.2011
CBD (2006). Perspectiva Mundial sobre Diversidad Biológica 2. Montreal. 81p.
Chapin III FS, Zavaleta ES, Eviner VT, Taylor RL, Vitousek PM, Reynolds HL, Hooper DU,
Lavorel S, Sala OE, Hobbie SE, Mack MC & Díaz S (2000). Consequences of
changing biodiversity. Nature 405: 234 – 242.
CIREN (2011). Determinación de la erosión actual y potencial del territorio de Chile.
http://www.ciren.cl/cirenxml/noticias/default.asp?a=5&id=647. Visitada el 25 de
Febrero de 2011.
Cob J, Sabja AM, Ríos D, Lara A, Donoso PJ, Arias L & Escobar B. (2010). Potencial de la
organogénesis como estrategia para la masificación in vitro de Persea lingue en la
zona centro-sur de Chile. BOSQUE 31 (3): 202-208.
Conger BV (1981). Cloning agricultural plants via in vitro techniques. CRC Press, Inc.
Boca Raton, Florida, 273.
De Fossard RA, Baker PK, Bourne RA (1977). The organ culture of nodes of four species
of Eucalyptus. Acta Horticulture, 78:157-65.
Deb CR & Tandom P (2004). Factors influencing initiation of embryogenic cultures in
Pinus kesiya Royle ex Gord. Indian Journal of Biotechnology 3:589-593.
Donoso C (1993). Bosques templados de Chile y Argentina. Variación, estructura y
dinámica. Ecología forestal. Editorial Universitaria. Santiago. 484 p.
Dodds JH (1991). In Vitro Methods for Conservation of Plant Genetic Resources.
Chapman & Hall. London 240 pp.
Engelman F (1997). In Vitro conservation Methods. Chapter 6. Biotecnology and Plant
Genetics Resources. Conservation and Use. Edited by JA Callow, BV Ford-Lloyd
and HJ Newbury. CAB International. 308 p.
Favier V, Falvey M, Rabatel A, Praderio E, Lopez D (2009). Interpreting discrepancies
between discharge and precipitation in high-altitude area of Chile’s Norte Chico
region (26–32 S). Water Resource Research 45(2):W02424
Ferreyra R & Sellés G (1997). Manejo del riego en condiciones de restricción hídrica.
Instituto de Investigaciones Agropecuaria-La Platina. Serie La Platina. N°67, 36p.
Finelstein RR & Crouch ML (1984). Precociously germinating rapeseed embryos retain
characteristics of embryyogeny. Planta 162:125-131.

39
García et al (2011). Preliminary notes on the ecology, distribution and conservation
status of Myrchianthes coquimbensis: A globally endangered endemic shrubs of
the Chilean Coastal Desert. Plant Ecology and Diversity: submitted.
Garreaud R, Barichivich J, Christie DA & Maldonado A (2008). Interannual variability of
the coastal fog at Fray Jorge relict forests in semiarid Chile. Journal of Geophysical
Research, Vol 113: G04011, doi:10.1029/2008JG000709
Garreaud R,Vuille M, Compagnucci R & Marengo J (2009). Present-day South American
climate. Palaeogeography, Palaeoclimatology, Palaeoecology 281 (3-4): 180-195.
Gomes F & Canhoto J (2003). Micropropagation of Eucalyptus nitens Maiden (Shining
Gum). In Vitro Cellular & Developmental Biology. Plant, 39:3: 316-321.
Gutiérrez AG, Barbosa O, Christie DA, Del-Val E, Ewing HA, Jones CG, Marquet PA,
Weathers KC, Armesto JJ (2007). Regeneration patterns and persistence of the
fog dependent Fray Jorge forest in semiarid Chile during the past two centuries.
Global Change Biology 14: 1–16.
Gutiérrez JR, Meserve PL, Kelt DA, Engilis Jr. A, Previtali MA, Milstead MB, Jaksic FM
(2010). Long-term research in Bosque Fray Jorge National Park: Twenty years
studying the role of biotic and abiotic factors in a Chilean semiarid scrubland.
Revista Chilena de Historia Natural 83: 69-98.
Hechenleitner P, Gardner M, Thomas P, Echeverria C, Escobar B, Brownless P, Martinez
C. ( 2005). Plantas amenazadas del Centro-Sur de Chile. Universidad Austral de
Chile. Real Jardín Botánico de Edimburgo. 187 p.
Infante L (1989). Estudio de germinación de palma chilena (Jubaea chilensis (Mol)
Baillon). Memoria Ingeniería Forestal. Santiago, Chile. Universidad de Chile.
Facultad de Ciencias Agrarias y Forestales. 132 p.
Jain SM & Häggman H (2007). Protocols for Micropropagation of Woody Trees and Fruits.
Springer-The Netherlands. 559p.
Kalthoff N, Fiebig-Wittmaack M, Meißner C, Kohler M, Uriarte M, Bischoff-Gauß I &
Gonzales E (2006). The energy balance, evapo-transpiration and nocturnal dew
deposition of an arid valley in the Andes. Journal of Arid Environments 65 (3):
420–443.
Kehr J (2001). High resolution spatial analysis of plant systems. Curr. Opin. Plant Biol.
4:197–201.
Lazo I, Gnocchio R, Cofre H, Vilina Y & Iriarte A (2008). Capítulo II. Nuestra diversidad
biológica. In: Biodiversidad de Chile. Patrimonio y desafíos. 2da edición. CONAMA.
Editado por Ocho Libros. 639 p.
Lu C, Ruan Y & Bridgen M (1995). Micropropagation procedures for Leontochir ovallei
Plant Cell, Tissue and Organ Culture, 42 (2): 219-221.
Luebert F & Pliscoff P (2006). Sinopsis bioclimática y vegetacional de Chile. Santiago de
Chile: Editorial Universitaria, 316 pp.
Muñoz C (1966). Flores silvestres de Chile. Edic. Univ. Chile-München, 245 p.
Muñoz M (2000). Novedades en la familia Alstroemeriaceae. Gayana Botanica 57(1): 55-
59.
Machuka J (2004). Agricultural genomics and sustainable development: perspectives and
prospects for Africa. African Journal of Biotechnology, 3:2: 127-35.
Matthews P (1999). Vegetative Propagation from Stem Cuttings, Leaves and Roots. En: A
Colour Atlas of Plant Propagation and Conservation. Bowes, B.G. (ed.), Manson
Publishing, London, pp. 58-68.
Moco S, Bino R, De Vos R, Vervoort J (2007). Metabolomics Technologies and metabolite
identification. Trends in Analytical Chemistry, 26:9:855-66.
Montpetit B (2003). Proteomics in the Post-Genomics Era: Weighing in the Data.
BioTeach Journal, 1:9-12.
Morini S (2000). In vitro culture of Osmunda regalis fern. Journal of Horticultural Science
& Biotechnology 75(1): 31-34.
Oliver F (2002). Metabolomics – the link between genotypes and phenotypes. Plant
Molecular Biology, 48: 155–171.
Pammenter NW, Berjak P, Farrant JM, Smith MT & Ross G (1994). Why do stored
hydrated recalcitrant seeds die?. Seed Science Researcher 4:187-191.

40
Pardos K (1984). Un programa de mejoramiento genético en Prosopis tamarugo y
Prosopis chilensis. Documento de Trabajo No. 1, CONAF/FAO, Santiago de Chile,
28 p.
Peacock B & Dokoozlian N (1997). Irrigation strategies for Crimson Seedless. California
Table Grape Commission. Viticulture Research.
Pierik RLM & Steegman HHM (1986). Adventitious plantlet regeneration from floral stem
explants of Nerine bowdenii W. Watts. Netherlands journal of agricultural science
34 (2):217-223.
Pitt J, Roth W, Lacor P, Smith AB, Blankenship M, Velasco P, De Felice F, Breslin P, Klein
WL (2009). Alzheimer's-associated Aβ oligomers show altered structure,
immunoreactivity and synaptotoxicity with low doses of oleocanthal. Toxicology
and Applied Pharmacology 240: 189–197.
R (2005). Development Core Team. R: A language and environment for statistical
computing, reference index version 2.2.1. R Foundation for Statistical Computing,
Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org.
Ravenna P, Teillier S, Macaya J, Rodríguez R, Zöllner O (1998). Categorías de
conservación de las plantas bulbosas nativas de Chile. Boletín MNHN 47: 47-68.
Razdan MK & Cocking EC (1997). eds. In Conservation of Plant Genetic Resources In
Vitro Vol. 1, M/S Science, USA, p. 153-174
Sabja AM; Ortiz O & Trivino C (2008). Avances de clonación in vitro de árboles adultos de
raulí (Nothofagus alpina Poepp. et Endl.) Oerst.) para propagación comercial.
Agrociencia 42 (5): 595-603.
Sala OE, F. Stuart Chapin III, Juan J. Armesto, Eric Berlow, Janine Bloomfield, Rodolfo
Dirzo, Elisabeth Huber-Sanwald, Laura F. Huenne e, Robert . ac son, Ann
inzig, Ri eemans, avid M. odge, Harold A. Mooney, Mart n Oesterheld, N.
LeRoy Poff, Martin T. Sykes, Brian H. Walker, Marilyn Walker, Diana H. Wall
(2000). Global Biodiversity Scenarios for the Year 2100, Science 287: 1770-1774
Sánchez-Olate M, Ríos D, Pedraza M, Pereira G, Castellano H & Escobar R (2004).
Propagación in vitro de Nothofagus procera((Poepp. et Endl.) Oerst.) a partir de
embriones aislados. BOSQUE 25 (1): 123-128.
Santibáñez F, Roa P & Santibáñez F (2008). Capítulo I. El medio físico. En: Biodiversidad
de Chile. Patrimonio y desafíos. 2da edición. CONAMA. Por Ocho Libros. 639 p.
Schmeda-Hirschmann G, Loyola JI, Sierra J, Retamal R, Rodríguez J (1992). Hypotensic
effect and enzyme inhibitory activity of Mapuche medicinal plant extracts.
Phytotherapy Research 6: 184-188.
Schmeda-Hirschmann G, Loyola JI, Razmilic I, Rodriguez J, Pacheco P, Theoduloz C
(1993). La Farmacopea Mapuche, una fuente de productos biológicamente activos.
UNIVERSUM 153-178.
Scherling C, Roscher C, Giavalisco P, Schulze E & Weckwerth W (2010). Metabolomics
Unravel Contrasting Effects of Biodiversity on the Performance of Individual Plant
Species. PLos ONE 5 (9), e12569.
Schwab W (2003). Metabolome diversity: too few genes, too many metabolites?
Phytochemistry 62: 837–849.
Shepherd LVT, Alexander CA, Sungurtas JA, McNicol JW, Stewart D &, Davies HV (2010).
Metabolomic analysis of the potato tuber life cycle. Metabolomics 6: 274–291.
Shirley Z-S, Taharat Y, Manashi B, Archana C, Joe AV & Debasis B (2007) Berry
anthocyanins as novel antioxidants in human health and disease prevention. Mol.
Nutr. Food Res. 51: 675 – 683.
Shiloach J, Reshamwala S, Noronha SB & Negrete A (2010). Analyzing metabolic
variations in different bacterial strains, historical perspectives and current trends –
example E. coli. Current Opinion in Biotechnology, 21: 21–26.
Squeo FA, Olivares N, Olivares S, Pollastri A, Aguirre E, Aravena R, Jorquera C &
Ehleringer JR (1999). Grupos funcionales en arbustos desérticos definidos en base
a las fuentes de agua utilizadas. Gayana Bot. 56:1-15.
Squeo FA, Arancio G & Gutiérrez JR (2001). Libro rojo de la flora nativa y de los sitios
prioritarios para su conservación: región de Coquimbo. Ediciones Universidad de
La Serena, La Serena. 372 pp.

41
Squeo FA, Arancio G & Gutiérrez JR (2008). Libro rojo de la flora nativa y de los sitios
prioritarios para su conservación: región de Atacama. Ediciones Universidad de La
Serena, La Serena. 466 pp.
Stoll A, Maldonado A, Gascoin S, Moncada X, Tracol Y, Rioseco T (2011). Water in the
desert: spatial-temporal influences on vegetation patterns in northern Chile
(20°S-32°S). Plant Ecology & Diversity, submitted
Taiz & Zeiger 2007. Plant Physiology. Fourth Edition. Sinauer Associations Inc. 764 p
The Arabidopsis Genome Initiative (2000). Analysis of the genome sequence of the
flowering plant Arabidopsis thaliana. Nature 408: 796-815.
Theoduloz C, Rodríguez JA, Pertino M & Schmeda-Hirschmann, G (2009). Antitumor
activity of jatrophone and jatropholone derivatives. Planta Medica 75, 1520-1522.
Troncoso H, Jamett F, Benavides A, Astorga M (2006). Caracterización química de aceites
de oliva producidos en la Región de Coquimbo. Instituto de Investigaciones
Agropecuaria – Intihuasi. Boletín INIANº 153, 30p.
Uribe-Moraga M & Cifuentes L (2004). Aplicación de técnicas de cultivo in vitro en la
propagación de Legrandia concinna. Bosque 25 (1): 129-135.
van Husen C (1967). Klimagliederung in Chile auf der Basis von Häufigkeitsverteilungen
der Niederschlagssummen. Freiburger Geographische Hefte 4: 0–113.
Watson JD & Crick FHC (1953). A Structure for Deoxyribose Nucleic Acid. Nature 171:
737-738.
Wang PJ & Charles A (1991). Micropropagation through meristem culture. In
biotechnology in agriculture and forestry. Vol. 17: 32-52. Hightech and
micropropagation. Edited by Y.P.S. Bajaja.. Springer Verlag Berlin Heidelberg
Venter Graig et al, (2001) The Sequence of the Human Genome. Science 291 (5507):
1304-1351.
Zhao Y, Li XL, Amarasinghe AAY & Yang YS (2009). Influence of psychological state of
fresh and dry explants on in vitro culture response in rice (Oryza sativa L.) The
Journal of Agricultural Sciences 4 (1): 1-9.
Zuloaga F, Morrone O& Belgrano MJ (eds.). 2008. Clusiaceae. Catálogo de las Plantas
vasculares del Cono Sur (Argentina, sur de Brasil, Chile, Paraguay y Uruguay).
Monogr. Syst. Bot. Missouri Bot. Gard. 107 (2):1932-1933.

42
8. ANEXOS
PROPAGACIÓN
ANEXO n° 1: Colecta de semillas de Garra de León (L. ovallei).
a) Las semillas de L. ovallei son depredadas por la fauna silvestre y colectadas
intensamente por floristas/viveristas. Es por ello que se recomienda proteger las
inflorescencias con una bolsa de papel, después que la inflorescencia se haya
formado.
b) En la Fig. 1A y 1B se muestran las inflorescencias de L. ovallei en el Parque
Nacional Llanos de Challe en Noviembre de 2011, el último año que se les vio
florecer. La Fig. 1C muestra la inflorescencia ya cubierta por la bolsa de papel
(Diciembre de 2011) y las Fig. 1D y 1E, muestran el excelente estado en que
quedan las inflorescencias protegidas por las bolsas de papel y que permitió
recuperar el 100% de semillas (Febrero de 2012).
c) La fig. 1F muestra una inflorescencia no protegida con bolsa de papel y cuyas
semillas fueron consumidas por algún animal silvestre.
d) En general, las semillas tienen una muy buena sanidad, no presentando larvas ni
presencia de microrganismos. Una vez colectadas, las semillas deben secarse
hasta lograr peso constante y pueden mantenerse en frío (4°C).o temperatura
ambiente hasta su utilización.
1A Parque Nacional Llanos de Challe. Noviembre 2011

43
1B
1C
Parque Nacional Llanos de Challe. Noviembre 2011
Método para colectar de semillas

44
1E
1D Método para colectar de semillas
El Método permitió controlar la indehiscencia explosiva.

45
1F Inflorescencia sin bolsas de papel cuyas semillas se perdieron o fueron
consumidas por animales y

46
ANEXO n° 2: Protocolo de Germinación in vitro de Garra de León (L ovallei)
a) Desinfección: sumergir las semillas en etanol 70% durante 1 minuto. Enjuagar
con agua destilada estéril. Luego embeber en Hipoclorito de Sodio 1,5% por 15
min. Realizar cuatro enjuagues con agua destilada estéril. Realizar un corte
superficial en la testa de cada semilla antes de sembrar.
b) Medio de siembra: A placas Petri de 90mm, agregar 30 mL de medio Murashige
and Skoog 1962 (MS), Sacarosa (2%), ácido giberélico (5mg/L), 6-
Bencilaminopurina (0,5 mg/L), Cloruro de Sodio (20 uM) y agar (8 g/L), pH 5,8.
Medir pH antes de agregar el agar
c) Mantener las placas con semillas durante dos meses a 21°C, fotoperiodo 16h/8h
(luz/oscuridad), intensidad de 120 umolm-2s-1 (Fig. 2A).
d) Una vez germinadas (Fig 2B y 2C), traspasar las plántulas a sustrato estéril
arena:vermiculita (2:1) y mantener en las mismas condiciones de crecimiento
durante dos meses. Las plántulas de L. ovallei son muy sensibles a la
deshidratación por lo cual es fundamental que se utilicen frascos plásticos, donde
se pueda ir graduando el contacto de las plantas con su medio externo (Fig. 2D).
Como protección, también se pueden utilizar bolsas de plástico, a las que se les
corta gradualmente sus esquinas.
e) Traspasar a invernadero, idealmente, con condiciones controladas de luz y
temperatura. Regar poco, y cubrir con una bolsa de plástico para minimizar la
deshidratación excesivamente. En la Fig. 2E se muestra una planta de 6 meses
crecida en invernadero con dos rizomas ya desarrollados. Hay que indicar que un
número importante de las plantas transferidas al invernadero sufre una
deshidratación severa, aún con los cuidados antes mencionados (Fig. 2F). Sin
embargo, está comprobado que si se continúa con el régimen de riego, al cabo de
unas semanas los rizomas volverán a crecer y darán inicio a nuevas plantas.
f) Con este protocolo se obtuvo un 13% de germinación de semillas, lejos el más
alto.

47
12
2A
2B
Germinación in vitro en la cámara de crecimiento con condiciones
controladas
Cotiledón y radícula de semillas germinación in vitro

48
2C
2D
Plantas de 5 meses aclimatadas al
interior de la cámara de crecimiento
Plantas en proceso de
aclimatación
Plántula de 1 mes de edad

49
2E
Plantas de seis meses de edad, con dos rizomas ya formados
Plantas de ocho meses de edad, transferidas a invernadero y
que sufrieron una severa deshidratación. Sin embargo, todas
estas plantas se recuperan al cabo de 1 mes
aproximadamente.
2F

50
ANEXO n° 3: Protocolo de multiplicación de rizomas de Garra de León (L.
ovallei)
a) Después de 8 meses (Fig. 3A), una planta de L. ovallei debería generar, en
promedio, cinco rizomas (Fig. 3B).
b) Saturar con abundante agua el sustrato de arena:vermiculita (2:1) en que se
encuentran las plantas de L. ovallei (Fig 3C).
c) Remover con cuidado la zona radicular e iniciar la propagación de las plántulas
cortando las secciones de tallos que unen cada rizoma entre sí. Procurar mantener
unida la respectiva estructura de reserva a su rizoma (Fig. 3D).
d) Trasladar el rizoma y su estructura de reserva a una nueva bolsa o maceta con
sustrato de arena:vermiculita (3:1) (Fig. 3E)
e) Regar 2 veces por semana hasta que comience a crecer una nueva planta.
Mantener el mismo régimen de riego por 4 semanas y luego distanciar el riego a 1
vez por semana.
3A
Plantas de ocho meses de edad,
aclimatada en invernadero

51
3C
3B Rizomas obtenidos de una planta de ocho meses de edad
Rizomas obtenidos de una planta de ocho meses de edad

52
3E Rizomas en pleno crecimiento, a 1 mes de haber sido dividido y plantado
3D Disección de rizomas

53
ANEXO n° 4: Protocolo de germinación semillas de Lucumillo (Myrcianthes
coquimbensis).
a) Colectar frutos maduros. Estos se reconocen por tener una tonalidad
rojo/naranja intenso, y porque se desprenden fácilmente del arbusto (Figs.
4A,. 4B)
b) Remover el mesocarpio (pulpa) de las semillas (Fig. 4C)
c) Sembrar directamente sobre un sustrato de tierra:arena (2:1) humedecido.
Sembrar a 1.5-2 cm de profundidad (Fig. 4D)
d) Regar diariamente la primera semana, las siguientes dos semanas hacerlo
cada dos días, para luego continuar con riegos 2 veces por semana y
mantener luego 1 vez por semana (Fig. 4E)
e) Con este protocolo se consiguió un 80% de germinación.
4A Detalle de las hojas y frutos maduros

54
4B
4C
Semillas maduras Cotiledón y
embrión Semillas inmaduras
Mesocarpio
a = Semilla con mesocarpio
a b c
b = Semilla con radícula deshidratada
c = Semilla sin mesocarpio

55
4E
4D Semilla germinada mostrando su gran cotiledón carnoso
Plántulas creciendo en sombreadero

56
ANEXO n° 5: Protocolo de propagación ex vitro por esquejes en Lucumillo (M.
coquimbensis)
a) Cortar esquejes de la mitad superior de ramas de lucumillo con 6 pares de yemas
(5A).
b) Lavar los esquejes durante 5 minutos con agua corriente.
c) Sumergir los esquejes en una solución antifúngica (Eco Opción o Captan) durante
15 minutos.
d) Cortar los esquejes dejándolos con dos pares de yemas. Mantener las hojas del par
superior de las yemas y cortar las hojas del par inferior.
e) Sumergir la base del explante en Ácido Indolbutírico (IBA Root) a 10.000 mg/L por
5 segundos.
f) Colocar los esquejes en un sustrato de tierra de hojas o vermiculita en una cama
de propagación a 21°C (Figs. 5B, 5C, 5D)
g) Mantener durante 60 días y traspasar los esquejes enraizados (Figs. 5D, 5E) a
bolsas o vasos plásticos con tierra de hojas (Fig. 5G).
h) Después de 30 días, las nuevas plantas deberían aumentar considerablemente su
masa radicular (Fig. 5H).
i) Con este protocolo se obtuvo un 85% de enraizamiento final.

57
5A
5B Cama caliente mostrando su sistema de calefacción
Esqueje antes de ser seccionado

58
5D
5C
Embebimiento de explantes en Ácido Indolbutírico (IBA root)
Explantes montados sobre la cama caliente

59
5E
5F
Explante enraizado de 1 mes de edad
Explantes enraizados de 2 meses de edad

60
5G
5H
G
Explantes enraizados transferidos a vasos de
plástico para iniciar su aclimatación
Explante mostrando su masa radicular en
crecimiento después de estar 1 mes en invernadero

61
ANEXO n° 6: Protocolo de propagación in vitro de Lucumillo (M. coquimbensis)
a) Cortar explantes de 3-5 cm que contengan al menos 1 par de yemas laterales
(Fig. 6A). Se recomienda utilizar como plantas madres, individuos jóvenes (6
meses a 1 ½ año de edad), desarrolladas a partir de semillas en invernadero.
La utilización de esquejes provenientes de plantas silvestres, resultó en una
contaminación permanente que fue imposible de erradicar (6B).
b) Lavar los explantes con agua corriente durante 5 minutos.
c) Desinfectar los explantes sumergiéndolos en etanol 70% durante 30 segundos.
Luego en Hipoclorito de Sodio 1,5% + 2 gotas tween por 10 minutos.
Posteriormente realizar tres enjuagues con agua destilada estéril.
d) Medio de cultivo para el establecimiento: En frascos de vidrio adicionar un
volumen final de 15ml de medio de establecimiento. Cada litro de medio de
cultivo está constituido por medio básico de Murashige & Skoog 1962 (MS),
sacarosa (2%), 6-bencilaminopurina (14,1 µM), Polivinilpirrolidona (PVP,
1mg/L) y Agar-Agar (8,0 g/L), pH 5,8 previo a la adición del agar. Autoclavar
el medio a 121°C, presión de 1Kg/cm2 y por 20 min. Traspasar el explante
desinfectado a este medio y mantener en cámara con temperatura 21°C,
fotoperiodo largo (16h luz/8h oscuridad), 70% H.R, 120 µmolm-2s-1.
e) A los 30 días, transferir los explantes con sus yemas brotadas a medio fresco
de cultivo. Mantener por otros 30 días y luego pasar a la etapa de
enraizamiento.
f) Enraizamiento: Colocar los brotes desarrollados en frascos de vidrio con 15 mL
de medio de enraizamiento. Cada litro de medio de enraizamiento está
constituido por sacarosa (3%), Polivinilpirrolidona (PVP, 1mg/L), Kinetina
(1mg/L), Ácido Indolbutírico (AIB, 5 mg/L) y Carbón activado (1g/L).
Autoclavar el medio según lo indicado en punto d. Traspasar el brote
desarrollado a este medio y mantener por 1 mes en cámara de crecimiento
con condiciones controladas de luz, temperatura y humedad relativa ya
descritas en el punto d. Pasado el mes, subcultivar en un medio fresco y
mantener por 1 mes más (Fig. 6C y 6D).
g) Transferencia a tierra y aclimatación: Extraer las plántulas de los tubos, lavar
raíces y extraer el agar. Plantar en vasos plástico cerrados con sustrato
autoclavado de arena:vermiculita (2:1) o sólo de vermiculita y mantener
durante 1 ½ mes en cámara de crecimiento, facilitando gradualmente el
intercambio gaseoso y con riegos cada 7 días con solución nutritiva Hoagland.
Luego transferir a invernadero (Fig. 6E).
h) i) Con este protocolo se obtuvo un 80% de enraizamiento final.
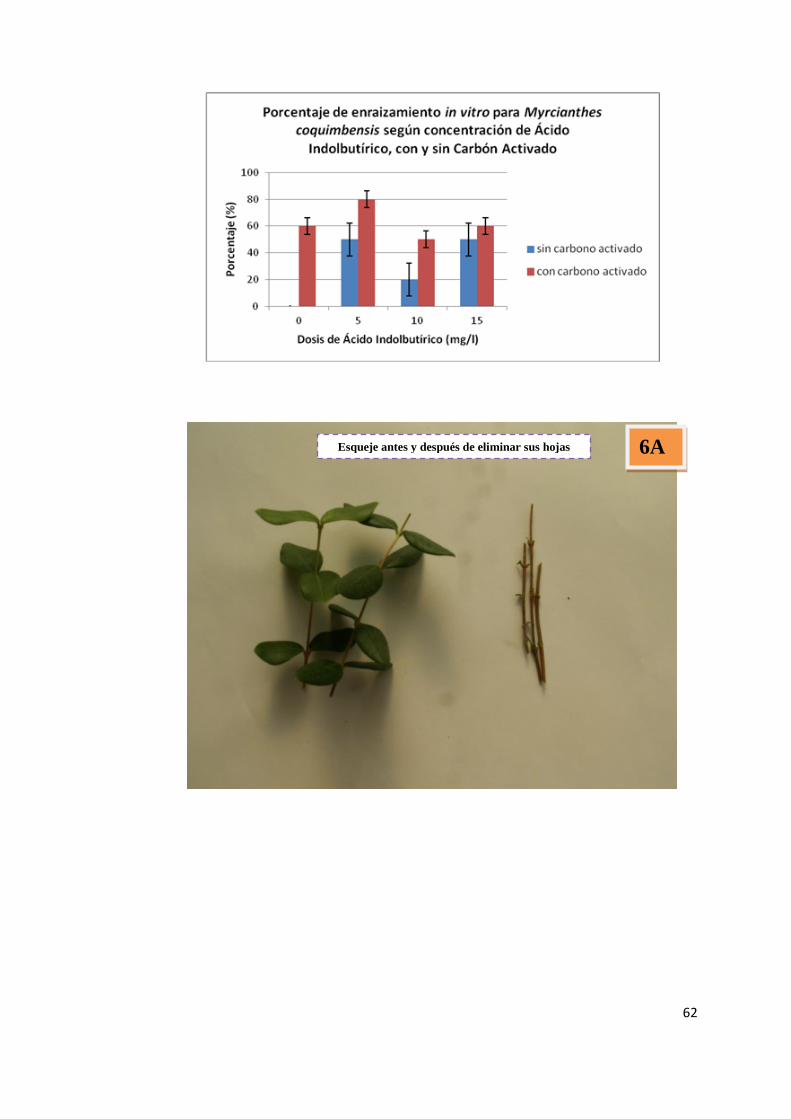
62
6A
G
Esqueje antes y después de eliminar sus hojas

63
6B
6C
Esqueje adulto contaminado a pesar de la desinfección realizada
Esqueje enraizado in vitro y en presencia de carbón activado

64
6D
G
Esqueje enraizado in vitro y en ausencia de carbón activado.
Las raíces son más cortas.
6E
G
Plántulas de 3 meses de edad ya aclimatadas en invernadero

65
ANEXO n° 7: Protocolo de germinación semillas de Algarrobo dulce (Prosopis
flexuosa)
a) Extraer las semillas de las vainas (Figs. 7A y 7B), eliminando las semillas
vanas o comidas por insectos. Almacenar en frío (4°C) por mínimo 1 mes.
b) Escarificar las semillas con ácido sulfurico (H2SO4) a temperatura ambiente
durante 3 minutos (Fig. 7C). Enjuagar con agua destilada por 2 minutos.
Secar con papel absorbente y guardar a 4°C por 24 horas.
c) Al siguiente día, poner las semillas a germinar en placas Petri con papel filtro
embebido en agua destilada en una cámara de crecimiento a 21ºC.
d) A las 48 horas se debería tener sobre un 50% de germinación (Fig. 7D).
e) Con este protocolo se consiguió un 95% de germinación.
7A
C
Frutos inmaduros de P. flexuosa
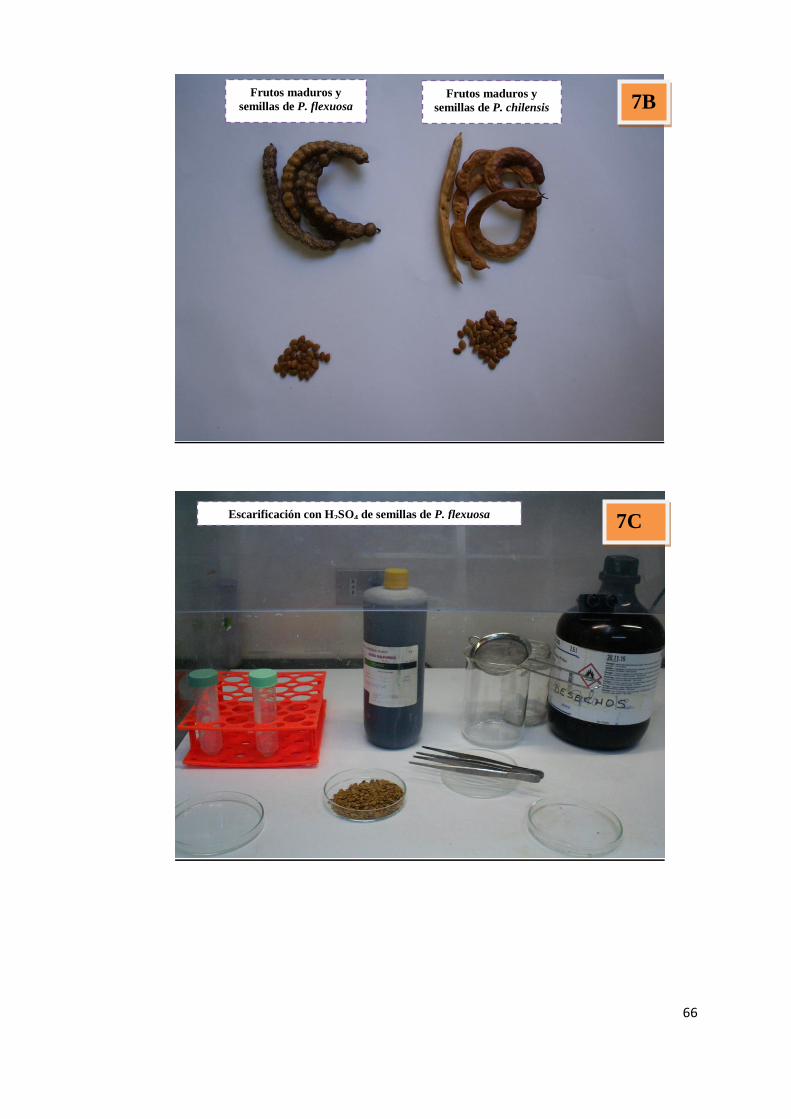
66
7B
C
Frutos maduros y
semillas de P. flexuosa Frutos maduros y
semillas de P. chilensis
7C
C
Escarificación con H2SO4 de semillas de P. flexuosa

67
7D
C
Semillas germinadas de P. flexuosa
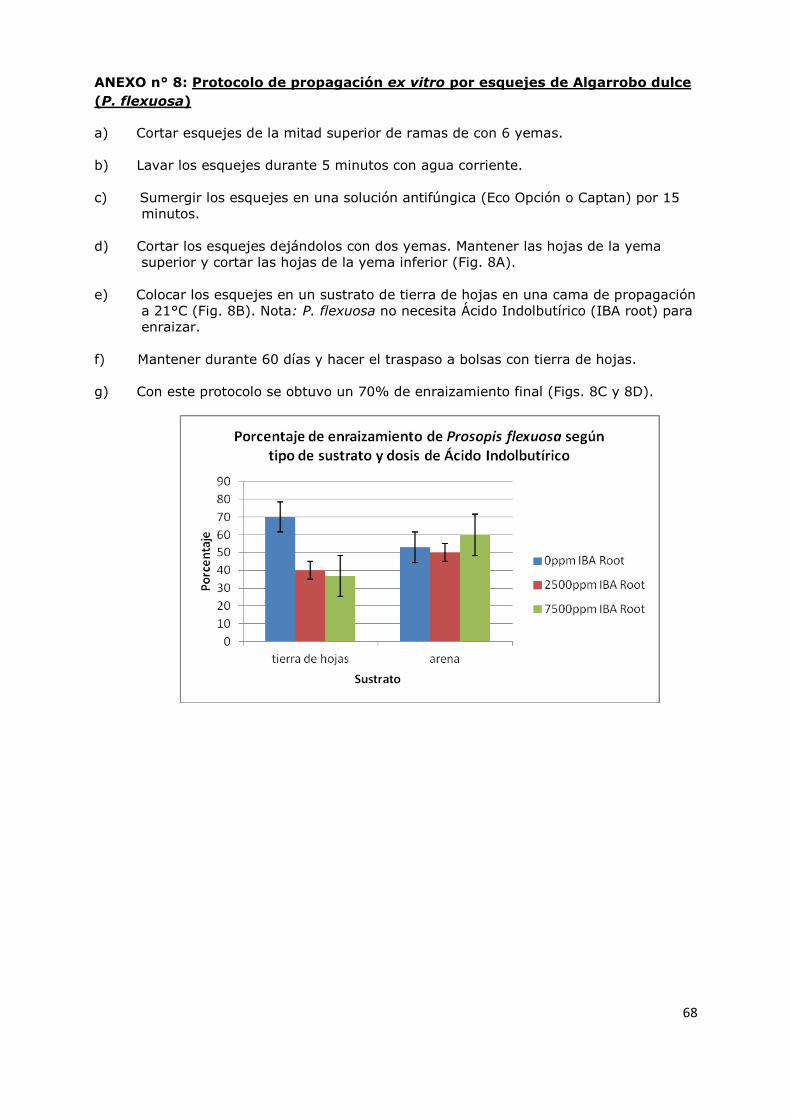
68
ANEXO n° 8: Protocolo de propagación ex vitro por esquejes de Algarrobo dulce
(P. flexuosa)
a) Cortar esquejes de la mitad superior de ramas de con 6 yemas.
b) Lavar los esquejes durante 5 minutos con agua corriente.
c) Sumergir los esquejes en una solución antifúngica (Eco Opción o Captan) por 15
minutos.
d) Cortar los esquejes dejándolos con dos yemas. Mantener las hojas de la yema
superior y cortar las hojas de la yema inferior (Fig. 8A).
e) Colocar los esquejes en un sustrato de tierra de hojas en una cama de propagación
a 21°C (Fig. 8B). Nota: P. flexuosa no necesita Ácido Indolbutírico (IBA root) para
enraizar.
f) Mantener durante 60 días y hacer el traspaso a bolsas con tierra de hojas.
g) Con este protocolo se obtuvo un 70% de enraizamiento final (Figs. 8C y 8D).

69
8B
8A
G
Esqueje de P. flexuosa antes y después de ser seccionado
Establecimiento de esquejes de P. flexuosa en la cama caliente
(zona amarilla demarcada)

70
8C
8D
Esquejes de P. flexuosa de 1 mes de edad
con un incipiente enraizamiento
Esquejes de P. flexuosa de 2 meses de edad ya enraizados

71
ANEXO n° 9: Protocolo de propagación in vitro de Algarrobo dulce (P. flexuosa)
a) Cortar explantes de 3-5 cm que contengan al menos 1 yema lateral. Se
recomienda utilizar como plantas madres, individuos jóvenes (1 año de edad),
desarrolladas a partir de semillas en invernadero (Fig. 9A).
b) Lavar los explantes con agua corriente durante 5 minutos.
c) Desinfectar los explantes sumergiéndolos en etanol 70% durante 1 minuto.
Luego en Hipoclorito de Sodio 1,5% + 2 gotas tween por 10 minutos.
Posteriormente realizar tres enjuagues con agua destilada estéril.
d) Medio de cultivo para el establecimiento: En frascos de vidrio adicionar un
volumen final de 15ml de medio de establecimiento. Cada litro de medio de
cultivo está constituido por medio básico de Murashige & Skoog 1962 (MS),
sacarosa (2%), 6-bencilaminopurina (4,4 µM), Polivinilpirrolidona (PVP, 1,5
g/L) y Agar-Agar (8,0 g/L), pH 5,8 previo a la adición del agar. Autoclavar los
medios a 121°C, presión de 1Kg/cm2 y por 20 min. Traspasar el explante
desinfectado a este medio y mantener en cámara con temperatura 21°C,
fotoperiodo largo (16h luz/8h oscuridad), 70% H.R, 120 µmolm-2s-1.
e) A los 30 días, transferir los explantes con sus yemas brotadas a medio fresco
de cultivo. Mantener por otros 30 días y luego pasar a la etapa de
enraizamiento.
f) Enraizamiento: Colocar los esquejes brotados en frascos de vidrio con 15 mL
de medio de enraizamiento. Cada litro de medio de cultivo está constituido por
medio MS, sacarosa (3%), Kinetina (1mg/L) y Ácido Indolbutírico (AIB, 10
mg/L). Autoclavar según lo indicado en punto d. Durante un mes mantener en
cámara de crecimiento con condiciones controladas de luz, temperatura y
humedad relativa ya descritas en el punto d (Fig. 9B y 9C).
g) Extraer la plántula de los tubos, lavar raíces y extraer el agar. Plantar en
vasos plástico cerrados con sustrato autoclavado de arena vermiculita (2:1) o
sólo de vermiculita y mantener durante 1 mes en cámara, facilitando
gradualmente el intercambio gaseoso y con riegos cada 7 días con solución
nutritiva Hoagland. Luego transferir a invernadero.
h) Con este protocolo se obtuvo un 70% de enraizamiento final (Fig. 9D).
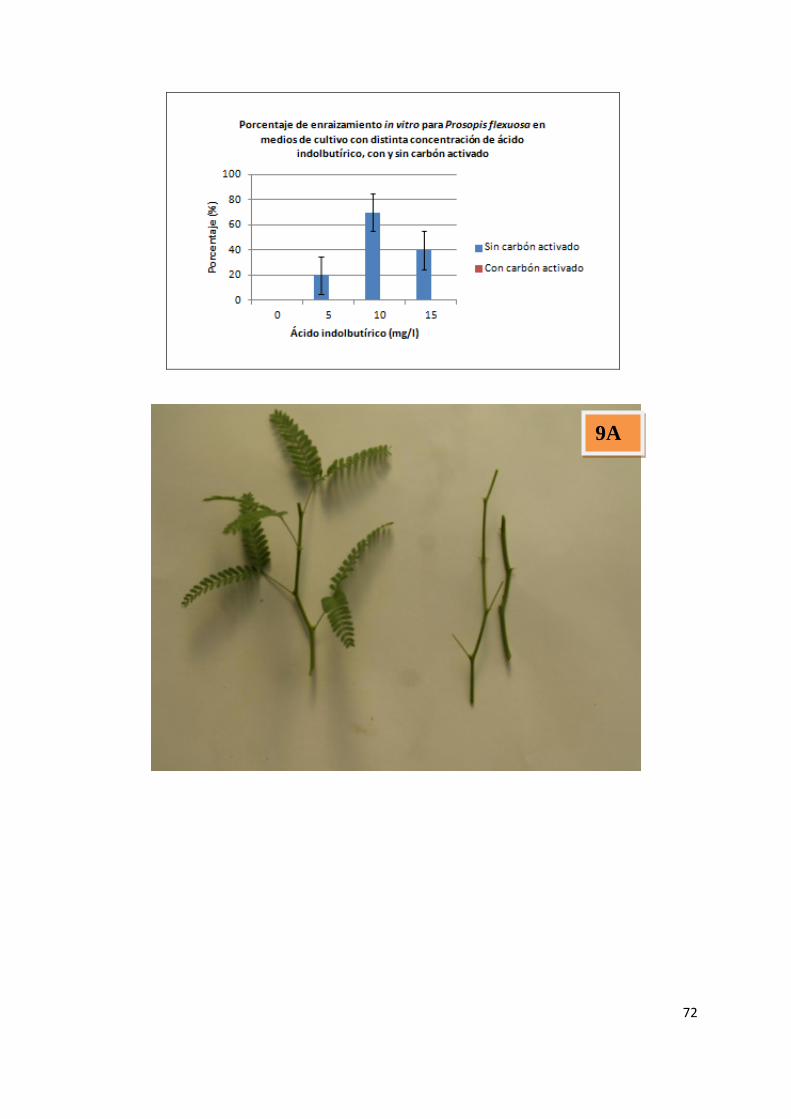
72
9A

73
9B
9C

74
9D

75
ANEXO n° 10: Protocolo de germinación semillas de Algarrobo (P. chilensis)
a) Extraer las semillas de las vainas (Fig. 10A). Eliminar las semillas vanas o
comidas por insectos. Almacenar en frío (4°C) por mínimo 1 mes.
b) Escarificar las semillas con ácido sulfúrico (H2SO4) a temperatura ambiente
durante 3 minutos. Enjuagar con agua destilada por 2 minutos. Secar con
papel absorbente y guardar a 4°C por 24 horas.
c) Al siguiente día, poner las semillas a germinar en placas Petri con papel filtro
embebido en agua destilada en una cámara de crecimiento a 21ºC.
d) A las 48 horas se debería tener sobre un 50% de germinación.
e) Con este protocolo se consiguió un 95% de germinación (Fig. 10B), y sobre el
90% de las semillas germinadas lograron desarrollarse exitosamente (Fig.
10C).
10A

76
10B
10C
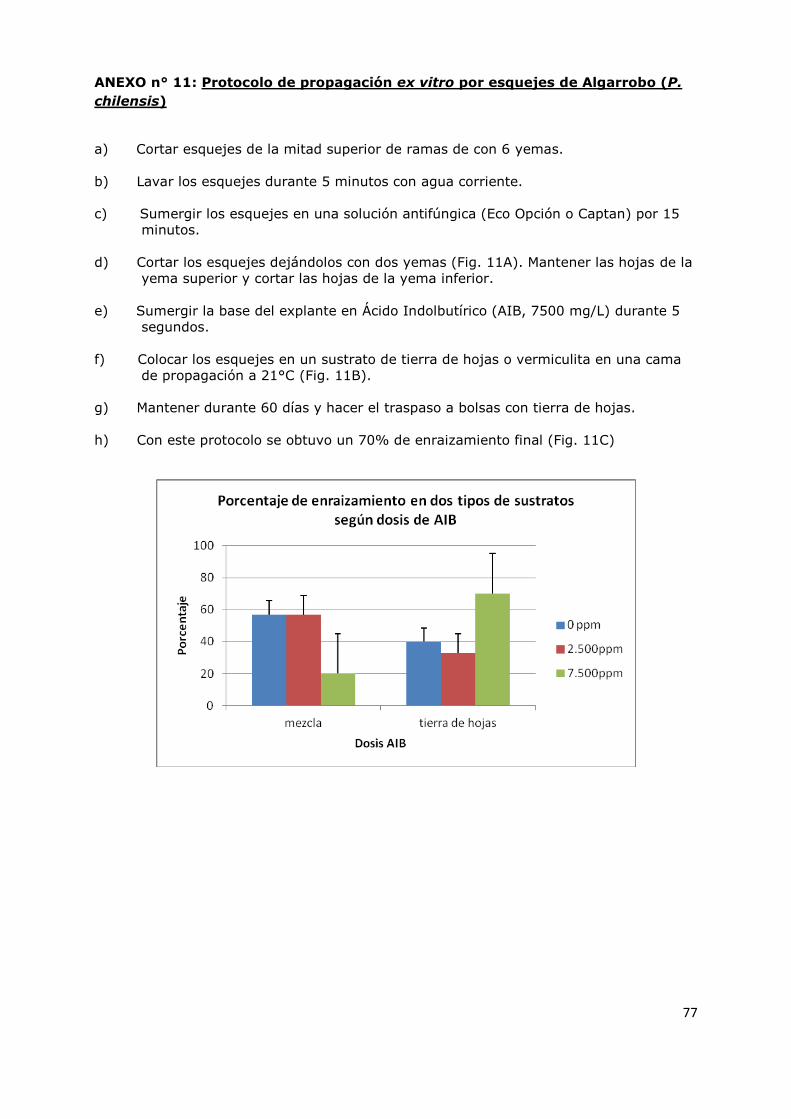
77
ANEXO n° 11: Protocolo de propagación ex vitro por esquejes de Algarrobo (P.
chilensis)
a) Cortar esquejes de la mitad superior de ramas de con 6 yemas.
b) Lavar los esquejes durante 5 minutos con agua corriente.
c) Sumergir los esquejes en una solución antifúngica (Eco Opción o Captan) por 15
minutos.
d) Cortar los esquejes dejándolos con dos yemas (Fig. 11A). Mantener las hojas de la
yema superior y cortar las hojas de la yema inferior.
e) Sumergir la base del explante en Ácido Indolbutírico (AIB, 7500 mg/L) durante 5
segundos.
f) Colocar los esquejes en un sustrato de tierra de hojas o vermiculita en una cama
de propagación a 21°C (Fig. 11B).
g) Mantener durante 60 días y hacer el traspaso a bolsas con tierra de hojas.
h) Con este protocolo se obtuvo un 70% de enraizamiento final (Fig. 11C)

78
11A
11B

79
11C

80
ANEXO n° 12: Protocolo de propagación in vitro de Algarrobo (P.
chilensis)
a) Cortar explantes de 3-5 cm que contengan al menos 1 yema lateral
provenientes de plantas jóvenes (1 año de edad), desarrolladas a partir de
semillas en invernadero.
b) Lavar los explantes con agua corriente durante 5 minutos.
c) Desinfectar los explantes sumergiéndolos en etanol 70% durante 1 minuto.
Luego en Hipoclorito de Sodio 1,5% + 2 gotas tween por 10 minutos.
Posteriormente realizar tres enjuagues con agua destilada estéril.
d) Medio de cultivo para el establecimiento: En frascos de vidrio adicionar 15 mL
de medio básico de Murashige and Skoog (MS), sacarosa (2%), 6-
bencilaminopurina (4,4 µM), PVP (1,5 g/l) y Agar-Agar (8,0 g/L), pH 5,8
previo a la adición del agar. Autoclavar los medios a 121°C, presión de
1Kg/cm2 y por 20 min. Mantener en cámara con temperatura 21°C,
fotoperiodo largo (16h luz/8h oscuridad), 70% H.R, 120 µmolm-2s-1 (Fig 12A).
e) Enraizamiento: En frascos de vidrio colocar los brotes en un medio de
enraizamiento con MS, sacarosa (3%), Kinetina (1mg/L), Ácido Indolbutírico
(AIB, 5 mg/L) y Carbón Activado (1g/L). Autoclavar según lo indicado en
punto d. Durante un mes mantener en cámara de crecimiento con condiciones
controladas de luz, temperatura y humedad relativa ya descritas en el punto
d.
f) Extraer la plántula de los tubos, lavar raíces y extraer agar. Plantar en vasos
plástico cerrados con sustrato autoclavado de arena vermiculita (2:1) o sólo
de vermiculita y mantener durante 1 mes en cámara, facilitando gradualmente
el intercambio gaseoso y con riegos cada 7 días con solución nutritiva
Hoagland. Luego transferir a invernadero.
g) Con este protocolo se obtuvo un 30% de enraizamiento final (Fig 12B).

81
12A
12B

82
ANEXO n° 13: Materiales usados en la parte de propagación
Campana de flujo laminar y reactivos típicos del cultivo in vitro
83
84
ANEXO n° 14: Poster 1 presentado en la VII Reunión de Biología Vegetal. Pucón
2012

85
ANEXO n° 15: Poster 2 presentado en la VII Reunión de Biología Vegetal. Pucón
2012

86
ANEXO n° 16: Poster 3 presentado en la VII Reunión de Biología Vegetal. Pucón
2012

87
ANEXO n° 17: Programa II Encuentro temático del FIBN, Iquique 2013

88
ANEXO n° 18: Certificado de aceptación trabajo oral Cristian Ibáñez. Tercer
Congreso de Flora Nativa. Santiago 2013.

89
ANEXO n° 19: Certificado de aceptación trabajo oral Fabiola Jamett. Tercer
Congreso de Flora Nativa. Santiago 2013.

90
ANEXO n° 20: Certificado de aceptación Poster 4 presentado al Tercer Congreso
de Flora Nativa. Santiago 2013.

91
ANEXO n° 21: Poster 5 presentado en la VIII Reunión de Biología Vegetal,
Pucón 2013.
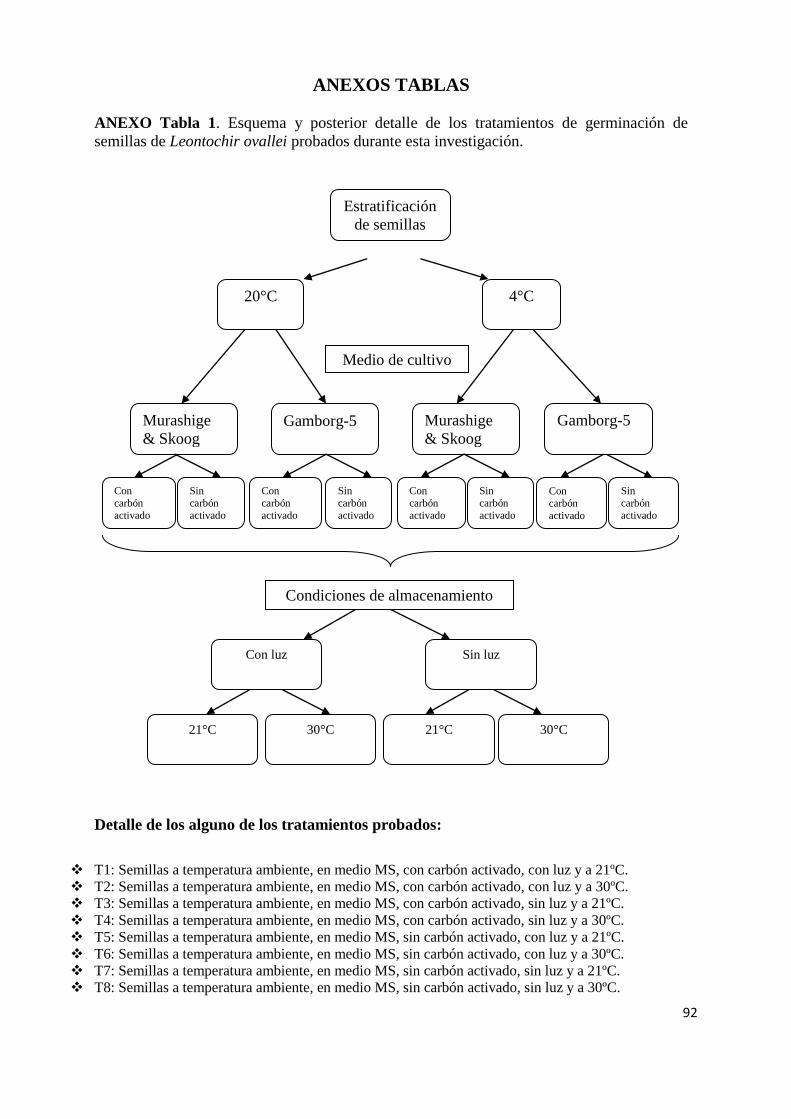
92
ANEXOS TABLAS
ANEXO Tabla 1. Esquema y posterior detalle de los tratamientos de germinación de
semillas de Leontochir ovallei probados durante esta investigación.
Detalle de los alguno de los tratamientos probados:
T1: Semillas a temperatura ambiente, en medio MS, con carbón activado, con luz y a 21ºC.
T2: Semillas a temperatura ambiente, en medio MS, con carbón activado, con luz y a 30ºC.
T3: Semillas a temperatura ambiente, en medio MS, con carbón activado, sin luz y a 21ºC.
T4: Semillas a temperatura ambiente, en medio MS, con carbón activado, sin luz y a 30ºC.
T5: Semillas a temperatura ambiente, en medio MS, sin carbón activado, con luz y a 21ºC.
T6: Semillas a temperatura ambiente, en medio MS, sin carbón activado, con luz y a 30ºC.
T7: Semillas a temperatura ambiente, en medio MS, sin carbón activado, sin luz y a 21ºC.
T8: Semillas a temperatura ambiente, en medio MS, sin carbón activado, sin luz y a 30ºC.
Estratificación
de semillas
4°C 20°C
Murashige
& Skoog Gamborg-5 Murashige
& Skoog
Gamborg-5
Con
carbón
activado
Sin
carbón
activado
Con
carbón
activado
Sin
carbón
activado
Con
carbón
activado
Sin
carbón
activado
Con
carbón
activado
Sin
carbón
activado
Medio de cultivo
Condiciones de almacenamiento
Con luz Sin luz
30°C 21°C 21°C 30°C
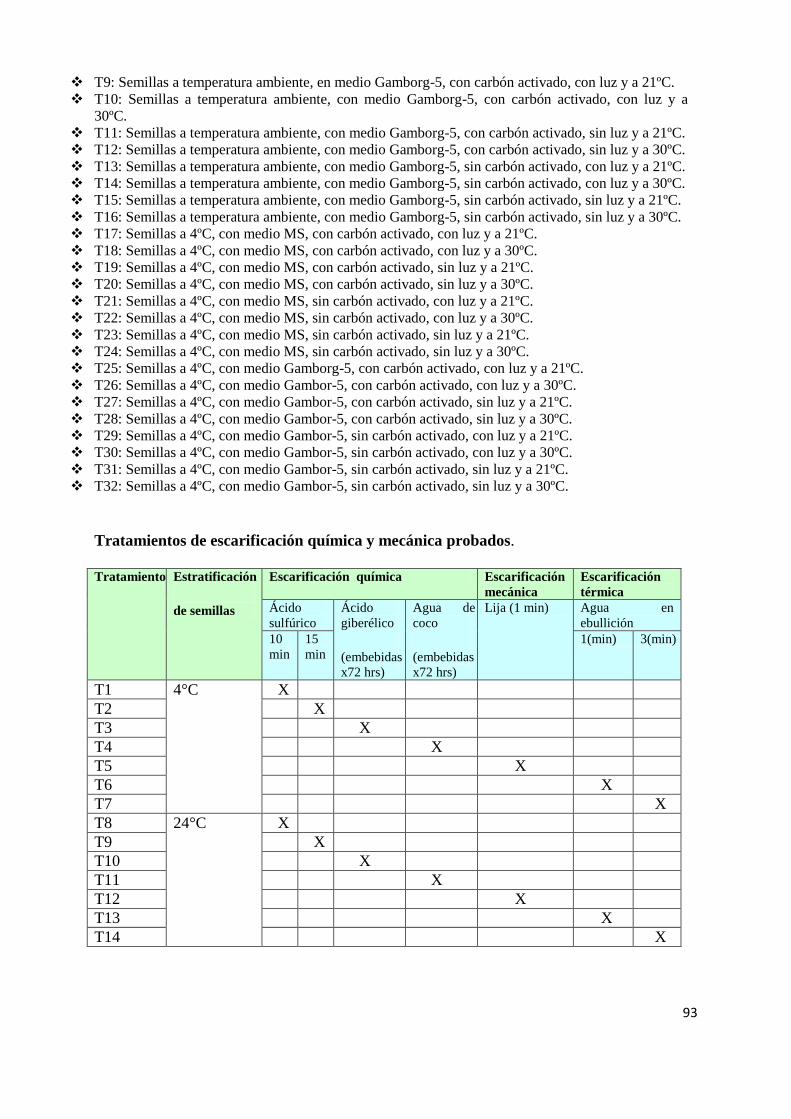
93
T9: Semillas a temperatura ambiente, en medio Gamborg-5, con carbón activado, con luz y a 21ºC.
T10: Semillas a temperatura ambiente, con medio Gamborg-5, con carbón activado, con luz y a
30ºC.
T11: Semillas a temperatura ambiente, con medio Gamborg-5, con carbón activado, sin luz y a 21ºC.
T12: Semillas a temperatura ambiente, con medio Gamborg-5, con carbón activado, sin luz y a 30ºC.
T13: Semillas a temperatura ambiente, con medio Gamborg-5, sin carbón activado, con luz y a 21ºC.
T14: Semillas a temperatura ambiente, con medio Gamborg-5, sin carbón activado, con luz y a 30ºC.
T15: Semillas a temperatura ambiente, con medio Gamborg-5, sin carbón activado, sin luz y a 21ºC.
T16: Semillas a temperatura ambiente, con medio Gamborg-5, sin carbón activado, sin luz y a 30ºC.
T17: Semillas a 4ºC, con medio MS, con carbón activado, con luz y a 21ºC.
T18: Semillas a 4ºC, con medio MS, con carbón activado, con luz y a 30ºC.
T19: Semillas a 4ºC, con medio MS, con carbón activado, sin luz y a 21ºC.
T20: Semillas a 4ºC, con medio MS, con carbón activado, sin luz y a 30ºC.
T21: Semillas a 4ºC, con medio MS, sin carbón activado, con luz y a 21ºC.
T22: Semillas a 4ºC, con medio MS, sin carbón activado, con luz y a 30ºC.
T23: Semillas a 4ºC, con medio MS, sin carbón activado, sin luz y a 21ºC.
T24: Semillas a 4ºC, con medio MS, sin carbón activado, sin luz y a 30ºC.
T25: Semillas a 4ºC, con medio Gamborg-5, con carbón activado, con luz y a 21ºC.
T26: Semillas a 4ºC, con medio Gambor-5, con carbón activado, con luz y a 30ºC.
T27: Semillas a 4ºC, con medio Gambor-5, con carbón activado, sin luz y a 21ºC.
T28: Semillas a 4ºC, con medio Gambor-5, con carbón activado, sin luz y a 30ºC.
T29: Semillas a 4ºC, con medio Gambor-5, sin carbón activado, con luz y a 21ºC.
T30: Semillas a 4ºC, con medio Gambor-5, sin carbón activado, con luz y a 30ºC.
T31: Semillas a 4ºC, con medio Gambor-5, sin carbón activado, sin luz y a 21ºC.
T32: Semillas a 4ºC, con medio Gambor-5, sin carbón activado, sin luz y a 30ºC.
Tratamientos de escarificación química y mecánica probados.
Tratamiento Estratificación
de semillas
Escarificación química Escarificación
mecánica
Escarificación
térmica
Ácido
sulfúrico
Ácido
giberélico
(embebidas
x72 hrs)
Agua de
coco
(embebidas
x72 hrs)
Lija (1 min) Agua en
ebullición
10
min
15
min
1(min) 3(min)
T1 4°C X
T2 X
T3 X
T4 X
T5 X
T6 X
T7 X
T8 24°C X
T9 X
T10 X
T11 X
T12 X
T13 X
T14 X