enzyme-antibody conjugation by a heterobifunctional reagent and its application in enzyme-linked...
TRANSCRIPT

Journal of Virological Methods, 10 (1985) 215-224
Elsevier
JVM 00375
215
ENZYME-ANTIBODY CONJUGATION BY A HETEROBIFUNCTIONAL
REAGENT AND ITS APPLICATION IN ENZYME-LINKED
IMMUNOSORBENT ASSAY (ELISA) FOR THE DETECTION OF
HEPATITIS B SURFACE ANTIGEN
DEEPAK A. GADKARI’. HOWARD A. FIELDS? and JAMES E. MAYNARD2
‘National .Instirute qf Virology, Post Office Box 1 I, Pune 41IOO1, India: *Division of Viral Diseases,* Center
for Ir$ectkms Diseases. Centers for Disease Control, Public Health Service. U.S. Department qf Health and
Human Services. 1600 Clifton Road, N.E.. Atlanta, GA 85014. U.S.A.
(Accepted 29 October 1984)
The heterobifunctional reagent 3-(2.pyridyldithio)propionate (SPDP) was used to prepare defined
conjugates composed of horseradish peroxidase (HRP) and goat anti-HBs IgG. The modification of HRP
and IgG ,wtth SPDP was dependent on both the SPDP: protein molar ratio and the pH of the buffer.
Conjugates were separated by a single affinity chromatographic step using concanavalin A-Sepharose 48
equilibrated with 0.1 M Tris-HCI buffer, pH 7.4, containing 0.5 M KCI and I mM EDTA. The conjugate
was eluted with 10 or 100 mM a-methyl-o-mannoside and appropriate pools were made reflecting various
HRP/IgCi molar ratios. Each pool was examined for performance in an enzyme-linked immunosorbent
assay for HBsAg. Conjugate composed of an HRP/lgG molar ratioof 2.5-4 yielded the greatest sensitivity.
SPDP conjugation conjugate purification ELISA HBsAp
INTRODUCTION
Since its introduction in 1971 by Engvall and Pearlman (1971) enzyme-linked
immunosorbent assay (ELISA) has been widely used for the measurement of bac-
terial, viral, parasitic and fungal antigens, as well as immunoglobulins, hormones,
drugs, serum proteins and tumor antigens (Yolken, 1982). Advances in substrate
selection, solid-phase adsorbents and instrumentation have resulted in ELISA achiev-
ing similar sensitivities as radioimmunoassay without the concomitant disadvan-
tages associated with the use of radioactive reagents. In 1976, Wolters et al. introduced
ELISA for the detection of hepatitis B surface antigen. Since this initial report, there
have been many publications on this subject (Caldwell and Barrett, 1977; Halbert and
Anken, 1977: Wei et al., 1977).
* World Health Organization Collaborating Centre for Reference and Research on Viral Hepatitis
0166-0934/85/$03.30 0 1985 Elsevier Science Publishers B.V. (Biomedical Division)

216
Various methods for conjugating IgG to HRP using either glutaraldehyde (Avra-
meus, 1969; Avrameus and Ternynck, 1971) or meta-periodate (Wilson and Nakane,
1978) have been utilized in developing ELISA. These methods have several drawbacks
including the production of large molecular weight conjugates, homopolymers, and
low enzyme yield. A heterobifunctional reagent, N-succinimidyl 3-(2-pyridyldithio)
propionate (SPDP), synthesized by Carlsson et al. (1978) has been successfully used
by Nilsson et al. (1981) to prepare defined conjugates of IgG and HRP without the
drawbacks associated with the other conjugation methods. This paper extends Nils-
son’s conjugation results and presents a novel one-step procedure for separating
conjugates into defined HRP : IgG molar ratios.
MATERIALS AND METHODS
Reagents
SPDP was purchased from Pharmacia Fine Chemicals (Piscataway, NJ). Stock
solutions were prepared in absolute ethanol and the concentrations were determined
by the method recommended by the manufacturer. SPDP solutions were aliquoted
and stored at -70°C until used. All other reagents were purchased or prepared as
previously described (Fields et al., 1981, 1983).
SPDP thiolation
HRP and IgG were thiolated at different SPDP: protein molar ratios. The degree of
substitution (DOS) (number of thiol groups per mole of protein) was determined by
the method of Grassetti and Murray (1967). Briefly, after reacting HRP or IgG with
SPDP at different molar ratios for 30 min, the modified protein was desalted on a I .6
X 30 cm column of Sephadex G-25 (Pharmacia Fine Chemicals) equilibrated with 0.1
M sodium acetate buffer, pH 4.6, containing 0.1 M NaCl (buffer A) to remove
N-hydroxysuccinimide released during the reaction. One molar dithiothreitol (DTT)
was added to the modified protein preparation to a final DTT concentration of 50
mM. After incubation for 30 min at room temperature, the reaction mixture was
desalted on a second Sephadex G-25 column equilibrated with 0.1 M Tris-HCl buffer,
pH 7.4, containing 0.5 M KC1 and 1 mM EDTA (buffer B). The eluted protein was
concentrated to a volume of 4 ml by ultrafiltration by means of a 25 mm diameter
YMlO ultrafilter and an 8 MC stirred cell (Amicon, Lexington, MA). Equal volumes
of thiolated protein and 1.5 mM 2,2’-dipyridyl disulfide (2-PDS) were added to a
cuvette and the concentration of liberated 2-thiopyridone (2-TP) was determined
spectrophotometrically at a wavelength of 343 nm after subtracting absorbance due to
thiolated protein alone and 2-PDS alone. One mole of liberated 2-TP is equivalent to 1
thiol group.
Conjugate preparation
SPDP (15-30 mM) was added dropwise to 15-20 mg of goat (gt) anti-HBs IgG

217
prepared in 0.1 M phosphate buffer, pH 7.2, containing 0.1 M NaCl to yield a molar
ratio (SPDP : IgG) of 54 : 1. The modified protein was desalted on G-25 equilibrated
with buffer B, concentrated by ultrafiltration and stored at 4°C. HRP was modified
with SPDP at a molar ratio of 50 : 1. The modified protein was desalted on Sephadex
G-25 equilibrated with buffer A, treated with 50 mM DTT, desalted on Sephadex
G-25 equilibrated with buffer B, concentrated to 4 ml by ultrafiltration, and used
immediately for the conjugation reaction. The modified IgG containing 2-pyridyl
disulphide residues and the thiolated HRP were mixed together and stirred gently for
16 h at 2.3”C. A 1.5-5 molar excess of thiolated HRP was used to favor the thiol-disul-
phide bond exchange reaction.
Separation of conjugate
A 0.9 X 16 cm column of concanavalin A-Sepharose 4B (Con A-Seph) (Sigma
Chemical Co., St. Louis, MO) equilibrated with 10 bed volumes of buffer B was used
to separate the conjugate. The conjugation mixture was chromatographed at a flow
rate of 5 ml/h. Unadsorbed protein was eluted with buffer B until the absorbance at
280 nm (A,,,) returned to baseline and then the conjugate was eluted with 10 and 100
mM a-methyl-o-mannoside (AMDM) prepared in buffer B. Fractions containing the
conjugate were selectively pooled, lyophilized, and stored at -20°C. The conjugate has
remained stable for more than 2 yr.
HRP: IgG molar ratio
The molar ratio of HRP : IgG was determined spectrophotometrically by measuring
the AZ80 and A,,,,. The extinction coefficients (Ey:z ) andmolecular WeightsforHRPand
IgG used were 1.95 and 4.0 X lo5 and 1.35 and 1.6 X 105, respectively. A standard
curve is presented in Fig. 1.
Enzyme-linked immunosorbent assay (ELBA)
ELISA was performed according to the method described by Fields et al. (1983).
Briefly, 0.125 ml of guinea pig anti-HBs IgG appropriately diluted in 0.05 M carbo-
nate buffer, pH 9.6, was added to flat bottom microtiter strips (Immulon I or II,
Dynatech, Alexandria, VA) for 16 h at 23°C. The wells were then washed 5 times with
phosphate-buffered saline (PBS), pH 7.2, containing 0.5% (w/v) bovine serum albu-
min (BSA), and 0.1 ml of a standard dilution of HBsAg-positive chimpanzee plasma
was added. After incubation at 45°C for 3 h the wells were washed 7 times and 0.1 ml of
various dilutions of conjugate were added to each well. After 2 h of incubation at 37°C
the wells were washed 10 times and 0.2 ml of substrate (H,O,) and chromophore
(o-phenylenediamine) prepared in phosphate-citrate buffer, pH 5.0, were added in the
dark. After 40 min at 23°C the reaction was stopped by the addition of 0.1 ml of 4 N
H,SO,. Absorbance was measured by reading the A,,, in each well using an automatic
Dynatech reader (MR-580). Results were scored by comparing positive to negative
(P/N) quotients. It has been previously demonstrated that the higher the P/N ratio,
the greater the sensitivity (Fields et al. 1983).

218
20-
5
I 5-
8 \lO- 8
ob O 5-
2 4 6 8 IO 12 14 16 I8 HRP/lgG molar ratio
0
Fig. I. Spectrophotometric determlnatlon 01. HRP/lpG molar ratios. The calculation was performed as
follows:
1) HRP protein concentration (me/ml) = A,,,,/E’\j’jl
2) IgG protein concentration (mg/ml) = (A 2R,,-A,,,,/RZ value for HRP)/E”,i’,,
3) HRPAgG molar ratio = (HRP protein concentration/molecular weight)/(IgG protein concentra-
tion/molecular *eight)
Pro rein assay
Protein concentration was determined by the Bio-Rad protein assay (Richmond,
CA) using protein standard I for IgG and a prepared HRP standard for HRP.
RESULTS
Degree of substitution
Fig. 2 demonstrates the degree of substitution (number of thiol groups/mole of
protein) as a function of the SPDP : IgG molar ratio. At a molar ratio of 2 : 1, a degree
of substitution (DOS) of 1.8 was obtained which increased linearly up to a DOS of 3.5
at a molar ratio of 18 : 1. The DOS could not, however, be increased further even at a
molar ratio of 54 : 1.
In an attempt to increase the DOS, we examined the SPDP thiolation reaction as a
function of pH (Fig. 3). The goat anti-HBs IgG was dialyzed against the appropriate
buffer before being reacted with SPDP at a molar ratio of 54 : 1, The buffers used were
as follows: (1) 0.1 M phosphate buffer, pH 7.2, (2) 0.1 M phosphate buffer, pH 8.0, and
(3) 0.1 M carbonate buffer, pH 9.0. All buffers contained 0.1 M NaCl. As the pH was
increased from 7.2 to 9.0 the DOS decreased from 7.5 to 1.5.
HRP was considerably more refractory to thiolation than IgG (Table 1). At a molar
ratio of 15 : I, a DOS of 0.8 1 was obtained. By increasing the molar ratio by a factor of
5, only a 2-fold increase of DOS was observed. For the preparation of conjugates a
SPDP: HRP molar ratio of 50: 1 was used which resulted in a DOS of close to 1.

219
5 IO 15 20 25 30 35 40 45 50 !
SPDP/ IgG molar ratlo 7.0 80
PH
9.0
Fig. 2. Degree of substitution of goat anti-HBs IgG as a function ofSPDP: I@ molar ratios obtained at pH
8.0. All values are averages of at least 3 separate experiments.
Fig. 3. Degree ofsubstitution ofgoat anti-HBs IgG asa function ofpH at a constant SPDP: IgG molar ratio
of 54: 1.
Separation of conjugate
SPDP-modified IgG was mixed with the modified and reduced HRP as described in
Materials and Methods. After the incubation period the conjugate was fractionated by
lectin affinity chromatography using Con A-Seph 4B (Fig. 4). The unbound fractions
(fractions 7-49) contained little anti-HBs-HRP conjugate activity based upon low
reactivity in the ELISA for HBsAg. This peak was subsequently rechromatographed
on a 2.6 X 87 cm gel filtration column of Bio-Gel A-O.5 m agarose equilibrated with
PBS and previously calibrated with HRP and iodinated IgG. This experiment de-
monstrated (data not shown) that the unbound fraction from the Con A-Seph 4B
chromatography was composed of free IgG and free HRP as indicated by their
appearances corresponding to their expected elution volumes. Conjugate was eluted
with 10 mM AMDM from the Con A-Seph 4B column when the absorbance returned
to baseline. Fractions 65-75 contained considerable conjugate activity based upon its
TABLE I
Degree of substitution in HRP by SPDP
SPDP/HRP Degree of
molar ratio Substitution
15: 1 0.81
25: I 0.80
50: 1 1.18
75: 1 1.65
All values are average of 2 or more experiments.

220
0 4 a I2 16 20 24 28 32 36 40 44 40 52 56 60 64 68 72 76 80 84 00 92
FRACTION NUMBER
Fig. 4. Separation of SPDP-prepared conjugates on concanavalin A-Sepharose 4B. The column (0.9 X 16
cm) was equilibrated with 0.1 M Tr~s-HCI buffer. pH 7.4, containing 0.5 M KCI and I mM EDTA (buffer
B). The flow rate was 5 ml/h and 2 ml size fractions were collected.
performance in an ELISA. When the AMDM concentration was increased to 100
mM, a second absorbance peak was observed between fractions 85 and 92. This peak
also displayed conjugate activity by ELISA. No more conjugate could be eluted
despite the presence of a brownish colored material characteristic of HRP adsorbed to
the top of the column.
ELISA
The assay was performed using conjugates obtained by pooling various fractions
eluted from the Con A-Seph 4B column. The HRP/IgG ratio was determined
according to the standard curve (Fig. 1). Table 2 lists the pooled fractions comprising
the various conjugate preparations and their corresponding HRP/IgG molar ratio.
Conjugates A-E were compared with a conjugate previously prepared by the sodium
meta-periodate method (Fields et al., 1983).
The performance of each conjugate was assessed by comparing a calculated P/N
TABLE 2
HRP/IgG molar ratios of selected pooled fractions from the concanavalin A-Sepharose 4B fractionation of
SPDP-prepared conjugates’
Conjugate Fraction Nos. ‘42,” &J HRP/lgG
pooled (Fig. 3) (molar ratio)
A 65-67 0.290 0.458 8.64
B 68-74 0.085 0.086 4.10
c 75-84 0.032 0.023 2.57
D 85-87 0.085 0.119 6.89
E 88-89 0.077 0.095 5.56
221
TABLE 3
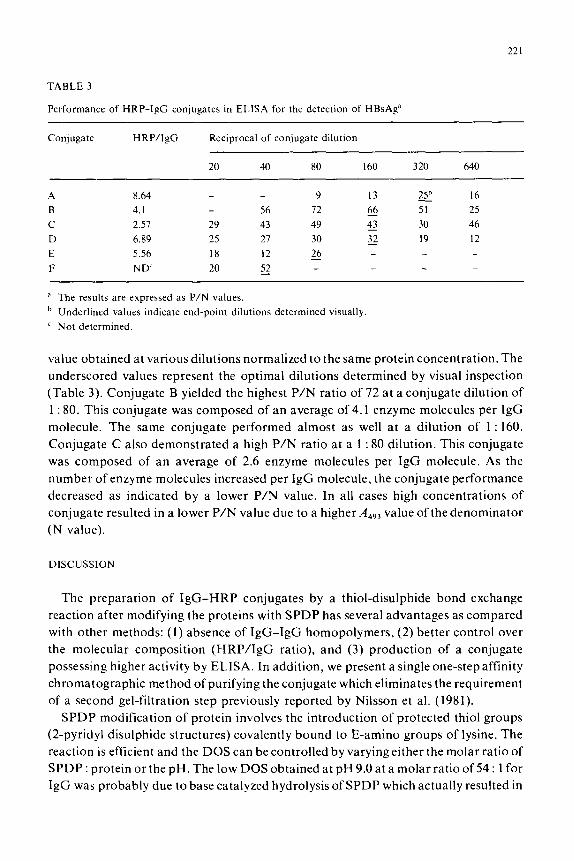
Performance of HRP-IgC conjugates in ELISA for the detection of HBsAg”
Conjugate HRP/IgG Reciprocal of conjugate dilution
20 40 80 160 320 640
A 8.64 9 13 25h 16
B 4. I 56 72 66 F 25
C 2.51 29 43 49 43 30 46
D 6.89 25 27 30 32 19 12 - E 5.56 18 12 26 F ND 20 52 r _
a The results are expressed as P/N values.
b Underlined values indicate end-point dilutions determined visually
’ Not determined.
value obtained at various dilutions normalized to the same protein concentration. The
underscored values represent the optimal dilutions determined by visual inspection
(Table 3). Conjugate B yielded the highest P/N ratio of 72 at a conjugate dilution of
1 : 80. This conjugate was composed of an average of 4.1 enzyme molecules per IgG
molecule. The same conjugate performed almost as well at a dilution of 1 : 160.
Conjugate C also demonstrated a high P/N ratio at a 1 : 80 dilution. This conjugate
was composed of an average of 2.6 enzyme molecules per IgG molecule. As the
number of enzyme molecules increased per IgG molecule, the conjugate performance
decreased as indicated by a lower P/N value. In all cases high concentrations of
conjugate resulted in a lower P/N value due to a higher Adq3 value of the denominator
(N value).
DISCUSSION
The preparation of IgG-HRP conjugates by a thiol-disulphide bond exchange
reaction after modifying the proteins with SPDP has several advantages as compared
with other methods: (1) absence of IgG-IgG homopolymers, (2) better control over
the molecular composition (HRP/IgG ratio), and (3) production of a conjugate
possessing higher activity by ELISA. In addition, we present a single one-step affinity
chromatographic method of purifying the conjugate which eliminates the requirement
of a second gel-filtration step previously reported by Nilsson et al. (1981).
SPDP modification of protein involves the introduction of protected thiol groups
(Zpyridyl disulphide structures) covalently bound to E-amino groups of lysine. The
reaction is efficient and the DOS can be controlled by varying either the molar ratio of
SPDP : protein or the pH. The low DOS obtained at pH 9.0 at a molar ratio of 54 : 1 for
IgG was probably due to base catalyzed hydrolysis of SPDP which actually resulted in

222
a lowered molar ratio of intact SPDP to IgG. The 2-pyridyl disulphide (2-PDS)
protecting groups can be removed by a single reduction procedure which spares native
protein disulphide bridges.
The DOS of proteins is dependent upon the number of accessible E-amino groups
of lysine. IgG possesses between 50 and 70 groups while HRP possesses only 4
(Carlsson et al., 1978). Even though IgG contains many lysine residues only an
average of 7.5 2-PDS groups could be introduced indicating that most ofthese groups
are not accessible for SPDP modification or that optimal conditions were not used. It
was, therefore, not surprising that HRP, possessing only 4 lysine residues, required a
molar ratio of 75 : 1 (SPDP: HRP) to achieve a modest DOS of 1.65.
The lectin concanavalin A has been shown to bind HRP (Bio-Rad, Richmond, CA,
1982). This observation has led to the use of immobilized Con A for the purification of
IgG-HRPconjugates (Lanneret al., 1978; Arends, 1979; Nilsson etal., 1981). Unreact-
ed IgG does not bind to Con A-Seph 4B whereas conjugates and unreacted HRP bind
to the lectin necessitating a subsequent gel-filtration step. The separation method
described in this report uses a Con A-Seph 4B column equilibrated with a buffer
containing Tris, KC1 and EDTA (buffer B). The reaction mixture also contained the
same buffer. Under these conditions unreacted IgG and HRP were not significantly
adsorbed to the column, although HRP was slightly retarded. Most of the conjugate,
however, was bound to the column and was eluted with AMDM. Conjugate eluted
with 10 mM AMDM was qualitatively similar and differed only quantitatively to the
conjugate eluted with 100 mM AMDM. The separation procedure was performed on
three separate trials and similar results were obtained demonstrating consistent
reproducibility.
Why unreacted HRP did not bind to the affinity column is not understood. It is
known that Ca2’ and Mn?’ are required for Con A activity. In the presence of EDTA
(buffer B) these ions are chelated which might interfere with the binding of unreacted
HRP. However, in the presence of EDTA the majority of the conjugate remained
bound to the affinity column. This might have occurred due to conformation changes
in HRP molecules as a result of the conjugation reaction. Although the precise
mechanism for the preferential binding of IgG-HRP conjugates to Con A-Seph is not
understood, the method described was highly reproducible and eliminated the require-
ment of a second gel-filtration step. Furthermore, this method yielded conjugates
with different molar ratios of HRP/IgG which could be separated by pooling selected
fractions. HRP/IgG molar ratios in these pools varied from 2.6 to 8.6. Despite an
average DOS of 7.5 for IgG, not all of the modified groups participated in a disulphide
bond formation with the reduced HRP. Some of the modified groups probably were
located within the IgG molecule and were not accessible for the reaction. For
large-scale preparations, however, conjugates can be eluted with 100 mM AMDM in a
single pool. This pool had an average HRP/IgG ratio of 5.2 and could be used at a
dilution of 1 : 160 in an ELISA (data not shown).
The ELISA data indicated that conjugates composed of 2.5-4 HRP molecules per

223
IgG molecule yielded the greatest sensitivity. This observation is based upon a
comparison of P/N ratios obtained with various conjugates differing in their
HRP/IgG molar ratios. It has been previously demonstrated (Fields et al., 1983) that
P/N ratios are directly related to sensitivity. The decreased P/N values obtained with
conjugates composed of >4 HRP molecules per IgG molecule may be due to a loss of
IgG affinity or specificity for HBsAg due to a stereoconformational change of the
antibody binding site. These high molecular weight conjugates yielded a higher
background (N value) which decreased the overall P/N quotient.
This paper not only supports and extends the results of Nilsson et al. (198 l), but
also points out some important differences. First, the conjugates reported in the
present study possessed an average HRP : IgG molar ratio of 4.1 as compared to 2.0 by
Nilsson. This difference is probably due to the slightly higher DOS ( 1.18) achieved for
HRP which may have r,esulted in some degree of intermolecular bridging due to more
than one reaction site on some of the HRP molecules. Indeed, some conjugates
possessed molar ratios of greater than 8.0 (Table 2). Secondly, this paper examines the
parameters of SPDP modification of IgG as a function of SPDP : protein molar ratios
and pH, Finally, this paper describes a one-step method to separate conjugates from
free reactants. The mechanism responsible for this separation, however, remains
enigmatic.
REFERENCES
Arends, J.. 1979. J. Immunol. Methods 25. 171.
Avrameas. S.. 1969. Immunochemistry 6. 43.
Avrameas. S. and Ternynck, T., 1971, Immunochemistry 8, 1175.
Belanger. L., Hamel. D., Dufour, D. and Pouliot. M., 1976. Clin. Chem. 22, IX.
Bio-Rad Catalogw. January 1982, Richmond. CA, 4X.
Caldwell. C.W. and Barrett. J.T.. 1977. Chn. Chim. .4cta 81, 305.
Carlsaon, .I.. Drevin, H. and Axen. R., 1978. Biochem. J.. 173. 723.
Enpvall. E. and Pearlman, P.. 1971. Immunochemistry 8. 871.
Fields, H.A.. Davis. C.L.. Drecsman. G.R.. Bradley, D.W. andMaynard,J.E.. 19XI.J. Immunol. Methods
47. 145.
Fields, H.A.. Daws, C.L.. Bradley. D.W. and Maynard, .J.E.. 1983, Bull. Wrld Hlth Org. 61. 135.
Glshaw. J.B.. Hu. M.W.. Singh, P., Miller. J.G. and Schneider. R.S.. 1977, Clin. Chem. 23, 1144.
Grassetti. D.R. and Murray, J.F., 1967. Arch. Biochem. Biophys. 119, 41.
Halbert. S.P. and Anken, M., 1977, J. Infect. Dis. 136, 5318.
Lanner. M.. BerquiSt. R. and Carlsson. J.. 1978. in: Affinity Chromatography (Prrganon Press) p. 237.
Masseyeff. R.. 1978, Stand. J. Immunol. 8 (Suppl. 7). 83.
Nilsson. P.. Berqulst. N.R. and Grundy, M.S.. 1981. J. Immunol. Methods 41. 81.
Pal, S.B.. 1978. Walter de Gryuter. Berlin.
Pepple. J.. Moyom. E.R. and Yolken, R.N.. 1980, J. Pediat. 97. 233.
Voller, A.. Bartlett. A. and Bidwell. D.E.. 1978, Trap. Med. Hyg. 70. 98.
Warren. R.C.. Bartlett. A.. Bidwell. D.E.. Richardson. M.D.. Voller, A.and White. L.O., 1977,Br.Mtd..J.
I. 11x3.
Wel. R. ct al.. 1977. Clan. Chem. 23. 813.

224
Wilson, B.M. and Nakane, P.K., 1978, in: Immunofluorescence and Related Staining Techniques (Elsevier/
North-Holland Biomedical Press) p. 215.
Walters, G., L. Kuijpers, Kalaki, J. and Schuurs, A., 1976, J. Clin. Pathol. 29. 873.
Yolken, R.H., 1982, Rev. Infect. Dis. 4, 35.