effects of synthetic micro- and nano-structured surfaces on cell behavior
TRANSCRIPT

*Corresponding author. Tel.: 001 608 265 8171.
Biomaterials 20 (1999) 573—588
Effects of synthetic micro- and nano-structured surfaceson cell behavior
R.G. Flemming!, C.J. Murphy!, G.A. Abrams!, S.L. Goodman", P.F. Nealey#,*!Department of Surgical Sciences, School of Veterinary Medicine, UW-Madison, 2015 Linden Drive West, Madison, WI 53706, USA
"Center for Biomaterials MC-1615, University of Conn. Health Center, Farmington, CT 06030, USA#Department of Chemical Engineering, School of Engineering, UW-Madison, 1415 Engineering Drive, Madison, WI, USA
Received 10 June 1998; accepted 30 September 1998
Abstract
Topographical cues, independent of biochemistry, generated by the extracellular matrix may have significant effects upon cellularbehavior. Studies have documented that substratum topography has direct effects on the ability of cells to orient themselves, migrate,and produce organized cytoskeletal arrangements. Basement membranes are composed of extracellular matrix proteins and foundthroughout the vertebrate body, serving as substrata for overlying cellular structures. The topography of basement membranes isa complex meshwork of pores, fibers, ridges, and other features of nanometer sized dimensions. Synthetic surfaces with topographicalfeatures have been shown to influence cell behavior. These facts lead to the hypothesis that the topography of the basement membraneplays an important role in regulating cellular behavior in a manner distinct from that of the chemistry of the basement membrane.This paper describes the topography of the basement membrane and reviews the fabrication of synthetic micro- and nano-structuredsurfaces and the effects of such textured surfaces on cell behavior. ( 1999 Elsevier Science Ltd. All rights reserved
Keywords: Basement membranes; Cell behavior
1. Introduction
Fundamental knowledge of cell—substrate interactionsis important for tissue engineering, in the development ofmedical implants, and the production of pharmaceu-ticals. Cell—substrate interaction may also explain differ-ences in cell behavior in vivo and in vitro. To gain insightinto these interactions, a logical approach is to investi-gate the substrates on which cells attach and grow inliving systems.
Basement membranes are found throughout the verte-brate body and serve as substrata for overlying cellularstructures. Basement membranes consist of extracellularmatrix (ECM) components, including fibrous collagen,hyaluronic acid, proteoglycans, laminin, and fibronectin.The effects of the surface chemistry on cell and tissuefunction has been explored extensively in the past fewdecades. Hyaluronic acid, for example, has been shownto inhibit cell—cell adhesion and promote cell migration,
and laminin can prevent cell migration [1, 2]. Fibronec-tin has been observed to allow greater translocation ofcells than does laminin [3], and excess ECM has beenshown to inhibit endothelial cell replication by causingincreased cell—ECM adhesion and cytoskeletal re-arrangements [4]. One well-understood mechanism inwhich components of the basement membrane modulatecell behavior is the activation of plasma membrane integ-rin receptors, such as RGD that bind to ligands on thebasement membrane [5—7]. In addition to mediating cellattachment, integrin receptors also act as signaling mol-ecules, activating intracellular pathways important in cellgrowth and survival [8—11]. In the cornea, componentsof the basement membrane have been shown to influencethe distribution of cytoskeletal elements and of their ownintegrin receptors [12, 13] as well as modulating prolifer-ation [14], migration [15, 16], and differentiation [17].
The mechanical and tensile properties of the basementmembrane also influence fundamental cell behaviors.Cells can sense restraining forces and respond bystrengthening cytoskeletal linkages [18]. The strength ofthese integrin—cytoskeleton links depend on both the
0142-9612/99/$ — see front matter ( 1999 Elsevier Science Ltd. All rights reserved.PII: S 0 1 4 2 - 9 6 1 2 ( 9 8 ) 0 0 2 0 9 - 9

Fig. 1. Scanning electron micrograph of a corneal epithelial basementmembrane of Macaque monkey. After the corneal epithelium wasstripped, the cornea was fixed in 2% glutaraldehyde, dried by thecritical point method, and imaged on Hitachi S-900 scanning electronmicroscope (Bar"1 lm).
matrix rigidity as well as its biochemical composition[18]. When cells bind to ligands on the basement mem-brane, the cell receptors can act as mechanochemicaltransducers, activating signal transduction pathwaysand modulating gene expression [19]. Fibroblasts at-tached to strained collagen matrix produced more tenas-cin and collagen XII than those attached to a morerelaxed matrix [19]. Resilience and deformability alsoinfluences in vitro migration and morphology of somecells [3].
In addition to biochemical and mechanical properties,basement membranes possess a complex, three-dimen-sional topography consisting of nanometer sized features.Physical topography is known to affect cell behavior. Forreviews on the subject of the influence of substratumtopography on cells, see Curtis and Clark [20], vonRecum and van Kooten [21], Curtis and Wilkinson [22],Singhvi et al. [23], and Clark [24]. Paul Weiss, amongothers, pioneered the field of ‘contact guidance’ duringthe 1930s, 1940s and 1950s [25]. Rosenberg claimed, asearly as 1962 and 1963, that nanometer sized featuresinfluenced cells [26, 27]. Despite recognition of the im-portance of substratum topography, relatively little isknown about the effects of topographical features ofnanometer scale on cell behavior. The purpose of thispaper is to: (1) describe the nanometer scale topographyof basement membranes, (2) review the techniques usedto fabricate synthetic micro- and nano-structured surfa-ces, and (3) discuss the effects of micro- and nano-struc-tured synthetic surfaces on cellular behavior.
2. Topography of the basement membrane
Basement membranes are comprised of a complexmixture of pores, ridges, and fibers which have sizes in thenanometer range. A scanning electron micrographof a corneal epithelial basement membrane is shown inFig. 1. Abrams and coworkers used scanning electronmicroscopy, transmission electron microscopy, andatomic force microscopy to measure the sizes of featureson the surface of the corneal epithelial basement mem-brane of the Macaque monkey [28]. The average featureheights were between 147 and 191 nm and the averagefiber width was 77 nm [28, 29]. They found that poresmade up 15% of the total surface area of the membraneand had an average diameter of 72 nm [28]. Similarfeatures were observed on the human corneal epithelialbasement membrane [30] and on MatrigelTM, a commer-cially available basement membrane matrix [31].
Shirato et al. studied the glomerular basement mem-brane of the rat kidney and observed a meshwork offibrils 5—9 nm thick and pores 11—30 nm wide on thelamina rara interna and 6—11 nm fibrils and 10—24 nmpores on the lamina rara externa [32]. Yamasaki andcoworkers examined the pores and fibers present in
bovine glomerular and tubular basement membranes[33]. They found pore diameters near 10 nm in theglomerular membranes and pores near 12 nm in dia-meter in the tubular membranes. Fibrous strands wereseen to have widths of 3—15 nm. Another study byHironaka et al. found that the glomerular, tubular, andBowman’s capsule basement membranes of the rat kid-ney all had mean fiber diameters of 6—7 nm while havingmean pore diameters of 9.7, 14.1, and 13.1 nm, respective-ly [34].
3. Fabrication of micro- and nano-structured surfaces
A list of the fabrication strategies employed to createsynthetic substrates with topography is given in Table 1.The majority of the studies used photolithography toproduce features with controlled dimensions and specificshapes. Among the first to employ photolithography tocreate controlled features for the study of cell behaviorwas Brunette et al. [35].
A simplified schematic diagram of the photolitho-graphic process is shown in Fig. 2. A substrate is coatedwith a thin polymeric film called a resist. The resist isexposed to light through a mask such that the lightirradiates only selected regions of the resist. A photo-chemical reaction in the irridiated regions renders thoseregions either more soluble or less soluble in a solventcalled a developer. Hence, immersion in the developeryields either a positive-tone image of the mask, or a
574 R.G. Flemming et al. / Biomaterials 20 (1999) 573—588
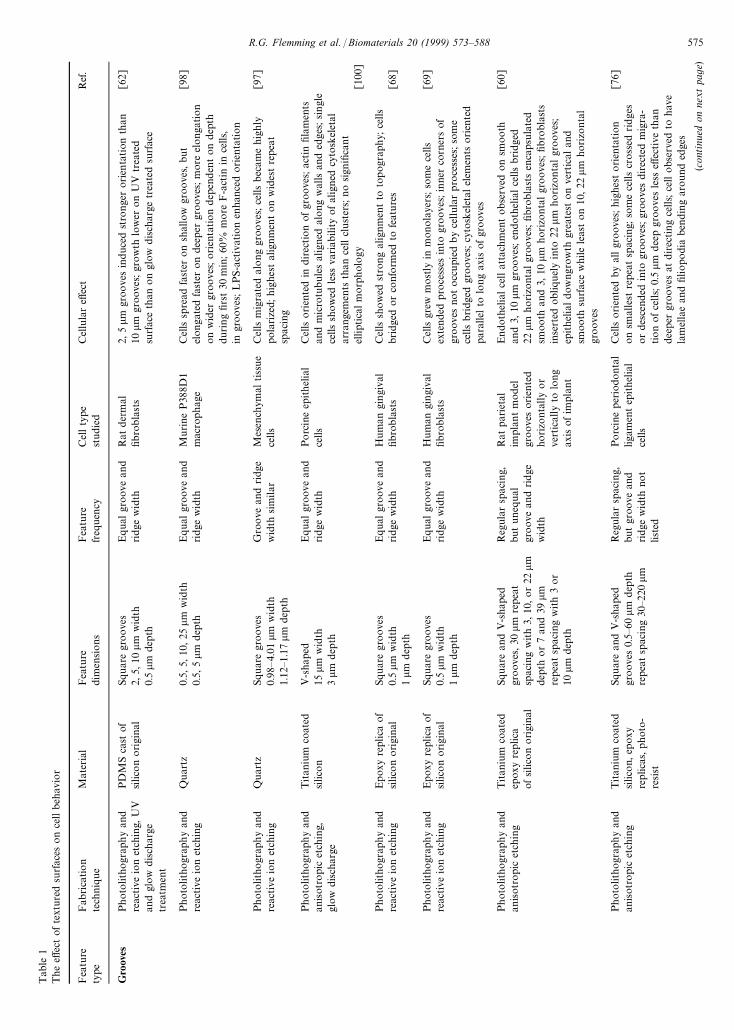
Tab
le1
The
effec
tof
text
ured
surfac
eson
cell
beh
avio
r
Fea
ture
Fab
rica
tion
Mat
eria
lFea
ture
Fea
ture
Cel
lty
pe
Cel
lula
reff
ect
Ref
.ty
pete
chniq
ue
dim
ensions
freq
uen
cyst
udi
ed
Gro
oves
Photo
litho
grap
hy
and
reac
tive
ion
etch
ing,
UV
and
glow
disch
arge
trea
tmen
t
PD
MS
cast
ofsilic
on
origi
nal
Squar
egr
oove
s2,
5,10
lmw
idth
0.5
lmde
pth
Equa
lgr
oov
ean
dridge
wid
thR
atder
mal
fibro
bla
sts
2,5
lmgr
oove
sin
duc
edst
ronge
ror
ienta
tion
than
10lm
groov
es;gr
ow
thlo
wer
on
UV
trea
ted
surfac
eth
anon
glow
disch
arge
trea
ted
surfac
e
[62]
Photo
litho
grap
hy
and
reac
tive
ion
etch
ing
Quar
tz0.
5,5,
10,25
lmw
idth
0.5,
5lm
dep
thEqua
lgr
oov
ean
dridge
wid
thM
urine
P38
8D1
mac
rophag
eC
ells
spre
adfa
ster
on
shal
low
groove
s,but
elon
gate
dfa
ster
on
deep
ergr
oov
es;m
ore
elong
atio
non
wid
ergr
oove
s;orien
tation
dep
enden
ton
dep
thdu
ring
firs
t30
min
;60
%m
ore
F-a
ctin
ince
lls,
ingr
oov
es;L
PS-a
ctiv
atio
nen
hance
dorien
tation
[98]
Photo
litho
grap
hy
and
reac
tive
ion
etch
ing
Quar
tzSq
uar
egr
oove
s0.
98—4.
01lm
wid
th1.
12—1
.17
lmde
pth
Gro
ove
and
ridg
ew
idth
sim
ilar
Mes
ench
ymal
tiss
ue
cells
Cel
lsm
igra
ted
along
groo
ves;
cells
bec
ame
high
lypo
larize
d;hig
hest
alig
nm
enton
wid
estre
pea
tsp
acin
g
[97]
Photo
litho
grap
hy
and
anisot
ropi
cet
chin
g,gl
owdisch
arge
Titan
ium
coat
edsilic
on
V-s
hape
d15
lmw
idth
3lm
dep
th
Equa
lgr
oov
ean
dridge
wid
thPorc
ine
epithel
ial
cells
Cel
lsorien
ted
indirec
tion
ofgr
oov
es;ac
tin
fila
men
tsan
dm
icro
tubule
sal
igne
dal
ong
wal
lsan
ded
ges;
singl
ece
llssh
ow
edle
ssva
riab
ility
ofal
igne
dcy
tosk
elet
alar
rang
emen
tsth
ance
llcl
ust
ers;
no
sign
ifica
nt
ellip
tica
lm
orp
hol
ogy
[100
]
Photo
litho
grap
hy
and
reac
tive
ion
etch
ing
Epox
yre
plic
aof
silic
on
origi
nal
Squar
egr
oove
s0.
5lm
wid
th1
lmdep
th
Equa
lgr
oov
ean
dridge
wid
thH
um
angi
ngi
val
fibro
bla
sts
Cel
lssh
ow
edst
rong
alig
nm
ent
toto
pog
raph
y;ce
llsbr
idge
dor
confo
rmed
tofe
ature
s[6
8]
Photo
litho
grap
hy
and
reac
tive
ion
etch
ing
Epox
yre
plic
aof
silic
on
origi
nal
Squar
egr
oove
s0.
5lm
wid
th1
lmdep
th
Equa
lgr
oov
ean
dridge
wid
thH
um
angi
ngi
val
fibro
bla
sts
Cel
lsgr
ewm
ost
lyin
mon
ola
yers
;so
me
cells
exte
nded
pro
cess
esin
togr
oove
s;in
ner
corn
ers
of
groov
esno
tocc
upi
edby
cellul
arpro
cess
es;s
om
ece
llsbridg
edgr
oov
es;cy
tosk
elet
alel
emen
tsorien
ted
para
llelto
long
axis
ofgr
oov
es
[69]
Photo
litho
grap
hy
and
anisot
ropi
cet
chin
gTitan
ium
coat
edep
oxy
replic
aof
silic
on
origi
nal
Squar
ean
dV
-sha
ped
groov
es,30
lmre
peat
spac
ing
with
3,10
,or
22lm
dept
hor
7an
d39
lmre
pea
tsp
acin
gw
ith
3or
10lm
dep
th
Reg
ular
spac
ing,
but
uneq
ual
groov
ean
dridge
wid
th
Rat
par
ieta
lim
plan
tm
odel
groov
esor
iente
dho
rizo
nta
lly
or
vert
ical
lyto
long
axis
ofim
pla
nt
Endo
thel
ialce
llat
tach
men
tob
serv
edon
smooth
and
3,10
lmgr
oove
s;en
doth
elia
lce
llsbridg
ed22
lmhorizo
ntal
groo
ves;
fibr
obla
sts
enca
psu
late
dsm
ooth
and
3,10
lmhor
izonta
lgr
oove
s;fibro
blas
tsin
sert
edob
liquel
yin
to22
lmhorizo
nta
lgr
oov
es;
epithel
ialdow
ngro
wth
grea
test
on
vert
ical
and
smoo
thsu
rfac
ew
hile
leas
ton
10,22
lmhor
izonta
lgr
oov
es
[60]
Photo
litho
grap
hy
and
anisot
ropi
cet
chin
gTitan
ium
coat
edsilic
on,
epox
yre
plic
as,p
hot
o-
resist
Squar
ean
dV
-sha
ped
groov
es0.
5—60
lmdep
thre
pea
tsp
acin
g30
—220
lm
Reg
ular
spac
ing,
but
groo
vean
dridge
wid
thno
tlis
ted
Porc
ine
per
iodo
nta
llig
amen
tep
ithe
lial
cells
Cel
lsorien
ted
byal
lgr
oov
es;h
ighe
storien
tation
onsm
alle
stre
peat
spac
ing;
som
ece
llscr
oss
edridge
sor
desc
ended
into
groo
ves;
groo
ves
direc
ted
mig
ra-
tion
ofce
lls;
0.5
lmdee
pgr
oove
sle
sseff
ective
than
deep
ergr
oove
sat
direc
ting
cells
;ce
llob
serv
edto
hav
ela
mel
lae
and
filio
podi
aben
din
gar
ound
edge
s
[76]
(con
tinu
edon
nextpa
ge)
R.G. Flemming et al. / Biomaterials 20 (1999) 573—588 575
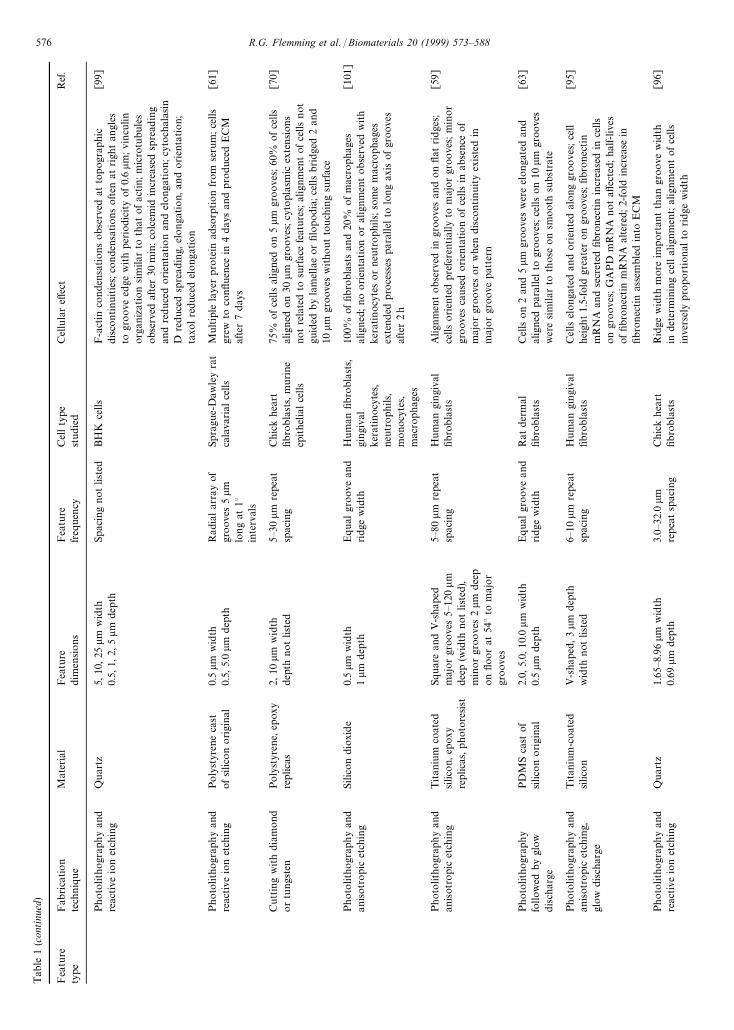
Tab
le1
(con
tinu
ed)
Fea
ture
Fab
rica
tion
Mat
eria
lFea
ture
Fea
ture
Cel
lty
pe
Cel
lula
reff
ect
Ref
.ty
pete
chniq
ue
dim
ensions
freq
uen
cyst
udi
ed
Photo
litho
grap
hy
and
reac
tive
ion
etch
ing
Quar
tz5,
10,25
lmw
idth
0.5,
1,2,
5lm
depth
Spac
ing
notlis
ted
BH
Kce
llsF-a
ctin
conde
nsa
tions
obs
erve
dat
topog
raphi
cdi
scontinui
ties
;con
dens
atio
nsoften
atrigh
tan
gles
togr
oov
eed
gew
ith
per
iodi
city
of0.
6lm
;vi
ncu
linor
ganiz
atio
nsim
ilar
toth
atofac
tin;m
icro
tubul
esob
serv
edaf
ter
30m
in;co
lcem
idin
crea
sed
spre
adin
gan
dre
duce
dorien
tation
and
elong
atio
n;cy
toch
alas
inD
reduc
edsp
read
ing,
elonga
tion,
and
orien
tation;
taxo
lre
duce
del
ong
atio
n
[99]
Photo
litho
grap
hy
and
reac
tive
ion
etch
ing
Poly
styr
ene
cast
ofsilic
on
origi
nal
0.5
lmw
idth
0.5,
5.0
lmdep
thR
adia
lar
ray
of
groov
es5
lmlo
ng
at1°
inte
rval
s
Spra
gue-
Daw
ley
rat
cala
varial
cells
Multip
lela
yer
pro
tein
adso
rption
from
seru
m;c
ells
grew
toco
nflue
nce
in4
days
and
pro
duce
dE
CM
afte
r7
days
[61]
Cut
ting
with
diam
ond
or
tung
sten
Poly
styr
ene,
epox
yre
plic
as2,
10lm
wid
thde
pth
not
liste
d5—
30lm
repe
atsp
acin
gC
hick
hea
rtfib
robla
sts,
murine
epithe
lialce
lls
75%
ofce
lls
alig
ned
on
5lm
groove
s;60
%ofce
llsal
igned
on
30lm
groove
s;cy
topl
asm
icex
tensions
not
rela
ted
tosu
rfac
efe
atur
es;al
ignm
ent
ofce
llsnot
guid
edby
lam
ella
eor
filo
pod
ia;c
ells
brid
ged
2an
d10
lmgr
oov
esw
ithout
touch
ing
surfac
e
[70]
Photo
litho
grap
hy
and
anisot
ropi
cet
chin
gSi
licon
dioxi
de
0.5
lmw
idth
1lm
dep
thEqua
lgr
oov
ean
dridge
wid
thH
um
anfib
robla
sts,
ging
ival
kera
tinocy
tes,
neut
roph
ils,
mono
cyte
s,m
acro
phag
es
100%
offib
robla
sts
and
20%
ofm
acro
pha
ges
alig
ned
;no
orien
tation
oral
ignm
ent
obse
rved
with
kera
tinocy
tes
or
neu
troph
ils;so
me
mac
roph
ages
exte
nded
pro
cess
espa
rallel
tolo
ng
axis
ofgr
oov
esaf
ter
2h
[101
]
Photo
litho
grap
hy
and
anisot
ropi
cet
chin
gTitan
ium
coat
edsilic
on,
epox
yre
plic
as,p
hot
ore
sist
Squar
ean
dV
-sha
ped
maj
or
groo
ves
5—12
0lm
deep
(wid
thnot
list
ed),
min
or
groo
ves
2lm
dee
pon
floor
at54
°to
maj
or
groov
es
5—80
lmre
peat
spac
ing
Hum
angi
ngi
val
fibro
bla
sts
Alig
nm
entobs
erve
din
groo
ves
and
on
flat
ridge
s;ce
llsorien
ted
pref
eren
tial
lyto
maj
orgr
oove
s;m
inor
groov
esca
use
dorien
tation
ofce
llsin
abse
nce
of
maj
or
groo
ves
or
whe
ndi
scon
tinuity
existe
din
maj
or
groo
vepat
tern
[59]
Photo
litho
grap
hy
follow
edby
glow
disch
arge
PD
MS
cast
ofsilic
on
origi
nal
2.0,
5.0,
10.0
lm
wid
th0.
5lm
depth
Equa
lgr
oov
ean
dridge
wid
thR
atder
mal
fibro
bla
sts
Cel
lson
2an
d5
lmgr
oove
sw
ere
elon
gate
dan
dal
igned
par
alle
lto
groov
es;c
ells
on10
lmgr
oove
sw
ere
sim
ilar
toth
ose
on
smoot
hsu
bst
rate
[63]
Photo
litho
grap
hy
and
anisot
ropi
cet
chin
g,gl
owdisch
arge
Titan
ium
-coa
ted
silic
on
V-s
hape
d,3
lmdep
thw
idth
not
liste
d6—
10lm
repe
atsp
acin
gH
um
angi
ngi
val
fibro
bla
sts
Cel
lsel
onga
ted
and
orien
ted
along
groo
ves;
cell
heig
ht1.
5-fo
ldgr
eate
ron
groove
s;fib
ronec
tin
mR
NA
and
secr
eted
fibr
onec
tin
incr
ease
din
cells
ongr
oov
es;G
APD
mR
NA
notaff
ecte
d;ha
lf-liv
esof
fibro
nec
tin
mR
NA
alte
red;
2-fo
ldin
crea
sein
fibro
nec
tin
asse
mbl
edin
toE
CM
[95]
Photo
litho
grap
hy
and
reac
tive
ion
etch
ing
Quar
tz1.
65—8
.96
lmw
idth
0.69
lmdep
th3.
0—32
.0lm
repea
tsp
acin
gC
hick
hea
rtfib
robla
sts
Rid
gew
idth
mor
eim
por
tant
than
groove
wid
thin
dete
rmin
ing
cell
alig
nm
ent;
alig
nm
ent
ofce
llsin
vers
ely
pro
por
tion
alto
ridge
wid
th
[96]
576 R.G. Flemming et al. / Biomaterials 20 (1999) 573—588
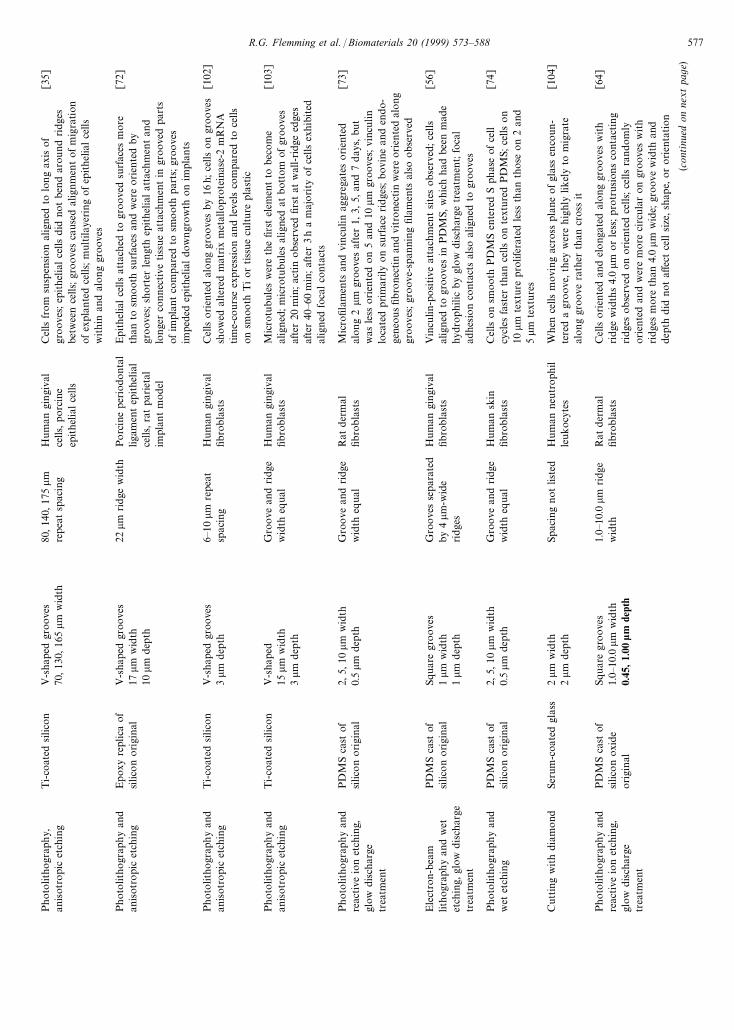
Photo
litho
grap
hy,
anisot
ropi
cet
chin
gTi-co
ated
silic
onV
-sha
ped
groo
ves
70,13
0,16
5lm
wid
th80
,14
0,17
5lm
repea
tsp
acin
gH
um
angi
ngi
val
cells
,por
cine
epithe
lialce
lls
Cel
lsfrom
susp
ension
alig
ned
tolo
ng
axis
of
groov
es;ep
ithel
ialce
llsdid
not
ben
dar
ound
ridge
sbe
twee
nce
lls;gr
oove
sca
used
alig
nm
entofm
igra
tion
ofex
plan
ted
cells
;m
ultila
yering
ofep
ithel
ialce
llsw
ithin
and
alon
ggr
oove
s
[35]
Photo
litho
grap
hy
and
anisot
ropi
cet
chin
gEpox
yre
plic
aof
silic
on
origi
nal
V-s
hape
dgr
oove
s17
lmw
idth
10lm
dep
th
22lm
ridge
wid
thPorc
ine
per
iodo
nta
llig
amen
tep
ithe
lial
cells
,rat
pariet
alim
plan
tm
odel
Epithel
ialce
lls
atta
ched
togr
oove
dsu
rfac
esm
ore
than
tosm
oot
hsu
rfac
esan
dw
ere
orien
ted
bygr
oov
es;sh
orte
rle
ngth
epithe
lialat
tach
men
tan
dlo
nge
rco
nnec
tive
tiss
ue
atta
chm
ent
ingr
oov
edpar
tsof
impl
antco
mpar
edto
smoot
hpar
ts;g
roove
sim
pede
dep
ithe
lialdo
wngr
ow
thon
impla
nts
[72]
Photo
litho
grap
hy
and
anisot
ropi
cet
chin
gTi-co
ated
silic
onV
-sha
ped
groo
ves
3lm
dep
th6—
10lm
repe
atsp
acin
gH
um
angi
ngi
val
fibro
bla
sts
Cel
lsorien
ted
along
groo
ves
by16
h;ce
llson
groove
ssh
ow
edal
tere
dm
atrix
met
allo
pro
tein
ase-
2m
RN
Atim
e-co
urs
eex
pres
sion
and
leve
lsco
mpar
edto
cells
onsm
ooth
Tior
tiss
ue
culture
pla
stic
[102
]
Photo
litho
grap
hy
and
anisot
ropi
cet
chin
gTi-co
ated
silic
onV
-sha
ped
15lm
wid
th3
lmdep
th
Gro
ove
and
ridg
ew
idth
equa
lH
um
angi
ngi
val
fibro
bla
sts
Mic
rotu
bul
esw
ere
the
first
elem
ent
tobe
com
eal
igned
;m
icro
tubu
les
alig
ned
atbot
tom
ofgr
oove
saf
ter
20m
in;ac
tin
obse
rved
first
atw
all-ridg
eed
ges
afte
r40
—60
min
;af
ter
3h
am
ajority
ofce
llsex
hibi
ted
alig
ned
foca
lco
ntac
ts
[103
]
Photo
litho
grap
hy
and
reac
tive
ion
etch
ing,
glow
disch
arge
trea
tmen
t
PD
MS
cast
ofsilic
on
origi
nal
2,5,
10lm
wid
th0.
5lm
depth
Gro
ove
and
ridg
ew
idth
equa
lR
atder
mal
fibro
bla
sts
Mic
rofil
amen
tsan
dvi
ncu
linag
greg
ates
orien
ted
alon
g2
lmgr
oove
saf
ter
1,3,
5,an
d7
day
s,bu
tw
asle
ssor
iente
don
5an
d10
lmgr
oov
es;vi
ncu
linlo
cate
dprim
arily
on
surfac
eridge
s;bov
ine
and
endo-
gene
ousfibro
nect
inan
dvi
trone
ctin
wer
eor
iente
dal
ong
groov
es;gr
oov
e-sp
anni
ng
fila
men
tsal
soob
serv
ed
[73]
Ele
ctro
n-bea
mlit
hog
raph
yan
dw
etet
chin
g,gl
owdisch
arge
trea
tmen
t
PD
MS
cast
ofsilic
on
origi
nal
Squar
egr
oove
s1
lmw
idth
1lm
dep
th
Gro
ove
sse
par
ated
by4
lm-w
ide
ridge
s
Hum
angi
ngi
val
fibro
bla
sts
Vin
culin
-positive
atta
chm
entsite
sob
serv
ed;ce
llsal
igned
togr
oove
sin
PD
MS,w
hic
hhad
been
mad
ehy
drop
hilic
by
glow
disc
har
getr
eatm
ent;
foca
lad
hesion
cont
acts
also
alig
ned
togr
oove
s
[56]
Photo
litho
grap
hy
and
wet
etch
ing
PD
MS
cast
ofsilic
on
origi
nal
2,5,
10lm
wid
th0.
5lm
depth
Gro
ove
and
ridg
ew
idth
equa
lH
um
ansk
infib
robla
sts
Cel
lson
smoo
thP
DM
Sen
tere
dS
phas
eof
cell
cycl
esfa
ster
than
cells
on
text
ured
PD
MS;
cells
on
10lm
text
ure
prol
ifera
ted
less
than
thos
eon
2an
d5
lmte
xture
s
[74]
Cut
ting
with
diam
ond
Seru
m-c
oate
dgl
ass
2lm
wid
th2
lmdep
thSp
acin
gnot
liste
dH
um
anne
utr
oph
ille
ukoc
ytes
Whe
nce
llsm
ovin
gac
ross
plan
eofgl
ass
enco
un-
tere
da
groove
,they
wer
ehig
hly
likel
yto
mig
rate
alon
ggr
oov
era
ther
than
cros
sit
[104
]
Photo
litho
grap
hy
and
reac
tive
ion
etch
ing,
glow
disch
arge
trea
tmen
t
PD
MS
cast
ofsilic
on
oxi
deor
igin
al
Squar
egr
oove
s1.
0—10
.0lm
wid
th0.
45,1.
00lm
dept
h
1.0—
10.0
lmridg
ew
idth
Rat
der
mal
fibro
bla
sts
Cel
lsorien
ted
and
elong
ated
along
groo
ves
with
ridge
wid
ths
4.0
lmor
less
;pro
trus
ions
cont
acting
ridge
sobse
rved
on
orien
ted
cells
;ce
llsra
ndo
mly
orie
nted
and
wer
em
ore
circ
ula
ron
groov
esw
ith
ridge
sm
ore
than
4.0
lm
wid
e;gr
oov
ew
idth
and
dept
hdid
not
affec
tce
llsize
,sha
pe,
ororien
tation
[64]
(con
tinu
edon
nextpa
ge)
R.G. Flemming et al. / Biomaterials 20 (1999) 573—588 577
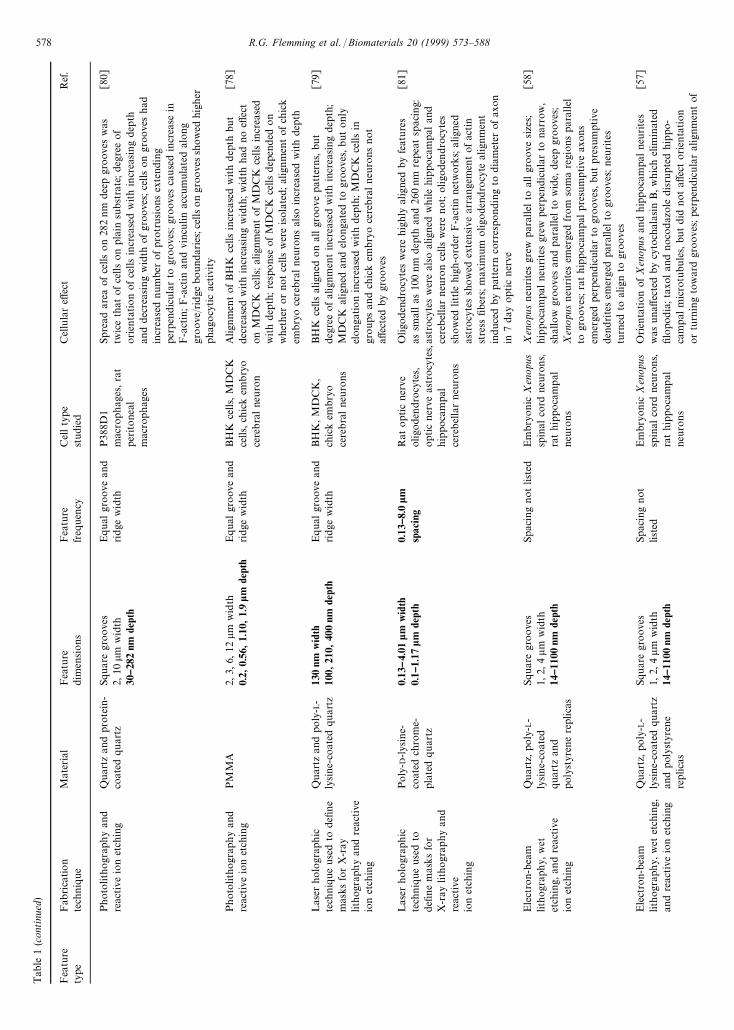
Tab
le1
(con
tinu
ed)
Fea
ture
Fab
rica
tion
Mat
eria
lFea
ture
Fea
ture
Cel
lty
pe
Cel
lula
reff
ect
Ref
.ty
pete
chniq
ue
dim
ensions
freq
uen
cyst
udi
ed
Photo
litho
grap
hy
and
reac
tive
ion
etch
ing
Quar
tzan
dpr
otei
n-co
ated
quar
tzSq
uar
egr
oove
s2,
10lm
wid
th30
–282
nmde
pth
Equa
lgr
oov
ean
dridge
wid
thP38
8D1
mac
rophag
es,ra
tpe
rito
neal
mac
rophag
es
Spre
adar
eaof
cells
on
282
nm
deep
groov
esw
astw
ice
that
ofce
lls
onpl
ain
subst
rate
;de
gree
of
orie
nta
tion
ofce
llsin
crea
sed
with
incr
easing
depth
and
dec
reas
ing
wid
thofgr
oove
s;ce
llson
groo
ves
had
incr
ease
dnum
ber
ofpro
trusions
exte
ndin
gpe
rpen
dic
ular
togr
oov
es;gr
oov
esca
use
din
crea
sein
F-a
ctin
;F
-act
inan
dvi
ncu
linac
cum
ula
ted
along
groov
e/ridge
bound
arie
s;ce
llson
groo
vessh
ow
edhi
gher
phag
ocy
tic
activi
ty
[80]
Photo
litho
grap
hy
and
reac
tive
ion
etch
ing
PM
MA
2,3,
6,12
lmw
idth
0.2,
0.56
,1.
10,1.
9lm
dept
hEqua
lgr
oov
ean
dridge
wid
thBH
Kce
lls,M
DC
Kce
lls,c
hic
kem
bry
oce
rebra
lne
uro
n
Alig
nm
entofB
HK
cells
incr
ease
dw
ith
dep
thbut
decr
ease
dw
ith
incr
easing
wid
th;w
idth
had
noeff
ect
onM
DC
Kce
lls;al
ignm
ent
ofM
DC
Kce
lls
incr
ease
dw
ith
dep
th;re
spons
eofM
DC
Kce
llsdep
ended
on
whe
ther
orno
tce
llsw
ere
isol
ated
;al
ignm
entofch
ick
embry
oce
rebr
alneu
rons
also
incr
ease
dw
ith
depth
[78]
Las
erholo
grap
hic
tech
niq
ue
used
tode
fine
mas
ksfo
rX
-ray
lithog
raph
yan
dre
active
ion
etch
ing
Quar
tzan
dpo
ly- L
-ly
sine
-coa
ted
quar
tz13
0nm
wid
th10
0,21
0,40
0nm
dept
hEqua
lgr
oov
ean
dridge
wid
thBH
K,M
DC
K,
chic
kem
bry
oce
rebra
lne
uro
ns
BH
Kce
llsal
igned
on
allgr
oove
pat
tern
s,bu
tde
gree
ofal
ignm
ent
incr
ease
dw
ith
incr
easing
dep
th;
MD
CK
alig
ned
and
elong
ated
togr
oove
s,but
only
elon
gation
incr
ease
dw
ith
dep
th;M
DC
Kce
llsin
group
san
dch
ick
embry
oce
rebr
alneu
rons
not
affec
ted
bygr
oov
es
[79]
Las
erholo
grap
hic
tech
niq
ue
used
tode
fine
mas
ks
for
X-r
aylith
ogr
aphy
and
reac
tive
ion
etch
ing
Poly
-D-lys
ine-
coat
edch
rom
e-pl
ated
quar
tz
0.13
–4.
01lm
wid
th0.
1–1.
17lm
dept
h0.
13–8
.0lm
spac
ing
Rat
optic
ner
veol
igode
ndr
ocy
tes,
optic
ner
veas
trocy
tes,
hippo
cam
pal
cere
bel
lar
neu
rons
Olig
oden
droc
ytes
wer
ehi
ghly
alig
ned
by
feat
ure
sas
smal
las
100
nmdep
than
d26
0nm
repe
atsp
acin
g:as
trocy
tes
wer
eal
soal
igned
while
hippo
cam
pal
and
cere
bel
lar
neu
ron
cells
wer
enot
;ol
igod
endr
ocyt
essh
ow
edlit
tle
hig
h-o
rder
F-a
ctin
netw
ork
s;al
igned
astr
ocy
tes
show
edex
tensive
arra
nge
men
tofac
tin
stre
ssfib
ers;
max
imum
olig
ode
ndro
cyte
alig
nm
ent
induc
edby
pat
tern
corr
espo
ndi
ngto
dia
met
erof
axon
in7
day
optic
ner
ve
[81]
Ele
ctro
n-bea
mlit
hog
raph
y,w
etet
chin
g,an
dre
active
ion
etch
ing
Quar
tz,po
ly-L
-ly
sine
-coa
ted
quar
tzan
dpo
lyst
yren
ere
plic
as
Squar
egr
oove
s1,
2,4
lm
wid
th14
–110
0nm
dept
h
Spac
ing
not
liste
dEm
bryo
nic
Xen
opus
spin
alco
rdne
uron
s,ra
thi
ppoca
mpal
neur
ons
Xen
opus
neu
rite
sgr
ewpar
alle
lto
allgr
oov
esize
s;hi
ppo
cam
pal
neurite
sgr
ewper
pendi
cula
rto
narr
ow,
shal
low
groov
esan
dpar
alle
lto
wid
e,de
epgr
oov
es;
Xen
opus
neu
rite
sem
erge
dfrom
som
are
gion
spar
alle
lto
groov
es;ra
thip
poca
mpa
lpr
esum
ptive
axon
sem
erge
dpe
rpen
dic
ular
togr
oove
s,bu
tpre
sum
ptive
dend
rite
sem
erge
dpa
ralle
lto
groove
s;neu
rite
stu
rned
toal
ign
togr
oove
s
[58]
Ele
ctro
n-bea
mlit
hog
raph
y,w
etet
chin
g,an
dre
active
ion
etch
ing
Quar
tz,po
ly-L
-ly
sine
-coa
ted
quar
tzan
dpol
ysty
rene
replic
as
Squar
egr
oove
s1,
2,4
lm
wid
th14
–110
0nm
dept
h
Spac
ing
not
liste
dEm
bryo
nic
Xen
opus
spin
alco
rdne
uron
s,ra
thi
ppoca
mpal
neur
ons
Orien
tation
ofX
enop
usan
dhip
poca
mpa
lneu
rite
sw
asuna
ffec
ted
bycy
toch
alas
inB
,whic
hel
imin
ated
filopo
dia
;tax
olan
dnoc
odaz
ole
disr
upte
dhip
po-
cam
pal
mic
rotu
bule
s,bu
tdid
not
affec
tor
ienta
tion
ortu
rnin
gto
war
dgr
oov
es;pe
rpen
dic
ula
ral
ignm
ent
of
[57]
578 R.G. Flemming et al. / Biomaterials 20 (1999) 573—588
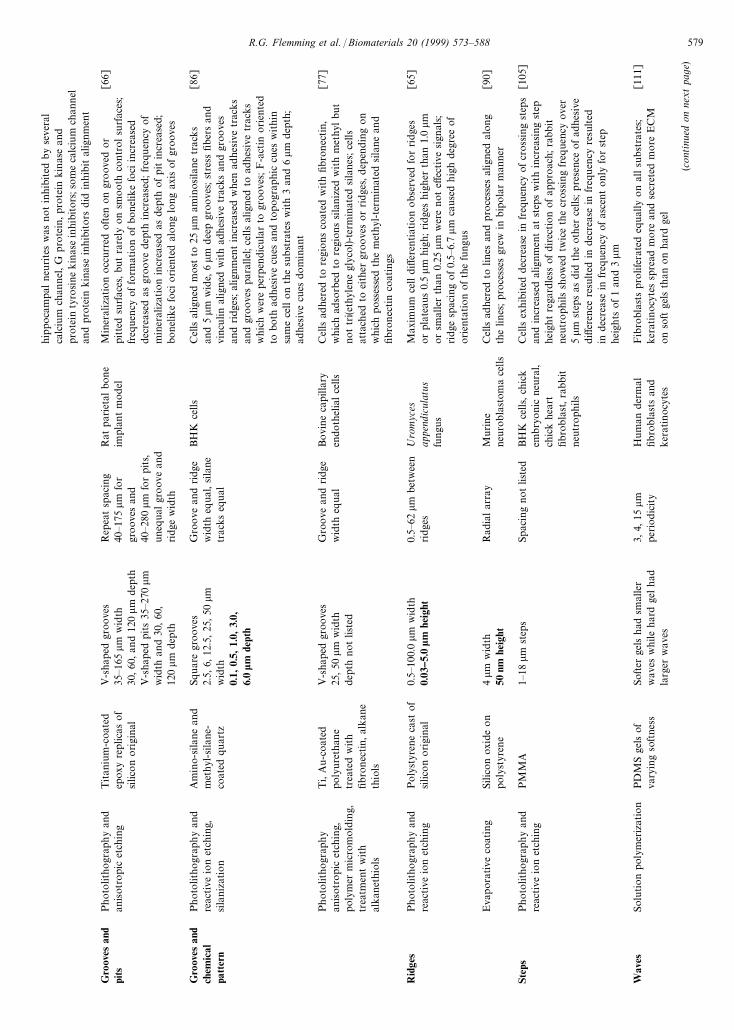
hippo
cam
palne
urite
sw
asno
tin
hib
ited
by
seve
ral
calc
ium
chan
nel
,Gpro
tein
,pro
tein
kinas
ean
dpr
otei
nty
rosine
kin
ase
inhi
bito
rs;s
om
eca
lciu
mch
annel
and
pro
tein
kin
ase
inhib
itor
sdid
inhib
ital
ignm
ent
Gro
oves
and
pits
Photo
litho
grap
hy
and
anisot
ropi
cet
chin
gTitan
ium
-coa
ted
epox
yre
plic
asof
silic
on
origi
nal
V-s
hape
dgr
oove
s35
—165
lmw
idth
30,60
,and
120
lmdep
thV
-sha
ped
pits
35—2
70lm
wid
than
d30
,60
,12
0lm
depth
Rep
eatsp
acin
g40
—175
lmfo
rgr
oov
esan
d40
—280
lmfo
rpits,
uneq
ual
groov
ean
dridge
wid
th
Rat
par
ieta
lbo
ne
impl
antm
odel
Min
eral
izat
ion
occ
urre
dof
ten
ongr
oov
edor
pitt
edsu
rfac
es,but
rare
lyon
smooth
contr
olsu
rfac
es;
freq
uency
offo
rmat
ion
ofbo
nel
ike
foci
incr
ease
dde
crea
sed
asgr
oove
dept
hin
crea
sed;freq
uen
cyof
min
eral
izat
ion
incr
ease
das
dept
hofpit
incr
ease
d;bo
nel
ike
foci
orien
ted
along
long
axis
ofgr
oove
s
[66]
Gro
oves
and
chem
ical
patter
n
Photo
litho
grap
hy
and
reac
tive
ion
etch
ing,
sila
niza
tion
Am
ino-
sila
nean
dm
ethyl
-sila
ne-
coat
edqu
artz
Squar
egr
oove
s2.
5,6,
12.5
,25
,50
lmw
idth
0.1,
0.5,
1.0,
3.0,
6.0
lmde
pth
Gro
ove
and
ridg
ew
idth
equa
l,sila
netr
acks
equal
BH
Kce
llsC
ells
alig
ned
mos
tto
25lm
amin
osila
netr
acks
and
5lm
wid
e,6
lmdee
pgr
oove
s;st
ress
fiber
san
dvi
nculin
alig
ned
with
adhes
ive
trac
ksan
dgr
oove
san
dridg
es;al
ignm
ent
incr
ease
dw
hen
adhe
sive
trac
ks
and
groov
espa
ralle
l;ce
llsal
igne
dto
adhe
sive
trac
ksw
hich
wer
eper
pendic
ula
rto
groo
ves;
F-a
ctin
orie
nted
tobo
thad
hes
ive
cues
and
topog
raphi
ccu
esw
ithi
nsa
me
cell
on
the
subst
rate
sw
ith
3an
d6
lmdep
th;
adhe
sive
cues
dom
inan
t
[86]
Photo
litho
grap
hy
anisot
ropi
cet
chin
g,pol
ymer
mic
rom
old
ing,
trea
tmen
tw
ith
alka
net
hiols
Ti,
Au-
coat
edpo
lyure
than
etr
eate
dw
ith
fibro
nec
tin,al
kan
eth
iols
V-s
hape
dgr
oove
s25
,50
lmw
idth
dept
hnot
liste
d
Gro
ove
and
ridg
ew
idth
equa
lBovi
neca
pilla
ryen
doth
elia
lce
llsC
ells
adhe
red
tore
gions
coat
edw
ith
fibro
nec
tin,
whic
had
sorb
edto
regi
ons
sila
niz
edw
ith
met
hylbu
tno
ttr
i(eth
ylen
egl
ycol
)-te
rmin
ated
sila
nes
;ce
llsat
tach
edto
eith
ergr
oove
sor
ridg
es,d
epen
din
gon
whi
chposs
esse
dth
em
ethy
l-te
rmin
ated
sila
nean
dfib
ronec
tin
coat
ings
[77]
Rid
ges
Photo
litho
grap
hy
and
reac
tive
ion
etch
ing
Poly
styr
ene
cast
of
silic
on
origi
nal
0.5—
100.
0lm
wid
th0.
03–5
.0lm
heig
ht0.
5—62
lmbe
twee
nridge
sº
rom
yces
appe
ndic
ulat
usfu
ngu
s
Max
imum
cell
diff
eren
tiat
ion
obs
erve
dfo
rridg
esor
plat
eaus
0.5
lm
high
;ridg
eshig
her
than
1.0
lmor
smal
ler
than
0.25
lm
wer
enot
effec
tive
sign
als;
ridg
esp
acin
gof0.
5—6.
7lm
cause
dhig
hde
gree
ofor
ienta
tion
ofth
efu
ngus
[65]
Eva
por
ativ
eco
atin
gSi
licon
oxid
eon
poly
styr
ene
4lm
wid
th50
nmhe
ight
Rad
ialar
ray
Murine
neur
obla
stom
ace
lls
Cel
lsad
here
dto
lines
and
pro
cess
esal
igne
dal
ong
the
lines
;pro
cess
esgr
ewin
bip
ola
rm
anne
r[9
0]
Step
sPhoto
litho
grap
hy
and
reac
tive
ion
etch
ing
PM
MA
1—18
lmst
eps
Spac
ing
not
liste
dBH
Kce
lls,ch
ick
embry
oni
cne
ural
,ch
ick
hear
tfib
robla
st,r
abbit
neut
roph
ils
Cel
lsex
hib
ited
dec
reas
ein
freq
uen
cyof
cros
sing
step
san
din
crea
sed
alig
nmen
tat
step
sw
ith
incr
easing
step
heig
htre
gard
less
ofdi
rect
ion
ofap
pro
ach;
rabbit
neut
roph
ilssh
owed
twic
eth
ecr
ossing
freq
uenc
yov
er5
lmst
eps
asdid
the
other
cells
;pre
senc
eofad
hesive
differ
ence
resu
lted
inde
crea
sein
freq
uen
cyre
sulted
inde
crea
sein
freq
uen
cyof
asce
nt
only
for
step
heig
hts
of1
and
3lm
[105
]
Wav
esSol
ution
pol
ymer
izat
ion
PD
MS
gels
ofva
ryin
gso
ftne
ssSo
fter
gels
had
smal
ler
wav
esw
hile
hard
gelhad
larg
erw
aves
3,4,
15lm
periodic
ity
Hum
ande
rmal
fibro
bla
sts
and
kera
tinocy
tes
Fib
robl
asts
prolife
rate
deq
ual
lyon
allsu
bst
rate
s;ke
ratinocy
tes
spre
adm
ore
and
secr
eted
mor
eEC
Mon
soft
gels
than
on
har
dge
l
[111
]
(con
tinu
edon
nextpa
ge)
R.G. Flemming et al. / Biomaterials 20 (1999) 573—588 579
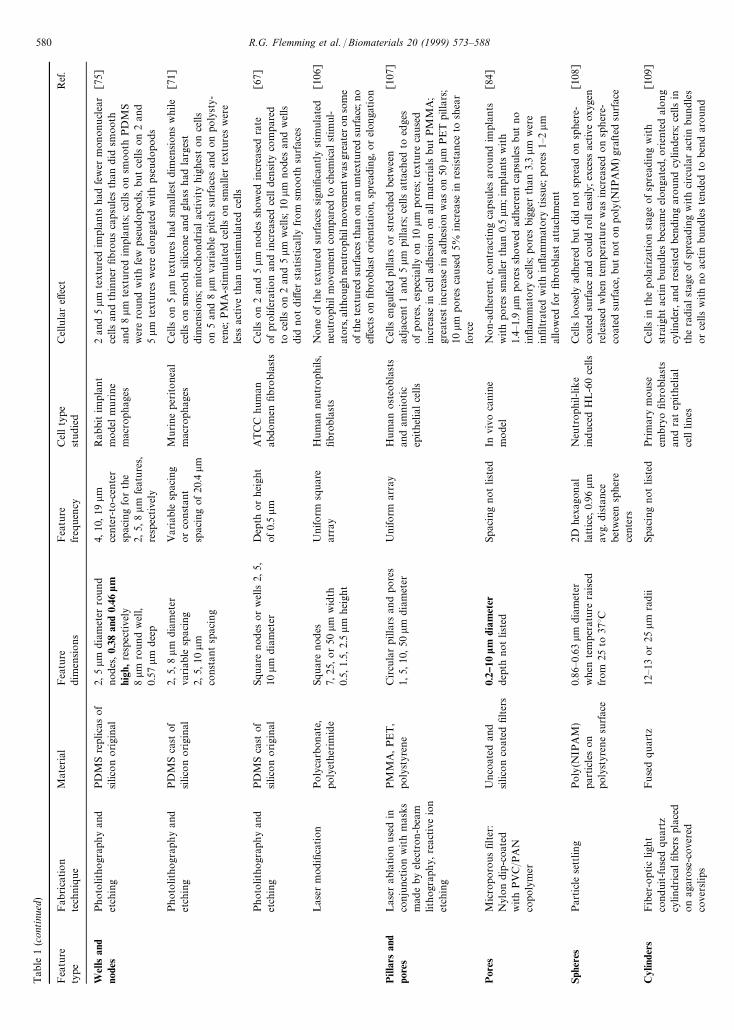
Tab
le1
(con
tinu
ed)
Fea
ture
Fab
rica
tion
Mat
eria
lFea
ture
Fea
ture
Cel
lty
pe
Cel
lula
reff
ect
Ref
.ty
pete
chniq
ue
dim
ensions
freq
uen
cyst
udi
ed
Wel
lsan
dno
des
Photo
litho
grap
hy
and
etch
ing
PD
MS
replic
asof
silic
on
origi
nal
2,5
lmdia
met
erro
und
nodes
,0.
38an
d0.
46lm
high
,re
spec
tive
ly8
lmro
und
wel
l,0.
57lm
dee
p
4,10
,19
lmce
nter
-to-c
ente
rsp
acin
gfo
rth
e2,
5,8
lm
feat
ure
s,re
spec
tive
ly
Rab
bit
impla
nt
mode
lm
urine
mac
rophag
es
2an
d5
lmte
xture
dim
pla
nts
had
few
erm
onon
ucl
ear
cells
and
thin
ner
fibro
usca
psul
esth
andi
dsm
ooth
and
8lm
text
ured
impla
nts;
cells
on
smoot
hP
DM
Sw
ere
roun
dw
ith
few
pseu
dopo
ds,bu
tce
llson
2an
d5
lmte
xture
sw
ere
elon
gate
dw
ith
pse
udopo
ds
[75]
Photo
litho
grap
hy
and
etch
ing
PD
MS
cast
ofsilic
on
origi
nal
2,5,
8lm
diam
eter
variab
lesp
acin
g2,
5,10
lmco
nsta
ntsp
acin
g
Var
iable
spac
ing
orco
nst
ant
spac
ing
of20
.4lm
Murine
per
itone
alm
acro
phag
esC
ells
on
5lm
text
ure
shad
smal
lest
dim
ensions
whi
lece
llson
smoo
thsilico
ne
and
glas
shad
larg
est
dim
ensions
;mitoch
ondr
ialac
tivi
tyhig
hest
once
llson
5an
d8
lmva
riab
lepitch
surfac
esan
don
poly
sty-
rene;
PM
A-s
tim
ula
ted
cells
on
smal
ler
text
ures
wer
ele
ssac
tive
than
unst
imul
ated
cells
[71]
Photo
litho
grap
hy
and
etch
ing
PD
MS
cast
ofsilic
on
origi
nal
Squar
enod
esor
wel
ls2,
5,10
lmdia
met
erD
epth
or
hei
ght
of0.
5lm
ATC
Chu
man
abdo
men
fibro
blas
tsC
ells
on
2an
d5
lmnodes
show
edin
crea
sed
rate
ofpr
olifer
atio
nan
din
crea
sed
cell
den
sity
com
par
edto
cells
on
2an
d5
lmw
ells;10
lmnode
san
dw
ells
did
notdiff
erst
atistica
llyfrom
smoo
thsu
rfac
es
[67]
Las
erm
odifi
cation
Poly
carb
onat
e,po
lyet
herim
ide
Squar
enod
es7,
25,or
50lm
wid
th0.
5,1.
5,2.
5lm
hei
ght
Unifo
rmsq
uar
ear
ray
Hum
anne
utr
oph
ils,
fibro
bla
sts
None
ofth
ete
xture
dsu
rfac
essign
ifica
ntly
stim
ula
ted
neut
roph
ilm
ovem
entco
mpar
edto
chem
ical
stim
ul-
ator
s,al
thoug
hne
utr
ophi
lmov
emen
twas
grea
teron
som
eof
the
text
ure
dsu
rfac
esth
anon
anun
text
ured
surfac
e;no
effec
tson
fibro
blas
tor
ient
atio
n,sp
read
ing,
orel
onga
tion
[106
]
Pill
ars
and
pore
sLas
erab
lation
use
din
conju
nct
ion
with
mas
ksm
ade
by
elec
tron-b
eam
lithog
raph
y,re
active
ion
etch
ing
PM
MA
,PE
T,
poly
styr
ene
Circu
lar
pill
ars
and
pore
s1,
5,10
,50
lmdia
met
erU
nifo
rmar
ray
Hum
anos
teobla
sts
and
amnio
tic
epithe
lialce
lls
Cel
lsen
gulfe
dpill
ars
or
stre
tche
dbe
twee
nad
jace
nt1
and
5lm
pill
ars;
cells
atta
ched
toed
ges
ofpo
res,
espe
cial
lyon
10lm
pore
s;te
xture
caus
edin
crea
sein
cell
adhes
ion
onal
lm
ater
ials
butPM
MA
;gr
eate
stin
crea
sein
adhe
sion
was
on
50lm
PE
Tpill
ars;
10lm
pore
sca
use
d5%
incr
ease
inre
sist
ance
tosh
ear
forc
e
[107
]
Por
esM
icro
poro
usfil
ter:
Nyl
ondi
p-c
oate
dw
ith
PV
C/P
AN
copoly
mer
Unco
ated
and
silic
on
coat
edfil
ters
0.2–
10lm
diam
eter
dept
hnot
liste
dSp
acin
gnot
liste
dIn
vivo
cani
nem
ode
lN
on-
adher
ent,
cont
ract
ing
caps
ule
sar
ound
impl
ants
with
pore
ssm
alle
rth
an0.
5lm
;im
pla
nts
with
1.4—
1.9
lmpo
res
show
edad
here
ntca
psul
esbut
no
inflam
mat
ory
cells
;por
esbi
gger
than
3.3
lmw
ere
infiltra
ted
with
infla
mm
ator
ytiss
ue;
pore
s1—
2lm
allo
wed
for
fibro
blas
tat
tach
men
t
[84]
Sphe
res
Par
ticl
ese
ttlin
gPoly
(NIP
AM
)pa
rtic
les
onpo
lyst
yren
esu
rfac
e
0.86
—0.6
3lm
diam
eter
whe
nte
mper
atur
era
ised
from
25to
37°C
2Dhe
xago
nal
latt
ice,
0.96
lm
avg.
dista
nce
betw
een
sphe
rece
nter
s
Neu
trop
hil-
like
induc
edH
L-6
0ce
llsC
ells
loos
ely
adher
edbu
tdi
dnot
spre
adon
sphe
re-
coat
edsu
rfac
ean
dco
uld
roll
easily
;exc
ess
active
oxyg
enre
leas
edw
hen
tem
pera
ture
was
incr
ease
don
spher
e-co
ated
surfac
e,bu
tnot
on
pol
y(N
IPA
M)gr
afte
dsu
rfac
e[108
]
Cyl
inde
rsFib
er-o
ptic
light
condui
t-fu
sed
quar
tzcy
lindr
ical
fibe
rspl
aced
onag
arose
-cov
ered
cove
rslip
s
Fuse
dquar
tz12
—13
or25
lmra
dii
Spac
ing
not
liste
dPrim
ary
mou
seem
bry
ofib
robl
asts
and
rat
epithe
lial
cell
lines
Cel
lsin
the
pola
riza
tion
stag
eofsp
read
ing
with
stra
ight
actin
bun
dle
sbe
cam
eel
onga
ted,
orien
ted
alon
gcy
linder
,and
resist
edben
ding
around
cylin
ders
;ce
llsin
the
radi
alst
age
ofsp
read
ing
with
circ
ula
rac
tin
bundle
sor
cells
with
no
actin
bun
dle
ste
nded
tobe
ndar
oun
d
[109
]
580 R.G. Flemming et al. / Biomaterials 20 (1999) 573—588
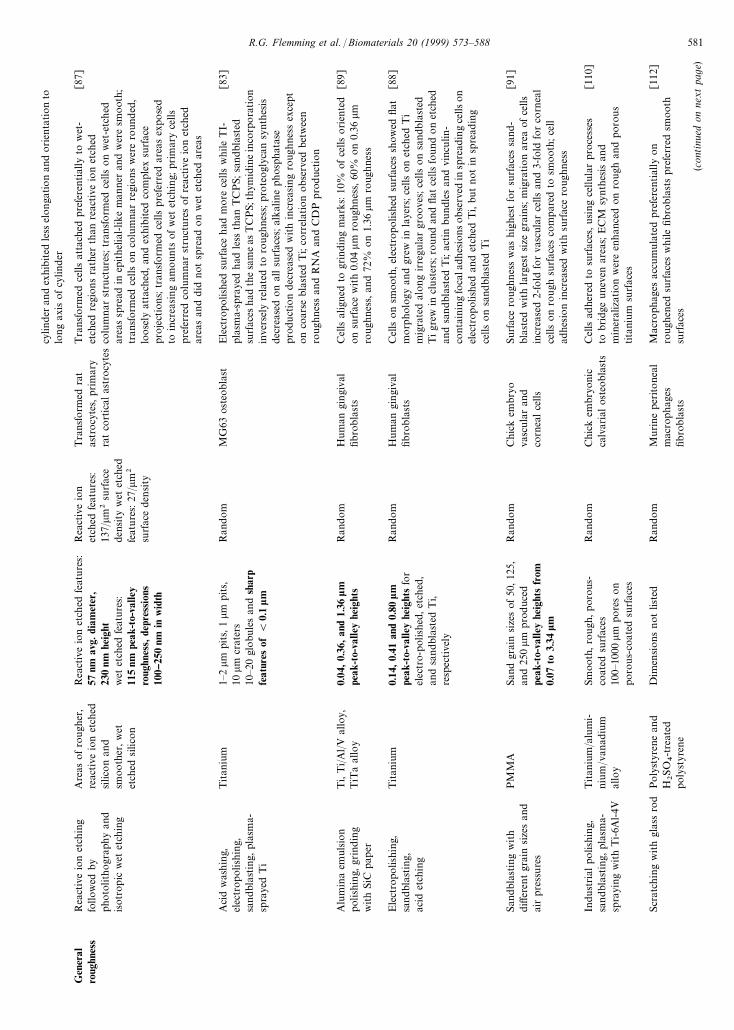
cylin
der
and
exhi
bited
less
elonga
tion
and
orien
tation
tolo
ng
axis
ofcy
linder
Gen
eral
roug
hnes
sR
eact
ive
ion
etch
ing
follow
edby
pho
tolith
ogr
aphy
and
isot
ropic
wet
etch
ing
Are
asofro
ugh
er,
reac
tive
ion
etch
edsilic
on
and
smoo
ther
,w
etet
ched
silic
on
Rea
ctiv
eio
net
ched
feat
ures
:57
nmav
g.di
amet
er,
230
nmhe
ight
wet
etch
edfe
ature
s:11
5nm
peak
-to-
valle
yro
ughn
ess,
depr
ession
s10
0–25
0nm
inwid
th
Rea
ctiv
eio
net
ched
feat
ure
s:13
7/lm
2su
rfac
ede
nsity
wet
etch
edfe
ature
s:27
/lm
2
surfac
ede
nsity
Tra
nsfo
rmed
rat
astr
ocy
tes,
prim
ary
rat
cort
ical
astr
ocy
tes
Tra
nsfo
rmed
cells
atta
ched
pref
eren
tial
lyto
wet
-et
ched
regi
ons
rath
erth
anre
active
ion
etch
edco
lum
nar
stru
ctur
es;t
rans
form
edce
llson
wet
-etc
hed
area
ssp
read
inep
ithel
ial-lik
em
anner
and
wer
esm
oot
h;
tran
sfor
med
cells
on
colu
mna
rre
gions
wer
ero
und
ed,
loose
lyat
tach
ed,an
dex
hibited
com
ple
xsu
rfac
epr
oje
ctio
ns;
tran
sform
edce
llspr
efer
red
area
sex
pose
dto
incr
easing
amoun
tsofw
etet
chin
g;pr
imar
yce
llspr
efer
red
colu
mna
rst
ruct
ure
sofre
active
ion
etch
edar
eas
and
did
not
spre
adon
wet
etch
edar
eas
[87]
Aci
dw
ashin
g,el
ectr
opol
ishin
g,sa
ndbl
asting
,pl
asm
a-sp
raye
dT
i
Titan
ium
1—2
lmpi
ts,1
lmpits,
10lm
crat
ers
10—2
0gl
obu
les
and
shar
pfe
atur
esof
(0.
1lm
Ran
dom
MG
63os
teobla
stEle
ctro
polis
hed
surfac
ehad
mor
ece
llsw
hile
TI-
plas
ma-
spra
yed
had
less
than
TC
PS;sa
ndbl
aste
dsu
rfac
eshad
the
sam
eas
TC
PS;
thym
idin
ein
corp
ora
tion
inve
rsel
yre
late
dto
rough
ness
;pr
ote
ogl
ycan
synt
hes
isde
crea
sed
on
allsu
rfac
es;al
kal
ine
phos
phat
ase
produ
ctio
ndec
reas
edw
ith
incr
easing
roug
hne
ssex
cept
onco
arse
blas
ted
Ti;
corr
elat
ion
obse
rved
betw
een
rough
nes
san
dR
NA
and
CD
Ppro
duc
tion
[83]
Alu
min
aem
ulsion
polish
ing,
grin
din
gw
ith
SiC
pap
er
Ti,
Ti/A
l/V
allo
y,TiT
aal
loy
0.04
,0.
36,an
d1.
36lm
peak
-to-
valle
yhe
ight
sR
andom
Hum
angi
ngi
val
fibro
bla
sts
Cel
lsal
igne
dto
grin
din
gm
arks
:10%
ofce
llsorien
ted
onsu
rfac
ew
ith
0.04
lm
rough
nes
s,60
%on
0.36
lmro
ughn
ess,
and
72%
on1.
36lm
roug
hne
ss
[89]
Ele
ctro
polis
hing,
sandbl
asting
,ac
idet
chin
g
Titan
ium
0.14
,0.
41an
d0.
80lm
peak
-to-
valle
yhe
ight
sfo
rel
ectr
o-po
lish
ed,et
ched
,an
dsa
ndb
last
edT
i,re
spec
tive
ly
Ran
dom
Hum
angi
ngi
val
fibro
bla
sts
Cel
lson
smoo
th,e
lect
ropolis
hed
surfac
essh
owed
flat
morp
holo
gyan
dgr
ewin
laye
rs;ce
llson
etch
edT
im
igra
ted
alon
girre
gula
rgr
oove
s;ce
llson
sand
bla
sted
Tigr
ewin
clust
ers;
round
and
flatce
llsfo
und
on
etch
edan
dsa
ndbl
aste
dT
i;ac
tin
bundl
esan
dvi
nculin
-co
ntai
ning
foca
ladh
esio
nsobse
rved
insp
read
ing
cells
onel
ectr
opol
ished
and
etch
edTi,
butnot
insp
read
ing
cells
on
sand
blas
ted
Ti
[88]
San
dbl
asting
with
diff
eren
tgr
ain
size
san
dai
rpre
ssure
s
PM
MA
Sand
grai
nsize
sof50
,12
5,an
d25
0lm
pro
duc
edpe
ak-t
o-va
lley
heig
hts
from
0.07
to3.
34lm
Ran
dom
Chi
ckem
bryo
vasc
ula
ran
dco
rnea
lce
lls
Surfac
ero
ugh
nes
sw
ashig
hest
for
surfac
essa
nd-
blas
ted
with
larg
est
size
grai
ns;m
igra
tion
area
ofce
llsin
crea
sed
2-fo
ldfo
rva
scul
arce
llsan
d3-
fold
for
corn
eal
cells
on
rough
surfac
esco
mpar
edto
smooth
;ce
llad
hesion
incr
ease
dw
ith
surfac
ero
ughn
ess
[91]
Indu
strial
polis
hing,
sandbl
asting
,pl
asm
a-sp
rayi
ng
with
Ti-6A
l-4V
Titan
ium
/alu
mi-
nium
/van
adiu
mal
loy
Smoo
th,ro
ugh
,por
ous
-co
ated
surfac
es10
0—10
00lm
pore
son
poro
us-
coat
edsu
rfac
es
Ran
dom
Chi
ckem
bryo
nic
calv
aria
los
teob
last
sC
ells
adhe
red
tosu
rfac
es,u
sing
cellul
arpro
cess
esto
brid
geun
even
area
s;EC
Msy
nth
esis
and
min
eral
izat
ion
wer
een
han
ced
onro
ugh
and
por
ous
tita
nium
surfac
es
[110
]
Scr
atch
ing
with
glas
sro
dPoly
styr
ene
and
H2S
O4-t
reat
edpo
lyst
yren
e
Dim
ension
snot
liste
dR
andom
Murine
per
itone
alm
acro
phag
esfib
robla
sts
Mac
roph
ages
accu
mula
ted
pref
eren
tial
lyon
rough
ened
surfac
esw
hile
fibro
bla
sts
pref
erre
dsm
oot
hsu
rfac
es
[112
]
(con
tinu
edon
nextpa
ge)
R.G. Flemming et al. / Biomaterials 20 (1999) 573—588 581
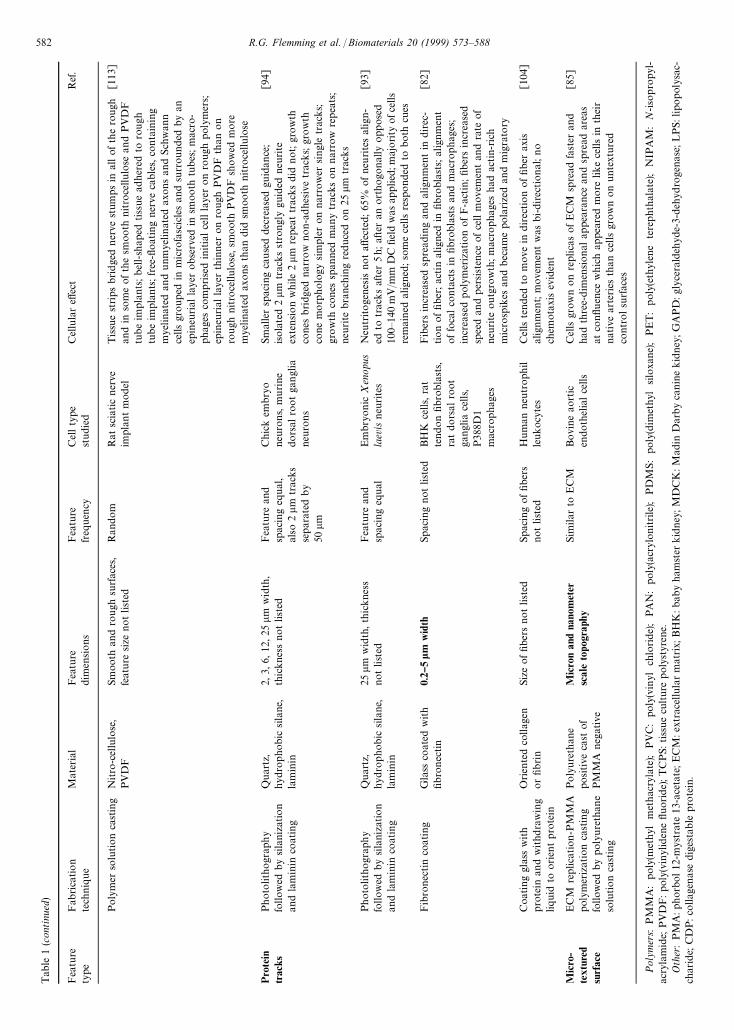
Tab
le1
(con
tinu
ed)
Fea
ture
Fab
rica
tion
Mat
eria
lFea
ture
Fea
ture
Cel
lty
pe
Cel
lula
reff
ect
Ref
.ty
pete
chniq
ue
dim
ensions
freq
uen
cyst
udi
ed
Poly
mer
solu
tion
cast
ing
Nitro
-cel
lulo
se,
PV
DF
Smoo
than
dro
ugh
surfac
es,
feat
ure
size
notlist
edR
ando
mR
atsc
iatic
ner
veim
plan
tm
odel
Tissu
est
rips
brid
ged
ner
vest
umps
inal
lofth
ero
ugh
and
inso
me
ofth
esm
ooth
nitro
cellu
lose
and
PV
DF
tube
impl
ants
;bel
l-sh
aped
tiss
ue
adher
edto
rough
tube
impl
ants
;fre
e-flo
atin
gner
veca
bles
,co
ntai
ning
mye
linat
edan
dunm
yelin
ated
axon
san
dSch
wan
nce
llsgr
ouped
inm
icro
fasc
icle
san
dsu
rroun
ded
byan
epin
eurial
laye
rob
serv
edin
smoo
thtu
bes
;m
acro
-ph
ages
com
prise
din
itia
lce
llla
yer
on
rough
poly
mer
s;ep
ineu
rial
laye
rth
inner
on
rough
PV
DF
than
on
rough
nitr
oce
llulo
se,s
moot
hP
VD
Fsh
ow
edm
ore
mye
linat
edax
ons
than
did
smoo
thni
troce
llulo
se
[113
]
Pro
tein
trac
ksPhoto
litho
grap
hy
follow
edby
sila
niza
tion
and
lam
inin
coat
ing
Quar
tz,
hydr
ophob
icsila
ne,
lam
inin
2,3,
6,12
,25
lmw
idth
,th
ickn
ess
notlis
ted
Fea
ture
and
spac
ing
equal
,al
so2
lm
trac
ksse
par
ated
by
50lm
Chi
ckem
bryo
neur
ons,
murine
dors
alro
otga
ngl
iane
urons
Smal
ler
spac
ing
cause
dde
crea
sed
guid
ance
;isola
ted
2lm
trac
ks
stro
ngly
guid
edne
urite
exte
nsion
while
2lm
repe
attr
acks
did
not;
grow
thco
nes
bridg
edna
rrow
non-a
dhe
sive
trac
ks;
grow
thco
nem
orp
holo
gysim
pler
onna
rrow
ersing
letr
acks
;gr
ow
thco
nes
span
ned
man
ytr
acks
on
nar
row
repea
ts;
neur
ite
bran
chin
gre
duce
don
25lm
trac
ks
[94]
Photo
litho
grap
hy
follow
edby
sila
niza
tion
and
lam
inin
coat
ing
Quar
tz,
hydr
ophob
icsila
ne,
lam
inin
25lm
wid
th,th
ickne
ssno
tlist
edFea
ture
and
spac
ing
equal
Em
bryo
nic
Xen
opus
laev
isneu
rite
sN
eutr
itoge
nesis
notaff
ecte
d;65
%of
neurite
sal
ign-
edto
trac
ks
afte
r5
h;af
ter
anor
thogo
nal
lyopp
osed
100—
140
mV
/mm
DC
field
was
applie
d;m
ajority
ofc
ells
rem
ained
alig
ned;s
ome
cells
resp
onde
dto
both
cues
[93]
Fib
ronec
tin
coat
ing
Gla
ssco
ated
with
fibro
nec
tin
0.2–
5lm
wid
thSp
acin
gnot
liste
dBH
Kce
lls,ra
tte
ndon
fibr
obla
sts,
rat
dors
alro
otga
ngl
iace
lls,
P38
8D1
mac
rophag
es
Fib
ers
incr
ease
dsp
read
ing
and
alig
nmen
tin
dire
c-tion
offibe
r;ac
tin
alig
ned
infibro
blas
ts;a
lign
men
tof
foca
lco
nta
cts
infibro
bla
sts
and
mac
rophag
es;
incr
ease
dpol
ymer
izat
ion
ofF-a
ctin
;fib
ers
incr
ease
dsp
eed
and
per
sist
ence
ofce
llm
ovem
ent
and
rate
ofne
urite
outg
row
th;m
acro
pha
ges
had
actin-
rich
mic
rosp
ikes
and
bec
ame
pola
rize
dan
dm
igra
tory
[82]
Coa
ting
glas
sw
ith
pro
tein
and
withd
raw
ing
liquid
toor
ientpro
tein
Orien
ted
collag
enor
fibrin
Size
offibe
rsno
tlist
edSp
acin
goffibe
rsno
tlist
edH
um
anne
utr
oph
ille
ukoc
ytes
Cel
lste
nde
dto
mov
ein
direc
tion
offiber
axis
alig
nm
ent;
mov
emen
tw
asbi-di
rect
ional
;no
chem
otax
isev
iden
t
[104
]
Mic
ro-
text
ured
surf
ace
EC
Mre
plic
atio
n-P
MM
Apo
lym
eriz
atio
nca
stin
gfo
llow
edby
poly
ure
than
eso
lution
cast
ing
Poly
ure
than
epo
sitive
cast
of
PM
MA
neg
ativ
e
Mic
ron
and
nano
met
ersc
ale
topo
grap
hySi
milar
toEC
MBovi
ne
aort
icen
doth
elia
lce
llsC
ells
grow
non
repl
icas
ofEC
Msp
read
fast
eran
dha
dth
ree-
dim
ension
alap
pear
ance
and
spre
adar
eas
atco
nfluen
cew
hic
hap
pear
edm
ore
like
cells
inth
eir
native
arte
ries
than
cells
grow
non
unte
xtur
edco
ntro
lsu
rfac
es
[85]
Pol
ymer
s:P
MM
A:
poly
(met
hyl
met
hac
ryla
te);
PV
C:
poly
(vin
ylch
loride)
;PA
N:
pol
y(ac
rylo
nitrile
);P
DM
S:
poly
(dim
ethyl
silo
xane)
;P
ET:
pol
y(et
hyle
ne
tere
pht
hal
ate)
;N
IPA
M:
N-iso
pro
pyl-
acry
lam
ide;
PV
DF:pol
y(vi
nyl
iden
efluor
ide)
;TC
PS:
tiss
ue
culture
poly
styr
ene.
Oth
er:
PM
A:p
hor
bol12
-mys
trat
e13
-ace
tate
;EC
M:e
xtra
cellu
lar
mat
rix;
BH
K:b
aby
ham
ster
kid
ney
;MD
CK
:Mad
inD
arby
canin
ekid
ney
;GA
PD
:gly
cera
ldeh
yde-
3-deh
ydro
gena
se;L
PS:l
ipopol
ysac
-ch
arid
e;C
DP:co
llage
nas
edi
gest
able
pro
tein
.
582 R.G. Flemming et al. / Biomaterials 20 (1999) 573—588

Fig. 3. Schematic diagram of the pattern transfer process. Regions of the substrate not protected by resist are etched isotropically or anisotropically.Three examples of possible etching results are shown: anisotropic, reactive ion etching of silicon substrate to yield square grooves, anisotropic, wetetching of silicon [1 0 0] in KOH to yield V-shaped grooves, and isotropic, wet etching of glass in HF (that undercuts the resist). After the etching iscomplete, the remaining resist is removed.
Fig. 2. Schematic diagram of the photolithographic process. A substra-te coated with a thin polymeric resist in irradiated through a mask.Depending on the type of resist, the irradiated regions become eithermore soluble or less soluble in a developer. Immersion in the developeryields either a positive-tone or negative-tone image of the mask.
negative-tone image of the mask. Due to diffraction lim-itations, the smallest lateral, or width, dimension that canbe achieved by photolithography without using phase-shift masks are of the same size scale as the wavelength ofthe light source used to make them. Current state-of-the-art photolithography employs UV light of wavelength248 nm and is able to achieve features with lateral dimen-sions of 220—250 nm.
The regions of the substrate not covered by resist canthen be etched isotropically or anisotropically, as shownin Fig. 3, to transfer the pattern in the resist to theunderlying substrate. A common topographical featureinvestigated by many researchers was grooves, structuresmade by etching the substrate using a photoresist pat-terned in the shape of lines. Depending on the type of
etchant used, grooves can be constructed to have crosssections of square wave, V-shape, or truncated V-shape[36, 37]. A few examples of different shapes of etchedstructures are depicted in Fig. 3. Repeat spacing is con-sidered the distance from the beginning of a feature to thebeginning of the next feature. A groove and its associatedridge would thus be considered one repeat unit. The etchdepth and the lateral, or width dimensions are alsoshown in Fig. 3. After the etching is complete, the remain-ing resist is removed. The final product is a three-dimen-sional positive or negative relief image duplicating theopaque and transparent regions of the light mask.
One problem encountered with some of the etchingprocesses is undercutting of the resist, yielding shapeswith different dimensions than the patterned resist andwith curved sides. In addition, some etchants produceroughened surfaces. These roughened surfaces sometimessuperimpose random nanometer sized features on top ofthe intended topography [38, 39]. The effects of roughen-ed surfaces on cell behavior have yet to be addressed inthe literature. For a more complete discussion of thelithographic and pattern transfer processes, the reader isdirected elsewhere [36, 37].
Other techniques used to produce features with con-trolled dimensions include glancing angle deposition[40, 41], laser ablation [42, 43], laser deposition [44],replica molding of X-ray lithography masters [45, 46],imprint lithography [47, 48], microcontact printing andetching [49, 50], and ink-jet printing [51, 52]. Some ofthese techniques are capable of producing nanometerscale features. Electron-beam lithography, for example, isthe most developed, high-resolution lithographic tech-nique known, and has been used to fabricate featuresas small as 50 nm over large areas [53—55]. Thesetechniques, for the most part, are not represented inTable 1 because no data has been reported in the literature
R.G. Flemming et al. / Biomaterials 20 (1999) 573—588 583

concerning the behavior of cells upon surfaces createdby these techniques. Cell studies have been conductedon surfaces created by electron-beam lithography, but inthese studies the lateral, or width dimension was micronscale [56—58].
In some of the studies summarized in Table 1, thematerial on which cells were cultured was quartz, silicondioxide, or titanium-coated silicon. In other studies poly-meric replicas of the textured surfaces were created bycasting or embossing techniques [56—77]. For example,the fifth entry in Table 1 lists the fabrication technique asphotolithography and reactive ion etching, and the mate-rial on which the cells were cultured as an epoxy replicaof the silicon original. This means that the original pat-tern was created in silicon, which was used to makea negative polymeric cast, which, in turn, was used tocreate a final, positive replica. In some cases, such as inthe first entry, the negative cast was simply used in thecell assays. If the entry lists a polymer ‘cast’ in the‘Material’ column, then the cells were cultured on a nega-tive replica of the original surface. If the entry lists a poly-mer ‘replica’, then cells were cultured on a positive rep-lica. Polymers used for making such casts and replicasinclude PDMS, epoxy, and polystyrene. Other polymersused in studies include PMMA, PET, polycarbonate,polyetherimide, poly (NIPAM), nitrocellulose, and PVDF.
Most of the studies to date address the effects ofsurface features with dimensions of 0.5 lm and greater.The relevance of these studies to topography found onbasement membranes is unclear since membrane topo-graphy is comprised of much smaller nanometer scalefeatures, as discussed in the previous section. A few inves-tigators have fabricated structures with control over atleast one dimensional less than 500 nm [57, 58, 64, 65, 75,78—89]. These studies are highlighted in bold type inTable 1. In most of those studies, the lateral dimension ofthe features was greater than 0.5 lm, and the depthdimension was less than 0.5 lm. The depth dimension iseasily controlled in the etching process. Using reactiveion etching, Webb et al. [81] and Clark et al. [79]achieved grooves with depths of 100 nm. In anotherstudy, Clark et al. fabricated grooves with depths of200 nm, but with lateral dimensions of 2—12 lm on a sur-face of PMMA using photolithography (for patterningthe lateral dimensions) and reactive ion etching (to con-trol the etch depth) [78]. Wojciak-Stothard and cowork-ers used photolithography and reactive ion etching toproduce silicon grooves 2—10 lm in width and withdepths of 30—282 nm [80]. Several series of polystyreneridges with heights of 30 nm and widths of 0.5—100 lmwere made by Hoch et al. via electron-beam lithographyand reactive ion etching [65]. Grooves as shallows as14 nm and 1, 2, or 4 lm wide were achieved by Rajniceket al. [57, 58]. Britland and coworkers also used photoli-thography followed by reactive ion etching to producegrooves of 100 nm in depth, but 2.5—50 lm in width [86].
Cooper et al. were able to create radial arrays of groovesonly 50 nm in height [90]. Schmidt and von Recumreport round nodes only 380—460 nm in height [75].
Only two studies were found in which cell behaviorwas investigated on surfaces with features of controlledshape and size, and lateral dimensions less than 0.5 lm.Webb et al. [81] and Clark et al. [79] both used a laserholographic method developed for fabrication of masksfor X-ray lithography to create patterns of grooves inquartz with widths of 130 nm and depths of 100—400 and100—1170 nm, respectively.
Other methods have been used to fabricate surfaceswith nanoscale features, but with no direct control overthe topographical shapes and dimensions. Turner et al.used reactive ion etching to produce columnar structureswith average diameters of 57 nm [87]. Martin et al.sandblasted then acid-etched titanium surfaces to createfine and coarse structures with sharp features (100 nmas well as larger pits and craters [83]. Kononen et al.,Lampin et al., and Eisenbarth et al. have used sandblast-ing [88, 91] and sanding [89], respectively, to producefeatures less than 0.5 lm in size. Polymer-coated micro-porous filters [84] and fibronectin-coated glass [82] havealso been used as textured surfaces and have been foundto have features as small as 200 nm. Goodman et al.replicated the surface topography of extracellular matrixvia poly(methylmethacrylate) polymerization casting,which produced an inverse replica, followed by poly-urethane solution casting, which yielded the final positivereplica [85]. The microtextured surface produced by thismethod contained micron and nanometer scale topo-graphical features. Sims and Albrecht have also achievedsome success in replicating the surface structure of fibril-lar collagen, but interactions of cells with such surfaceshave not been reported [92].
Protein tracks have been used as guidance cues forcells. Laminin tracks have been patterned by adsorptionto hydrophobic silane tracks, which had been patternedusing photolithography [93, 94]. In another study, fibersof fibronectin as small as 200 nm in diameter were ad-hered to glass [82].
4. Effects of micro- and nano-structured surfaceson cell behavior
Studies of the interactions between substrate topogra-phy and cells have encompassed a wide variety of celltypes and substratum features including grooves, ridges,steps, pores, wells, nodes, and adsorbed protein fibers.Table 1 summarizes the literature, providing a list offeature type, fabrication technique employed, substratummaterial, feature size and spacing, cell type studied, andthe cellular effect generated by the surface features.
Grooves are the most common feature type employedin the study of the effects of surface structure on cells
584 R.G. Flemming et al. / Biomaterials 20 (1999) 573—588

[35, 56—64, 68—70, 72—74, 76, 78—81, 90, 95—104]. Typi-cally, the grooves are arrayed in regular, repeating pat-terns, often with equal groove and ridge width. The crosssections of the groove are often of the square wave,V-shape, or truncated V-shape [59, 80, 95].
In general, investigations of grooved surfaces revealedthat cells aligned to the long axis of the grooves [35,56—64, 68—70, 72—74, 76, 78—81, 95—104], often with or-ganization of actin and other cytoskeletal elements in anorientation parallel to the grooves [56, 69, 73, 80, 81, 99,100, 103]. The organization of cytoskeletal elements wasobserved to occur in some cases with actin and micro-tubules aligned along walls and edges [69, 98, 100]. Woj-ciak-Stothard et al. noted that F-actin condensationsappeared at topographic discontinuities, often at rightangles to groove edges [98]. They also observed thatmicrotubules formed after 30 min and that vinculin or-ganization was similar to that of actin. In addition, somecells were observed to have lamellae and filipodia be-nding around edges. Similar results were obtained byOakley and Brunette, who found that microtubules werethe first element to align to grooves, followed by actin[103]. Many studies found that the depth of grooves wasmore important than their width in determining cellorientation [78—80, 96]. Orientation often increased withincreasing depth, but decreased with increasing groovewidth. Repeat spacing also played a role, with orientationdecreasing at higher repeat spacing [76, 96]. In otherwords, as ridge width or groove width increased, theorientation phenomena of cells on grooves diminished.Dunn and Brown reported that ridge width is a moreimportant factor than groove width [96].
Certain hierarchical effects have been encountered aswell. Brunnette exposed cells to a pattern of major andminor grooves, in which the minor grooves were placedon the floor of the major grooves at an angle of 54° to themajor grooves [59]. Brunnette found that the cellsoriented preferentially to the major grooves, althoughcells did align to the minor grooves when major grooveswere not present or when a discontinuity existed in thepattern of major grooves. Britland et al. investigated thebehavior of cells upon grooves that were overlaid withadhesive tracks of silane and observed that the cellsresponded to both the topographical and adhesive cues,but that the adhesive cues had the dominant effect[86].
Although most cell types studied exhibited alignmentto grooves, some did not align. One study conducted byMeyle et al. found that, while 100% of fibroblasts alignedto grooves, alignment was observed in only 20% ofmacrophages and not at all in keratinocytes or neuro-phils [101]. Webb et al. noticed that oligodendrocytesand astrocytes were aligned by grooves, but that hippo-campal and cerebellar neuron cells were not aligned [81].In addition, although the oligodendrocytes were aligned,they exhibited little high-order F-actin organization. On
the other hand, the aligned astrocytes showed extensiveorganization of actin stress fibers. Rajnicek and McCraigobserved that Xenopus neurites grew parallel to grooves,but that rat hippocampal neurites grew perpendicular togrooves [57].
When ridges are placed far apart, relative to cellulardimensions, they may be considered individually as stepsor cliffs which the cell must encounter and traverse. Onlya few studies are available in the literature that deal withthis type of substratum topography: the study by Hochet al., which investigated the effect of ridges on funguscells [65], and the study by Clark et al., in which severaldifferent cell types were exposed to steps of varyingheights [105]. As might be expected, cell alignment in-creased with increasing step height.
There are some studies in the literature investiga-ting the behavior of cells on other synthetic features,including wells and nodes [67, 71, 75, 106, 107], pores[84, 107], spheres [108], and cylinders [109]. Green andcoworkers found that nodes of 2 and 5 lm resulted inincreased cell proliferation compared to 10 lm nodes andsmooth surfaces [67]. Cambell and von Recum examinedthe effects of pore size and hydrophobicity in their studyinvolving a canine in vivo implant model [84]. They usednylon mesh coated with polyvinylchloride/polyacrylonit-rile with or without an additional silicone coating andfound that pore size played a larger role than materialhydrophobicity in determining tissue response, withpores of 1—2 lm allowing for direct fibroblast attachment[84]. Fujimoto et al. investigated the behavior of cells onspheres and observed that cells responded to a change insphere size produced by an increase in temperature[108]. The cells released excess active oxygen whensphere diameters shrunk as a result of the temperaturebeing increased from 25 to 37°C.
There were several studies in the literature in whichtextured surfaces were created by techniques which yieldtopographical features that were less definable than thosepreviously discussed, but nevertheless equally interesting.The behavior of cells on sandblasted surfaces has beenstudied, although the observed trends seem less clearthan those on controlled morphologies, such as grooves.In general, adhesion, migration areas, and ECM produc-tion were greater on rougher surfaces, or those surfacessandblasted with larger grain sizes [83, 91, 110]. Turneret al. compared cell behavior on reactive ion and wetetched surfaces and found that primary cells behaved ina manner opposite that of a transformed cell line [87].The primary cells preferred the narrow, columnar struc-tures created by the reactive ion etching, but the trans-formed cells preferred the smoother, wet etched surfaces.
A few studies were found in which protein tracks wereemployed as guidance cues for several cell types, includ-ing neural cells [82, 93, 94]. Isolated tracks were found toprovide stronger guidance than repeated tracks [94].One interesting result from these studies was that cells
R.G. Flemming et al. / Biomaterials 20 (1999) 573—588 585

remained aligned to laminin tracks even when an ortho-gonal DC field was applied [93].
Lastly, Goodman et al. used polymer casting to repli-cate the topographical features of the extracellular matrix[85]. Goodman and coworkers observed that endothelialcells cultured on the ECM textured replicas spread fasterand had appearance more like cells in their native arteriesthan did cells grown on untextured surfaces [85].
5. Conclusions
Topographical cues, independent of biochemistry, gen-erated by the ECM may have significant effects uponcellular behavior. Clearly, substratum topography hasdirect effects on the abilities of cells to orient themselves,migrate, and produce organized cytoskeletal arrange-ments, as documented by the studies summarized here.However, the relevance of such studies to the behavior ofcells adhering to basement membranes is unclear sincethe topographical features of basement membranes havebeen shown to possess much smaller features in thenanometer size range [28—34]. The fact that basementmembranes are composed of unique and intricate topo-graphies into which cells adhere and extend processes,coupled with the fact that topographical features havebeen shown to influence cell behavior, leads to the hy-pothesis that the topography of the basement membraneis important in regulating cellular behavior in a mannerdistinct from that of the chemistry of the basement mem-brane.
It is therefore concluded, that, in order to fully under-stand the role that substrate topography plays in theregulation of cell behavior, smaller, denser, nanometerscale features must be fabricated. Techniques such aslaser holography and X-ray lithography offer the clearestway to synthesize topographical features with controlledlateral dimensions in the nanometer size range. Sincereactive ion etching is already a proven method for con-trolling etch depth, a coupling of etching techniques withstate-of-the-art lithographic techniques to create denser,nanometer scale features is encouraged in order to morefully understand the effects of substrate topography oncellular behavior.
References
[1] Coopman PJ, Bracke ME, Lissitzky JC et al. Influence of base-ment membrane molecules on directional migration of humanbreast cell lines in vitro. J Cell Sci 1991;98:395—401.
[2] Darnell J, Lodish H, Baltimore D. Multicellularity: cell—cell andcell—matrix interactions. Molecular Cell Biology. New York:Scientific American Books Inc., 1990:903—51.
[3] Carnegie JA, Cabaca O. Extracellular matrix composition andresilience: two parameters that influence the in vitro migrationand morphology of rat inner cell mass-derived cells. Biol Reprod1993;48:287—99.
[4] Podestra F, Roth T, Ferrara F, Lorenzi M. Cytoskeletal changesinduced by excess extracellular matrix impair endothelial cellreplication. Diabetologica 1997;40:879—86.
[5] Dua HS, Gomes JAP, Singh A. Corneal epithelial wound heal-ing. Br J Ophthalmol 1994;78:401—8.
[6] Juliano RL, Haskill S. Signal transduction from the extracellularmatrix. J Cell Biol 1993;120:577—85.
[7] Mousa SA, Cheresh DA. Recent advances in cell adhesion mol-ecules and extracellular matrix proteins: potential clinical ap-plications. DDT 1997;2:187—99.
[8] Chen CS, Mrksich M, Huang S, Whitesides G, Ingber DE. Geo-metric control of cell life and death. Science 1997;276: 1425—8.
[9] Clark EA, Brugge JS. Integrins and signal transduction path-ways: the road taken. Science 1995;268:233—9.
[10] Hynes RO. Integrins: versatility, modulation, and signaling incell adhesion. Cell 1992;69:11—25.
[11] Ruoslahti E. Stretching is good for a cell. Science 1997; 276:1345—6.[12] Grushkin-Lerner LS, Kewalramani R, Trinkaus-Randall V. Ex-
pression of integrin receptors on plasma membranes of primarycorneal epithelial cells is matrix specific. Exp Eye Res1997;64:323—34.
[13] Wu XY, Svoboda KKH, Trinkaus-Randall V. Distribution of F-actin, vinculin and integrin subunits (alpha6 and beta4) in re-sponse to corneal substrata. Exp Eye Res 1995;60:445—58.
[14] Trinkaus-Randall VCJ, Newton A, Vadasz A, Leibowitz HM,Franzblau C. Development of a biopolymeric keratoprosthesismaterial: evaluation in vitro and in vivo. Invest Ophthalmol VisSci 1988;29:293—300.
[15] Watanabe K, Nakagawa S, Nishida T. Stimulatory effects onfibronectin and EGF on migration of corneal epithelial cells.Invest Ophthalmol Vis Sci 1987;28:205—11.
[16] Watanabe K, Nakagawa S, Nishida T. Chemotactic and hap-totactic activities of fibronectin for cultured rabbit corneal epi-thelial cells. Invest Ophthalmol Vis Sci 1988;29:572—7.
[17] Reid TW. Growth control of cornea and lens epithelial cells.Prog Retinal Eye Res 1994;13:507—54.
[18] Choquet D, Felesenfiled DP, Sheetz MP. Extracellular matrixrigidity causes strengthening of integrin-cytoskeleton linkages.Cell 1997;88:39—48.
[19] Chiquet M, Matthisson M, Koch M, Tannheimer M, Chiquet-Ehrismann R. Regulation of extracellular matrix synthesis bymechanical stress. Biochem Cell Biol 1996;74:737—44.
[20] Curtis ASG, Clark P. The effects of topographic and mechanicalproperties of materials on cell behavior. Crit Rev Biocompata-bility 1990;5:343—62.
[21] von Recum AF, van Kooten TG. The influence of micro-topo-graphy on cellular response and the implications for siliconeimplants. J Biomater Sci Polym Ed 1995;7:181—98.
[22] Curtis A, Wilkinson C. Topographical control of cells. Bio-materials 1997;18:1573—83.
[23] Singhvi R, Stephanopoulos G, Wang DIC. Review: effects ofsubstratum morphology on cell physiology. Biotechnol Bioeng1994;43:764—71.
[24] Clark P. Cell behavior on micropatterned surfaces. BiosensBioelec 1994;9:657—61.
[25] Weiss P. Cell contact. Int Rev Cytol 1958;7:391—423.[26] Rosenberg MD. Long-range interactions between cell and sub-
stratum. Proc Natl Acad Sci 1962;48:1342—9.[27] Rosenberg MD. Cell guidance by alterations in monomolecular
films. Science 1963;139:411—2.[28] Abrams G, Goodman SL, Nealey PF, Franco M, Murphy CJ.
Nanoscale topography of the basement membrane underlyingthe corneal epithelium of the Rhesus Macaque. Cell Tissue Res,submitted.
[29] Abrams GA, Goodman SL, Nealey PF, Murphy CJ. Nanoscaletopography of the extracellular matrix underlying the cornealepithelium of the non-human primate. IOVS 1997;38:350.
586 R.G. Flemming et al. / Biomaterials 20 (1999) 573—588

[30] Abrams GA, Goodman SL, Nealey PF, Murphy CJ. Nanoscaletopography of the anterior and posterior basement membranesof the human cornea. IOVS 1998.
[31] Abrams GA, Goodman SL, Nealey PF, Murphy CJ. Nanoscaletopography of Matrigel. IOVS 1998;39:5160.
[32] Shirato I, Tomino Y, Koide H, Sakai T. Fine structure of theglomerular basement membrane of the rat kidney visualized byhigh resolution scanning electron microscopy. Cell Tissue Res1991;266:1—10.
[33] Yamasake Y, Makino H, Ota Z. Meshwork structures in bovineglomerular and tubular basement membrane as revealed byultra-high resolution scanning electron microscopy. Nephron1994;66:189—99.
[34] Hironaka K, Makino H, Yamsaki Y, Ota Z. Renal basementmembranes by ultrahigh resolution scanning electron micro-scopy. Kidney Int 1993;43:334—45.
[35] Brunette DM, Kenner GS, Gould TRL. Grooved titanium surfa-ces orient growth and migration of cells from human gingivalexplants. J Dent Res 1983;62:1045—8.
[36] Madou M. Fundamentals of microfabrication. Boca Raton, NewYork: CRC Press, 1997.
[37] Thompson LF, Willson CG, Bowden MJ. Introduction tomicrolithography. Washington DC: American Chemical So-ciety, 1995.
[38] Pearton SJ. High ion density dry etching of compound semicon-ductors. Mater Sci Eng 1996:B40:101—18.
[39] Perez-Murano F, Barniol N, Abadal G, Ye JH, Aymerich X,Cane C. Nanomodification of silicon (1 0 0) surface with scann-ing-tunneling-microscopy using polysilicon on silicon structure.Mater Sci Technol 1995;11:85—9.
[40] Robbie K, Brett MJ. Sculptured thin films and glancing angledeposition: growth mechanics and applications. J Vac Sci Tech-nol A 1997;15:1460—5.
[41] Robbie K, Sit JC, Brett MJ. Advanced techniques for glancingangle deposition. J Vac Sci Technol B 1998;16:1115—22.
[42] Phillips HM, Sauerbrey RA. Excimer-laser-produced nanostruc-tures in polymers. Opt Eng 1993;32:2424—36.
[43] Schleicher B, Friedlander SK. Characterization of nanostruc-tured carbon microwires fabricated by an aerosol process.J Appl Phys 1995;78:6046—9.
[44] McClelland JJ, Scholten RE, Palm EC, Celotta RJ. Laser-focused atomic deposition. Science 1993;262:877—80.
[45] Xia Y, Kim E, Zhao XM, Rogers JA, Prentiss M, WhitesidesGM. Complex optical surfaces formed by replica moldingagainst elastomeric surfaces. Science 1996;273:347—439.
[46] Xia Y, McClelland JJ, Gupta R et al. Replica molding usingpolymeric materials: a practical step toward nanomanufactur-ing. Adv Mater 1997;9:147—9.
[47] Chou SY, Krauss PR, Renstrom PJ. Imprint of sub-25 nm viasand trenches in polymers. Appl Phys Lett 1995;67:3114—6.
[48] Chou SY, Krauss PR, Renstrom PJ. Imprint lithography with25-nanometer resolution. Science 1996;272:85—7.
[49] Xia Y, Kim E, Whitesides GM. Microcontact printing of al-kanethiols on silver and its application in microfabrication.J Electrochem Soc 1996;143:1070—8.
[50] Xia Y, Kim E, Mrksich M, Whitesides GM. Microcontact print-ing of alkanethiols on copper and its application in microfabri-cation. Chem Mater 1996;8:601—3.
[51] Blanchard AP, Kaiser RJ, Hood LE. High-density oligonucleo-tide arrays. Biosens Bioelec 1996;11:687—90.
[52] Giordano RA, Wu BM, Borland SW, Cima LG, Sachs EM,Cima MJ. Mechanical properties of dense polylactic acid struc-tures fabricated by three dimensional printing. J Biomater SciPolym Ed 1996;8:63—75.
[53] Gold J, Nilsson B, Kasemo B. Microfabricated metal and oxidefibers for biological applications. J Vac Sci Technol A 1995;13:2683—44.
[54] Gold J, Kasemo B. Morphological and chemical characteriza-tion of microfabricated fibres for biological applications. JMater Sci Mater Med 1997;8:251—63.
[55] Terris BD, Mamin HJ, Best ME, Logan JA, Rugar D. Nanoscalereplication for scanning probe data storage. Appl Phys Lett1996;69:4262—4.
[56] Meyle J, Gultig K, Brich M, Hammerle H, Nisch W. Contactguidance of fibroblasts on biomaterial surfaces. J Mater SciMater Med 1994;5:463—6.
[57] Rajnicek AM, McCraig CD. Guidance of CNS growth cones bysubstratum grooves and ridges: effects of inhibitors of the cyto-skeleton, calcium channels and signal transduction pathways.J Cell Sci 1997;110:2915—24.
[58] Rajnicek AM, Britland S, McCraig CD. Contact guidance ofCNS neurites on grooved quartz: influence of groove dimen-sions, neuronal age and cell type. J Cell Sci 1997;110:2905—13.
[59] Brunette DM. Fibroblasts on micromachined substrata orienthierarchically to grooves of different dimensions. Exp Cell Res1986;164:11—26.
[60] Chehroudi B, Gould TRL, Brunette DM. Titanium-coated mi-cromachined grooves of different dimensions affect epithelialand connective-tissue cells differently in vivo. J Biomed MaterRes 1990;24:1203—19.
[61] Chesmel KD, Black J. Cellular responses to chemical and mor-phologic aspects of biomaterial surfaces. I. A novel in-vitromodel system. J Biomed Mater Res 1995;29:1089—99.
[62] den Braber ET, de Ruijter JE, Smits HTJ, Ginsel LA, von RecumAF, Jansen JA. Effect of parallel surface microgrooves andsurface energy on cell growth. J Biomed Mater Res 1995;29:511—8.
[63] den Braber ET, de Ruijter JE, Smits HTJ, Ginsel LA, von RecumAF, Jansen JA. Quantitative analysis of cell proliferation andorientation on substrata with uniform parallel surface micro-grooves. Biomaterials 1996;17:1093—9.
[64] den Braber ET, de Ruijter JE, Smits HTJ, Ginsel LA, von RecumAF, Jansen JA. Quantitative analysis of fibroblast morphologyon microgrooved surfaces with various groove and ridge dimen-sions. Biomaterials 1996;17:2037—44.
[65] Hoch HC, Staples RC, Whitehead B, Comeau J, Wolf ED.Signaling for growth orientation and cell differentiation by sur-face topography in Uromyces. Science 1997;235:1659—62.
[66] Chehroudi B, McDonnell D, Brunette DM. The effects of mi-cromachined surfaces on formation of bonelike tissue on subcu-taneous implants as assessed by radiography and computerimage processing. J Biomed Mater Res 1997;34:279—90.
[67] Green AM, Jansen JA, ven der Waerden JPCM, von Recum AF.Fibroblast response to microtextured silicone surfaces: textureorientation into or out of the surface. J Biomed Mater Res1994;28:647—53.
[68] Meyle J, Wolburg H, von Recum AF. Surface micromorphologyand cellular interactions. J Biomater Appl 1993;7:362—74.
[69] Meyle J, Gultig K, Wolburg H, von Recum AF. Fibroblastanchorage to microtextured surfaces. J Biomed Mater Res1993;27:1553—7.
[70] Ohara PT, Buck RC. Contact guidance in vitro: a light, trans-mission, and scanning electron microscopic study. Exp Cell Res1979;121:235—49.
[71] Schmidt JA, von Recum AF. Macrophage response to microtex-tured silicone. Biomaterials 1992;13:1059—69.
[72] Chehroudi B, Gould TR, Brunette DM. Effects of a groovedepoxy substratum on epithelial cell behavior in vitro and in vivo.J Biomed Mater Res 1988;22:459—73.
[73] den Braber ET, de Ruitjer JE, Ginsel LA, von Recum AF, JansenJA. Orientation of ECM protein deposition, fibroblast cyto-skeleton, and attachment complex components on siliconemicrogrooved surfaces. J Biomed Mater Res 1998;40:291—300.
R.G. Flemming et al. / Biomaterials 20 (1999) 573—588 587

[74] van Kooten TG, Whitesides JF, von Recum AF. Influence ofsilicone (PDMS) surface texture on human skin fibroblast prolif-eration as determined by cell cycle analysis. J Biomed Mater Res(Appl Biomater) 1998;43:1—14.
[75] Schmidt JA, von Recum AF. Texturing of polymer surfaces atthe cellular level. Biomaterials 1991;12:385—9.
[76] Brunette DM. Spreading and orientation of epithelial cells ongrooved substrata. Exp Cell Res 1986;167:203—17.
[77] Mrksich M, Chen CS, Xia Y, Dike LE, Ingber DE. Controllingcell attachment on contoured surfaces with self-assembledmonolayers of alkanethiolates on gold. Proc Natl Acad Sci1996;93:10775—8.
[78] Clark P, Connolly P, Curtis ASG, Dow JAT, Wilkinsin CDW.Topographical control of cell behavior. II. multiple groovedsubstrata. Development 1990;108:635—44.
[79] Clark P, Connolly P, Curtis ASG, Dow JAT, Wilkinson CDW.Cell guidance by ultrafine topography in vitro. J Cell Sci1991;99:73—7.
[80] Wojciak-Stothard B, Curtis A, Monaghan W, MacDonald K,Wilkinson C. Guidance and activation of murine macrophagesby nanometric scale topography. Exp Cell Res 1996;223:426—35.
[81] Webb A, Clark P, Skepper J, Compston A, Wood A. Guidanceof oligodendrocytes and their progenitors by substratumtopography. J Cell Sci 1995;108:2747—60.
[82] Wojciak-Stothard B, Denyer M, Mishra M, Brown RA. Ad-hesion, orientation, and movement of cells cultured on ultrathinfibronectin fibers. In Vitro Cell Dev Biol Animal 1997;33:110—7.
[83] Martin JY, Schwartz Z, Hummert TW et al. Effect of titaniumsurface roughness on proliferation, differentiation, and proteinsynthesis of human osteoblast-like cells (MG63). J BiomedMater Res 1995;29:389—401.
[84] Campbell CE, Von Recum AF. Microtopography and soft tissueresponse. J Invest Surg 1989;2:51—74.
[85] Goodman SL, Sims PA, Albrecht RM. Three-dimensional ex-tracellular matrix textured biomaterials. Biomaterials 1996;17:2087—95.
[86] Britland S, Morgan H, Wojiak-Stodart B, Riehle M, Curtis A,Wilkinson C. Synergistic and hierarchical adhesive and topo-graphic guidance of BHK cells. Exp Cell Res 1996;228:313—25.
[87] Turner S, Kam L, Isaacson M, Craighead HG, Shain W, TurnerJ. Cell attachment on silicon nanostructures. J Vac Sci TechnolB 1997;15:2848—54.
[88] Kononen M, Hormia M, Kivilahti J, Hautaniemi J, Thesleff I.Effect of surface processing on the attachment, orientation, andproliferation of human gingival fibroblasts on titanium. J Bio-med Mater Res 1992;26:1325—41.
[89] Eisenbarth E, Meyle J, Nachtigail W, Breme J. Influence of thesurface structure of titanium materials on the adhesion of fibro-blasts. Biomaterials 1996;17:1399—403.
[90] Cooper A, Munden HR, Brown GL. The growth of mouseneuroblastoma cells in controlled orientations on thin films ofsilicon monoxide. Exp Cell Res 1976;103:435—9.
[91] Lampin M, Warocquier-Clerout R, Legris C, Degrange M,Sigot-Luizard MF. Correlation between substratum roughnessand wettability, cell adhesion, and cell migration. J BiomedMater Res 1997;36:99—108.
[92] Sims PA, Albrecht RM. Corrosion casting in the reproduction ofthe micro-surface topography of fibrillar collagen. MicrosMicroanal 1998;4:916—7.
[93] Britland S, McCraig C. Embryonic Xenopus neurites integrateand respond to simultaneous electrical and adhesive guidancecues. Exp Cell Res 1996;225:31—8.
[94] Clark P, Britland S, Connolly P. Growth cone guidance andneuron morphology on micropatterned laminin surfaces. J CellSci 1993;105:203—12.
[95] Chou L, Firth JD, Uitto V-J. Brunette DM. Substratum surfacetopography alters cell shape and regulates fibronectin mRNAlevel, mRNA stability, secretion and assembly in human fibro-blasts. J Cell Sci 1995;108:1563—73.
[96] Dunn GA, Brown AF. Alignment of fibroblasts on groovedsurfaces described by a simple geometric transformation. J CellSci 1986;83:313—40.
[97] Wood A. Contact guidance on microfabricated substrata: theresponse of teleost fin mesenchyme cells to repeating topo-graphical patterns. J Cell Sci 1988;90:667—81.
[98] Wojciak-Stothard B, Madeja Z, Korohoda W, Curtis A, Wilkin-son C. Activation of macrophage-like cells by multiple groovedsubstrata. Topographical control of cell behavior. Cell Biol Int1995;19:485—90.
[99] Wojciak-Stothard B, Curtis ASG, Monaghan W, McGrath M,Sommer I, Wilkinson CDW. Role of the cytoskeleton in thereaction of fibroblasts to multiple grooved substrata. Cell MotilCytoskeleton 1995;31:147—58.
[100] Oakley C, Brunette DM. Response of single, pairs, and clustersof epithelial cells to substratum topography. Biochem Cell Biol1995;73:473—89.
[101] Meyle J, Gutlig K, Nisch W. Variation in contact guidance byhuman cells on a microstructured surface. J Biomed Mater Res1995;29:81—8.
[102] Chou L, Firth JD, Uitto VJ, Brunette DM. Effects of titaniumsubstratum and grooved surface topography on metallo-proteinase-2 expression in human fibroblasts. J Biomed MaterRes 1998;39:437—45.
[103] Oakley C, Brunette DM. The sequence of alignment of micro-tubules, focal contacts and actin filaments in fibroblasts spread-ing on smooth and grooved titanium substrata. J Cell Sci1993;106:343—54.
[104] Wilkinson PC, Shields JM, Haston WS. Contact guidance ofhuman neutrophil leukocytes. Exp Cell Res 1982;140:55—62.
[105] Clark P, Connolly P, Curtis ASG, Dow JAT, Wilkinson CDW.Topographical control of cell behavior: I. Simple step cues.Development 1987;99:439—48.
[106] Hunt JA, Williams RL, Tavakoli SM, Riches ST. Laser surfacemodification of polymers to improve biocompatibility. J MaterSci Mater Med 1995;6:813—7.
[107] Fewster SD, Coombs RRH, Kitson J, Zhou S. Precise ultrafinesurface texturing of implant materials to improve cellular ad-hesion and biocompatibility. Nanobiol 1994;3:201—14.
[108] Fujimoto K, Takahashi T, Miyaki M, Kawaguchi H. Cell activa-tion by the micropatterned surface with settling particles. J Bio-mater Sci Polym Ed 1997;8:879—91.
[109] Rovensky YA, Samoilov VI. Morphogenetic response of cul-tured normal and transformed fibroblasts, and epitheliocytes, toa cylindrical substratum surface. Possible role for the actinfilament bundle pattern. J Cell Sci 1994;107:1255—63.
[110] Groessner-Screiber B, Tuan RS. Enhanced extracellular matrixproduction and mineralization by osteoblasts cultured on tita-nium surfaces in vitro. J Cell Sci 1992;101:209—17.
[111] Rosdy M, Grisoni B, Clauss LC. Proliferation of normal humankeratinocytes on silicone substrates. Biomaterials 1991;12:511—7.
[112] Rich A, Harris AK. Anomalous preferences of cultured macro-phages for hydrophobic and roughened substrata. J Cell Sci1981;50:1—7.
[113] Geunard V, Valenti RF, Aebischer P. Influence of surface textureof polymeric sheets on peripheral nerve regeneration in a two-compartment guidance system. Biomaterials 1991;12:259—63.
588 R.G. Flemming et al. / Biomaterials 20 (1999) 573—588